 Hailiang Yu1
Hailiang Yu1 Qi Wang1
Qi Wang1 Jianqiang Tang1Liyue Dong1
Jianqiang Tang1Liyue Dong1 Guojun Dai1*
Guojun Dai1* Tao Zhang1
Tao Zhang1 Genxi Zhang1
Genxi Zhang1 Kaizhou Xie1Hongsheng Wang2
Kaizhou Xie1Hongsheng Wang2 Zhenhua Zhao3
Zhenhua Zhao3- 1College of Animal Science and Technology, Yangzhou University, Yangzhou, China
- 2Technical Research Department, Jiangsu Jinghai Poultry Group Co., Ltd., Haimen, China
- 3Poultry Institute, Chinese Academy of Agricultural Sciences, Yangzhou, China
Background: Coccidiosis is an intestinal parasitic disease caused by Eimeria protozoa, which endangers the health and growth of animals, and causes huge economic losses to the poultry industry worldwide every year. Studies have shown that poultry gut microbiota plays an important role in preventing the colonization of pathogens and maintaining the health of the host. Coccidia infection also affects host gene expression. However, the underlying potential relationship between gut microbiome and host transcriptome during E. tenella infection in chickens remain unclear.
Methods: In this study, metagenomic and transcriptome sequencing were applied to identify microbiota and genes in cecal contents and cecal tissues of infected (JS) and control (JC) chickens on day 4.5 postinfection (pi), respectively.
Results: First, microbial sequencing results of cecal contents showed that the abundance of Lactobacillus, Roseburia sp. and Faecalibacterium sp decreased significantly after E. tenella infection (P < 0.05), while the abundance of Alistipes and Prevotella pectinovora increased significantly (P < 0.05). Second, transcriptome sequencing results showed that a total of 434 differentially expressed mRNAs were identified, including 196 up-regulated and 238 down-regulated genes. These differentially expressed genes related to inflammation and immunity, such as GAMA, FABP1, F2RL1 and RSAD2, may play an important role in the process of host resistance to coccidia infection. Functional studies showed that the enriched pathways of differentially expressed genes included the TGF-beta signaling pathway and the ErbB signaling pathways. Finally, the integrated analysis of gut microbiome and host transcriptome suggested that Prevotella pectinovora associated with FABP1, Butyricicoccus porcorum and Colidextribacter sp. associated with RSAD2 were involved in the immune response upon E. tenella infection.
Conclusion: In conclusion, this study provides valuable information on the microbiota and key immune genes after chicken E. tenella infection, with the aim of providing reference for the impact of coccidia infection on cecal microbiome and host.
1 Introduction
Coccidiosis is a poultry intestinal parasitic disease caused by Eimeria protozoa, which seriously affects the development of the global poultry industry. Studies have shown that the annual global economic loss due to coccidiosis exceeds $14 billion (Blake et al., 2020; Wickramasuriya et al., 2023). Economic costs mainly include the prevention and treatment of coccidiosis (coccidiosis drugs and high vaccine costs) and subclinical symptoms of chicken populations. Infected chickens cause enormous economic losses to the poultry industry, including reduced body weight gain, feed conversion and egg production (Burrell et al., 2019; Yu et al., 2020). At present, due to the prohibition of antibiotics and food safety issues, the use of traditional anticoccidial drugs is limited and the alternative strategies for coccidiosis require a lot of attention (Yu et al., 2021a).
The gastrointestinal system of poultry, like mammals, is a complex physiological structure that plays crucial roles in digestion and host health. The gut microbiota is able to digest food ingested by animals and provide essential nutrients for growth, development, reproduction and daily activities of the organism (Baldwin et al., 2018; Madlala et al., 2021). In addition, when pathogens invade, gut microbiota can inhibit the colonization of pathogens and play an important role in maintaining intestine homeostasis and host health (Stanley et al., 2014; Clavijo and Flórez, 2018). Compared to other segments of the gut, such as duodenum, jejunum and ileum, the cecum has the highest number, richness and diversity of gut microbiota (Yeoman et al., 2012; Rubio, 2019), which play an important role in the repeated fermentation and absorption of nutrients. E. tenella mainly invades the cecal tissue of chickens, leading to swelling and bleeding of the cecum tissues and changes of cecal microbiota, which ultimately reduce the production performance of chickens (Yu et al., 2022). Studies have shown that gut microbiota play an important role in the resistance of poultry to coccidia infections. Butyrate alleviates the severity of E. tenella infection in chickens (Chen et al., 2020). Li et al. (2017) showed that the addition of Lactobacillus acidophilus to the diet enhanced the inflammatory response in poultry and modulated the innate and acquired immune response. In addition, in ovo supplementation with probiotics protected early chicks against Eimeria infection (Pender et al., 2016). Therefore, microbiota plays an important role in birds’ resistance to pathogens such as coccidiosis and maintaining healthy growth and development.
In recent years, the development of high-throughput sequencing technologies and omics analysis have significantly expanded our knowledge of the gut microbiota of animals. However, the detailed function of the gut microbiota in infected diseases (such as coccidiosis) and the interactions between the microbiota and the host remain unclear. In this study, the metagenomic and transcriptome sequencing technologies were used to sequence the cecal contents and cecal tissue of infected group and control group chickens on day 4.5 pi to explore the effect of coccidia infection on cecal microbiota and host gene expression, respectively and interaction analysis between microbiota and host was also carried out. The purpose of this study is to provide a reference for the impact of coccidia infection on cecal microbiome and host.
2 Materials and methods
2.1 Animals and oocysts
A total of twenty 1-day-old healthy Chinese yellow-feathered broiler chicks with similar size were randomly selected. The experimental chicks were kept in a clean, pathogen-free animal room, with free access to food and water. At the age of 18 days, all the chicks were divided into two groups. Each bird in the infection group was orally fed with 3.5×104 sporulated oocysts (water was not provided within 6 hours after the challenge to ensure that the oocysts were ingested by the birds), and the chicks of control group were fed with the same amount of normal saline. E. tenella oocysts, a Yangzhou strain in China, were a gift from the Parasite Laboratory in the Veterinary Medicine College of Yangzhou University. The collection, purification and sporulation of fresh oocysts were described in previous articles (Yu et al., 2022).
2.2 Sample collection
On 4.5 days (108 h) post infection, 4 individuals of female chicks were randomly selected from each group and euthanized by rapid cervical dislocation. After the cecum was isolated, the intestinal contents were collected immediately in cryovials. The cecum tissues were washed with sterilized PBS for excess intestinal contents and mucus, and placed in cryopreservation tubes. All samples were immediately stored in liquid nitrogen and then transferred to a -80°C refrigerator for later use. The animal experiment protocols were approved by the Animal Welfare Committee of Yangzhou University (permit number: SYXK (Su) IACUC 2012-0029).
2.3 Metagenomic sequencing analysis of cecal microbiome
2.3.1 DNA isolation and library construction
Total DNA of chicken cecal microorganisms was extracted using a QIAamp® Fast DNA Stool Mini Kit (Qiagen, Hilden, Germany) according to the manufacturer’s instructions. The DNA integrity of the sample was detected by agarose gel electrophoresis, and the DNA concentration was measured with a NanoDrop2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA). Library construction was performed using the TruSeq Nano DNA LT Sample Preparation Kit (Illumina, San Diego, CA, USA). The library construction and sequencing were carried out by OE Biomedical Technology Co., Ltd (Shang hai, China).
2.3.2 Metagenome sequencing analysis
The library was sequenced using the llumina Novaseq 6000 sequencing platform, and 150 bp paired-end reads were generated (the sequencing depth/volume is 10 G). The raw data is preprocessed using Trimmomatic (v 0.36) (Bolger et al., 2014), including removing adapters, low-quality Reads and low-quality bases; The filtered reads were aligned to the chicken genome (Gallus gallus bGalGal1.mat.broiler. GRCg7b) using bowtie2 (v 2.2.9), and the aligned reads were removed. Then Metagenome assembly was performed on the valid reads using the splicing software MEGAHIT (v 1.1.2) (Li et al., 2015). ORF prediction of assembled scaffolds using prodigal was performed and translated into amino acid sequences. The non-redundant gene sets (in-house version) were built for all predicted genes using CDHIT (v 4.5.7). The Clean reads of each sample were aligned against the non-redundant gene set (95% identity) use bowtie2 (v 2.2.9), and the abundant information of the gene in the corresponding sample was counted.
Species annotations were obtained through the corresponding taxonomic information database of NR library, and then the abundance of the species was comprehensively calculated using the corresponding abundance of gene. To construct the abundance profile of taxonomy level, abundance statistics of samples were performed at different level, including Domain, Kingdom, Phylum, Class, Order, Family, Genus and Species. The gene set representative sequence (amino acid sequence) was annotated with COG, NR, SWISSPROT, KEGG and GO database with an e-value of 1e-5 using DIAMOND (v 0.9.7) (Buchfink et al., 2015). The taxonomy abundance spectrum or functional abundance spectrum were subjected to PCA analysis and plotted using prcomp function and ggplot2 package in R.
2.3.3 Predictive function of differential cecal microbiota
To reveal the predictive function of differential microbiota after coccidia infection, eggNOG (Evolutionary genealogy of genes: Non-supervised Orthologous Groups) and were used for pathway enrichment analysis of differential cecal microbiota. The eggNOG database used the Smith-Waterman alignment algorithm to annotate the function of constructed Orthologous Groups. KEGG database is a comprehensive database for annotating gene functions (DIAMOND software), the core of which is kegg pathway and kegg orthology database.
2.4 Transcriptome sequencing analysis
2.4.1 RNA isolation
Total RNA from cecum tissues were extracted with TRIzol reagent (Invitrogen, CA, USA) following the manufacturer’s recommended procedure. The NanoDrop 2000 spectrophotometer (Thermo Fisher Scientific, Waltham, MA, USA) was used to measured RNA purity and concentration, and the Agilent 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA) was used to evaluate RNA integrity. The Host transcriptome sequencing can be performed on the basis of meeting RNA quality standards (RNA integrity number>=7 and 28S/18S>=0.7).
2.4.2 Library construction and Sequencing
Then the libraries were constructed using TruSeq Stranded Total RNA with Ribo-Zero Gold (illumina, Cat.No. RS-122-2301) according to the manufacturer’s instructions. The libraries were sequenced on an Illumina HiSeq X Ten platform, 150 bp paired-end reads were generated (the sequencing depth/volume is 6 G). Raw reads of fastq format were firstly processed using fastp and the low-quality reads were removed to obtain the clean reads. Then the clean reads were mapped to the chicken reference genome using HISAT2 (Kim et al., 2015). For mRNAs, FPKM (Roberts et al., 2011) of each gene was calculated using Cufflinks (Trapnell et al., 2010), and the read counts of each gene were obtained by HTSeq-count (Anders et al., 2015). Differential expression analysis was performed using the DESeq2 package in R. P-value < 0.05 was set as the threshold for significantly differential expression.
2.4.3 Functional enrichment analysis of differentially expressed genes
After the differentially expressed genes were obtained, GO enrichment analysis was performed to describe their functions. GO enrichment contains three class: biological process, cellular component and molecular function. Moreover, the KEGG database was used to conduct pathway enrichment analysis of differential expressed genes. GO terms and KEGG pathways were considered to be significantly enriched based on a P-value <0.05.
2.5 Integrated analysis of gut microbiome and host transcriptome
To explore the potential relationship between cecal microbiome and hosts after E. tenella infection, Spearman correlation coefficient was used to calculate the correlation between differentially microbiota and differentially expressed genes. Spearman correlation coefficient |r| > 0.8 and P-value < 0.05 are considered as significantly relationship pairs. The correlation heatmap of cecal microbiota and differentially expressed genes were constructed using the R package, and the co-expression network map were constructed using Cytoscape (v3.8.2) (Smoot et al., 2011).
2.6 Correlation analysis of gut microbiome after E. tenella infection
Moreover, to explore the potential relationship between cecal microbiome and hosts after E. tenella infection, this study also selected the top 30 microbiotas with the total abundance at the genus level, and used the Spearman correlation coefficient to explore the correlation between dominant genus. The R package was used to generate the correlation heatmap of the dominant genus.
2.7 Histopathology
The cecal tissues from JC and JS groups were cut into approximately 0.5 cm pieces and fixed with 4% tissue fixative. After embedding in paraffin and sectioning (5 μm), the sections were stained with hematoxylin-eosin. Then, the slices were observed and imaged under a microscope (Nikon, Japan).
2.8 Statistical analysis
The difference between different groups was analyzed using Wilcoxon rank-sum test. The linear discriminant analysis effect size (LEfSe) method (Segata et al., 2011) was used to compare the taxonomy abundance spectrum or functional abundance spectrum and Linear discriminant analysis (LDA) score > 2.0 and P-value <0.05 were considered as significant difference. In addition, the correlation analysis was constructed to investigate the interaction between microbiome and host transcriptome, and Spearman coefficient |r| > 0.8 and P-value < 0.05 were shown.
3 Results
3.1 Chicken models of E. tenella infection
On day 4.5 pi, all chickens were euthanized and cecal tissues were harvested. As expected, the cecal tissues of the chickens in JS group had severe intestinal bleeding, and the cecum wall was obviously swollen and thickened, while the intestinal tissue structure of the chickens in JC group was intact (Supplementary Figure S1). In addition, the pathological analysis of cecal tissue in this study showed that the tissue layers of JC group chickens had a clear structure, the mucosal epithelium was intact, and the epithelial cells were tightly arranged. Extensive mucosal layer necrosis and ulcers appeared in the cecal tissue of JS group chickens (Supplementary Figure S2). The results of PCA plot also showed that the cecal microbiota of the JS group was different from that of the JC group (Supplementary Figure S3). Thus, these data indicated that the chicken model of E. tenella infection was successfully constructed.
3.2 Effects of E. tenella infection on cecal microbiota
To reveal the impact of E. tenella infection on host cecal microbiota, we analyzed the changes of cecal microbiota at the phylum, genus, and species level (Figure 1, Supplementary Figure 4). At the phylum level, Bacteroidetes, Firmicutes, Proteobacteria and Actinobacteria were the most dominant phylum in the chicken cecal contents (Figure 1A). Moreover, we found that the abundance of Firmicutes, Proteobacteria and Actinobacteria decreased, while the abundance of Bacteroidetes increased after E. tenella infection. At the genus level, Bacteroides, Alistipes, Phocaeicola, and Clostridium were the most dominant genus in the chicken cecal contents (Figure 1B). In addition, Clostridia bacterium, Rikenellaceae bacterium, Alistipes sp. CAG:831, and Bacilli bacterium at the species level had the relatively high abundance in chicken cecal microbiota (Figure 1C).

Figure 1 Comparison of microbial community composition between JC and JS groups. (A), phylum. (B), genus. (C), species. JC, control group; JS, infected group.
3.3 Differential abundance analysis of cecal microbiota after chicken E. tenella infection
To identify the important microbiota during the process of E. tenella infection in chickens, we performed differential abundance analysis of cecal microbiota in different groups by using the Wilcoxon rank-sum test. The results of the top 30 species with significant differences were shown in Figure 2. At genus level, E. tenella infection significantly reduced the abundance of Lactobacillus, Colidextribacter, Pseudobutyrivibrio, Shuttleworthia, and Weissella in the cecal microbial community (P <0.05, Figure 2A, C). At the species level, the abundance of Lactobacillus reuteri, Lactobacillus crispatus, Intestinimonas butyriciproducens, Butyricicoccus porcorum, and Roseburia sp. in the cecal microbiota were significantly reduced when chickens were infected by E. tenella (P <0.05, Figure 2B, D). However, the abundance of Prevotella pectinovora increased significantly (P <0.05, Figure 2D). Taken together, these results suggest that coccidiosis infection alters the composition of the cecal microbiota.
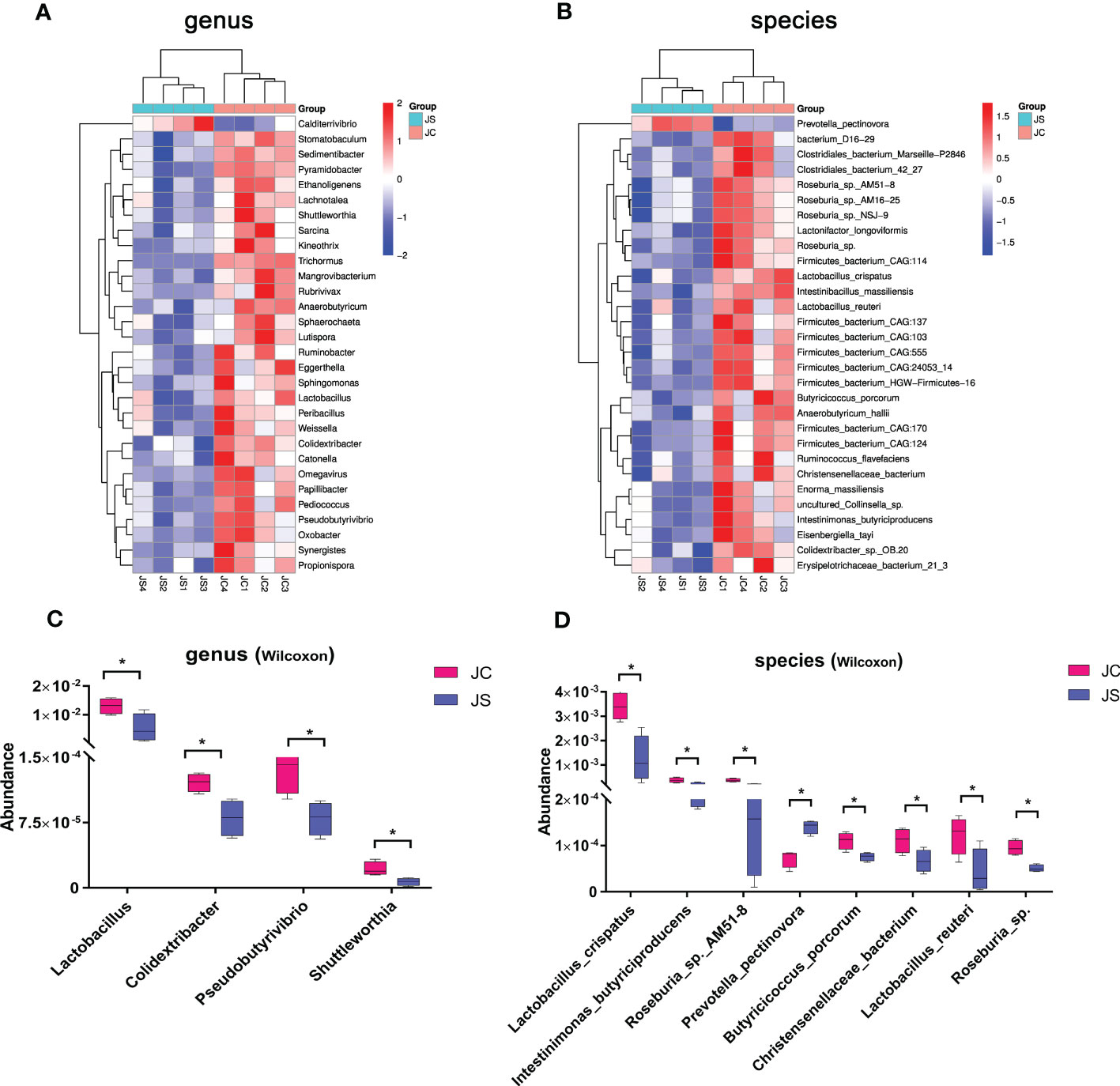
Figure 2 Differential abundance analysis of cecal microbiota after chicken E. tenella infection. (A, B) Hierarchical clustering plots of top 30 microbiota with significant differences at genus level (A) and species level (B). (C, D) The relative abundance of key microbiota at genus level (C) and species level (D). The bar represents the maximum and minimum values of a set of data. Statistical analyses were performed using Wilcoxon rank-sum test (*P < 0.05). JC, control group; JS, infected group.
3.4 LEfSe analysis of differentially microbiota in chicken cecal contents after E. tenella infection
LEfSe analysis, also known as LDA EFfect Size analysis, is an analytical method to count and find biomarkers with significant differences between different groups. To further understand and reveal the effects of E. tenella infection on the microbial community of chicken cecum, we performed LEfSe analysis of cecal microbiota in the infected and control chickens (Figure 3A). Similarly, we found that the abundance of lactobacillus salivarius, lactobacillus aviarius, lactobacillus johnsonii, and lactobacillus crispatus decreased during E. tenella infection (Figures 3C-F). In addition, the abundance of Roseburia sp, Faecalibacterium sp and Ruminococcus gauvreauii also decreased (Figures 3G-I). However, the abundance of Alistipes increased after coccidia infection (Figure 3B).
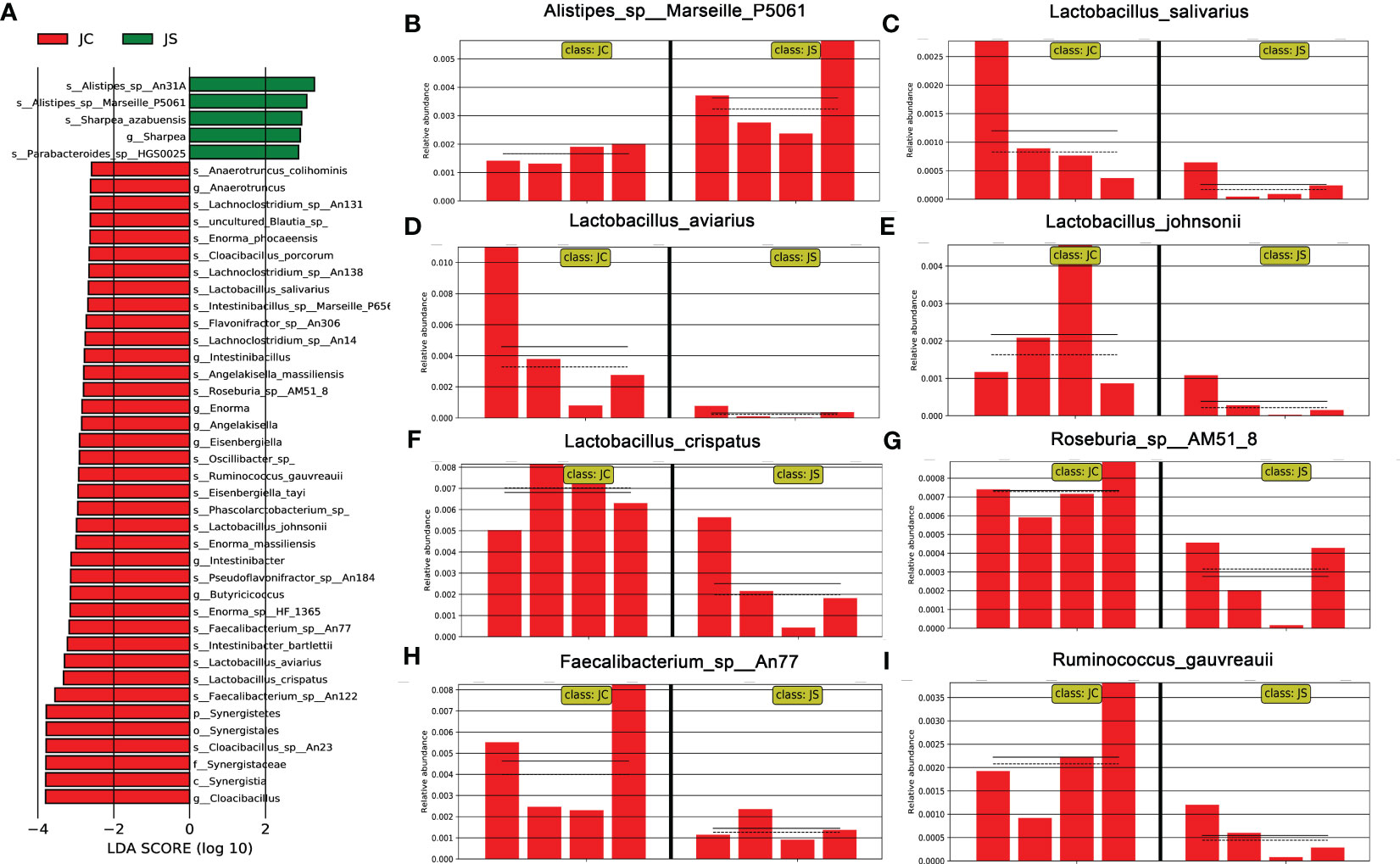
Figure 3 Linear discriminant analysis effect size (LEfSe) analysis of the cecal microbiota in different groups. (A) LDA score of differential cecal microbiota. LDA score > 2.0 and P < 0.05 were considered to be significantly differences. The green and red horizontal color bars represent microbial enrichment in the JS and JC groups, respectively. JC, control group; JS, infected group. (B-I) Histogram of relative abundance of differential cecal microbiota. (B) Alistipes_sp:Marseille_P5061. (C) Lactobacillus_salivarius. (D) lactobacillus_aviarius. (E) lactobacillus_johnsonii. (F) lactobacillus_crispatus. (G) Roseburia_sp:AM51_8. (H) Faecalibacterium_sp:An77. (I) Ruminococcus_gauvreauii.
3.5 Functional enrichment analysis of differential microbiota after chicken E. tenella infection
To reveal the potential function of these differential microbiota, we performed functional pathway enrichment analysis. The eggNOG analysis results showed that the significantly enriched pathways mainly include signal transduction mechanisms and defense mechanisms, etc (Figure 4A). The KEGG enrichment analysis indicated that differential microbiota may be involved in pathways such as cell growth and death, immune system and immune diseases, and environmental adaptation (Figure 4B).
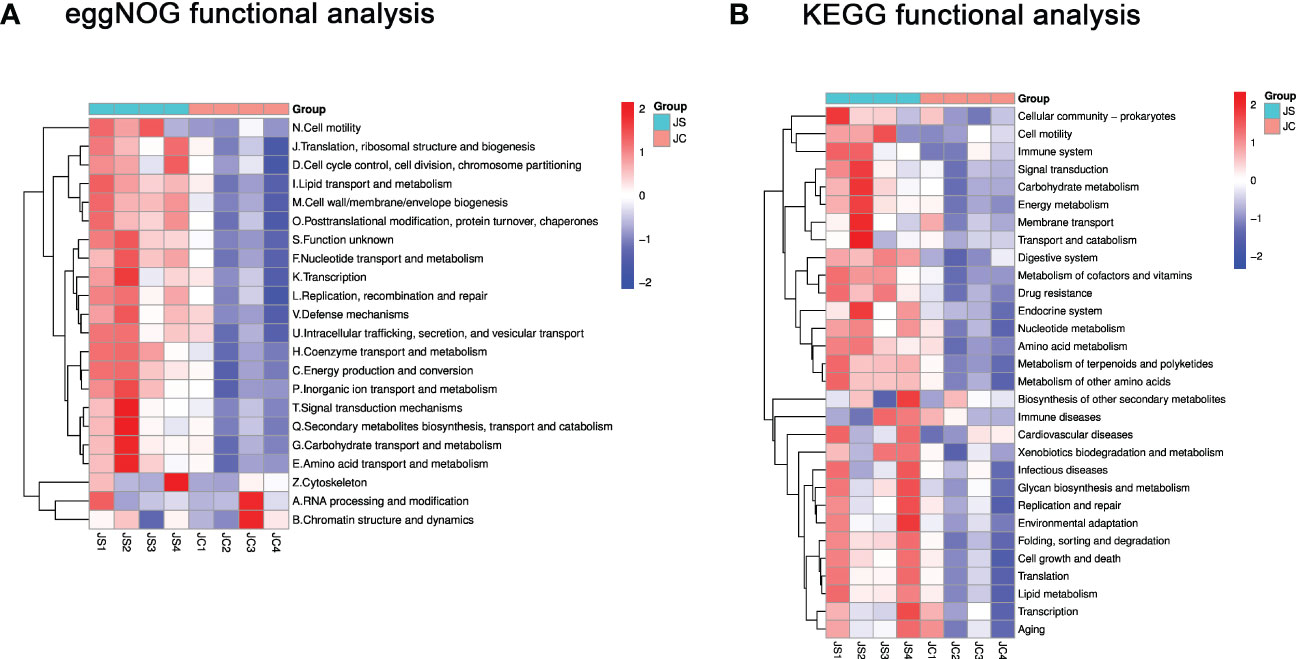
Figure 4 Functional enrichment analysis of differential cecal microbiota after chicken E. tenella infection. (A) Hierarchical clustering plots of eggNOG functional analysis. (B) Hierarchical clustering plots of KEGG functional analysis. Red and blue colors represent increased and decreased levels, respectively. JC, control group; JS, infected group.
3.6 Transcriptome sequencing analysis of chicken cecum tissues after E. tenella infection
To investigate the effect of E. tenella infection on the host transcriptome, RNA-seq was carried out to detect differentially expressed mRNAs in chicken cecal tissues from the control and infection groups on day 4.5 pi, respectively. After removing the most discrete individuals from each group, the results showed that a total of 434 genes were differentially expressed in chicken cecal tissues after E. tenella infection, including 196 up-regulated genes and 238 down-regulated genes (Figure 5A). Hierarchical cluster analysis displayed that expression patterns of significantly differentially expressed mRNA between the control group and the treatment group were different (Figure 5B). The key differential expressed genes such as GAMA, FABP1, F2RL1 and RSAD2 may play an important role in host resistance to coccidiosis infection.
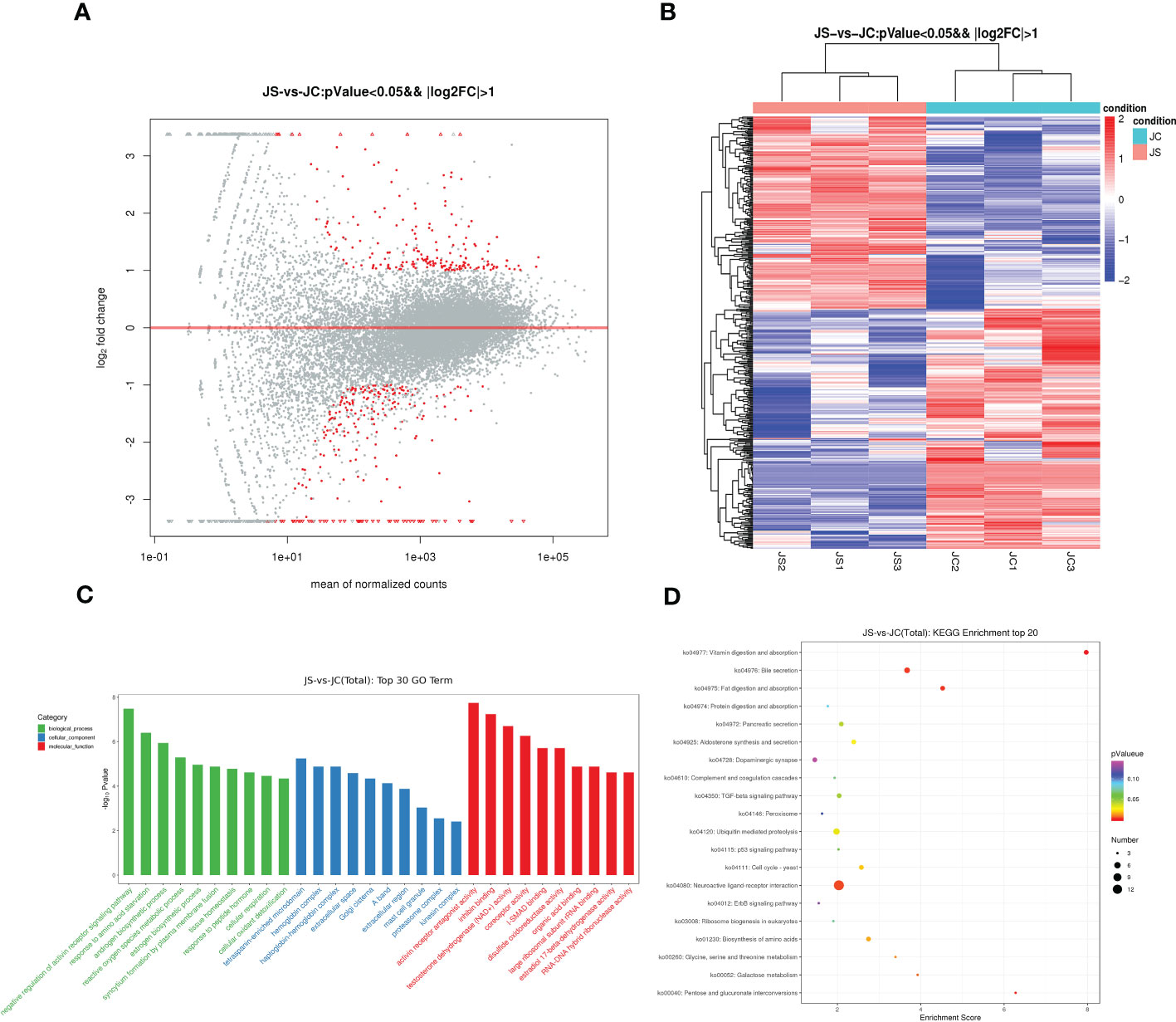
Figure 5 Transcriptome sequencing analysis of chicken cecal tissues after E. tenella infection. (A) The MA plot of differentially expressed mRNAs between JC and JS groups. Red dots represent differentially expressed genes, while gray dots represent genes with insignificant differences (196 up-regulated genes and 238 down-regulated genes). (B) Hierarchical clustering plots of differentially expressed mRNAs. Red and blue colors represent increased and decreased levels, respectively. (C) GO term analysis of differentially expressed mRNAs between JC and JS groups (Top 30). (D) KEGG pathway analysis of differentially expressed mRNAs between JC and JS groups (Top 20). The size of the circle represents the enriched number of differentially expressed genes and color represents significance. GO terms and KEGG pathways were considered to be significantly enriched based on a P-value <0.05. JC, control group; JS, infected group.
Moreover, to further reveal the function of differentially expressed genes, we performed both GO and KEGG pathway enrichment analysis. The top 30 significantly enriched GO terms were shown in Figure 5C. The results showed that the differentially expressed genes were mainly enriched in cellular oxidant detoxification, tissue homeostasis and negative regulation of activin receptor signaling pathway. The results of the significantly enriched top 20 KEGG pathways showed that the TGF-beta signaling pathway and ErbB signaling pathway were significantly enriched in JC and JS groups (Figure 5D).
3.7 Interaction analysis of gut microbiome and host transcriptome
Through metagenomic sequencing analysis, we found that chicken E. tenella infection affect the changes in the abundance of cecal microbiota. In addition, we also identified a total of 434 differentially expressed genes in cecal tissues after E. tenella infection by using transcriptome sequencing. To explore the potential relationship between intestinal microbiota and host transcriptome after E. tenella infection, we use Spearman’s correlation coefficients to conduct the interaction analysis between differential microbiota and differentially expressed genes. As shown in the Figures 6A, B, Prevotella pectinovora has a significant positive correlation with FABP1 (r=0.94, P <0.05), LECT2 (r=0.88, P <0.05) and SERPINB1 (r=0.88, P <0.05). GZMA has a significant negative correlation with Roseburia sp. NSJ-9 (r=0.94, P <0.05) and Roseburia sp. AM51-8 (r=0.95, P <0.05). RSAD2 was significantly negatively correlated with Butyricicoccus porcorum (r=0.88, P <0.05) and Colidextribacter sp. OB.20 (r=0.99, P <0.01). Notably, LOC107052718, an unknown gene, was significantly positively correlated with 11 cecal microbiota (P <0.05). SERPINB1 was only significantly positively correlated with Prevotella pectinovora, but had a significant negative correlation with the other 8 microbiota (P <0.05). These molecular markers may play an important role after E. tenella infection in chicken.
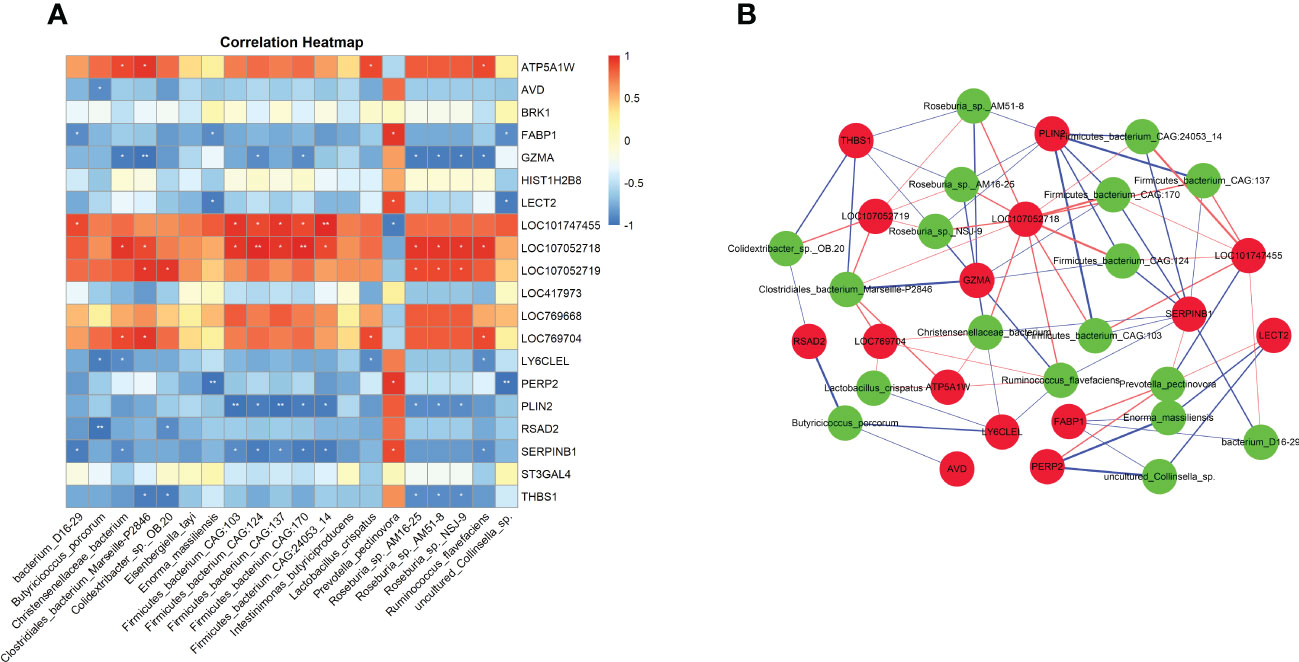
Figure 6 The interaction analysis of gut microbiome and host transcriptome. (A) Heatmap of Spearman’s correlation coefficients between differential microbiota and differential expressed mRNAs. Each row represents a differentially expressed mRNAs and each column represents a differentially microbiota. Red indicates positive correlations while blue indicates negative. The darker the color, the higher the correlation was. Spearman coefficient |r| > 0.8 and P < 0.05 were considered a significant difference. Correlation significance, *P < 0.05, **P < 0.01. (B) Association network of differential microbiota and differential expressed mRNAs. Red circles represent differentially expressed genes and green circles represent microbes. Red edges represent positive correlations and blue edges represent negative correlations. Edge thickness represents the strength of the correlation.
3.8 Correlation analysis of gut microbiome after E. tenella infection
In addition, to reveal potential associations between cecal microbiota after coccidia infection, we selected the top 30 species in total abundance at the genus level and constructed the correlation analysis between dominant genus according to Spearman’s correlation coefficients (Figure 7). The results showed a significant negative correlation between Alistipes and Butyricicoccus (r=0.83, P <0.05) and a significant positive correlation between Ruminococcus and Eubacterium (r=0.73, P <0.05).
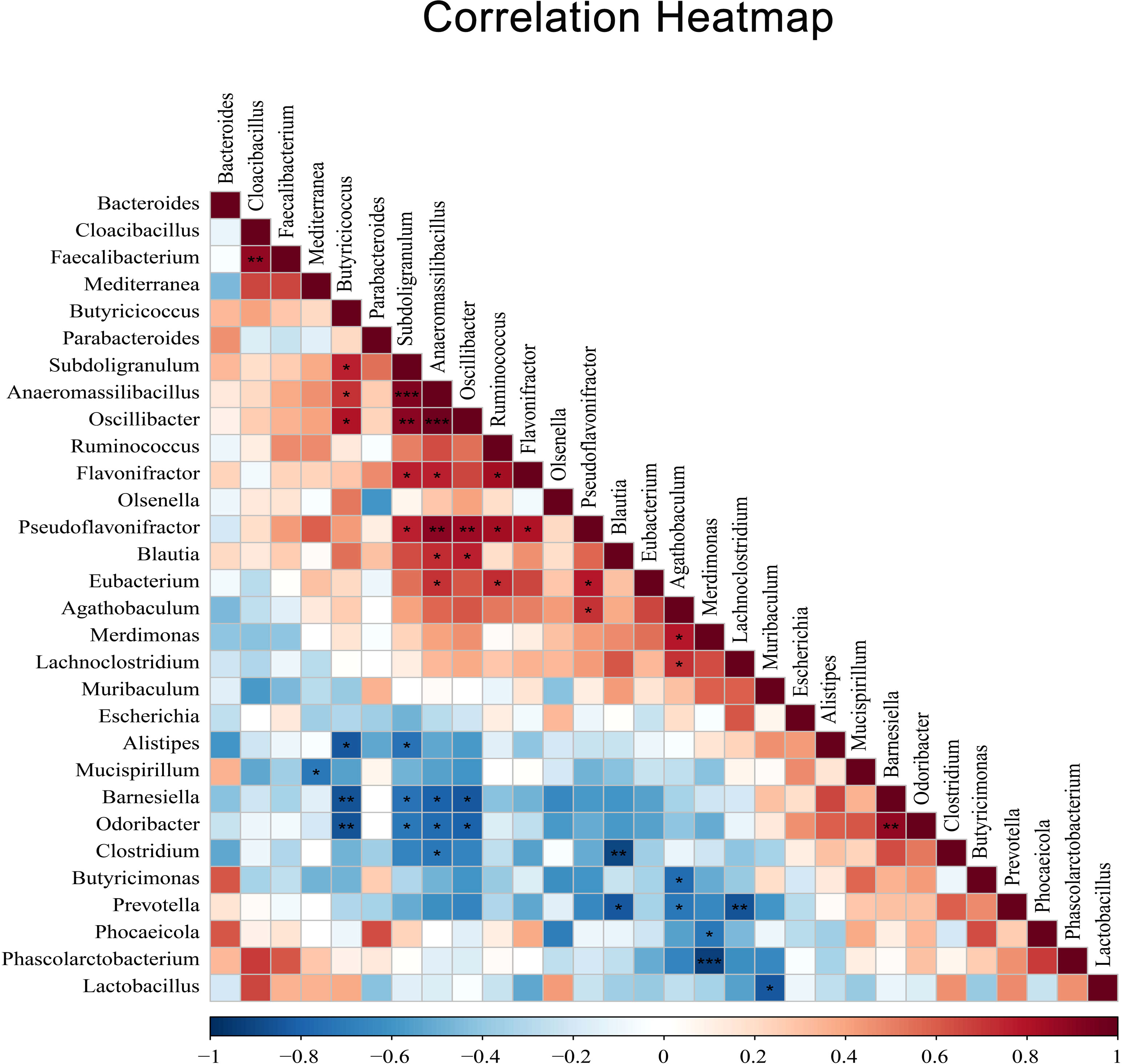
Figure 7 The heatmap of Spearman’s correlation coefficients between different dominant genus. Red indicates positive correlations while blue indicates negative. The darker the color, the higher the correlation was. Spearman’s correlations of different dominant genus were tested with a coefficient |r| > 0.8 and P < 0.05. Correlation significance, *P < 0.05, **P < 0.01, ***P < 0.001.
4 Discussion
The gastrointestinal microflora plays an important role in the nutrient absorption and immune protection process of poultry (Baldwin et al., 2018). Therefore, it is meaningful to study the effect of E. tenella infection on the cecal microbiota of chickens. In this study, metagenomic sequencing analysis of the cecal contents after E. tenella infection showed that Bacteroidetes, Firmicutes, Proteobacteria, and Actinobactereia were the most dominant phylum in the chicken cecal contents, which was consistent with previous studies consistent (Waite and Taylor, 2015; Zhou et al., 2020). Studies have shown that a gastrointestinal tract microbial of chickens with high diversity is expected to be more stable and healthier than a low diversity one (Clemente et al., 2012). In this study, compared with the control group, the proportion of Firmicutes, Proteobacteria, Actinobactereia, etc. at the phylum level in the infected group decreased, and the abundance of most genera such as Faecalibacterium, Lactobacillus, and Mediterranea also decreased. Therefore, E. tenella infection changed the diversity of cecal microbiota and affected the health of the host.
To screen out the important microbiota after E. tenella infection, we used Wilcoxon rank-sum test to analyze the significant difference of cecal microbiota between the JC and JS groups. The results showed that the abundance of Lactobacillus and Roseburia significantly decreased during E. tenella infection, while the abundance of Prevotella pectinovora significantly increased. In addition, we performed LEfSe analysis (an analysis method for biomarkers that find significant differences between different groups) on the cecal microbiota of the JC and JS groups. the results also showed that the abundance of Lactobacillus (includes salivarius, aviarius, johnsonii, and crispatus), Roseburia sp and Faecalibacterium sp decreased significantly, while the abundance of Alistipes increased after coccidia infection. Lactobacillus are an important component of the intestinal microbiota of mammals and birds, with functions such as resisting colonization by harmful bacteria, regulating inflammatory responses, and protecting the intestinal barrier (Zhao et al., 2021). Current evidence suggests that Lactobacillus play an important role in the anti-inflammatory immune response. Studies have shown that Lactobacillus casei and Lactobacillus johnsonii regulate the production of IL-12 in macrophages to adapt to infection by pathogenic bacteria (Teame et al., 2020). The peptidoglycan Ls33 purified from Lactobacillus salivarius also exerts anti-inflammatory effects by inducing the production of IL-10 (Fernandez et al., 2011). Lactobacillus acidophilus deficient in lipoteichoic acid downregulates the expression of IL-12 and TNF-a in dendritic cells and promotes IL-10 production to reduce intestinal inflammation (Mohamadzadeh et al., 2011). Roseburia is one of the commensal bacteria in the intestinal microbiota that produce short-chain fatty acids such as butyric acid, which maintain intestinal immunity and improve disease resistance (Tamanai-Shacoor et al., 2017). Roseburia has been shown to stimulate intestinal cells to secrete immune cytokines that promote the activation of regulatory T (Treg) cells (Nie et al., 2021). Butyrate produced by Roseburia promotes the production of IL-10, TGF-β, and IFN-r cytokines to influence the differentiation of Treg cytokines (Hegazy et al., 2017). Our previous study showed that Treg cells and related cytokines were up-regulated in cecum tissue after chicken coccidia infection, and are involved in the regulation of intestinal inflammation (Yu et al., 2021b). Therefore, coccidiosis infection causes structural destruction of intestinal tissues and changes in the intestinal microbiota, and butyrate produced by Roseburia may also be involved in the host’s inflammatory response by regulating the production of Treg cytokines. Alistipes are a relatively new microbiota isolated from the intestinal microbial community. Current research suggests that Alistipes reduce the inflammatory response in ulcerative colitis (UC) and inflammatory bowel disease (IBD) (Parker et al., 2020) and are also involved in cancer disease (Iebba et al., 2018). In poultry, Alistipes have been identified as producers of propionate in the chicken cecal microbiota (Polansky et al., 2015), and low levels of short-chain fatty acids such as butyric acid, propionic acid in the gastrointestinal system are associated with disease and inflammatory responses (Zaky et al., 2021). Our study showed the abundance of Alistipes in cecal microbiota increased significantly after E. tenella infection, which is consistent with the findings of Jebessa et al. (Jebessa et al., 2022). These evidence reveals that Alistipes may also be involved in the host immune response to chicken coccidia infection. Moreover, our study found that Prevotella are elevated after E. tenella infection in chickens. It was shown that elevated abundance of Prevotella enhances Th17-mediated mucosal inflammatory response and promotes Th17 cell immune factor production and neutrophil recruitment (Larsen, 2017). Th17 cell-mediated immune responses play an important role in the process of coccidia infection (Kim et al., 2012; Min et al., 2013), and thus elevated Prevotella may play an important role in immune response to chicken coccidia infection.
Eimeria infections have a serious impact on the physiological health of broiler chickens. The sporulated oocysts multiply in the cecum tissues and release a large number of sporozoites, which destroy intestinal epithelial cells and cause the blood vessels to rupture (Burrell et al., 2019; Madlala et al., 2021). Coccidia infection elicits a rapid response from the host immune system and some important genes play an important role in the inflammatory response to coccidial infection in chicken (Yu et al., 2021b). In this study, transcriptome sequencing of chicken cecal tissue after E. tenella infection was conducted to identify some important functional genes. A total of 434 genes were differentially expressed in cecal tissue, including immune-related genes such as GAMA, FABP1, F2RL1 and RSAD2. GAMA interacts with the band 3 receptor to promote erythrocyte invasion by malaria parasites (Lu et al., 2022). A Toxoplasma gondii ortholog of plasmodium GAMA contributes to parasite attachment and cell invasion (Huynh and Carruthers, 2016). F2RL1 can regulate M1 macrophage polarization and inflammatory responses through the FOXO1 signaling pathway (Chen et al., 2019). Furthermore, high expression of F2RL1 in the intestine has been associated with intestinal inflammatory and oncogenic processes (Ke et al., 2020). Therefore, these genes related to immune may be involved in the host’s immune response against coccidioides. Functional enrichment analysis of these differentially expressed genes showed that the TGF-beta and ErbB signaling pathways were significantly enriched. TGF-beta signaling pathway was involved in host’s immune tolerance and liver fibrosis during Echinococcus multilocularis infection (Wang et al., 2013). Moreover, TGF-β1-induced Smad signaling pathway played a critical role in the activation of HSCs during liver fibrosis induced by Clonorchis sinensis infection (Yan et al., 2015). Studies have shown that Cryptosporidium parvum maintains intracellular survival by activating the host cellular EGFR (ErbB1)-PI3K/Akt signaling pathway (Yang et al., 2023). Therefore, these pathways may also play an important role in the process of coccidia infection.
To unravel the interactions between the gut microbiota and the host transcriptome after E. tenella infection in chicken. In this study, Spearman correlation coefficient was used to analyze the association of transcriptome and metagenomics. The results showed that FABP1 was significantly associated with Prevotella pectinovora. FABP1 is a fatty acid binding gene that plays an important role in regulating intestinal fatty acid metabolic pathways. Studies have shown that FABP1 regulates key lipid-sensitive pathways in macrophages and thus participates in the intestinal inflammatory response (Levy et al., 2001). In addition, the expression of the FABP1 gene is downregulated in poultry infected with necrotizing enteritis (NE), which in turn reduces fatty acid utilization and intestinal immunity (Gharib-Naseri et al., 2021). Therefore, the results of our study suggest that FABP1 and Prevotella pectinovora may be jointly involved in host immunity and metabolism-related processes following coccidial infection. In addition, RSAD2 is significantly associated with Butyricicoccus and Colidextribacter. RSAD2 is necessary for dendritic cell maturation (Jang et al., 2018). Moreover, RSAD2 can regulate B cell activity through the NF-kB signaling pathway (Zhu et al., 2021). Butyricicoccus is also an important intestinal probiotic that has a positive effect in intestinal inflammatory diseases. Notably, SERPINB1 was only significantly positively correlated with Prevotella pectinovora, but had a significant negative correlation with the other 8 microbiota. SerpinB1, a protease inhibitor and neutrophil survival factor, is widely involved in the inflammatory response of diseases (Benarafa, 2011; Gong et al., 2011). Research by Zhao et al. (2014) shows that SerpinB1 regulates homeostatic expansion of IL-17+ γδ and CD4+ Th17 cells. The results of this study suggest that the expression of SerpinB1 may affect the changes of cecal microbiota during coccidia infection. LOC107052718, an unknown gene, was significantly positively correlated with 11 cecal microbiota. Although there is no report on the function of the LOC107052718 gene, our results reveal that LOC107052718 may play an important role in the infection of E. tenella, and the specific function and regulatory mechanism need further study.
In conclusion, the results of this study indicated that E. tenella infection significantly altered the abundance of chicken cecal microbiota and also affected the regulation of host genes. Moreover, we also revealed some potential relationships between differentially expressed genes and microbiota after coccidia infection. These results provide a reference for our understanding of the impact of coccidiosis on the host and the relationship between cecal microbiota and the host gene expression.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: NCBI; PRJNA678759; PRJNA943112.
Ethics statement
The animal study was reviewed and approved by the Animal Welfare Committee of Yangzhou University (permit number: SYXK (Su) IACUC 2012-0029).
Author contributions
HY and GD designed the study and wrote the manuscript. HY, QW and JT performed the experiments, collected the data. HY and QW analyzed the transcriptome data and prepared the figures. LD provided technical support for the sequencing details. KX, TZ, GZ, HW and ZZ reviewed the manuscript. All authors contributed to the article and approved the submitted version.
Funding
This research was supported by the Jiangsu Agricultural Industry Technology System (JATS[2023]490), the Postgraduate Research & Practice Innovation Program of Jiangsu Province (KYCX22_3528, Yangzhou University), the Yangzhou University International Academic Exchange Foundation, the Priority Academic Program Development of Jiangsu Higher Education Institutions (PAPD), and the China Agriculture Research System (CARS-41-G23).
Acknowledgments
The authors would like to thank all members of this work for their advice and technical assistance.
Conflict of interest
Hongsheng Wang is employed by Jiangsu Jinghai Poultry Group Co., Ltd.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2023.1191939/full#supplementary-material
References
Anders, S., Pyl, P. T., Huber, W. (2015). HTSeq–a Python framework to work with high-throughput sequencing data. Bioinformatics 31, 166–169. doi: 10.1093/bioinformatics/btu638
Baldwin, S., Hughes, R. J., Van, T. T. H., Moore, R. J., Stanley, D. (2018). At-Hatch administration of probiotic to chickens can introduce beneficial changes in gut microbiota. PloS One 13 (3), e0194825. doi: 10.1371/journal.pone.0194825
Benarafa, C. (2011). The SerpinB1 knockout mouse a model for studying neutrophil protease regulation in homeostasis and inflammation. Methods Enzymol. 499, 135–148. doi: 10.1016/B978-0-12-386471-0.00007-9
Blake, D. P., Knox, J., Dehaeck, B., Huntington, B., Rathinam, T., Ravipati, V., et al. (2020). Re-calculating the cost of coccidiosis in chickens. Vet. Res. 51 (1), 115. doi: 10.1186/s13567-020-00837-2
Bolger, A. M., Lohse, M., Usadel, B. (2014). Trimmomatic: a flexible trimmer for illumina sequence data. Bioinformatics 30 (15), 2114–2120. doi: 10.1093/bioinformatics/btu170
Buchfink, B., Xie, C., Huson, D. H. (2015). Fast and sensitive protein alignment using DIAMOND. Nat. Methods 12 (1), 59–60. doi: 10.1038/nmeth.3176
Burrell, A., Tomley, F. M., Vaughan, S., Marugan-Hernandez, V. (2019). Life cycle stages, specific organelles and invasion mechanisms of eimeria species. Parasitology 147 (3), 263–278. doi: 10.1017/S0031182019001562
Chen, L., Gao, B., Zhang, Y., Lu, H., Li, X., Pan, L., et al. (2019). PAR2 promotes M1 macrophage polarization and inflammation via FOXO1 pathway. J. Cell. Biochem. 120 (6), 9799–9809. doi: 10.1002/jcb.28260
Chen, H., Zhao, X., Zhao, G., Huang, H., Li, H., Shi, C., et al. (2020). Dissection of the cecal microbial community in chickens after eimeria tenella infection. Parasit. Vectors. 13 (1), 56. doi: 10.1186/s13071-020-3897-6
Clavijo, V., Flórez, M. J. V. (2018). The gastrointestinal microbiome and its association with the control of pathogens in broiler chicken production: a review. Poult. Sci. 97 (3), 1006–1021. doi: 10.3382/ps/pex359
Clemente, J. C., Ursell, L. K., Parfrey, L. W., Knight, R. (2012). The impact of the gut microbiota on human health: an integrative view. Cell 148 (6), 1258–1270. doi: 10.1016/j.cell.2012.01.035
Fernandez, E. M., Valenti, V., Rockel, C., Hermann, C., Pot, B., Boneca, I. G., et al. (2011). Anti-inflammatory capacity of selected lactobacilli in experimental colitis is driven by NOD2-mediated recognition of a specific peptidoglycan-derived muropeptide. Gut 60 (8), 1050–1059. doi: 10.1136/gut.2010.232918
Gharib-Naseri, K., Heras-Saldana, S. L., Kheravii, S., Qin, L., Wang, J., Wu, S. (2021). Necrotic enteritis challenge regulates peroxisome proliferator-1 activated receptors signaling and β-oxidation pathways in broiler chickens. Anim. Nutr. 7 (1), 239–251. doi: 10.1016/j.aninu.2020.08.003
Gong, D., Farley, K., White, M., Hartshorn, K. L., Benarafa, C., Remold-O'Donnell, E. (2011). Critical role of serpinB1 in regulating inflammatory responses in pulmonary influenza infection. J. Infect. Dis. 204 (4), 592–600. doi: 10.1093/infdis/jir352
Hegazy, A. N., West, N. R., Stubbington, M. J. T., Wendt, E., Suijker, K. I. M., Datsi, A., et al. (2017). Circulating and tissue-resident CD4(+) T cells with reactivity to intestinal microbiota are abundant in healthy individuals and function is altered during inflammation. Gastroenterology 153 (5), 1320–1337. doi: 10.1053/j.gastro.2017.07.047
Huynh, M., Carruthers, V. B. (2016). A Toxoplasma gondii Ortholog of Plasmodium GAMA contributes to parasite attachment and cell invasion. mSphere. 1 (1), e00012–16. doi: 10.1128/mSphere.00012-16
Iebba, V., Guerrieri, F., Gregorio, V. D., Levrero, M., Gagliardi, A., Santangelo, F., et al. (2018). Combining amplicon sequencing and metabolomics in cirrhotic patients highlights distinctive microbiota features involved in bacterial translocation, systemic inflammation and hepatic encephalopathy. Sci. Rep. 8 (1), 8210. doi: 10.1038/s41598-018-26509-y
Jang, J., Lee, J., Jung, Na., Choi, S., Park, S., Yoo, J., et al. (2018). Rsad2 is necessary for mouse dendritic cell maturation via the IRF7-mediated signaling pathway. Cell. Death. Dis. 9 (8), 823. doi: 10.1038/s41419-018-0889-y
Jebessa, E., Guo, L., Chen, X., Bello, S. F., Cai, B., Girma, M., et al. (2022). Influence of eimeria maxima coccidia infection on gut microbiome diversity and composition of the jejunum and cecum of indigenous chicken. Front. Immunol. 13. doi: 10.3389/fimmu.2022.994224
Ke, Z., Wang, C., Wu, T., Wang, W., Yang, Y., Dai, Y. (2020). 3PAR2 deficiency enhances myeloid cell-mediated immunosuppression and promotes colitis-associated tumorigenesis. Cancer Lett. 469, 437–446. doi: 10.1016/j.canlet.2019.11.015
Kim, W. H., Jeong, J., Park, A. R., Yim, D., Kim, Y. H., Kim, K. D., et al. (2012). Chicken IL-17F: identification and comparative expression analysis in eimeria-infected chickens. Dev. Comp. Immunol. 38 (3), 401–409. doi: 10.1016/j.dci.2012.08.002
Kim, D., Langmead, B., Salzberg, S. L. (2015). HISAT: a fast spliced aligner with low memory requirements. Nat. Methods 12 (4), 357–360. doi: 10.1038/nmeth.3317
Larsen, J. M. (2017). The immune response to prevotella bacteria in chronic inflammatory disease. Immunology 151 (4), 363–374. doi: 10.1111/imm.12760
Levy, E., Ménard, D., Delvin, E., Stan, S., Mitchell, G., Lambert, M., et al. (2001). The polymorphism at codon 54 of the FABP2 gene increases fat absorption in human intestinal explants. J. Biol. Chem. 276 (43), 39679–39684. doi: 10.1074/jbc.M105713200
Li, D., Liu, C. M., Luo, R., Sadakane, K., Lam, T. (2015). MEGAHIT: an ultra-fast single-node solution for large and complex metagenomics assembly via succinct de bruijn graph. Bioinformatics 31 (10), 1674–1676. doi: 10.1093/bioinformatics/btv033
Li, Z., Wang, W., Liu, D., Guo, Y. (2017). Effects of lactobacillus acidophilus on gut microbiota composition in broilers challenged with clostridium perfringens. PloS One 12 (11), e0188634. doi: 10.1371/journal.pone.0188634
Lu, J., Chu, R., Yin, Y., Yu, H., Xu, Q., Yang, B., et al. (2022). Glycosylphosphatidylinositol-anchored micronemal antigen (GAMA) interacts with the band 3 receptor to promote erythrocyte invasion by malaria parasites. J Biol Chem. 298 (4), 101765. doi: 10.1016/j.jbc.2022.101765
Madlala, T., Okpeku, M., Adeleke, M. A. (2021). Understanding the interactions between eimeria infection and gut microbiota, towards the control of chicken coccidiosis: a review. Parasite 28, 48. doi: 10.1051/parasite/2021047
Min, W., Kim, W. H., Lillehoj, E. P., Lillehoj, H. S. (2013). Recent progress in host immunity to avian coccidiosis: IL-17 family cytokines as sentinels of the intestinal mucosa. Dev. Comp. Immunol. 41 (3), 418–428. doi: 10.1016/j.dci.2013.04.003
Mohamadzadeh, M., Pfeiler, E. A., Brown, J. B., Zadeh, M., Gramarossa, M., Managlia, E., et al. (2011). Regulation of induced colonic inflammation by lactobacillus acidophilus deficient in lipoteichoic acid. Proc. Natl. Acad. Sci. U. S. A. 108 (Suppl 1), 4623–4630. doi: 10.1073/pnas.1005066107
Nie, K., Ma, K., Luo, W., Shen, Z., Yang, Z., Xiao, M., et al. (2021). Roseburia intestinalis: a beneficial gut organism from the discoveries in genus and species. Front. Cell. Infect. Microbiol. 11. doi: 10.3389/fcimb.2021.757718
Parker, B. J., Wearsch, P. A., Veloo, A. C. M., Rodriguez-Palacios, A. (2020). The genus alistipes: gut bacteria with emerging implications to inflammation, cancer, and mental health. Front. Immunol. 11. doi: 10.3389/fimmu.2020.00906
Pender, C. M., Kim, S., Potter, T. D., Ritzi, M. M., Young, M., Dalloul, R. A. (2016). Effects of in ovo supplementation of probiotics on performance and immunocompetence of broiler chicks to an eimeria challenge. Benef. Microbes 7 (5), 699–705. doi: 10.3920/BM2016.0080
Polansky, O., Sekelova, Z., Faldynova, M., Sebkova, A., Sisak, F., Rychlik, I. (2015). Important metabolic pathways and biological processes expressed by chicken cecal microbiota. Appl. Environ. Microbiol. 82 (5), 1569–1576. doi: 10.1128/AEM.03473-15
Roberts, A., Trapnell, C., Donaghey, J., Rinn, J. L., Pachter, L. (2011). Improving RNA-seq expression estimates by correcting for fragment bias. Genome Biol. 12 (3), R22. doi: 10.1186/gb-2011-12-3-r22
Rubio, L. A. (2019). Possibilities of early life programming in broiler chickens via intestinal microbiota modulation. Poult. Sci. 98 (2), 695–706. doi: 10.3382/ps/pey416
Segata, N., Izard, J., Waldron, L., Gevers, D., Miropolsky, L. (2011). Garrett, W. S., et al. Metagenomic biomarker discovery and explanation. Genome Biol. 12 (6), R60. doi: 10.1186/gb-2011-12-6-r60
Smoot, M. E., Ono, K., Ruscheinski, J., Wang, P., Ideker, T. (2011). Cytoscape 2.8: new features for data integration and network visualization. Bioinformatics 27 (3), 431–432. doi: 10.1093/bioinformatics/btq675
Stanley, D., Wu, S., Rodgers, N., Swick, R. A., Moore, R. J. (2014). Differential responses of caecal microbiota to fishmeal, eimeria and clostridium perfringens in a necrotic enteritis challenge model in chickens. PloS One 9 (8), e104739. doi: 10.1371/journal.pone.0104739
Tamanai-Shacoor, Z., Smida, I., Bousarghin, L., Loreal, O., Meuric, V., Fong, S., et al. (2017). Roseburia spp.: a marker of health? Future. Microbiol. 12, 157–170. doi: 10.2217/fmb-2016-0130
Teame, T., Wang, A., Xie, M., Zhang, Z., Yang, Y., Ding, Q., et al. (2020). Paraprobiotics and postbiotics of probiotic lactobacilli, their positive effects on the host and action mechanisms: a review. Front. Nutr. 7. doi: 10.3389/fnut.2020.570344
Trapnell, C., William, B. A., Pertea, G., Mortazavi, A., Kwan, G., Baren, M. J., et al. (2010). Transcript assembly and quantification by RNA-seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 28 (5), 511–515. doi: 10.1038/nbt.1621
Waite, D. W., Taylor, M. W. (2015). Exploring the avian gut microbiota: current trends and future directions. Front. Microbiol. 6. doi: 10.3389/fmicb.2015.00673
Wang, J., Zhang, C., Wei, X., Blagosklonov, O., Lv, G., Lu, X., et al. (2013). TGF-β and TGF-β/Smad signaling in the interactions between echinococcus multilocularis and its hosts. PloS One 8 (2), e55379. doi: 10.1371/journal.pone.0055379
Wickramasuriya, S. S., Park, I., Lee, Y., Richer, L. M., Przybyszewski, C., Gay, C. G., et al. (2023). Orally delivered bacillus subtilis expressing chicken NK-2 peptide stabilizes gut microbiota and enhances intestinal health and local immunity in coccidiosis-infected broiler chickens. Poult. Sci. 102 (5), 102590. doi: 10.1016/j.psj.2023.102590
Yan, C., Wang, L., Li, B., Zhang, B., Zhang, B., Wang, Y., et al. (2015). The expression dynamics of transforming growth factor-β/Smad signaling in the liver fibrosis experimentally caused by clonorchis sinensis. Parasit. Vectors. 48, 70. doi: 10.1186/s13071-015-0675-y
Yang, H., Zhang, M., Wang, X., Gong, P., Zhang, N., Zhang, X., et al. (2023). Cryptosporidium parvum maintains intracellular survival by activating the host cellular EGFR-PI3K/Akt signaling pathway. Mol. Immunol. 154, 69–79. doi: 10.1016/j.molimm.2023.01.002
Yeoman, C. J., Chia, N., Patricio, J., Maksim, S., Goldenfeld, N. D., White, B. A. (2012). The microbiome of the chicken gastrointestinal tract. anim. Health Res. Rev. 13 (1), 89–99. doi: 10.1017/S1466252312000138
Yu, H., Mi, C., Wang, Q., Dai, G., Zhang, T., Zhang, G., et al. (2022). The lnRNA profiling reveals lncRNA BTN3A2 inhibits the host inflammatory response to eimeria tenella infection in chickens. Front. Immunol. 13. doi: 10.3389/fimmu.2022.891001
Yu, H., Mi, C., Wang, Q., Zou, W., Dai, G., Zhang, T., et al. (2021a). Comprehensive analyses of circRNA expression profiles and function prediction in chicken cecums after eimeria tenella infection. Front. Cell. Infect. Microbiol. 11. doi: 10.3389/fcimb.2021.628667
Yu, H., Zou, W., Mi, C., Wang, Q., Dai, G., Zhang, T., et al. (2021b). Research note: expression of T cell-related cytokines in chicken cecal and spleen tissues following eimeria tenella infection in vivo. Poult. Sci. 100 (7), 101161. doi: 10.1016/j.psj.2021.101161
Yu, H., Zou, W., Wang, X., Dai, G., Zhang, T., Zhang, G., et al. (2020). Research note: correlation analysis of interleukin-6, interleukin-8, and CC motif chemokine ligand 2 gene expression in chicken spleen and cecal tissues after eimeria tenella infection in vivo. Poult. Sci. 99 (3), 1326–1331. doi: 10.1016/j.psj.2019.10.071
Zaky, A., Glastras, S. J., Wong, M. Y. W., Pollock, C. A., Saad, S. (2021). The role of the gut microbiome in diabetes and obesity-related kidney disease. Int. J. Mol. Sci. 22 (17), 9641. doi: 10.3390/ijms22179641
Zhao, P., Hou, L., Farley, K., Sundrud, M. S., Remold-O'Donnell, E. (2014). SerpinB1 regulates homeostatic expansion of IL-17+ γδ and CD4+ Th17 cells. J. Leukoc. Biol. 95 (3), 521–530. doi: 10.1189/jlb.0613331
Zhao, X., Zhong, X., Liu, X., Wang, X., Gao, X. (2021). Therapeutic and improving function of lactobacilli in the prevention and treatment of cardiovascular-related diseases: a novel perspective from gut microbiota. Front. Nutr. 8. doi: 10.3389/fnut.2021.693412
Zhou, B., Jia, L., Wei, S., Ding, H., Yang, J., Wang, H. (2020). Effects of eimeria tenella infection on the barrier damage and microbiota diversity of chicken cecum. Poult. Sci. 99 (3), 1297–1305. doi: 10.1016/j.psj.2019.10.073
Keywords: chicken, E. tenella, cecal microbiome, transcriptome, integrated analysis
Citation: Yu H, Wang Q, Tang J, Dong L, Dai G, Zhang T, Zhang G, Xie K, Wang H and Zhao Z (2023) Comprehensive analysis of gut microbiome and host transcriptome in chickens after Eimeria tenella infection. Front. Cell. Infect. Microbiol. 13:1191939. doi: 10.3389/fcimb.2023.1191939
Received: 23 March 2023; Accepted: 18 May 2023;
Published: 06 June 2023.
Edited by:
Luis-Miguel Ortega-Mora, Complutense University of Madrid, SpainReviewed by:
Po-Yu Liu, National Sun Yat-sen University, TaiwanMusa Hassan, University of Edinburgh, United Kingdom
Samiru Sudharaka Wickramasuriya, Agricultural Research Service (USDA), United States
Copyright © 2023 Yu, Wang, Tang, Dong, Dai, Zhang, Zhang, Xie, Wang and Zhao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guojun Dai, ZGFpZ2pAeXp1LmVkdS5jbg==