 Silvia Valeria Padilla-García1
Silvia Valeria Padilla-García1 Abraham Loera-Muro2
Abraham Loera-Muro2 Martín Humberto Muñoz-Ortega3
Martín Humberto Muñoz-Ortega3 David Alejandro Hernández-Marín4
David Alejandro Hernández-Marín4 Javier Ventura-Juárez1
Javier Ventura-Juárez1 Sandra Luz Martínez-Hernández4*
Sandra Luz Martínez-Hernández4*- 1Department of Morphology, Center for Basic Sciences, Autonomous University of Aguascalientes, Aguascalientes, Ags, Mexico
- 2Secihti-Center for Biological Research of the Northwest, SC, La Paz, Mexico
- 3Department of Chemistry, Center for Basic Sciences, Autonomous University of Aguascalientes, Aguascalientes, Ags, Mexico
- 4Department of Microbiology, Center for Basic Sciences, Autonomous University of Aguascalientes, Aguascalientes, Ags, Mexico
Introduction: The pathological progression of cirrhosis disrupts the gut-liver axis. Lactococcus lactis (L. lactis) exhibits immunomodulatory properties and an ability to enhance intestinal barrier function. It has been demonstrated that tamsulosin has antifibrotic and anti-inflammatory effects in hepatic injury models. This study evaluated the effect of a tamsulosin and L. lactis co-treatment on the recovery of microbiota and intestinal barrier integrity in a Wistar rat liver cirrhosis model.
Material and methods: Male Wistar rats were administered CCl4 intraperitoneally for 4 weeks. Subsequently, rats received tamsulosin, L. lactis, or both, orally for 2 weeks. The intestinal microbiota was assessed using 16S rRNA gene sequencing. Intestinal barrier integrity was evaluated using qPCR and Western blot for proteins ZO-1, occludin, and claudin-2. Bacterial translocation was evaluated by endotoxin concentration, bacterial DNA, and microbial culture of extraintestinal tissues. Finally, hepatic, intestinal histology, and liver function markers were analyzed.
Results: L. lactis and its combination with tamsulosin (T/L. lactis) increased microbial diversity and promoted a balanced gut microbiota characterized by a Firmicutes predominance followed by Proteobacteria and reduced Clostridia and Gammaproteobacteria levels. L. lactis group upregulated ZO-1 and occludin expression, while no significant changes were observed with tamsulosin or T/L. actis groups, nonetheless, intestinal morphology resembled that of healthy controls. Bacterial translocation analysis revealed no endotoxins, bacterial DNA, or bacteria in extraintestinal tissues. Both treatments also improved hepatic and intestinal histology, with partial liver function recovery.
Conclusión: Findings such as reduced bacterial translocation, lower systemic endotoxin levels, improved intestinal morphology, and modulation of gut microbiota composition suggest that both agents (L. lactis and tamsulosin), particularly in combination, exert positive effects on the intestinal barrier in cirrhosis.
1 Introduction
Hepatic cirrhosis, is a chronic progressive disease with significant systemic consequences, arising from various hepatic insults that trigger inflammation and fibrosis. Its complications range from mild to severe, potentially leading to multi-organ failure and death (Roehlen et al., 2020). Cirrhosis develops after prolonged injury, which leads to the replacement of healthy liver parenchyma with fibrotic tissue and regenerative nodules, ultimately causing portal hypertension. The disease progresses from an asymptomatic stage (compensated cirrhosis) to a symptomatic one (decompensated cirrhosis), whose complications frequently require hospitalization (Ginès et al., 2021). The intestine is among the hepatic cirrhosis-impacted organs, where increased intestinal permeability leads to significant modifications in gut microbiota composition (Lee and Suk, 2020), as well as changes in the quantity and quality of mucus lining the intestinal epithelium, along with dysmotility and damage to the epithelial barrier. Cirrhotic patients exhibit an overgrowth of potentially pathogenic bacteria accompanied by a reduction in autochthonous bacterial populations (Fukui, 2021). Pathological bacterial translocation plays a critical role in the onset of various infections. In cirrhotic patients, it contributes to the development of spontaneous bacterial peritonitis and is associated with multi-organ failure in critically ill individuals, as well as progressive hepatic hemodynamic deterioration (Garcı、a-Tsao, 2001; Wiest et al., 2014). Gut alterations associated with liver cirrhosis arise from the bidirectional interaction and complex crosstalk between the gut and the liver (Albillos et al., 2020). Recent studies have explored the use of various probiotic supplements as potential therapeutic interventions for cirrhosis, with some reporting beneficial effects in affected patients. L. lactis is a Gram-positive, spherical, homolactic, non-sporulant, and facultative anaerobic bacterium (Parapouli et al., 2013). It plays a relevant role in the intestine as an immunomodulator by promoting the production of anti-inflammatory cytokines, strengthening the intestinal barrier, and competing against pathogens (Laguna et al., 2024). Previous studies from our group have reported that oral administration of L. lactis may serve as a potential strategy to prevent and protect against liver injury. L. lactis attenuated hepatic cirrhosis by preventing steatosis and fibrosis, decreasing serum aspartate aminotransferase (AST) and alanine aminotransferase (ALT) levels, downregulating hepatic IL-1β, and enhancing anti-inflammatory responses through increased expression of Foxp3 and IL-10 (Delgado-Venegas et al., 2021).
On the other hand, tamsulosin is a selective α1-adrenergic antagonist with a prolonged therapeutic effect, commonly prescribed for the management of genitourinary conditions. It primarily acts on the smooth muscle of the urethra, bladder neck, and prostate, inducing muscle relaxation and thereby improving urinary flow and reducing symptoms associated with lower urinary tract obstruction (Dunn et al., 2002; Song et al., 2020). Pharmacological inhibition using nonselective α- and β-adrenergic receptor antagonists has been shown to suppress hepatic stellate cell (HSC) proliferation and attenuate fibrogenesis (Liu et al., 2009). Studies have shown that tamsulosin facilitates improved recovery of hepatic architecture, accompanied by a reduction in fibrosis and NF-κB activation (Soleil et al., 2022) in cirrhotic rats. Primary HSCs were found to express adrenoreceptors α-1B, α-1D, β1, and β2 by RT-PCR analysis. Neurotransmitters (noradrenaline and adrenaline) stimulate and activate hepatic stellate cells by binding to cells through α1-adrenergic receptors, triggering a cellular response that leads to the release of inflammatory cytokines (Oben and Diehl, 2004; Sigala et al., 2013). Previous studies have shown that the α1-adrenoceptor blocker doxazosin decreases the fibrogenic activity of activated HSCs, which is associated with the induction of cellular senescence via α1-AR antagonism, suggesting that α1-AR is a potential treatment for liver fibrosis. The adrenoblocker downregulated collagen I and ACTA2 and increased PPAR-γ expression both in the presence and absence of TGF-β, confirming that doxazosin delays stellate cell activation (Serna-Salas et al., 2022). Tamsulosin also directly modulates the HSC, reducing quiescent and activated forms (Buendía-Delgado et al., 2022) and promoting anti-inflammatory processes (Alabdali and Ibrahim, 2023; Dunn et al., 2002; Song et al., 2020).
To date, no studies have investigated the effects of tamsulosin on intestinal barrier permeability or gut microbiota composition in experimental cirrhosis models. Furthermore, it remains unknown whether co-administration of this drug with L. lactis may enhance intestinal integrity, influence bacterial translocation, and thereby mitigate the effects of cirrhosis.
2 Materials and methods
2.1 Animal conditions
Wistar rats were obtained from the Central Bioterium of the Autonomous University of Aguascalientes and kept in the local Animal Core Facility at 25°C with 12-h light/dark cycles. The rats were fed Laboratory Chow (Purina, Mexico) and provided tap water ad libitum. All animals were treated with fenbendazole (55 mg), toltrazuril (20 mg), and praziquantel (10 mg) (1 ml/kg) for 3 days. Finally, animals were kept under standard and hygienic conditions to acclimate for 7 days before starting the cirrhosis induction.
2.2 Lactococcus lactis strain and culture conditions
L. lactis strain NZ9000 was grown in 10% glucose-M17 broth (BD Difco, Sparks, MD, USA) at 30°C. Cells were harvested at 8,000 xg for 5 min. Finally, cells suspensions were adjusted to 1×109 CFU/ml in sterile water based on an optical density (OD) at 600 nm of 1.0. Doses of 1×109 CFU in 250 µl per animal were prepared from fresh cultures.
2.3 Experimental design
A total of 30 male Wistar rats (150–200 g) were randomly divided into 6 groups (n=5): (i) control (healthy), (ii) cirrhotic (diseased), (iii) placebo (endogenous recovery), (iv) L. lactis (treated with L. lactis), (v) tamsulosin (treated with tamsulosin), (vi) T/L. lactis (treated with tamsulosin and L. lactis). To induce liver cirrhosis, carbon tetrachloride (CCl4, Sigma-Aldrich, Darmstadt, Germany) was administered intraperitoneally at a 0.4 g/kg dose, three times per week for four weeks (Macías-Pérez et al., 2019). Once the induction was completed, treatments were started. Orally, for 2 weeks, 0.8 mg/kg/day of tamsulosin hydrochloride (Pharmalife LTC) and L. lactis (1×109 CFU in 250 µl) were suspended in sterile purified water and administered separately once daily, using a curved stainless steel esophageal cannula (18x3 mm, Cadence Science). At the end of the treatments, all rats were euthanized with sodium pentobarbital overdose (≥100 mg/kg) applied intraperitoneally, until rapid loss of consciousness, thus minimizing the stress and anxiety experienced by the animal, while monitoring respiratory and cardiac signs until their absence. All animal experiments were approved by the Ethics Committee for the use of animals in teaching and research at UAA (CEADI−UAA, UAA: Autonomous University of Aguascalientes, AUT−B−C−1121−077), following the Mexican Official Standard NOM−062−ZOO−1999 (Muñoz-LIO, 2001) and the guidelines of the National Institutes of Health for the care and use of Laboratory animals (National Research Council, 2011) samples were taken from the portal vein and by cardiac puncture, and under sterile conditions, samples of intestinal tissue, liver, mesenteric lymph node (MLN), and spleen were collected for further analysis. Additionally, fecal samples were directly collected from the intestine and stored at -80°C until processing (Figure 1).
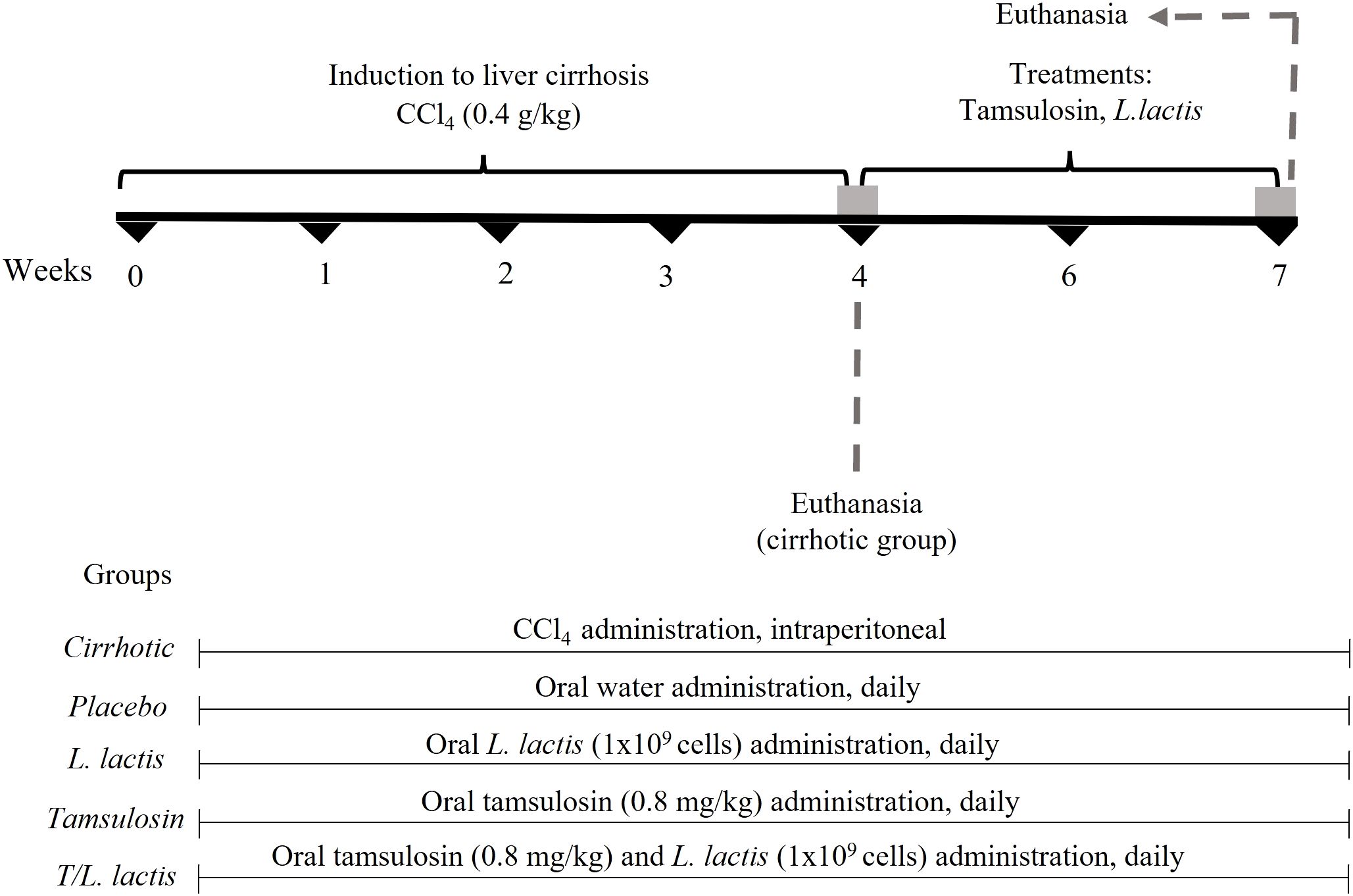
Figure 1. Treatment protocol with tamsulosin and L. lactis. CCl4 (carbon tetrachloride) was administered intraperitoneally at a dose of 0.4 g/kg, three times per week for 4 weeks. Tamsulosin hydrochloride (tamsulosin) (0.8 mg/kg/day) and L. lactis (NZ9000) (1×109 cells/ml/day) were administered orally for 2 weeks. At the end of the treatment period, all animals were euthanized, and tissue samples were collected.
2.4 Analysis of intestinal bacterial community
Fecal samples were collected at the end of the study and placed in sterile containers. Bacterial DNA was isolated using the Quick-DNA™ Fecal/Soil Microbe Miniprep Kit (Zymo Research, OC, California) according to the manufacturer’s specifications. The DNA pellet was resuspended in 50 µl of ultra-pure distilled water. The integrity of total DNA (100–200 ng) for each sample was verified by 1.0% agarose gel electrophoresis, and its quality was assessed using a Bio-Drop-LITE spectrophotometer (Isogen Lifescience, UT, Netherlands). The extracted DNA was stored at -80°C until sequencing. From the total extracted DNA, the V3 region of the 16S rDNA gene in prokaryotes (bacteria and archaea) was amplified using the V3-338F (5′-ACTCCTACGGGAGGCAGC-3′) and V4-806R (5′ GGACTACHVHHHTWTCTAAT-3′) primers. The amplified regions were sequenced on the Illumina MiniSeq platform 2×150 (300 cycles) (Illumina) at the Centro de Investigación en Alimentos y Desarrollo (CIAD, Mazatlan, Sinaloa, Mexico). Raw reads were filtered, trimmed, and dereplicated; paired-end reads were merged, denoised, and chimeras were removed using the DADA2 (version 1.8) pipeline within QIIME2 (version 2020.2). Taxonomic assignment of amplicon sequence variants (ASVs) was performed using the SILVA-132 database. Sequences assigned to eukaryotes and archaea were discarded. Alpha diversity metrics were calculated, including relative taxon abundance, Shannon and Simpson diversity indices, and Pielou’s evenness index. Additionally, the UniFrac distance matrix was computed (QIIME2, version 2020.2).
2.5 Intestinal permeability: ZO-1, occludin and claudin-2 determination
The effects of treatments with tamsulosin and L. lactis on the mRNA and protein levels of ZO-1, occludin, and claudin-2 were evaluated using quantitative PCR (qPCR) and Western Blot analysis, respectively. For qPCR analysis, total RNA was extracted using the Direct-zol™ RNA MiniPrep Kit (Zymo Research), following the manufacturer’s instructions. The RNA was quantified using the BioDrop (Isogen Life Science B.V.). For cDNA synthesis, reverse transcription was performed with 1 µg of RNA using the GoScript™ Reverse Transcription System (Promega). Subsequently, qPCR was performed using the Maxima SYBR Green/ROX qPCR Master Mix (2X) (Thermo Fisher Scientific) with the StepOne™ Real-Time PCR System (Applied Biosystems). The primers for the ZO-1, occludin, claudin-2, and β-actin genes are shown in Table 1. Thermocycling conditions were as follows: 50°C for 2 min, 95°C for 3 min, followed by 40 cycles of 95°C for 40 s and 60°C for 45 s. The relative expression levels were normalized to β-actin, and differences were determined using the 2−ΔΔCq method (Livak and Schmittgen, 2001). The specificity of the PCR product was confirmed by gel electrophoresis (1.0%). For Western blot analysis, intestinal samples were lysed on ice in RIPA buffer (Pierce RIPA buffer, Thermo Scientific). After homogenization and centrifugation at 14,000 rpm for 15 min at 4°C, protein concentration was measured using a standard Bradford assay (Bio-Rad). 50 μg of total protein was separated on polyacrylamide gels (30% acrylamide/bis-acrylamide) at 4% and 12% (Bio-Rad) and transferred to a PVDF membrane (Bio-Rad). The membranes were incubated with anti-rat ZO-1 (1:1000, Santa Cruz Biotechnology), anti-mouse occludin (1:300, Santa Cruz Biotechnology), anti-mouse claudin-2 (1:400, Santa Cruz Biotechnology), and anti-rabbit β-actin (1:10 000, Abcam) antibodies overnight at 4°C. Afterwards, the primary antibody was removed, and the membranes were re-incubated with secondary antibodies: goat anti-mouse IgG (HRP, 1:5000, Thermo Fisher Scientific) and anti-rat IgG (HRP, 1:10,000, Abcam). The blots were developed with the Clarity Western ECL substrate (Bio-Rad) to obtain chemiluminescence images.
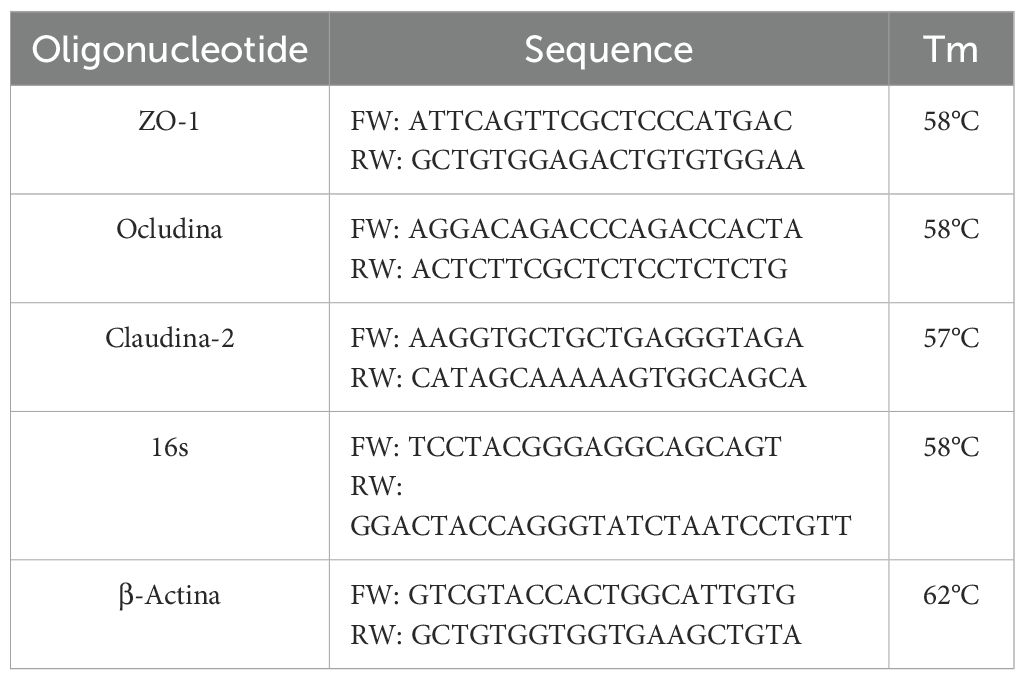
Table 1. Oligonucleotides used in this study.
2.6 Bacterial translocation assay
Tissues from MLN, spleen, and liver, previously obtained, were homogenized under sterile conditions. 100 µl of the homogenate from each tissue was collected and inoculated into different tubes containing M17 broth (BD Difco, Sparks, MD, USA), brain-heart broth (BD Bioxon, Mexico), thioglycolate (BD Bioxon, Mexico), and standard count agar (BD Bioxon, Mexico). Subsequently, they were plated onto selective media such as MacConkey, mannitol salt, and blood agar. After 48 hours of incubation at 37°C, the number of viable colonies on each plate was counted, and specific microorganisms were identified through biochemical tests. The positivity observed in these organs was considered indicative of the passage of bacteria to the portal or systemic circulation. On the other hand, bacterial DNA was determined using 1 ml of blood extracted from the portal vein. Blood was processed using the Quick-DNA™ Miniprep Plus kit (Zymo Research, OC, California), following the manufacturer’s instructions. DNA concentration was determined using the Bio-Drop-LITE spectrophotometer (Isogen Lifescience, UT, Netherlands). The 16S rDNA region was amplified by PCR in a thermal cycler (Swift MiniPro, Singapore). The PCR thermal cycles were as follows: 95°C for 3 min, 95°C for 45 s, 58°C for 35 s, 72°C for 12 s, and 72°C for 3 min. The primers set are shown in Table 1. Furthermore, endotoxin levels were measured through the LAL assay (Pierce Chromogenic Endotoxin Quantification Kit, Thermo Scientific, Waltham, MA, USA) as per the manufacturer’s instructions. The concentration of endotoxin (EU/ml) was calculated using a standard curve, and the absorbance of each well was measured at 450 nm using a microplate reader.
2.7 Histopathological analysis
Intestinal and liver samples were evaluated using hematoxylin-eosin (H&E) staining to assess architectural and morphological tissue changes. Sirius Red staining, observed under polarized light microscopy, was used to identify collagen fiber deposits (type I = red, type III = green). Tissue samples were fixed in 4% paraformaldehyde (PFA) for 24–48 hours before paraffin embedding. Serial sections (5 µm thick) of paraffin-embedded tissue were cut using a microtome (Leica RM 2125RT). Paraffin sections were then deparaffinized by heating at 60°C, washed with xylene as the deparaffinizing agent, and rehydrated through a graded ethanol series before staining. Histological preparations were visualized using a Zeiss Axioscope 40/40 FL microscope and analyzed with Image Pro Plus Software 4.5.1 (Media Cybernetics, Bethesda, MD, USA). For intestinal tissue, histomorphometric parameters were assessed in H&E-stained sections by measuring villus height (5 villi per slice and 3 slices per sample) and crypt depth (5 crypts per slice and 3 slices per sample).
2.8 Markers of liver damage
Serum samples were analyzed to determine liver damage, with quantification of levels of ALT, AST, and alkaline phosphatase (AP) using a spectrophotometric semiautomatic BTS-350 analyzer (Biosystems, Quezon City, Philippines).
2.9 Statistical analysis
Statistical analysis was performed using GraphPad Prism 8.0.2 software (GraphPad Software, San Diego, CA, USA). Data are presented as mean ± standard error of the mean (SEM). Differences between groups were assessed using two-way analysis of variance (ANOVA), followed by Tukey’s post hoc test. For non-parametric data, a Kruskal–Wallis test was used for multiple group comparisons, followed by Dunn’s post hoc test, or a Mann–Whitney U test for comparisons between two groups. A p-value < 0.05 was considered statistically significant. PERMANOVA analysis was performed using the ‘adonis’ function from the ‘vegan’ package.
3 Results
3.1 Effect of tamsulosin and L. lactis on intestinal barrier integrity
3.1.1 Intestine histological changes
In the histopathological analysis performed on intestinal samples, the intact group displayed a mucosa with a surface composed of normal goblet cells, abundant crypts, mucous cells, and microvilli. Additionally, a characteristic lamina propria was observed, rich in lymphoid cells, bordered by the muscularis mucosae. In the cirrhotic group, a thinning of both the epithelium and lamina propria was noted, along with a loss of the brush border structure and a reduction in goblet cells (Figure 2A). Villus height and crypt depth were reduced (147.95 µm and 106.50 µm, respectively) compared to the intact group (246.58 µm, p < 0.001 and 197.6 µm, respectively). The placebo group exhibited a more uniform structure; however, villus height (164.2 µm) and crypt depth (143.8 µm) were also reduced. Intestinal morphology of the L. lactis, tamsulosin, and T/L. lactis groups closely resembled that of the intact group, with villus height (x̅, 221.49 µm) and crypt depth (x̅, 210.88 µm) remaining unchanged across all three groups (Figure 2B).
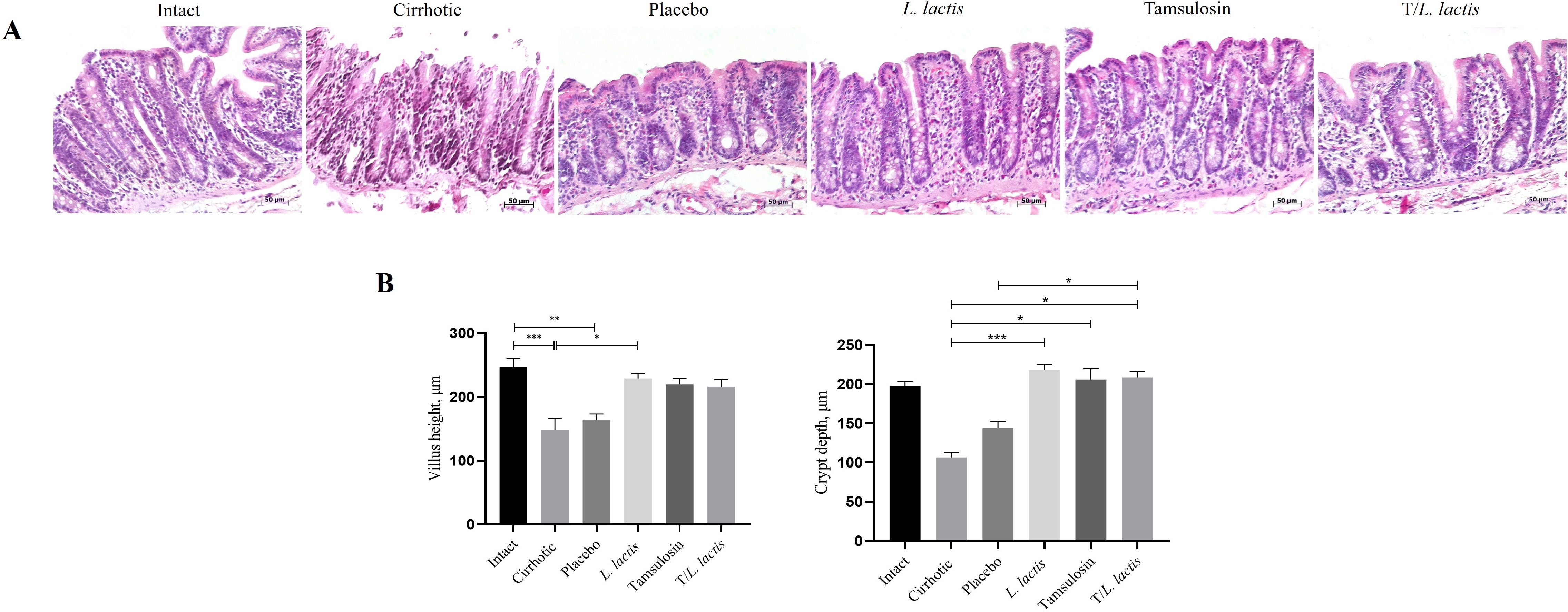
Figure 2. Analysis of intestinal morphology. (A) Representative histological images were obtained at a magnification of 20x. (B) Analysis of intestinal villus height and crypt depth. The results are presented as mean ± standard error of the mean. Samples from each rat were analyzed in triplicate. The statistical analysis was performed with the Kruskal-Wallis and Tukey post-test methods, where the values of *p < 0.05, **p < 0.01, and ***p < 0.001 were considered significant.
3.1.2 mRNA and protein expression of ZO-1, occludin and claudin-2 in intestinal tissue
RT-qPCR and Western blot analyses were performed to evaluate the effect of tamsulosin and L. lactis on the ZO-1, occludin, and claudin-2 expression. The mRNA levels of ZO-1, occluding, and claudin-2 were reduced in the cirrhotic (1.02-fold, 1.62-fold, 0.81) and placebo (1.49-fold, 1.92-fold, 1.43-fold) groups, and their expression was lower than in control animals (2.40-fold, 2.50-fold, 2.17-fold). Regarding the groups that received the different treatments, we observed that L. lactis enhanced the mRNA expression of ZO-1 (3.94-fold; p < 0.05: L. lactis vs cirrhotic) and claudin-2 (2.97-fold; p < 0.05: L. lactis vs cirrhotic). T/L. lactis showed significant differences in occludin expression (4.63-fold; p < 0.05: T/L. lactis vs cirrhotic). The tamsulosin group did not show changes in the expression of ZO-1, occludin, and claudin-2 (Figure 3A). Finally, densitometric analysis showed that ZO-1 protein expression exhibited significant differences only in the L. lactis group (0.9-fold; p < 0.05: L. lactis vs cirrhotic, p < 0.001: L. lactis vs placebo, p < 0.001: L. lactis vs T/L. lactis), although not reaching the levels seen in the intact (1.0-fold) group. Similarly, a significant increase in occludin expression was observed in the L. lactis group (1.22-fold; p < 0.05, p < 0.01) compared to the cirrhotic, placebo, tamsulosin, and T/L. lactis groups. As for claudin-2, no significant differences were observed among the treatment groups (Figure 3B).
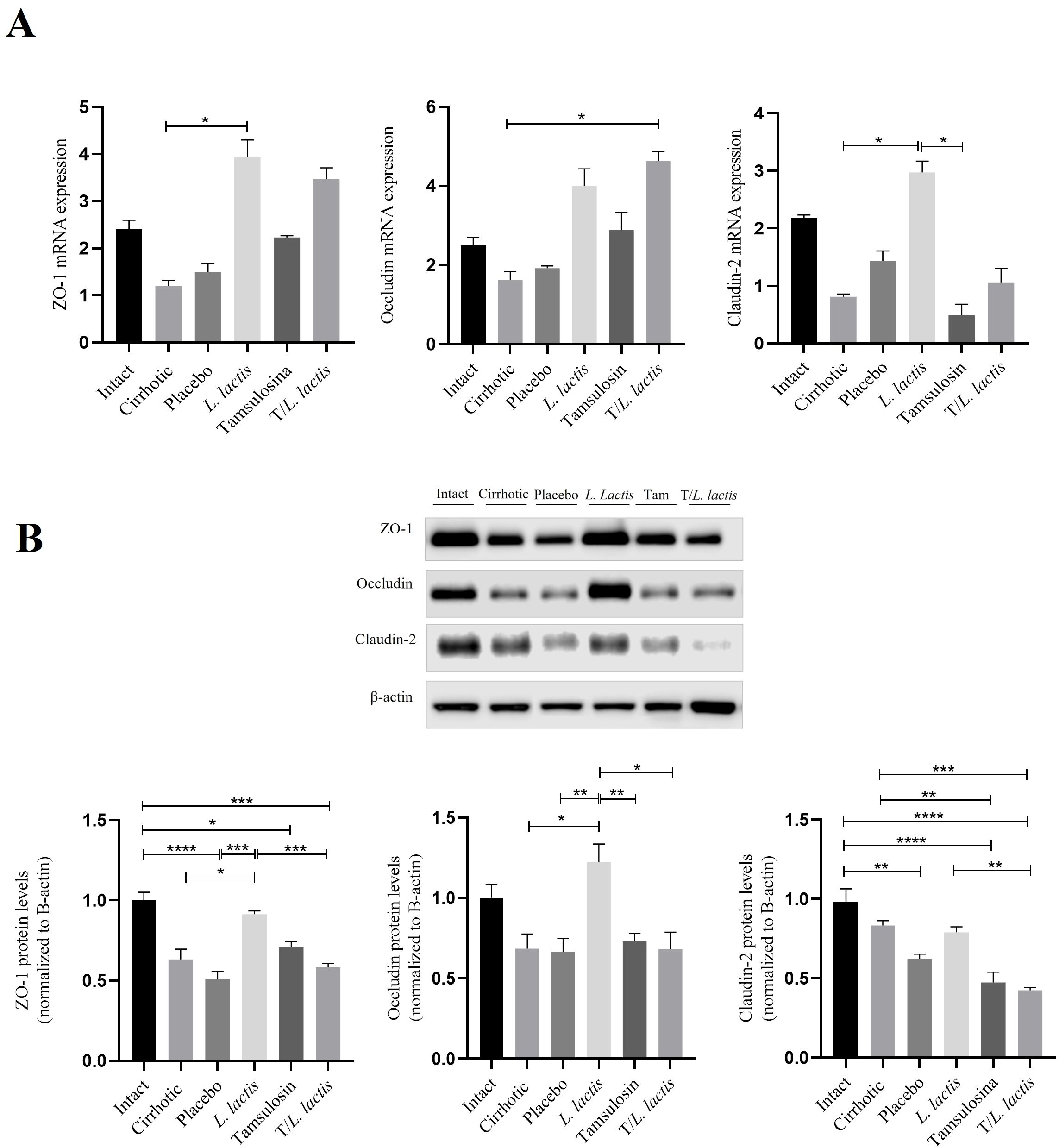
Figure 3. Intestinal Barrier: mRNA and Protein Expression Analysis. (A) mRNA of ZO-1, occludin, and claudin-2. (B) Densitometric analysis of ZO-1, occludin, and claudin-2 in intestinal tissue. Data corresponds to the mean ± SEM of three independent experiments. The statistical analysis was performed with the Kruskal-Wallis and Tukey post-test methods, where the values of *p < 0.05, **p < 0.01, ***p < 0.001, and ****p < 0.0001 were considered significant.
3.1.3 Bacterial translocation
The evaluation of bacterial translocation demonstrated the absence of bacterial growth in the intact group. In the cirrhotic and placebo groups, microorganisms were found in MLN, spleen, and liver. The isolated microorganisms were of enteric origin, with Escherichia coli being the most common (Table 2). In the L. lactis, tamsulosin, and T/L. lactis groups, no bacteria were detected in the MLN, spleen, or liver. On the other hand, blood samples processed for bacterial DNA identification showed the presence of the 16S gene in the cirrhotic and placebo groups. In the treatment groups, the 16S gene was not identified (Figure 4A). Finally, the results of plasma endotoxin measurement showed only concentrations of 0.65 EU/ml and 0.32 EU/ml in the cirrhotic and placebo groups, respectively. Animals treated with L. lactis, tamsulosin, and T/L. lactis did not show endotoxin concentrations (Figure 4B).
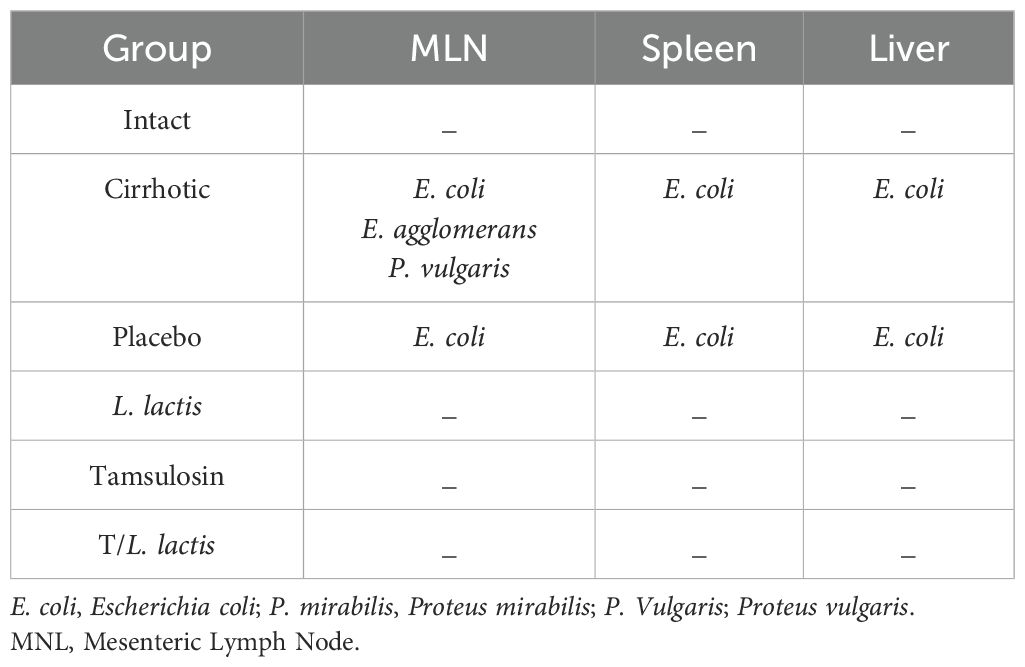
Table 2. Bacteria species isolated from mesenteric lymph nodes, spleen, and liver.
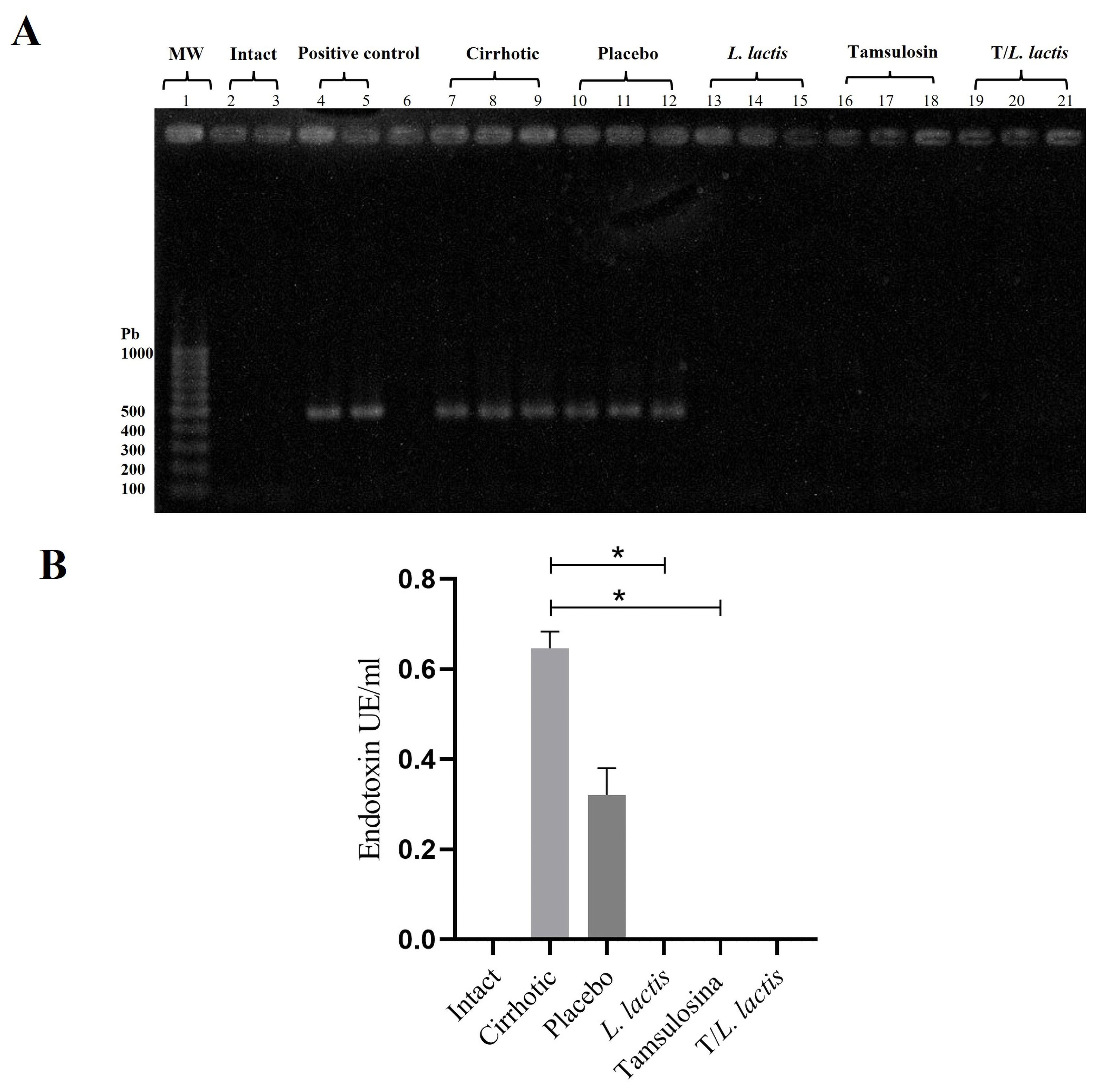
Figure 4. Effects on bacterial translocation. (A) PCR amplification of the 16S rRNA gene from bacterial DNA. (B) Serum endotoxin levels measured using a Kinetic Turbidimetric LAL assay. No amplification of the 16S rRNA gene and no detectable endotoxin levels were observed in the groups treated with tamsulosin, L. lactis, or their combination. The statistical analysis was performed using the Kruskal–Wallis and Tukey post-test methods, where the value of *p < 0.05 were considered significant.
3.2 Effect of tamsulosin and L. lactis on gut microbiota composition
3.2.1 Changes in gut bacterial diversity
Pielou’s evenness index revealed distinct patterns in microbial community structure among the analyzed groups. The L. lactis group exhibited higher microbial community evenness. T/L. lactis group showed slightly lower evenness, compared to the L. lactis group. The tamsulosin group presented a reduction in microbial community evenness compared with untreated groups (cirrhotic and placebo). According to the Shannon and Simpson indices, the L. lactis and T/L. lactis groups exhibited higher diversity levels. The tamsulosin group showed reduced microbial diversity compared with the cirrhotic and placebo groups (Figure 5A). Beta diversity was assessed using a UniFrac distance matrix and visualized via principal coordinate analysis (PCoA) to explore differences in microbial community composition. The PCoA revealed two distinct clusters: one cluster (blue ellipse) included the cirrhotic, placebo, and tamsulosin groups, indicating similar microbial community structures among them; the second cluster (green ellipse) comprised samples from the intact, L. lactis, and T/L. lactis groups, which exhibited greater similarity in microbial composition among themselves and clear differentiation from the cirrhotic group (Figure 5B).
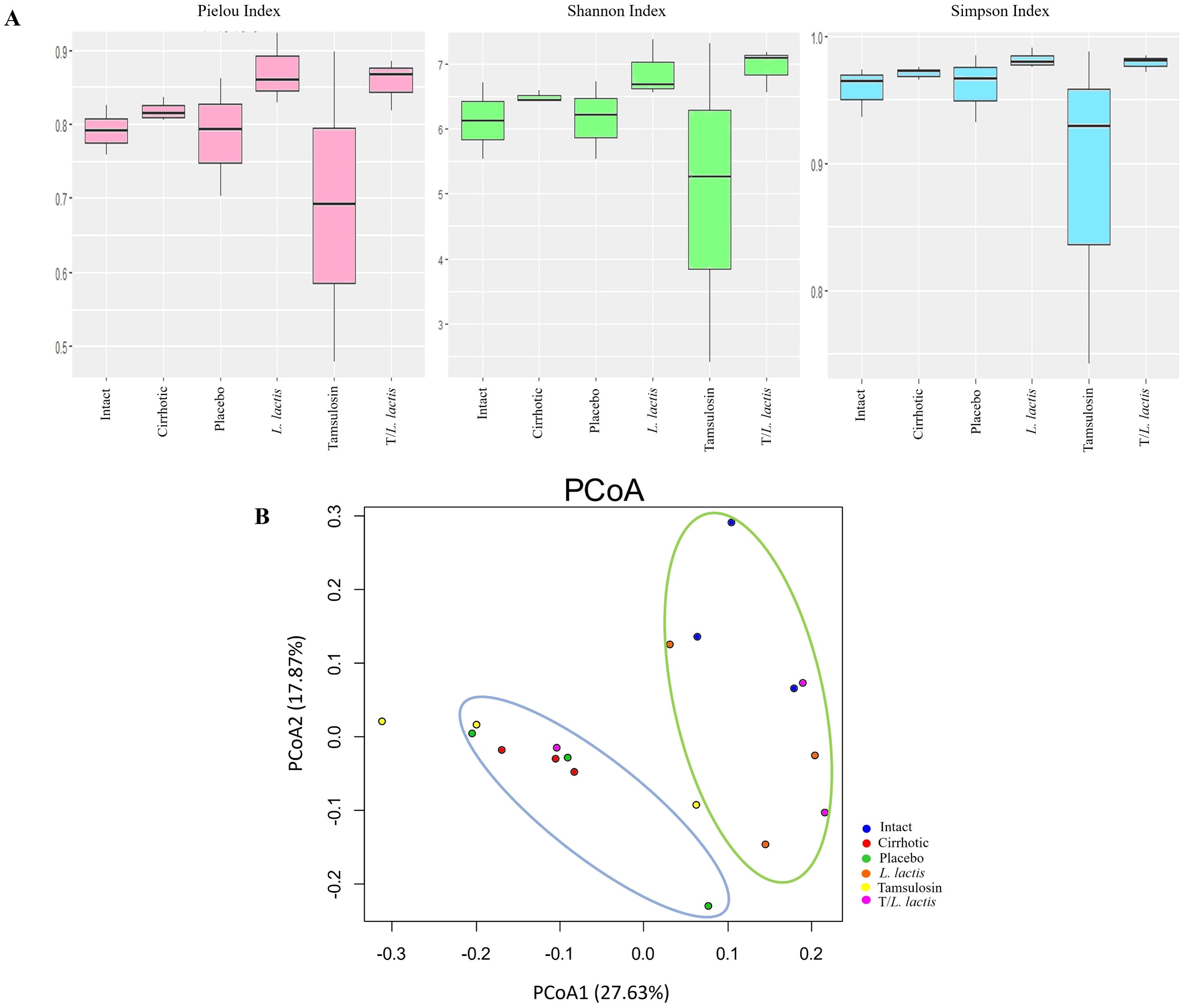
Figure 5. α and β diversity of gut microbiota between intact, cirrhotic, placebo, L. lactis, tamsulosin, and T/L. lactis groups presented by box plot. (A) Alpha diversity analysis showed that the L. lactis and T/L. lactis groups exhibited higher microbial evenness and diversity (Pielou, Shannon, and Simpson indices), while the tamsulosin group showed reduced values compared to the cirrhotic and placebo groups. (B) Beta diversity analysis, based on UniFrac distances and visualized by PCoA, revealed two distinct clusters: one (blue ellipse) comprising the cirrhotic, placebo, and tamsulosin groups, and a second cluster (green ellipse) grouping the intact, L. lactis, and T/L. lactis subjects. Similarities in microbial community composition were observed within each cluster, with clear differentiation between them.
3.2.2 Changes in gut microbial community composition
The analysis of the relative abundance of bacterial populations in the different rat groups revealed phylum-level alterations in the cirrhotic group compared to the control group: Firmicutes (52.96% vs. 90.76%), Proteobacteria (24.47% vs. 2.12%), and Bacteroidetes (21.72% vs. 6.23%). The placebo group exhibited results similar to those of the cirrhotic group: Firmicutes (68.96%), Proteobacteria (21.47%), and Bacteroidetes (8.72%). Regarding the L. lactis group vs. the cirrhotic group: Firmicutes (84.32% vs. 52.96%), Proteobacteria (0.6% vs. 24.47%), and Bacteroidetes (14.28% vs. 21.72%). The tamsulosin group vs. the cirrhotic group showed the following relative abundances: Firmicutes (71.32% vs. 52.96%), Proteobacteria (4.65% vs. 24.47%), and Bacteroidetes (23.41% vs. 21.72%). Finally, for the T/L. lactis group vs. the cirrhotic group, the proportions were as follows: Firmicutes (87.68% vs. 52.96%), Proteobacteria (1.73% vs. 24.47%), and Bacteroidetes (10.12% vs. 21.72%). The proportion of Firmicutes in the cirrhotic group was lower compared to all other groups, while Proteobacteria and Bacteroidetes were found to be increased (Figure 6A). At the class level, Clostridia were predominant in all groups. In the control group, Clostridia showed a proportion of 43.12%, followed by Gammaproteobacteria (47.09%), Bacteroidia (8.31%), and other classes (1.48%). In the cirrhotic and placebo groups, there was a predominant increase in Clostridia (77.13% and 87.94%, respectively) and Bacteroidia (14.64% and 4.87%, respectively). In the group treated with L. lactis, Clostridia were reduced (56.21%) compared to the other groups, except for the T/L. lactis group (48.76%). Gammaproteobacteria were found at 0.3%, Bacteroidia at 32.60%, and Campylobacteria at 8.15%. In the tamsulosin and the T/L. lactis-treated groups, the proportion of Clostridia were 74.45% and 48.76%, respectively; Gammaproteobacteria were 2.83% and 14.23%, and Bacteroidia were 16.87% and 28.98%, respectively (Figure 6B). Finally, family-level analysis revealed the absence of Clostridiaceae but the presence of Peptostreptococcaceae (21.12%) Prevotellaceae (22.09%), Lachnospiraceae (9.03%) Oscillospiraceae (10.14%), and Ruminococcaceae (3.33%) in the control group. In the cirrhotic and placebo groups, an increase in Clostridiaceae (27.45% and 25.61%, respectively) was observed, along with a decrease in Peptostreptococcaceae (9.02%,19.56%) and Lachnospiraceae (6.31% and 10.04%). In the L. lactis treated group, Clostridiaceae (3.17%) and Peptostreptococcaceae (7.45%) decreased, while Prevotellaceae (28,32%), Lachnospiraceae (22.78%) and Oscillospiraceae (20.01%) increased. The tamsulosin group showed an increase in Clostridiaceae (27.27%), and Peptostreptococcaceae (37.11%) was found in higher proportions compared to the other groups, whereas Lachnospiraceae (2.50%) and Oscillospiraceae (9.98%) exhibited reduced relative abundances. The T/L. lactis group exhibited a decrease in Clostridiaceae (0.23%) and Peptostreptococcaceae (3.42%). Prevotellaceae (17.11%), Lachnospiraceae (10.15%), and Oscillospiraceae (13.61%) exhibited higher abundance (Figures 6C, Table 3).
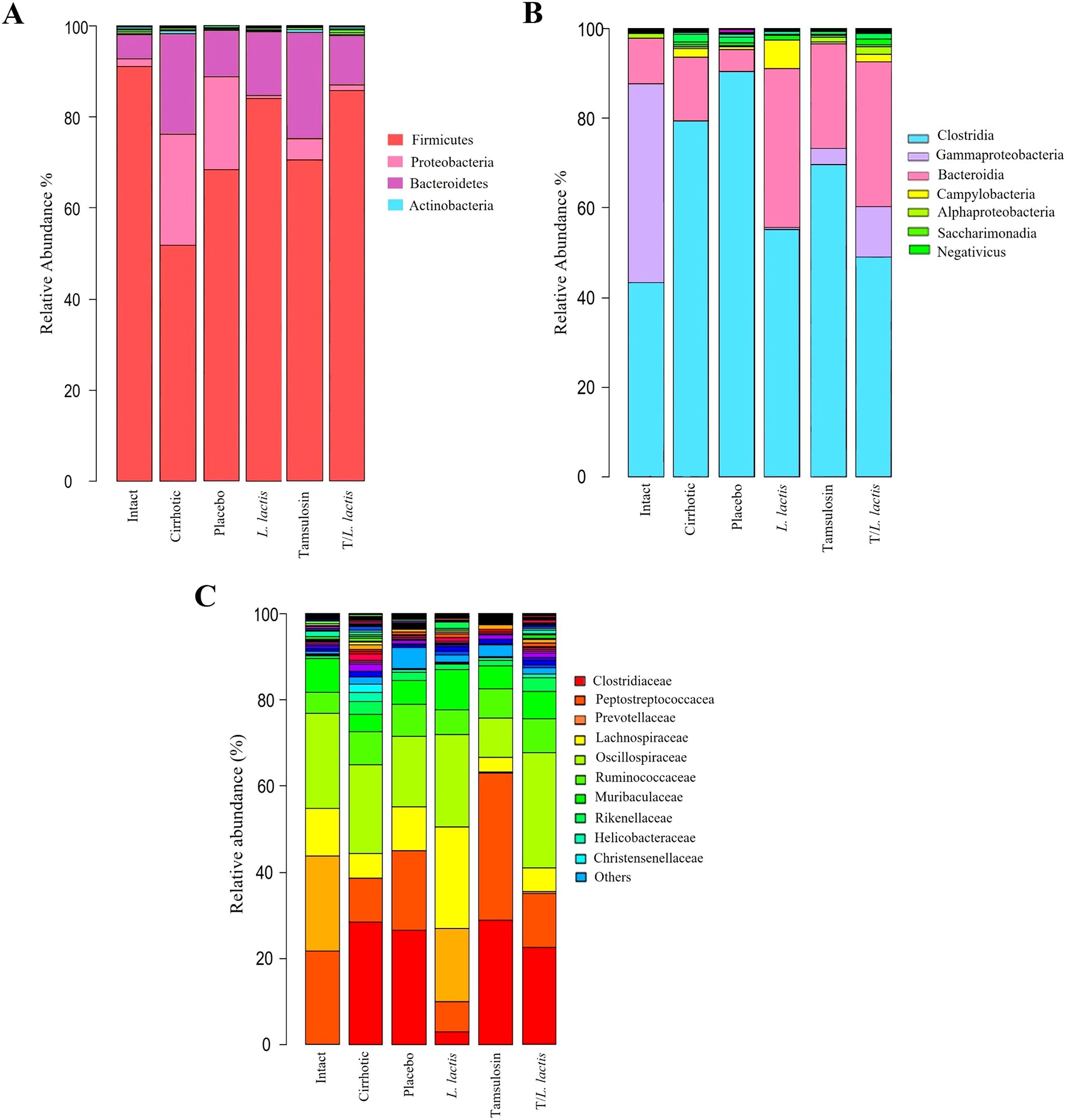
Figure 6. Change in intestinal microbial composition. Relative abundance bar plots of intestinal microbiota at the phylum, class, and family levels (A) L. lactis and T/L. lactis showed a dominance of Firmicutes, followed by Proteobacteria, and (B) reduced levels of Clostridia and Gammaproteobacteria were observed. (C) At the family level, L. lactis and T/L. lactis showed reduced abundances of Clostridiaceae and Peptostreptococcaceae, an increase of Prevotellaceae and Lachnospiraceae. Tamsulosin reduced Firmicutes and increased Clostridia and Clostridiaceae abundances.
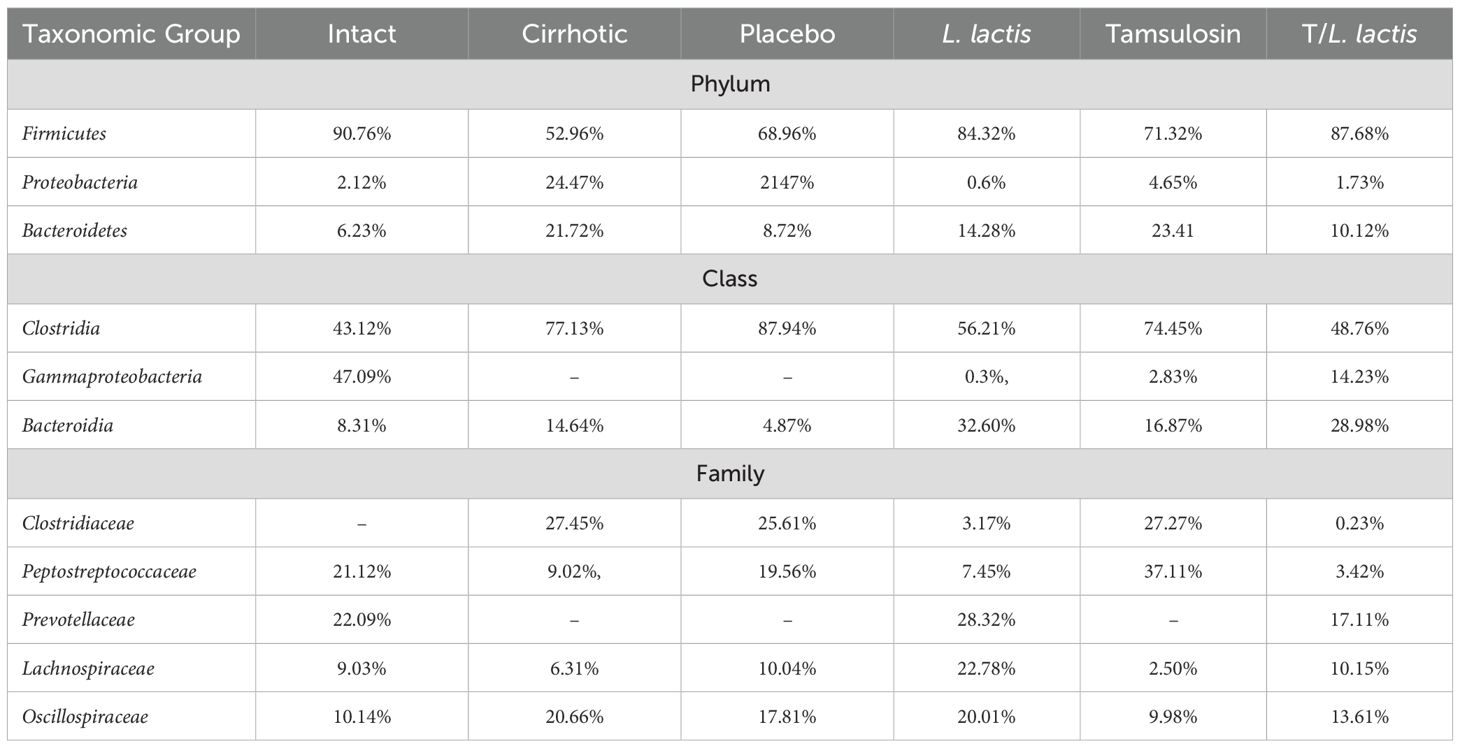
Table 3. Relative abundance (%) of major microbial taxa identified by 16S rRNA sequencing.
At the genus level, a higher abundance of Prevotella spp., was observed in the L. lactis and T/L. lactis groups compared to the cirrhotic, placebo, and tamsulosin groups. In contrast, the genera Blautia spp, Clostridioides spp., Helicobacter spp., Bacteroides spp., and Ruminococcus spp., were predominantly detected in the placebo group. Regarding the genera Colidextrinbacter spp., Alloprevotella spp., Phascolarctobacterium spp., Parabacteroides spp., Solobacterium spp., Clostridium sensu stricto, Clostridia_UCG-014, Butyricicoccus spp., and Parasutterella spp., a very low or undetectable abundance was observed in all groups (Figure 7).
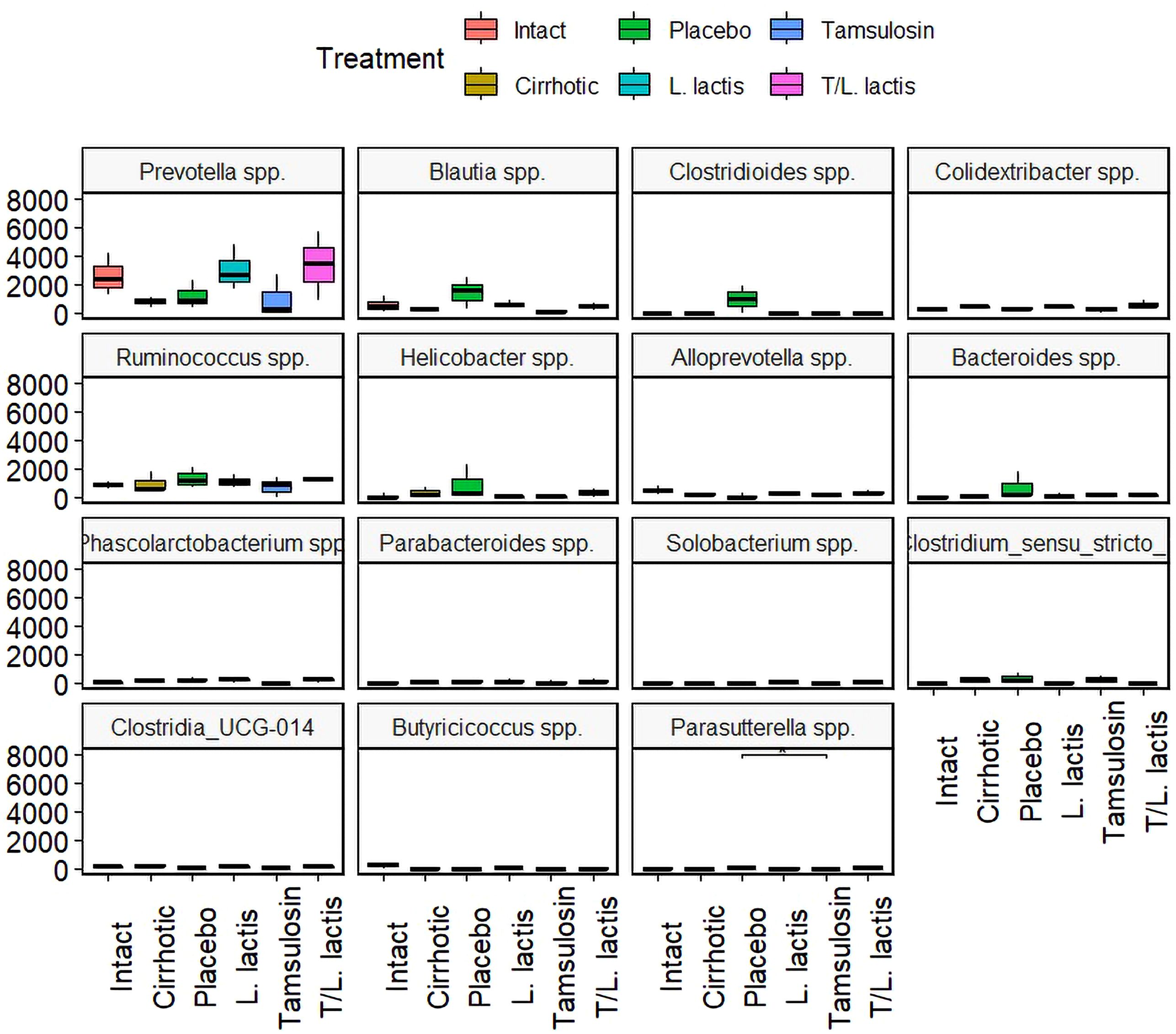
Figure 7. Change in intestinal microbial composition. Relative abundance box plot of intestinal microbiota at the genus level. Prevotella spp., showed higher abundance in the L. lactis and T/L. lactis groups. In contrast, the placebo group exhibited elevated levels of Blautia spp., Clostridioides spp., Helicobacter spp., Bacteroides spp., and Ruminococcus spp. The remaining genera displayed low or undetectable abundance across all groups.
3.3 Effect on hepatic damage
To evaluate the effect of tamsulosin and L. lactis administration on liver damage induced by cirrhosis, histopathological analyses and biochemical assessments of hepatic injury markers were performed. Histopathological analysis of liver tissue samples demonstrated distinct morphological differences across experimental groups. In the intact group, hepatic lobules exhibited a well-organized structure characterized by radially arranged hepatocyte cords surrounding the centrilobular vein and a typical distribution of peripheral collagen adjacent to the portal triad. In contrast, the cirrhotic group displayed a markedly disorganized architecture, with hepatocytes showing macrovesicular and microvesicular steatosis. Additionally, prominent type I collagen fibers (visualized via Sirius Red staining) were observed in both perivascular and lamellar regions. The placebo group revealed regenerative nodules and hepatocytes exhibiting ballooning degeneration, with nuclei displaying karyorrhexis and karyolysis; type I collagen distribution resembled the cirrhotic groups. In the groups treated with tamsulosin and L. lactis, discontinuous nodules were observed, accompanied by pyknotic hepatocytes and regions of reduced tissue damage relative to the cirrhotic and placebo groups. Notably, the tamsulosin and L. lactis (T/L. lactis) combined treatment group exhibited significant hepatic parenchyma recovery, with hepatocytes approaching a morphological appearance comparable to the intact group and a diminished presence of collagen fibers. (Figure 8A). Regarding the determination of serum markers of liver damage ALT, AST, and AP, cirrhotic animals showed significantly higher ALT(98.0 U/L; p < 0.01), AST (479.4 U/L, p < 0.001) and AP (667.5 U/L; p < 0.01) levels compared to the intact group (ALT: 34.8 U/L, AST: 82.6 U/L, and AP: 150.8 U/L). Rats treated with L. lactis, tamsulosin, and the T/L. lactis exhibited lower ALT (L. lactis; 43.8 U/L, tamsulosin; 42.8 U/L, T/L. lactis; 43.3 U/L) and AST (L. lactis; 230.8 U/L, tamsulosin; 108.8 U/L, T/L. lactis; 112.8 U/L) levels. Currently, AP was significantly reduced only in the tamsulosin group (109.0 U/L, p < 0.01) (Figure 8B).
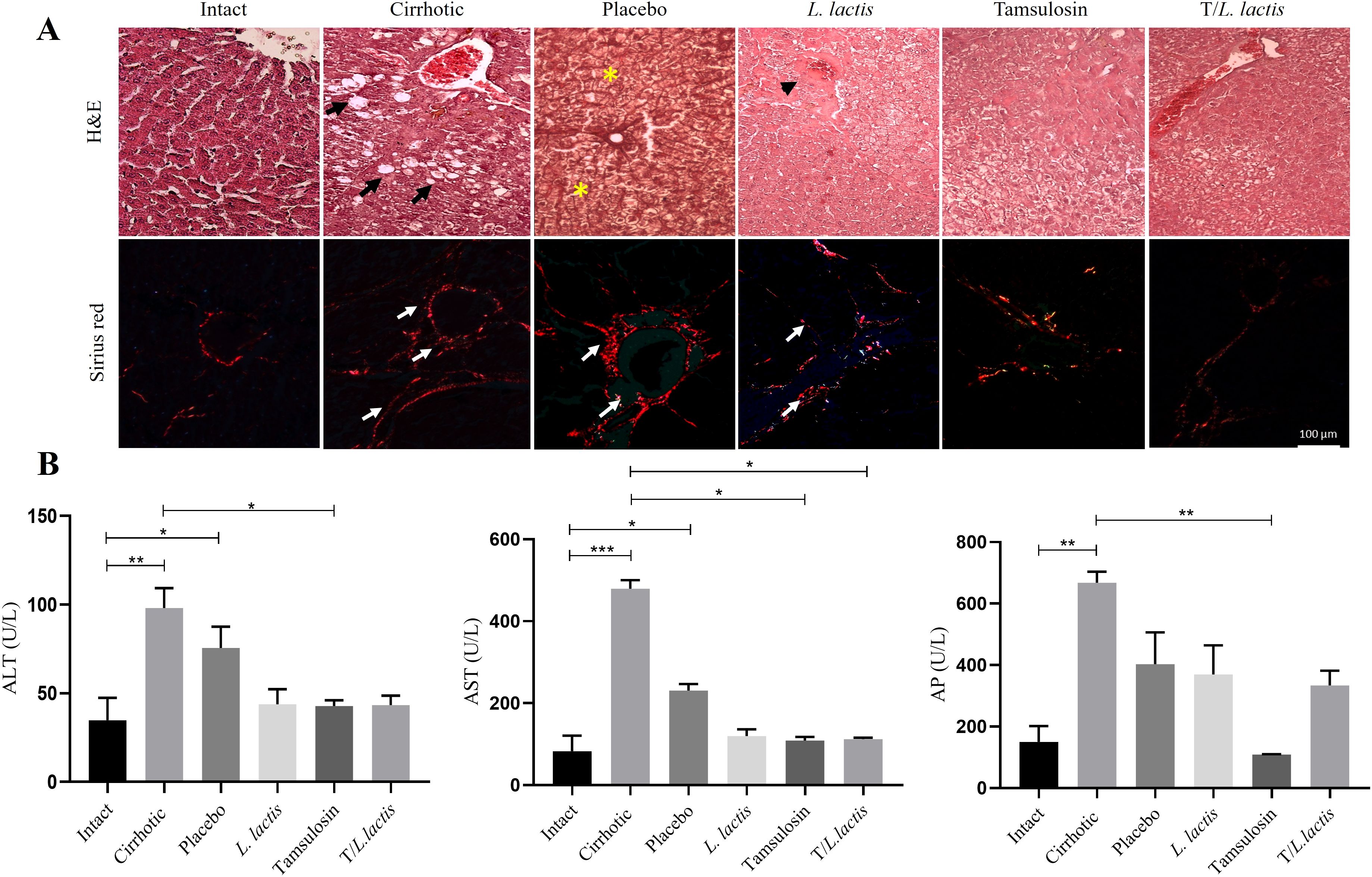
Figure 8. Effects of tamsulosin and L. lactis administration on liver morphology and function. (A) Analysis of the histological structure of the hepatic parenchyma after treatment, carried out with H&E and Sirius red. White arrow indicated fibrotic area (type I collagen fibers), black arrow indicated macrovesicular and microvesicular steatosis, yellow asterisks indicated regeneration nodules, black arrowhead indicated discontinuous nodules with pyknotic hepatocytes. Magnification, 20x. (B) The serum levels of ALT, AST, and AP were used as markers of liver damage. A decrease in ALT and AST levels was observed in the groups that received tamsulosin, L. lactis, and their combination. AP levels decreased significantly in the tamsulosin group. The results are presented as mean ± standard error of the mean. Samples from each rat were analyzed in triplicate. The statistical analysis was performed with the Kruskal-Wallis and Tukey post-test methods, where the values of *p < 0.05, **p < 0.01, and ***p < 0.001 were considered significant.
4 Discussion
In patients with liver cirrhosis, a dysregulation of the liver-gut axis has been found, where splanchnic hyperemia, intestinal dysmotility, and alterations in the intestinal mucosal barrier are observed, as well as changes in gastric function and microcirculation (Tsiaoussis et al., 2015). Jejunal biopsy studies have shown a reduction in the number and length of microvilli, along with thickening, suggesting a decreased absorptive surface area (Wakim-Fleming et al., 2008). In our study, we observed that cirrhotic rats exhibited marked intestinal damage, including epithelial and lamina propria thinning, and significant decreases in villus height and crypt depth. In contrast, the intestinal morphology in the L. lactis, tamsulosin, and combination treatment (T/L. lactis) groups closely resembled that of the intact controls, with preserved villus height and crypt depth. Consistent with previous findings showing that probiotic administration in TAA-induced cirrhotic rats significantly increased crypt depth, suggesting a protective or restorative effect on intestinal architecture (Jantararussamee et al., 2021). On the other hand, alterations in tight junction (TJ) proteins such as ZO-1, occludin, and claudin have been linked to increased intestinal permeability and systemic endotoxemia (Assimakopoulos et al., 2012). According to our analysis, ZO-1 and occludin expression were observed only in the group treated with L. lactis. Some studies have shown that probiotic supplementation can enhance intestinal barrier integrity (Rosenfeldt et al., 2004) by increasing the expression of tight junction proteins such as ZO-1, occludin, and claudins, conferring protective effects against epithelial barrier disruption (Karczewski et al., 2010; Miyauchi et al., 2012; Ukena et al., 2007; Wagnerberger et al., 2013). However, in our study, we did not observe such an effect in the T/L. lactis group. Further studies are required to confirm these findings.
Direct evidence regarding the effects of tamsulosin on the intestinal mucosa is limited; existing studies demonstrate that this α1-adrenergic antagonist enhances microcirculatory blood flow in highly vascularized tissues, such as the bladder, within ischemia-reperfusion models (Mizuno et al., 2010). Improved submucosal perfusion has been associated with reduced epithelial injury and accelerated tissue regeneration. Given that cirrhosis-associated intestinal hypoperfusion contributes significantly to epithelial barrier dysfunction (Di Pascoli et al., 2017), it is plausible that tamsulosin exerts protective effects on the intestinal mucosa through similar vascular mechanisms.
A randomized study conducted in male patients with lower urinary tract symptoms (LUTS) and benign prostatic hyperplasia (BPH) demonstrated that after four weeks of treatment with tamsulosin, stool consistency shifted toward softer forms, and constipation scores significantly decreased. These findings suggest that tamsulosin positively influences gastrointestinal transit, likely through the relaxation of intestinal smooth muscle (Matsumoto et al., 2020). Tamsulosin’s α1-adrenergic blockade targets receptors located on vascular and smooth muscle, including intestinal sphincters and smooth muscle layers, which mediate contraction. By blocking these receptors, tamsulosin promotes vasodilation, smooth muscle relaxation, and reduces sympathetic tone. In the setting of cirrhosis, such effects could improve intestinal perfusion and motility, thereby reducing luminal stasis and bacterial overgrowth. Additionally, emerging evidence suggests that tamsulosin may possess anti-inflammatory properties (Alabdali and Ibrahim, 2023), which could further contribute to the preservation of mucosal architecture. Nevertheless, further research is required to confirm these effects in the intestinal context directly.
Bacterial translocation is a key factor in the development of infectious complications in patients with cirrhosis. It contributes to systemic inflammation and worsening of liver function, thereby accelerating disease progression. For this to occur, significant changes must take place, involving modifications in the gut microbiota, damage to the epithelial barrier, and bacterial overgrowth (Simbrunner et al., 2023). The Enterobacteriaceae family (E. coli, Klebsiella spp.), enterococci, and Streptococcus spp., are the most observed organisms in bacterial translocation in humans. A study in mice demonstrated that Gram-negative bacteria translocate in large quantities to the MLN, instead of Gram-positive and obligate anaerobic bacteria translocate at much lower levels (Skinner et al., 2020). In our study, we found bacteria in MLNs, spleen, and liver. Specifically, we identified the presence of Escherichia coli (E. coli), Klebsiella pneumoniae (K. pneumoniae), Enterobacter agglomerans (E. agglomerans), and Proteus vulgaris (P. vulgaris) in the cirrhotic and placebo groups, but not in the groups treated with tamsulosin, L. lactis, or their combination. Similarly, endotoxin levels and bacterial DNA in blood were found in the cirrhotic and placebo groups. In contrast, endotoxins and bacterial DNA were not detected in the groups treated with L. lactis, tamsulosin, and T/L. lactis groups. Endotoxemia plays a central role in the pathophysiology of liver cirrhosis, as it reflects the abnormal translocation of bacterial endotoxins such as lipopolysaccharide (LPS) from the gut into the systemic circulation. This process results from increased intestinal permeability and the dysbiosis commonly observed in cirrhotic patients. Probiotic strains such as Bifidobacterium bifidum and Lactobacillus rhamnosus have been shown to mitigate bacterial translocation and lower systemic LPS levels by enhancing epithelial barrier function (Han et al., 2016). Several studies have shown that L. lactis can attenuate LPS-induced endotoxemia by reducing proinflammatory cytokines such as TNF-α and IL-6, preserving intestinal integrity, and limiting microbial translocation (Fu et al., 2024). These results reinforce the potential role of L. lactis in modulating host immune responses and preventing endotoxin-mediated complications, particularly in conditions characterized by gut barrier dysfunction, such as liver cirrhosis. Although tamsulosin has not been extensively investigated in intestinal models, the absence of endotoxemia and bacterial translocation in our study suggests a protective effect on the gut barrier. The tamsulosin has not been extensively investigated in intestinal models; the absence of endotoxemia and bacterial translocation in our study suggests an indirect protective effect on the intestinal barrier, as the attenuation of tamsulosin-induced liver inflammation could help reduce systemic inflammatory stress, promoting a more stable environment for the intestinal microbiota (Alabdali and Ibrahim, 2023). Numerous studies have reported alterations in the composition of the intestinal microbiome in various liver diseases, where, in addition to a reduction in species diversity, bacterial overgrowth occurs in the intestine, partly due to decreased intestinal motility (Tsiaoussis et al., 2015). In cirrhotic patients, a decrease in the α-diversity of the gut microbiota has been observed (Oh et al., 2020), and alterations in β-diversity have also been reported, particularly in individuals with NAFLD-related cirrhosis (Behary et al., 2021). These findings suggest that cirrhosis is associated with a less diverse and less stable gut microbiome. In this study, we found that L. lactis group exhibited increased α-diversity and a more uniform microbial composition, with a more stable gut environment. The T/L. lactis group also showed increased α-diversity, although with less compositional uniformity compared to L. lactis group. In addition, the microbial profiles found resembled those of the healthy group. Moreover, both groups exhibited microbial community structures (β-diversity) that more closely resembled those of healthy controls. In contrast, the tamsulosin group displayed reduced α-diversity and greater variability in microbial composition. Firmicutes and Bacteroidetes are the two most prominent bacterial phyla in the gastrointestinal tract, and their relative abundance, expressed as the Firmicutes/Bacteroidetes (F/B) ratio, has been recognized as an intestinal homeostasis key indicator. An increased proportion of Bacteroidetes and Proteobacteria is associated with inflammation (Stojanov et al., 2020). In our study, the untreated and tamsulosin groups exhibited a decrease in Firmicutes alongside an increase in Proteobacteria and Bacteroidetes. In contrast, in the groups receiving L. lactis and T/L. lactis, we observed a significant increase in Firmicutes and Bacteroidetes, and a decrease in Proteobacteria.
The class Clostridia was predominant in the cirrhotic and placebo groups, whereas in the groups that received treatment (with L. lactis and T/L. lactis), the abundance of Clostridia was markedly reduced, accompanied by an increase in the class Bacteroidia. Members of this class, particularly the genus Bacteroides, are well-known intestinal commensals that protect the gut from pathogenic bacteria and support the nutritional needs of other microorganisms in the gut, playing essential roles in the digestion of complex polysaccharides and the modulation of the immune system (Cheng et al., 2022). At the family level, potentially beneficial autochthonous taxa such as Lachnospiraceae, Ruminococcaceae, and Clostridiales XIV are reduced while potentially pathogenic taxa, including Staphylococcaceae, Enterobacteriaceae, and Enterococcaceae are increased (Bajaj et al., 2015; Chen et al., 2011). We observed a similar decrease in Lachnospiraceae and Ruminococcaceae in the cirrhotic group. In the group that received L. lactis, Lachnospiraceae increased, while Ruminococcaceae remained unchanged. The significance of Lachnospiraceae lies in its production of butyrate, which, along with other short-chain fatty acids, inhibits intestinal inflammation and maintains the intestinal barrier (Vacca et al., 2020). At the genus level, an increase in Prevotella spp., was observed in the groups treated with L. lactis and T/L. lactis, compared to the placebo, tamsulosin, and cirrhotic groups. In contrast, the genera Blautia spp., Clostridioides spp., Helicobacter spp., Ruminococcus spp., and Bacteroides spp., showed greater relative abundance in the placebo group, which could reflect a microbial imbalance characteristic of cirrhosis without intervention. Other genera such as Colidextrinbacter spp., Alloprevotella spp., Phascolarctobacterium spp., Parabacteroides spp., Solobacterium spp., Clostridium sensu stricto, Clostridia_UCG-014, Butyricicoccus spp., and Parasutterella spp., showed very low or undetectable levels in all groups, with no appreciable differences between treatments. The observed improvements in microbial composition following L. lactis administration suggests that this strain may exert beneficial modulatory effects on gut microbiota in cirrhotic conditions. L. lactis is known to compete with pathogenic bacteria through the production of antimicrobial peptides such as nisin and the acidification of the intestinal environment, thereby limiting the expansion of Proteobacteria and other opportunistic taxa (Bernbom et al., 2006). Additionally, L. lactis may indirectly promote the growth of beneficial commensals such as Lachnospiraceae and Prevotella spp., which are involved in short-chain fatty acid (SCFA) production and immune modulation (Ćesić et al., 2023; Winiarska-Mieczan et al., 2022).
The liver is continuously exposed to gut-derived microbial and metabolic signals through the portal vein, making it particularly sensitive to changes in intestinal homeostasis. The microbial components, such as lipopolysaccharide (LPS), activate hepatic immune responses via pattern recognition receptors like TLR4 (Seki et al., 2007). This leads to increased expression of pro-inflammatory cytokines, including IL-1β and TNF-α, and the activation of hepatic stellate cells (HSCs), contributing to fibrosis (Krenkel and Tacke, 2017). The progression of liver fibrosis is characterized by the distortion of normal hepatocyte structure, the formation of nodules, impaired blood flow, portal hypertension, hepatocellular carcinoma, and liver failure (Aydin and Akcali, 2018). Treatment with tamsulosin, L. lactis, and their combination was associated with a substantial recovery of liver architecture, with hepatocyte structures closely resembling those observed in the intact group. Furthermore, biochemical markers of liver injury showed that ALT and AST were lower in the L. lactis, tamsulosin, and T/L. lactis groups, while AP showed significant reduction only in the tamsulosin group. Tamsulosin has demonstrated antifibrotic and immunomodulatory effects in various experimental models. Its ability to reduce the expression of proinflammatory cytokines such as TNF-α and IL-6 (Ali Hasan and Abdulkhaliq Ibrahim, 2023) as well as profibrotic factors like TGF-β1 (Alrasheed et al., 2023) suggests a beneficial influence on the chronic hepatic inflammatory microenvironment. The use of L. lactis in a cirrhosis model prevented steatosis and fibrosis, and reduced serum AST and ALT. Likewise, an immunoregulatory process was observed since a notable decrease in the hepatic expression of IL-1β; this could be due to an increase in intestinal IL-10 as a result of the induced damage, which possibly has a hepatic effect due to the close relationship between the liver and the intestine (Delgado-Venegas et al., 2021).
In conclusion, the results of this study reinforce the crucial role of the gut–liver axis in the pathophysiology of liver cirrhosis and highlight the therapeutic potential of L. lactis and tamsulosin. The co-administration of L. lactis and tamsulosin promoted the restoration of microbial diversity, improved intestinal barrier integrity, and reduced bacterial translocation and endotoxemia. Complementarily, tamsulosin contributed to the preservation of liver architecture and significantly decreased hepatic enzyme levels. These findings suggest that modulating the gut microbiota and systemic factors through probiotic and pharmacological strategies may represent a promising approach to treat chronic liver diseases. However, additional studies with larger sample sizes are necessary to confirm these effects and clarify underlying mechanisms.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Ethics statement
The animal study was approved by Ethics Committee for the use of animals in teaching and research at UAA (CEADI−UAA, UAA: Autonomous University of Aguascalientes, AUT−B−C−1121−077). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
SP-G: Methodology, Investigation, Writing – original draft. AL-M: Supervision, Validation, Investigation, Writing – review & editing. MM-O: Investigation, Validation, Writing – review & editing, Data curation. DH-M: Validation, Supervision, Conceptualization, Writing – review & editing, Software. JV-J: Formal analysis, Funding acquisition, Conceptualization, Investigation, Validation, Writing – review & editing, Supervision. SM-H: Investigation, Writing – review & editing, Supervision, Funding acquisition, Conceptualization, Writing – original draft, Validation, Project administration, Methodology.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the PIBB24-3 grant of the Autonomous University of Aguascalientes for MSL. The master's fellowship for PSV (CVU Number: 1092421) was awarded by SECIHTI.
Acknowledgments
We thank Dr. Raúl O. Martínez-Rincón (Secihti-Center for Biological Research of the Northwest, SC.) for his valuable assistance in the analysis and graphical representation of bacterial genera. We also appreciate the support from M.Sc. Norma Adela Carrasco Esparza in the identification of bacterial strains.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alabdali, H. and Ibrahim, M. (2023). Study the anti-inflammatory effects of tamsulosin by the evaluation of inflammatory cells and lung histopathology in an airway inflammation model in rats. Bull. Pharm. Sci. Assiut 46, 633–645. doi: 10.21608/bfsa.2023.301290
Albillos, A., de Gottardi, A., and Rescigno, M. (2020). The gut-liver axis in liver disease: Pathophysiological basis for therapy. J. Hepatol. 72, 558–577. doi: 10.1016/j.jhep.2019.10.003
Ali Hasan, W. and Abdulkhaliq Ibrahim, M. (2023). An investigation of anti-inflammatory effects of famotidine in rats by evaluating inflammatory cells and lung histopathology: an airway model. . JAHP 12, 55–63. doi: 10.17582/journal.jahp/2024/12.1.55.63
Alrasheed, N. M., Alammari, R. B., Alshammari, T. K., Alamin, M. A., Alharbi, A. O., Alonazi, A. S., et al. (2023). α1A Adrenoreceptor blockade attenuates myocardial infarction by modulating the integrin-linked kinase/TGF-β/Smad signaling pathways. BMC Cardiovasc. Disord. 23, 153. doi: 10.1186/s12872-023-03188-w
Assimakopoulos, S. F., Tsamandas, A. C., Tsiaoussis, G. I., Karatza, E., Triantos, C., Vagianos, C. E., et al. (2012). Altered intestinal tight junctions’ expression in patients with liver cirrhosis: a pathogenetic mechanism of intestinal hyperpermeability. Eur. J. Clin. Invest. 42, 439–446. doi: 10.1111/j.1365-2362.2011.02609.x
Aydin, M. M. and Akcali, K. C. (2018). Liver fibrosis. Turk J. Gastroenterol. 29, 14–21. doi: 10.5152/tjg.2018.17330
Bajaj, J. S., Betrapally, N. S., Hylemon, P. B., Heuman, D. M., Daita, K., White, M. B., et al. (2015). Salivary microbiota reflects changes in gut microbiota in cirrhosis with hepatic encephalopathy. Hepatology 62, 1260–1271. doi: 10.1002/hep.27819
Behary, J., Amorim, N., Jiang, X.-T., Raposo, A., Gong, L., McGovern, E., et al. (2021). Gut microbiota impact on the peripheral immune response in non-alcoholic fatty liver disease related hepatocellular carcinoma. Nat. Commun. 12, 187. doi: 10.1038/s41467-020-20422-7
Bernbom, N., Licht, T. R., Brogren, C.-H., Jelle, B., Johansen, A. H., Badiola, I., et al. (2006). Effects of lactococcus lactis on composition of intestinal microbiota: role of nisin. Appl. Environ. Microbiol. 72, 239–244. doi: 10.1128/AEM.72.1.239-244.2006
Buendía-Delgado, R., Guerrero-Alba, R., Martínez-Hernández, S., Muñoz-Ortega, M., and Medina-Pizaño, M. (2022). In vitro evaluation of the antifibrogenic effect of tamsulosin during its interaction with activated stellate cells. Ann. Hepatol. 27, 100852. doi: 10.1016/j.aohep.2022.100852
Ćesić, D., Lugović Mihić, L., Ozretić, P., Lojkić, I., Buljan, M., Šitum, M., et al. (2023). Association of gut lachnospiraceae and chronic spontaneous urticaria. Life 13, 1280. doi: 10.3390/life13061280
Chen, Y., Yang, F., Lu, H., Wang, B., Chen, Y., Lei, D., et al. (2011). Characterization of fecal microbial communities in patients with liver cirrhosis. Hepatology 54, 562–572. doi: 10.1002/hep.24423
Cheng, J., Hu, J., Geng, F., and Nie, S. (2022). Bacteroides utilization for dietary polysaccharides and their beneficial effects on gut health. Food Sci. Hum. Wellness 11, 1101–1110. doi: 10.1016/j.fshw.2022.04.002
Delgado-Venegas, C. S., Martínez-Hernández, S. L., Cervantes-García, D., Montes de Oca-Luna, R., de Jesús Loera-Arias, M., Mata-Martínez, M. G., et al. (2021). Modulating effects of the probiotic Lactococcus lactis on the hepatic fibrotic process induced by CCl4 in Wistar rats. Exp. Ther. Med. 21, 339. doi: 10.3892/etm.2021.9770
Di Pascoli, M., Sacerdoti, D., Pontisso, P., Angeli, P., and Bolognesi, M. (2017). Molecular mechanisms leading to splanchnic vasodilation in liver cirrhosis. J. Vasc. Res. 54, 92–99. doi: 10.1159/000462974
Dunn, C. J., Matheson, A., and Faulds, D. M. (2002). Tamsulosin: A review of its pharmacology and therapeutic efficacy in the management of lower urinary tract symptoms. Drugs Aging 19, 135–161. doi: 10.2165/00002512-200219020-00004
Fu, Y., Zhang, S., Yue, Q., An, Z., Zhao, M., Zhao, C., et al. (2024). The preventative effects of Lactococcus Lactis metabolites against LPS-induced sepsis. Front. Microbiol. 15. doi: 10.3389/fmicb.2024.1404652
Fukui, H. (2021). Leaky gut and gut-liver axis in liver cirrhosis: clinical studies update. Gut Liver 15, 666–676. doi: 10.5009/gnl20032
Garcı、a-Tsao, G. (2001). Bacterial translocation: cause or consequence of decompensation in cirrhosis? J. Hepatol. 34, 150–155. doi: 10.1016/S0168-8278(00)00006-4
Ginès, P., Krag, A., Abraldes, J. G., Solà, E., Fabrellas, N., and Kamath, P. S. (2021). Liver cirrhosis. Lancet 398, 1359–1376. doi: 10.1016/S0140-6736(21)01374-X
Han, C., Ding, Z., Shi, H., Qian, W., Hou, X., and Lin, R. (2016). The role of probiotics in lipopolysaccharide-induced autophagy in intestinal epithelial cells. Cell Physiol. Biochem. 38, 2464–2478. doi: 10.1159/000445597
Jantararussamee, C., Rodniem, S., Taweechotipatr, M., Showpittapornchai, U., and Pradidarcheep, W. (2021). Hepatoprotective effect of probiotic lactic acid bacteria on thioacetamide-induced liver fibrosis in rats. Probiotics Antimicro. Prot. 13, 40–50. doi: 10.1007/s12602-020-09663-6
Karczewski, J., Troost, F. J., Konings, I., Dekker, J., Kleerebezem, M., Brummer, R.-J. M., et al. (2010). Regulation of human epithelial tight junction proteins by Lactobacillus plantarum in vivo and protective effects on the epithelial barrier. Am. J. Physiol. Gastrointest Liver Physiol. 298, G851–G859. doi: 10.1152/ajpgi.00327.2009
Krenkel, O. and Tacke, F. (2017). Liver macrophages in tissue homeostasis and disease. Nat. Rev. Immunol. 17, 306–321. doi: 10.1038/nri.2017.11
Laguna, J. G., Freitas, A. D. S., Barroso, F. A. L., De Jesus, L. C. L., De Vasconcelos, O. A. G. G., Quaresma, L. S., et al. (2024). Recombinant probiotic Lactococcus lactis delivering P62 mitigates moderate colitis in mice. Front. Microbiol. 15. doi: 10.3389/fmicb.2024.1309160
Lee, N. Y. and Suk, K. T. (2020). The role of the gut microbiome in liver cirrhosis treatment. IJMS 22, 199. doi: 10.3390/ijms22010199
Liu, N., Zhang, X., Liang, C., Yao, D., Liu, L., Zhao, D., et al. (2009). Dynamic changes of alpha-AR, beta1-AR and beta2-AR expression during hepatic fibrogenesis. Zhonghua Gan Zang Bing Za Zhi 17, 653–656.
Livak, K. J. and Schmittgen, T. D. (2001). Analysis of relative gene expression data using real-time quantitative PCR and the 2–ΔΔCT method. Methods 25, 402–408. doi: 10.1006/meth.2001.1262
Macías-Pérez, J. R., Vázquez-López, B. J., Muñoz-Ortega, M. H., Aldaba-Muruato, L. R., Martínez-Hernández, S. L., Sánchez-Alemán, E., et al. (2019). Curcumin and α/β -adrenergic antagonists cotreatment reverse liver cirrhosis in hamsters: participation of nrf-2 and NF- κ B. J. Immunol. Res. 2019, 1–12. doi: 10.1155/2019/3019794
Matsumoto, S., Kasamo, S., and Hashizume, K. (2020). Influence of alpha-adrenoceptor antagonists therapy on stool form in patients with lower urinary tract symptoms suggestive of benign prostatic hyperplasia. Low Urin Tract Symptoms 12, 86–91. doi: 10.1111/luts.12289
Miyauchi, E., O’Callaghan, J., Buttó, L. F., Hurley, G., Melgar, S., Tanabe, S., et al. (2012). Mechanism of protection of transepithelial barrier function by Lactobacillus salivarius: strain dependence and attenuation by bacteriocin production. Am. J. Physiol. Gastrointest Liver Physiol. 303, G1029–G1041. doi: 10.1152/ajpgi.00003.2012
Mizuno, H., Yamamoto, T., Okutsu, H., Ohtake, A., Sasamata, M., Matsukawa, Y., et al. (2010). Effect of tamsulosin on bladder microcirculation in a rat ischemia-reperfusion model, evaluated by pencil lens charge-coupled device microscopy system. Urology 76, 1266.e1–1266.e5. doi: 10.1016/j.urology.2010.06.039
Muñoz, L. I. O. (2001). Norma Oficial Mexicana NOM−062−ZOO−1999, especificaciones técnicas para la producción, cuidado y uso de los animales de laboratorio. Diario Oficial de la Federación (Ciudad de México: Secretaría de Agricultura, Ganadería, Desarrollo Rural, Pesca y Alimentación).
National Research Council (2011). National research council (US) committee for the update of the guide for the care and use of laboratory animals: guide for the care and use of laboratory animals. 8th ed (Whashington, D.C: The National academies press).
Oben, J. A. and Diehl, A. M. (2004). Sympathetic nervous system regulation of liver repair. Anat. Rec. 77. doi: 10.1002/ar.a.20081
Oh, T. G., Kim, S. M., Caussy, C., Fu, T., Guo, J., Bassirian, S., et al. (2020). A universal gut-microbiome-derived signature predicts cirrhosis. Cell Metab. 32, 901. doi: 10.1016/j.cmet.2020.10.015
Parapouli, M., Delbès-Paus, C., Kakouri, A., Koukkou, A.-I., Montel, M.-C., and Samelis, J. (2013). Characterization of a wild, novel nisin a-producing Lactococcus strain with an L. lactis subsp. cremoris genotype and an L. lactis subsp. lactis phenotype, isolated from Greek raw milk. Appl. Environ. Microbiol. 79, 3476–3484. doi: 10.1128/AEM.00436-13
Roehlen, N., Crouchet, E., and Baumert, T. F. (2020). Liver fibrosis: mechanistic concepts and therapeutic perspectives. Cells 9, 875. doi: 10.3390/cells9040875
Rosenfeldt, V., Benfeldt, E., Valerius, N. H., Paerregaard, A., and Michaelsen, K. F. (2004). Effect of probiotics on gastrointestinal symptoms and small intestinal permeability in children with atopic dermatitis. J. Pediatr. 145, 612–616. doi: 10.1016/j.jpeds.2004.06.068
Seki, E., De Minicis, S., Österreicher, C. H., Kluwe, J., Osawa, Y., Brenner, D. A., et al. (2007). TLR4 enhances TGF-β signaling and hepatic fibrosis. Nat. Med. 13, 1324–1332. doi: 10.1038/nm1663
Serna-Salas, S. A., Arroyave-Ospina, J. C., Zhang, M., Damba, T., Buist-Homan, M., Muñoz-Ortega, M. H., et al. (2022). α-1 Adrenergic receptor antagonist doxazosin reverses hepatic stellate cells activation via induction of senescence. Mech. Ageing Dev. 201, 111617. doi: 10.1016/j.mad.2021.111617
Sigala, B., McKee, C., Soeda, J., Pazienza, V., Morgan, M., Lin, C., et al. (2013). Sympathetic nervous system catecholamines and neuropeptide y neurotransmitters are upregulated in human NAFLD and modulate the fibrogenic function of hepatic stellate cells. Plos One 95. doi: 10.1371/journal.pone.0072928
Simbrunner, B., Caparrós, E., Neuwirth, T., Schwabl, P., Königshofer, P., Bauer, D., et al. (2023). Bacterial translocation occurs early in cirrhosis and triggers a selective inflammatory response. Hepatol. Int. 17, 1045–1056. doi: 10.1007/s12072-023-10496-y
Skinner, C., Thompson, A. J., Thursz, M. R., Marchesi, J. R., and Vergis, N. (2020). Intestinal permeability and bacterial translocation in patients with liver disease, focusing on alcoholic etiology: methods of assessment and therapeutic intervention. Therap Adv. Gastroenterol. 13, 1756284820942616. doi: 10.1177/1756284820942616
Soleil, S., Marichal-Cansino, B., Martínez-Hernández, S., Ventura-Juárez, J., and Muñoz-Ortega, M. (2022). Role of tamsulosin in recovery from thioacetamide-induced subchronic liver damage in a Wistar rat model. Ann. Hepatol. 27, 100866. doi: 10.1016/j.aohep.2022.100866
Song, Y., Chen, G., Huang, P., Hu, C., and Liu, X. (2020). Effects of tamsulosin combined with solifenacin on lower urinary tract symptoms: evidence from a systematic review, meta-analysis, and trial sequential analysis of randomized controlled trials. Front. Pharmacol. 11. doi: 10.3389/fphar.2020.00763
Stojanov, S., Berlec, A., and Štrukelj, B. (2020). The influence of probiotics on the firmicutes/bacteroidetes ratio in the treatment of obesity and inflammatory bowel disease. Microorganisms 8, 1715. doi: 10.3390/microorganisms8111715
Tsiaoussis, G. I., Assimakopoulos, S. F., Tsamandas, A. C., Triantos, C. K., and Thomopoulos, K. C. (2015). Intestinal barrier dysfunction in cirrhosis: Current concepts in pathophysiology and clinical implications. World J. Hepatol. 7, 2058–2068. doi: 10.4254/wjh.v7.i17.2058
Ukena, S. N., Singh, A., Dringenberg, U., Engelhardt, R., Seidler, U., Hansen, W., et al. (2007). Probiotic Escherichia coli Nissle 1917 inhibits leaky gut by enhancing mucosal integrity. PloS One 2, e1308. doi: 10.1371/journal.pone.0001308
Vacca, M., Celano, G., Calabrese, F. M., Portincasa, P., Gobbetti, M., and De Angelis, M. (2020). The controversial role of human gut lachnospiraceae. Microorganisms 8, 573. doi: 10.3390/microorganisms8040573
Wagnerberger, S., Spruss, A., Kanuri, G., Stahl, C., Schröder, M., Vetter, W., et al. (2013). Lactobacillus casei Shirota protects from fructose-induced liver steatosis: a mouse model. J. Nutr. Biochem. 24, 531–538. doi: 10.1016/j.jnutbio.2012.01.014
Wakim-Fleming, J., Zein, N. N., Bennett, A., Lopez, R., Santisi, J., and Carey, W. D. (2008). Histological abnormalities of the small bowel mucosa in cirrhosis and portal hypertension. WJG 14, 6370. doi: 10.3748/wjg.14.6370
Wiest, R., Lawson, M., and Geuking, M. (2014). Pathological bacterial translocation in liver cirrhosis. J. Hepatol. 60, 197–209. doi: 10.1016/j.jhep.2013.07.044
Keywords: tamsulosin, L. lactis, microbiota, intestinal permeability, bacterial translocation, cirrhosis
Citation: Padilla-García SV, Loera-Muro A, Muñoz-Ortega MH, Hernández-Marín DA, Ventura-Juárez J and Martínez-Hernández SL (2025) Microbiota composition and intestinal barrier function modulated by tamsulosin and Lactococcus lactis in a cirrhosis rat model. Front. Cell. Infect. Microbiol. 15:1624065. doi: 10.3389/fcimb.2025.1624065
Received: 06 May 2025; Accepted: 18 July 2025;
Published: 02 September 2025.
Edited by:
Xiong Jiang, Hubei Three Gorges Polytechnic, ChinaReviewed by:
Yongfang Xie, Chongqing University of Post and Telecommunications, ChinaNageena Qayyum, Northwest A&F University, China
Copyright © 2025 Padilla-García, Loera-Muro, Muñoz-Ortega, Hernández-Marín, Ventura-Juárez and Martínez-Hernández. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Sandra Luz Martínez-Hernández, bHV6Lm1hcnRpbmV6QGVkdS51YWEubXg=