 Côme J. Thieulent1
Côme J. Thieulent1 Melissa Laverack2
Melissa Laverack2 Mariano Carossino1,3Brittany Cronk2
Mariano Carossino1,3Brittany Cronk2 Leonardo Cardia Caserta2
Leonardo Cardia Caserta2 Diego G. Diel2
Diego G. Diel2 Udeni B. R. Balasuriya1*
Udeni B. R. Balasuriya1*- 1Department of Pathobiological Sciences, School of Veterinary Medicine, Louisiana State University, Baton Rouge, LA, United States
- 2Department of Population Medicine and Diagnostic Sciences, Cornell University College of Veterinary Medicine, Ithaca, NY, United States
- 3Louisiana Animal Disease Diagnostic Laboratory (LSU Diagnostics), School of Veterinary Medicine, Louisiana State University, Baton Rouge, LA, United States
Canine Infectious Respiratory Disease Complex (CIRDC), caused by a diverse range of viral and bacterial pathogens, is the leading cause of respiratory illness in dogs. In the winter of 2023–2024, the United States experienced a noticeable increase in cases consistent with CIRDC. This study investigated the potential association of emerging pathogens with CIRDC cases. It involved the analysis of 50 clinical specimens collected from CIRDC-suspected dogs from six US states between December 2023 and February 2024. All clinical cases presented with respiratory illness characterized mainly by coughing (78%), nasal and ocular discharges (30%), and sneezing (22%). Specimens were tested for 12 known CIRDC-associated pathogens using a previously described panel of one-step TaqMan® multiplex qPCR/RT-qPCR assays designed to detect eight viral and four bacterial pathogens. Specimens were also subjected to next-generation sequencing (NGS) to confirm qPCR/RT-qPCR results and identify potential emerging pathogens. In this study, 64% of samples tested positive for various canine respiratory pathogens. Mycoplasma canis was the most frequently detected agent (n = 20), followed by M. cynos (n = 9), canine respiratory coronavirus (n = 3), canine parainfluenza virus (n = 3), and Bordetella bronchiseptica (n = 3). Additionally, canine adenovirus type 2, canine herpesvirus 1, and canine distemper virus were also detected in some samples. NGS also identified canine calicivirus, canine circovirus, and, for the first time, vientovirus in a CIRDC-affected dog. This study indicates that CIRDC cases observed in the winter of 2023–2024 were not associated with the emergence of any new pathogens. The clinical relevance of the detection of vientovirus in a single dog remains unknown.
1 Introduction
Canine Infectious Respiratory Disease Complex (CIRDC), commonly known as kennel cough, is a highly prevalent and contagious respiratory condition in the dog population worldwide (1, 2). Outbreaks most commonly occur in settings where dogs are housed or gathered nearby, such as boarding facilities, animal shelters, and dog daycares (3, 4). CIRDC manifests as a multifactorial syndrome characterized by acute respiratory signs, including coughing, nasal or ocular discharge, sneezing, fever, and respiratory distress, that generally lasts up to 2 weeks (1). The disease is primarily transmitted through direct dog-to-dog contact and airborne transmission via respiratory secretions (2). Most infections are self-limiting, with affected dogs typically recovering within a few days to a few weeks. CIRDC is a complex disease involving a wide array of viral and bacterial agents, including canine adenovirus 2 (CAdV-2) (5), canine distemper virus (CDV; Morbilivirus canis) (6), canine herpesvirus 1 (CaHV-1; Varicellovirus canidalpha1) (7), canine influenza virus (CIV) (8), canine pneumovirus (CnPnV) (9, 10), canine parainfluenza virus (CPiV) (11), canine respiratory coronavirus (CRCoV) (12, 13), Bordetella bronchiseptica (14), Mycoplasma canis (15), M. cynos (16), and Streptococcus equi zooepidemicus (17, 18). Additionally, sporadic infections with other canine viruses were also reported, including canine calicivirus, canine circovirus, and canine hepacivirus (19–22). Co-infections and certain host factors can contribute to more severe clinical outcomes (23–26). Vaccines targeting specific pathogens associated with CIRD, such as B. bronchiseptica, CAdV-2, CDV, and CPiV, are available in the United States and are commonly administered to puppies (27). Nevertheless, outbreaks continue to be frequently reported (28, 29).
Accurate aetiologic diagnosis of CIRDC is challenging due to the overlapping clinical presentations among pathogens and the frequent occurrence of asymptomatic carriers. For example, B. bronchiseptica, M. cynos, CaHV-1, and CRCoV are often found in both healthy and clinically ill dogs (12, 30). As such, their contribution to the clinical signs observed in dogs remains uncertain. Asymptomatic dogs significantly contribute to the spread of infections and illnesses among susceptible dogs and may also serve as a reservoir for pathogens between disease outbreaks. Most pathogens have a short incubation period, typically from a few days up to 2 weeks (2). However, given the similarity in clinical signs induced by these pathogens, diagnosis cannot rely solely on history and clinical signs. Thus, laboratory confirmation is essential to confirm clinical diagnosis by accurate pathogen identification. Molecular diagnostic methods such as polymerase chain reaction (PCR) and quantitative PCR (qPCR or reverse transcriptase-qPCR [RT-qPCR]) are widely used for detecting CIRDC agents (31–33). These assays offer high sensitivity and specificity, enabling the detection of active infections. Multiplex qPCR/RT-qPCR platforms, in particular, allow simultaneous detection of multiple pathogens in clinical specimens, making them well-suited for investigating the complex etiology of CIRDC (26, 34). Furthermore, next-generation sequencing (NGS) has become a rapid and comprehensive technique for identifying and characterizing multiple pathogens, including the detection of novel agents in clinical specimens (35). Therefore, these novel molecular techniques (multiplex qPCR/RT-qPCR and NGS) offer high sensitivity, specificity, and quick turnaround time compared to classical virology and bacterial isolation and characterization methods.
In late 2023, veterinarians in several states across the US reported cases of an unusual respiratory illness in dogs, referred to as atypical canine infectious respiratory disease complex (aCIRDC). These cases raised alarms about the possible emergence of a new pathogen (36–39). Unlike typical infections seen in previous years, affected dogs experienced prolonged and severe respiratory clinical signs. Notably, the animals often tested negative for common pathogens associated with canine infectious respiratory disease (CIRD). The atypical nature of CIRDC was further underscored by its poor response to standard antibiotic treatments and a noticeable rise in mortality rates (40). This study aimed to investigate a potential emerging pathogen associated with CIRDC cases during the winter of 2023–2024. A comprehensive one-step multiplex qPCR/RT-qPCR assay was employed to detect the most common CIRDC pathogens, including CAdV-2, CDV, CaHV-1, CIV, CnPnV, CPiV, CRCoV, B. bronchiseptica, M. canis, M. cynos, and S. equi subsp. zooepidemicus, and SARS-CoV-2. Furthermore, specimens were tested for the possible presence of novel or emerging pathogens by NGS.
2 Materials and methods
2.1 Clinical specimens
Between December 2023 and February 2024, a total of 50 clinical specimens (pooled nasal and nasopharyngeal swabs) from dogs suspected of having CIRDC were submitted for routine diagnostic testing to the Louisiana Animal Disease Diagnostic Laboratory (LSU Diagnostics) in Baton Rouge, Louisiana, United States (Table 1). Primary practicing veterinarians submitted all specimens to LSU Diagnostics for routine testing; thus, IACUC approval or owner consent was not required for this study. Only samples collected from dogs with respiratory signs were included in this study, regardless of age, body weight, reproductive status, sex, or breed. Both nasal and oropharyngeal specimens were collected by the primary veterinarian using 15 cm sterile flocked collection swabs (VWR, Radnor, PA). The specimens were pooled in 2 mL of BHI Broth (Hardy Diagnostics, Santa Maria, CA) and shipped overnight at 4°C to the LSU Diagnostics. The diagnostic laboratory provided collection swabs and BHI broth along with instructions on how to collect the specimens for the primary practicing veterinarians to ensure optimal sample collection and consistency. The specimens in BHI Broth were packed in ice, and either hand-delivered or submitted vial overnight courier service to LSU Diagnostics for molecular testing. Upon receipt, the samples were immediately processed for molecular diagnostic testing (i.e., multiplex qPCR/RT-qPCR). The remaining BHI broth from each sample was then aliquoted into 500 μL aliquots and stored at −80°C for further use in NGS analysis. Virus and bacteria isolation were not performed.
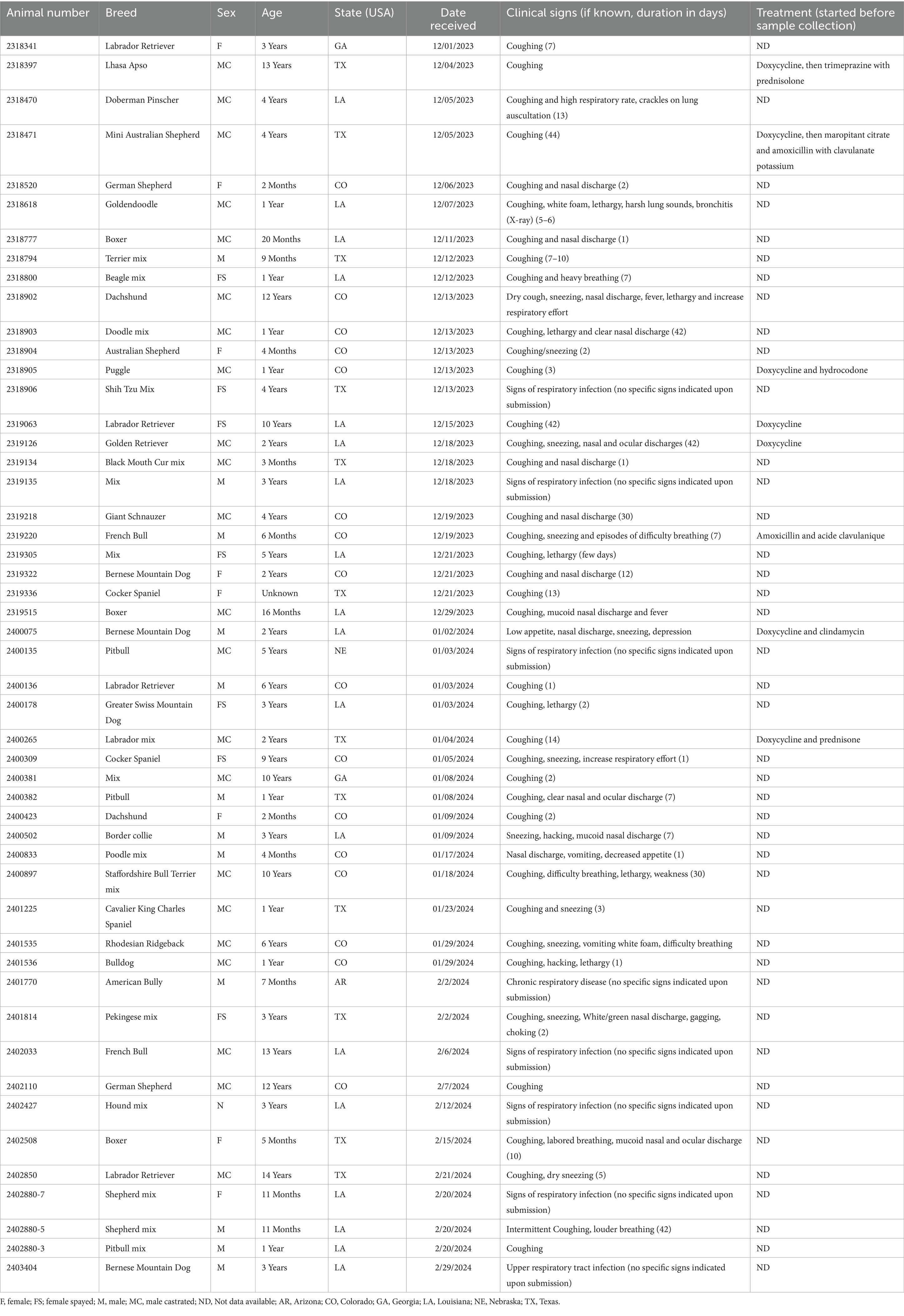
Table 1. Signalment of the samples included in this study (n = 50).
2.2 Detection of canine respiratory pathogens using one-step multiplex qPCR/RT-qPCR
Each BHI Broth tube with the swabs was vortexed, spun down, and nucleic acid was extracted using the taco™ mini nucleic acid automatic extraction system (GeneReach, Taichung, Taiwan) following the manufacturer’s recommendations. Nucleic acid was extracted from 100 μL of BHI Broth and then eluted in 100 μL of elution buffer following the manufacturer’s instructions. The extracted nucleic acid from each sample was immediately used in multiplex qPCR/RT-qPCR testing, and the remaining nucleic acid was archived at −80°C. Routine diagnostic testing was conducted using multiplex one-step qPCR/RT-qPCR assays to detect 12 canine respiratory pathogens, including CAdV-2, CDV, CaHV-1, CIV, CPiV, CPnV, CRCoV, SARS-CoV-2, B. bronchiseptica, M. canis, M. cynos, and S. equi subsp. zooepidemicus, as previously described (26). Briefly, five μl of template nucleic acid per reaction was mixed with the master mix in a final volume of 25 uL reaction and ran on a 7,500 Fast Real-Time PCR System (Applied Biosystems, Waltham, MA) with the following thermal profile: a reverse transcription step (20 min. at 50°C) followed by an initial activation step (15 min. of at 95°C) and 40 cycles of denaturation and annealing/extension (45 sec. at 94°C and 75 s. at 60°C). The cycle threshold (Ct) cut-off for the detection of positive/negative samples was used as previously described (26).
2.3 Detection of canine respiratory pathogens using next-generation sequencing
After nucleic acid extraction for real-time RT-qPCR/qPCR, the remaining BHI Broth samples were aliquoted (approximately 500 μL per vial) and stored at −80°C. Approximately 500 μL of the frozen archived samples were shipped on dry ice to the Animal Health and Diagnostic Center (AHDC) at Cornell University, NY, a leading NGS laboratory for NGS as previously described (41). The NGS protocol applied was specifically optimized for virus detection and was used here exclusively for research purposes, rather than routine diagnostic testing. Upon receipt, samples were subjected to an enzymatic cocktail treatment composed of 10 × DNase I buffer, DNase I, Turbo DNase, RNase Cocktail™ (Thermo Fisher Scientific, Waltham, MA), Baseline-ZERO DNase (Lucigen, Middleton, WI), Benzonase (Sigma-Aldrich, Saint-Louis, MO) and RNase ONE™ Ribonuclease (Promega, Madison, WI) to deplete host and bacterial nucleic acids. Additionally, Mengovirus strain vMC0 (Mengo Extraction control kit, bioMérieux, Marcy-l’Étoile, France) was added to each sample at a concentration of 2.17 × 106 copies/ml, serving as an exogenous internal control for the NGS sample preparation, library preparation, and sequencing reactions (42). The enzyme cocktail, Mengovirus, and sample mixture were incubated for 90 min at 37°C. After enzymatic treatment, nucleic acid extraction was performed using the QIAamp® MinElute® Virus Spin Kit (Qiagen, Hilden, Germany). Purified nucleic acids were subjected to sequence-independent, single-primer amplification (SISPA) procedures modified from a previously reported protocol (43). In brief, 11 μL of nucleic acids were used in a reverse transcription reaction with 100 pmol of primer FR20RV-12 N (5′-GCCGGAGCTCTGCAGATATCNNNNNNNNNNNN-3′) using SuperScript™ IV reverse transcriptase (Thermo Fisher Scientific), followed by second-strand synthesis using the Klenow Fragment of DNA polymerase (New England Biolabs, Ipswich, MA) with primer FR20RV-12 N at 10 pmol. After purification using Agencourt AMPure XP beads (Beckman Coulter, Brea, CA), SISPA PCR amplification was conducted with TaKaRa Taq DNA Polymerase (Takara, Kusatsu, Japan) using the primer FR20RV (5′-GCCGGAGCTCTGCAGATATC-3′) at 10 pmol. Sequencing libraries were prepared using 143 ng of double-stranded DNA as input to the SQK-LSK109 kit and barcoded individually using the EXP-NBD196 Native barcodes (Oxford Nanopore Technologies (ONT), Oxford, United Kingdom). The sequencing was performed on the FLO-MIN106 MinION flow cell r9.4.1 using the GridION Sequencer (ONT). High-accuracy base calling was performed by the GridION with the parameters “--require_barcodes_both_ends” and “--detect_mid_strand_barcodes.” Fastq reads were then filtered by size and quality using Chopper (44), host-removed using NanoLyse (45), and classified using Kraken (v2.1.0) (46, 47) followed by relative abundance estimation using Bracken. Bioinformatics data analysis was performed using Base2Bio (Oshkosh, WI).
2.4 Detection of vientovirus by standard PCR and phylogenetic analysis
The presence of the vientovirus in original clinical sample was confirmed by two distinct PCR assays targeting capsid (Cap) and the replicative (Rep) open reading frames in a total volume of 25 μL containing 12.5 μL of AccuStart II PCR ToughMix (2X) (Quantabio, Beverly, MS), 2.5 μL of a forward and reverse primer mix (5 μM; F_Cap: GATATGCATCAAGAAAGAGAGTTTATCG; R_Cap: ATTCTTAACACCTTTGCCAGAAATC; F_Rev: CCGTCTAGTAATCTGAGGAGGA; R_Rev: TGTGGTTTCATGGAGATACAGG), 5 μL of nuclease-free water and 5 μL of DNA template. Thermal cycling was performed on a Mastercycler® × 50 – PCR Thermocycler (Eppendorf, Hamburg, Germany) using the following conditions: an initial denaturation step (3 min. at 94°C) and 40 cycles of denaturation, annealing, and extension (30 s. at 94°C, 30 s. at 60°C, 30 s. at 70°C). The PCR products were analyzed by gel electrophoresis on a 1% agarose gel (ThermoFisher Scientific, Waltham, MA), and the expected band sizes were as follows: Cap: 200 bp; Rev.: 218 bp. The PCR products were submitted for Sanger sequencing to Eurofins Genomics (Louisville, KY).
3 Results
3.1 Clinical history
A total of 50 pooled nasal and oropharyngeal swab samples from dogs with respiratory illness were submitted to LSU Diagnostic Laboratory—24 in December 2023, 15 in January 2024, and 11 in February 2024 (Table 1). Out of the 50 samples, 35 were collected from male (70%) and 15 were collected from female (30%) dogs. The age ranged from 2 months to 13 years (average = 3.8 years). Samples were received from six states, including Arizona (AR; n = 1), Colorado (CO; n = 16), Georgia (GA; n = 2), Louisiana (LA; n = 18), Nebraska (NE; n = 1), and Texas (TX; n = 12). Among the clinical signs reported by the veterinarians, coughing was the most prevalent (n = 39; 78%), followed by nasal and ocular discharges (n = 15; 30%), sneezing (n = 11; 22%), lethargy (n = 7; 14%) and high respiratory rate/heavy breathing (n = 7; 14%). Other clinical signs, including fever (n = 2; 4%), vomiting (n = 2; 4%), decreased appetite (n = 2; 4%), choking (n = 2; 4%), hacking (n = 2; 4%) and weakness (n = 1; 2%) were also reported. For six of the dogs, signs of respiratory infection were indicated, but no specific signs were provided in the submission forms. Clinical signs were reported up to 44 days before specimens were collected (Interquartile range [IQR] = 11).
3.2 Pathogen identification using multiplex qPCR/RT-qPCR assays
Thirty samples (60%) were positive for at least one of the tested pathogens, while twenty samples (40%) were negative (Table 2). Eight different pathogens were detected, including M. canis (n = 20; 40%), M. cynos (n = 9; 18%), CRCoV (n = 3; 6%), CPiV (n = 3; 6%), B. bronchiseptica (n = 3; 6%), CAdV-2 (n = 1; 2%), CaHV-1 (n = 1; 2%) and CDV (n = 1; 2%). Eight samples were positive for more than one pathogen, including seven positive samples for two pathogens (CRCoV and M. canis [n = 2], CPiV and M. canis [n = 2], M. cynos and M. canis [n = 3]) and one sample positive for five pathogens (CDV, CRCoV, CaHV-1, B. bronchiseptica, and M. canis). M. canis was detected in all the samples where more than a single pathogen was identified.
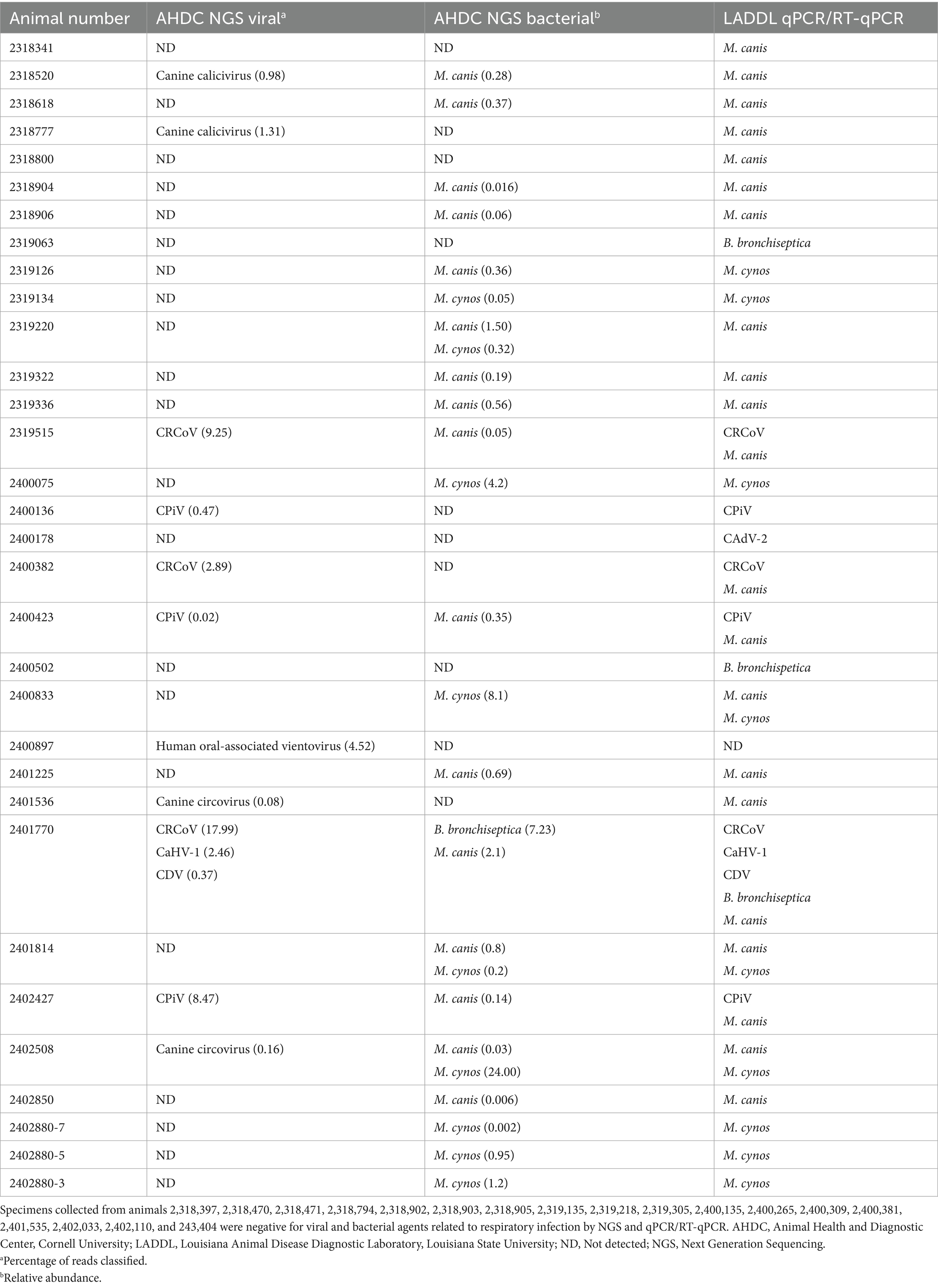
Table 2. Detection of viral and bacterial agents in samples collected from dogs with respiratory disorders using NGS and qPCR/RT-qPCR.
3.3 Pathogen identification using next-generation sequencing
NGS results matched those of qPCR/RT-qPCR in forty-one samples (82%), while discrepancies were noted in nine samples (18%) (Table 2). Among these, one sample was positive for M. cynos by qPCR, but M. canis was identified by NGS, and another sample was positive for M. canis by qPCR, while both M. canis and M. cynos were identified by NGS. In the remaining seven specimens with discrepancies, NGS did not detect B. bronchiseptica, M. canis, and CAdV-2, while qPCR/RT-qPCR did. Importantly, NGS identified canine calicivirus (n = 2) and canine circovirus (n = 2), two viruses that were not included in the multiplex qPCR/RT-PCR panel. Both canine calicivirus-positive samples and one of the canine circovirus-positive samples were also positive for M. canis, while the second circovirus-positive sample tested positive for M. cynos. Additionally, vientovirus was identified in one sample collected from a dog that presented with coughing, difficulty breathing, lethargy, and weakness for 30 days, with the sample tested being collected at that timepoint. No other pathogens were detected in this specimen. The specimens that yielded contradicting results with multiplex qPCR/RT-qPCR versus NGS were retested to confirm the validity of the test results. As expected, the results remained unchanged in repeat testing and NGS. Finally, in 18 (36%) of the samples, no pathogens were detected by either multiplex real-time qPCR/RT-qPCR or by NGS.
3.4 Confirmation of vientovirus DNA by standard PCR and phylogenetic analysis of capsid and replicative genes
To confirm the presence of vientovirus DNA in sample 2,400,897, two specific PCR assays targeting the capsid (Cap) and replicative (Rep) genes were developed based on the full-genome sequence obtained by NGS and the sequence was deposited in GenBank (Vientovirus/USA/CO/2024/2400897; GenBank accession #: PQ450187). These assays detected vientovirus in the original specimen but not in any other samples (Supplementary Figure 1A) and were further validated by Sanger sequencing. Phylogenetic analysis showed high similarity to known vientovirus isolates (Supplementary Figures 1B,C). To rule out human DNA contamination, species-level classification of 500 raw reads revealed that over 60% aligned with Canis lupus familiaris, 0.51% corresponded to the human oral-associated vientovirus AV, and only 0.25% mapped to human DNA (Supplementary Figure 1D).
4 Discussion
CIRDC is a complex infectious disease in dogs, attributable to single or synergistic actions of diverse viral and bacterial agents (1, 23). A rapid detection of the implicated pathogens is important to provide the most appropriate treatment strategy and implement proper biosecurity measures to prevent the spread of respiratory infections. Moreover, the detection of emerging pathogens is essential to understand their epidemiology and to inform owners and public health officials. An increase in the number of CIRDC cases in the US was perceived in late 2023 and was relayed by several public news outlets (36–39). Between December 2023 and February 2024, 50 nasal and nasopharyngeal swab specimens were collected from dogs experiencing respiratory disease and submitted for diagnostic testing to LSU Diagnostics. These samples were tested for common canine respiratory pathogens by multiplex qPCR/RT-qPCR and by NGS to attempt the identification of potential emerging pathogens.
When considering both qPCR/RT-qPCR and NGS results, 32 samples (64%) were positive for one pathogen known or suspected to induce respiratory infections in dogs. M. canis and M. cynos are the most common pathogens identified from CIRDC dogs, which is consistent with previous findings (24, 26, 34). Additionally, cases of CRCoV, CPiV, and B. bronchiseptica were reported, aligning with earlier reports (23, 48). The use of NGS has the advantage of allowing the identification of pathogens not included in the qPCR/RT-qPCR diagnostic panels. Here, canine calicivirus, canine circovirus and, for the first time, vientovirus (family: Redondoviridae) were detected in dogs with CIRDC. Canine calicivirus (CaCV) is an unclassified virus in the family Caliciviridae. CaCV was isolated and characterized for the first time in 1985 from the feces of a diarrheic dog in the US (49). Clinical signs induced by CaCV are diverse and range from the absence of clinical signs to severe diseases characterized by bloody diarrhea, hemorrhagic gastroenteritis, vomiting, and depression (20). However, CaCV has not been detected in respiratory tract specimens before. Additionally, experimentally infected dogs with different strains of CaCV failed to induce respiratory clinical signs (20, 49). Canine circovirus (species: Circovirus canine; genus: Circovirus; family: Circoviridae) was detected for the first time in 2011 in the serum of healthy dogs (50). The pathogenicity of canine circovirus is currently poorly understood, with the virus being mainly detected in fecal samples. Canine circovirus infection has been associated with clinical profiles of acute gastroenteritis, hemorrhagic diarrhea, signs of vasculitis, lymphadenitis, thrombocytopenia, neutropenia, and lymphopenia (51). Canine circovirus has also been associated with respiratory signs in dogs such as dyspnea, nasal discharge, cough, or rales (52). However, the virus has also been detected in samples of asymptomatic dogs, so its role in clinical disease and its pathogenesis is not clearly defined. It is interesting to note that M. canis, which is commonly found in the respiratory tract of healthy dogs, was detected in all samples that tested positive for CaCV and canine circovirus. Although it is not yet clear whether M. canis contributes to secondary infections in the upper airways, it is generally considered a commensal or opportunistic cofactor in respiratory and urogenital tract diseases in dogs (15). In this study, vientovirus DNA was found in a nasal/pharyngeal swab from one dog with prolonged respiratory illness (30 days) with no other detectable pathogens. Vientovirus (Torbevirus viento), a member of the Redondoviridae family (53), has so far been identified only in human respiratory samples through metagenomics, with no confirmed detection in animals (such as pigs, chickens, ducks, and dogs) or the environment (freshwater, marine environments, or soil) (54, 55). The transmission of this virus from humans to animals or animals to humans has not yet been demonstrated. Although prior infection with common respiratory agents cannot be ruled out for the dog positive for vientovirus DNA, the relevance of this virus to the clinical signs remains uncertain. Both NGS and PCR confirmed its detection. However, sample collection occurred late in the illness, and no other samples were found to be positive. While human DNA contamination was considered, it is unlikely due to the low human read counts and the inability to detect this virus in other specimens, simultaneously processed and tested by the same technicians. However, the detected viral sequence showed greater than 90% similarity to human-associated strains. Without supporting pathological or serological data, the role of the vientovirus virus in canine disease remains unclear.
Our results indicate that dogs from different states were infected with several known canine pathogens during the winter of 2023–2024, originating from six states in the US. Previously, studies have focused on detecting the nasal virome in dogs, which identified similar pathogens, including a novel Taupapillomavirus (CPV21–23) (56). Similarly, studies have focused on identifying and comparing the canine microbiome, which has revealed numerous bacterial taxa (57). It is important to highlight that the NGS protocol employed in this study was specifically optimized for viral detection, showing concordant results with the multiplex qPCR/RT-qPCR. Although the NGS protocol was not optimized for bacterial identification, it was applied for this purpose, which may partly explain the differences observed between the two methods in detecting bacterial pathogens, particularly the reduced bacterial detection by NGS. The nucleic acid extraction method, which includes a step to deplete host nucleic acids, may have contributed to the failure to detect certain bacteria. While our findings highlight several pathogens detected during the outbreak, it is important to consider factors that may influence diagnostic test results and the interpretation of negative findings. The quality of test results depends on the timely collection of appropriate samples during the infection. A negative qPCR/RT-qPCR or NGS test result means that the target virus or bacteria were not detected in the specimen. This could be due to the sample being collected too early or too late in the course of infection, resulting in insufficient target DNA or RNA for detection, or because no infection was present. In this study, 38% of the samples yielded negative results. The absence of pathogen identification in these cases could be attributed to several factors, including, but not limited to, lower respiratory tract infections, nucleic acids depletion techniques used during samples preparation for NGS (i.e., specific for bacteria detection), pathogen clearance before sample collection, antibiotic treatment by veterinarians, or non-infectious causes that the attending veterinarians may have considered. Based on available information, antibiotic treatment had been initiated in eight dogs before sample collection, which may have influenced the results by reducing the detectability of primary bacterial pathogens at the time of sampling. Follow-up was not possible for these cases, which limited the ability to gather additional clinical data or repeat testing to clarify the etiology of illness. Further investigation into the other respiratory causes of these negative cases could provide valuable insights and may be an important area for future research. Given the limited number of samples available during the Winter 2023–2024 outbreak (n = 50), it is essential to note that the relatively small sample size does not represent the overall US canine population, and the findings of this study should be considered exploratory. Nevertheless, the results provide valuable preliminary insights that may inform future, larger-scale investigations into CIRDC-associated pathogens. Additionally, the vaccine status of these animals was unknown. For these reasons, the association of other known or new respiratory pathogens in CIRDC cases across the country cannot be ruled out. Finally, nasal and pharyngeal swabs were pooled together before performing the diagnostic assays; therefore, the comparison of the two collection methods was not possible in this study. The SISPA method is a valuable tool for viral and bacterial metagenomic studies (58–60); however, like other NGS methods, it has limitations that affect its sensitivity. A key challenge is the requirement for high viral loads for adequate amplification, as lower concentrations lead to non-specific products and poor genome recovery. SISPA is also prone to amplification biases, frequently resulting in incomplete viral genome assemblies, particularly when viral titers are low (61). Additionally, background contamination from host nucleic acids can reduce sensitivity, necessitating stringent preprocessing and enrichment strategies.
While Mycoplasma species were detected at a high rate in these samples, we believe it is unlikely that they were the primary pathogen responsible for the outbreak. Instead, it may have acted as a secondary pathogen, contributing to the overall disease burden. Future studies focusing on viral and bacterial co-infections, as well as the timing of pathogen detection relative to clinical signs, would be valuable in further elucidating the role of Mycoplasma in CIRDC.
In summary, our results suggest that the increase in CIRDC cases observed in the winter of 2023–2024 is unlikely to be attributed to a single pathogen or the emergence of a novel viral or bacterial pathogen. No conclusive evidence of a new causative agent was identified in this study.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Ethics statement
Ethical approval was not required for the study involving animals in accordance with the local legislation and institutional requirements because samples were submitted for routine diagnostic testing to the Louisiana Animal Disease Diagnostic Laboratory by primary practicing veterinarians. Thus, IACUC approval or owner consent was not required for this study.
Author contributions
CT: Validation, Investigation, Formal analysis, Methodology, Data curation, Conceptualization, Visualization, Writing – original draft. ML: Writing – review & editing. MC: Methodology, Supervision, Writing – review & editing, Conceptualization, Funding acquisition, Validation. BC: Writing – review & editing. LC: Writing – review & editing. DD: Writing – review & editing. UB: Methodology, Validation, Writing – review & editing, Supervision, Conceptualization, Funding acquisition.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work has been financed by self-generated funds from Udeni B. R. Balasuriya and Mariano Carossino (PG008671).
Acknowledgments
The authors kindly acknowledge the technical staff of the Molecular Biology Laboratory at the Louisiana Animal Disease Diagnostic Laboratory (LSU Diagnostics) for testing specimens initially as routine diagnostic specimens and sharing them with the authors to conduct this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2025.1617147/full#supplementary-material
References
1. Buonavoglia, C, and Martella, V. Canine respiratory viruses. Vet Res. (2007) 38:355–73. doi: 10.1051/vetres:2006058
2. Reagan, KL, and Sykes, JE. Canine infectious respiratory disease. Vet Clin North Am Small Anim Pract. (2020) 50:405–18. doi: 10.1016/j.cvsm.2019.10.009
3. Mitchell, JA, Cardwell, JM, Leach, H, Walker, CA, Le Poder, S, Decaro, N, et al. European surveillance of emerging pathogens associated with canine infectious respiratory disease. Vet Microbiol. (2017) 212:31–8. doi: 10.1016/j.vetmic.2017.10.019
4. Erles, K, Dubovi, EJ, Brooks, HW, and Brownlie, J. Longitudinal study of viruses associated with canine infectious respiratory disease. J Clin Microbiol. (2004) 42:4524–9. doi: 10.1128/JCM.42.10.4524-4529.2004
5. Ditchfield, J, Macpherson, LW, and Zbitnew, A. Association of canine adenovirus (Toronto a 26/61) with an outbreak of Laryngotracheitis (“kennel cough”). Can Vet J. (1962) 3:238–46. Available online at: https://pmc.ncbi.nlm.nih.gov/articles/PMC1585919/
6. Martella, V, Elia, G, and Buonavoglia, C. Canine Distemper Virus. Vet Clin N Am Small Anim Pract. (2008) 38:787–97. doi: 10.1016/j.cvsm.2008.02.007
7. Evermann, JF, Ledbetter, EC, and Maes, RK. Canine reproductive, respiratory, and ocular diseases due to canine herpesvirus. Vet Clin N Am Small Anim Pract. (2011) 41:1097–120. doi: 10.1016/j.cvsm.2011.08.007
8. Wasik, BR, Voorhees, IEH, and Parrish, CR. Canine and feline influenza. Cold Spring Harb Perspect Med. (2021) 11:a038562. doi: 10.1101/cshperspect.a038562
9. Renshaw, R, Laverack, M, Zylich, N, Glaser, A, and Dubovi, E. Genomic analysis of a pneumovirus isolated from dogs with acute respiratory disease. Vet Microbiol. (2011) 150:88–95. doi: 10.1016/j.vetmic.2011.01.013
10. Thieulent, CJ, Carossino, M, Peak, L, Wolfson, W, Li, G, and Balasuriya, UBR. Coding-complete genome sequences of two strains of canine pneumovirus derived from dogs with upper respiratory disease in the United States. Microbiol Resour Announc. (2024) 13:e0105723–3. doi: 10.1128/mra.01057-23
11. Weese, JS, and Stull, J. Respiratory disease outbreak in a veterinary hospital associated with canine parainfluenza virus infection. Can Vet J. (2013) 54:79–82. Available online at: https://pmc.ncbi.nlm.nih.gov/articles/PMC3524821/
12. Mitchell, JA, Brooks, HW, Szladovits, B, Erles, K, Gibbons, R, Shields, S, et al. Tropism and pathological findings associated with canine respiratory coronavirus (CRCoV). Vet Microbiol. (2013) 162:582–94. doi: 10.1016/j.vetmic.2012.11.025
13. De Luca, E, Álvarez-Narváez, S, Baptista, RP, Maboni, G, Blas-Machado, U, and Sanchez, S. Epidemiologic investigation and genetic characterization of canine respiratory coronavirus in the Southeastern United States. J Vet Diagn Invest. (2024) 36:46–55. doi: 10.1177/10406387231213662
14. Schulz, BS, Kurz, S, Weber, K, Balzer, H-J, and Hartmann, K. Detection of respiratory viruses and Bordetella bronchiseptica in dogs with acute respiratory tract infections. Vet J. (2014) 201:365–9. doi: 10.1016/j.tvjl.2014.04.019
15. Chalker, VJ, Owen, WMA, Paterson, C, Barker, E, Brooks, H, Rycroft, AN, et al. Mycoplasmas associated with canine infectious respiratory disease. Microbiology (Reading). (2004) 150:3491–7. doi: 10.1099/mic.0.26848-0
16. Hong, S, and Kim, O. Molecular identification of Mycoplasma cynos from laboratory beagle dogs with respiratory disease. Lab Anim Res. (2012) 28:61–6. doi: 10.5625/lar.2012.28.1.61
17. Priestnall, S, and Erles, K. Streptococcus zooepidemicus: an emerging canine pathogen. Vet J. (2011) 188:142–8. doi: 10.1016/j.tvjl.2010.04.028
18. Chalker, VJ, Brooks, HW, and Brownlie, J. The association of Streptococcus equi subsp. zooepidemicus with canine infectious respiratory disease. Vet Microbiol. (2003) 95:149–56. doi: 10.1016/S0378-1135(03)00155-X
19. El-Attar, LMR, Mitchell, JA, Brooks Brownlie, H, Priestnall, SL, and Brownlie, J. Detection of non-primate hepaciviruses in UK dogs. Virology. (2015) 484:93–102. doi: 10.1016/j.virol.2015.05.005
20. Renshaw, RW, Griffing, J, Weisman, J, Crofton, LM, Laverack, MA, Poston, RP, et al. Characterization of a Vesivirus associated with an outbreak of acute hemorrhagic gastroenteritis in domestic dogs. J Clin Microbiol. (2018) 56:e01951-17. doi: 10.1128/JCM.01951-17
21. Binn, LN, Norby, EA, Marchwicki, RH, Jarman, RG, Keiser, PB, and Hang, J. Canine caliciviruses of four serotypes from military and research dogs recovered in 1963−1978 belong to two phylogenetic clades in the Vesivirus genus. Virol J. (2018) 15:39. doi: 10.1186/s12985-018-0944-4
22. Dankaona, W, Mongkholdej, E, Satthathum, C, Piewbang, C, and Techangamsuwan, S. Epidemiology, genetic diversity, and association of canine circovirus infection in dogs with respiratory disease. Sci Rep. (2022) 12:15445. doi: 10.1038/s41598-022-19815-z
23. Decaro, N, Mari, V, Larocca, V, Losurdo, M, Lanave, G, Lucente, MS, et al. Molecular surveillance of traditional and emerging pathogens associated with canine infectious respiratory disease. Vet Microbiol. (2016) 192:21–5. doi: 10.1016/j.vetmic.2016.06.009
24. Maboni, G, Seguel, M, Lorton, A, Berghaus, R, and Sanchez, S. Canine infectious respiratory disease: new insights into the etiology and epidemiology of associated pathogens. PLoS One. (2019) 14:e0215817. doi: 10.1371/journal.pone.0215817
25. More, GD, Cave, NJ, Biggs, PJ, Acke, E, and Dunowska, M. A molecular survey of canine respiratory viruses in New Zealand. N Z Vet J. (2021) 69:224–33. doi: 10.1080/00480169.2021.1915211
26. Thieulent, CJ, Carossino, M, Peak, L, Strother, K, Wolfson, W, and Balasuriya, UBR. Development and validation of a panel of one-step 4-plex qPCR/RT-qPCR assays for simultaneous detection of SARS-CoV-2 and other pathogens associated with canine in-fectious respiratory disease complex. Viruses. (2023) 15:1881. doi: 10.3390/v15091881
27. Day, MJ, Horzinek, MC, Schultz, RD, and Squires, RA. Wsava guidelines for the vaccination of dogs and cats. J Small Anim Pract. (2016) 57:E1–E45. doi: 10.1111/jsap.2_12431
28. Andrukonis, A, Brown, KM, Hall, NJ, and Protopopova, A. Intake vaccinations reduced signs of canine respiratory disease during an outbreak at an animal shelter. Front Vet Sci. (2021) 8:627580. doi: 10.3389/fvets.2021.627580
29. Erles, K, and Brownlie, J. Investigation into the causes of canine infectious respiratory disease: antibody responses to canine respiratory coronavirus and canine herpesvirus in two kennelled dog populations. Arch Virol. (2005) 150:1493–504. doi: 10.1007/s00705-005-0533-x
30. Okonkowski, LK, Szlosek, D, Ottney, J, Coyne, M, and Carey, SA. Asymptomatic carriage of canine infectious respiratory disease complex pathogens among healthy dogs. J Small Anim Pract. (2021) 62:662–8. doi: 10.1111/jsap.13378
31. Wang, R, Zhang, W, Ye, R, Pan, Z, Li, G, and Su, S. One-step multiplex TaqMan probe-based method for real-time PCR detection of four canine diarrhea viruses. Mol Cell Probes. (2020) 53:101618. doi: 10.1016/j.mcp.2020.101618
32. Decaro, N, Amorisco, F, Desario, C, Lorusso, E, Camero, M, Bellacicco, AL, et al. Development and validation of a real-time PCR assay for specific and sensitive detection of canid herpesvirus 1. J Virol Methods. (2010) 169:176–80. doi: 10.1016/j.jviromet.2010.07.021
33. Tizolova, A, Brun, D, Guiso, N, and Guillot, S. Development of real-time PCR assay for differential detection of Bordetella bronchiseptica and Bordetella parapertussis. Diagn Microbiol Infect Dis. (2014) 78:347–51. doi: 10.1016/j.diagmicrobio.2013.12.020
34. Dong, J, Tsui, WNT, Leng, X, Fu, J, Lohman, M, Anderson, J, et al. Development of a three-panel multiplex real-time PCR assay for simultaneous detection of nine canine respiratory pathogens. J Microbiol Methods. (2022) 199:106528. doi: 10.1016/j.mimet.2022.106528
35. Nafea, AM, Wang, Y, Wang, D, Salama, AM, Aziz, MA, Xu, S, et al. Application of next-generation sequencing to identify different pathogens. Front Microbiol. (2024) 14:1329330. doi: 10.3389/fmicb.2023.1329330
36. American Veterinary Medical Association. (2024). Questions remain as canine respiratory disease cases fall. Available online at: https://www.avma.org/news/questions-remain-canine-respiratory-disease-cases-fall (accessed August 29, 2024).
37. Ramspacher, A. Q&a: What to know about a mysterious dog illness sweeping the country. (2023). Available online at: https://news.virginia.edu/content/qa-what-know-about-mysterious-dog-illness-sweeping-country (accessed August 29, 2024).
38. Phillips, A. Dog respiratory illness maps show its spread to 16 states. Newsweek (2023). Available online at: https://www.newsweek.com/dog-respiratory-illness-map-spread-16-1852084 (Accessed August 29, 2024).
39. Baisas, L. An unknown respiratory illness is sickening dogs in the US, but ‘don’t panic’. (2023). Available online at: https://www.popsci.com/health/respiratory-illness-dogs/?fbclid=IwAR3Kwqie0faYWYrgK-2Oc6ENkqaQkM_LEiglFkA7KCWRMlyMsYDwO9krFDo (Accessed August 29, 2024).
40. Tanaka, R. (2023). WesternU College of Veterinary Medicine statement on the Atypical Canine Infectious Respiratory Disease Outbreak (aCIRD). WesternU News. Available online at: https://news.westernu.edu/westernu-college-of-veterinary-medicine-statement-on-the-atypical-canine-infectious-respiratory-disease-outbreak-acird/ (Accessed July 24, 2025).
41. Caserta, LC, Frye, EA, Butt, SL, Laverack, M, Nooruzzaman, M, Covaleda, LM, et al. Spillover of highly pathogenic avian influenza H5N1 virus to dairy cattle. Nature. (2024) 634:669–676. doi: 10.1038/s41586-024-07849-4
42. Van Borm, S, Fu, Q, Winand, R, Vanneste, K, Hakhverdyan, M, Höper, D, et al. Evaluation of a commercial exogenous internal process control for diagnostic RNA virus metagenomics from different animal clinical samples. J Virol Methods. (2020) 283:113916. doi: 10.1016/j.jviromet.2020.113916
43. Chrzastek, K, Lee, D-H, Smith, D, Sharma, P, Suarez, DL, Pantin-Jackwood, M, et al. Use of sequence-independent, single-primer-amplification (SISPA) for rapid detection, identification, and characterization of avian RNA viruses. Virology. (2017) 509:159–66. doi: 10.1016/j.virol.2017.06.019
44. De Coster, W, and Rademakers, R. NanoPack2: population-scale evaluation of long-read sequencing data. Bioinformatics. (2023) 39:btad311. doi: 10.1093/bioinformatics/btad311
45. De Coster, W, D’Hert, S, Schultz, DT, Cruts, M, and Van Broeckhoven, C. NanoPack: visualizing and processing long-read sequencing data. Bioinformatics. (2018) 34:2666–9. doi: 10.1093/bioinformatics/bty149
46. Wood, DE, Lu, J, and Langmead, B. Improved metagenomic analysis with kraken 2. Genome Biol. (2019) 20:257. doi: 10.1186/s13059-019-1891-0
47. Lu, J, Rincon, N, Wood, DE, Breitwieser, FP, Pockrandt, C, Langmead, B, et al. Metagenome analysis using the kraken software suite. Nat Protoc. (2022) 17:2815–39. doi: 10.1038/s41596-022-00738-y
48. Matsuu, A, Yabuki, M, Aoki, E, and Iwahana, M. Molecular detection of canine respiratory pathogens between 2017 and 2018 in Japan. J Vet Med Sci. (2020) 82:690–4. doi: 10.1292/jvms.20-0017
49. Schaffer, FL, Soergel, ME, Black, JW, Skilling, DE, Smith, AW, and Cubitt, WD. Characterization of a new calicivirus isolated from feces of a dog. Arch Virol. (1985) 84:181–95. doi: 10.1007/BF01378971
50. Kapoor, A, Dubovi, EJ, Henriquez-Rivera, JA, and Lipkin, WI. Complete genome sequence of the first canine circovirus. J Virol. (2012) 86:7018–8. doi: 10.1128/JVI.00791-12
51. Gomez-Betancur, D, Vargas-Bermudez, DS, Giraldo-Ramírez, S, Jaime, J, and Ruiz-Saenz, J. Canine circovirus: an emerging or an endemic undiagnosed enteritis virus? Front Vet Sci. (2023) 10:1150636. doi: 10.3389/fvets.2023.1150636
52. Thaiwong, T, Wise, AG, Maes, RK, Mullaney, T, and Kiupel, M. Canine circovirus 1 (CaCV-1) and canine parvovirus 2 (CPV-2): recurrent dual infections in a Papillon breeding Colony. Vet Pathol. (2016) 53:1204–9. doi: 10.1177/0300985816646430
53. Abbas, A, Taylor, LJ, Collman, RG, and Bushman, FDICTV Report Consortium. ICTV virus taxonomy profile: redondoviridae: this article is part of the ICTV virus taxonomy profiles collection. J Gen Virol. (2021) 102:jgv001526. doi: 10.1099/jgv.0.001526
54. Lázaro-Perona, F, Dahdouh, E, Román-Soto, S, Jiménez-Rodríguez, S, Rodríguez-Antolín, C, De La Calle, F, et al. Metagenomic detection of two Vientoviruses in a human sputum sample. Viruses. (2020) 12:327. doi: 10.3390/v12030327
55. Abbas, AA, Taylor, LJ, Dothard, MI, Leiby, JS, Fitzgerald, AS, Khatib, LA, et al. Redondoviridae, a family of small, circular DNA viruses of the human Oro-respiratory tract associated with periodontitis and critical illness. Cell Host Microbe. (2019) 25:719–729.e4. doi: 10.1016/j.chom.2019.04.001
56. Altan, E, Seguin, MA, Leutenegger, CM, Phan, TG, Deng, X, and Delwart, E. Nasal virome of dogs with respiratory infection signs include novel taupapillomaviruses. Virus Genes. (2019) 55:191–7. doi: 10.1007/s11262-019-01634-6
57. Tress, B, Dorn, ES, Suchodolski, JS, Nisar, T, Ravindran, P, Weber, K, et al. Bacterial microbiome of the nose of healthy dogs and dogs with nasal disease. PLoS One. (2017) 12:e0176736. doi: 10.1371/journal.pone.0176736
58. Djikeng, A, Halpin, R, Kuzmickas, R, DePasse, J, Feldblyum, J, Sengamalay, N, et al. Viral genome sequencing by random priming methods. BMC Genomics. (2008) 9:5. doi: 10.1186/1471-2164-9-5
59. Wright, MS, Stockwell, TB, Beck, E, Busam, DA, Bajaksouzian, S, Jacobs, MR, et al. SISPA-Seq for rapid whole genome surveys of bacterial isolates. Infect Genet Evol. (2015) 32:191–8. doi: 10.1016/j.meegid.2015.03.018
60. Soria-Villalba, A, Pesantes, N, Jiménez-Hernández, N, Pons, J, Moya, A, and Pérez-Brocal, V. Comparison of experimental methodologies based on bulk-metagenome and virus-like particle enrichment: pros and cons for representativeness and reproducibility in the study of the Fecal human Virome. Microorganisms. (2024) 12:162. doi: 10.3390/microorganisms12010162
Keywords: canine respiratory pathogens, canine infectious respiratory disease complex, CIRDC, qPCR, RT-qPCR, next-generation sequencing, vientovirus
Citation: Thieulent CJ, Laverack M, Carossino M, Cronk B, Caserta LC, Diel DG and Balasuriya UBR (2025) Detection of pathogens in dogs with respiratory disease during winter 2023–2024 using multiplex qPCR/RT-qPCR assays and next-generation sequencing. Front. Vet. Sci. 12:1617147. doi: 10.3389/fvets.2025.1617147
Edited by:
Jesús Hernández, National Council of Science and Technology (CONACYT), MexicoReviewed by:
Kanisht Batra, Lala Lajpat Rai University of Veterinary and Animal Sciences, IndiaBrianna Stenger, North Dakota State University, United States
Anthony Loor, Universidad de las Américas, Ecuador
Copyright © 2025 Thieulent, Laverack, Carossino, Cronk, Caserta, Diel and Balasuriya. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Udeni B. R. Balasuriya, YmFsYXN1cml5YTFAbHN1LmVkdQ==
†ORCID: Côme J. Thieulent, orcid.org/0000-0001-7402-5989
Mariano Carossino, orcid.org/0000-0003-3864-5915
Udeni B. R. Balasuriya, orcid.org/0000-0003-0609-678X