 Emory G. Barrett
Emory G. Barrett Richard A. Lutz
Richard A. Lutz- Department of Marine and Coastal Sciences, Rutgers University, New Brunswick, NJ, United States
The search for extraterrestrial life has historically focused on photosynthetic organisms but following the discovery of deep-sea hydrothermal vents on Earth and the variety of microbes with unconventional metabolic pathways that inhabit them, astrobiology has expanded to encompass the search for extraterrestrial chemoautotrophs. Although bombarded by Jovian radiation and encased in a thick shell of ice, Jupiter’s moon Europa could very well be a sanctuary for extraterrestrial microbial life. It is widely accepted that underneath the ice lies a deep, salty ocean, and, despite possibly anoxic conditions, high pressures, and lack of sunlight, many physical and chemical properties of Europa are analogs of extreme environments on Earth, such as hydrothermal vents or subterranean radiogenic ecosystems. By drawing comparisons between Europa and Earth, it can be seen how processes such as serpentization, water radiolysis, and volcanic activity induced by tidal heating or Jovian radiation could very well be providing energy to chemoautotrophs inhabiting various habitats on the ocean floor. Understanding the possibilities of chemoautotrophic life on Europa can be extended to other planetary bodies, such as Mars and Enceladus, allowing for their consideration under a new perspective. Future missions such as the Europa Clipper, scheduled to arrive in 2030, will provide further clarity on the moon’s habitability and potentially bring science closer to the discovery of extraterrestrial biota.
1 Introduction
Extraterrestrial life has long been of interest to humanity, both scientists and the public alike (Nascimento-Dias and Martinez-Frias, 2023). Rover and satellite missions to planets and moons in our solar system have so far been unsuccessful at documenting extraterrestrial life. Still, numerous discoveries of planetary objects with the potential to support life, either currently or in the past, have been made (Kasting et al., 2014). Searches for extraterrestrial life usually focus on detecting patterns on other planets that parallel life on Earth, such as searching for liquid water, determining the location of a “habitable zone,” or identifying biosignatures–compounds produced by or useful to familiar forms of life, revolving around photosynthesis as a means of primary production (Kasting et al., 2014; Seeburger et al., 2023). Of course, the search for extraterrestrial biosignatures is limited to what one expects to find. As new metabolic pathways are discovered on Earth, less obvious signs of life may begin to present themselves on other planetary bodies. An example of this is the search for hydrothermal activity on planetary objects (Vance et al., 2007). Since the discovery of hydrothermal vents in 1977, science has expanded its understanding of life to encompass unconventional ideas of what forms of extraterrestrial life could be possible (Hessler and Smithey, 1983). Searching for geologic and chemical signatures of hydrothermal vent activity can allow science to expand its horizons past the limitations of photosynthesis-powered ecosystems. This is because the presence of hydrothermal activity or natural unstable isotopes on planetary objects with liquid water could suggest extraterrestrial ecosystems powered by pathways alternative to photosynthesis, such as chemosynthesis, which is commonly observed on our own planet, even in the most unfavorable conditions (Chyba and Phillips, 2001). While searching for analogs to life on Earth is a straightforward and classical method of researching exobiology, it is imperative to maintain an agnostic perspective on extraterrestrial life. The profound diversity of metabolisms seen on our planet, including those discovered at hydrothermal vents, reminds us that while entertaining the thought of extraterrestrial life, we must leave room for the potential for unfamiliar metabolic strategies (Nealson and Popa, 2005). If no room is left for the consideration of completely unfamiliar life strategies, the scope of potential discovery will be stunted.
Much interest in astronomy has been directed at the planet Jupiter’s system since Galileo’s discovery of the four larger Jovian moons in 1,610 (Schmidt, 2020). Each of the four Galilean moons has its distinct characteristics, with the third moon from the planet, Europa, being a prime example of an ice moon and potentially an “ocean world” - a planetary body with an ocean covering the entirety of its surface. Blasted by radiation from Jupiter and millions of miles from the sun’s warmth, the dry, icy surface of Europa seems to be completely uninhabitable (Altair et al., 2018; Weber et al., 2023). As hostile as the surface is, evidence shows that underneath the radiation and the 12–17 km of ice, there is likely to be a deep, salty ocean of liquid water and a rocky mantle, all surrounding a metallic core (Chyba and Phillips, 2001; Walker and Rhoden, 2022). Compelling evidence for the presence of this subsurface ocean of liquid water on Europa has allowed for much scientific discussion surrounding possibilities of extraterrestrial life, sparking research into the possibility of a variety of lifeforms on Europa and other planetary bodies like it (Howell and Pappalardo, 2020). From hydrothermal vent ecosystems to radiation-driven pathways to photosynthesis in near-surface brine pockets, there is a mountain of research regarding the astrobiology of Europa, all spinning on a singular linchpin, the presence of this cryptic subsurface ocean (Chyba and Phillips, 2001).
2 Evidence of liquid water on Europa
2.1 Europan topography
While liquid water has yet to be definitively confirmed to exist on Europa, there lies a laundry list of compelling evidence sufficient to treat the concept as near fact. One line of evidence for the presence of water under the surface of Europa is the moon’s topography. The ice surface of Europa is heavily textured, with clusters of pits, domes, and spots, called “chaos terrains.” These chaos terrains appear to be where ice may have broken while floating on a liquid subsurface, similar to icebergs (Kivelson et al., 2000). It is theorized that these chaos terrains suggest thermal structuring of the ice and, therefore, a liquid ocean underneath the moon’s icy exterior (Van Dover, 2000). Two of the largest (greater than 50 km across) chaos terrains are called Thera Macula and Thrace Macula, and it is highly likely that their formation is associated with liquid water and brines (Figure 1). The ice of these sunken chaos terrains is composed of jagged, chaotic ice, and is younger than ice elsewhere on the surface. The telltale reddish color spilling from the borders is characteristic of high salt content, formed as liquid water carrying the salts freezes upon reaching the surface of the ice, expelling highly concentrated brine that leeches into the surrounding terrain (Schmidt, 2020). This could be caused by the upward motion of subsurface water seeping through cracks in a thin ice crust and then refreezing at the surface, or through the formation and upward movement of “melt pockets” of relatively warm ice within a thicker ice crust (Figure 2) (Chyba and Phillips, 2001; Schmidt, 2020).
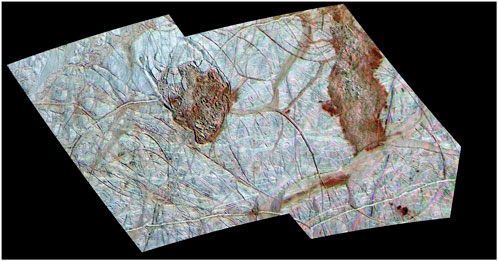
Figure 1. Chaos terrains on the surface of Europa. Thera Macula (left) and Thrace Macula (right) show jagged terrain and a leeching reddish color, associated with salt content. Credit: NASA/JPL/University of Arizona PIA02099.
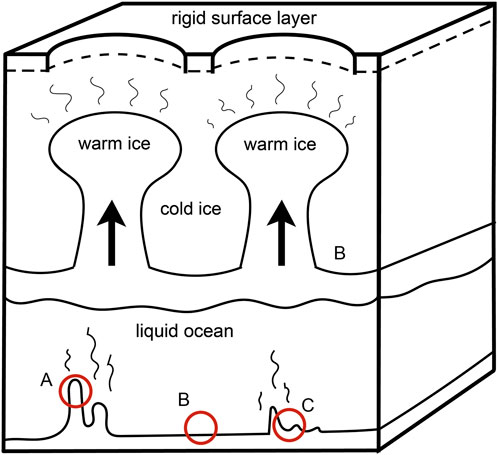
Figure 2. Diapiric rising of warm ice, showing thermal structuring of Europa’s ice layer. A popular theory of chaos terrain formation. Below the ice, heat rises from theorized hydrothermal structures. Potential habitats for microbial life are circled. (A) Hydrothermal vent, (B) water radiolysis site, (C) serpentization site. Modified from Pappalardo et al. (1998).
2.2 Magnetic field
Especially compelling lines of evidence for the presence of a liquid water ocean beneath the ice crust of Europa are the magnetic field results collected when the Galileo spacecraft passed near the moon (Chyba and Phillips, 2001). Jupiter’s magnetosphere is the strongest of the planets in our solar system, and its interactions with the Jovian moons can uncover some of their interesting conductive qualities (Schmidt, 2020). Europa is heavily influenced by Jupiter’s magnetic field, but the Galileo spacecraft was able to pick up fluctuations in the moon’s magnetic field that were unrelated to fluctuations in Jupiter’s magnetosphere. This showed that the moon had an induced magnetic dipole, produced by what is presumed to be a spherical layer with sufficiently high conductivity (Kivelson et al., 2000). In other words, the conductor responsible for the induced magnetic field on Europa is similar to the molten iron layer of Earth, which is responsible for its magnetic field. On a small moon covered in ice, such as Europa, this phenomenon could be easily and simply explained by the presence of a subsurface salty ocean (Chyba and Phillips, 2001). Both lines of evidence are quite compelling and allow us confidence in considering what lifeforms may be present on Europa, as we understand that the most fundamental ingredient for life, water, is likely present on the moon in great volumes.
2.3 Heating
The presence of a subsurface ocean on Europa is only possible with sufficient heating, which could be obtained by the moon through two main pathways. The first is radiogenic heating from within the moon’s core, and the second is through tidal heating caused by gravity (Chyba and Phillips, 2001). Radiogenic heating is simpler and more similar to the geothermal heating we experience on Earth (Van Dover, 2000). While this mode of heating is what allows for geothermal activity on Earth, it is unlikely that a moon as small and uninsulated as Europa has sufficient radiogenic heating leftover from its formation to maintain a liquid ocean on its own (Schmidt, 2020). Tidal heating is more complicated, and given Europa’s role as a Jovian moon, it is a much more significant contributor to any geothermal heating and activity (Reynolds et al., 1987). To understand tidal heating on Europa, we can first understand tidal heating on a more extreme example, Europa’s sister moon Io.
Io is the closest to Jupiter of the Galilean moons, and therefore experiences extreme radiation and is under significant influence of the Jovian magnetosphere, expressed in its role as the most volcanically active body in our solar system (Schmidt, 2020). Due to its synchronous orbit around Jupiter and resonant state with Europa and Ganymede, Io experiences intense tidal heating. Tidal heating of a satellite occurs when an external time-varying force of gravity is exerted upon the satellite, causing it to deform, heating it with frictional energy (Matsuyama et al., 2022). The Earth’s moon also experiences tidal heating while orbiting the Earth, but the tidal heating experienced by it is much less significant, as the Earth is much smaller than Jupiter and the presence of a malleable liquid layer (such as Io’s magma oceans) allows for the most significant heating (Matsuyama et al., 2022). When considering the severity of tidal flexion experienced by Io due to its uneven elliptical orbit and the sheer magnitude of the planet Jupiter, it is clear that the surface of Io would be a landscape of magma and volcanism, which is shockingly different than the icy terrain of its sister, Europa (Matsuyama et al., 2022). This understanding of tidal heating’s effects on Io’s geological activity is critical to understanding how Europa can maintain a liquid ocean underneath its ice crust, as Io’s condition is highly suggestive of similar volcanic activity on Europa itself. Tidal heating has been calculated to account for an estimated 86% of Europa’s internal heating and can likely support a liquid subsurface ocean (Chyba and Phillips, 2001; Reynolds et al., 1987).
3 The Europan ocean
Europa’s ocean is more like Earth’s ocean than other ocean worlds. Its ocean is estimated to be about 100,000 m deep, which is extremely deep compared to the average depth of 3,682 m across oceans found on Earth (Charette and Smith, 2010; Naganuma and Uematsu, 1998). This staggering depth raises questions about the hydrostatic pressure at Europa’s seafloor, with such extreme pressures potentially posing challenges for the existence of life as we know it. Largely depending on seawater chemistry, the pressure at Europa’s seafloor is estimated to be between 130 and 260 MPa, which is comparable to the pressure of an Earthlike ocean at 13–26 km (Naganuma and Uematsu, 1998). Life on Earth is exposed to no more than 150 MPa naturally, with laboratory studies showing bacterial survival in laboratory experiments up to 1,000–2,000 MPa (Hazael et al., 2016). If familiar Earth life can survive pressures as extreme as these, it is not out of the question to speculate that an extraterrestrial form of life could withstand similar. While deeper than Earth’s oceans, Europa’s sole ocean is far shallower than the oceans on some other ocean worlds, such as Ganymede, with Europa having a rock-to-water ratio much more similar to that of Earth, made of almost 90% rock (Vance et al., 2007). Another critical similarity to Earth and a significant difference from ocean worlds like Ganymede is that Europa’s ocean comes into direct contact with the rocky seafloor below (Schmidt, 2020). This detail is extremely important, as the contact between the water and rock can allow for the potential of many geochemical processes similar to those encountered on Earth’s seafloors that allow for processes which can support chemosynthetic communities in hydrothermal vent ecosystems. If these processes are possible on Europa, they could very well support a diverse variety of chemoautotrophic microorganisms on Europa’s seafloor as they do on Earth (Chyba and Phillips, 2001; Reynolds et al., 1987; Schmidt, 2020; Vance et al., 2007).
3.1 Volcanic activity
Following the discovery of hydrothermal vents on Earth in 1977 (Hessler and Smithey, 1983), astrobiologists have shifted their focus from the search for extraterrestrial photosynthetic organisms to the search for extraterrestrial chemoautotrophs, focusing on potential energy sources such as hydrogen gas or on biomolecules (Van Dover, 2000; Weber et al., 2023). While chemoautotrophs can be found in a variety of environments on Earth from insects’ digestive tracts to deep gold mines to anoxic muds, the geological parallels that can be drawn between hydrothermal vents on Earth and those on other planetary bodies make for these deep-sea chemosynthetic ecosystems a prime example that can easily be applied to the search for life on harsh or hostile ocean worlds like Europa (Chivian et al., 2008; Huber et al., 1982; Van Dover, 2000). Due to its tidal heating and passive cooling of radiogenic heat, Europa is thought to potentially experience hydrothermal activity, so long as this energy is sufficient to act upon the rocky portion of the moon, pulling and cracking at the rocky seafloor and allowing for cold salt water to seep into the moon’s interior (Schmidt, 2020; Van Dover, 2000; Walker and Rhoden, 2022; Weber et al., 2023). If sufficient volcanic activity exists on Europa, similar to Io, this could mean ecosystems powered by anaerobic reactions such as sulfate reduction, similar to those at many Earth vents, might exist at depth on Europa (Matsuyama et al., 2022; Weber et al., 2023; Van Dover, 2000). Many anaerobic species at hydrothermal vents on Earth are able to withstand extreme conditions, including a strain of the hyperthermophilic archaea (Methanopyrus kandleri) isolated from the Central Indian Ridge, able to grow at temperatures up to 122 °C under 40 MPa pressure (Clarke, 2014). Another example is the anaerobic, chemoautotrophic, and hyperthermophilic bacterium Phorcysia thermohydrogeniphila that reduces sulfide while residing within polymetallic sulfide chimneys of hydrothermal vents, living comfortably in temperatures up to 80 °C (Pérez-Rodríguez et al., 2012). For something similar to inhabit the oceans of Europa, it would require the presence of sulfur compounds in the rocky interior, which is plausible given the detection of hydrated sulfur compounds within chaos terrains and the suggestion of volcanic activity (Weber et al., 2023). If such compounds are indeed present, the seeping of ocean water into cracks in the Europan seafloor could bring these compounds to the crust-water interface, allowing for an anaerobic sulfate-reducing microorganism to metabolize and live either on the surface of the seafloor or within a vent fluid chimney, similarly to many species on Earth (Pérez-Rodríguez et al., 2012; Van Dover, 2000). When searching for analogs to life on Earth in seemingly desolate places, metabolisms such as sulfate reduction that could very well date back close to the origins of life on Earth should be granted priority, as parallel conditions could favor the origin of something similar elsewhere (Shen and Buick, 2004).
3.2 Serpentization
Even without volcanic activity, the rock-water interface still allows for consideration of an ecosystem reliant on the compounds and energy formed through serpentization. Serpentization is a process by which ferromagnesian silicate minerals, such as olivine and pyroxenes, formed at a planetary body’s creation come into contact with water and hydrolyze, commonly producing serpentine group minerals (Holm et al., 2015; Schwander et al., 2023). In doing so, this reaction releases significant amounts of heat, hydrogen gas, and methane, as well as a handful of biologically important trace metals (Vance et al., 2007). The production of hydrogen gas and methane is especially important, as these compounds are critical ingredients in many chemoautotrophic processes such as methane oxidation, methanogenesis, and sulfur reduction (Vance et al., 2007; Van Dover, 2000). It is highly possible that life on Earth originated at hydrothermal vents, and serpentization is a key process supporting this theory. 3.8 billion years ago, the first life on Earth was likely to be bacteria and or archaea reliant on serpentization in the deep ocean for the hydrogen gas needed for their metabolisms (Schwander et al., 2023). Modern methanogens on Earth are known for their resilience in extreme environments, such as the thermophilic archaea Methanococcus thermolithotrophicus isolated from geothermally heated sea sediments. If life on Earth really did originate hydrothermally through serpentization, it is entirely possible that it could have originated via the same pathway on another planetary body, especially considering the wide range of conditions that permit the process (Holm et al., 2015). In order for serpentization to support life on Europa, sufficient contact between ferromagnesian silicate minerals and liquid water would be needed, as well as a supply of carbon dioxide to support continued methanogenesis, to be a possibility at the Europan seafloor (Schwander et al., 2023). While we are still unable to confirm the occurrence of serpentization on Europa due to limited data regarding the moon’s composition, the possibility is a leading theory in Europan theoretical astrobiology and one that should be considered further upon further understanding of the minerals at the moon’s seafloor (Holm et al., 2015; Schwander et al., 2023). The existence and persistence of hydrogen gas-reliant life on Earth in such unusual conditions supports the idea that life could exist at extraterrestrial extremes as well, potentially in seafloor ecosystems similar to the Lost City hydrothermal system here on Earth (Huber et al., 1982; Schwander et al., 2023).
3.3 Radiogenic chemosynthesis
Along with traditional chemosynthetic pathways, metabolic pathways powered by radiogenic processes should be considered, especially for a moon experiencing as much radiation as Europa (Chyba and Phillips, 2001). Most of the focus on Europan radiolysis has been at the icy surface, where bombardment of charged particles from the Jovian magnetosphere could produce oxidants. These oxidants could greatly increase the chances for life’s existence on the moon, the challenge being that the compounds would have to somehow enter the ocean through the thick ice layer (Altair et al., 2018). For example, the possibility of methane oxidation being a viable metabolic pathway on Europa is dependent largely on the diffusion of compounds from the ice crust into the subsurface ocean (Chyba and Phillips, 2001). Methane oxidation is an aerobic pathway common on Earth, requiring oxygen (Van Dover, 2000). Oxygen is far from likely to be present in Europa’s ocean in the same way it is on Earth due to a lack of photosynthetic organisms. If molecular oxygen generated from water ice through the bombardment with Jovian radiation is able to enter the ocean below, methane generated from serpentization could allow for methane oxidation to occur (Weber et al., 2023). Without sufficient ice turnover or diffusion of oxidants, it is unlikely that any compounds produced at the surface would be able to enter the ocean and support life through oxidizing pathways. This is not to say that surface-ocean exchange on Europa is impossible, as some rate of turnover is highly supported by evidence. One such possibility is driven by the sinking of impact-generated melt chambers, which could sink through the ice shell, delivering oxidants from the surface and creating porous ice for the drainage of surface brines (Carnahan et al., 2022). Europa’s surface is relatively young, with the chaos terrains containing ice that is only 30–90 million years old (Schmidt, 2020). This could be a result of the thermal structuring theory, considering sub-surface melt chambers of warmer ice, allowing for compounds in the surface ice to mix into the ocean below in a slushy matrix (Chyba and Phillips, 2001). This is unlikely to be a reliable method of oxidant transportation, as resurfacing in these chaos terrains would extrapolate out to be a 10,000-to-100,000-year timescale for the turnover of only 10%–35% of the surface ice (Chyba and Phillips, 2001; Schmidt, 2020). Because surface-ocean exchange is a seemingly unreliable and lengthy process on Europa, it is essential to consider anaerobic pathways on Europa independent of surface chemistry.
A way to consider radiolysis on Europa independent of surface ocean exchange is through water radiolysis at depth. This kind of radiolysis would occur as a reaction between water and fissionable materials, heavily relying on direct contact between the seafloor and ocean, much like serpentization (Altair et al., 2018; Vance et al., 2007). Considering life powered by water radiolysis in the deep ocean is an interesting perspective, as it rules out the most commonly known sources of energy: solar radiation, geothermal heating, galactic cosmic rays, and planetary radiation (Altair et al., 2018). On Earth, radiolysis of water produces hydrogen gas in subsurface oceanic sediments, which have been found to support microbial communities. Experimental results indicate that this process may account for up to 10% of metabolic respiration at certain low organic carbon sites, becoming more significant in areas where organic-fueled respiration is low, potentially in the center of ocean gyres. This process has important implications for extraterrestrial life, as the only drivers of this simple metabolic pathway are the decay of elements such as uranium and thorium, and the presence of water (Blair et al., 2007). Considerations for this kind of radiolysis are being applied to the study of Martian astrobiology, with recent models outlining production rates of hydrogen gas in various Martian sediments and rocks. These results have been shown to be comparable to some production rates on Earth, and similar models could be constructed for other planetary bodies, such as Europa, once more data has been collected about the moon’s geochemistry (Dzaugis et al., 2018).
Another way in which water hydrolysis could support extraterrestrial life is through the production of sulfate as a secondary product. This could allow for sulfate reduction reactions similar to those found in some ecosystems on Earth (Weber et al., 2023). An example of a sulfate-reducing species reliant on water radiolysis is the terrestrial bacterium Candidatus Desulforudis audaxviator. This species was discovered 2.8 km below the Earth’s surface in a South African gold mine. Completely independent of the sun and anaerobic, this bacterium was the only organism found in the water of this mine, forming a single-species ecosystem (Chivian et al., 2008). Inspired by this discovery, a 2018 study investigated the possibility of an anaerobic, sulfate-reducing, water radiolysis-driven pathway to sustain life on Europa. This model showed that this hypothetical pathway could produce more than enough energy to sustain a microbial community on the seafloor of Europa, contributing to the ever-evolving list of potential pathways that could sustain life on the icy moon (Altair et al., 2018).
4 Other planetary bodies with potential to support life
4.1 Enceladus
Europa is a current focus of astrobiology as its vast amounts of liquid water, tidal heating, chemical composition, and rocky seafloor are characteristic supporters of chemosynthetic life. Now that a variety of pathways that could potentially support life on Europa have been identified, this framework can be applied to other moons and planets to see if they hold potential for microbial life. Similar to Europa, Saturn’s ice moon Enceladus has been of recent interest to astrobiologists. Enceladus’s ice shell is thick, but with thinner regions at the poles that jet plumes of water vapor (Weber et al., 2023). These plumes contain a myriad of compounds indicative of hydrothermal activity, including silica particles and organic molecules. These compounds not only provide direct evidence of hydrothermal activity but could also support potential metabolic pathways on the moon’s seafloor (Schmidt, 2020; Weber et al., 2023). Once hydrated, acetylene can create compounds relevant to methanogenic and sulfate-reducing anaerobes, including hydrogen gas, suggesting a handful of metabolic pathways (Weber et al., 2023). Unlike Europa, Enceladus receives little radiation and has little to no surface-ocean exchange, meaning the production of oxidants through radiation is even less likely than on Europa (Postberg et al., 2016).
4.2 Mars
A historically popular subject for astrobiology among scientists and the public alike is the planet Mars. Following the discovery of hydrothermal vents on Earth, the search for life on Mars has shifted from the search for familiar photosynthetic organisms to fossilized remains of hydrothermal vent ecosystems or ancient minerals indicative of hydrothermal activity (Van Dover, 2000). Through the study of its landforms, it is understood that Mars likely once had complex surface hydrology 3.8 billion years ago, with oceans producing fluvial landforms still seen today (Gillespie et al., 2024). A wide variety of evidence for past hydrothermal systems has been identified on Mars, from silica-rich deposits linked to volcanism, minerals associated with serpentization, and evidence of magma intrusion (Ojha et al., 2021). A compelling line of evidence for very early hydrothermal activity on Mars comes from a Martian meteorite containing impact-shocked zircon. This meteorite suggests magmatic hydrothermal activity on Mars as far back as 4.45 billion years ago, relatively close to the planet’s formation (Gillespie et al., 2024). The presence of this ancient system allows us to consider other forms of ancient hydrothermal activity on Mars, with one possibility being radiogenic. Unlike systems mentioned before, radiogenic hydrothermal systems powered by the decay of elements with half-lives of billions of years can remain active for far longer than hydrothermal systems powered by other means, up to one billion years (Ojha et al., 2021). If a magmatic or radiogenic system were present on Mars in its earlier years, that would allow for more time for microbial communities to be formed and establish, hopefully leaving us clues about ancient extraterrestrial life forms.
5 Discussion
Recent discussion and research into potential life on Jupiter’s icy moon Europa is well warranted, as the moon is currently the most promising location for undiscovered extraterrestrial life in our solar system (Howell and Pappalardo, 2020). Although bombarded by Jovian radiation and encased in a thick shell of ice, the salty ocean underneath could very well be a sanctuary for extraterrestrial microbial life (Weber et al., 2023). Despite possibly anoxic conditions, high pressures, and lack of sunlight, many physical and chemical properties of Europa are analogs of extreme environments on Earth, such as hydrothermal vents, serpentization sites, and subterranean radiogenic ecosystems. Along with these high-temperature systems, the diversity of chemosynthetic microorganisms on Earth includes a variety of low-temperature environments as well. While this review focuses on high-temperature systems in contact with rocky seafloor substrates, as they are commonly on Earth and continually linked to the origins of life on our planet, low-temperature systems on the Europan seafloor should be considered as well in a broader context to maintain an open and agnostic perspective, with recent research showing valuable applications to astrobiology (Jones et al., 2018; Robador, 2024). For example, within the low-temperature subsurface fluids of the oceanic crust, sulfate-reducing bacteria related to the aforementioned Candidatus Desulforudis have been found, and are suspected to help mediate the flow of organic materials through the upper crustal fluids, and may even influence marine and global biogeochemical cycling (Robador et al., 2015). Processes such as these on Europa, including serpentization, water radiolysis, and volcanic activity induced by tidal heating, could very well be providing energy to chemoautotrophs, unusual organisms alien even when observed on Earth (Chyba and Phillips, 2001; Schmidt, 2020). While the existence of life on Europa is entirely theoretical, there is hope that missions such as the launch of NASA’s Europa Clipper spacecraft will finally bring concrete answers and explore the moon’s habitability. Scheduled to reach Europa in 2030, the spacecraft is equipped with a variety of instruments to analyze its composition and chemistry. These instruments include a surface dust analyzer to analyze ejected particles, an imaging system, a near-infrared spectrometer to understand surface composition, and an ice-penetrating radar that will allow for complex mapping of the ice shell and ocean’s structure (Howell and Pappalardo, 2020). Existing concepts for Europa lander missions have been put on hold, with NASA electing to focus on indirect exploratory methods, such as those outlined by the Europa Clipper. Although direct access to subsurface materials would be ideal, remote sensing has its place in the current scope of exploration, given the limited surface exchange and thickness of the ice shell (National Aeronautics and Space Administration, 2019). With what little information regarding Europa’s properties and composition gleaned from high-level overview flybys like the Galileo spacecraft, there is much data to be collected and contributed to the field of astrobiology (Kivelson et al., 2000). Having detailed data from an Europan-specific mission will help astrobiologists worldwide continue the search for extraterrestrial life on the moon, and if life is found, allow for the search to expand to other planetary bodies, even those outside our solar system.
Author contributions
EB: Writing – original draft, Writing – review and editing. RL: Writing – review and editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Altair, T., de Avellar, M. G. B., Rodrigues, F., and Galante, D. (2018). Microbial habitability of europa sustained by radioactive sources. Sci. Rep. 8, 260. doi:10.1038/s41598-017-18470-z
Blair, C. C., D'Hondt, S., Spivack, A. J., and Kingsley, R. H. (2007). Radiolytic hydrogen and microbial respiration in subsurface sediments. Astrobiology 7 (6), 951–970. doi:10.1089/ast.2007.0150
Carnahan, E., Vance, S. D., Cox, R., and Hesse, M. A. (2022). Surface-to-ocean exchange by the sinking of impact generated melt chambers on europa. Geophys. Res. Lett. 49 (24), e2022GL100287. doi:10.1029/2022GL100287
Charette, M. A., and Smith, W. H. F. (2010). The volume of earth’s ocean. Oceanography 23 (2), 112–114. doi:10.5670/oceanog.2010.51
Chivian, D., Brodie, E. L., Alm, E. J., Culley, D. E., Dehal, P. S., DeSantis, T. Z., et al. (2008). Environmental genomics reveals a single-species ecosystem deep within Earth. Science 322 (5899), 275–278. doi:10.1126/science.1155495
Chyba, C. F., and Phillips, C. B. (2001). Possible ecosystems and the search for life on europa. Proc. Natl. Acad. Sci. 98 (3), 801–804. doi:10.1073/pnas.98.3.801
Clarke, A. (2014). The thermal limits to life on Earth. Int. J. Astrobiol. 13 (2), 141–154. doi:10.1017/S1473550413000438
Dzaugis, M., Spivack, A. J., and D'Hondt, S. (2018). Radiolytic H2 production in martian environments. Astrobiology 18 (9), 1137–1146. doi:10.1089/ast.2017.1654
Gillespie, J., Cavosie, A. J., Fougerhouse, D., Ciobanu, C. L., Rickard, W. D., Saxey, D. W., et al. (2024). Zircon trace element evidence for early hydrothermal activity on Mars. Sci. Adv. 10 (47), eadq3694. doi:10.1126/sciadv.adq3694
Hazael, R., Meersman, F., Ono, F., and McMillan, P. F. (2016). Pressure as a limiting factor for life. Life 6 (3), 34. doi:10.3390/life6030034
Hessler, R. R., and Smithey, W. M. (1983). “The distribution and community structure of megafauna at the Galapagos rift hydrothermal vents,” in Hydrothermal processes at seafloor spreading centers (Boston, MA: Springer US), 735–770.
Holm, N., Oze, C., Mousis, O., Waite, J., and Guilbert-Lepoutre, A. (2015). Serpentinization and the formation of H2 and CH4 on celestial bodies (planets, moons, comets). Astrobiology 15 (7), 587–600. doi:10.1089/ast.2014.1188
Howell, S. M., and Pappalardo, R. T. (2020). NASA’s europa Clipper—A mission to a potentially habitable ocean world. Nat. Commun. 11 (131), 1311. doi:10.1038/s41467-020-15160-9
Huber, H., Thomm, M., König, H., Thies, G., and Setter, K. O. (1982). Methanococcus thermolithotrophicus, a novel thermophilic lithotrophic methanogen. Archives Microbiol. 132, 47–50. doi:10.1007/BF00690816
Jones, R. M., Goordial, J. M., and Orcutt, B. N. (2018). Low energy subsurface environments as extraterrestrial analogs. Front. Microbiol. 9 (1605), 1605. doi:10.3389/fmicb.2018.01605
Kasting, J. F., Kopparapu, R., Ramirez, R. M., and Harman, C. E. (2014). Remote life-detection criteria, habitable zone boundaries, and the frequency of Earth-like planets around M and late K stars. Proc. Natl. Acad. Sci. 111 (35), 12641–12646. doi:10.1073/pnas.1309107110
Kivelson, M. G., Khurana, K. K., Russel, C. T., Volwerk, M., Walker, R. J., and Zimmer, C. (2000). Galileo magnetometer measurements: a stronger case for a subsurface ocean at europa. Science 289 (5483), 1340–1343. doi:10.1126/science.289.5483.1340
Matsuyama, I. N., Steinke, T., and Nimmo, F. (2022). Tidal heating in Io. Elements 18 (6), 374–378. doi:10.2138/gselements.18.6.374
Naganuma, T., and Uematsu, H. (1998). Dive europa: a search-for-life initiative. Biol. Sci. Space 12 (2), 126–130. doi:10.2187/bss.12.126
Nascimento-Dias, B. L., and Martinez-Frias, J. (2023). Brief review about history of astrobiology. Int. J. Astrobiol. 22 (1), 67–78. doi:10.1017/s1473550422000386
National Aeronautics and Space Administration (2019). Management of NASA’s europa mission. Available online at: https://oig.nasa.gov/wp-content/uploads/2024/02/IG-19-019.pdf.
Nealson, K. H., and Popa, R. (2005). “Metabolic diversity in the microbial world: relevance to exobiology,” in Micro-organisms and Earth systems. Editors G. M. Gadd, and H. M. Lappin-Scott (Cambridge University Press), 151–171. doi:10.1017/CBO9780511754852.009
Ojha, L., Karunatillake, S., Karimi, S., and Buffo, J. (2021). Amagmatic hydrothermal systems on Mars from radiogenic heat. Nat. Commun. 12, 1754. doi:10.1038/s41467-021-21762-8
Pappalardo, R. T., Head, J. W., Greeley, R., Sullivan, R. J., Pilcher, C., Schubert, G., et al. (1998). Geological evidence for solid-state convection in Europa's ice shell. Nature 391 (6665), 365–368. doi:10.1038/34862
Pérez-Rodríguez, I., Grosche, A., Massenburg, L., Starovoytov, V., Lutz, R. A., and Vetriani, C. (2012). Phorcysia thermohydrogeniphila gen. nov., sp. Nov., a thermophilic, chemolithoautotrophic, nitrate-ammonifying bacterium from a deep-sea hydrothermal vent. Int. J. Syst. Evol. Microbiol. 62 (10), 2388–2394. doi:10.1099/ijs.0.035642-0
Postberg, F., Tobie, G., and Dambeck, T. (2016). Under the sea of enceladus. Sci. Am. 315 (4), 38–45. doi:10.1038/scientificamerican1016-38
Reynolds, R. T., McKay, C. P., and Kasting, J. F. (1987). Europa, tidally heated oceans, and habitable zones around giant planets. Adv. Space Res. 7 (5), 125–132. doi:10.1016/0273-1177(87)90364-4
Robador, A. (2024). The subseafloor crustal biosphere: ocean’s hidden biogeochemical reactor. Front. Microbiol. 15, 1495895. doi:10.3389/fmicb.2024.1495895
Robador, A., Jungbluth, S. P., LaRowe, D. E., Bowers, R. M., Rappé, M. S., Amend, J. P., et al. (2015). Activity and phylogenetic diversity of sulfate-reducing microorganisms in low-temperature subsurface fluids within the upper oceanic crust. Front. Microbiol. 5 (748), 748. doi:10.3389/fmicb.2014.00748
Schmidt, B. E. (2020). “The astrobiology of Europa and the Jovian system,” in Planetary astrobiology (Tucson: University of Arizona). doi:10.2458/azu_uapress_9780816540068-ch008
Schwander, L., Brabender, M., Mrnjavac, N., Wimmer, J. L. E., Preiner, M., and Martin, W. F. (2023). Serpentinization as the source of energy, electrons, organics, catalysts, nutrients and pH gradients for the origin of LUCA and life. Front. Microbiol. 14, 1257597. doi:10.3389/fmicb.2023.1257597
Seeburger, R., Higgins, P. M., Whiteford, N. P., and Cockell, C. S. (2023). Linking methanogenesis in low-temperature hydrothermal vent systems to planetary spectra: methane biosignatures on an archean-earth-like exoplanet. Astrobiology 23 (4), 415–430. doi:10.1089/ast.2022.0127
Shen, Y., and Buick, R. (2004). The antiquity of microbial sulfate reduction. Earth-Science Rev. 64 (3), 243–272. doi:10.1016/S0012-8252(03)00054-0
Van Dover, C. L. (2000). The ecology of deep-sea hydrothermal vents. Princeton University Press. doi:10.2307/j.ctv1zm2v35
Vance, S. D., Harnmeijer, J., Kimura, J., Hussmann, H., deMartin, B., and Brown, J. M. (2007). Hydrothermal systems in small ocean planets. Astrobiology 7 (6), 987–1005. doi:10.1089/ast.2007.0075
Walker, M. E., and Rhoden, A. R. (2022). Tidal heating at europa using the multifrequency analysis of tidal heating toolkit. Planet. Sci. J. 3 (7), 149. doi:10.3847/psj/ac6df0
Keywords: Europa, astrobiology, hydrothermal vents, radiogenic ecosystems, chemoautotrophy
Citation: Barrett EG and Lutz RA (2025) Europa’s ocean: potential for extraterrestrial chemoautotrophy. Front. Astron. Space Sci. 12:1694079. doi: 10.3389/fspas.2025.1694079
Received: 27 August 2025; Accepted: 16 October 2025;
Published: 24 October 2025.
Edited by:
Josep M. Trigo-Rodríguez, Spanish National Research Council (CSIC), SpainReviewed by:
Alberto Robador, University of Southern California, United StatesCopyright © 2025 Barrett and Lutz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Richard A. Lutz, cmx1dHpAbWFyaW5lLnJ1dGdlcnMuZWR1