 Jorge Vasconez-Gonzalez1
Jorge Vasconez-Gonzalez1 Isaac A. Suárez-Sangucho1
Isaac A. Suárez-Sangucho1 Esteban Acosta-Muñoz1
Esteban Acosta-Muñoz1 Luis Paz y Miño1
Luis Paz y Miño1 Domenic Borja-Mendoza1John Altamirano Alexander-Castillo1Julia Saa1
Domenic Borja-Mendoza1John Altamirano Alexander-Castillo1Julia Saa1 Natasha Salazar-Calvopiña1
Natasha Salazar-Calvopiña1 Paúl Cárdenas2
Paúl Cárdenas2 Andrés López-Cortés3
Andrés López-Cortés3 Esteban Ortiz-Prado1*
Esteban Ortiz-Prado1*- 1One Health Research Group, Faculty of Health Science, Universidad de Las Americas, Quito, Ecuador
- 2Institute of Microbiology, Universidad San Francisco de Quito, Quito, Ecuador
- 3Cancer Research Group (CRG), Faculty of Medicine, Universidad de Las Américas, Quito, Ecuador
The human T-cell lymphotropic virus type 1 (HTLV-1) is a highly oncogenic retrovirus recognized as the causative agent of adult T-cell leukemia/lymphoma (ATLL) and HTLV-1-associated myelopathy/tropical spastic paraparesis (HAM/TSP). Among the key risk factors for ATLL development are high proviral load, reduced anti-Tax immune responses, and elevated levels of soluble interleukin-2 receptor. Unlike classical oncogenic viruses, HTLV-1 does not encode proto-oncogenes but instead drives cellular transformation through a combination of mechanisms, including viral gene dysregulation, chromatin remodeling, epigenetic reprogramming, persistent clonal expansion, immune evasion, and RNA-based modifications. Despite growing understanding of these molecular pathways, an effective prophylactic vaccine against HTLV-1 remains unavailable. However, several vaccine strategies including viral vector platforms, mRNA-based candidates, peptide vaccines, and dendritic cell-based approaches have shown promise in preclinical models. In this review, we provide a comprehensive synthesis of current knowledge on HTLV-1 oncogenesis, highlight the roles of viral proteins such as Tax and HBZ in immune evasion, and critically examine the state of vaccine development efforts aimed at controlling this neglected human retrovirus.
1 Introduction
Human T-lymphotropic viruses (HTLVs) constitute a family of human retroviruses classified as oncoviruses due to their ability to induce malignant transformation. To date, four distinct types have been identified: HTLV-1, HTLV-2, HTLV-3, and HTLV-4 (Eusebio-Ponce et al., 2019; Bryan and Tadi, 2025). Among them, HTLV-1 is the most clinically significant and was the first human retrovirus to be discovered. It was isolated in 1980 from T-cell lines derived from patients with cutaneous T-cell lymphoma and adult T-cell leukemia (Eusebio-Ponce et al., 2019; Forlani et al., 2021). Notably, it was also the first retrovirus conclusively linked to human disease (Coffin, 2015).
HTLV-1 is associated with several diseases, most notably adult T-cell leukemia/lymphoma (ATLL) and HTLV-1-associated myelopathy/tropical spastic paraparesis (HAM/TSP) (Kannian and Green, 2010). Although it can infect a variety of immune and endothelial cell types including T cells, B cells, monocytes, dendritic cells, and endothelial cells its transforming capacity is restricted to primary T lymphocytes, which play a central role in initiating and sustaining the adaptive immune response (Macatonia et al., 1992; Hanon et al., 2000; Forlani et al., 2021).
HTLV-1 is transmitted through parenteral, sexual, and vertical routes. Effective transmission notably requires direct transfer of infected cells, as the cell-free virus is largely non-infectious due to host cellular barriers that hinder viral propagation (Giam and Semmes, 2016; Eusebio-Ponce et al., 2019). In fact, free HTLV-1 virions exhibit low infectivity for most cell types. Following transmission, the majority of infected individuals remain asymptomatic throughout life. However, approximately 3%–5% of cases may progress to ATLL, a highly aggressive hematologic malignancy (Osame et al., 1986; Giam and Semmes, 2016).
2 HTLV-1 structure
HTLVs are enveloped retroviruses with an approximate diameter of 80 to 100 nm. HTLV-1, the most clinically relevant member of this group, contains a single-stranded RNA genome enclosed within an icosahedral capsid. This capsid is surrounded by a host-derived proteolipid bilayer that incorporates viral surface glycoproteins essential for cell entry (Verdonck et al., 2007; IARC Working Group on the Evaluation of Carcinogenic Risks to Humans, 2012; Bryan and Tadi, 2025).
Unlike other retroviruses, HTLV-1 particles are often polymorphic, which may partly explain their limited infectivity in the cell-free state (Meissner et al., 2017). The capsid protein (CA) consists of N- and C-terminal domains connected by a flexible linker and shows distinct structural characteristics compared with HIV-1 and equine infectious anemia virus (EIAV), including a dependence on the oxidation state of cysteine residues that may influence oligomerization and capsid assembly (Khorasanizadeh et al., 1999). Internally, HTLV-1 encodes a classic retroviral genomic structure, including long terminal repeat (LTR) sequences and three structural genes: gag, pol, and env. In addition, the pX region, contains regulatory genes (tax, rex) and accessory genes (p12, p13, p30, and HBZ) (IARC Working Group on the Evaluation of Carcinogenic Risks to Humans, 2012; Bryan and Tadi, 2025). This organization was first elucidated through complete sequencing of the integrated provirus in leukemic cells (Seiki et al., 1983; Haseltine et al., 1985).
The Gag precursor is cleaved by viral protease into p19 (matrix), p24 (capsid), and p15 (nucleocapsid), whereas the Env precursor undergoes cellular protease-mediated cleavage into gp46 (surface glycoprotein) and gp21 (transmembrane protein) (Figure 1) (IARC Working Group on the Evaluation of Carcinogenic Risks to Humans, 2012; Bryan and Tadi, 2025). These proteins are critical for viral assembly, infectivity, and cell-to-cell transmission, which occurs predominantly through virological synapses rather than free virion dissemination (Lairmore et al., 2011). The Tax protein (40 kDa) is a potent transactivator of viral and cellular genes involved in transformation and proliferation, whereas Rex (27 kDa) regulates the nuclear export and stability of viral RNAs (Gaudray et al., 2002; Lambert, 2009). The HBZ protein, transcribed from the antisense strand, plays key roles in immune evasion (Uchiyama, 1997), cell survival, and oncogenesis (Romanelli et al., 2013).
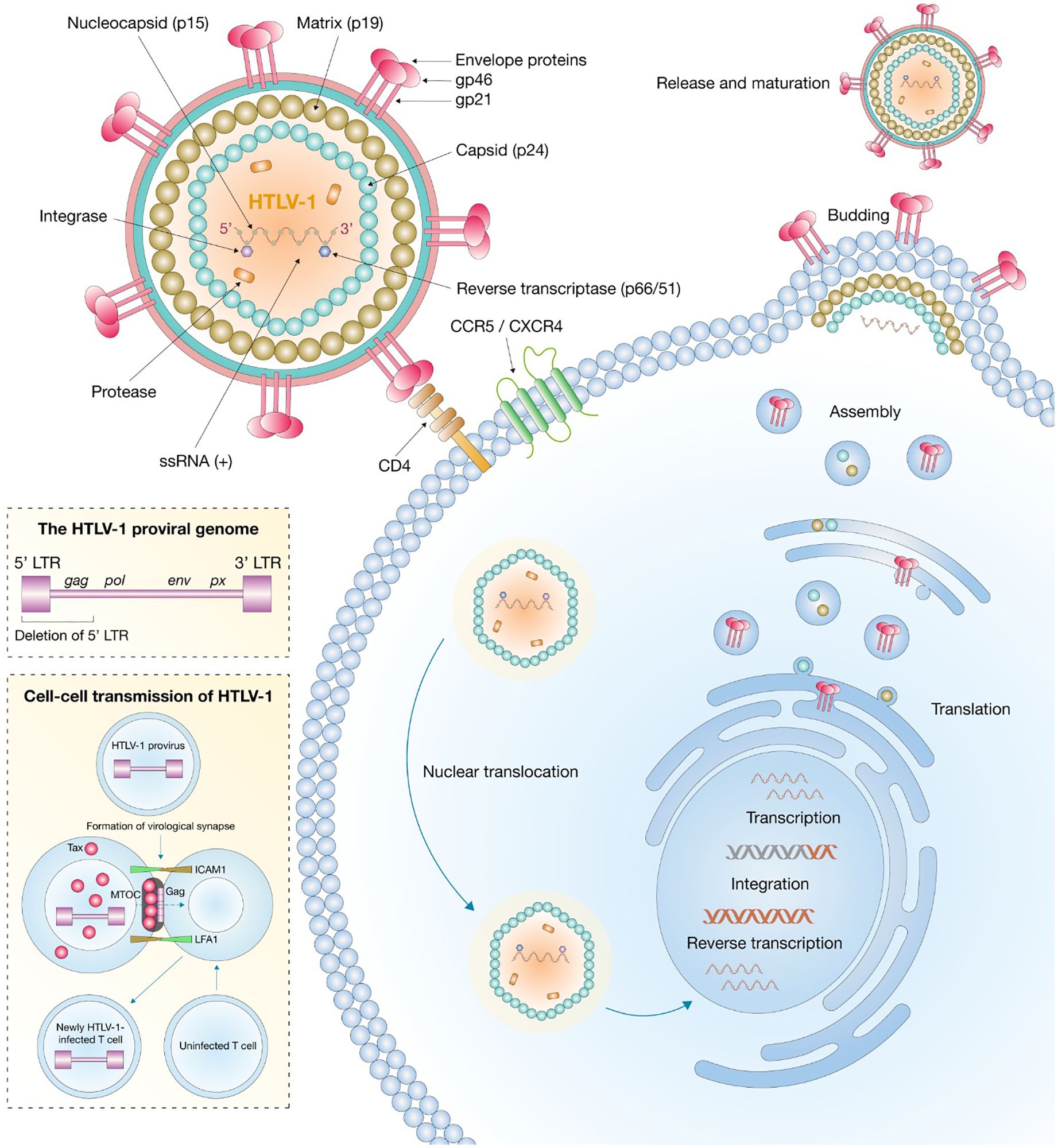
Figure 1. Structure and replication of HTLV-1.
3 Epidemiology
It is estimated that between 5 and 10 million people are currently infected with HTLV-1 worldwide, although the true burden is likely underestimated due to limited screening in endemic regions (Gessain et al., 2023; Mendoza et al., 2024). Approximately 50% of these infections occur in equatorial Africa, where prevalence varies markedly between regions (Mendoza et al., 2024). In adult populations, seroprevalence ranges from 0.3% to 3%, with significantly higher rates in older individuals (Gessain et al., 2023). In some African communities, particularly among older women, prevalence can reach 10%–25% (Gessain et al., 2023). The geographic distribution of HTLV-1 is highly heterogeneous. While Eastern and Northern Africa exhibit lower prevalence rates, other regions—including the southwest of Japan, parts of South America, the Caribbean, Australo-Melanesia, and focal areas in the Middle East—are considered endemic zones (Tsukasaki and Tobinai, 2013; Eusebio-Ponce et al., 2019). In Indigenous Australian populations, HTLV-1 infection has reached hyperendemic levels, with reported seroprevalence as high as 33.6%, particularly among older men (Einsiedel et al., 2016). In Europe, data remain limited; however, in England and Wales, it was estimated that 36,300 individuals were living with HTLV-1 as of 2021 (Mendoza et al., 2024). Prevalence in Israel has also shown geographic variability, with higher rates among donors from Eastern Europe, Latin America, and the Middle East (Stienlauf et al., 2009). Major routes of transmission include breastfeeding, sexual contact, and exposure to infected blood. The risk of transmission is heightened in settings where public health infrastructure does not support routine screening or educational interventions (Radygina and Mochalova, 2024).
4 Transmission mechanisms of HTLV-1
4.1 Mother-to-child transmission of HTLV-1
Mother-to-child transmission represents a significant route for the dissemination of HTLV-1, accounting for approximately 15% to 25% of all infections (Bittencourt, 1998; Martin-Latil et al., 2012). The main way that vertical transmission occurs is through extended breastfeeding, particularly when it continues beyond 6 months. The risk of transmission in these cases can range from 4% to 31%, depending on how long and how often the infant is breastfed (Ngoma et al., 2019; Miyazawa et al., 2021; Vesterbacka et al., 2024). Various factors related to both the mother and the child can affect how likely this transmission is to happen. One of the most important factors is the level of proviral load in the mother’s breast milk—higher levels are strongly linked to a greater chance of the infant becoming infected (Hisada et al., 2002; Van Tienen et al., 2012). While it is less common, transmission can also happen during pregnancy or childbirth, with estimates suggesting this occurs in approximately 0.5% to 7% of cases (Denis et al., 1992; Berteau et al., 1994).
4.2 Sexual transmission of HTLV-1
Sexual transmission is one of the main ways adults acquire HTLV-1, especially in areas where the virus is not common (Paiva and Casseb, 2014; Martel and Gotuzzo, 2022). The virus is transmitted more efficiently from men to women than the other way around, which is likely because seminal fluid tends to carry a higher proviral load than cervicovaginal secretions (Murphy et al., 1989; Ayerdi et al., 2023). According to epidemiological studies, the estimated transmission rate among heterosexual couples is around 0.6 cases per 100 person-years (Roucoux et al., 2005). Risk factors that increase susceptibility include having multiple sexual partners, engaging in unprotected intercourse, the presence of genital ulcers, and a history of sexually transmitted infections (STIs), such as syphilis, which is associated with a 40-fold increase in HTLV-1 seropositivity (Kaplan et al., 1996; Martel and Gotuzzo, 2022). A longer duration of sexual relationships and higher viral loads further enhance the likelihood of transmission (Kaplan et al., 1996; Paiva and Casseb, 2014).
4.3 Transmission through blood and medical procedures
HTLV-1 is a bloodborne virus that is mainly spread through transfusions of contaminated blood products and through needle-sharing among people who inject drugs (Hewagama et al., 2014; Oliveira et al., 2018). This route of transmission is highly efficient because the virus is integrated into the DNA of infected lymphocytes, which are passed directly to the recipient (Cassar and Gessain, 2017; Kalinichenko et al., 2022). In addition to recreational drug use, medical procedures have also played a major role in the spread of HTLV-1 (Pépin et al., 2010). HTLV-1 can also be transmitted through solid organ transplants, which has led to calls for universal screening of both donors and recipients in regions where the virus is common (Osman, 2003; Yara et al., 2009). While less frequent, healthcare workers have occasionally been infected through occupational exposures such as needlestick injuries, highlighting the ongoing importance of strict biosafety measures (Weiss et al., 1985).
5 Oncological mechanisms of HTLV-1
5.1 Multifaceted oncogenic role of HTLV-1 Tax
Tax acts as a viral transactivator protein that plays a key role in regulating HTLV-1 gene expression. It forms a complex with pCREB, which then binds to viral DNA and recruits’ coactivators like CBP/p300 to help activate transcription. Tax also promotes the recruitment of positive transcription elongation factor b (P-TEFb), which supports the elongation phase of transcription (Zhou et al., 2006; Nyborg et al., 2010). In addition, Tax facilitates the assembly of transcriptional machinery at the viral long terminal repeat (LTR) by helping recruit TBP, general transcription factors, and RNA polymerase II. This is achieved through the formation of quaternary complexes that include histone-modifying enzymes, ultimately aiding in the production of viral RNA (de la Fuente and Kashanchi, 2004). Tax also enhances gene expression by increasing the binding of activating transcription factor 2 and CRE-binding protein to the 21-base pair repeat enhancer regions (Franklin et al., 1993). Furthermore, it interacts with BRG1, which helps bring in the core transcriptional machinery, leading to higher transcription activity and greater viral replication (Easley et al., 2010).
The stability of the Tax protein plays a key role in maintaining persistent HTLV-1 infection and contributing to its ability to cause cancer. Several mechanisms have been identified that help keep Tax stable within infected cells. For example, research has shown that Tax can interfere with the ubiquitin system, which not only prevents its own degradation but also activates NF-κB signaling by promoting the activity of ubiquitin kinases (Lavorgna and Harhaj, 2014). It was also found that heat shock protein 90 (HSP90) protects Tax from being broken down by the proteasome, helping to stabilize it and enhance NF-κB pathway activation (Gao and Harhaj, 2013). They also noted that when HSP90 is inhibited, Tax levels drop significantly in HTLV-1-transformed cells, highlighting the potential of HSP90 inhibitors as a possible therapeutic approach (Gao and Harhaj, 2013). Moreover, it has been shown that even the transient expression of Tax is sufficient to activate anti-apoptotic mechanisms, which remain active even when Tax expression decreases. Even low levels of Tax can activate NF-κB and induce IL-10 production, both of which play a crucial role in the leukemogenesis of ATLL. On the other hand, low-level Tax expression is sufficient for the survival of primary ATLL cells (Mahgoub et al., 2018; Hleihel et al., 2023). This evidence highlights Tax as a promising therapeutic target, as the absence of Tax renders primary ATLL cells unable to survive, despite the presence of somatic mutations and HBZ (Hleihel et al., 2023).
Tax supports the proliferation of T cells by interfering with key regulatory proteins like cyclin D2 and D3 (Figure 2). This interference drives continuous, uncontrolled growth of mature T cells (Azran et al., 2004). Tax also suppresses the activity of the tumor-suppressor protein p53, disrupting normal cell cycle checkpoints and contributing to genomic instability an early step in the development of ATLL (Pise-Masison and Brady, 2005; Tabakin-Fix et al., 2006; Currer et al., 2012). Additionally, Tax can inhibit the transcription of human telomerase reverse transcriptase (hTERT), resulting in reduced telomerase activity in affected cells (Gabet et al., 2003). It also downregulates DNA polymerase β, a key enzyme in DNA repair processes, thereby impairing several repair pathways, including mismatch repair, base excision repair, and nucleotide excision repair (Azran et al., 2004). On top of that, Tax contributes to genomic instability by disrupting the function of checkpoint proteins like MDC1 and ATM (Mühleisen et al., 2014; Mohanty and Harhaj, 2020).
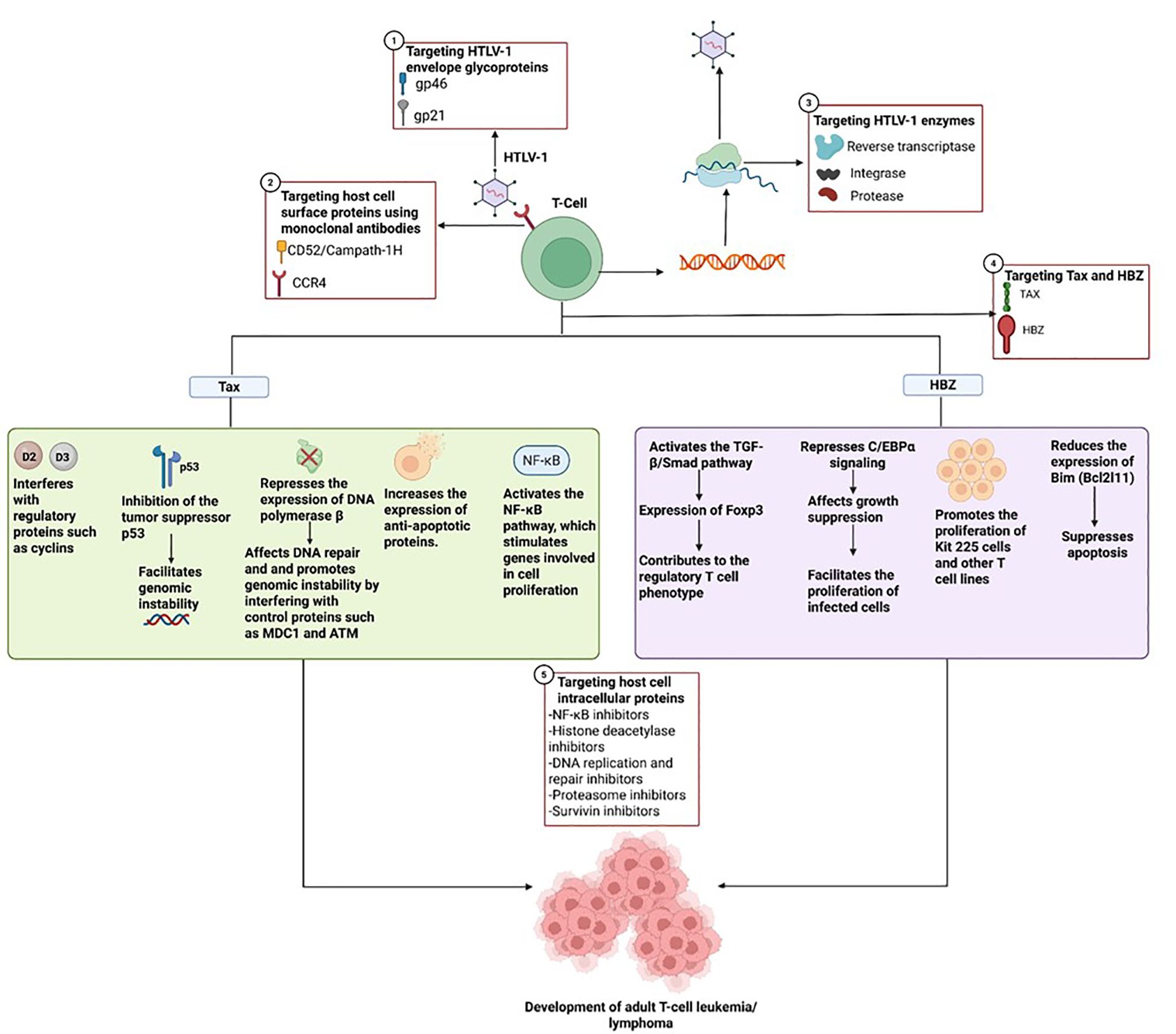
Figure 2. Oncogenic role of HTLV-1 mediated by Tax and HBZ, and possible therapeutic targets covering different stages ranging from T-cell infection by the virus, through the replication process, to mechanisms involved in oncogenesis.
In addition, Tax increases the expression of anti-apoptotic proteins from the Bcl-2 family (Mühleisen et al., 2014; Mohanty and Harhaj, 2020) and upregulates XIAP through the activation of the NF-κB pathway (Kawakami et al., 1999). Tax-driven activation of NF-κB occurs through both the canonical and non-canonical signaling pathways. In this process, Tax binds to NEMO, which helps recruit the IKK complex and results in sustained NF-κB activation (Cheng et al., 2012; Mohanty and Harhaj, 2020). The IKK complex composed of IKKα, IKKβ, and NEMO triggers phosphorylation events of IκB and NF-Kb (Hinz and Scheidereit, 2014). More specifically, IKK phosphorylates IκB, marking it for ubiquitination and subsequent degradation, which allows NF-κB to move into the nucleus (Mohanty and Harhaj, 2020). A non-canonical NF-κB pathway also exists and is activated by signals such as B-cell activating factor, lymphotoxin-beta receptor, and CD40 (Mohanty and Harhaj, 2020).
Tax also promotes cell survival by linking IKK signaling to autophagy pathways, further contributing to cellular transformation (Cheng et al., 2012). Beyond NF-κB, it activates PI3K and AP-1, promotes trimethylation of histone H3K27, and engages EZH2, leading to gene silencing (Peloponese and Jeang, 2006; Fujikawa et al., 2016). Tax directly interacts with NF-κB subunits (p50, p52, RelA, c-Rel) and NF-κB precursors (p100, p105), enabling their activation and nuclear translocation (Li and Gaynor, 2018). Through these interactions, Tax enhances transcriptional activity and supports T-cell transformation (Mohanty and Harhaj, 2020).
Tax also activates TRAF6 and stabilizes MCL-1 via K63-linked polyubiquitination, conferring resistance to apoptosis (Mohanty and Harhaj, 2020). Ubiquitination at lysines K263, K280, and K284 regulates Tax stability and NF-κB activity through platforms that recruit IKK and TAK1 (Lavorgna and Harhaj, 2014). The K63-linked polyubiquitination of Tax enables the recruitment of kinases such as IKK and TAK1, facilitating the activation of NF-κB. Furthermore, Tax contributes to sustained NF-κB activation by interacting with both NEMO and Optineurin (Lavorgna and Harhaj, 2014). In response to DNA damage, Tax is monoubiquitinated at lysine residues K280 and K284, a modification that leads to its dissociation from nuclear bodies containing SC35. This change promotes the export of Tax from the nucleus through the CRM1-dependent pathway (Gatza et al., 2007; Lavorgna and Harhaj, 2014).
Tax can also be modified through SUMOylation, primarily at lysine residues K280 and K284. This modification plays a key role in regulating Tax’s movement from the cytoplasm into the nucleus, where it can interact with p300, RelA, and NEMO (Lavorgna and Harhaj, 2014; Mohanty and Harhaj, 2020). In addition, phosphorylation of Tax contributes to gene expression by activating the ATF/CREB and NF-κB pathways and promotes its accumulation in nuclear bodies activation (Mohanty and Harhaj, 2020). Mohanty et al. observed in their study that Tax can also be phosphorylated by VEGFR2. This phosphorylation protects Tax from degradation by autophagy and also stabilizes it. Therefore, drugs targeting VEGFR2 represent a promising therapeutic strategy, as its inhibition leads to apoptosis of infected T cells through Tax degradation. This, in turn, disrupts the NF-κB and JAK/STAT signaling pathways (Mohanty et al., 2024). Acetylation also plays a role when Tax is acetylated, it interacts with p300 to further support NF-κB activation (Mohanty and Harhaj, 2020).
Finally, Tax also contributes to immune evasion. It has been shown to inhibit interferon-β-inducing adaptors, disrupting their role as cellular sensors and reducing the production of type I interferons (Hyun et al., 2015). Tax also interferes with type I IFN signaling pathways mediated by RIG-I and MDA-5, making cells more vulnerable to viral infections (Hyun et al., 2015). Additionally, Tax has been found to interact with RIPK1, which affects the production of IFN-α, and with STING, thereby impairing the induction of IFN-β. These combined actions help create an immunosuppressive environment (Mohanty and Harhaj, 2023).
5.2 HBZ and its multifaceted role in oncogenesis
HBZ is a nuclear protein that consists of three main structural domains: an activation domain (AD) at the N-terminus, a central domain (CD), and a basic leucine zipper (bZIP) domain (Ma et al., 2017). Within the activation domain, two LXXLL-like motifs interact with the KIX domain of the transcriptional coactivators CBP/p300, which are crucial for chromatin remodeling and transcriptional regulation. These motifs are particularly important for HBZ’s ability to activate the TGF-β/Smad signaling pathway. This activation leads to the induction of Foxp3 expression, promoting a regulatory T cell phenotype (Figure 2) (Ma et al., 2016; Zhao, 2016).
HBZ has also been shown to antagonize Tax-mediated transcriptional activation through the 5′ LTR promoter, likely via interactions with CREB and CBP/p300 (Yoshida et al., 2008; Matsuoka and Green, 2009). Additionally, it can repress C/EBPα signaling, a pathway implicated in growth suppression by physically interacting with C/EBPα and impairing its DNA-binding capacity. This repression facilitates the proliferation of HTLV-1-infected cells, a key event in leukemogenesis (Zhao and Matsuoka, 2012; Zhao, 2016). HBZ also stimulates the expression of JunD and promotes the production of its alternative isoform, ΔJunD, through the nuclear retention of RPS25. This cooperation with JunD and Sp1 enhances transcription from the 3′ LTR and upregulates the expression of hTERT, thereby reinforcing proliferative and survival signaling (Zhao and Matsuoka, 2012; Gazon et al., 2018).
Apart from its role as a protein, HBZ mRNA itself plays a significant role in oncogenesis. It has been found to promote the proliferation of Kit 225 cells and other T-cell lines by forming stem-loop structures that activate E2F1 and its downstream targets, thereby encouraging progression from the G1 to the S phase of the cell cycle (Satou et al., 2006). In addition to promoting cell growth, HBZ RNA also helps cells evade apoptosis by downregulating Bim (Bcl2l11), a key pro-apoptotic gene. This occurs through its interference with FoxO3a, a transcription factor that normally activates Bim and FasL, by preventing its proper localization to the nucleus and its ability to bind DNA (Tanaka-Nakanishi et al., 2014).
HBZ also contributes to oncogenesis by upregulating oncogenic microRNAs (oncomiRs) at the posttranscriptional level. Higher levels of HBZ expression have been associated with the overexpression of specific oncomiRs in CD4+ T cells from individuals infected with HTLV-1, a pattern that supports the development of malignant features (Vernin et al., 2014; Mitagami et al., 2015).
Additionally, HBZ plays a significant role in immune evasion. It weakens CD4+ T-cell responses and disrupts interferon-gamma (IFN-γ) signaling by suppressing the canonical NF-κB and AP-1 pathways. These actions contribute to immune tolerance and enable the virus to persist in the host (Zhao, 2016). This protein has also been reported to stimulate the production of co-inhibitory molecules, enhance FOXP3 expression in coordination with TGF-β signaling, and increase IL-10 production, all of which contribute to the suppression of immune activation. Furthermore, by downregulating canonical NF-κB signaling, HBZ helps prevent the expression of cellular senescence markers in infected cells (Tan et al., 2022; Mohanty and Harhaj, 2023).
5.3 Additional proteins involved in oncogenesis
Rex is a 27-kDa phosphoprotein that functions as a posttranscriptional regulator, enhancing the expression of both spliced and unspliced viral mRNAs. It is also essential for maintaining the proper balance between these mRNAs, a process critical for the production of infectious viral particles (Ahmadi Ghezeldasht et al., 2013). The accessory protein p13 has been shown to induce an inward K+ current within mitochondria, which leads to an increase in the production of reactive oxygen species (ROS). In addition, p13 has been reported to enhance the effects of pro-apoptotic stimuli, including Fas ligand (FasL) and C2-ceramide (Silic-Benussi et al., 2010).
On the other hand, 12-kDa protein (p12I) is necessary for the infection of primary lymphocytes and plays an important role during the early stages of HTLV-1 infection. Additionally, by binding to the heavy chain of MHC I, p12I renders it susceptible to degradation and may reduce the presentation of viral peptide MHC I complexes on the surface of infected cells, thereby protecting them from lysis by cytotoxic T lymphocytes (Ahmadi Ghezeldasht et al., 2013).
The p30 protein inhibits DNA repair via homologous recombination by targeting the MRE11/RAD50/NBS1 complex and instead promotes the error-prone non-homologous end joining (NHEJ) DNA repair pathway. This shift facilitates the accumulation of mutations in the host genome and increases the cumulative risk of cellular transformation (Baydoun et al., 2011).
5.4 Role of the JAK/STAT, PI3K/AKT, and NF-κB signaling pathways
Signaling through hormone and cytokine receptors is the most widely accepted model for JAK activation; this process leads to the aggregation of JAK proteins within lipid rafts of the cell membrane (Bellon and Nicot, 2024). Once activated, this pathway results in a loss of phosphatase expression. Additionally, IL-2 has been shown to promote T-cell proliferation, whereas IL-13 exerts both proliferative and anti-apoptotic effects via the JAK/STAT pathway (Islam et al., 2021; Bellon and Nicot, 2024).
It has been reported that the p12 protein interacts with the interleukin-2 receptor β chain and the IL-2Rγ chains. This interaction activates the Janus kinase/signal transducer and activator of transcription 5 (JAK/STAT5) signaling pathway, which is involved in the infection of quiescent primary T cells and the formation of syncytia (Kannian and Green, 2010).
On the other hand, the PI3K/AKT signaling pathway has been identified as playing a role in cell survival, as well as in regulating cell cycle progression. Moreover, AKT activation leads to the inhibition of p53. Jeong et al. indicate that p53 siRNA contributes to cell survival by preventing LY294002-induced apoptosis (Jeong et al., 2008). In addition, Tax regulates the expression of Bcl-3 by activating the PI3K/AKT signaling pathway, a mechanism that has been linked to the increased proliferation of HTLV-1-infected T cells (Saito et al., 2010; Marino-Merlo et al., 2023).
The activation of the NF-κB pathway is essential for both the survival and proliferation of HTLV-1-infected T cells. Both the canonical and non-canonical branches of this pathway contribute to the upregulation of genes involved in cell proliferation, cell cycle progression, and anti-apoptotic functions (Harhaj and Giam, 2018; Zhao et al., 2022). However, sustained NF-κB activation also leads to the accumulation of R-loops nucleic acid structures that, when resolved, can result in DNA double-strand breaks (Giam and Pasupala, 2022). These breaks contribute to genomic instability, including insertions, deletions, and other structural alterations. Over time, such mutations may inactivate tumor-suppressor genes or disrupt regulatory feedback within the IKK/NF-κB pathway, promoting the development of adult ATLL (Giam and Pasupala, 2022).
6 HTLV-1 vaccine platforms
6.1 Viral vector vaccines
Viral vectors represent a relatively recent approach in vaccine development, using engineered viruses to deliver specific immunogens into the host. These vectors can trigger innate immune responses without requiring an adjuvant. Because they are capable of infecting host cells and expressing foreign antigens, they promote effective antigen presentation and activate major histocompatibility complex (MHC) pathways, ultimately eliciting a strong cellular immune response (McCann et al., 2022). This platform offers several advantages, including efficient expression of intracellular antigens, the induction of a potent cytotoxic T-cell response, and the stimulation of innate immunity resulting in the production of interferons and pro-inflammatory cytokines (Santana et al., 2023).
A variety of viral vectors have been employed to express HTLV-1 genes, including adenovirus (AdV), cytomegalovirus (CMV), vaccinia virus (VV), variola virus (VARV), and baculovirus (BV) (Letafati et al., 2024). One of the earliest attempts to develop a vaccine against HTLV-1 was carried out by Shida et al., who engineered recombinant vaccinia viruses to carry the HTLV-1 envelope gene. This gene was inserted into the hemagglutinin (HA) gene locus of the vaccinia virus, which served as a novel insertion site for foreign genetic material (Shida et al., 1987). By 1992, Ford et al. designed recombinant vaccinia viruses named RVV E1, RVV E2, and RVV E3 to express three different versions of the HTLV-1 envelope proteins (Ford et al., 1992) (Table 1).
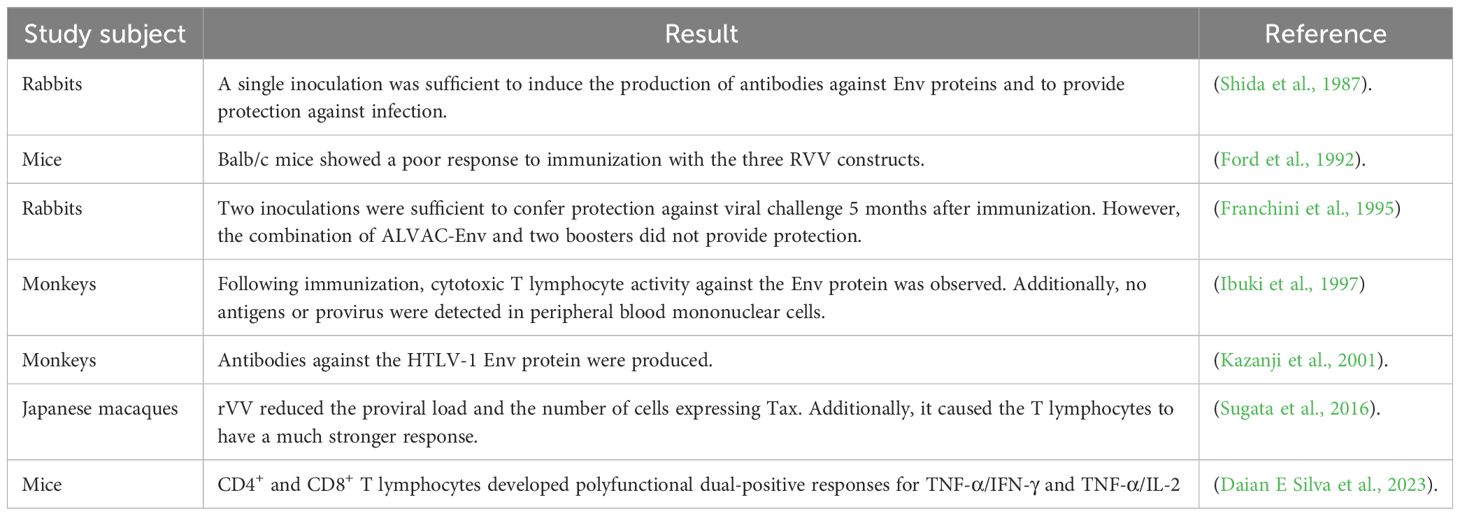
Table 1. HTLV-1 viral vector vaccines.
In 1995, the full-length envelope protein of the HTLV-I (1711) virus was expressed using highly attenuated poxvirus vaccine vectors ALVAC and NYVAC. These vectors were used to immunize white rabbits; however, even after booster doses, no sustained immunity was observed, as the animals became infected with HTLV-1 after a few months (Franchini et al., 1995). Two years later, the recombinant vaccinia virus WR-SFB5env, which synthesizes the gp46 envelope protein, was developed. It was observed that WR-SFB5env expressed high levels of gp46 and was capable of inducing anti-HTLV-I Env antibodies, including neutralizing antibodies, which were maintained at high levels for up to 136 weeks (Ibuki et al., 1997). For their part, Kazanji et al. evaluated the immunogenicity and efficacy of two NYVAC-based vaccine candidates. They found that sensitizing the animals with a single dose of env DNA, followed by immunization with the NYVAC-HTLV-1 gag and env vaccine at months 6, 7, and 8, protected all three animals against exposure to HTLV-1-infected cells (Kazanji et al., 2001).
In another study involving vaccination with recombinant vaccinia virus (rVV) expressing either the Tax or HBZ protein, administered to rhesus monkeys and mice, it was observed that rVV expressing HBZ could induce HBZ-specific CD4+ and CD8+ T-cell responses. However, compared with the Tax protein, the immunogenicity of HBZ was low (Sugata et al., 2016). More recently, in 2023, a recombinant modified vaccinia Ankara virus (MVA-HBZ) and a plasmid DNA vector (pcDNA3.1(+)-HBZ) expressing a multiepitope protein based on HBZ peptides were developed. The results highlight the potential of the HBZ multiepitope protein, expressed from both the plasmid DNA and the poxviral vector, as candidates for a therapeutic vaccine (Daian E Silva et al., 2023).
6.2 Protein/peptide vaccines
Peptide vaccines are composed of the most immunogenic protein fragments and are capable of stimulating the humoral immune response through antibody production, thereby reducing the risk of adverse effects (Santana et al., 2023). The peptides present in this type of vaccine have the ability to stimulate antibody production, and when combined with adjuvants, they enhance immune responses. Their efficacy has been demonstrated in vaccine development trials (Letafati et al., 2024). Vaccination with peptides based on T- and B-cell epitopes, constructed by conjugating gp46 (aa 181–210) with a branched poly-lysine oligomer, was tested in rats and rabbits and demonstrated high levels of neutralizing antibodies against HTLV-1 (Seighali et al., 2023).
A study conducted in rabbits revealed that synthetic gp46 peptides containing amino acids 190–199 and 180–204 are effective in generating neutralizing antibody responses. Upon reinfection with HTLV-1, no detectable provirus was found in these vaccinated rabbits (Tanaka et al., 1994). The use of synthetic peptides is a promising strategy for the development of a vaccine targeting the envelope of this pathogen (Santana et al., 2023). The most recent vaccines are described in Table 2.
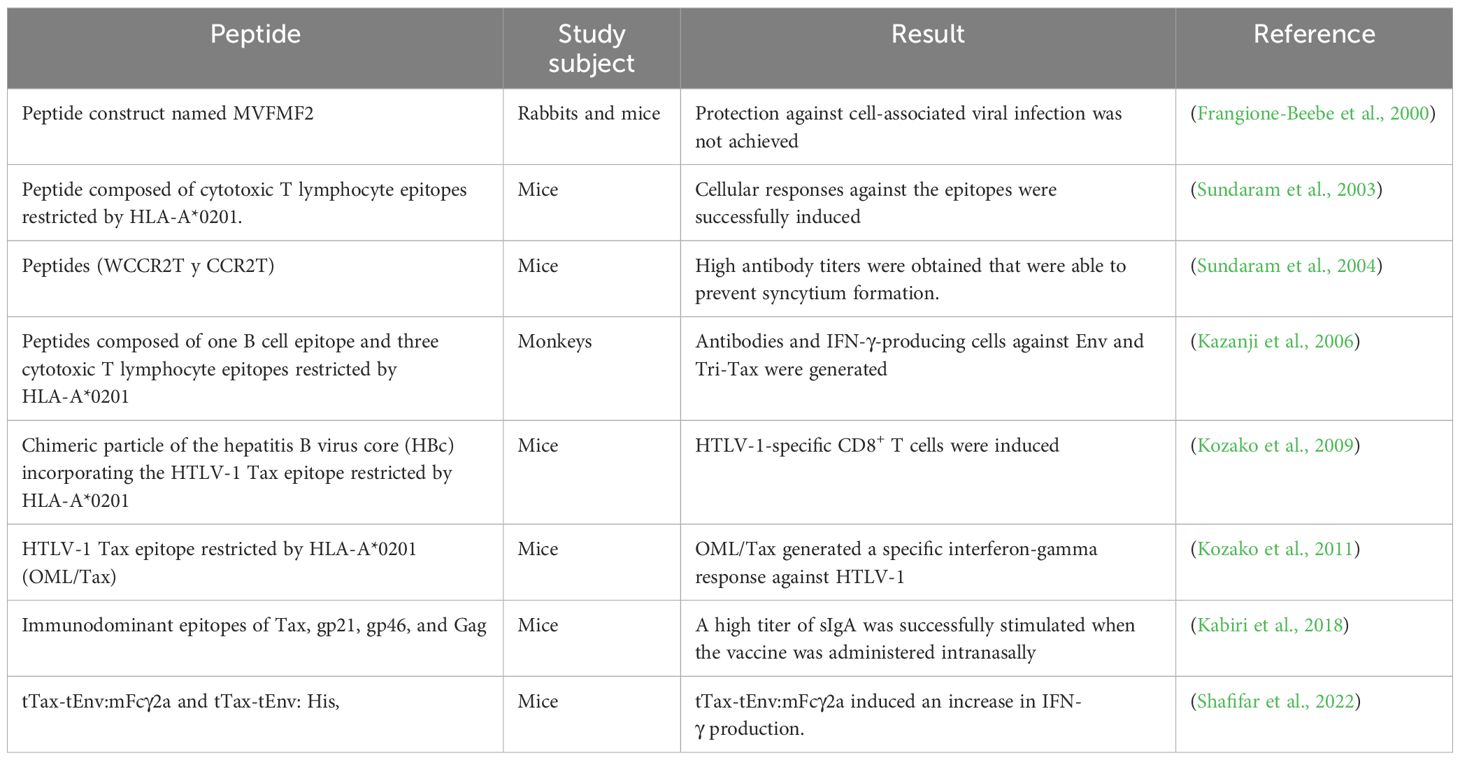
Table 2. HTLV-1 protein/peptide vaccines.
6.3 DNA vaccines
This type of vaccine is capable of generating adaptive immune responses comparable with those produced after administration of attenuated pathogen vaccines while maintaining the safety profile of subunit vaccines. Consequently, they are suitable for administration in immunocompromised individuals (Kozak and Hu, 2024). This type of vaccine is composed of a plasmid responsible for encoding an immunogen, which is administered into the body to generate a targeted immune response (Letafati et al., 2024).
One of the first studies of this type of vaccine against HTLV-1 was conducted in 1987, where the immunization of animals with products from the HTLV-I env gene produced in Escherichia coli was evaluated. The results revealed that animals immunized with the env product produced antibodies against HTLV-I gp68 and gp46, and their sera were found to strongly inhibit syncytium formation in a feline fibroblast cell line induced by HTLV-I (Nakamura et al., 1987). Later, in 1997, two plasmids encoding the full HTLV-1 envelope proteins were evaluated, with or without recombinant Baculovirus gp62 protein boosts, in BALB/c mice. The authors concluded that a single DNA inoculation expressing the HTLV-I env gene could stimulate memory B-cell clones capable of responding effectively to subsequent encounters with HTLV-I envelope proteins and a specific helper T-cell response in mice (Grange et al., 1997). By the year 2000, Armand et al. concluded, after evaluating two plasmids, that the choice of vectors would be crucial for the design of genetic vaccines against HTLV-I (Armand et al., 2000) (Table 3).

Table 3. HTLV-1 DNA vaccines.
6.4 Dendritic cell-based vaccines
Dendritic cell-based constructs have been proposed as therapeutic vaccines capable of inducing antigen-specific CD8+ T cell responses (Seighali et al., 2023; Letafati et al., 2024). Sagar et al. were among the first to suggest a dendritic cell-based vaccine candidate against HTLV-1, which utilized the Tax (11-19) epitope and effectively induced antigen-specific CD8+ T cells. Moreover, administration of Freund’s adjuvant reduced TGF-β levels and enhanced the CD8+ T cell response (Sagar et al., 2014) (Table 4).
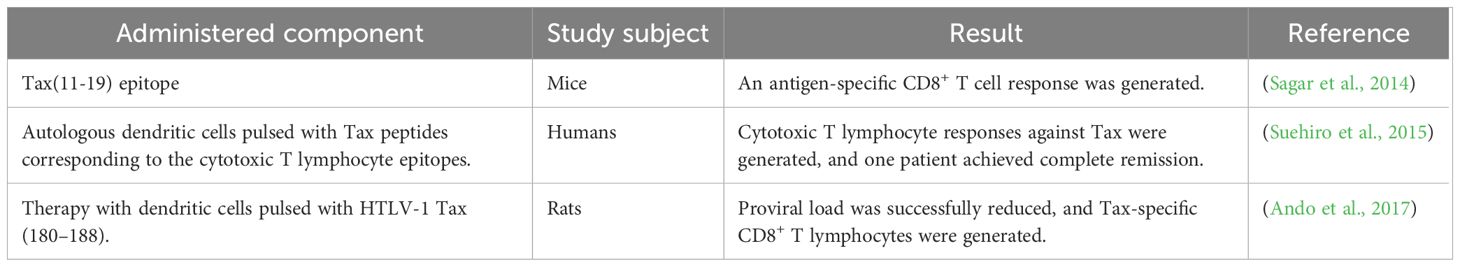
Table 4. HTLV-1 dendritic cell-based vaccines.
6.5 mRNA vaccines
mRNA vaccines have been shown to possess high potency, safety, and efficacy (Gote et al., 2023). These types of vaccines exert their protective effects by inoculating target cells with mRNA fragments related to a viral protein, enabling the cells to express this protein in order to trigger recognition by the immune system (Letafati et al., 2024).
In 2024, a codon-optimized mRNA encoding the HTLV-1 envelope (Env) was developed and evaluated. Results in animal models revealed that three rabbits were partially protected and three were fully protected against HTLV-1 exposure. It was also observed that in those immunized with Env mRNA-LNP, the proviral load and viral gene expression were significantly lower, in addition to showing an increase in CD4+/IFN-γ+ and CD8+/IFN-γ+ T cells (Tu et al., 2024).
6.6 HTLV-1 vaccine challenges
Among the main challenges in developing an effective vaccine against HTLV-1 are the virus’s ability to infect and persist in CD4+ T cells, as well as its capacity for cell-to-cell transmission, which facilitates immune evasion (Letafati et al., 2024). Furthermore, understanding the mechanisms by which the immune response can control HTLV-1 infection is necessary for vaccine design. However, with this type of virus, there is an additional challenge due to the limited knowledge about its protective mechanisms and pathogenesis (Santana et al., 2023). It is essential to thoroughly understand the gp46 subunit and its roles and configuration, as it may contribute to the development of successful vaccines. Therefore, the development of tools for its research and understanding is crucial (Letafati et al., 2024).
7 New therapies
7.1 Mogamulizumab
Is a humanized monoclonal antibody against CCR4 (Figure 2) that leads to antibody-dependent elimination of CCR4+ cells. It is used for the treatment of T-cell lymphomas (Meyerowitz et al., 2023). In a multicenter, randomized, phase 3 study in which 34 and 33 patients were randomized to the mogamulizumab and placebo arms, respectively, the results revealed that the mogamulizumab arm showed a significant reduction in HTLV-1 proviral load (−59.39% ± 29.91% vs. placebo 2.32% ± 36.31%) (Sato et al., 2024).
On the other hand, in an uncontrolled phase 1–2a study involving patients with HAM/TSP, it was observed that by day 15, the proviral load was reduced by 64.9%. By day 29, the number of HTLV-1 proviral copies per milliliter of cerebrospinal fluid (CSF) decreased by 41.4% (Sato et al., 2018). Additionally, in a study evaluating long-term safety and efficacy, the results revealed that 19% of participants developed neutralizing antibodies, and after 4 years, the proviral load decreased by 60.7% in peripheral blood and 66.3% CSF (Sato et al., 2023).
7.2 EZH2 inhibitors
Koseki et al. reported in their study that EZH2 and EZH1/2 inhibitors are capable of reducing proviral loads and increasing IL-10 levels. They also trigger the inhibition of proliferation in infected cells and enhance early apoptosis, as indicated by annexin-V(+) 7-aminoactinomycin D(−) staining (Koseki et al., 2023). It has been described that it is possible to reverse the epigenetic alteration and even eliminate leukemic cells infected with the virus through pharmacological inhibition of EZH2. Moreover, by inhibiting both EZH2 and DZNep, apoptosis of ATLL cells can be induced (Fujikawa et al., 2016; Letafati et al., 2025).
7.3 Pomalidomide
In T-lymphocyte cell lines immortalized and infected with HTLV-1, pomalidomide increases the expression of MHC-I, ICAM-1, and B7-2/CD86 and also enhances susceptibility to cytotoxicity mediated by NK cells (Davis et al., 2019). In a Rhesus macaque model, Gutowska et al. observed that pomalidomide induced immune activation, marked by increased proliferation of CD4+, CD8+, and NK cells. It was also reported that the specific humoral response against the virus was stronger, and there was an increase in the levels of antibodies directed against viral antigens (Gutowska et al., 2022).
7.4 Alcoholic extract from Eucalyptus camaldulensis
Abu-Jafar et al., in their study, demonstrated that the tannin extract of Eucalyptus camaldulensis inhibited the activation of NF-κB, SRF-dependent promoters, and the HTLV-1 LTR induced by Tax, and was even capable of preventing the degradation of IκBα (Abu-Jafar et al., 2020).
7.5 (E)-3-Phenyl-5-(phenylamino)-2-styryl-1,3,4-thiadiazol-3-ium chloride derivatives
Sousa-Pereira et al. reported IC50 values for all compounds in the range of 1.51–7.70 μM in both HTLV-1-infected and uninfected cells. These compounds may induce necrosis after 24 h in Jurkat and MT2 cell lines (Sousa-Pereira et al., 2020).
8 Conclusion
Although this virus was the first retrovirus identified as a causal agent of diseases in humans, it is often overlooked and, at times, forgotten, even becoming an unattended public health problem. The cellular and molecular processes through which HTLV-1 leads to the development of ATLL are complex, as they involve a series of mechanisms such as genomic integration, epigenetic reprogramming, persistent clonal expansion, and RNA-based modifications and alternative splicing, processes in which various proteins, including Tax and HBZ, play a role. Understanding the oncogenic mechanisms of this virus is crucial for developing therapies to prevent cancer associated with this infection and for developing effective vaccines. Various platforms have been studied to create a compelling and safe vaccine in recent years, but a safe vaccine that can prevent HTLV-1 infection has not yet been produced. It is also important to emphasize the limited number of clinical trials published regarding their development.
Author contributions
JV-G: Investigation, Methodology, Writing – original draft, Writing – review & editing. IS-S: Investigation, Software, Writing – original draft. EA-M: Software, Writing – original draft. LM: Resources, Writing – original draft. DB-M: Resources, Writing – original draft. JA-C: Methodology, Writing – original draft. JS: Data curation, Investigation, Writing – original draft. NS-C: Methodology, Writing – original draft. PC: Validation, Writing – review & editing. AL-C: Software, Writing – original draft. EO-P: Funding acquisition, Resources, Supervision, Validation, Visualization, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research, and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted without any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abu-Jafar, A., Suleiman, M., Nesim, N., and Huleihel, M. (2020). The effect of alcoholic extract from Eucalyptus camaldulensis leaves on HTLV-1 Tax activities. Cell Cycle Georget. Tex 19, 1768–1776. doi: 10.1080/15384101.2020.1779455
Ahmadi Ghezeldasht, S., Shirdel, A., Assarehzadegan, M. A., Hassannia, T., Rahimi, H., Miri, R., et al. (2013). Human T lymphotropic virus type I (HTLV-I) oncogenesis: molecular aspects of virus and host interactions in pathogenesis of adult T cell leukemia/lymphoma (ATL). Iran. J. Basic Med. Sci. 16, 179–195.
Ando, S., Hasegawa, A., Murakami, Y., Zeng, N., Takatsuka, N., Maeda, Y., et al. (2017). HTLV-1 tax-specific CTL epitope-pulsed dendritic cell therapy reduces proviral load in infected rats with immune tolerance against tax. J. Immunol. Baltim. Md 1950 198, 1210–1219. doi: 10.4049/jimmunol.1601557
Armand, M.-A., Grange, M.-P., Paulin, D., and Desgranges, C. (2000). Targeted expression of HTLV-I envelope proteins in muscle by DNA immunization of mice. Vaccine 18, 2212–2222. doi: 10.1016/S0264-410X(99)00565-4
Ayerdi, O., Benito, R., Ortega, D., Aguilera, A., Montiel, N., Pintos, I., et al. (2023). HTLV infection in persons with sexually transmitted diseases in Spain. Front. Immunol. 14. doi: 10.3389/fimmu.2023.1277793
Azran, I., Schavinsky-Khrapunsky, Y., and Aboud, M. (2004). Role of Tax protein in human T-cell leukemia virus type-I leukemogenicity. Retrovirology 1, 20. doi: 10.1186/1742-4690-1-20
Baydoun, H. H., Pancewicz, J., and Nicot, C. (2011). Human T-lymphotropic type 1 virus p30 inhibits homologous recombination and favors unfaithful DNA repair. Blood 117, 5897–5906. doi: 10.1182/blood-2010-08-304600
Bellon, M. and Nicot, C. (2024). HTLV-1 tax tug-of-war: cellular senescence and death or cellular transformation. Pathogens 13, 87. doi: 10.3390/pathogens13010087
Berteau, F., Martin-Prével, Y., and Bedjabaga, I. (1994). ÉTUDE de la TRANSMISSION VERTICALE DU HUMAN T-CELL LEUKEMIA VIRUS EN ZONE D’ENDÉMIE. Bull. Soc. Path Ex 87, 217–221.
Bittencourt, A. L. (1998). Vertical transmission of HTLV-I/II: a review. Rev. Inst. Med. Trop. São Paulo 40, 245–251. doi: 10.1590/S0036-46651998000400008
Bryan, E. S. and Tadi, P. (2025). “Human T-cell lymphotropic virus,” in StatPearls (StatPearls Publishing, Treasure Island (FL). Available online at: http://www.ncbi.nlm.nih.gov/books/NBK560825/.
Cassar, O. and Gessain, A. (2017). Serological and molecular methods to study epidemiological aspects of human T-cell lymphotropic virus type 1 infection. Methods Mol. Biol. Clifton NJ 1582, 3–24. doi: 10.1007/978-1-4939-6872-5_1
Cheng, H., Ren, T., and Sun, S. (2012). New insight into the oncogenic mechanism of the retroviral oncoprotein Tax. Protein Cell 3, 581–589. doi: 10.1007/s13238-012-2047-0
Coffin, J. M. (2015). The discovery of HTLV-1, the first pathogenic human retrovirus. Proc. Natl. Acad. Sci. U. S. A. 112, 15525–15529. doi: 10.1073/pnas.1521629112
Currer, R., Van Duyne, R., Jaworski, E., Guendel, I., Sampey, G., Das, R., et al. (2012). HTLV tax: a fascinating multifunctional co-regulator of viral and cellular pathways. Front. Microbiol. 3, 406. doi: 10.3389/fmicb.2012.00406
Daian E Silva, D. S. O., Cox, L. J., Rocha, A. S., Lopes-Ribeiro, Á., Souza, J. P. C., Franco, G. M., et al. (2023). Preclinical assessment of an anti-HTLV-1 heterologous DNA/MVA vaccine protocol expressing a multiepitope HBZ protein. Virol. J. 20, 304. doi: 10.1186/s12985-023-02264-z
Davis, D. A., Shrestha, P., Aisabor, A. I., Stream, A., Galli, V., Pise-Masison, C. A., et al. (2019). Pomalidomide increases immune surface marker expression and immune recognition of oncovirus-infected cells. Oncoimmunology 8, e1546544. doi: 10.1080/2162402X.2018.1546544
de la Fuente, C. and Kashanchi, F. (2004). The expanding role of Tax in transcription. Retrovirology 1, 19. doi: 10.1186/1742-4690-1-19
Denis, F., Verdier, M., and Bonis, J. (1992). Vertical transmission of HTLV-I. Pathol. Biol. (Paris) 40, 714–719.
Easley, R., Carpio, L., Guendel, I., Klase, Z., Choi, S., Kehn-Hall, K., et al. (2010). Human T-lymphotropic virus type 1 transcription and chromatin-remodeling complexes. J. Virol. 84, 4755–4768. doi: 10.1128/jvi.00851-09
Einsiedel, L., Woodman, R. J., Flynn, M., Wilson, K., Cassar, O., and Gessain, A. (2016). Human T-Lymphotropic Virus type 1 infection in an Indigenous Australian population: epidemiological insights from a hospital-based cohort study. BMC Public Health 16, 787. doi: 10.1186/s12889-016-3366-5
Eusebio-Ponce, E., Anguita, E., Paulino-Ramirez, R., and Javier Candel, F. (2019). HTLV-1 infection: An emerging risk. Pathogenesis, epidemiology, diagnosis and associated diseases. Rev. Esp. Quimioter. 32, 485–496.
Ford, C. M., Arp, J., Palker, T. J., King, E. E., and Dekaban, G. A. (1992). Characterization of the antibody response to three different versions of the HTLV-I envelope protein expressed by recombinant vaccinia viruses: induction of neutralizing antibody. Virology 191, 448–453. doi: 10.1016/0042-6822(92)90208-7
Forlani, G., Shallak, M., Accolla, R. S., and Romanelli, M. G. (2021). HTLV-1 infection and pathogenesis: new insights from cellular and animal models. Int. J. Mol. Sci. 22, 8001. doi: 10.3390/ijms22158001
Franchini, G., Tartaglia, J., Markham, P., Benson, J., Fullen, J., Wills, M., et al. (1995). Highly attenuated HTLV type Ienv poxvirus vaccines induce protection against a cell-associated HTLV type I challenge in rabbits. AIDS Res. Hum. Retroviruses 11, 307–313. doi: 10.1089/aid.1995.11.307
Frangione-Beebe, M., Albrecht, B., Dakappagari, N., Rose, R. T., Brooks, C. L., Schwendeman, S. P., et al. (2000). Enhanced immunogenicity of a conformational epitope of human T-lymphotropic virus type 1 using a novel chimeric peptide. Vaccine 19, 1068–1081. doi: 10.1016/S0264-410X(00)00340-6
Franklin, A. A., Kubik, M. F., Uittenbogaard, M. N., Brauweiler, A., Utaisincharoen, P., Matthews, M. A., et al. (1993). Transactivation by the human T-cell leukemia virus Tax protein is mediated through enhanced binding of activating transcription factor-2 (ATF-2) ATF-2 response and cAMP element-binding protein (CREB). J. Biol. Chem. 268, 21225–21231. doi: 10.1016/S0021-9258(19)36914-5
Fujikawa, D., Nakagawa, S., Hori, M., Kurokawa, N., Soejima, A., Nakano, K., et al. (2016). Polycomb-dependent epigenetic landscape in adult T-cell leukemia. Blood 127, 1790–1802. doi: 10.1182/blood-2015-08-662593
Gabet, A.-S., Mortreux, F., Charneau, P., Riou, P., Duc-Dodon, M., Wu, Y., et al. (2003). Inactivation of hTERT transcription by Tax. Oncogene 22, 3734–3741. doi: 10.1038/sj.onc.1206468
Gao, L. and Harhaj, E. W. (2013). HSP90 protects the human T-cell leukemia virus type 1 (HTLV-1) tax oncoprotein from proteasomal degradation to support NF-κB activation and HTLV-1 replication. J. Virol. 87, 13640–13654. doi: 10.1128/jvi.02006-13
Gatza, M. L., Dayaram, T., and Marriott, S. J. (2007). Ubiquitination of HTLV-I Tax in response to DNA damage regulates nuclear complex formation and nuclear export. Retrovirology 4, 95. doi: 10.1186/1742-4690-4-95
Gaudray, G., Gachon, F., Basbous, J., Biard-Piechaczyk, M., Devaux, C., and Mesnard, J.-M. (2002). The complementary strand of the human T-cell leukemia virus type 1 RNA genome encodes a bZIP transcription factor that down-regulates viral transcription. J. Virol. 76, 12813–12822. doi: 10.1128/JVI.76.24.12813-12822.2002
Gazon, H., Barbeau, B., Mesnard, J.-M., and Peloponese, J.-M. (2018). Hijacking of the AP-1 signaling pathway during development of ATL. Front. Microbiol. 8. doi: 10.3389/fmicb.2017.02686
Gessain, A., Ramassamy, J.-L., Afonso, P. V., and Cassar, O. (2023). Geographic distribution, clinical epidemiology and genetic diversity of the human oncogenic retrovirus HTLV-1 in Africa, the world’s largest endemic area. Front. Immunol. 14. doi: 10.3389/fimmu.2023.1043600
Giam, C.-Z. and Pasupala, N. (2022). NF-κB-induced R-loops and genomic instability in HTLV-1-infected and adult T-cell leukemia cells. Viruses 14, 877. doi: 10.3390/v14050877
Giam, C.-Z. and Semmes, O. J. (2016). HTLV-1 infection and adult T-cell leukemia/lymphoma—A tale of two proteins: tax and HBZ. Viruses 8, 161. doi: 10.3390/v8060161
Gote, V., Bolla, P. K., Kommineni, N., Butreddy, A., Nukala, P. K., Palakurthi, S. S., et al. (2023). A comprehensive review of mRNA vaccines. Int. J. Mol. Sci. 24, 2700. doi: 10.3390/ijms24032700
Grange, M.-P., Armand, M.-A., Audoly, G., Thollot, D., and Desgranges, C. (1997). Induction of neutralizing antibodies against HTLV-I envelope proteins after combined genetic and protein immunizations in mice. DNA Cell Biol. 16, 1439–1448. doi: 10.1089/dna.1997.16.1439
Gutowska, A., McKinnon, K., Sarkis, S., Doster, M. N., Bissa, M., Moles, R., et al. (2022). Transient viral activation in human T cell leukemia virus type 1-infected macaques treated with pomalidomide. Front. Med. 9. doi: 10.3389/fmed.2022.897264
Hanon, E., Stinchcombe, J. C., Saito, M., Asquith, B. E., Taylor, G. P., Tanaka, Y., et al. (2000). Fratricide among CD8(+) T lymphocytes naturally infected with human T cell lymphotropic virus type I. Immunity 13, 657–664. doi: 10.1016/s1074-7613(00)00065-0
Harhaj, E. W. and Giam, C.-Z. (2018). NF-κB signaling mechanisms in HTLV-1-induced adult T-cell leukemia/lymphoma. FEBS J. 285, 3324–3336. doi: 10.1111/febs.14492
Haseltine, W. A., Sodroski, J. G., and Patarca, R. (1985). “Structure and Function of the Genome of HTLV,” in Human T-Cell Leukemia Virus. Ed. Vogt, P. K. (Springer Berlin Heidelberg, Berlin, Heidelberg), 177–209. doi: 10.1007/978-3-642-70113-9_12
Hewagama, S., Krishnaswamy, S., King, L., Davis, J., and Baird, R. (2014). Human T-cell lymphotropic virus type 1 exposures following blood-borne virus incidents in central Australia 2002-2012. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc Am. 59, 85–87. doi: 10.1093/cid/ciu227
Hinz, M. and Scheidereit, C. (2014). The IκB kinase complex in NF-κB regulation and beyond. EMBO Rep. 15, 46–61. doi: 10.1002/embr.201337983
Hisada, M., Maloney, E. M., Sawada, T., Miley, W. J., Palmer, P., Hanchard, B., et al. (2002). Virus markers associated with vertical transmission of human T lymphotropic virus type 1 in Jamaica. Clin. Infect. Dis. 34, 1551–1557. doi: 10.1086/340537
Hleihel, R., Skayneh, H., de Thé, H., Hermine, O., and Bazarbachi, A. (2023). Primary cells from patients with adult T cell leukemia/lymphoma depend on HTLV-1 Tax expression for NF-κB activation and survival. Blood Cancer J. 13, 67. doi: 10.1038/s41408-023-00841-7
Hyun, J., Ramos, J. C., Toomey, N., Balachandran, S., Lavorgna, A., Harhaj, E., et al. (2015). Oncogenic human T-cell lymphotropic virus type 1 tax suppression of primary innate immune signaling pathways. J. Virol. 89, 4880–4893. doi: 10.1128/JVI.02493-14
IARC Working Group on the Evaluation of Carcinogenic Risks to Humans (2012). “HUMAN T-CELL LYMPHOTROPIC VIRUS TYPE 1,” in Biological Agents (International Agency for Research on Cancer). Available online at: https://www.ncbi.nlm.nih.gov/books/NBK304341/.
Ibuki, K., Funahashi, S. I., Yamamoto, H., Nakamura, M., Igarashi, T., Miura, T., et al. (1997). Long-term persistence of protective immunity in cynomolgus monkeys immunized with a recombinant vaccinia virus expressing the human T cell leukaemia virus type I envelope gene. J. Gen. Virol. 78, 147–152. doi: 10.1099/0022-1317-78-1-147
Islam, S., Espitia, C. M., Persky, D. O., Carew, J. S., and Nawrocki, S. T. (2021). Targeting JAK/STAT signaling antagonizes resistance to oncolytic reovirus therapy driven by prior infection with HTLV-1 in models of T-cell lymphoma. Viruses 13, 1406. doi: 10.3390/v13071406
Jeong, S.-J., Dasgupta, A., Jung, K.-J., Um, J.-H., Burke, A., Park, H. U., et al. (2008). PI3K/AKT inhibition induces caspase-dependent apoptosis in HTLV-1 transformed cells. Virology 370, 264–272. doi: 10.1016/j.virol.2007.09.003
Kabiri, M., Sankian, M., Hosseinpour, M., and Tafaghodi, M. (2018). The novel immunogenic chimeric peptide vaccine to elicit potent cellular and mucosal immune responses against HTLV-1. Int. J. Pharm. 549, 404–414. doi: 10.1016/j.ijpharm.2018.07.069
Kalinichenko, S., Komkov, D., and Mazurov, D. (2022). HIV-1 and HTLV-1 transmission modes: mechanisms and importance for virus spread. Viruses 14, 152. doi: 10.3390/v14010152
Kannian, P. and Green, P. L. (2010). Human T lymphotropic virus type 1 (HTLV-1): molecular biology and oncogenesis. Viruses 2, 2037–2077. doi: 10.3390/v2092037
Kaplan, J. E., Khabbaz, R. F., Murphy, E. L., Hermansen, S., Roberts, C., Lal, R., et al. (1996). Male-to-female transmission of human T-cell lymphotropic virus types I and II: association with viral load. JAIDS J. Acquir. Immune Defic. Syndr. 12, 193–201. doi: 10.1097/00042560-199606010-00014
Kawakami, A., Nakashima, T., Sakai, H., Urayama, S., Yamasaki, S., Hida, A., et al. (1999). Inhibition of caspase cascade by HTLV-I tax through induction of NF-kappaB nuclear translocation. Blood 94, 3847–3854. doi: 10.1182/blood.V94.11.3847
Kazanji, M., Bomford, R., Bessereau, J.-L., Schulz, T., and de Thé, G. (1997). Expression and immunogenicity in rats of recombinant adenovirus 5 DNA plasmids and vaccinia virus containing the HTLV-I-env gene. Int. J. Cancer 71, 300–307. doi: 10.1002/(SICI)1097-0215(19970410)71:2<300::AID-IJC27>3.0.CO;2-J
Kazanji, M., Heraud, J.-M., Merien, F., Pique, C., de Thé, G., Gessain, A., et al. (2006). Chimeric peptide vaccine composed of B- and T-cell epitopes of human T-cell leukemia virus type 1 induces humoral and cellular immune responses and reduces the proviral load in immunized squirrel monkeys (Saimiri sciureus). J. Gen. Virol. 87, 1331–1337. doi: 10.1099/vir.0.81582-0
Kazanji, M., Tartaglia, J., Franchini, G., de Thoisy, B., Talarmin, A., Contamin, H., et al. (2001). Immunogenicity and protective efficacy of recombinant human T-cell leukemia/lymphoma virus type 1 NYVAC and naked DNA vaccine candidates in squirrel monkeys (Saimiri sciureus). J. Virol. 75, 5939–5948. doi: 10.1128/jvi.75.13.5939-5948.2001
Khorasanizadeh, S., Campos-Olivas, R., and Summers, M. F. (1999). Solution structure of the capsid protein from the human T-cell leukemia virus type-I. J. Mol. Biol. 291, 491–505. doi: 10.1006/jmbi.1999.2986
Koseki, A., Araya, N., Yamagishi, M., Yamauchi, J., Yagishita, N., Takao, N., et al. (2023). EZH1/2 dual inhibitors suppress HTLV-1-infected cell proliferation and hyperimmune response in HTLV-1-associated myelopathy. Front. Microbiol. 14. doi: 10.3389/fmicb.2023.1175762
Kozak, M. and Hu, J. (2024). DNA vaccines: their formulations, engineering and delivery. Vaccines 12, 71. doi: 10.3390/vaccines12010071
Kozako, T., Fukada, K., Hirata, S., White, Y., Harao, M., Nishimura, Y., et al. (2009). Efficient induction of human T-cell leukemia virus-1-specific CTL by chimeric particle without adjuvant as a prophylactic for adult T-cell leukemia. Mol. Immunol. 47, 606–613. doi: 10.1016/j.molimm.2009.09.006
Kozako, T., Hirata, S., Shimizu, Y., Satoh, Y., Yoshimitsu, M., White, Y., et al. (2011). Oligomannose-coated liposomes efficiently induce human T-cell leukemia virus-1-specific cytotoxic T lymphocytes without adjuvant. FEBS J. 278, 1358–1366. doi: 10.1111/j.1742-4658.2011.08055.x
Lairmore, M. D., Anupam, R., Bowden, N., Haines, R., Haynes, R. A., Ratner, L., et al. (2011). Molecular determinants of human T-lymphotropic virus type 1 transmission and spread. Viruses 3, 1131. doi: 10.3390/v3071131
Lambert, P. (2009). “Oncogenic viruses,” in Encyclopedia of microbiology, 3rd ed. Ed. Schaechter, M. (Academic Press, Oxford), 421–429. doi: 10.1016/B978-012373944-5.00308-4
Lavorgna, A. and Harhaj, E. W. (2014). Regulation of HTLV-1 tax stability, cellular trafficking and NF-κB activation by the ubiquitin-proteasome pathway. Viruses 6, 3925–3943. doi: 10.3390/v6103925
Letafati, A., Bahari, M., Salahi Ardekani, O., Nayerain Jazi, N., Nikzad, A., Norouzi, F., et al. (2024). HTLV-1 vaccination Landscape: Current developments and challenges. Vaccine X 19, 100525. doi: 10.1016/j.jvacx.2024.100525
Letafati, A., Mehdigholian Chaijani, R., Edalat, F., Eslami, N., Askari, H., Askari, F., et al. (2025). Advances in epigenetic treatment of adult T-cell leukemia/lymphoma: a comprehensive review. Clin. Epigenet. 17, 39. doi: 10.1186/s13148-025-01841-z
Li, X. H. and Gaynor, R. B. (2018). Regulation of NF-κB by the HTLV-1 tax protein. Gene Expr. 7, 233–245.
Ma, G., Yasunaga, J., and Matsuoka, M. (2016). Multifaceted functions and roles of HBZ in HTLV-1 pathogenesis. Retrovirology 13, 16. doi: 10.1186/s12977-016-0249-x
Ma, Y., Zhang, B., Wang, D., Qian, L., Song, X., Wang, X., et al. (2017). HTLV-1 basic leucine zipper factor downregulates cyclin D1 expression via interactions with NF-κB. Int. J. Mol. Med. 39, 764–770. doi: 10.3892/ijmm.2017.2868
Macatonia, S. E., Cruickshank, J. K., Rudge, P., and Knight, S. C. (1992). Dendritic cells from patients with tropical spastic paraparesis are infected with HTLV-1 and stimulate autologous lymphocyte proliferation. AIDS Res. Hum. Retroviruses 8, 1699–1706. doi: 10.1089/aid.1992.8.1699
Mahgoub, M., Yasunaga, J.-I., Iwami, S., Nakaoka, S., Koizumi, Y., Shimura, K., et al. (2018). Sporadic on/off switching of HTLV-1 Tax expression is crucial to maintain the whole population of virus-induced leukemic cells. Proc. Natl. Acad. Sci. U. S. A. 115, E1269–E1278. doi: 10.1073/pnas.1715724115
Marino-Merlo, F., Grelli, S., Mastino, A., Lai, M., Ferrari, P., Nicolini, A., et al. (2023). Human T-cell leukemia virus type 1 oncogenesis between active expression and latency: A possible source for the development of therapeutic targets. Int. J. Mol. Sci. 24, 14807. doi: 10.3390/ijms241914807
Martel, M. and Gotuzzo, E. (2022). HTLV-1 is also a sexually transmitted infection. Front. Public Health 10. doi: 10.3389/fpubh.2022.840295
Martin-Latil, S., Gnädig, N. F., Mallet, A., Desdouits, M., Guivel-Benhassine, F., Jeannin, P., et al. (2012). Transcytosis of HTLV-1 across a tight human epithelial barrier and infection of subepithelial dendritic cells. Blood 120, 572–580. doi: 10.1182/blood-2011-08-374637
Matsuoka, M. and Green, P. L. (2009). The HBZ gene, a key player in HTLV-1 pathogenesis. Retrovirology 6, 71. doi: 10.1186/1742-4690-6-71
McCann, N., O’Connor, D., Lambe, T., and Pollard, A. J. (2022). Viral vector vaccines. Curr. Opin. Immunol. 77, 102210. doi: 10.1016/j.coi.2022.102210
Meissner, M. E., Mendonça, L. M., Zhang, W., and Mansky, L. M. (2017). Polymorphic nature of human T-cell leukemia virus type 1 particle cores as revealed through characterization of a chronically infected cell line. J. Virol. 91, e00369–e00317. doi: 10.1128/JVI.00369-17
Mendoza, C., Taylor, G., Gessain, A., Thoma-Kress, A. K., Bangham, C., Vesterbacka, J., et al. (2024). Virology, pathogenesis, epidemiology and clinical management of HTLV-1 infection. Proceedings of the 30th HTLV European research network (HERN 2023). NeuroImmune Pharmacol. Ther. 3, 61–69. doi: 10.1515/nipt-2023-0025
Meyerowitz, E. A., Mukerji, S. S., Kyle Harrold, G., Erdil, R. M., Chen, S. T., Rudmann, E. A., et al. (2023). Mogamulizumab for treatment of human T-lymphotropic virus type 1-associated myelopathy/tropical spastic paraparesis: A single-center US-based series. Clin. Infect. Dis. Off. Publ. Infect. Dis. Soc Am. 77, 851–856. doi: 10.1093/cid/ciad281
Mitagami, Y., Yasunaga, J.-I., Kinosada, H., Ohshima, K., and Matsuoka, M. (2015). Interferon-γ Promotes inflammation and development of T-cell lymphoma in HTLV-1 bZIP factor transgenic mice. PloS Pathog. 11, e1005120. doi: 10.1371/journal.ppat.1005120
Miyazawa, T., Hasebe, Y., Murase, M., Sakurai, M., Itabashi, K., and Yonemoto, N. (2021). The effect of early postnatal nutrition on human T cell leukemia virus type 1 mother-to-child transmission: A systematic review and meta-analysis. Viruses 13, 819. doi: 10.3390/v13050819
Mohanty, S. and Harhaj, E. W. (2020). Mechanisms of oncogenesis by HTLV-1 tax. Pathogens 9, 543. doi: 10.3390/pathogens9070543
Mohanty, S. and Harhaj, E. W. (2023). Mechanisms of innate immune sensing of HTLV-1 and viral immune evasion. Pathogens 12, 735. doi: 10.3390/pathogens12050735
Mohanty, S., Suklabaidya, S., Lavorgna, A., Ueno, T., Fujisawa, J., Ngouth, N., et al. (2024). The tyrosine kinase KDR is essential for the survival of HTLV-1-infected T cells by stabilizing the Tax oncoprotein. Nat. Commun. 15, 5380. doi: 10.1038/s41467-024-49737-5
Mühleisen, A., Giaisi, M., Köhler, R., Krammer, P. H., and Li-Weber, M. (2014). Tax contributes apoptosis resistance to HTLV-1-infected T cells via suppression of Bid and Bim expression. Cell Death Dis. 5, e1575–e1575. doi: 10.1038/cddis.2014.536
Murphy, E. L., Figueroa, J. P., Gibbs, W. N., Brathwaite, A., Holding-Cobham, M., Waters, D., et al. (1989). Sexual transmission of human T-lymphotropic virus type I (HTLV-I). Ann. Intern. Med. 111, 555–560. doi: 10.7326/0003-4819-111-7-555
Nakamura, H., Hayami, M., Ohta, Y., Ishikawa, K., Tsujimoto, H., Kiyokawa, T., et al. (1987). Protection of cynomolgus monkeys against infection by human T-cell leukemia virus type-I by immunization with viral env gene products produced in Escherichia coli. Int. J. Cancer 40, 403–407. doi: 10.1002/ijc.2910400320
Ngoma, A. M., Mutombo, P. B., Omokoko, M. D., Mvika, E. S., Nollet, K. E., and Ohto, H. (2019). Prevalence and molecular epidemiology of human T-lymphotropic virus type 1 among women attending antenatal clinics in sub-saharan africa: A systematic review and meta-analysis. Am. J. Trop. Med. Hyg. 101, 908–915. doi: 10.4269/ajtmh.19-0182
Nyborg, J. K., Egan, D., and Sharma, N. (2010). The HTLV-1 Tax protein: revealing mechanisms of transcriptional activation through histone acetylation and nucleosome disassembly. Biochim. Biophys. Acta 1799, 266–274. doi: 10.1016/j.bbagrm.2009.09.002
Ohashi, T., Hanabuchi, S., Kato, H., Tateno, H., Takemura, F., Tsukahara, T., et al. (2000). Prevention of adult T-cell leukemia-like lymphoproliferative disease in rats by adoptively transferred T cells from a donor immunized with human T-cell leukemia virus type 1 tax-coding DNA vaccine. J. Virol. 74, 9610–9616. doi: 10.1128/jvi.74.20.9610-9616.2000
Oliveira, P. D., Kachimarek, A. C., and Bittencourt, A. L. (2018). Early onset of HTLV-1 associated myelopathy/tropical spastic paraparesis (HAM/TSP) and adult T-cell leukemia/lymphoma (ATL): systematic search and review. J. Trop. Pediatr. 64, 151–161. doi: 10.1093/tropej/fmx039
Osame, M., Usuku, K., Izumo, S., Ijichi, N., Amitani, H., Igata, A., et al. (1986). HTLV-I associated myelopathy, a new clinical entity. Lancet Lond. Engl. 1, 1031–1032. doi: 10.1016/s0140-6736(86)91298-5
Osman, H. K. (2003). Humas T-cell lymphotropic virus type 1: Is it time to bcbeen organ donors and recipients? Transplantation 75, 1–2. doi: 10.1097/00007890-200301150-00001
Paiva, A. and Casseb, J. (2014). Sexual transmission of human T-cell lymphotropic virus type 1. Rev. Soc Bras. Med. Trop. 47, 265–274. doi: 10.1590/0037-8682-0232-2013
Peloponese, J.-M. and Jeang, K.-T. (2006). Role for akt/protein kinase B and activator protein-1 in cellular proliferation induced by the human T-cell leukemia virus type 1 tax oncoprotein *. J. Biol. Chem. 281, 8927–8938. doi: 10.1074/jbc.M510598200
Pépin, J., Labbé, A.-C., Mamadou-Yaya, F., Mbélesso, P., Mbadingaï, S., Deslandes, S., et al. (2010). Iatrogenic transmission of human T cell lymphotropic virus type 1 and hepatitis C virus through parenteral treatment and chemoprophylaxis of sleeping sickness in colonial Equatorial Africa. Clin. Infect. Dis. 51, 777–784. doi: 10.1086/656232
Pise-Masison, C. A. and Brady, J. N. (2005). Setting the stage for transformation: HTLV-1 Tax inhibition of p53 function. Front. Biosci. 10, 919–930. doi: 10.2741/1586
Radygina, L. V. and Mochalova, L. V. (2024). Diseases caused by human T-lymphotropic virus type 1 (HTLV-1). Microbiol. Indep. Res. J. 11, 57–69. doi: 10.18527/2024115769
Romanelli, M. G., Diani, E., Bergamo, E., Casoli, C., Ciminale, V., Bex, F., et al. (2013). Highlights on distinctive structural and functional properties of HTLV Tax proteins. Front. Microbiol. 4, 271. doi: 10.3389/fmicb.2013.00271
Roucoux, D. F., Wang, B., Smith, D., Nass, C. C., Smith, J., Hutching, S. T., et al. (2005). A prospective study of sexual transmission of human T lymphotropic virus (HTLV)–I and HTLV-II. J. Infect. Dis. 191, 1490–1497. doi: 10.1086/429410
Sagar, D., Masih, S., Schell, T., Jacobson, S., Comber, J. D., Philip, R., et al. (2014). In vivo immunogenicity of Tax(11-19) epitope in HLA-A2/DTR transgenic mice: implication for dendritic cell-based anti-HTLV-1 vaccine. Vaccine 32, 3274–3284. doi: 10.1016/j.vaccine.2014.03.087
Saito, K., Saito, M., Taniura, N., Okuwa, T., and Ohara, Y. (2010). Activation of the PI3K–Akt pathway by human T cell leukemia virus type 1 (HTLV-1) oncoprotein Tax increases Bcl3 expression, which is associated with enhanced growth of HTLV-1-infected T cells. Virology 403, 173–180. doi: 10.1016/j.virol.2010.04.018
Santana, C. S., de Andrade, F., da Silva, G. C. S., de Souza Nascimento, J. O., Campos, R. F., Giovanetti, M., et al. (2023). Advances in preventive vaccine development against HTLV-1 infection: A systematic review of the last 35 years. Front. Immunol. 14. doi: 10.3389/fimmu.2023.1073779
Sato, T., Coler-Reilly, A. L. G., Yagishita, N., Araya, N., Inoue, E., Furuta, R., et al. (2018). Mogamulizumab (Anti-CCR4) in HTLV-1–associated myelopathy. N. Engl. J. Med. 378, 529–538. doi: 10.1056/NEJMoa1704827
Sato, T., Nagai, M., Watanabe, O., Misu, T., Takenouchi, N., Ohkubo, R., et al. (2024). Multicenter, randomized, double-blind, placebo-controlled phase 3 study of mogamulizumab with open-label extension study in a minimum number of patients with human T-cell leukemia virus type-1-associated myelopathy. J. Neurol. 271, 3471–3485. doi: 10.1007/s00415-024-12239-x
Sato, T., Yamauchi, J., Yagishita, N., Araya, N., Takao, N., Ohta, Y., et al. (2023). Long-term safety and efficacy of mogamulizumab (anti-CCR4) for treating virus-associated myelopathy. Brain J. Neurol. 146, 3181–3191. doi: 10.1093/brain/awad139
Satou, Y., Yasunaga, J., Yoshida, M., and Matsuoka, M. (2006). HTLV-I basic leucine zipper factor gene mRNA supports proliferation of adult T cell leukemia cells. Proc. Natl. Acad. Sci. U. S. A. 103, 720–725. doi: 10.1073/pnas.0507631103
Seighali, N., Shafiee, A., Rafiee, M. A., Aminzade, D., and Mozhgani, S.-H. (2023). Human T-cell lymphotropic virus type 1 (HTLV-1) proposed vaccines: a systematic review of preclinical and clinical studies. BMC Infect. Dis. 23, 320. doi: 10.1186/s12879-023-08289-7
Seiki, M., Hattori, S., Hirayama, Y., and Yoshida, M. (1983). Human adult T-cell leukemia virus: complete nucleotide sequence of the provirus genome integrated in leukemia cell DNA. Proc. Natl. Acad. Sci. 80, 3618–3622. doi: 10.1073/pnas.80.12.3618
Shafifar, M., Mozhgani, S.-H., Razavi Pashabayg, K., Mosavat, A., Karbalaei, M., Norouzi, M., et al. (2022). Selective APC-targeting of a novel Fc-fusion multi-immunodominant recombinant protein (tTax-tEnv:mFcγ2a) for HTLV-1 vaccine development. Life Sci. 308, 120920. doi: 10.1016/j.lfs.2022.120920
Shida, H., Tochikura, T., Sato, T., Konno, T., Hirayoshi, K., Seki, M., et al. (1987). Effect of the recombinant vaccinia viruses that express HTLV-I envelope gene on HTLV-I infection. EMBO J. 6, 3379–3384. doi: 10.1002/j.1460-2075.1987.tb02660.x
Silic-Benussi, M., Biasiotto, R., Andresen, V., Franchini, G., D’Agostino, D. M., and Ciminale, V. (2010). HTLV-1 p13, a small protein with a busy agenda. Mol. Aspects Med. 31, 350–358. doi: 10.1016/j.mam.2010.03.001
Sousa-Pereira, D., Silva de Oliveira, T., Paiva, R. O., Chaves, O. A., Netto-Ferreira, J. C., Echevarria-Lima, J., et al. (2020). Synthetic (E)-3-phenyl-5-(phenylamino)-2-styryl-1,3,4-thiadiazol-3-ium chloride derivatives as promising chemotherapy agents on cell lines infected with HTLV-1. Molecules 25, 2537. doi: 10.3390/molecules25112537
Stienlauf, S., Yahalom, V., Schwartz, E., Shinar, E., Segal, G., and Sidi, Y. (2009). Epidemiology of human T-cell lymphotropic virus type 1 infection in blood donors, Israel. Emerg. Infect. Dis. 15, 1116–1118. doi: 10.3201/eid1507.080796
Suehiro, Y., Hasegawa, A., Iino, T., Sasada, A., Watanabe, N., Matsuoka, M., et al. (2015). Clinical outcomes of a novel therapeutic vaccine with Tax peptide-pulsed dendritic cells for adult T cell leukaemia/lymphoma in a pilot study. Br. J. Haematol. 169, 356–367. doi: 10.1111/bjh.13302
Sugata, K., Yasunaga, J., Miura, M., Akari, H., Utsunomiya, A., Nosaka, K., et al. (2016). Enhancement of anti-STLV-1/HTLV-1 immune responses through multimodal effects of anti-CCR4 antibody. Sci. Rep. 6, 27150. doi: 10.1038/srep27150
Sundaram, R., Lynch, M. P., Rawale, S. V., Sun, Y., Kazanji, M., and Kaumaya, P. T. P. (2004). De novo design of peptide immunogens that mimic the coiled coil region of human T-cell leukemia virus type-1 glycoprotein 21 transmembrane subunit for induction of native protein reactive neutralizing antibodies*. J. Biol. Chem. 279, 24141–24151. doi: 10.1074/jbc.M313210200
Sundaram, R., Sun, Y., Walker, C. M., Lemonnier, F. A., Jacobson, S., and Kaumaya, P. T. P. (2003). A novel multivalent human CTL peptide construct elicits robust cellular immune responses in HLA-A∗0201 transgenic mice: implications for HTLV-1 vaccine design. Vaccine 21, 2767–2781. doi: 10.1016/S0264-410X(03)00179-8
Tabakin-Fix, Y., Azran, I., Schavinky-Khrapunsky, Y., Levy, O., and Aboud, M. (2006). Functional inactivation of p53 by human T-cell leukemia virus type 1 Tax protein: mechanisms and clinical implications. Carcinogenesis 27, 673–681. doi: 10.1093/carcin/bgi274
Tan, B. J. Y., Sugata, K., Ono, M., and Satou, Y. (2022). HTLV-1 persistence and leukemogenesis: A game of hide-and-seek with the host immune system. Front. Immunol. 13. doi: 10.3389/fimmu.2022.991928
Tanaka, Y., Tanaka, R., Terada, E., Koyanagi, Y., Miyano-Kurosaki, N., Yamamoto, N., et al. (1994). Induction of antibody responses that neutralize human T-cell leukemia virus type I infection in vitro and in vivo by peptide immunization. J. Virol. 68, 6323–6331. doi: 10.1128/jvi.68.10.6323-6331.1994
Tanaka-Nakanishi, A., Yasunaga, J., Takai, K., and Matsuoka, M. (2014). HTLV-1 bZIP factor suppresses apoptosis by attenuating the function of FoxO3a and altering its localization. Cancer Res. 74, 188–200. doi: 10.1158/0008-5472.CAN-13-0436
Tsukasaki, K. and Tobinai, K. (2013). Biology and treatment of HTLV-1 associated T-cell lymphomas. Best Pract. Res. Clin. Haematol. 26, 3–14. doi: 10.1016/j.beha.2013.04.001
Tu, J. J., King, E., Maksimova, V., Smith, S., Macias, R., Cheng, X., et al. (2024). An HTLV-1 envelope mRNA vaccine is immunogenic and protective in New Zealand rabbits. J. Virol. 98, e01623–e01623. doi: 10.1128/jvi.01623-23
Uchiyama, T. (1997). Human T cell leukemia virus type I (HTLV-I) and human diseases. Annu. Rev. Immunol. 15, 15–37. doi: 10.1146/annurev.immunol.15.1.15
Van Tienen, C., McConkey, S. J., De Silva, T. I., Cotten, M., Kaye, S., Sarge-Njie, R., et al. (2012). Maternal proviral load and vertical transmission of human T cell lymphotropic virus type 1 in Guinea-bissau. AIDS Res. Hum. Retroviruses 28, 584–590. doi: 10.1089/aid.2011.0219
Verdonck, K., González, E., Dooren, S. V., Vandamme, A.-M., Vanham, G., and Gotuzzo, E. (2007). Human T-lymphotropic virus 1: recent knowledge about an ancient infection. Lancet Infect. Dis. 7, 266–281. doi: 10.1016/S1473-3099(07)70081-6
Vernin, C., Thenoz, M., Pinatel, C., Gessain, A., Gout, O., Delfau-Larue, M.-H., et al. (2014). HTLV-1 bZIP factor HBZ promotes cell proliferation and genetic instability by activating oncomiRs. Cancer Res. 74, 6082–6093. doi: 10.1158/0008-5472.CAN-13-3564
Vesterbacka, J., Svensson, A.-K., and Nowak, P. (2024). HTLV in Sweden. AIDS Rev. 26, 13647. doi: 10.24875/AIDSRev.24000002
Weiss, S. H., Saxinger, W. C., Rechtman, D., Grieco, M. H., Nadler, J., Holman, S., et al. (1985). HTLV-III infection among health care workers: association with needle-stick injuries. JAMA 254, 2089–2093. doi: 10.1001/jama.1985.03360150065025
Yara, S., Fujita, J., and Date, H. (2009). Transmission of human T-lymphotropic virus type I by bilateral living-donor lobar lung transplantation. J. Thorac. Cardiovasc. Surg. 138, 255–256. doi: 10.1016/j.jtcvs.2008.12.050
Yoshida, M., Satou, Y., Yasunaga, J., Fujisawa, J., and Matsuoka, M. (2008). Transcriptional control of spliced and unspliced human T-cell leukemia virus type 1 bZIP factor (HBZ) gene. J. Virol. 82, 9359–9368. doi: 10.1128/jvi.00242-08
Zhao, T. (2016). The role of HBZ in HTLV-1-induced oncogenesis. Viruses 8, 34. doi: 10.3390/v8020034
Zhao, T. and Matsuoka, M. (2012). HBZ and its roles in HTLV-1 oncogenesis. Front. Microbiol. 3. doi: 10.3389/fmicb.2012.00247
Zhao, T., Wang, Z., Fang, J., Cheng, W., Zhang, Y., Huang, J., et al. (2022). HTLV-1 activates YAP via NF-κB/p65 to promote oncogenesis. Proc. Natl. Acad. Sci. U. S. A. 119, e2115316119. doi: 10.1073/pnas.2115316119
Keywords: HTLV-1, human T-cell leukemia virus type 1, oncogenesis, viral oncogenes, immune evasion, retrovirus, cancer
Citation: Vasconez-Gonzalez J, Suárez-Sangucho IA, Acosta-Muñoz E, Miño LPy, Borja-Mendoza D, Alexander-Castillo JA, Saa J, Salazar-Calvopiña N, Cárdenas P, López-Cortés A and Ortiz-Prado E (2025) Human T-cell leukemia virus type 1: oncogenic potential and vaccine development strategies. Front. Cell. Infect. Microbiol. 15:1587802. doi: 10.3389/fcimb.2025.1587802
Received: 05 March 2025; Accepted: 17 July 2025;
Published: 15 August 2025.
Edited by:
Robert Breiman, Emory University, United StatesReviewed by:
Yue Han, Shanghai Jiao Tong University, ChinaSuchitra Mohanty, The Pennsylvania State University, United States
Copyright © 2025 Vasconez-Gonzalez, Suárez-Sangucho, Acosta-Muñoz, Miño, Borja-Mendoza, Alexander-Castillo, Saa, Salazar-Calvopiña, Cárdenas, López-Cortés and Ortiz-Prado. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Esteban Ortiz-Prado, ZXN0ZWJhbi5vcnRpei5wcmFkb0B1ZGxhLmVkdS5lYw==