 Jörg Schaller
Jörg Schaller1 Introduction
The origin of life on Earth is an unsolved mystery that has engaged many of the best minds in the Earth and biological sciences. Since the Miller-Urey experiment (Miller, 1953) showed the formation of organic molecules required for life, scientists have tried to simulate the molecular precursors of life and to explain how they assembled into replicating forms. There are several competing hypotheses about how this may have occurred (Ferris, 1999). All these hypotheses suggest conditions or material suitable as template to allow formation of rudimentary DNA or RNA, as well as vesicles, membranes and protocells. Here I focus on the clay-template hypothesis proposed by Cairns-Smith in 1966 (Cairns-Smith, 1966), which suggests that the polymerized crystal structures of (alumino)silicates that form clay minerals provide ideal surfaces on which large organic molecules could have been assembled. I argue that disproportionation of amorphous silicon monoxide under hydrogen-rich conditions may have allowed abiogenesis.
2 Mineral composition during origin of life
Different silicate minerals (e.g., achondrite, olivine, feldspar, pyroxenes) (McCord and Gaffey, 1974) or crystalline clay minerals such as smectites (Baird et al., 1977) exist on Earth as well as on Mars, Moon, meteorites, asteroids and comets. The initial mineral evolution on Earth was driven by volcanism, degassing, fractional crystallization, and associated large-scale fluid-rock interactions (Hazen et al., 2008). Deploying the idea of the clay-template hypothesis (Kloprogge and Hartman, 2022), one could assume that prebiotic molecules assembled (i) in interstellar space, (ii) after arriving on Earth, or (iii) during water-mediated weathering of magmatic minerals on the early Earth which results in the formation of secondary minerals. Despite the attractiveness of the clay-template hypothesis, no research has been able to explain the process needed to assemble the precursors of life on crystalline clays.
Today, amorphous clay minerals in soils on Earth show a larger chemical activity, and play a more important role in sorbing organic and inorganic compounds than crystalline clays (Cairns-Smith, 1982), by ligand exchange, cation bridging, van der Waals force, and hydrogen bonding (Shang, 2023). Some of the amorphous compounds present on today’s Earth have been identified in Martian soils as well. Such amorphous compounds offer a loosely coordinated 3-D structure which provides a more flexible template, compared to the 2-D structure of crystalline clays, allowing for more complex polymerization of organic molecules that can act as pre-cursors of life.
It is well known that silica gels, a form of amorphous silica connected to water, exhibit ideal properties for binding DNA, which is the reason why this material is commonly used for DNA extraction (Vogelstein and Gillespie, 1979; Chase and Hills, 1991). Such amorphous silicas are prone to adsorb amino acids and potentially force the formation of nucleic acids, the backbone of RNA and DNA structures. Amorphous silica polymers enhance amino acid polymerization and other prebiotic organic reactions like vesicle, membrane, and protocell formation (Cho et al., 2024; Samrout et al., 2024; Jenewein et al., 2025) that would have been necessary to the genesis of replicating biomolecules. Hence, amorphous silica may serve as a catalyst for the origin of life.
Chemical and physical conditions on early Earth were likely far different from the present and may have favored stability of amorphous silica phases. For example, hydrogen may have been present in the early Earth crust as meteorites were found to contain large share of hydrogen (Barrett et al., 2025) and due to hydrogen formation by low temperature water-rock reactions (Mayhew et al., 2013), limiting oxidation. Early Earth may have had a much higher hydrogen content compared to present conditions as light elements like hydrogen are lost from the Earth due to meteor impact (Biersteker and Schlichting, 2019) and if not bound to other elements, could escape to outer space because Earth gravitation is not able to prevent losses (Hunten and Donahue, 1976). It can be assumed that hydrogen loss would have favored a shift toward more oxidizing conditions even before the strong shift associated with photosynthesis. Another example for the different conditions of early Earth compared to present Earth is the presence of silicon monoxide (SiO) in interstellar space (Gibb et al., 2007), which is the most common oxide of silicon in the universe (Cherchneff, 2013). Hence, such material may have also been present in early Earth during the time of origin of life and the presence of hydrogen may have prevented immediate oxidation to amorphous SiO2.
3 Importance of disproportion of amorphous silicon monoxide
Disproportion of amorphous silicon monoxide to elemental silicon and silicon dioxide (Hirata et al., 2016) would have been favored by strong reducing conditions on early Earth, potentially offering conditions to support abiogenesis. The lower the temperature, the slower the disproportionation process (Mamiya et al., 2001). The disproportion of amorphous silicon monoxide into Si and SiO2 will result in distances between elemental Si and a neighboring SiO2 unit at molecular level of about ∼0.4 nm (Hirata et al., 2016) which is comparable in scale to the distance of nucleotides of DNA (∼0.34 nm) (Alberts et al., 2014) (Figure 1).
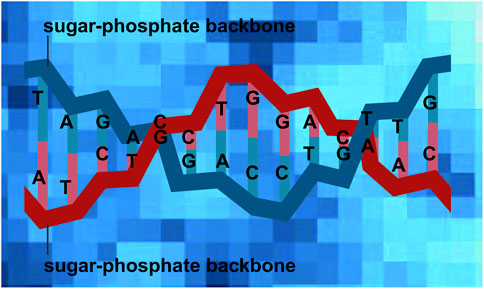
Figure 1. Small-scale pattern (∼0.4 nm) of elemental Si and SiO2 from SiO disproportionation, adapted from Hirata et al. (2016), which is on the same scale as the distance between nucleotides of DNA (∼0.34 nm). One should note that the number for SiO disproportionation, ∼0.4 nm, is based on the resolution limit of the analytical method used and may in fact be lower. Different blue shades of squares in the background indicate different Si species from amorphous elemental Si (light blue) to amorphous SiO2 (dark blue). The red and blue lines represent DNA backbones.
At a scale of about 0.4 nm the formation of a protective SiO2 layer of a few atoms thickness, preventing further oxidation, is negligible. Because of the missing protections from a SiO2-layer the single Si atoms distributed at the surfaces of the amorphous solids may be oxidized to SiO2 even under low temperature conditions (Greenwood and Earnshaw, 2012). However, under hydrogen rich conditions this direct oxidation of SiO to SiO2 may be restricted. A potential process may have started by the absorption of amino acids already in meteorites (Cronin and Pizzarello, 1983). The amino acids adsorbed to the amorphous material (in the immediate vicinity of a Si atom) may cause oxidation of the Si to SiO2, as the formation of SiO2 is thermodynamically favored. During oxidation of Si the amino acid groups will be transformed into amines or N-containing heterocycles, the latter offering a base structure for the formation of nucleic acid. Note: variations in the spatial arrangement of the Si atoms (or Si-Si dimers) and the SiO2 units across the surface of the amorphous material may allow formation of an innumerable number of different amines or heterocyclic acid structures (Hohl et al., 2003; Hirata et al., 2016).
Life seems to have originated only once in Earth history. There are no intermediates or leftovers from the ancient origin of life (like rudimentary DNA or RNA trapped in minerals) found in current Earth environments. Consequently, the process must have occurred under conditions different from present Earth conditions. I suggest that amorphous silicon monoxide was both available (SiO rich meteorite input) and chemically favored because of the presence of free hydrogen. If any amorphous phase formed from SiO was involved in the origin of life, no leftovers would occur. This is because oxidation of Si to SiO2 and restructuring of the Si template due to structural rearrangement by crystallization of the amorphous material to crystalline minerals would have destroyed all organic residues. Additionally, if amorphous silicon monoxide was involved in the origin of life, such abiogenesis could have happened during a short period in Earth history, as the amorphous silicon monoxide will have been disproportionated and oxidized to silicon dioxide eventually. With amorphous silicon dioxide (eventually forming after SiO disproportionation) supporting vesicle, membrane and protocell formation (Cho et al., 2024; Jenewein et al., 2025), necessary environmental conditions to support the survival of replicating biomolecules might have ensured Abiogenesis.
Author contributions
JS: Visualization, Writing – original draft, Writing – review and editing, Conceptualization.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Acknowledgments
I like to thank Oliver Chadwick (UC Santa Barbara), Ruth Ellerbrock (ZALF) and the reviewer for their comprehensive comments which helped a lot to improve this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alberts, B., Heald, R., Johnson, A., Morgan, D., Raff, M., Roberts, K., et al. (2014). Molecular biology of the cell garland science.
Baird, A., Castro, A., Clark, B., Toulmin, P., Rose, H., Keil, K., et al. (1977). The Viking X ray fluorescence experiment: sampling strategies and laboratory simulations. J. Geophys. Res. 82, 4595–4624. doi:10.1029/js082i028p04595
Barrett, T. J., Bryson, J. F., and Geraki, K. (2025). The source of hydrogen in earth's building blocks. Icarus 436, 116588. doi:10.1016/j.icarus.2025.116588
Biersteker, J. B., and Schlichting, H. E. (2019). Atmospheric mass-loss due to giant impacts: the importance of the thermal component for hydrogen–helium envelopes. Mon. Notices R. Astronomical Soc. 485, 4454–4463. doi:10.1093/mnras/stz738
Cairns-Smith, A. G. (1966). The origin of life and the nature of the primitive gene. J. Theor. Biol. 10, 53–88. doi:10.1016/0022-5193(66)90178-0
Cairns-Smith, A. G. (1982). Genetic takeover and the mineral origins of life. New York: Cambridge University Press.
Chase, M. W., and Hills, H. H. (1991). Silica gel: an ideal material for field preservation of leaf samples for DNA studies. Taxon 40, 215–220. doi:10.2307/1222975
Cherchneff, I. (2013). The chemistry of dust formation in red supergiants. Eur. Astron. Soc. Publ. Ser. 60, 175–184. doi:10.1051/eas/1360020
Cho, C. J., An, T., Lai, Y.-C., Vázquez-Salazar, A., Fracassi, A., Brea, R. J., et al. (2024). Protocells by spontaneous reaction of cysteine with short-chain thioesters. Nat. Chem. 17, 148–155. doi:10.1038/s41557-024-01666-y
Cronin, J. R., and Pizzarello, S. (1983). Amino acids in meteorites. Adv. space Res. 3, 5–18. doi:10.1016/0273-1177(83)90036-4
Ferris, J. P. (1999). Prebiotic synthesis on minerals: bridging the prebiotic and RNA worlds. Biol. Bull. 196, 311–314. doi:10.2307/1542957
Gibb, A. G., Davis, C. J., and Moore, T. J. T. (2007). A survey of SiO 5 → 4 emission towards outflows from massive young stellar objects. Mon. Notices R. Astronomical Soc. 382, 1213–1224. doi:10.1111/j.1365-2966.2007.12455.x
Hazen, R. M., Papineau, D., Bleeker, W., Downs, R. T., Ferry, J. M., McCoy, T. J., et al. (2008). Mineral evolution. Am. Min. 93, 1693–1720. doi:10.2138/am.2008.2955
Hirata, A., Kohara, S., Asada, T., Arao, M., Yogi, C., Imai, H., et al. (2016). Atomic-scale disproportionation in amorphous silicon monoxide. Nat. Commun. 7, 11591. doi:10.1038/ncomms11591
Hohl, A., Wieder, T., Van Aken, P., Weirich, T., Denninger, G., Vidal, M., et al. (2003). An interface clusters mixture model for the structure of amorphous silicon monoxide (SiO). J. Non-Crystalline Solids 320, 255–280. doi:10.1016/s0022-3093(03)00031-0
Hunten, D., and Donahue, T. M. (1976). “Hydrogen loss from the terrestrial planets,”4. Palo Alto, Calif: Annual Reviews, Inc., 265–292. doi:10.1146/annurev.ea.04.050176.001405
Jenewein, C., Maíz-Sicilia, A., Rull, F., González-Souto, L., and García-Ruiz, J. M. (2025). Concomitant formation of protocells and prebiotic compounds under a plausible early Earth atmosphere, 122. Washington, D.C, United States: Proceedings of the National Academy of Sciences.e2413816122
Kloprogge, J. T., and Hartman, H. (2022). Clays and the origin of life: the experiments. Life 12, 259. doi:10.3390/life12020259
Mamiya, M., Takei, H., Kikuchi, M., and Uyeda, C. (2001). Preparation of fine silicon particles from amorphous silicon monoxide by the disproportionation reaction. J. Cryst. Growth 229, 457–461. doi:10.1016/S0022-0248(01)01202-7
Mayhew, L. E., Ellison, E., McCollom, T., Trainor, T. P., and Templeton, A. S. (2013). Hydrogen generation from low-temperature water–rock reactions. Nat. Geosci. 6, 478–484. doi:10.1038/ngeo1825
McCord, T. B., and Gaffey, M. J. (1974). Asteroids: surface composition from reflection spectroscopy. Science 186, 352–355. doi:10.1126/science.186.4161.352
Miller, S. L. (1953). A production of amino acids under possible primitive earth conditions. Science 117, 528–529. doi:10.1126/science.117.3046.528
Samrout, O. E., Berlier, G., and Lambert, J. F. (2024). Amino acid polymerization on silica surfaces. ChemPlusChem 89, e202300642. doi:10.1002/cplu.202300642
Shang, H. (2023). Mineral evolution facilitated Earth’s oxidation. Commun. Earth and Environ. 4, 213. doi:10.1038/s43247-023-00824-3
Keywords: abiogenesis, amorphous silica, disproportionation, origin of life, clay-hypothesis
Citation: Schaller J (2025) Potential misassumptions on the clay hypothesis in origin of life: a geochemical perspective. Front. Astron. Space Sci. 12:1678046. doi: 10.3389/fspas.2025.1678046
Received: 01 August 2025; Accepted: 29 August 2025;
Published: 09 September 2025.
Edited by:
Raffaele Saladino, University of Tuscia, ItalyReviewed by:
Haitao Shang, The University of Texas at El Paso, United StatesCopyright © 2025 Schaller. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jörg Schaller, Sm9lcmcuU2NoYWxsZXJAemFsZi5kZQ==