 Marte Glambek1,2*
Marte Glambek1,2* Morten Kjos3
Morten Kjos3 Marita T. Mårli3Zhian Salehian3
Marita T. Mårli3Zhian Salehian3 Steinar Skrede1,2Audun Sivertsen4
Steinar Skrede1,2Audun Sivertsen4 Bård R. Kittang2,5
Bård R. Kittang2,5 Oddvar Oppegaard1,2
Oddvar Oppegaard1,2- 1Department of Medicine, Haukeland University Hospital, Bergen, Norway
- 2Department of Clinical Science, Faculty of Medicine, University of Bergen, Bergen, Norway
- 3Faculty of Chemistry, Biotechnology and Food Science, Norwegian University of Life Sciences, Ås, Norway
- 4Department of Microbiology, Haukeland University Hospital, Bergen, Norway
- 5Department of Internal Medicine, Haraldsplass Deaconess Hospital, Bergen, Norway
Background: Streptococcus dysgalactiae (SD) is a potent pathogen associated with infections in a broad range of host species. Notably, a substantial proportion of SD isolates exhibit reduced susceptibility to tetracycline but lack identifiable resistance determinants. In the present study, we wanted to explore the genetic basis for this low-grade resistance to tetracycline.
Methods: Genome-wide association studies were performed on a collection of 407 SD genomes to identify potential novel resistance determinants. Two strains of SD, belonging to each of the subspecies dysgalactiae and equisimilis were used for mutagenesis. Natural transformation was exploited to knock out resistance gene candidates, and the resultant mutants were compared with their respective wildtypes regarding susceptibility to tetracycline, doxycycline, minocycline, tigecycline, erythromycin, gentamicin, clindamycin and ciprofloxacin.
Results: We identified a two gene operon, herein designated trexAB, significantly associated with reduced susceptibility to tetracycline. The proteins encoded by the operon were predicted in silico to constitute a heterodimeric efflux transporter. The knockout of trexAB led to a 16- to 32-fold reduction in minimum inhibitory concentration (MIC) for tetracycline and a 4-fold reduction in MIC for tigecycline in the investigated strains. No differences between mutants and wildtypes were observed for other antibiotics included in the test panel. Whole genome alignment of mutants and their respective wildtypes revealed no differences other than the expected differences caused by the knockout.
Conclusion: We have characterized a novel operon causing low-grade resistance to tetracycline in SD. The MIC distribution of trexAB-positive isolates is intersected by the current EUCAST susceptibility breakpoint, and our findings are relevant for future revisions and determinations of adequate breakpoints for tetracycline in S. dysgalactiae.
Introduction
Tetracyclines were among the first broad-spectrum antibiotics discovered. The limited number of side-effects together with the availability of oral formulations, made tetracyclines attractive choices in both clinical and agricultural settings. The tetracyclines are divided into three different generations, where the first generation comprises tetracycline, oxytetracycline and chlortetracycline, the second generation includes minocycline and doxycycline, and the glycylcycline tigecycline constitutes the third generation (Thaker et al., 2010).
The tetracyclines inhibit bacterial protein synthesis by binding to the bacterial ribosome, disturbing the bacteria’s ability to synthesize proteins. Bacterial resistance against tetracycline occurs by different mechanisms, which mainly fit into the categories of efflux systems, ribosomal protection and drug destruction (Thaker et al., 2010). The more than 60 unique tetracycline resistance determinants characterized to date, indicate that drug efflux is the main resistance strategy in gram-negative bacteria, whereas ribosomal protection is the most common mechanism in gram-positive bacteria (Roberts, 2024). To be defined as a unique resistance gene in this setting, the sequence homology to genes of known function must be lower than 79% at the amino acid level (Levy et al., 1999).
We recently characterized antimicrobial susceptibility patterns of Streptococcus dysgalactiae (SD), a gram-positive pathogen known to infect a broad range of host species (Glambek et al., 2024). We explored resistance in a One Health perspective, including both S. dysgalactiae subspecies dysgalactiae (SDSD) associated with bovine and ovine infections, and S. dysgalactiae subspecies equisimilis (SDSE) predominantly targeting other animals and humans. Surprisingly, we observed a trimodal distribution of minimum inhibitory concentrations (MIC) values to tetracycline, and a relative high proportion of low-grade tetracycline resistant isolates without an identifiable genetic resistance determinant.
The central cluster of SD isolates had MIC values ranging between 0.5 and 4 μg/ml (herein referred to as the transition zone), and thus encircled the EUCAST breakpoint between sensitive and resistant, suggestive of low-grade phenotypic resistance. Canonical tet-genes were identified in nearly all isolates with MIC values above the transition zone, whereas isolates in the central cluster generally did not encode identifiable resistance genes.
In the present study, we explore the underlying mechanism for this low-grade resistance using genome-wide association studies and mutant construction. We report the identification of a novel two-gene operon associated with the low-grade resistance phenotype, likely encoding proteins that together function as an ABC efflux transporter.
Materials and methods
Bacterial isolates
A collection of 407 SD strains procured from human and animal associated infections in Norway during 2018–2019 was investigated in this study. The isolates have previously been whole genome sequenced, examined for antibiotic resistance genes and susceptibility tested for tetracycline (Glambek et al., 2024).
In accordance with the phenotypic definition proposed by Vieira et al (Vieira et al., 1998), we defined SDSD in silico as genomes harboring the Lancefield group C-antigen operon, lacking the streptolysin S operon (corresponding to an α- or nonhemolytic reaction on blood agar), and lacking the streptokinase gene (inferring that streptokinase activity on human plasminogen does not occur). All other genomes were classified as SDSE (Glambek et al., 2024).
In silico analysis
DBGWAS was used to search for genetic variants associated with low-grade tetracycline resistance (Jaillard et al., 2018). Strains harboring known tetracycline resistance genes were excluded from the analyses, and minimum inhibitory concentration (MIC) level was used as phenotype-indicator, with a MIC phenoThreshold of 0.5 μg/ml. Genetic variants identified by DBGWAS were mapped to genomic location and inspected using the Geneous Prime v 2024.0 software. The predicted function of annotated genes at matching loci was evaluated by screening for conserved functional domains using CD search (Wang et al., 2022) and InterProScan 102.0 (Blum et al., 2024), with default settings. Searches for homologue genes and proteins to our candidate tetracycline resistance genes were done using megaBLASTn and BLASTp, respectively.
A core genome single-nucleotide polymorphism phylogeny was generated by CSI Phylogeny at the Center for Genomic Epidemiology (Kaas et al., 2014) using default settings and the SDSE type strain NCTC13762 as a reference. The resulting maximum likelihood phylogenetic tree was visualized and annotated using the Interactive Tree of Life platform, iTol v6 (Letunic and Bork, 2021).
Screening of global collection of SD genomes
For comparative analysis, we downloaded a global collection of SD genomes from published epidemiological studies available from GenBank and PubMLST. These included human associated isolates collected in Australia (Xie et al., 2024), Canada (Lother et al., 2017) and Japan (Shinohara et al., 2023), bovine isolates from Canada (Vélez et al., 2017), as well as swine and horse isolates from Italy, US and Portugal (Pinho et al., 2016; Cinthi et al., 2023). The genomes were de novo assembled using SPAdes v 5.14 (Bankevich et al., 2012), annotated using RAST v 1.073 (Aziz et al., 2008), and screened for the presence of candidate genes using the Geneious Prime v 2024.0 software.
Growth conditions and susceptibility testing
In the knockout experiments, bacteria were grown in airtight tubes in C-medium (Lacks and Hotchkiss, 1960) or on brain heart infusion (BHI) agar plates at 37 °C in 10% CO2. For selection of knock out mutants, kanamycin was added to the BHI agar to a final concentration of 400 μg/ml.
Knockout mutants and their respective wild types were examined for susceptibility to tetracycline, doxycycline, minocycline, tigecycline, erythromycin, gentamicin, clindamycin and ciprofloxacin according to the NORM protocol (NORM, 2019). Briefly, isolates were plated on Mueller-Hinton agar supplemented with defibrinated horse blood and β-NAD. MIC-levels were determined using MIC-gradient strips.
Synthetic peptides
Synthetic nature quorum sensing peptide pheromones, XIP1 (aa: EFDWWNLG) and XIP2 (aa: QVDWWRL) were purchased from Thermo Scientific.
Construction of deletion fragment and deletion of trexAB by natural transformation
A linear DNA fragment for homologous recombination to delete trexAB was assembled using overlap extension PCR (Higuchi et al., 1988). Amplicons of approximately 2 kb length of flanking sequences to the trexAB-operon in addition to a core sequence consisting of the so-called Janus cassette (Sung et al., 2001), encoding a kanamycin resistance cassette and a rpsL-allele, were made by PCR and extracted from agarose gel. The trexAB upstream and downstream fragments were merged to the 5’ end and 3’ end of the Janus cassette, respectively, making a DNA construct to create genetic knockouts (Higuchi et al., 1988). Primers used are listed in Table 1.

Table 1. Primers.
Two SD isolates were selected for functional studies of the trexAB operon, the human associated isolate iSDSE_NORM6 of subspecies equisimilis, and the bovine associated isolate SDSD24 of subspecies dysgalactiae. These strains contained an intact trexAB operon and displayed low grade resistance to tetracycline, without possessing any validated tetracycline resistance genes. These strains also possessed a complete and intact apparatus for competence and natural transformation (Mårli et al., 2024).
The natural transformation procedure was adapted from the protocol described by Mårli and co-workers (Mårli et al., 2024). Briefly, overnight cultures of isolates to be transformed were diluted in C-medium to an initial OD600 of 0.05 for further incubation until reaching OD600 0.2. The cultures were again diluted to OD600 0.03 and finally grown to OD600 0.05 before approximately 400 ng of DNA-construct and 250 ng of XIP1 (iSDSE_NORM6) or XIP2 (SDSD24) was added to 1 ml culture. Cultures were further incubated at 37°C for 3–4 hours and then plated on BHI agar containing 400 μg/ml kanamycin and grown overnight for selection of kanamycin resistant mutants. Cultures without the added DNA-construct, were used as negative controls.
Whole genome sequencing
Genomic DNA was purified using MagNA Pure extraction kit (Roche Life Science). Whole genome sequencing of knockout mutant strains was performed at Haukeland University Hospital on an Illumina 4,000 HiSeq system to produce 150 bp paired end reads, as previously described (Oppegaard et al., 2017). The genomes of mutant and wildtype strains were aligned with Mauve, and manually compared for insertion, deletion, and mutation events.
Results
Identification of tetracycline resistance determinants
Genome-wide associating studies with DBGWAS identified fifteen genetic regions with at least one polymorphism significantly associated with variations in MIC-level. Only two regions were associated with an increased MIC-level. One of these was a single C/G synonymous mutation in the YidA sugar phosphatase gene, which was deemed unlikely to confer tetracycline resistance. The other was 149 overlapping significant hits constituting a two-gene operon of unknown function. This operon was observed in 167 of the 172 strains with MIC values in the transition zone, but only in 13 of the 184 strains with lower MIC values (Figure 1), among which 5 strains contained an intact operon, and the remaining 8 strains had one or both genes truncated (Supplementary Table).
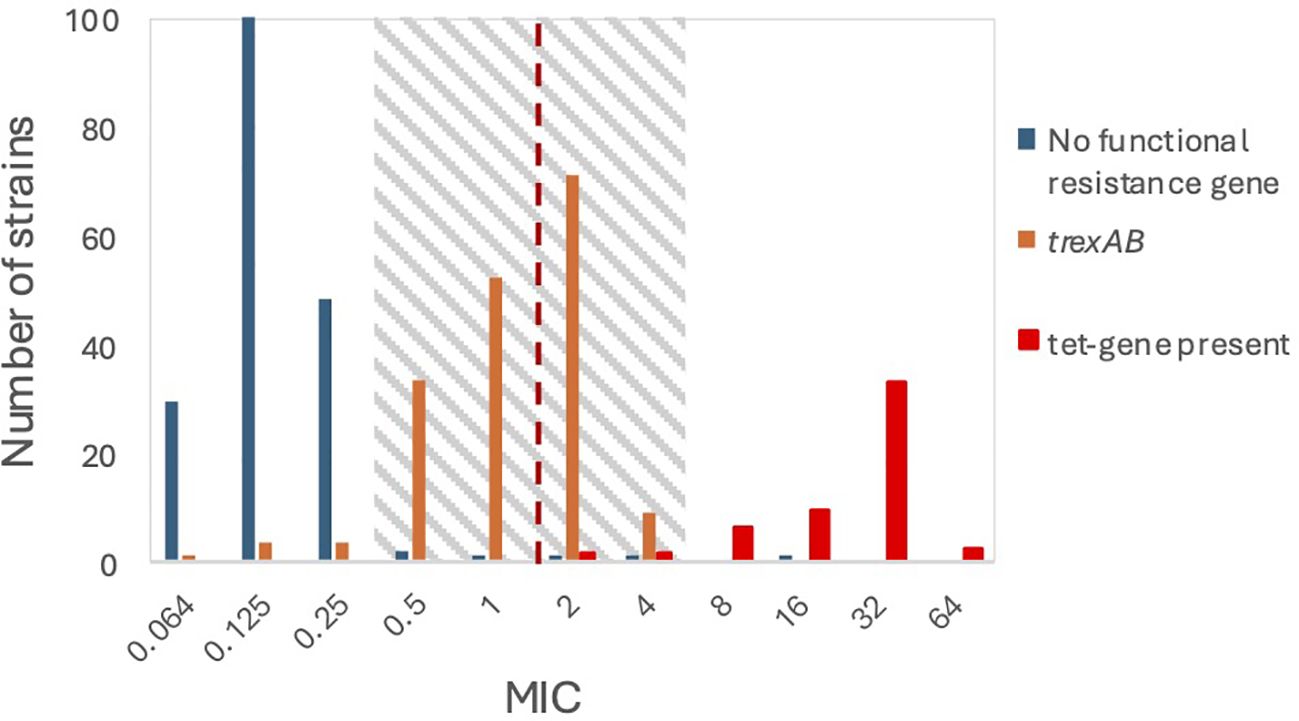
Figure 1. Susceptibility in relation to genes known or presumed to be associated with resistance to tetracycline. The dotted line represents the EUCAST breakpoint between susceptibility and resistance. The MIC transition zone is highlighted with grey shading.
We compared the products of the two novel genes to previously characterized tetracycline resistance determinants using BLASTp. A low-level homology to TetA(46) and TetB(46) was detected (55% and 59% pairwise amino acid sequence identity, respectively), a tetracycline efflux pump encoded by a two gene operon in Streptococcus australis (Warburton et al., 2013). We thus decided to further explore the potential role in tetracycline resistance of our newly discovered operon, herein designated trexAB (Tetracycline Resistance EffluX, gene A and gene B).
Genetic characterization of the trexAB operon
The operon comprised the trexA gene (1722 base pairs) and trexB gene (1743 base pairs). Both genes showed high interstrain homology within our collection of SD, with nucleotide sequence similarity ranging from 98 to 100%.
Screening the predicted proteins for conserved domains revealed that both TrexA and TrexB harbored MdlB superfamily domains (COG1132), which represent a group of well-characterized ABC-type multidrug transport systems. Typically, these systems are composed of two proteins constituting a dimer spanning the cell membrane, actively exporting toxic substances out of the cell, fueled by the energy generated from hydrolyzing ATP to ADP. In line with this, InterProScan detected the presence of several transmembrane regions gathered in a transmembrane domain (TMD) and an ABC transporter domain constituting the P-loop nucleoside triphosphate hydrolase domain in both TrexA and TrexB (Figure 2).
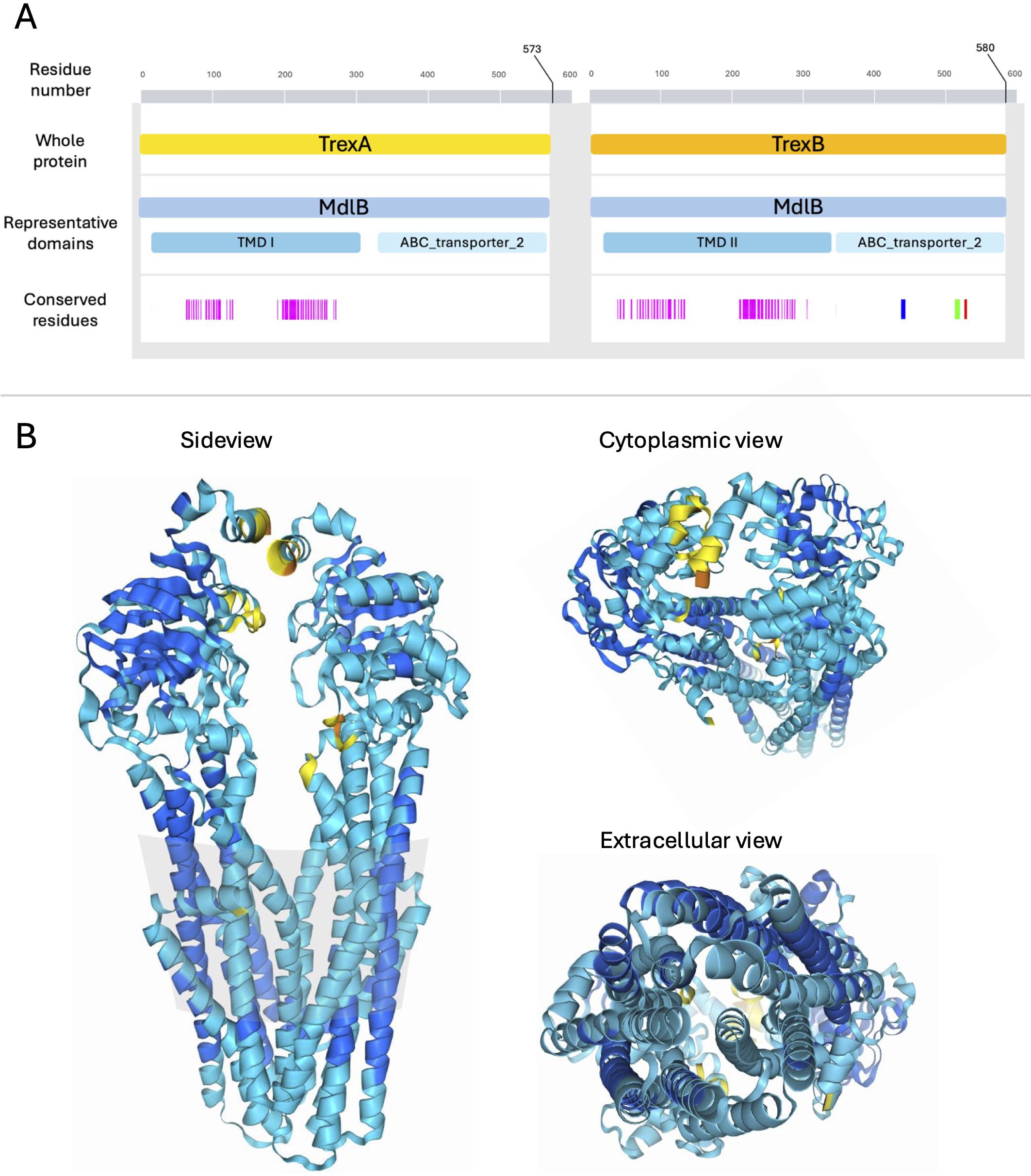
Figure 2. Domains and predicted structure of TrexAB. (A) The amino acid sequence of TrexA and TrexB where the numbering of amino acids in the primary sequence is marked in grey scale. Representative domains are highlighted in light blue shades. Conserved residues in TrexA and TrexB determined by InterPro-search representing the heterodimeric interface, the Walker A/P-loop motif, the ABC transporter signature motif, and the Walker B motif are highlighted in pink, blue, green and red, respectively. (B) Structures of the heterodimeric TrexAB predicted using Alpha Fold 3. The colors in this model represent per-atom prediction confidence where dark blue, light blue, yellow and orange represent very high, confident, low and very low accuracy, respectively.
Genomic context of trexAB
The trexAB operon had a conserved genomic location between an operon containing four genes of the mevalonate pathway and the s5nA-nucleotidase gene. In strains lacking trexAB, the same genomic location was found to be occupied by a three-gene operon of unknown function (Figure 3). The predicted proteins of these three genes all appear to be involved in signal transduction mechanisms, harboring the domains belonging to YjbM superfamily (COG2357), OmpR superfamily (COG0745) and BaeS superfamily (COG0642), respectively. All isolates harbored one of the two operons, and they were mutually exclusive. The genetic region was highly conserved, and we did not find any association between trexAB or the alternative operon and known conjugative mobile genetic elements. Differently, IS-elements were found immediately upstream or downstream of trexAB in 55 of the isolates.
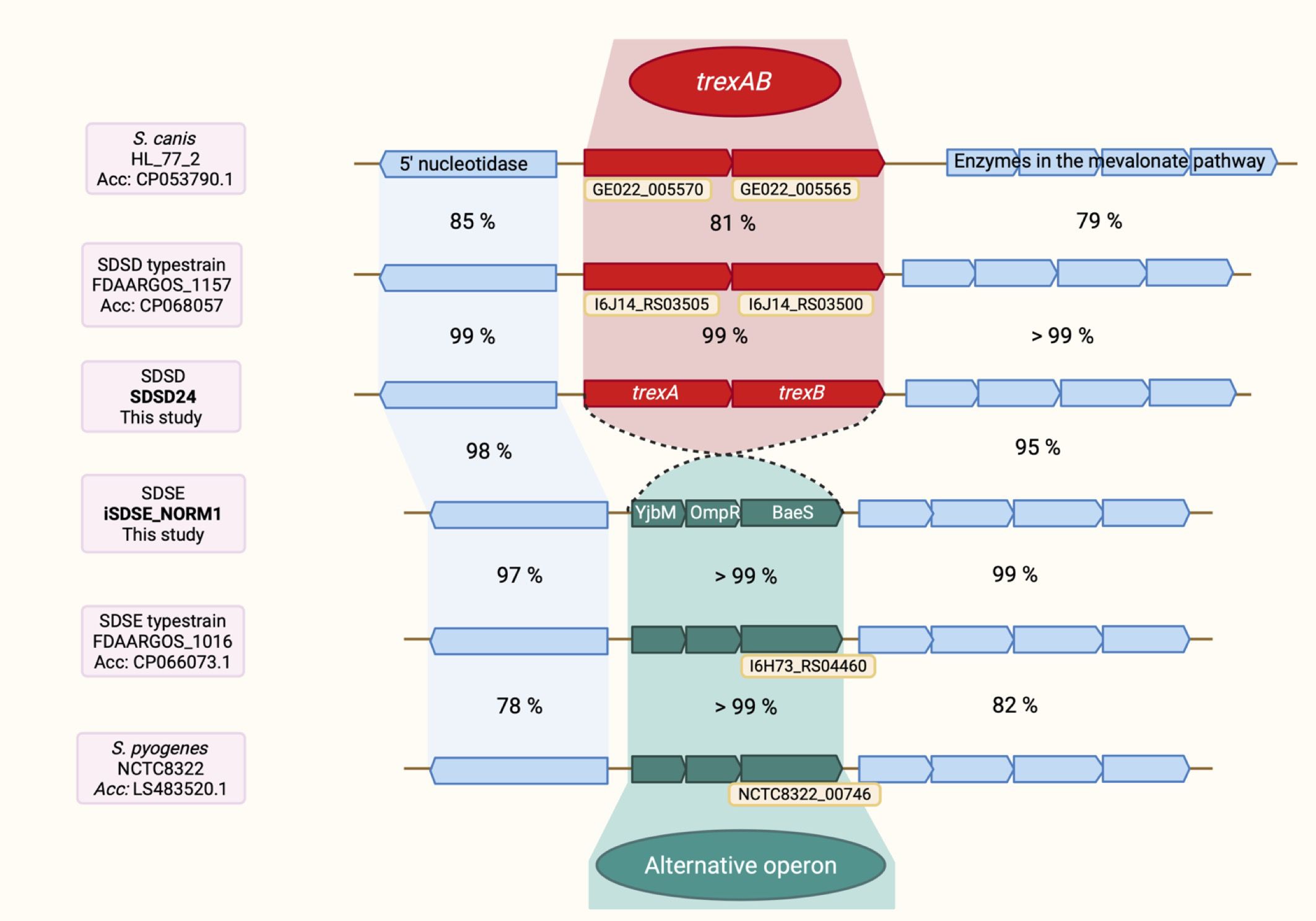
Figure 3. The chromosomal location of trexAB. trexAB has the closest homolog in S. canis, with a sequence identity of 81% at the nucleotide level. The alternative operon is found in S. pyogenes in addition to S. dysgalactiae. The sequence identity of the alternative operon between S. pyogenes and S. dysgalactiae is significantly higher than the sequence identity of trexAB and its closest homolog in S. canis. The close resemblance of the alternative operon in S. pyogenes and S. dysgalactiae is also in contrast with the flanking regions, which has a much lower sequence identity between the two species. Corresponding ProteinID (to locus tags): “TrexA”: QKG75729.1 (GE022_005570), QQT04366.1 (I6J14_03505); “TrexB”: QKG75728.1 (GE022_005565), QQT04365.1 (I6J14_03500); Alt. operon, 3. gene: QQC56251.1 (I6H73_04460), SRX87322.1 (NCTC8322_00746). Created with Biorender.com, https://app.biorender.com/illustrations/6836d76968fa6ae428cd3cb3.
Distribution of trexAB
The trexAB operon was distributed among SD collected from all ecovars. Notably, all strains (83 SDSD and 2 SDSE) originating from cattle and sheep possessed this operon, as did all isolates (SDSE) from pigs and horses, while only 69 out of 274 isolates (SDSE) associated with humans harbored trexAB. The operon was limited to specific phylogenetic clades of human associated SD, predominantly belonging to multilocus sequence type 29 clonal complex. Among the SD isolates (SDSE) procured from dogs, four of 20 isolates were lacking trexAB. However, phylogenetically these four isolates clustered with human-associated SD isolates also lacking trexAB (Figure 4).
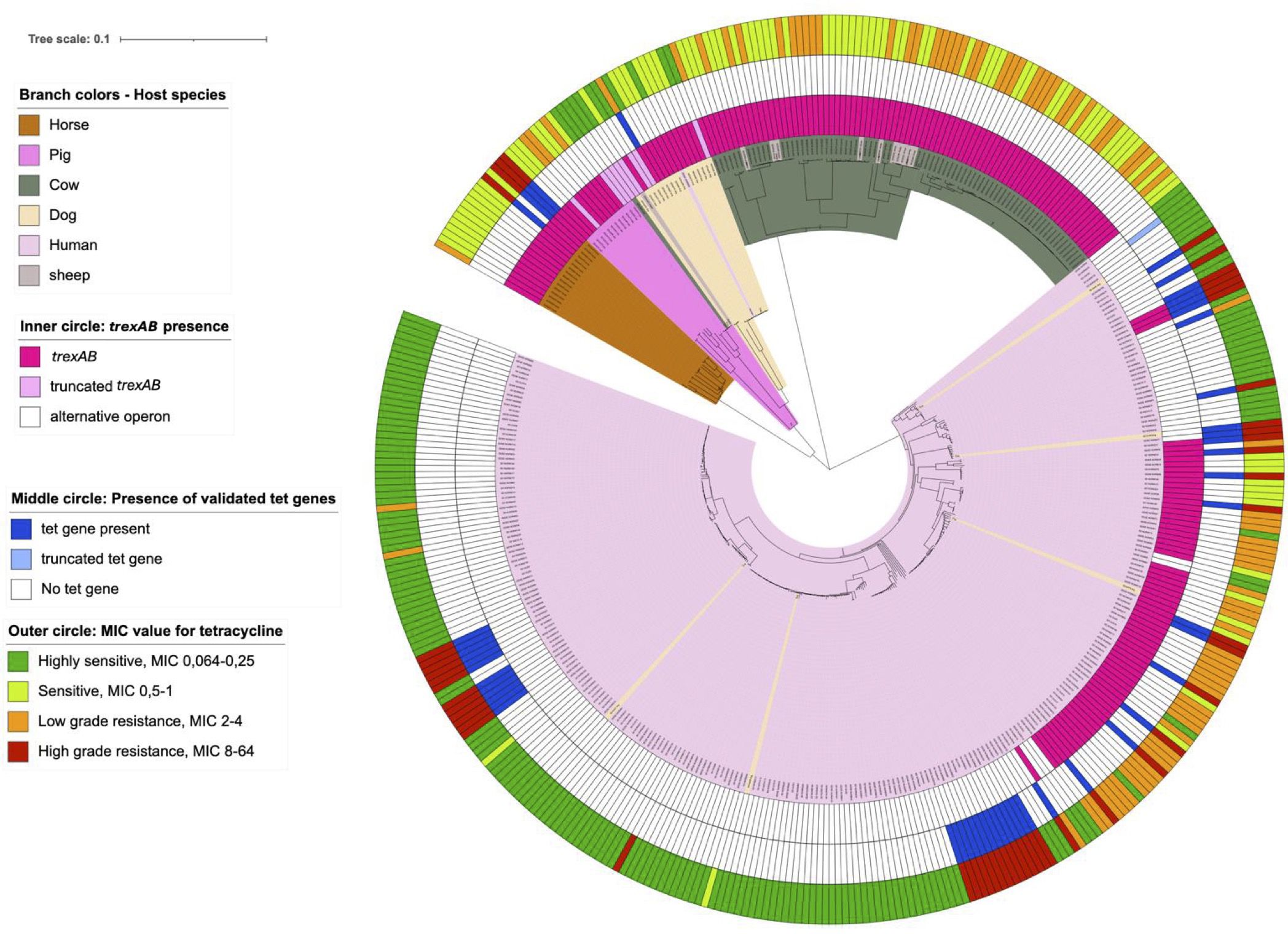
Figure 4. The relationship between genotypic and phenotypic tetracycline resistance and their linkage to phylogenetic distribution. Scale indicates substitutions per site. The phylogenetic tree is constructed based on a core genome single nucleotide polymorphism alignment on maximum likelihood method.
Further examination of collections of SD genomes available online revealed results in accordance with the findings regarding our strain collection. We found trexAB to be present in 22% (n = 294) isolates originating from Australia, 23% (n = 137) isolates from Japan and 7% (n = 122) isolates from Canada, all isolates associated with human infection or carriage. The trexAB carriage of SD isolates from livestock was higher and found in 100% of SD isolates in three different collections of isolates from cattle (n = 86), swine (n = 97) and horses (n = 14), respectively. These animal associated SD were isolated from widespread geographical areas, including locations in China, North America and Europe, confirming a global distribution of trexAB.
In BLASTp searches for TrexA and TrexB homologs in other bacterial species, the closest match was found in Streptococcus canis, with 100% query coverage and sequence identity of 88% and 89% on the aa level regarding TrexA and TrexB, respectively. Homologs with limited sequence identity (60 – 75%) were also detected in other animal associated streptococcal species, such as Streptococcus suis, Streptococcus phocae and Streptococcus iniae. No homologs were detected among typical human pathogenic species. Interestingly, the only significant homology found to the gene products of the alternative operon was in Streptococcus pyogenes, with both query coverage and sequence identity approximating 100% both on the aa and on the nucleotide level (Figure 3).
Knockout of trexAB
Successful construct of SDSD24ΔtrexAB and iSDSE_NORM6ΔtrexAB knockout mutants was confirmed by whole genome sequencing, and the difference between wild type and mutant was limited to the expected exchange of trexAB with the kanamycin cassette. In susceptibility testing for tetracycline, minocycline, tigecycline, doxycycline, erythromycin, gentamicin, clindamycin and ciprofloxacin, both transformant strains showed an increased susceptibility for tetracycline and tigecycline compared to their respective wild types, with a 16- to 32-fold reduction in MIC for tetracycline and a 4-fold reduction in MIC for tigecycline (Table 2). For iSDSE_NORM6ΔtrexAB, a small difference was noted also regarding susceptibility to minocycline, with MIC of 0.032 μg/ml compared to 0.064 μg/ml for the wild type. For all other antibiotics tested, no difference in susceptibility between mutant and wild type was observed.
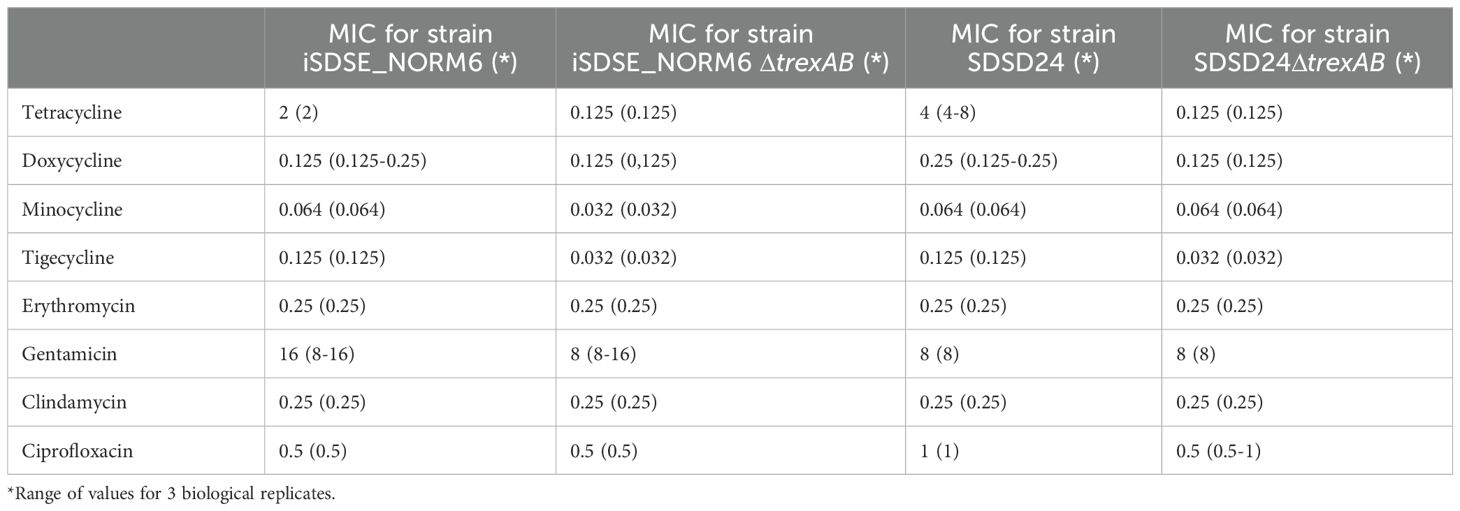
Table 2. MIC values for wildtype and knockout strains.
Discussion
In the present study, we demonstrate that a two-gene operon designated trexAB is associated with reduced susceptibility to tetracycline in SD, and that a targeted knockout of the operon lead to a 16- to 32-fold decrease in tetracycline MIC-values compared to the wildtype strains. The operon has a widespread dissemination and conserved chromosomal location without indications of being located on a mobile genetic element.
Notably, the trexAB-operon appears to have a skewed phylogenetic ecological distribution. Whereas only 25% of the human associated isolates in our collection contained the operon, almost all animal-associated isolates were found to carry trexAB. A similar distribution is evident in epidemiological collections of SD genomes available in GenBank and PubMLST.
Interestingly, the skewed distribution of trexAB between human- and animal-associated isolates is in line with the detection of the closest homolog to trexAB in a dog-associated species, S. canis, while the alternative operons dissemination is limited to the strictly human pathogen S. pyogenes. Combined with the mutual exclusivity of these two operons, this could point to interspecies horizontal genetic exchange and SD evolution occurring within their respective ecological niches. A similar evolutionary phenomenon has also been inferred in several previous genomic studies (Bessen et al., 2005; Ward et al., 2009; Porcellato et al., 2021). Moreover, extensive exchange of genetic material between SD and S. pyogenes has previously been documented in silico (Xie et al., 2024), as have adaptations of SD to host species through presumed tailored genetic content specific to SD of each host species (Porcellato et al., 2021). Nevertheless, further studies are needed to elucidate the potential origin of these operons.
In silico predictions of the domain architecture of the two amino acid sequences encoded in trexAB, revealed typical features of a heterodimeric multidrug resistance transporter (MDR) transporter (Lubelski et al., 2004; Matsuo et al., 2007; Torres et al., 2009; Reilman et al., 2014; Hürlimann et al., 2016). These transporters are typically shown capable of exporting a selection of substances across the cell membrane. Susceptibility testing for several antibiotics demonstrated an effect by trexAB only for tetracycline and to a lesser extent tigecycline, which provides a tenuous foundation for interpreting trexAB as a multidrug transporter. However, we only evaluated trexAB in relation to antibiotics, whereas tests of other MDR transporters have included a wider range of noxious substances like ethidium bromide, safranin, doxorubicin, pyrroles and acriflavine (Orelle et al., 2019). Thus, a broader selection of substrates for efflux by trexAB than demonstrated here is possible.
The level of resistance to tetracycline caused by trexAB seems to be modest, and the impact on susceptibility to tigecycline even more so. As such, the clinical significance of harboring this operon on treatment efficacy is uncertain. Notwithstanding, future vigilance towards potential treatment failures is warranted. Regardless of clinical impact, interpretation of tetracycline resistance in trexAB-positive isolates undoubtably represent a challenge, as the MIC distribution of this population is intersected by the current EUCAST breakpoint. Due to inherent technical and analytical variability in susceptibility testing, such isolates will thus be interchangeably classified as resistant or susceptible to tetracycline. Notably, in 2023 EUCAST removed the category “Susceptible, increased exposure” for tetracycline in beta-hemolytic streptococcal species, including SD. They argued that a pharmacodynamic and pharmacokinetic rationale for an intermediate category was not evident, and lowered the breakpoint for resistance from “above 2 μg/ml” to “above 1 μg/ml”. Considering the widespread distribution of low-grade resistant trexAB-positive SD isolates, the implications for dosage and clinical efficacy need to be carefully evaluated before further revising the EUCAST breakpoints.
A limitation of the knockout experiments in this study is the fact that both resistance genes were removed in one maneuver, making it difficult to decipher the individual contribution of TrexA and TrexB. However, several others have documented the need for the contribution from both half transporters for the function of a heterodimeric efflux pump (Matsuo et al., 2007; Garvey et al., 2010; Warburton et al., 2013). In addition, our collection contained isolates where trexA alone was truncated, which in each case was associated with full susceptibility to tetracycline.
Another potential limitation is the use of MIC-strips for susceptibility testing, as broth microdilution or disc diffusion are the reference methods proposed by EUCAST (EUCAST, 2025). Nevertheless, a distribution of tetracycline susceptibility encircling the current breakpoint is evident also in studies of SD using broth microdilution methodology (McDougall et al., 2014; Jensen et al., 2024). Moreover, we have disc diffusion data for one third of the isolates in the present study, and the susceptibility distribution is congruent with the MIC-strip results (data not shown).
In conclusion, we have investigated the cause for low grade tetracycline resistance in S. dysgalactiae and found the underlying genetic factor to be the two gene operon trexAB encoding a hitherto uncharacterized ABC transporter. The tetracycline MIC distribution of the trexAB-positive isolates is intersected by the current EUCAST breakpoint, and the clinical implications of this should be subject to scrutiny.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author contributions
MG: Conceptualization, Data curation, Formal analysis, Writing – original draft, Writing – review & editing. MK: Conceptualization, Project administration, Supervision, Writing – review & editing. MM: Methodology, Supervision, Writing – review & editing. ZS: Methodology, Supervision, Writing – review & editing. SS: Funding acquisition, Supervision, Writing – review & editing. AS: Writing – review & editing. BK: Writing – review & editing. OO: Conceptualization, Data curation, Formal analysis, Funding acquisition, Methodology, Project administration, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by grants from the Norwegian surveillance program for antimicrobial resistance (NORM) (Grant number: 19/08, to OO). The genomic Core Facility (GCF) at the University of Bergen, which is part of the NorSeq consortium, provided services on the whole genome sequencing in this study; GCF is supported by major grants from the Research Council of Norway (grant no. 245979/F50) and Trond Mohn Foundation (grant no. BFS2017TMT04/BFS2017TMT08). This study was received also financial contributions from the Western Norway Regional Health Authority (grant no. 912231). SS received grants from the Norwegian Research Council under the frames of NordForsk (Project: 90456, PerAID) and ERA PerMed (Project: 2018-151, PerMIT).
Acknowledgments
We thank the staff at Microbiological Department at Haukeland University Hospital for excellent technical assistance and for providing access to their laboratory facilities.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. As a dictionary for word expressions. Not for generating text or images.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2025.1583926/full#supplementary-material
References
Aziz, R. K., Bartels, D., Best, A. A., DeJongh, M., Disz, T., Edwards, R. A., et al. (2008). The RAST server: rapid annotations using subsystems technology. BMC Genom. 9, 75. doi: 10.1186/1471-2164-9-75
Bankevich, A., Nurk, S., Antipov, D., Gurevich, A. A., Dvorkin, M., Kulikov, A. S., et al. (2012). SPAdes: A new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 19, 455–477. doi: 10.1089/cmb.2012.0021
Bessen, D. E., Manoharan, A., Luo, F., Wertz, J. E., and Robinson, D. A. (2005). Evolution of transcription regulatory genes is linked to niche specialization in the bacterial pathogen Streptococcus pyogenes. J. Bacteriol. 187, 4163–4172. doi: 10.1128/jb.187.12.4163-4172.2005
Blum, M., Andreeva, A., Florentino, L. C., Chuguransky, S. R., Grego, T., Hobbs, E., et al. (2024). InterPro: the protein sequence classification resource in 2025. Nucleic Acids Res. 53, D444–D456. doi: 10.1093/nar/gkae1082
Cinthi, M., Massacci, F. R., Coccitto, S. N., Albini, E., Cucco, L., Orsini, M., et al. (2023). Characterization of a prophage and a defective integrative conjugative element carrying the optrA gene in linezolid-resistant Streptococcus dysgalactiae subsp. equisimilis isolates from pigs, Italy. J. Antimicrob. Chemother. 78, 1740–1747. doi: 10.1093/jac/dkad164
EUCAST (2025).European Committee on Antimicrobial Susceptibility Testing. Breakpoint tables for interpretation of MICs and zone diameters version 15.0, 2025. Available online at: https://www.eucast.org/clinical_breakpoints (Accessed February 24, 2025).
Garvey, M. I., Baylay, A. J., Wong, R. L., and Piddock, L. J. V. (2010). Overexpression of patA and patB, Which Encode ABC Transporters, Is Associated with Fluoroquinolone Resistance in Clinical Isolates of Streptococcus pneumoniae. Antimicrob. Agents Chemother. 55, 190–196. doi: 10.1128/aac.00672-10
Glambek, M., Skrede, S., Sivertsen, A., Kittang, B. R., Kaci, A., Jonassen, C. M., et al. (2024). Antimicrobial resistance patterns in Streptococcus dysgalactiae in a One Health perspective. Front. Microbiol. 15. doi: 10.3389/fmicb.2024.1423762
Higuchi, R., Krummel, B., and Saiki, R. (1988). A general method of in vitro preparation and specific mutagenesis of DNA fragments: study of protein and DNA interactions. Nucleic Acids Res. 16, 7351–7367. doi: 10.1093/nar/16.15.7351
Hürlimann, L. M., Corradi, V., Hohl, M., Bloemberg, G. V., Tieleman, D. P., and Seeger, M. A. (2016). The heterodimeric ABC transporter efrCD mediates multidrug efflux in enterococcus faecalis. Antimicrob. Agents Chemother. 60, 5400–5411. doi: 10.1128/aac.00661-16
Jaillard, M., Lima, L., Tournoud, M., Mahé, P., Belkum, A., Lacroix, V., et al. (2018). A fast and agnostic method for bacterial genome-wide association studies: Bridging the gap between k-mers and genetic events. PLoS Genet. 14, e1007758. doi: 10.1371/journal.pgen.1007758
Jensen, V. F., Damborg, P., Norström, M., Nonnemann, B., Slettemeås, J. S., Smistad, M., et al. (2024). Estimation of epidemiological cut-off values for eight antibiotics used for treatment of bovine mastitis caused by Streptococcus uberis and Streptococcus dysgalactiae subsp. dysgalactiae. Vet. Microbiol. 290, 109994. doi: 10.1016/j.vetmic.2024.109994
Johnsborg, O., Eldholm, V., Bjørnstad, M. L., and Håvarstein, L. S. (2008). A predatory mechanism dramatically increases the efficiency of lateral gene transfer in Streptococcus pneumoniae and related commensal species. Mol. Microbiol. 69, 245–253. doi: 10.1111/j.1365-2958.2008.06288.x
Kaas, R. S., Leekitcharoenphon, P., Aarestrup, F. M., and Lund, O. (2014). Solving the problem of comparing whole bacterial genomes across different sequencing platforms. PLoS One 9, e104984. doi: 10.1371/journal.pone.0104984
Lacks, S. and Hotchkiss, R. D. (1960). A study of the genetic material determining an enzyme activity in Pneumococcus. Biochim. Biophys. Acta 39, 508–518. doi: 10.1016/0006-3002(60)90205-5
Letunic, I. and Bork, P. (2021). Interactive Tree Of Life (iTOL) v5: an online tool for phylogenetic tree display and annotation. Nucleic Acids Res. 49, gkab301–. doi: 10.1093/nar/gkab301
Levy, S. B., McMurry, L. M., Barbosa, T. M., Burdett, V., Courvalin, P., Hillen, W., et al. (1999). Nomenclature for new tetracycline resistance determinants. Antimicrob. Agents Chemother. 43, 1523–1524. doi: 10.1128/aac.43.6.1523
Lother, S. A., Demczuk, W., Martin, I., Mulvey, M. R., Dufault, B., Lagacé-Wiens, P., et al. (2017). Clonal clusters and virulence factors of group C and G streptococcus causing severe infections, manitoba, Canada 2012–2014 - volume 23, number 7—July 2017 - emerging infectious diseases journal - CDC. Emerg. Infect. Dis. 23, 1079–1088. doi: 10.3201/eid2307.161259
Lubelski, J., Mazurkiewicz, P., Merkerk, R., Konings, W. N., and Driessen, A. J. M. (2004). ydaG and ydbA of Lactococcus lactis Encode a Heterodimeric ATP-binding Cassette-type Multidrug Transporter*. J. Biol. Chem. 279, 34449–34455. doi: 10.1074/jbc.m404072200
Mårli, M. T., Oppegaard, O., Porcellato, D., Straume, D., and Kjos, M. (2024). Genetic modification of Streptococcus dysgalactiae by natural transformation. mSphere 9, e0021424. doi: 10.1128/msphere.00214-24
Matsuo, T., Chen, J., Minato, Y., Ogawa, W., Mizushima, T., Kuroda, T., et al. (2007). SmdAB, a heterodimeric ABC-type multidrug efflux pump, in Serratia marcescens. J. Bacteriol. 190, 648–654. doi: 10.1128/jb.01513-07
McDougall, S., Hussein, H., and Petrovski, K. (2014). Antimicrobial resistance in Staphylococcus aureus, Streptococcus uberis and Streptococcus dysgalactiae from dairy cows with mastitis. N. Z. Vet. J. 62, 68–76. doi: 10.1080/00480169.2013.843135
NORM (2019).NORM/NORM-VET 2018. Usage of antimicrobial agents and occurrence of antimicrobial resistance in Norway. Tromsø/Oslo 2019. Available online at: https://www.unn.no/4a79d7/siteassets/documents/kompetansetjenester–sentre-og-fagrad/norm—norsk-overvakingssystem-for-antibiotikaresistens-hos-mikrober/rapporter/norm-norm-vet-2018.pdf (Accessed 2019).
Oppegaard, O., Mylvaganam, H., Skrede, S., Lindemann, P. C., and Kittang, B. R. (2017). Emergence of a Streptococcus dysgalactiae subspecies equisimilis stG62647-lineage associated with severe clinical manifestations. Sci. Rep. 7, 7589. doi: 10.1038/s41598-017-08162-z
Orelle, C., Mathieu, K., and Jault, J.-M. (2019). Multidrug ABC transporters in bacteria. Res. Microbiol. 170, 381–391. doi: 10.1016/j.resmic.2019.06.001
Pinho, M. D., Erol, E., Ribeiro-Gonçalves, B., Mendes, C. I., Carriço, J. A., Matos, S. C., et al. (2016). Beta-hemolytic Streptococcus dysgalactiae strains isolated from horses are a genetically distinct population within the Streptococcus dysgalactiae taxon. Sci. Rep. 6, 31736. doi: 10.1038/srep31736
Porcellato, D., Smistad, M., Skeie, S. B., Jørgensen, H. J., Austbø, L., and Oppegaard, O. (2021). Whole genome sequencing reveals possible host species adaptation of Streptococcus dysgalactiae. Sci. Rep. 11, 17350. doi: 10.1038/s41598-021-96710-z
Reilman, E., Mars, R. A. T., van Dijl, J. M., and Denham, E. L. (2014). The multidrug ABC transporter BmrC/BmrD of Bacillus subtilis is regulated via a ribosome-mediated transcriptional attenuation mechanism. Nucleic Acids Res. 42, 11393–11407. doi: 10.1093/nar/gku832
Roberts, M. (2024).Tetracycline genes Modified 1. Available online at: https://faculty.washington.edu/marilynr/tetweb4.pdf (Accessed February 10, 2025).
Shinohara, K., Murase, K., Tsuchido, Y., Noguchi, T., Yukawa, S., Yamamoto, M., et al. (2023). Clonal Expansion of Multidrug-Resistant Streptococcus dysgalactiae Subspecies equisimilis Causing Bacteremia, Japan 2005–2021 - Volume 29, Number 3—March 2023 - Emerging Infectious Diseases journal - CDC. Emerg. Infect. Dis. 29, 528–539. doi: 10.3201/eid2903.221060
Sung, C. K., Li, H., Claverys, J. P., and Morrison, D. A. (2001). An rpsL Cassette, Janus, for Gene Replacement through Negative Selection in Streptococcus pneumoniae. Appl. Environ. Microbiol. 67, 5190–5196. doi: 10.1128/aem.67.11.5190-5196.2001
Thaker, M., Spanogiannopoulos, P., and Wright, G. D. (2010). The tetracycline resistome. Cell. Mol. Life Sci. 67, 419–431. doi: 10.1007/s00018-009-0172-6
Torres, C., Galián, C., Freiberg, C., Fantino, J.-R., and Jault, J.-M. (2009). The YheI/YheH heterodimer from Bacillus subtilis is a multidrug ABC transporter. Biochim. Biophys. Acta (BBA) - Biomembr. 1788, 615–622. doi: 10.1016/j.bbamem.2008.12.012
Vélez, J. R., Cameron, M., Rodríguez-Lecompte, J. C., Xia, F., Heider, L. C., Saab, M., et al. (2017). Whole-Genome Sequence Analysis of Antimicrobial Resistance Genes in Streptococcus uberis and Streptococcus dysgalactiae Isolates from Canadian Dairy Herds. Front. Vet. Sci. 4. doi: 10.3389/fvets.2017.00063
Vieira, V. V., Teixeira, L. M., Zahner, V., Momen, H., Facklam, R. R., Steigerwalt, A. G., et al. (1998). Genetic relationships among the different phenotypes of Streptococcus dysgalactiae strains. Int. J. Syst. Evol. Micr 48, 1231–1243. doi: 10.1099/00207713-48-4-1231
Wang, J., Chitsaz, F., Derbyshire, M. K., Gonzales, N. R., Gwadz, M., Lu, S., et al. (2022). The conserved domain database in 2023. Nucleic Acids Res. 51, D384–D388. doi: 10.1093/nar/gkac1096
Warburton, P. J., Ciric, L., Lerner, A., Seville, L. A., Roberts, A. P., Mullany, P., et al. (2013). TetAB(46), a predicted heterodimeric ABC transporter conferring tetracycline resistance in Streptococcus australis isolated from the oral cavity. J. Antimicrob. Chemother. 68, 17–22. doi: 10.1093/jac/dks351
Ward, P. N., Holden, M. T., Leigh, J. A., Lennard, N., Bignell, A., Barron, A., et al. (2009). Evidence for niche adaptation in the genome of the bovine pathogen Streptococcus uberis. BMC Genom. 10, 54. doi: 10.1186/1471-2164-10-54
Xie, O., Zachreson, C., Tonkin-Hill, G., Price, D. J., Lacey, J. A., Morris, J. M., et al. (2024). Overlapping Streptococcus pyogenes and Streptococcus dysgalactiae subspecies equisimilis household transmission and mobile genetic element exchange. Nat. Commun. 15, 3477. doi: 10.1038/s41467-024-47816-1
Keywords: antibiotic resistance, tetracycline, Streptococcus dysgalactiae, natural transformation, ABC transporter
Citation: Glambek M, Kjos M, Mårli MT, Salehian Z, Skrede S, Sivertsen A, Kittang BR and Oppegaard O (2025) TrexAB, a novel tetracycline resistance determinant in Streptococcus dysgalactiae. Front. Cell. Infect. Microbiol. 15:1583926. doi: 10.3389/fcimb.2025.1583926
Received: 26 February 2025; Accepted: 11 June 2025;
Published: 30 June 2025.
Edited by:
Daniela Visaggio, Roma Tre University, ItalyReviewed by:
Yahan Wei, The University of Texas Rio Grande Valley, United StatesKarl Pedersen, Aarhus University, Denmark
Copyright © 2025 Glambek, Kjos, Mårli, Salehian, Skrede, Sivertsen, Kittang and Oppegaard. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Marte Glambek, bWFydGUuZ2xhbWJla0B1aWIubm8=