 Ana Fernández-Bravo1
Ana Fernández-Bravo1 Laura Camuña-Pardo1†
Laura Camuña-Pardo1† Marta Sanchis1,2†
Marta Sanchis1,2† Youssef Ahmiane1†
Youssef Ahmiane1† Javier Capilla1
Javier Capilla1 Josepa Gené1*
Josepa Gené1*- 1Mycology and Environmental Microbiology Unit, Faculty of Medicine and Health Science, Rovira i Virgili University, & Pere Virgili Health Research Institute (IISPV), Reus, Spain
- 2Serra Húnter Fellow, Mycology and Environmental Microbiology Unit, Faculty of Medicine and Health Science, Rovira i Virgili University, & Pere Virgili Health Research Institute (IISPV), Reus, Spain
Phialophora submersa is a recently described black yeast species (Chaetothyriales), isolated from freshwater sediments in Catalonia (Spain). It is closely related to P. americana and P. verrucosa, two opportunistic pathogens known to cause subcutaneous infections in humans and animals. This study investigates the pathogenic potential of P. submersa, its in vitro susceptibility to clinically relevant antifungal agents, and its response to various cellular stressors. Using a murine macrophage (J774A.1) infection model, we evaluated phagocytosis, intracellular survival, cell damage, and the expression of six immune-related genes (TNF-α, CCL20, RELA, TP53, NLRP3, IL-1β), in comparison with P. americana and P. verrucosa. The results showed that P. submersa induced higher phagocytosis rates in murine macrophages than the P. verrucosa, although lower than P. americana. Cell damage, intracellular survival, and expression of the immune-related genes were higher after macrophage infection with P. verrucosa than with P. submersa and P. americana, which exhibited comparable profiles. All three species displayed similar antifungal susceptibility profiles, being susceptible to most azoles (except fluconazole), terbinafine, and echinocandins (with reduced efficacy against P. verrucosa), but showed moderate resistance to flucytosine, amphotericin B, and olorofim. The resistance of P. submersa to stress was strain-dependent, with only one strain exhibiting notable resistance to multiple stressors. This research provides new insights into the biology of P. submersa, including its potential as a human pathogen, and the molecular factors that could drive an infection process.
Introduction
Phialophora is an ascomycetous genus belonging to the family Herpotrichiellaceae within the order Chaetothyriales. It comprises a group of melanized species characterized by brown, septate, and branched hyphae, and the production of conidia from flask-shaped or elongated conidiogenous cells with characteristic vase- to funnel-shaped darkly pigmented collarettes (De Hoog et al., 2020). Most Phialophora species are commonly found in the environment, where they play a role in the decomposition of plant material; however, they are also recognized for their pathogenic potential in humans (Li et al., 2017; De Hoog et al., 2020; Song et al., 2021). Among them, P. verrucosa and P. americana are well-documented causative agents of subcutaneous phaeohyphomycosis in both humans and animals. These species have also been associated, albeit less frequently, with chromoblastomycosis, a chronic fungal infection characterized by verrucous skin lesions containing brownish multiseptated fungal cells in the subcutaneous tissue (Singh et al., 1997; Borrás et al., 2022; Martini et al., 2022; Yiallouris et al., 2024). Phialophora infections usually occur through traumatic inoculation of fungal elements via skin, which are able to survive and proliferate into the tissue in immunocompromised, but also in apparently healthy individuals (Tong et al., 2013; De Hoog et al., 2020; Ahmed et al., 2021; Ply et al., 2023). Other opportunistic species in the genus recently associated with human disease are P. chinensis, P. expanda, P. macrospora, and P. tarda (De Hoog et al., 2020; Ahmed et al., 2021; Pruvot et al., 2022; Ply et al., 2023). Notably, multiple clinical cases involving Phialophora species have been linked to patients with immune system impairments, particularly those involving CARD9 (Caspase Recruitment Domain-containing protein 9) deficiencies (Liang et al., 2015; Ahmed et al., 2021; Song et al., 2021).
In addition to the clinical relevance, Phialophora species have been investigated for their ecological functions, particularly their capacity to degrade lignin and other complex organic compounds in their natural environments (Kwasna et al., 2021; Yasanthika et al., 2025). This dual role of Phialophora, as both environmental decomposers and opportunistic pathogens, makes the genus particularly intriguing from both medical mycology and environmental microbiology perspectives (Yiallouris et al., 2024). The presence of melanin in the cell wall enables Phialophora species to thrive in hostile environments, including low oxygen conditions and high irradiation exposure, and likely enhances their resistance to certain host immune responses (Guarro et al., 1999; Passero et al., 2021).
A recent survey exploring ascomycete diversity in freshwater sediments revealed Phialophora strains in both oligotrophic and eutrophic areas of several Spanish rivers, thereby broadening our understanding of the ecological niches occupied by potentially pathogenic fungi (Torres-Garcia et al., 2023). Approximately half of the isolates were identified as P. americana, while the remaining isolates represented a new lineage for the genus that was named P. submersa. Despite the genetic divergence, the two species exhibited only subtle morphological differences to be distinguished (Torres-Garcia et al., 2023). However, variations in their cardinal temperatures for growth were noted; P. americana displayed the optimum and maximum temperatures at 30 and 37 °C, as is typical for many other species in the genus, whereas P. submersa these were 25 and 30 °C, respectively (Li et al., 2017; Torres-Garcia et al., 2023). Irrespective of the lack of in vitro thermotolerance at 35─37 °C of P. submersa, but considering the clinical relevance of the Phialophora species, we consider interesting to investigate the pathogenic potential of P. submersa, particularly in comparison to the well-established human opportunists P. verrucosa and P. americana.
This study investigates the pathogenic mechanisms of the environmental species P. submersa, focusing on its ability to survive phagocytosis, trigger immune responses, and adapt to diverse conditions. By comparing it with P. verrucosa and P. americana, we highlight key similarities and differences in pathogenicity among Phialophora species. We also evaluate its susceptibility to commercial antifungal drugs and its response to stressors affecting the cell wall, membrane, and osmotic balance. These findings offer new insights into the biology of P. submersa and its potential role as a human pathogen.
Materials and methods
Phialophora strains
All Phialophora strains used in this study were isolated from freshwater sediments, as described by Torres-García et al (Torres-Garcia et al., 2023), with the exception of P. verrucosa, which was sourced from the fungal collection at the Westerdijk Fungal Biodiversity Institute (CBS, Utrecht, the Netherlands). The former strains have been deposited in the fungal culture collection of the Medicine Faculty at the Rovira i Virgili University (FMR, Reus, Spain), and their metadata are summarized in Table 1.
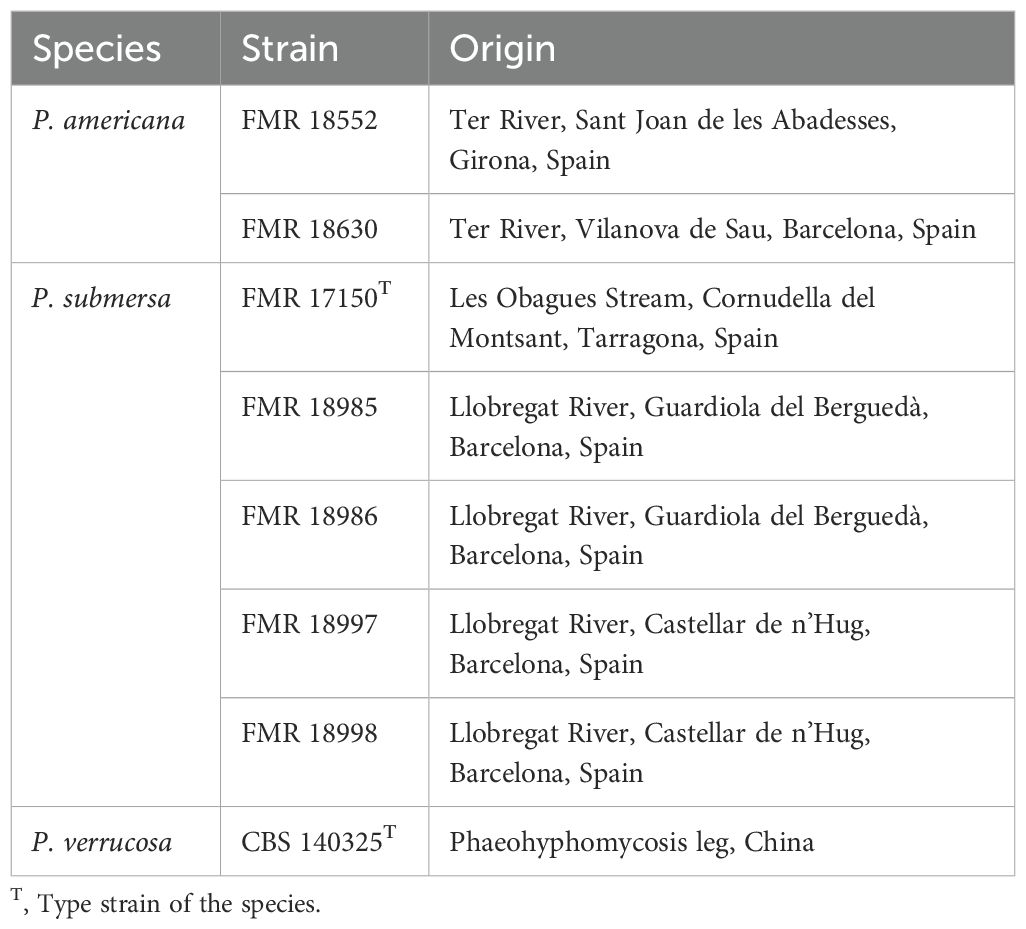
Table 1. Phialophora strains used in the study.
Cell lines and conditions
The J774.1A mouse macrophage cell line (BALB/C) was chosen for the infection experiments. Cells were cultured as adherent cells in DMEM (PAA Laboratories GmbH), supplemented with 10% fetal bovine serum (FBS, PAA Laboratories GmbH, Munich, Germany) and 1% penicillin-streptomycin (P/S, PAA Laboratories GmbH, Munich, Germany), and incubated at 37°C with 5% CO2. Prior to infection assays, cells were seeded in tissue culture plates using DMEM lacking FBS and P/S at a density of 0.5 × 106 cells/mL, aiming to achieve a final concentration of 1 × 106 cells/mL after 24 h (Fernández-Bravo and Figueras, 2022).
Infection
The J774.1A macrophage cell line was infected with P. americana (FMR 18552, FMR 18630), P. submersa (FMR 17150, FMR 18986), and P. verrucosa (CBS 140325) (Table 1). Fungal strains were grown overnight in DMEM without FBS and P/S, at a multiplicity of infection (MOI) of 10, representing the ratio of conidia to target cells. Cultures were incubated at 37°C in a 5% CO2 atmosphere and sampled at specific time points depending on the experimental conditions.
Phagocytosis and intracellular survival
J7741.A cells were seeded in 24-well plates and infected with the different Phialophora strains at an MOI of 10 and were incubated for 1 h at 37°C with 5% CO2. After infection, macrophages were washed with PBS, and the number of colonies associated with the cells was determined by serial dilution and plating. Infected macrophages were washed to eliminate extracellular conidia. Following the wash, the number of intracellular conidia was determined. The percentage of phagocytosis was calculated based on the colony counts after 1 h post-infection in comparison with the inoculum (Fernández-Bravo et al., 2019).
For the intracellular survival assay, macrophages were infected with the strains as described, and fungal loads were measured 1 h post-infection. The remaining infected macrophages were then incubated in fresh DMEM for an additional 4 to 6 h at 37°C with 5% CO2. After incubation, cells were washed with PBS, and the number of intracellular conidia was counted. The percentage of fungal survival was calculated based on the number of colonies after the 4 to 6 h incubation period in comparison with 1 h post-infection (Fernández-Bravo et al., 2019).
Cell damage assay
After infection of J7741.A cells at an MOI of 10 with the cultures, supernatants were collected at 3 and 5 h post-infection. Cell damage was assessed by quantifying the lactate dehydrogenase (LDH) enzyme released into the supernatants using the Cytox 96 Non-Radioactive Cytotoxicity Assay (Promega, Madison, WI, USA), following the manufacturer’s instructions. A standard curve was generated using bovine recombinant LDH (Sigma-Aldrich, St. Louis, MO, USA), and LDH levels in the samples were determined by extrapolation from this curve (Fernández-Bravo et al., 2024).
Analysis of the expression of the genes related to the immune system
Six genes known to be involved in the innate immune response against extracellular pathogens were chosen to assess their transcriptional activity in J774A.1 cells after exposure to Phialophora strains, with results compared to non-infected cells. The primers used for gene expression analysis were those described by Murciano et al. (Murciano et al., 2015) and Zhao et al. (Zhao et al., 2017), as listed in Table 2. The genes selected encode for cytokines and chemokines (TNF-α and CCL20), apoptosis (TP53), transcription factors (RELA), and pyroptosis-related proteins (NLRP3 and IL-1β). After infecting the J774A.1 cells for 4 h at a MOI of 10, the cells were washed twice with PBS. RNA was then extracted using the GenElute™ Mammalian Total RNA Miniprep Kit (Sigma-Aldrich). cDNA synthesis from the total RNA was performed using the iScript cDNA Synthesis Kit (Bio-Rad Laboratories, Inc., Hercules, CA, USA). Quantitative real-time PCR was carried out on the cDNA with Power SYBR® Green PCR Mastermix (Applied Biosystems®, Life Technologies, Glasgow, UK), using a StepOnePlus™ real-time PCR system (Applied Biosystems®). Thermal cycling conditions were as follows: initial 5-minute denaturation at 94°C, followed by 40 cycles consisting of 30 seconds at 94°C, 30 seconds at 61°C, 30 seconds at 72°C, and a final extension of 20 seconds at 80°C. The threshold cycle (CT) values were used to calculate the relative RNA expression of the target genes, with glyceraldehyde-3-phosphate dehydrogenase (GAPDH) serving as the reference housekeeping gene. Relative gene expression was determined using the delta-delta Ct (2–ΔΔCt) method, based on real-time PCR signal quantification, as previously described (Fernández-Bravo and Figueras, 2022).
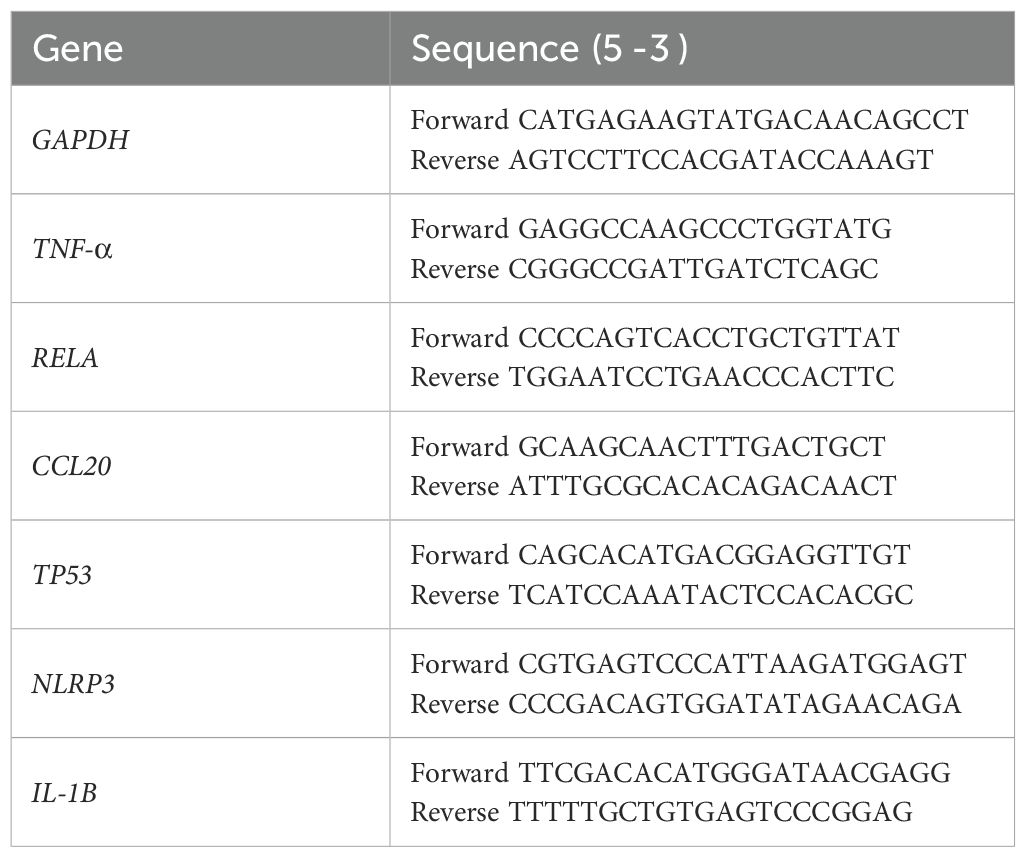
Table 2. Primers used to target gene expression (Murciano et al., 2015; Fernández-Bravo and Figueras, 2022).
Detection of Caspase 1 by Western blotting
Macrophages were infected with Phialophora spp. (MOI 10) for 3 h under standard culture conditions. After incubation, cells were washed twice with cold PBS and lysed on ice using RIPA buffer (Thermo Scientific) supplemented with protease inhibitor cocktail (Roche) and 1 mM PMSF. Lysates were incubated on ice for 30 minutes with occasional vortexing, and centrifuged at 14,000 × g for 15 minutes at 4°C. The supernatants were collected, and protein concentration was determined using a BCA protein assay kit (Thermo Scientific, Life Technologies). Equal amounts of total protein (20–30 µg) were mixed with Laemmli sample buffer containing 5% β-mercaptoethanol, boiled at 95°C for 5 minutes, and separated on a 12–15% SDS-PAGE gel. Proteins were transferred onto a PVDF membrane (Millipore, Madrid, Spain) using a wet transfer system at 100 V for 90 minutes at 4 °C.
Membranes were blocked in 5% non-fat dry milk in TBS-T (Tris-buffered saline with 0.1% Tween-20) for 1 h at room temperature, then incubated overnight at 4°C with primary antibodies against cleaved Caspase 1 (Casp 1) p20 (Bio-Rad, Madrid, Spain). After washing, membranes were incubated with HRP-conjugated secondary antibodies (anti-rabbit IgG, 1:3000) for 1 h at room temperature. Band intensities were measured with the ImageJ software program (NIH, MD, USA) (Fernández-Bravo and Figueras, 2022). GAPDH was used as a loading control in the Western blot.
Biofilm formation
To investigate biofilm formation following macrophage infection, J774A.1 macrophage cells were seeded in 24-well plates at a density of 1x105 cells per well, and incubated overnight at 37°C with 5% CO2 to allow cell attachment. The following day, the cells were infected with the Phialophora strains at MOI 10 for 1 h. After the incubation period, macrophages were washed twice with PBS to remove unbound conidia. Fresh culture medium was then added, and the cells were incubated for an additional 24 h to promote biofilm development.
To assess biofilm formation, the cells were washed gently with PBS to eliminate non-adherent conidia. Biofilms were stained with a 0.1% Gentian violet solution for 20─30 min at room temperature. After staining, excess dye was removed by washing with PBS. The attached biofilm was then solubilized by adding 95% ethanol to each well and incubating for 30 min at room temperature to release the Gentian violet. The optical density (OD) of the solubilized biofilm was measured at 570 nm using a microplate reader to quantify the biofilm biomass (Soussa et al., 2022).
Antifungal susceptibility pattern
The in vitro susceptibility of Phialophora spp. was performed using broth microdilution method according to the reference Clinical and Laboratory Standards Institute (CLSI) guidelines, document M38 3rd edition (Clinical and Laboratory Standards Institute (CLSI), 2017). Aspergillus flavus ATCC204304 was included as a quality control strain. Minimum inhibitory concentrations (MICs) were determined for fluconazole (FLZ, Sigma-Aldrich, St. Louis, MO, EEUU), itraconazole (ITZ, St. Louis, MO, EEUU), posaconazole (PSZ, Sigma-Aldrich, St. Louis, MO, EEUU), voriconazole (VRZ, Sigma-Aldrich, St. Louis, MO, EEUU), amphotericin B (AMB, Sigma-Aldrich, St. Louis, MO, EEUU), 5-fluorocytosine (5-FC, Sigma-Aldrich, St. Louis, MO, EEUU), olorofim (OLF, F2G, Manchester, UK), and terbinafine (TRB, Sigma-Aldrich, St. Louis, MO, EEUU). For the echinocandins, anidulafungin (AND, Pfizer Inc., Madrid, Spain), caspofungin (CSP, Merk & Co., Inc., Rahway, EEUU), and micafungin (MCF, Sigma-Aldrich, St. Louis, MO, EEUU) minimal effective concentrations (MECs) were determined. MICs and MECs were read after 8 days of incubation at 30°C, considering the maximum temperature for growth of P. submersa (Torres-Garcia et al., 2023).
Stress sensitivity assay
We assessed the cellular stress sensitivity of five P. submersa strains (Table 1). To do so, 10 μl of conidial suspensions (5x105 conidia/mL) were cultured by inoculating a single drop onto supplemented potato dextrose agar (PDA) plates and incubated for two weeks. Osmotic stress was tested by supplementing PDA with sorbitol (2M and 3M), potassium chloride (0,5M and 1M), or sodium chloride (0,5M and 1M). Cell wall-destabilizing stress was induced using calcofluor white (CFW; 200 and 250 μg/mL) and Congo Red (CR; 1 and 1,25 mg/mL). Membrane stress conditions were done with sodium dodecyl sulphate (SDS; 50 and 100 μg/mL). Sensitivity to the different stresses was evaluated by measuring radial growth after two weeks of incubation at 25°C using the software ImageJ (version 1.54g).
Statistical analysis
All experiments were replicated at least 3 times. Statistical analyses were performed by using t-test and 1-way ANOVA with Tukey post hoc test, or by 2-way ANOVA with Sidak post hoc test with GraphPad Prism 6.0 (GraphPad Software, CA, USA). P value of <0.05 was considered statistically significant.
Results
Phagocytosis and intracellular survival after macrophage infections with the fungal strains
The phagocytic activity of J774A.1 macrophages was significantly influenced by the Phialophora species involved in the infection. Upon infection with P. verrucosa, the percentage of phagocytosis was notably lower compared to the other two species tested (p > 0.05). In contrast, infection with P. americana led to the highest levels of phagocytosis, with macrophages efficiently engulfing the fungal cells. The phagocytic activity observed in macrophages infected with P. submersa was intermediate, with phagocytosis levels falling between those of P. verrucosa and P. americana (Figure 1).
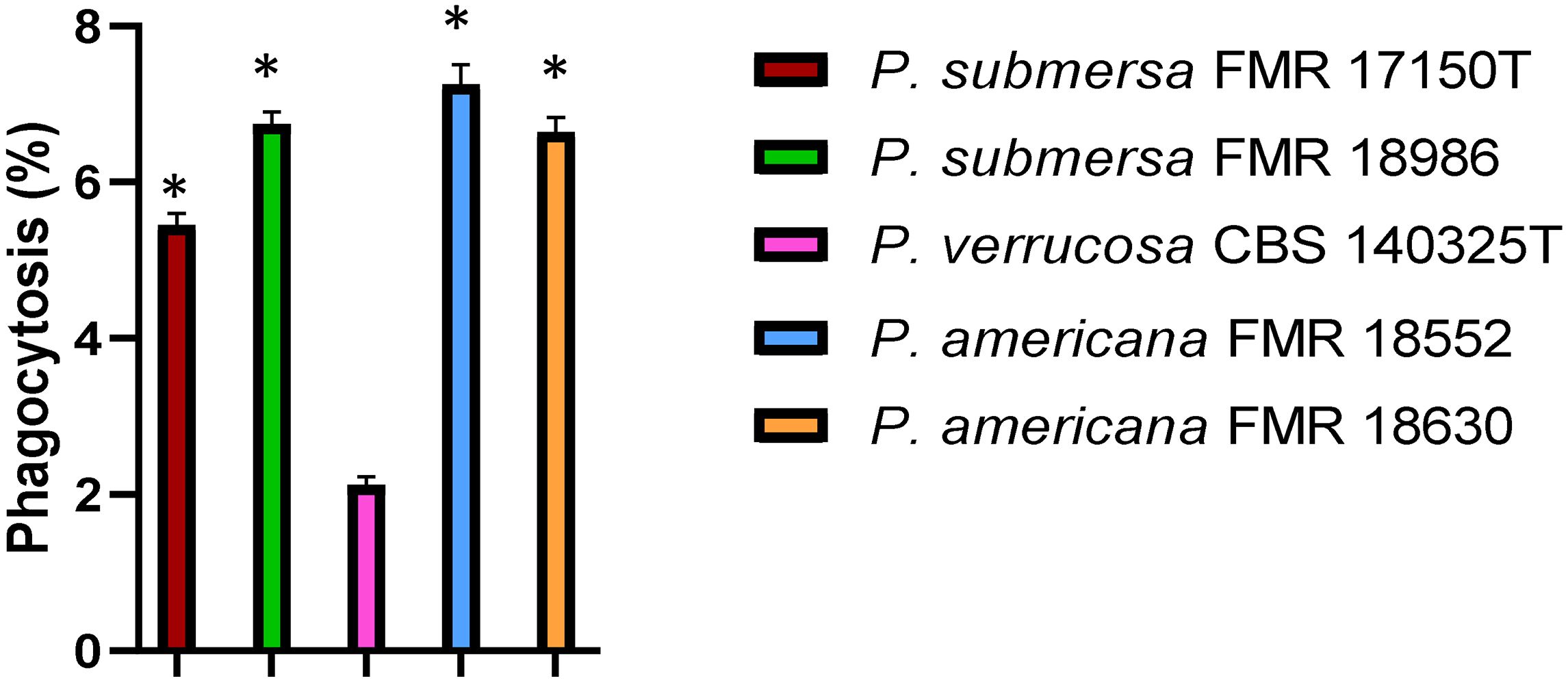
Figure 1. Phagocytosis assay after infection with the Phialophora strains at 1 h post-infection at MOI 10. Data were analyzed by using the Student’s two-tailed t-test and by 1- way ANOVA with Tukey post hoc. Asterisks indicate statistically a significant difference P < 0.05*.
Intracellular survival was assessed at 4 and 6 h post-infection, revealing that, regardless of the infecting Phialophora species, survival was higher at 4 h and decreased significantly by 6 h (p < 0.05). Among the species tested, P. verrucosa showed the highest intracellular survival, followed by P. submersa, with P. americana, irrespective of the strain used (p < 0.05) (Figure 2).
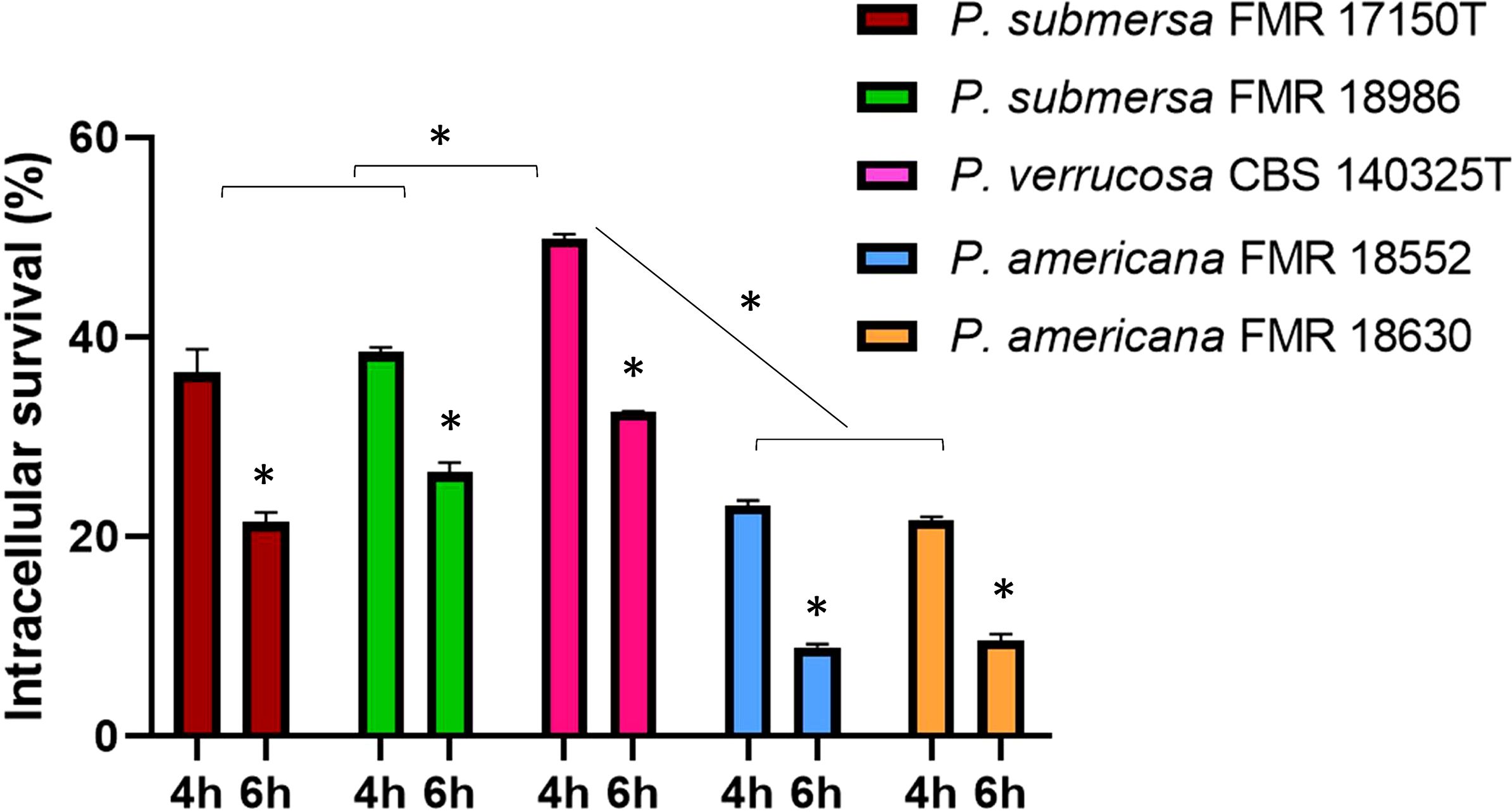
Figure 2. Intracellular survival after infection with the Phialophora strains at 4 and 6 h post-infection at MOI 10. Data were analyzed by using the 2- way ANOVA with Sidak post hoc test. Asterisks indicate a statistically significant difference P < 0.05*.
Cell damage after macrophage infection at different times
Cellular damage, measured by LDH enzyme release, was lower at 3 h post-infection but increased significantly by 5 h (p < 0.05), regardless of the Phialophora species infecting the macrophages. Among the species tested, the highest level of cellular damage was observed following infection with P. verrucosa, followed by P. submersa, and finally P. americana, irrespective of the strain used (Figure 3).
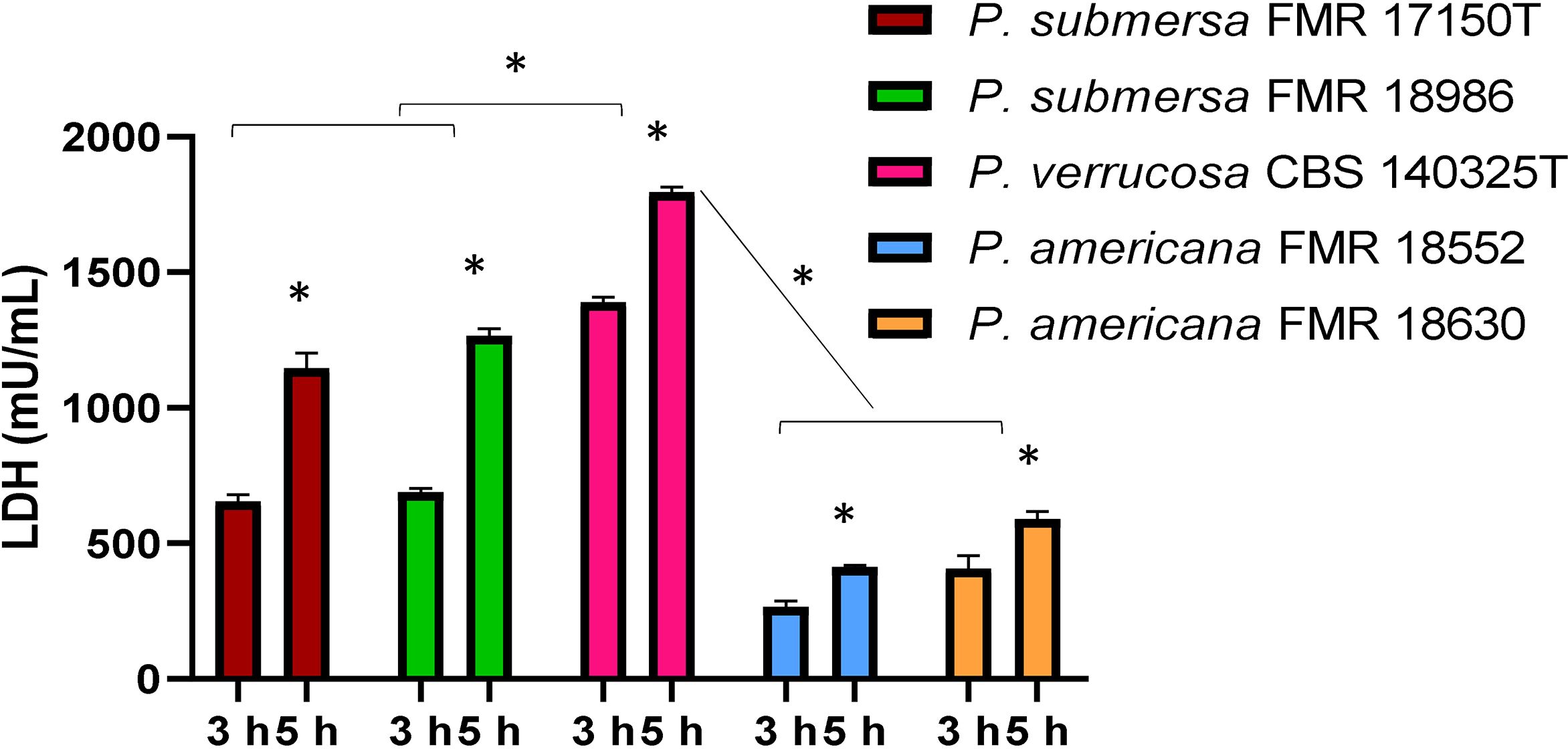
Figure 3. Detected J774.1A cell damage induced at 3 and 5 h by the Phialophora infections at MOI 10 in cells, measured by the release of lactate dehydrogenase (LDH) enzyme. Data were analyzed by using the 2- way ANOVA with Sidak post hoc test. Asterisks indicate a statistically significant difference P < 0.05*.
Macrophage cell line (J774.1A) gene-expression after infection with the fungal strains
Genes for cytokines and chemokines
All infections resulted in the expression of genes encoding the cytokine TNF-α, and the chemokine CCL20 in J774A.1 macrophages, with significant differences (p < 0.05) in transcription patterns compared to non-infected cells (Figure 4). Furthermore, the expression of TNF-α and CCL20 were notably higher following infection with P. verrucosa compared to the strains of P. submersa and P. americana (p < 0.05). Additionally, the expression levels of both genes were significantly higher after infection with P. submersa compared to P. americana. No significant differences were found between strains of the same species (Figure 4).
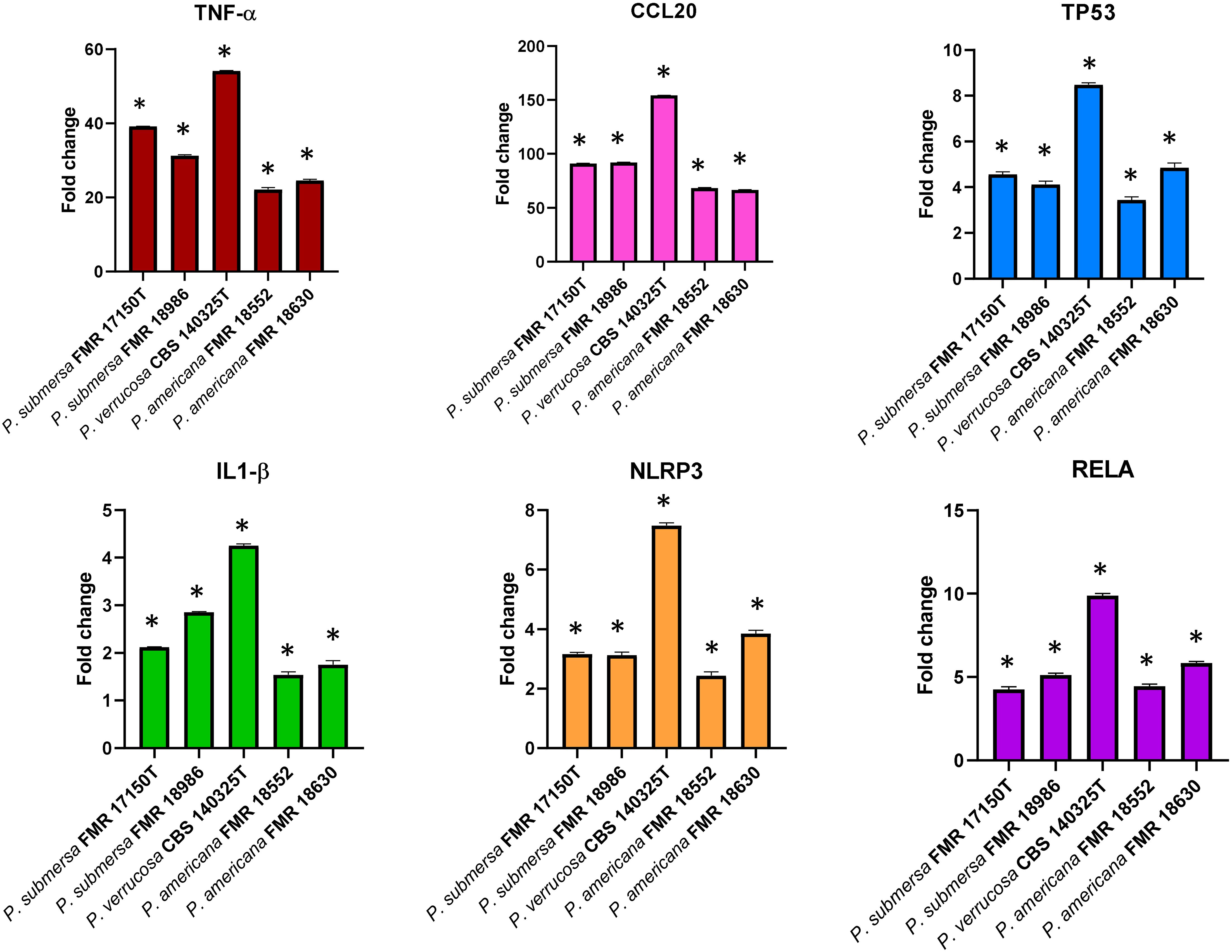
Figure 4. Gene expression profile of J7741. A cells in relation to the non-infected cells induced by Phialophora infections at MOI 10 determined by RT-qPCR. Transcript levels of the genes were normalized to the expression of the GAPDH gene. Expression fold change with respect to the non-infected cells was calculated using the comparative ΔΔCt method. Data were analyzed by using the Student’s two-tailed t-test and by 2- way ANOVA. Asterisks indicate a statistically significant difference P < 0.05*.
Gene involved in apoptosis
The transcriptional expression of the TP53 apoptosis gene was significantly elevated after infection with all three species in comparison with the non-infected cells. P. verrucosa caused a greater overexpression of TP53 compared with P. submersa and P. americana (p <0.05) (Figure 2). No significant differences in TP53 expression were observed between infections with P. americana and P. submersa, independently of the strain used (Figure 4).
Genes related to the inflammasome and pyroptosis
The NLRP3 and IL-1β genes, which are involved in pyroptosis, a type of cell death mediated by inflammasome activation, were significantly upregulated in J774A.1 cells following infections with all strains compared to non-infected controls (p < 0.05). The expression levels of NLPR3 were significantly highest after P. verrucosa infection, followed by P. submersa and P. americana. The expression levels of IL-1β were also higher after P. verrucosa infection, but were similar after infection with P. americana and P. submersa (Figure 4).
Genes related to the transcription factor RELA
All infections resulted in higher expression levels of the RELA gene, which is related to cytokine production, in comparison with non-infected cells (p < 0.05). However, P. verrucosa infection led to the highest overexpression of the RELA gene, followed by P. submersa and P. americana. Regardless of the strains, these levels were higher after infection with P. submersa than P. americana (Figure 4).
Detection of Casp 1 after Phialophora infection
The activation of Casp 1 were analyzed by Western blotting using an antibody that recognized the protein. The results demonstrated that, in macrophages J7741.A, all strains induced de activation of Casp 1 at 3 h postinfection. However, the band intensity was higher after infection with P. verrucosa (Figure 5).

Figure 5. Activation of Caspase 1 by Phialophora strains in macrophages measured by Western blotting.
Biofilm formation assay with gentian violet staining after macrophage infection
Biofilm formation by Phialophora strains on previously adherent macrophage cultures was assessed by Gentian violet staining. The results showed that the P. verrucosa strain produced significantly more biofilm (p > 0.05) compared to the other strains tested, while P. submersa strains exhibited a higher biofilm production than those of P. americana (Figure 6). This test demonstrated that Phialophora species have a notable capacity for biofilm formation, with production levels varying among species, and P. americana displaying the lowest biofilm-forming ability.
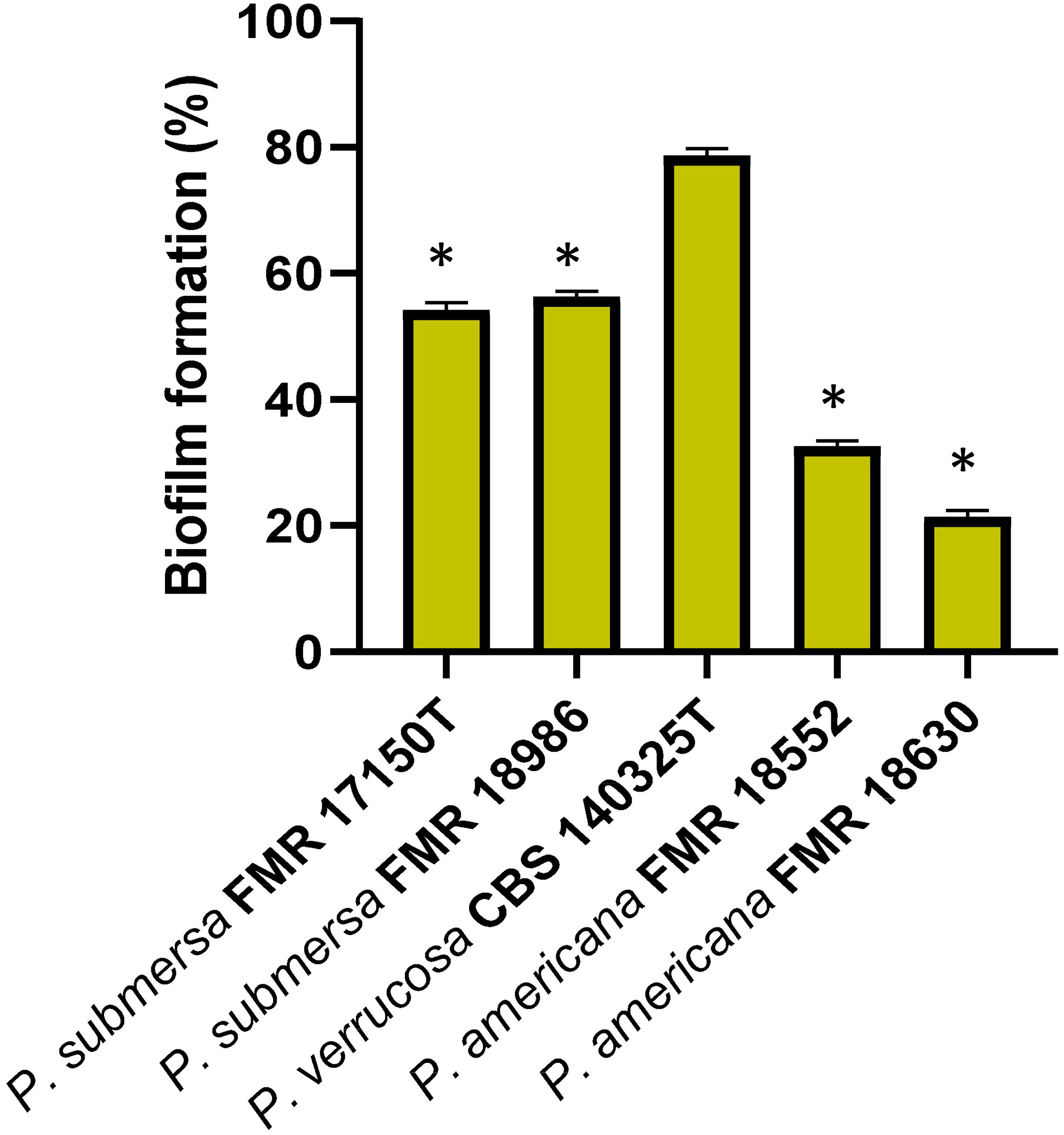
Figure 6. Percentage of biofilm formation after 24 h post-infection with the Phialophora strains. Data were analyzed by using the Student’s two-tailed t-test and by 1- way ANOVA with Tukey post hoc. Asterisks indicate statistically significant difference P < 0.05*.
Antifungal susceptibility
The antifungal susceptibility results for the three Phialophora species (Table 3) showed that TBF exhibited the highest antifungal activity against all strains tested, with MIC values ranging from 0.12 µg/mL to 0.5 µg/mL. The lowest MIC value was observed against P. verrucosa (MIC = 0.12 µg/mL). All azoles demonstrated potent antifungal activity (MIC ≤ 0.5 µg/mL) across the species tested, with the exception of FLZ, which showed poor activity (MIC ≥ 64 µg/mL), and a single P. americana strain (FMR18630) that exhibited a MIC of 1 µg/ml for VRZ. The modal MIC of AMB was 2 µg/ml against P. submersa, while P. americana and P. verrucosa showed slightly higher resistance, with a MIC of 4 µg/mL. Susceptibility to OLF and 5-FC was also low in all Phialophora strains tested, with MIC values ranging from 1 to ≥16 µg/mL. Echinocandins showed strain-dependent MECs, with values ranging from 0.06 to 4 µg/mL across all species, except for P. verrucosa, which was resistant to the three tested echinocandins (MEC ≥ 16 µg/mL). Notably, MCF was the only echinocandin that demonstrated antifungal activity against the P. americana and P. submersa strains tested, with MIC values between 0.06 and 0.5 µg/mL.
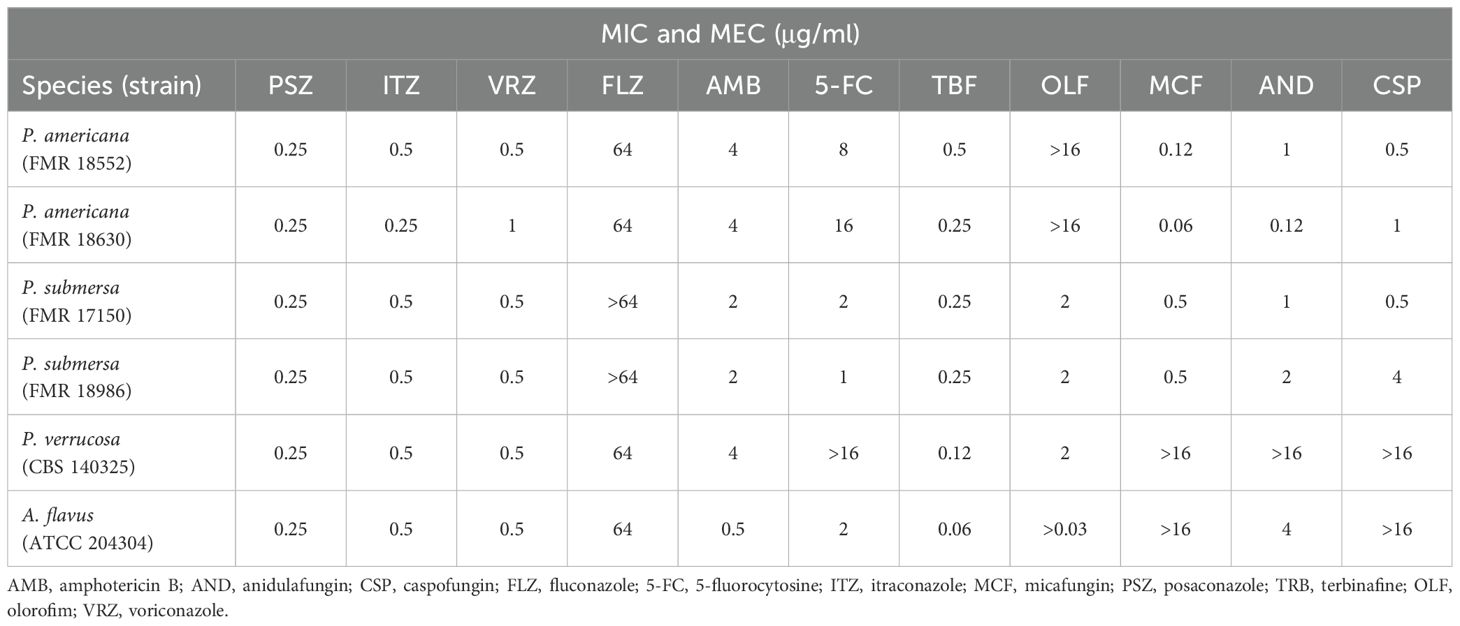
Table 3. In vitro antifungal susceptibility profiles of the Phialophora species tested.
Stress response and adaptation of P. submersa
Among the five tested strains of P. submersa, FMR 17150 exhibited the highest resistance to a range of stress conditions, except for cationic and salt stress. None of the strains showed growth on PDA supplemented with 3M sorbitol or 1M NaCl, indicating complete growth inhibition under these conditions (100% inhibition). In contrast, growth on PDA supplemented with 1M KCl was limited, with only FMR 18986 capable of growing under this condition. Under cell wall-destabilizing stress conditions (CFW and CR), FMR 17150 showed notable resistance, displaying even enhanced growth compared to control conditions. However, the other strains were sensitive to CR, showing reduced growth. All strains demonstrated resistance to membrane-destabilizing stress (SDS) and oxidative stress (H2O2) conditions, suggesting a general resilience of P. submersa to these environmental challenges (Figure 7).
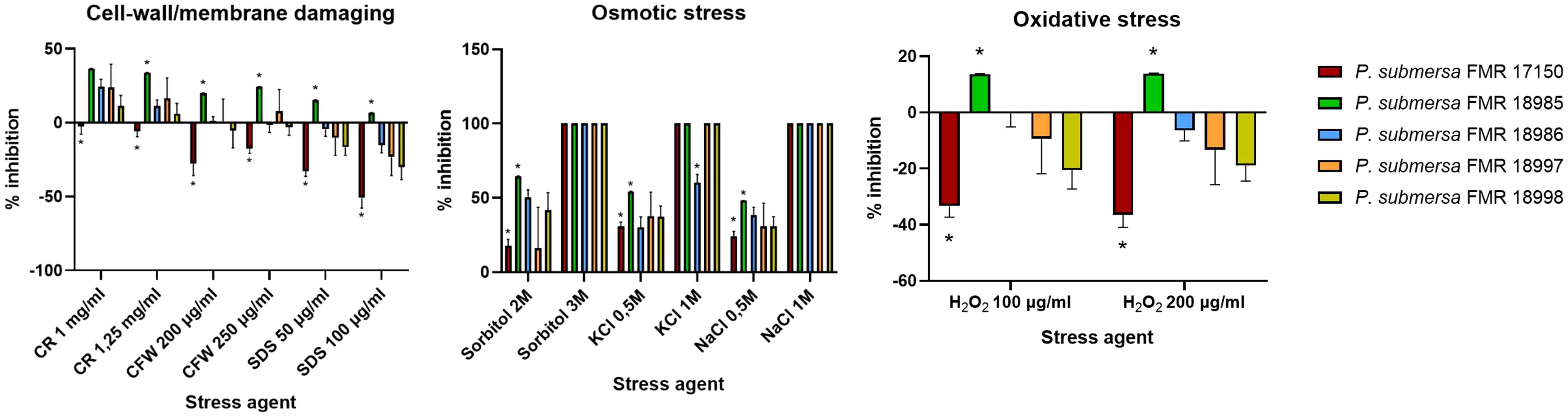
Figure 7. Rate inhibition of growth in Phialophora submersa under different stress conditions. Data were analyzed using 2-way ANOVA. Asterisks indicate a statistically significant difference P < 0.05*.
Discussion
The findings of this study provide new insights into the pathogenic potential and host-pathogen interactions of Phialophora species, with a particular focus on P. americana, P. submersa, and P. verrucosa. These species exhibited varying abilities to infect macrophages, form biofilms, and induce cellular damage, highlighting their different virulence profiles. Additionally, we observed changes in the transcription levels of key immune system genes, which may play a role in the host’s response to infection and further elucidate the pathogenesis of these species.
Phagocytosis assays revealed that macrophages internalized P. verrucosa significantly less efficiently than P. americana, with P. submersa showing intermediate uptake, irrespective of the strains tested. This supports previous findings indicating that fungal species differ in their ability to evade innate immune responses, with some fungi expressing surface proteins that reduce recognition by immune cells (Gordon and Taylor, 2005; Mira et al., 2017). The reduced phagocytosis level observed with P. verrucosa may be attributed to its ability to inhibit immune cell recognition through the modulation of host signaling pathways or the expression of surface proteins that evade immune detection (Gordon and Taylor, 2005). In contrast, the higher phagocytic uptake of P. americana implies a less effective evasion strategy, correlating with its lower intracellular survival.
In terms of intracellular survival, P. verrucosa demonstrated the highest survival at 4 h post-infection, followed by P. submersa and P. americana. This pattern aligns with the hypothesis that more virulent species can better withstand phagolysosomal conditions through mechanisms such as inhibition of phagosomal acidification or antioxidant defense systems (Borges et al., 2018).
In addition to the observed differences in phagocytosis and survival, transcriptional analysis of key immune-related genes also revealed significant variations in the macrophage response to these Phialophora species. The cytokines and chemokines TNF-α and CCL20, which play critical roles in inflammation and immune cell recruitment, were unregulated following infection with all Phialophora species, demonstrating that these pathogens elicit an inflammatory response from the host immune system. Notably, P. verrucosa induced the highest levels of TNF-α and CCL20 expression, suggesting a more robust inflammatory response. These cytokines are known to be involved in the recruitment of additional immune cells, potentially contributing to the persistence of infection (Cugini et al., 2021). On the other hand, P. americana and P. submersa induced lower levels of these cytokines, which could reflect differences in their ability to modulate host immune responses or evade immune detection. The upregulation of the apoptosis-related gene TP53 and the transcription factor gene RELA in response to all Phialophora species further emphasizes the immune response activation, as TP53 and RELA are key regulators of cell death pathways that can influence pathogen survival within the host (Maderna and Godson, 2003). RELA/p65 plays a central role in the regulation of inflammatory and immune responses. Upon activation of the NF-κB pathway, RELA translocates to the nucleus, where it drives the transcription of various pro-inflammatory genes, including TNF-α or IL-1β and CCL20, a chemokine involved in the recruitment of CCR6+ immune cells. The promoter region of CCL20 contains NF-κB binding motifs, and its expression is known to be directly regulated by RELA (Maderna and Godson, 2003; Borges et al., 2018; Cugini et al., 2021).
The pyroptosis-related genes NLRP3 and IL-1β were also significantly upregulated in response to infection, indicating the activation of inflammasomes and the subsequent release of pro-inflammatory cytokines that are crucial for the host’s defense against infection (Borges et al., 2018). The highest expression of IL-1β was observed following the infection of P. verrucosa, followed by P. submersa, which may suggest a heightened inflammatory response when these species are present in the infection. To further investigate this pathway, we performed a Western blot analysis to assess Casp 1 activation. While only minor differences were observed among most species, P. verrucose notably induced higher levels of cleaved Casp 1, aligning with its strong IL-1β expression. These results support the idea that inflammasome activation, particularly via Casp 1, may be species-dependent and more prominent during P. verrucosa infection (Borges et al., 2018).
Interestingly, recent comparative genomic analyses by Song et al (Song et al., 2021). identified the presence of multiple genes in Phialophora species potentially involved in immune evasion and host-pathogen interactions. Among these, CARD9-related pathways were highlighted as particularly relevant, given the central role of CARD9. CARD9 acts as a key adaptor protein in the signaling cascade triggered by C-type lectin receptors, leading to NF-κB activation and pro-inflammatory cytokine production. Mutations or deficiencies in CARD9 are known to predispose individuals to deep and disseminated fungal infections (Sobianski-Herman et al., 2024). The conservation of CARD9-interacting domains in virulent Phialophora strains, including P. verrucosa and P. americana, suggests that these fungi may exploit or modulate CARD9-mediated pathways to influence host immune responses. This could partially explain the differential expression of inflammatory markers observed in our macrophage infection assays and further supports the hypothesis that Phialophora species have evolved distinct mechanisms to interact with innate immunity depending on their ecological origin and pathogenic potential. Further research is necessary to confirm this issue.
Biofilm formation is a critical virulence factor correlated with the pathogenicity of many fungi, as it protects from host immune responses and antimicrobial treatments (Cugini et al., 2021). In our study, P. verrucosa produced the highest biofilm biomass, with P. americana forming the least amount of biofilm. This is consistent with the notion that biofilm formation contributes to fungal virulence by protecting cells from host defenses, including macrophages (Nobile and Johnson, 2015). Strong biofilm formation in P. verrucosa may partly explain its higher intracellular survival, as biofilm cells are known to exhibit increased resistance to both phagocytosis and antimicrobial agents (Finkel and Mitchell, 2011). The intermediate biofilm formation observed with P. submersa suggests that the ability to form biofilms may vary among Phialophora species, which could influence their virulence, survival in host tissues, and clinical outcomes in case of infection.
Cellular damage, assessed via LDH release, increased over time for all species, with P. verrucosa inducing the greatest damage, followed by P. submersa and P. americana. This test is indicative of host cell membrane disruption, often a consequence of the inflammatory response triggered by fungal infection (Maderna and Godson, 2003). This aligns with other virulence metrics, emphasizing the link between inflammation, biofilm-associated protection, and host cell disruption. The higher levels of cellular damage induced by P. verrucosa may reflect its more aggressive interaction with host cells, potentially involving the secretion of virulence factors that disrupt cell membranes or induce necrosis. In contrast, P. americana caused less cellular damage, possibly due to its less aggressive interaction with macrophages or a less pronounced inflammatory response. The intermediate cellular damage induced by P. submersa further supports the notion that this species exhibits virulence traits that are more similar to P. americana than to P. verrucosa.
Stress response and adaptation to a changing environment are critical for the survival of fungal species in dynamic natural environments and host niches (Brown et al., 2014; Jacobsen et al., 2018). Exposure of microorganisms to specific incremental stress enhances their resistance to that stress factor and other stresses (Gasch, 2003). Our results show that stress conditions involving the cell wall and membrane have a bigger impact on growth than the other challenging conditions. CR, a β-1,3-glucan-binding agent, and CFW, a chitin-binding agent, directly counteract the cell wall stress response itself. Different studies have shown that strains with lowered chitin levels in their walls seem more resistant to CR and CFW (Ram and Klis, 2006), being strain-dependent, as our results show. Interestingly, only the strain FMR17150 of P. submersa showed higher mycelial growth in the presence of CR and CFW.
Antioxidant defense is crucial in maintaining redox homeostasis and consist in several enzymatic activities, such are SODs, catalases, and thioredoxin/glutathione systems, among others, as well as non-enzymatic molecules such melanin. The antioxidant armamentarium in fungi is directly linked to fungal pathogenesis by attenuating host cellular ROS activity (Yaakoub et al., 2022). Overall, P. submersa, which is a more virulent species than P. americana, has shown, in general, a high resistance to cellular stresses. Resistance to SDS, a common detergent that permeates cell membranes and activates stress responses (Schroeder and Ikui, 2019), as seem for oxidative stress and CFW tests. However, results must be interpreted in a strain-dependent way and not as species characteristic since P. submersa FMR 17150 showed the most resistance to SDS, H2O2, CR and CFW, correlating with its strong biofilm formation. These traits likely enhance intracellular persistence (Matsumoto et al., 2023).
Finely, our antifungal susceptibility testing did not reveal clear species-specific patterns, although P. verrucosa was notably less susceptible to echinocandins. This may be linked to the clinical origin of the P. verrucosa strain used in our study, in contrast to the environmental origin of the other strains. Supporting this, Li et al. (Li et al., 2014) reported that clinical strains identified as P. verrucosa exhibited higher resistance than those environmental strains. Nonetheless, this data should be approached with caution, because clinical or environmental strains identified as P. verrucosa have later been reclassified mainly as P. americana and P. chinensis (Li et al., 2017; De Hoog et al., 2020; Ahmed et al., 2021). Recent literature suggests that P. verrucosa has yet to be reliably isolated from environmental sources, like its counterparts P. expanda and P. tarda (Li et al., 2017; De Hoog et al., 2020; Ahmed et al., 2021; Song et al., 2021).
Conclusions
This study focuses on the pathogenicity of P. submersa compared to P. verrucosa and P. americana on macrophage infections. By integrating macrophage infection assays, gene expression analyses, stress response evaluation, and antifungal susceptibility testing, we delineate distinct virulence profiles and stress tolerance mechanisms that could reflect their ecological origins and pathogenic capabilities. Phialophora verrucosa exhibited the highest virulence, with strong biofilm formation, extensive cellular damage, and superior intracellular survival. In contrast, P. americana showed the lowest virulence, with minimal biofilm production, lower cellular damage, and reduced intracellular macrophage survival. Phialophora submersa displayed an intermediate virulence profile, with moderate biofilm production and intracellular persistence. The immune response to these black yeasts also differed across species, with variations in the expression of cytokines, chemokines, apoptosis-related genes, and pyroptosis markers, further reflecting their differing virulence profiles. This suggests that the immune response is closely tied to the species-specific pathogenicity of Phialophora. All these findings suggest that, despite its environmental origin and its limitation to grow at human body temperature, P. submersa presents pathogenic potential to cause infection. These findings support the hypothesis of Song et al (Song et al., 2021), which suggests that all Phialophora strains, either clinical or environmental, possess a comparable potential to cause infection in humans. Understanding all these factors, particularly addressed to the stress resistance observed in P. submersa strains, may help inform the development of targeted therapeutic strategies and provide insights into the role of these fungi in infections.
Variations in stress response and resistance patterns observed in P. submersa may be attributed to the genetic variability detected among strains of this species, as well as its ability to adapt to the environmental conditions of its native habitats (Table 1; Torres-Garcia et al., 2023). Further genomic investigations could help to clarify and confirm this correlation.
Data availability statement
The original contributions presented in the study are included in the article, further inquiries can be directed to the corresponding author. Data regarding fungal strains presented in the study are deposited in the NCBI GenBank repository, accession numbers: ON009864; ON009944; ON009866; ON009946; ON009860; ON009940; ON009861; ON009941; KF881960; KF971761.
Ethics statement
Ethical approval was not required for the studies on humans in accordance with the local legislation and institutional requirements because only commercially available established cell lines were used.
Author contributions
AF-B: Visualization, Data curation, Validation, Conceptualization, Investigation, Methodology, Supervision, Software, Resources, Formal Analysis, Writing – original draft, Writing – review & editing. LC-P: Data curation, Formal Analysis, Writing – original draft, Methodology, Investigation. MS: Investigation, Methodology, Writing – review & editing, Resources, Data curation. YA: Formal Analysis, Methodology, Writing – review & editing, Investigation. JC: Writing – review & editing, Methodology, Investigation. JG: Visualization, Writing – original draft, Data curation, Investigation, Conceptualization, Funding acquisition, Resources, Supervision, Writing – review & editing, Project administration.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was supported by the I+D+I project PID2021-128068NB-100 funded by MCIN/AEI/10.78413039/501100011033 and by “ERDF A way of making Europe”.
Acknowledgments
LC-P and YA are grateful to University Rovira i Virgili and Diputació de Tarragona for the Martí Franquès doctoral grant.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ahmed, S. A., Bonifaz, A., González, G. M., Moreno, L. F., Menezes da Silva, N., Vicente, V. A, et al. (2021). Chromoblastomycosis caused by Phialophora-proven cases from Mexico. J Fungi 7, 95. doi: 10.3390/jof7020095
Borges, S. A., Ferreira, F. A., and Tavares, A. C. (2018). Mechanisms of fungal survival in the host: A focus on Candida albicans. Cell Microbiol. 20, e12829. doi: 10.1111/cmi.12829
Borrás, P., Messina, F., Abrantes, R., Iachini, R., Minatel, L., and Santiso, G. (2022). First report of phaeohyphomycosis caused by Phialophora americana in a domestic cat from Argentina. J Feline Med Sur Open Rep. 8, 20551169221077611. doi: 10.1177/20551169221077611
Brown, A. J., Budge, S., Kaloriti, D., Tillmann, A., Jacobsen, M. D., Yin, Z, et al. (2014) Stress adaptation in a pathogenic fungus. J Exp Biol. 217(Pt 1), 144–55. doi: 10.1242/jeb.088930
Clinical and Laboratory Standards Institute (CLSI) (2017). “Reference method for broth dilution antifungal susceptibility testing of filamentous fungi,” in CLSI standard M38, 3rd ed (CLSI, Wayne, PA).
Cugini, C., Ramasubbu, N., Tsiagbe, V. K., and Fine, D. H. (2021). Dysbiosis from a microbial and host perspective relative to oral health and disease. Front Microbiol. 12, 617485. doi: 10.3389/fmicb.2021.617485
De Hoog, G. S., Guarro, J., Gené, J., Ahmed, S.A., Al-Hatmi, A.M.S., Figueras, M.J, et al. (2020). “Atlas of clinical fungi: the ultimate bench tool for diagnostics,” in Part α: introductions, lower fungi, basidiomycetes, yeasts, filamentous ascomycetes (A-B)/part β: filamentous ascomycetes (C-Z), 4th ed (Foundation Atlas, Hilversum, The Netherlands), 1599.
Fernández-Bravo, A. and Figueras, M. J. (2022). Immune response of the monocytic cell line THP-1 against six Aeromonas spp. Front. Immunol. 13. doi: 10.3389/fimmu.2022.875689
Fernández-Bravo, A., Kilgore, P. B., Andersson, J. A., Blears, E., Figueras, M. J., Hasan, N. A., et al. (2019). T6SS and ExoA of flesh-eating Aeromonas hydrophila in peritonitis and necrotizing fasciitis during mono- and polymicrobial infections. Proc. Natl. Acad. Sci. U.S.A. 116, 24084–24092. doi: 10.1073/pnas.1914395116
Fernández-Bravo, A., Recio, G., and Figueras, M. J. (2024). Interactions between Aeromonas caviae and Yersinia enterocolitica isolated from a case of diarrhea: evaluation of antimicrobial susceptibility and immune response of infected macrophages. Front. Microbiol. 15. doi: 10.3389/fmicb.2024.1328766
Finkel, J. S. and Mitchell, A. P. (2011). “Biofilm formation in Candida albicans,” in Candida and candidiasis, eds. Calderone, R. A. and Clancy, C. J. pp 299–315. doi: 10.1128/9781555817176.ch19
Gasch, A. P. (2003). “The environmental stress response: a common yeast response to diverse environmental stresses.” in Yeast Stress Responses. Topics in Current Genetics, vol 1, eds. Hohmann, S. and Mager, W. H. (Heidelberg: Springer, Berlin). doi: 10.1007/3-540-45611-2_2
Gordon, S. and Taylor, P. R. (2005). Monocyte and macrophage heterogeneity. Nat. Rev. Immunol. 5, 953–964. doi: 10.1038/nri1733
Guarro, J., Gené, J., and Stchigel, A. M. (1999). Developments in fungal taxonomy. Clin. Microbiol. Rev. 12, 454–500. doi: 10.1128/cmr.12.3.454
Jacobsen, M. D., Beynon, R. J., Gethings, L. A., Claydon, A. J., Langridge, J. I., Vissers, J. P. C., et al. (2018). Specificity of the osmotic stress response in Candida albicans highlighted by quantitative proteomics. Sci Rep 8, 14492. doi: 10.1038/s41598-018-32792-6
Kwasna, H., Szewczyk, W., Baranowska, M., Gallas, E., Wisniewska, M., and Behnke-Borowczyk, J. (2021). Mycobiota associated with the vascular wilt of poplar. Plants, 10, 892. doi: 10.3390/plants10050892
Li, Y., Xiao, J., de Hoog, G. S., Wang, X., Wan, Z., Yu, J., et al. (2017). Biodiversity and human-pathogenicity of Phialophora verrucosa and relatives in Chaetothyriales. Persoonia. 38, 1–19. doi: 10.3767/003158517X692779
Li, Y., Wan, Z., and Li, R. (2014). In vitro activities of nine antifungal drugs and their combinations against Phialophora verrucosa. Antimicrob. Agents Chemother. 58, 5609–5612. doi: 10.1128/AAC.02875-14
Liang, P., Wang, X., Wang, R., Wan, Z., Han, W., Li, R., et al. (2015). CARD9 deficiencies linked to impaired neutrophil functions against Phialophora verrucosa. Mycopathologia 179, 347–357. doi: 10.1007/s11046-015-9877-2
Maderna, P. and Godson, C. (2003). Phagocytosis of apoptotic cells and the resolution of inflammation. Biochim Biophys Acta, 1639, 141–151. doi: 10.1016/j.bbadis.2003.09.004
Martini, F., Seehusen, F., Krudewig, C., Beckmann, K. M., Favrot, C., Fischer, N., et al. (2022). Phaeohyphomycosis caused by Phialophora americana in a dog. Vet. Dermatol. 33, 446–449. doi: 10.1111/vde.13096
Matsumoto, Y., Sugiyama, Y., Nagamachi, T., Yoshikaw, A., and Sugita, T. (2023) Hog1-mediated stress tolerance in the pathogenic fungus Trichosporon asahii. Sci Rep. 13, 13539. doi: 10.1038/s41598-023-40825-y
Mira, N., Liu, H., and Gordon, M. A. (2017). Phagocytosis and immune evasion by fungi. J. Fungal Immunol. 12, 295–310. doi: 10.1016/j.jfungim.2017.05.003
Murciano, C., Hor, L. I., and Amaro, C. (2015). Host–pathogen interactions in Vibrio vulnificus: responses of monocytes and vascular endothelial cells to live bacteria. Future Microbiol 10, 471–487. doi: 10.2217/fmb.14.136
Nobile, C. J. and Johnson, A. D. (2015). Candida biofilms and human disease. Annu. Rev. Microbiol. 69, 71–92. doi: 10.1146/annurev-micro-091014-104330
Passero, L. F. D., Cavallone, I. N., and Belda, W., Jr. (2021). Reviewing the etiologic agents, microbe-host relationship, immune response, diagnosis, and treatment in chromoblastomycosis. J. Immunol. Res. 2021 (1), 9742832. doi: 10.1155/2021/9742832
Ply, B., Cañete-Gibas, C. F., Sanders, C., Wiederhold, N. P., Dandar, R. A., Sheppard, J. D., et al. (2023). Phialophora chinensis fungal keratitis: An initial case report and species identification. Am. J. Ophthalmol. Case Rep. 29, 101800. doi: 10.1016/j.ajoc.2023.101800
Pruvot, C., Messagier, A. L., Garcia-Hermoso, D., Lebailly, F., Aglae, C., and Desbois-Nogard, N. (2022). First case of subcutaneous cystic phaeohyphomycosis due to Phialophora chinensis in a kidney transplant recipient in Martinique. Med. Mycol Case Rep. 39, 18–22. doi: 10.1016/j.mmcr.2022.12.003
Ram, A. L. and Klis, F. M. (2006). Identification of fungal cell wall mutants using susceptibility assays based on calcofluor white and Congo red. Nat. Protoc. 1, 2253–2256. doi: 10.1038/nprot.2006.397
Schroeder, L. and Ikui, A. E. (2019). Tryptophan confers resistance to SDS-associated cell membrane stress in Saccharomyces cerevisiae. PLoS One 14, e0199484. doi: 10.1371/journal.pone.0199484
Singh, N., Chang, F. Y., Gayowski, T., and Marino, I. R. (1997). Infections due to dematiaceous fungi in organ transplant recipients: case report and review. Clin Infect Dis. 24, 369–374. doi: 10.1093/clinids/24.3.369
Sobianski-Herman, T., Azevedo, C. M. P. S., Bocca, A. L., Lima, J. F. S., Beato-Souza, C. P., Razzolini, E., et al. (2024). CARD9 mutations and T cell immune response in patients with chromoblastomycosis. One Health Mycology 1, 14–22. doi: 10.63049/OHM.24.11.3
Song, Y., Menezes Da Silva, N., Vicente, V. A., Quan, Y., Teixeira, M. M., Gong, J., et al. (2021). Comparative genomics of opportunistic Phialophora species involved in divergent disease types. Mycoses 64, 555–568. doi: 10.1111/myc.13244
Soussa, I. S., Mello, T. P., Pereira, E. P., Granato, M. Q., Alviano, C. S., Santos, A. L. S., et al. (2022). Biofilm formation by chromoblastomycosis fungi Fonsecaea pedrosoi and Phialophora verrucosa: involvement with antifungal resistance. J Fungi 8, 963. doi: 10.3390/jof8090963
Tong, Z., Chen, S. C., Chen, L., Dong, B., Li, R., Hu, Z., et al. (2013). Generalized subcutaneous phaeohyphomycosis caused by Phialophora verrucosa: report of a case and review of literature. Mycopathologia 175, 301–306. doi: 10.1007/s11046-013-9626-3
Torres-Garcia, D., García, D., Réblová, M., Jurjević, Ž., Hubka, V., and Gené, J. (2023). Diversity and novel lineages of black yeasts in Chaetothyriales from freshwater sediments in Spain. Persoonia 51, 194–228. doi: 10.3767/persoonia.2023.51.05
Yaakoub, H., Mina, S., Calenda, A., Bouchara, J. P., and Papon, N. (2022). Oxidative stress response pathways in fungi. Cell Mol Life Sci 79, 333. doi: 10.1007/s00018-022-04353-8
Yasanthika, W. A. E., Chethana, K. W. T., Fatimah, A.-O., Wanasinghe, D. N., Tennakoon, D. S., Samarakoon, M. C., et al. (2025). Genera of soil Ascomycota and an account on soil-inhabiting species isolated from Thailand. Mycosphere 16, 1530–1860. doi: 10.5943/mycosphere/16/1/10
Yiallouris, A., Pana, Z. D., Marangos, G., Tzyrka, I., Karanasios, S., Georgiou, I., et al. (2024). Fungal diversity in the soil mycobiome: implications for One health. One Health 18, 100720. doi: 10.1016/j.onehlt.2024.100720
Keywords: Phialophora, environmental species, macrophages, immune response, virulence, antifungal susceptibility, stress responses
Citation: Fernández-Bravo A, Camuña-Pardo L, Sanchis M, Ahmiane Y, Capilla J and Gené J (2025) Pathogenic characterization of Phialophora submersa, a new black yeast isolated from freshwater sediments in Spain. Front. Cell. Infect. Microbiol. 15:1620047. doi: 10.3389/fcimb.2025.1620047
Received: 29 April 2025; Accepted: 23 June 2025;
Published: 14 July 2025.
Edited by:
Nicolas Papon, Université d’Angers, FranceReviewed by:
Prashant R. Desai, University of Wisconsin-Madison, United StatesBilin Dong, No.1 Hospital of Wuhan, China
Copyright © 2025 Fernández-Bravo, Camuña-Pardo, Sanchis, Ahmiane, Capilla and Gené. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Josepa Gené, am9zZXBhLmdlbmVAdXJ2LmNhdA==
†ORCID: Laura Camuña-Pardo, orcid.org/0009-0008-7952-0979
Marta Sanchis, orcid.org/0000-0001-7251-6792
Youssef Ahmiane, orcid.org/0000-0002-1060-2051