 Felix Schweitzer1,2
Felix Schweitzer1,2 Lara-Jasmin Schröder2,3
Lara-Jasmin Schröder2,3 Alina Friedrichs1,4
Alina Friedrichs1,4 Viktoria Gudi3
Viktoria Gudi3 Thomas Skripuletz3
Thomas Skripuletz3 Imke Steffen1,4†
Imke Steffen1,4† Martin Palus5,6
Martin Palus5,6 Daniel Růžek5,6,7
Daniel Růžek5,6,7 Albert Osterhaus1,2
Albert Osterhaus1,2 Chittappen Kandiyil Prajeeth1*
Chittappen Kandiyil Prajeeth1*- 1Research Center for Emerging Infections and Zoonoses, University of Veterinary Medicine, Hannover, Germany
- 2Center for Systems Neuroscience (ZSN), Hannover, Germany
- 3Department of Neurology, Hannover Medical School, Hannover, Germany
- 4Institute of Biochemistry, University of Veterinary Medicine, Hannover, Germany
- 5Institute of Parasitology, Biology Centre of the Czech Academy of Sciences, Ceske Budejovice, Czechia
- 6Laboratory of Emerging Viral Infections, Veterinary Research Institute, Brno, Czechia
- 7Department of Experimental Biology, Faculty of Science, Masaryk University, Brno, Czechia
Introduction: The genus Orthoflavivirus of the Flaviviridae family includes several notable pathogens such as mosquito-borne West-Nile virus (Orthoflavivirus nilense, WNV) and Tick-borne encephalitis virus (Orthoflavivirus encephalitidis, TBEV) that are highly neurotropic and may cause severe neurological disease leading to lifelong disabilities, coma and death. These viruses have developed mechanisms to breach the compact blood-brain barrier (BBB) and establish infection within the central nervous system (CNS). Nevertheless, neuroinvasive mechanisms of orthoflaviviruses remain poorly understood. Complex anatomy of the CNS and the organization of the BBB is a major challenge to study neuroinvasion of orthoflaviviruses in vivo. Therefore, in vitro BBB models are useful tools to study direct interaction of viruses with the endothelial barrier.
Methods: In this study, we employed an in vitro transwell BBB model comprising primary mouse brain microvascular endothelial cells and astrocytes to compare the ability of mosquito-borne and tick-borne orthoflaviviruses to cross a compact endothelial barrier and reach the basolateral compartment of the transwell system. The influence of virus inoculation on the barrier properties was determined by measuring transendothelial electrical resistance (TEER).
Results: The results demonstrate that while pathogenic WNV and TBEV cross the endothelial barrier the ability of low pathogenic Usutu virus (USUV) and Langat virus (LGTV) was inconsistent. All viruses tested display virus replication within the endothelial cells. Nevertheless, virus replication did not affect the barrier function of endothelial cells as demonstrated by sustained TEER and absence of leakage of high molecular weight dextran molecules through the endothelial barrier even at several hours post infection.
Discussion: Our findings indicate that orthoflaviviruses can infect the endothelial cells, replicate within them without affecting the cells and its barrier function. Nevertheless, only pathogenic WNV and TBEV showed the ability to cross the endothelial barrier and reach the basolateral compartment.
1 Introduction
Changing climatic conditions and spread of vectors capable of transmitting disease have led to the emergence of several viral diseases in areas where such cases were previously not reported (Pradier et al., 2012; Mora et al., 2022; Ruscher et al., 2023). This includes viruses of the genus Orthoflavivirus, which consists of notable pathogens transmitted through arthropod vectors. Amongst these are mosquito-borne West-Nile virus (Orthoflavivirus nilense, WNV) and Tick-borne encephalitis virus (Orthoflavivirus encephalitidis, TBEV), known for their neurotropism and thus responsible for severe debilitating neurological disease (Mandl, 2005; Lim et al., 2011).
The genome of orthoflaviviruses is organized in a single open reading frame, which codes for one polyprotein and is further processed into three structural and seven non-structural proteins. The structural and non-structural proteins play pivotal role in the pathogenesis of orthoflaviviruses (Kuno et al., 1998; Neufeldt et al., 2018; Postler et al., 2023). Several orthoflaviviruses share a certain degree of amino acid sequence homology and exhibit varying levels of immune cross reactivity (Mandl et al., 1991; Iacono-Connors and Schmaljohn, 1992; Heinz and Stiasny, 2012; Rathore and St. John, 2020). Hence, they are grouped into distinct serogroups, which are also linked to their respective arthropod vectors. Within the mosquito-borne serogroup, the cluster of Japanese encephalitis virus (Orthoflavivirus japonicum, JEV) and its relatives, such as WNV, further includes Usutu virus (Orthoflavivirus usutuense, USUV). The latter causes disease in birds, but is virtually non-pathogenic in immunocompetent humans (Agliani et al., 2023). Similarly, in the tick-borne serogroup, Langat virus (Orthoflavivirus langatense, LGTV) is a close relative of TBEV sharing significant homology and is low- or non-pathogenic in humans. Hence, LGTV has been tested as live attenuated vaccine in the past. However, following vaccine trials conducted in early 1970’s using live LGTV, neurological complications were reported in few vaccinees. This highlighted the risk of using such attenuated strains as vaccines without proper understanding of the mechanism of viral neuropathogenesis (Smorodintsev and Dubov, 1986; Gritsun et al., 2003).
Studies have shown that after introduction to the vertebrate host, these viruses replicate at the peripheral sites and reach the central nervous system (CNS) following a viraemic phase (Ruzek et al., 2019; Habarugira et al., 2020). Since exchange across the compact blood-brain barrier (BBB) is highly regulated, invading pathogens or cellular infiltrates are usually excluded and hence the BBB forms an anatomical barrier to protect the CNS from adverse effects. While endothelial cells constitute the principal structural component of the BBB, astrocytes also are a critical part as they provide essential growth factors needed for maintaining barrier integrity. The compactness of the BBB is mainly attributed to tight-junction proteins, such as occludin, claudin-5 and zonula occludens protein (ZO-)1 that tightly bind the endothelial cells together. As a result pathogens are prevented from using paracellular spaces to gain access into the CNS (Kadry et al., 2020; Dunton et al., 2021). Nevertheless, several neurotropic viruses have evolved mechanisms to circumvent the compact BBB and enter the CNS. These include non-destructive mechanisms by which the viruses cross endothelial cells without causing any significant damage to the BBB. In this case, viruses use transcellular routes where they enter endothelial cells on the luminal side and are released into the CNS on the opposite side (Cain et al., 2019). On the other hand, viruses that have evolved more disruptive mechanisms do so either by modulating the expression of tight junction proteins as a consequence of inflammatory reaction or by inducing cell death by replicating within the endothelial cells. This leads to a loss of barrier integrity, thus allowing the viruses to take paracellular routes to reach the CNS (Wang et al., 2004; Rochfort et al., 2014). Based on the current evidence it remains speculative if initial neuroinvasion is non-disruptive, and the breakdown of the BBB during the course of infection is a consequence of subsequent neuroinflammation upon virus replication within the CNS.
In this study, we employed an in vitro transwell endothelial-astrocyte barrier model, to gain insights into the neuroinvasive mechanisms of mosquito- and tick-borne orthoflaviviruses such as WNV, USUV, TBEV and LGTV displaying varying degrees of pathogenicity for animals and humans.
2 Materials and methods
2.1 Ethics statement
Mice were maintained and bred in-house in compliance with European guidelines (EU directive on animal testing 2010/63/EU) and German Animal Welfare Law. The sacrifice of neonatal mice for collecting brain was performed after obtaining approvals from animal welfare authorities (approval no. TiHo-T-2023-12, TiHo-T-2024-14).
2.2 Viruses
WNV (strain NY99) was obtained from Martin Groschup, Friedrich-Loeffler-Institute, Greifswald, Germany and initial viral stocks were generated by propagating it in Vero cells. The source and propagation of TBEV (strain Neudoerfl) and LGTV (strain TP21) used in this study was previously described (Petry et al., 2021; Kubinski et al., 2023). USUV was isolated from a dead blackbird in Hannover, Germany and grown in PK-15 cells. Phylogenetic analysis showed that this isolate clustered along with Africa 2 strains of USUV (Störk et al., 2021). All viruses were propagated on BHK-21 cells by using an initial inoculum of 5×105 TCID50/ml in T75 flasks. At three days post infection (dpi) culture supernatants were harvested, centrifuged at 1000xg for 10 minutes to remove cell debris and subsequently concentrated by centrifugation through amicon® Ultra-15 30k centrifugal filters (Merck Millipore, Darmstadt, Germany) at 2500xg for 15 minutes. Viral titers of the stocks generated were determined using the 50% tissue culture infectious dose (TCID50) on BHK-21 cells. Viral stocks were stored at -80°C until further use.
2.3 Primary mouse brain microvascular endothelial cells
C57BL/6 mouse primary brain microvascular endothelial cells (mBMEC) were obtained from CellBiologics Inc. (Chicago, IL, USA) and maintained in culture as per the manufacturer’s instructions. Initially, cells were grown in T25 culture flasks coated with gelatine-based coating solution (GCS, 0.1% ready-to-use solution; CellBiologics, Chicago, IL, USA) in complete mouse endothelial cell medium (CECM; CellBiologics, Chicago, IL, USA) until confluent. Following this, cells were harvested by trypsinization, transferred into T75 culture flasks and grown until confluent, and subsequently passaged in a similar manner. At the time of cell harvest, several aliquots of cells from each passage were frozen in CECM with additional fetal bovine serum (FBS, final 50%vol.; Thermo Fisher Scientific gibco, Waltham, MA, USA) and dimethyl sulfoxide (DMSO, final 10%vol.; Carl Roth, Karlsruhe, Germany) and stored at -150°C until further use. For the experiments endothelial cells were used between passage 3 to 5.
2.4 Primary astrocyte isolation
Primary mouse astrocytes were harvested from neonatal mouse brain mixed glia cultures as previously described (Prajeeth et al., 2018). Briefly, brains were collected from neonatal mice (P1-P4) after decapitation. Subsequently meninges were removed, and tissue was digested by enzymatic treatment with trypsin-EDTA (0.05%; Thermo Fisher Scientific gibco, Waltham, MA, USA) followed by treatment with bovine pancreas DNase (1mg/ml; Roche Merck, Darmstadt, Germany). Enzymatic reaction was stopped by application of soybean trypsin-inhibitor (10mg/ml; 50%vol.; Sigma-Aldrich Merck, Darmstadt, Germany)-based triturating solution (1%w/vol. bovine serum albumin (BSA; Sigma-Aldrich Merck, Darmstadt, Germany), 0.03%vol. DNase (Sigma-Aldrich Merck, Darmstadt, Germany) in PBS) and the suspension was filtered through a 70µm pore size sieve. Single cell suspensions obtained were seeded in poly-L-lysine (PLL, 100µg/ml; Sigma-Aldrich Merck, Darmstadt, Germany)-coated T75 flasks and cultured in Dulbecco’s Modified Eagle Medium (DMEM) supplemented with 4.5g/l D-glucose and L-glutamine (Thermo Fisher Scientific gibco, Waltham, MA, USA), FBS (final 10%vol.; Thermo Fisher Scientific gibco, Waltham, MA, USA) and 10,000 units of penicillin together with 10mg/ml streptomycin (PenStrep; Thermo Fisher Scientific gibco, Waltham, MA, USA) (MGP+ medium). After 14 to 21 days, flasks were treated with 10µM cytosine β-D-arabinofuranoside (ara-C; Thermo Fisher Scientific gibco, Waltham, MA, USA) for three days to eliminate proliferating microglia and oligodendrocyte precursor cells from the culture and the unaffected astrocytes cell layer was harvested by mild trypsinization (0.05% trypsin-EDTA for 10 min. at 37°C). At this point aliquots of cells were made in MGP+ medium with additional FBS (final 50%vol.; Thermo Fisher Scientific gibco, Waltham, MA, USA) and DMSO (final 10%vol.; Carl Roth, Karlsruhe, Germany) and stocks were stored at -150°C until further use in experiments. For each experiment, a fresh stock was thawed and cultured in a T75 flaks until confluency was reached, before cells were harvested as described above and adjusted cell solution was seeded onto transwell inserts as described below.
2.5 BHK-21 cells
Baby hamster kidney (BHK-)21 (C-13) cells (ATCC, CCL-10; Manassas, VA, USA) were cultured in minimal essential medium (including Earl’s salts and L-glutamine, Thermo Fisher Scientific gibco, Waltham, MA, USA) supplemented with PenStrep (1%vol.) and HEPES buffer solution (final concentration of 20mM; Thermo Fisher Scientific Gibco, Waltham, MA, USA) (MEM+H). For TCID50 assays, cells were harvested from cultures by trypsinization and seeded in 96-well plates at a density of 2×104 cells per well, one day before assay.
2.6 In vitro transwell barrier model
As described by (Marshall et al., 2024). mBMECs and astrocytes were either cultured alone or in co-culture to establish a compact barrier model. For this purpose, polyester membranes of 24-well transwell inserts (pore size of 0.4µm; Corning® costar®, Corning, NY, USA) were coated with GCS on the apical side and PLL on the basolateral side. For mBMECs-astrocytes co-cultures, 5×104 astrocytes were seeded onto the PLL-coated basal side of the inserts and cultured for three days in MGP+ medium. At this point the apical side of the same inserts was coated with GCS and 5×104 mBMECs were seeded in CECM onto the apical side. A day later inserts were transferred into a cellZscope-device (cellZscope+®, nanoAnalytics, Münster, Germany) and cultured until they were confluent. The cellZscope allowed real-time monitoring of transendothelial electrical resistance (TEER), values of which correspond to compactness and monolayer formation respectively. A plateau was observed in TEER at around 4 days in culture where the measured TEER was approximately 20-25Ωcm2 and at this point the cells formed a tight barrier. Supplementary Figure 1 created using Biorender depicts the barrier model consisting of endothelial monoculture (Supplementary Figure 1A) and endothelial-astrocyte coculture on transwell inserts (Supplementary Figure 1B).
2.7 Barrier permeability assays
The compactness of the barrier model on transwell inserts was further assessed using a fluorescein isothiocyanate (FITC)-dextran barrier permeability assay as described in (Schweitzer et al., 2025). For this purpose, 70kDa FITC-dextran solution (Sigma Aldrich, St. Louis, MI, USA) was added at a final concentration of 1mg/ml to the apical side of inserts cultured with mBMEC in mono- or in co-culture with astrocytes after a desired TEER value (>20Ωcm2) was reached. Subsequently, culture supernatants from the basolateral compartments were collected at 0, 30, 60, 120 and 240 minutes after FITC-dextran application. For certain experiments the sampling of culture supernatants was done at 300 minutes after FITC-dextran application. Subsequently, fluorescence in the culture supernatants was measured (excitation wavelength: 485nm, emission wavelength: 535nm; Infinite® 200 Pro plate reader, TECAN Group, Männedorf, Switzerland). To quantify translocated FITC-dextran a standard curve was generated by measuring fluorescence of a 2-fold dilution series of the FITC-dextran stock solution. Similarly, barrier permeability assays were also performed using inserts collected at 24 hours post inoculation (hpi) and 48hpi to assess the effect of virus inoculation.
2.8 Virus inoculation and translocation of barrier model
To the compact mBMEC-astrocyte co-cultures on transwell inserts 2.5×105 TCID50 of WNV, TBEV, USUV, or LGTV were added respectively, to the apical compartment in 100µl of serum-free CECM. After one hour of inoculation at 37°C, the inoculum and basolateral supernatants were sampled and inserts were washed twice with CECM (apical side) and MGP+ medium (basolateral side), respectively, before fresh media were added to apical and basolateral compartments. Inserts were returned to the cellZscope and TEER was monitored over a period of up to 48hpi. At 24hpi and 48hpi, respectively, culture supernatants were collected from apical and basolateral compartments of respective inserts and viral titers within the culture supernatants were determined by TCID50 assays as described below. For determining the barrier integrity post infection, inserts were taken for the above-described barrier permeability assays.
2.9 Virus replication in endothelial cells
For assessing viral replication in brain endothelial cells, mBMECs (1×105) were seeded into each well of 8-well chamber slides (SARSTEDT, Nümbrecht, Germany) coated with GCS. The following day, cells were infected with 5×105 TCID50 of WNV, TBEV, USUV or LGTV respectively, for one hour at 37°C and 5% CO2 in serum-free CECM. After thoroughly washing the initial inoculum with serum-free CECM culture supernatants were collected from respective wells at regular intervals from 0hpi to 72hpi and at 144hpi. Viral titers in culture supernatants were determined using TCID50 assays. At each timepoint the respective chamber slide was washed thoroughly, fixed with paraformaldehyde (4% in PBS) for 20 minutes and used for immunocytochemistry analyses. Fixed cells were stored at 4°C in double-distilled water until immunocytochemistry assays were performed.
2.10 Virus quantification
Quantification of infectious viral particles of all sampled supernatants was done by TCID50 assays on BHK-21 cells and calculations were performed according to the method described by Reed and Muench (Reed and Muench, 1938). For this, supernatants were thawed and diluted in MEM+H (supplemented with 2% FBS instead of 10% FBS) as follows. Supernatants of the mBMEC monoculture infections in 8-well chamber slides were diluted 1:10, viral inoculums collected from the apical compartment of the inserts used in the transwell experiments were diluted 1:5, while post-infection apical compartment supernatants were diluted 1:2. Supernatants collected from the basolateral compartment at each time-point were not further diluted. Of these (diluted/undiluted) supernatants, 100µl per well were applied onto BHK-21 cells cultured in 96-well plates and serial dilutions were performed by transferring 10µl into the following well. Viral titers were quantified by microscopic read-out of the cytopathic effect (CPE) four days after supernatant application.
2.11 Data processing and statistical analysis
For comparison of insert-culture techniques, TEER values obtained at 72h and 120h after start of measurements were extracted and used for statistical analyses. A repeated measures (RM) one-way analysis of variance (ANOVA) with subsequent Tukey’s test for multiple comparison was performed on these values. For post-inoculation TEER presentation, TEER values obtained by automated cellZscope measurements of 2 inserts in 3 independent experimental replicates were synchronized by measurement timestamps after virus inoculation. Subsequently, TEER values obtained from virus-inoculated inserts were normalized to those obtained from mock-inoculated inserts by subtracting pre-inoculation differences, followed by normalization per timepoint to obtain relative TEER values. TEER values obtained at 24hpi and 48hpi were extracted and used for statistical analyses. FITC-dextran concentrations of the last taken measurements during barrier permeability assay (240 and 300 minutes after FITC-dextran application, respectively) were used for statistical analyses. Extracted TEER values and FITC-dextran concentrations obtained from virus-inoculated cultures were tested for significant difference to mock-inoculated inserts by performing a RM one-way ANOVA with subsequent Dunnett’s test. Viral titers determined in mBMEC mono-culture supernatants collected at 0hpi were considered as baseline viral titers and subsequently subtracted from respective viral titers determined in supernatants collected at the following timepoints. For statistical analyses of viral titers measured via TCID50 assay, obtained values were log10-transformed (Y=log10[Y]) and a RM two-way ANOVA was performed on the log10-transformed data. Subsequently, a Tukey’s test for multiple comparison was performed for each timepoint. For all statistical tests performed a p-value below 0.05 was considered statistically significant. All statistical analyses were performed using GraphPad Prism 10 software.
3 Results
3.1 In vitro barrier model
To investigate the effects of orthoflaviviruses on barrier properties of brain endothelial cells in vitro, a transwell model was used in which compact monolayers of primary mouse brain microvascular endothelial cells (mBMEC) were cultured on the apical side of transwell inserts. To further enhance their barrier properties primary astrocytes were co-cultured on the basolateral side of the membrane. TEER measurements and barrier integrity assays were used to assess the compactness of the established barrier. TEER measurements showed significantly higher TEER in those inserts where mBMEC were co-cultured with astrocytes compared to those where only mBMEC were cultured on the transwell membrane. Monocultures with astrocytes alone did only show negligible electrical resistance (Figure 1A). Furthermore, co-cultures were less permeable to high molecular weight FITC-dextran (70 kda) compared to monocultures. The overall FITC-dextran concentration detected in the basolateral compartment of the co-cultures even after 240 minutes remained below 1% of the FITC-dextran concentration added to the apical compartment (Figure 1). Therefore, the mBMEC-astrocyte co-culture system was used for subsequent experiments.
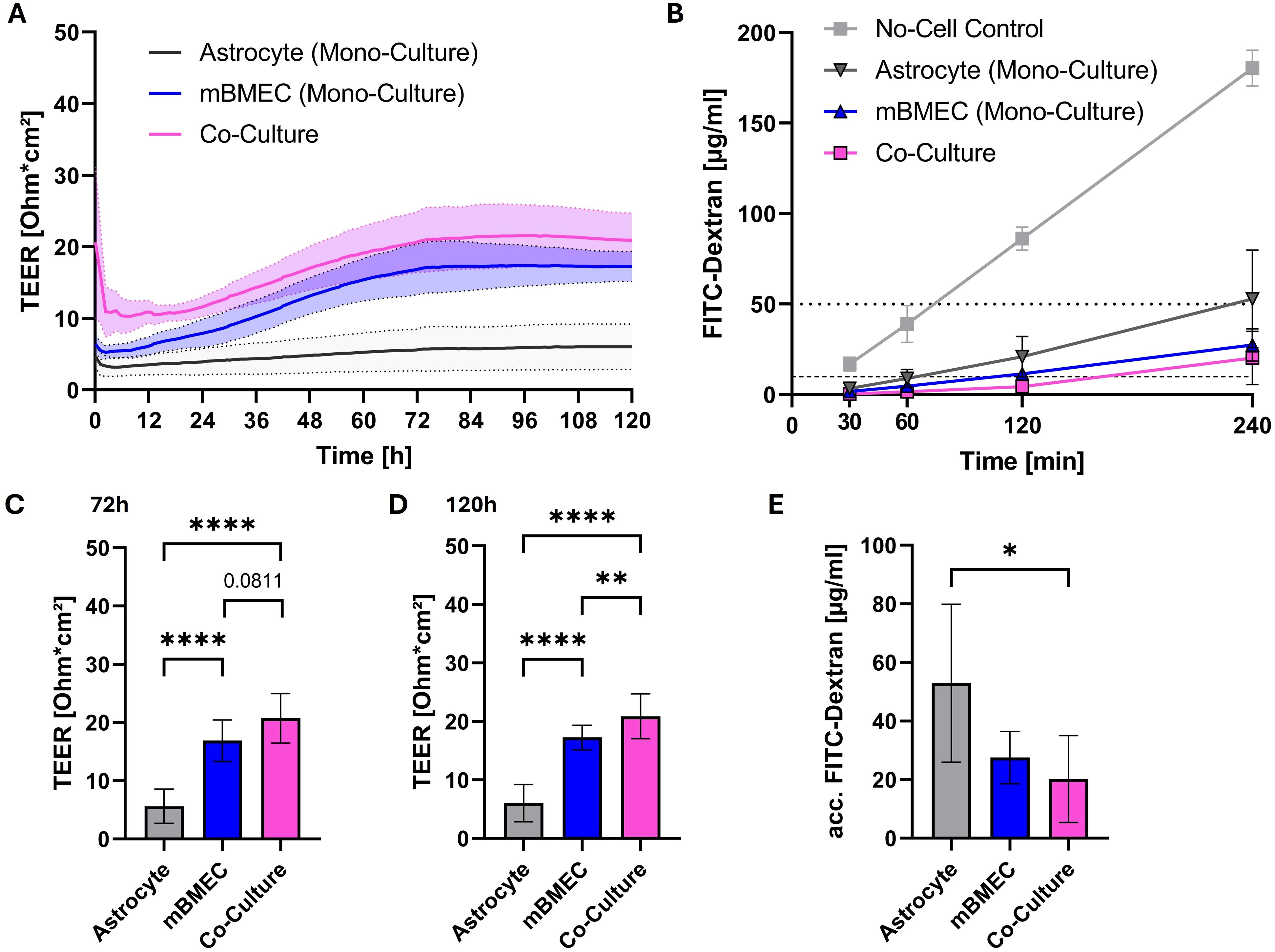
Figure 1. Transendothelial electrical resistance (TEER) monitored over time after introduction of transwell inserts to the cellZscope the day after seeding of mouse brain microvascular endothelial cells (mBMEC) and four days after astrocyte seeding. Presented are the mean values of n=6 inserts (continuous line) with the standard deviation (SD; dotted lines and filled area) (A). TEER values obtained at 72h (C) and 120h (D) of incubation in the cellZscope were statistically tested with a repeated measures one-way analysis of variance (ANOVA), followed by Tukey’s test for multiple comparison (C, D). Barrier permeability assay of inserts after 120h of incubation in the cellZscope. Presented are the values of n=3 inserts with the SD. Horizontal dotted line indicates 1% of FITC-dextran stock solution, whereas horizontal dashed line indicates 0.1% of applied FITC-dextran concentration (B). FITC-dextran concentrations determined at 240 minutes after apical application were statistically tested with a repeated measures one-way ANOVA, followed by Tukey’s test for multiple comparison (E). Asterisks indicate statistical significance as follows: p < 0.05*, p < 0.01**, p < 0.0001****.
3.2 Inoculation with orthoflaviviruses did not impact barrier properties
TEER values peaked around four days following culture of mBMEC on the apical side and astrocytes on the basolateral side of the transwell insert. At this point inserts were inoculated with WNV, TBEV, USUV, or LGTV on the apical side and the effect of the viruses on the barrier properties was assessed over a period of 48h post inoculation. Relative to mock inoculated inserts, a subtle increase in the TEER was observed initially in all inserts that were exposed to WNV, TBEV and USUV (Figure 2A). Only for LGTV inoculated inserts the increase in TEER was statistically significant compared to mock inoculated inserts (Supplementary Figure 2, p=0.0266 at 24hpi and p=0.0263 at 48hpi). Furthermore, the barrier integrity of the mBMEC-astrocyte barrier was maintained for all tested viruses as permeability of 70kDa FITC-dextran was highly restricted even at 24- and 48hpi (Figure 2B; Supplementary Figure 3). Application of staurosporine, an apoptosis-inducing agent, onto control inserts with a compact mBMEC-astrocyte barrier, in turn, led to substantial decrease of the TEER (Supplementary Figure 4). These findings suggest that the mBMEC-astrocyte barrier was intact and was not affected by infection with mosquito- (WNV & USUV) or tick-borne (TBEV & LGTV) orthoflaviviruses.
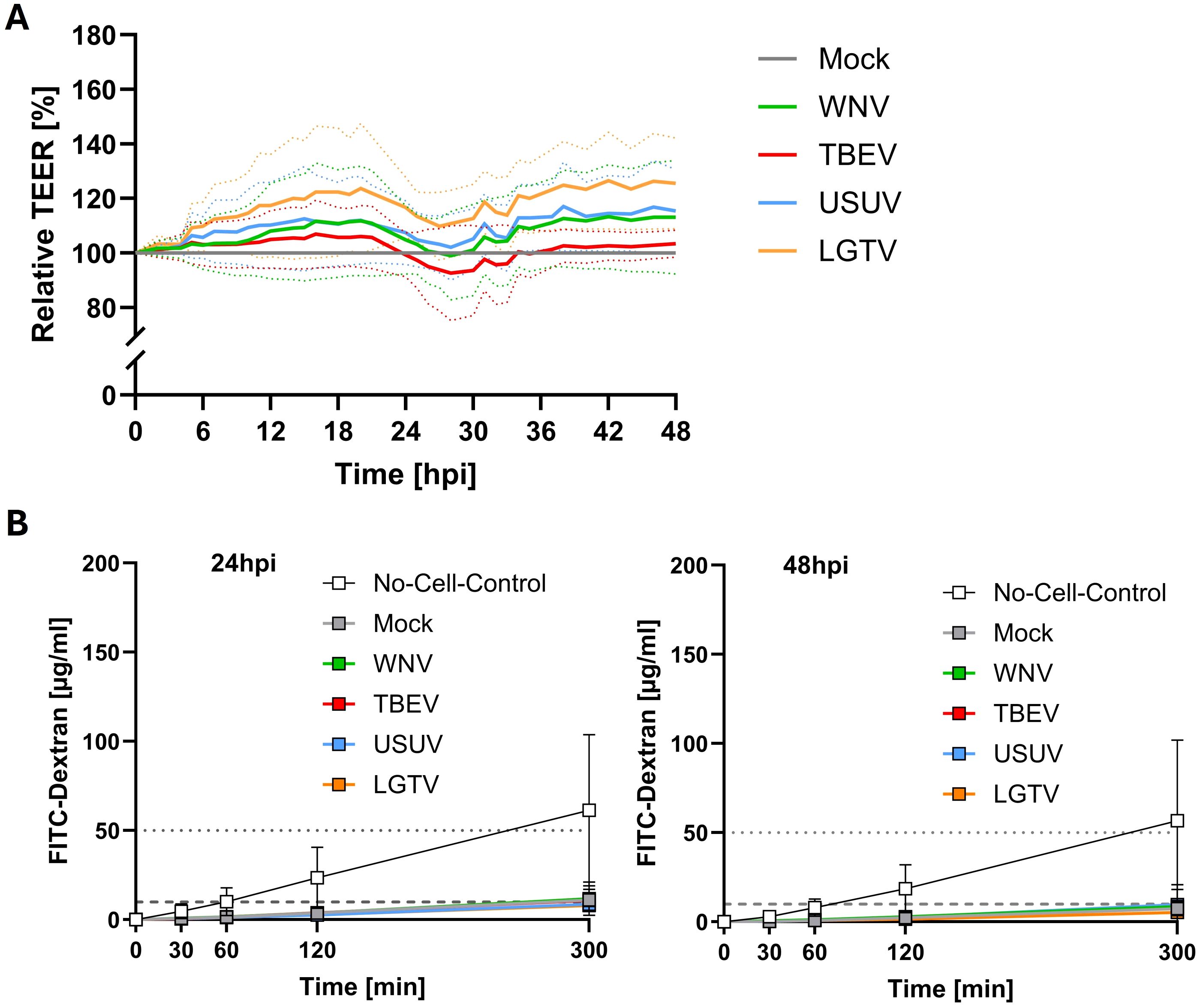
Figure 2. Relative transendothelial electrical resistance (TEER) monitored over time after inoculation of compact mBMEC-endothelial barriers on transwell inserts with West-Nile virus (WNV), Tick-borne encephalitis virus (TBEV), Usutu virus (USUV) and Langat virus (LGTV). Presented are the mean values of N=3 independent experiments (continuous line) with the standard deviation (SD; dotted lines) (A). Barrier permeability assay of inserts at 24 and 48 (B) hours post inoculation with TBEV, LGTV, WNV and USUV. Presented are the mean values of N=3 independent experiments with the standard deviation (SD). Horizontal dotted line indicates 1% of FITC-dextran stock solution, whereas horizontal dashed line indicates 0.1% of applied FITC-dextran concentration (B).
3.3 Translocation of mosquito- and tick-borne orthoflaviviruses across endothelial barrier follow different kinetics
The findings so far indicate that orthoflaviviruses do not compromise the barrier integrity of the endothelial cells. We further investigated whether these viruses were prevented from crossing the endothelial barrier or employ non-disruptive transcellular routes to reach the basolateral compartments. For this purpose, one hour after virus inoculation on the apical side, inserts were thoroughly washed to remove residual input virus. Culture supernatants were then collected from both the apical and basolateral compartments at 24 and 48hpi and the viral titers were quantified using TCID50 assays. As shown in Figure 3A, at 24hpi all viruses were detected in the culture supernatants of the apical compartment indicating viral uptake and subsequent re-release into the same compartment. By 48hpi, increased titers of WNV, TBEV and USUV were observed in the apical compartment to be significantly higher than titers of LGTV (p=0.0002), which was not detected at this timepoint (Figure 3A). No viral particles were detected in the basolateral culture supernatants at 0hpi indicating that none of the viruses crossed the endothelial barrier during the inoculation period. At 24hpi no TBEV or LGTV was detected in the basolateral compartment whereas WNV was detected in one and USUV was detected in two out of three replicates. At 48hpi relatively high titers of WNV and TBEV were detected in the basolateral compartment and no USUV and LGTV were detectable at this timepoint (Figure 3B). Overall, compared to lower-pathogenic USUV and LGTV, WNV and TBEV showed the potential to cross the endothelial astrocyte barrier in significantly higher amounts (p<0.0001) nevertheless with delayed kinetics.
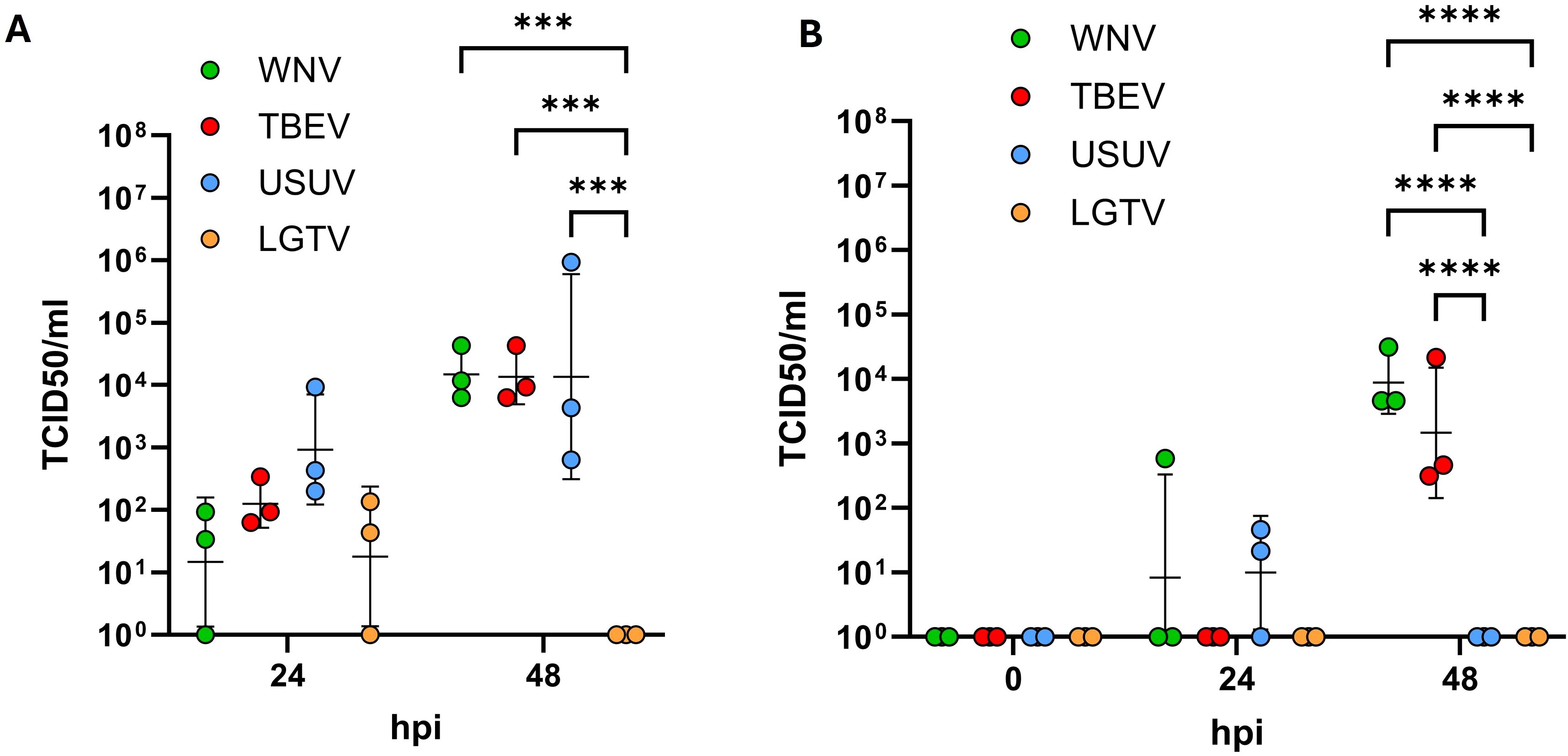
Figure 3. Quantification of viral particles in the apical (A) and basolateral (B) compartments via tissue culture infectious dose 50 (TCID50) assays. Assays were performed on supernatants collected at 24 and 48 hours post inoculation with West-Nile virus (WNV), Tick-borne encephalitis virus (TBEV), Usutu virus (USUV) and Langat virus (LGTV). Presented are the mean values of N=3 independent experiments with the standard deviation (SD). Original data was log-transformed and tested for statistical significance using a repeated measure two-way analysis of variance (ANOVA) followed by Tukey’s test for multiple comparison. Asterisks indicate statistical significance as follows: p < 0.001***, p < 0.0001****.
3.4 Orthoflaviviruses productively replicate within mBMEC mono-cultures
From the above results it is evident that WNV and TBEV demonstrated relatively superior ability to cross the endothelial-astrocyte barrier. To assess if this is due to different capacities of pathogenic versus less-pathogenic orthoflaviviruses to infect and replicate within the endothelial cells, mBMEC were inoculated with 5105 TCID50 WNV, TBEV, USUV and LGTV. Viral titers were determined from the culture supernatants collected at various time points post inoculation using TCID50 assays. The results suggest that all viruses replicate within the mBMEC albeit with different efficacies. Despite thorough washing to remove initial inoculum, small quantities of residual input virus were still detected in culture supernatants at 0hpi. Hence, for calculating the actual titers resulting from virus replication the background resulting from residual viruses in the culture supernatants was subtracted. As demonstrated in Figure 4 for all tested viruses, increase in the viral titers was observed in the culture supernatants over the period of 144hpi indicating virus replication within mBMEC. Interestingly, TBEV showed slightly higher capacity to replicate within mBMEC compared to LGTV, WNV and USUV. Taken together these results indicate that both mosquito- and tick-borne orthoflaviviruses can infect and replicate within the endothelial cells, but also that the ability to cross the endothelial cell is not dependent on virus replication.
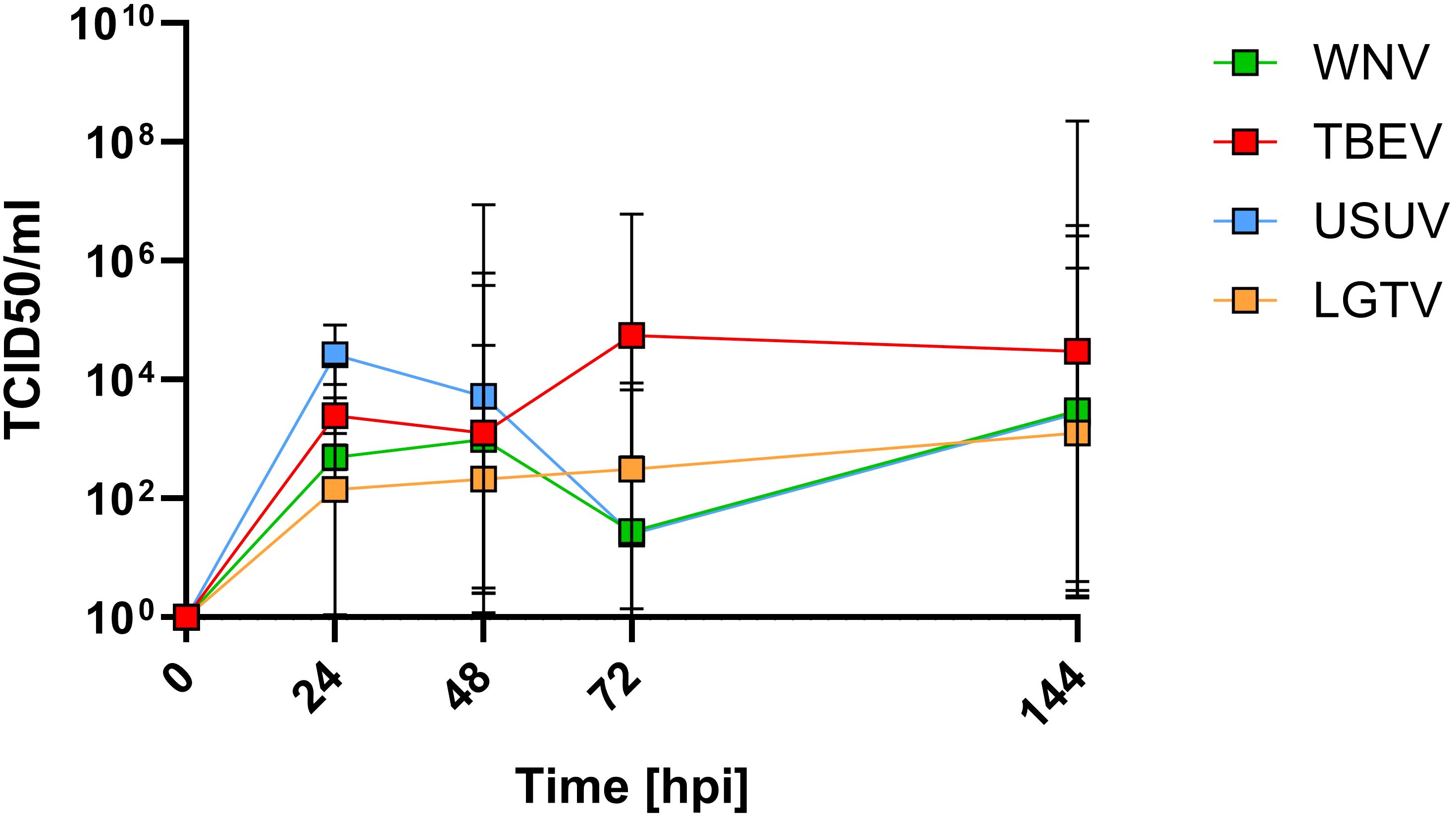
Figure 4. Quantification of infectious virus in the supernatants of mBMEC monocultures inoculated with TBEV, LGTV, WNV and USUV. TCID50 assays were performed on supernatants collected at different timepoints post inoculation. Presented are the mean values of N=3 independent experiments with the standard deviation (SD).
4 Discussion
In this study we compared the ability of neurotropic orthoflaviviruses with varying degrees of pathogenicity in humans and other mammalian species to cross a compact endothelial barrier and demonstrate that both mosquito- and tick-borne orthoflaviviruses cross a primary mouse endothelial cell barrier without compromising its integrity. The CNS is protected by compact and highly impermeable anatomical barriers such as the BBB, and as an initial step invading pathogens have to cross this barrier. Certain viruses such as ZIKV and Nipah virus (NiV) have been shown to infect and destroy the endothelial cells leading to vasculitis (Papa et al., 2017). Other viruses can cause BBB breakdown by triggering strong inflammation or endothelial cell death (German et al., 2006; Mishra et al., 2009; de Vries and Harding, 2023). In accordance with few previous reports we also demonstrate that WNV and TBEV cross the endothelial barrier and this occurs without compromising the endothelial barrier (Verma et al., 2009, 2010; Palus et al., 2017; Marshall et al., 2024). These findings are consistent with our recent findings which demonstrated that both TBEV and LGTV are capable of crossing human brain endothelial cell barrier without affecting its integrity (Schweitzer et al., 2025).
The BBB is a complex structure comprised of several other cell types such as astrocytes and pericytes in addition to tightly connected endothelial cells that essentially form a compact barrier. This implies that astrocyte and pericytes that are in close association with endothelial cells may be equally important in maintaining the integrity of the BBB (Pivoriūnas and Verkhratsky, 2021). In our preliminary analysis aimed at establishing a reliable barrier model, we have observed that transwell inserts with endothelial cell-astrocyte co-cultures provided relatively more compact barriers as demonstrated by improved TEER and decreased permeability compared to the inserts with endothelial cell mono-cultures. Several studies have shown that invading pathogens can modulate the properties of astrocytes and pericytes, which subsequently may contribute to the breakdown of the barrier. The best studied example among orthoflaviviruses is JEV which has been shown to infect both astrocytes and pericytes and induce proinflammatory factors such as IL-6 and matrix metalloproteases 2 and 9 which ultimately led to breakdown of endothelial barrier (Chen et al., 2014; Chang et al., 2015). Considering this, we employed an in vitro barrier model consisting of primary mouse brain endothelial cells and astrocytes to compare neuroinvasion of highly pathogenic WNV and TBEV and relatively non-pathogenic USUV and LGTV. For the viruses tested here, we did not see any notable changes in barrier properties despite of the presence of astrocytes. Similarly, in a recent study Marshall et al. used an in vitro BBB model consisting of human brain endothelial cells, astrocytes and pericytes to investigate neuroinvasion of WNV and USUV and demonstrated the ability of these viruses to cross the human brain endothelial cell barrier at different magnitudes without disrupting it (Marshall et al., 2024). TBEV strains Neudoerfl and Hypr with different pathogenic potentials were also shown to cross a human brain endothelial barrier model without altering its barrier properties (Palus et al., 2017). Interestingly, pathogenic viruses WNV and TBEV crossed the endothelial-astrocyte barrier despite an intact barrier whereas their non- or low-pathogenic counterparts for humans, USUV and LGTV respectively, were less efficient in overcoming the endothelial barrier and finding their way into basolateral compartment. The inability of 70 kDa FITC-dextran molecules to pass through endothelial barrier before and after infection supports the hypothesis that the viruses use transcellular routes to reach basolateral compartments rather than leaking through paracellular spaces. In this direction, using an in vitro human endothelial barrier model consisting of human cerebral microvascular endothelial cells (hCMEC/D3), we have demonstrated that TBEV and LGTV inoculated onto the apical side of the endothelial barrier transmigrate through hCMEC/D3 cells and are released on the basolateral side (Schweitzer et al., 2025). This process occurs without virus replication in the hCMEC/D3 cells. In contrast, in the current study we have observed that all viruses tested, WNV, TBEV, USUV and LGTV are capable of replicating within the mouse endothelial cells albeit with different efficacies. Though the ability to replicate within the endothelial cells was not significantly different between pathogenic and low- or non-pathogenic viruses the latter were inefficient in crossing the endothelial barrier. This implies that neuroinvasion most likely is not defined by the ability of viruses to replicate within the endothelial cells. Our observations are in line with the findings made by Marshall et al. where different efficiencies of WNV and USUV in crossing the endothelial cell barrier were attributed to their pathogenicity (Marshall et al., 2024). Immunofluorescence staining with antibodies directed against double stranded RNA (dsRNA) further supports the claim of efficient replication of pathogenic viruses within endothelial cells. We observed relatively higher dsRNA signal over time for TBEV and WNV, than the relatively low dsRNA signal for LGTV. Interestingly, dsRNA signals obtained for USUV are similar to those obtained for WNV (Supplementary Figure 5).
To our knowledge, this is the first study directly comparing the neuroinvasive properties of mosquito- and tick-borne orthoflaviviruses. Additionally, highly pathogenic viruses and closely related less pathogenic strains were used for comparison. Structurally both groups of orthoflaviviruses are similar, however, they cluster in distant genetic lineages (Neufeldt et al., 2018) and exhibit variations in viral proteins. For example, non-structural protein (NS)1 of mosquito-borne orthoflaviviruses such as DENV, JEV, WNV and ZIKV was shown to directly affect the integrity of human endothelial cell barriers in vitro (Puerta-Guardo et al., 2019). Although NS1 is conserved among orthoflaviviruses, its glycosylation sites differ between mosquito- and tick-borne orthoflaviviruses. These differences have been linked to complement interactions, endothelial glycocalyx layer disruption and neurovirulence (da Fonseca et al., 2017; Perera et al., 2023). Furthermore, NS1 of orthoflaviviruses is associated with vascular leakage (Puerta-Guardo et al., 2019). For this reason, we expected differences in the abilities of mosquito- and tick-borne orthoflaviruses to impact barrier properties. However, this proved not to be the case. A major limitation of in vitro endothelial barrier models is the low TEER values achieved, which does not accurately reflect the compactness of BBB observed in vivo. This is because BBB integrity in vivo is governed by several molecular and cellular factors and the extracellular matrix. Therefore, it is essential that such models are validated with appropriate methods, such as assessing the permeability of dextran molecules, to rule out the possibility of leakage through paracellular spaces. Nevertheless, well characterized and validated in vitro BBB models are useful tools to study direct effects of invading pathogens on the BBB.
Overall, the data obtained from this study suggests that initial neuroinvasive mechanisms employed by not only mosquito- but also tick-borne orthoflaviviruses are non-destructive and the viruses can enter the CNS without compromising the BBB. This has been demonstrated in vivo using a mouse model of TBEV infection, in which virus was detected in the brain before the BBB was compromised (Ruzek et al., 2011). Similarly, an increased permeability of the BBB is not necessary for a fatal outcome of WNV infection in mice (Morrey et al., 2008). In the absence of early BBB disruption during the initial neuroinvasion by pathogenic orthoflaviviruses, virus replication within target cells, innate immune sensing pathways and ensuing neuroinflammation are likely contributors to their neuropathogenesis. Finally, the endothelial cell-astrocyte co-culture model may provide an ideal platform to test and compare the neuroinvasive mechanisms of not only mosquito- and tick-borne orthoflaviviruses but also neurotropic viruses belonging to other families.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material. Further inquiries can be directed to the corresponding author.
Ethics statement
The animal study was approved by Institutional Review Board of University of Veterinary Medicine Hannover (approval no. TiHo-T-2023-12, TiHo-T-2024-14). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
FS: Formal analysis, Methodology, Investigation, Writing – original draft, Writing – review & editing, Visualization. L-JS: Writing – review & editing, Methodology, Investigation. AF: Investigation, Writing – review & editing, Methodology. VG: Methodology, Investigation, Writing – review & editing. TS: Writing – review & editing, Investigation, Methodology, Resources, Conceptualization, Supervision. IS: Resources, Supervision, Writing – review & editing, Conceptualization, Methodology. MP: Funding acquisition, Resources, Supervision, Methodology, Writing – review & editing, Conceptualization. DR: Methodology, Conceptualization, Supervision, Funding acquisition, Resources, Writing – review & editing. AO: Resources, Writing – review & editing, Supervision, Funding acquisition, Conceptualization. CP: Investigation, Writing – review & editing, Supervision, Conceptualization, Resources, Formal analysis, Data curation, Visualization, Methodology.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was funded by grants obtained from the EU Innovative Medicines Initiative (IMI) under Horizon2020 program for IM2PACT (Grant ID 807015). Further funding was provided by the Deutsche Forschungsgemeinschaft (DFG, German Research Foundation)—398066876/GRK 2485/2, Research Training Group VIPER. MP and DR were supported by the Czech Science Foundation (grant No. 23-08039S). We acknowledge financial support by the Open Access Publication Fund of the University of Veterinary Medicine Hannover, Foundation.
Acknowledgments
We acknowledge Kristin Laudeley for providing excellent technical support.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fcimb.2025.1624636/full#supplementary-material
Glossary
ANOVA: Analysis of variance
ara-C: Cytosine β-D-arabinofuranoside
BBB: Blood-brain barrier
BHK-21: Baby hamster kidney cells
BSA: Bovine serum albumin
CECM: Complete mouse endothelial cell medium
CNS: Central nervous system
CPE: Cytopathic effect
DMEM: Dulbecco’s Modified Eagle Medium
DMSO: Dimethyl sulfoxide
dsRNA: Double-stranded ribonucleic acid
FBS: Fetal bovine serum
FITC: Fluorescein isothiocyanate
GCS: Gelatin-based coating solution
hCMEC/D3: Human cerebral microvascular endothelial cells
hpi: Hours post inoculation
IL-6: Interleukin 6
JEV: Japanese encephalitis virus
LGTV: Langat virus
mBMEC: Mouse brain microvascular endothelial cells
MEM+H: Minimal essential medium supplemented with HEPES buffer
MGP+: Mixed glia culture preparation medium
MMP: Matrix metalloproteases
NiV: Nipah virus
NS1: Non-structural protein 1 (of orthoflaviviruses)
PLL: Poly-L-lysine
RM: Repeated measures
TBEV: Tick-borne encephalitis virus
TCID50: 50% Tissue culture infectious dose
TEER: Transendothelial electrical resistance
USUV: Usutu virus
WNV: West-Nile virus
ZIKV: Zika virus
ZO-1: Zonula occludens protein 1.
References
Agliani, G., Giglia, G., Marshall, E. M., Gröne, A., Rockx, B. H. G., and van den Brand, J. M. A. (2023). Pathological features of West Nile and Usutu virus natural infections in wild and domestic animals and in humans: A comparative review. One Health 16, 100525. doi: 10.1016/j.onehlt.2023.100525
Cain, M. D., Salimi, H., Diamond, M. S., and Klein, R. S. (2019). Mechanisms of pathogen invasion into the central nervous system. Neuron 103, 771–783. doi: 10.1016/j.neuron.2019.07.015
Chang, C.-Y., Li, J.-R., Chen, W.-Y., Ou, Y.-C., Lai, C.-Y., Hu, Y.-H., et al. (2015). Disruption of in vitro endothelial barrier integrity by Japanese encephalitis virus-Infected astrocytes. Glia 63, 1915–1932. doi: 10.1002/glia.22857
Chen, C.-J., Ou, Y.-C., Li, J.-R., Chang, C.-Y., Pan, H.-C., Lai, C.-Y., et al. (2014). Infection of pericytes in vitro by Japanese encephalitis virus disrupts the integrity of the endothelial barrier. J. Virol. 88, 1150–1161. doi: 10.1128/jvi.02738-13
da Fonseca, N. J., Lima Afonso, M. Q., Pedersolli, N. G., de Oliveira, L. C., Andrade, D. S., and Bleicher, L. (2017). Sequence, structure and function relationships in flaviviruses as assessed by evolutive aspects of its conserved non-structural protein domains. Biochem. Biophys. Res. Commun. 492, 565–571. doi: 10.1016/j.bbrc.2017.01.041
de Vries, L. and Harding, A. T. (2023). Mechanisms of neuroinvasion and neuropathogenesis by pathologic flaviviruses. Viruses 15, 261. doi: 10.3390/v15020261
Dunton, A. D., Göpel, T., Ho, D. H., and Burggren, W. (2021). Form and function of the vertebrate and invertebrate blood-brain barriers. Int. J. Mol. Sci. 22, 12111. doi: 10.3390/ijms222212111
German, A. C., Myint, K. S. A., Mai, N. T. H., Pomeroy, I., Phu, N. H., Tzartos, J., et al. (2006). A preliminary neuropathological study of Japanese encephalitis in humans and a mouse model. Trans. R. Soc. Trop. Med. Hygiene 100, 1135–1145. doi: 10.1016/j.trstmh.2006.02.008
Gritsun, T. S., Lashkevich, V. A., and Gould, E. A. (2003). Tick-borne encephalitis. Antiviral Res. 57, 129–146. doi: 10.1016/S0166-3542(02)00206-1
Habarugira, G., Suen, W. W., Hobson-Peters, J., Hall, R. A., and Bielefeldt-Ohmann, H. (2020). West nile virus: an update on pathobiology, epidemiology, diagnostics, control and “One health” Implications. Pathogens 9, 589. doi: 10.3390/pathogens9070589
Heinz, F. X. and Stiasny, K. (2012). Flaviviruses and their antigenic structure. J. Clin. Virol. 55, 289–295. doi: 10.1016/j.jcv.2012.08.024
Iacono-Connors, L. C. and Schmaljohn, C. S. (1992). Cloning and sequence analysis of the genes encoding the nonstructural proteins of langat virus and comparative analysis with other flaviviruses. Virology 188, 875–880. doi: 10.1016/0042-6822(92)90545-Z
Kadry, H., Noorani, B., and Cucullo, L. (2020). A blood–brain barrier overview on structure, function, impairment, and biomarkers of integrity. Fluids Barriers CNS 17, 69. doi: 10.1186/s12987-020-00230-3
Kubinski, M., Beicht, J., Zdora, I., Saletti, G., Kircher, M., Petry-Gusmag, M., et al. (2023). Cross-reactive antibodies against Langat virus protect mice from lethal tick-borne encephalitis virus infection. Front. Immunol. 14. doi: 10.3389/fimmu.2023.1134371
Kuno, G., Chang, G.-J. J., Tsuchiya, K. R., Karabatsos, N., and Cropp, C. B. (1998). Phylogeny of the genus flavivirus. J. Virol. 72, 73–83. doi: 10.1128/JVI.72.1.73-83.1998
Lim, S. M., Koraka, P., Osterhaus, A. D. M. E., and Martina, B. E. E. (2011). West nile virus: immunity and pathogenesis. Viruses 3, 811–828. doi: 10.3390/v3060811
Mandl, C. W. (2005). Steps of the tick-borne encephalitis virus replication cycle that affect neuropathogenesis. Virus Res. 111, 161–174. doi: 10.1016/j.virusres.2005.04.007
Mandl, C. W., Iacono-Connors, L., Wallner, G., Holzmann, H., Kunz, C., and Heinz, F. X. (1991). Sequence of the genes encoding the structural proteins of the low-virulence tick-borne flaviviruses Langat TP21 and Yelantsev. Virology 185, 891–895. doi: 10.1016/0042-6822(91)90567-U
Marshall, E. M., Koopmans, M., and Rockx, B. (2024). Usutu virus and West Nile virus use a transcellular route of neuroinvasion across an in vitro model of the human blood–brain barrier. NPJ Viruses 2, 1–9. doi: 10.1038/s44298-024-00034-4
Mishra, M. K., Dutta, K., Saheb, S. K., and Basu, A. (2009). Understanding the molecular mechanism of blood–brain barrier damage in an experimental model of Japanese encephalitis: Correlation with minocycline administration as a therapeutic agent. Neurochemistry Int. 55, 717–723. doi: 10.1016/j.neuint.2009.07.006
Mora, C., McKenzie, T., Gaw, I. M., Dean, J. M., von Hammerstein, H., Knudson, T. A., et al. (2022). Over half of known human pathogenic diseases can be aggravated by climate change. Nat. Clim. Change 12, 869–875. doi: 10.1038/s41558-022-01426-1
Morrey, J. D., Olsen, A. L., Siddharthan, V., Motter, N. E., Wang, H., Taro, B. S., et al. (2008). Increased blood–brain barrier permeability is not a primary determinant for lethality of West Nile virus infection in rodents. J. Gen. Virol. 89, 467–473. doi: 10.1099/vir.0.83345-0
Neufeldt, C. J., Cortese, M., Acosta, E. G., and Bartenschlager, R. (2018). Rewiring cellular networks by members of the Flaviviridae family. Nat. Rev. Microbiol. 16, 125–142. doi: 10.1038/nrmicro.2017.170
Palus, M., Vancova, M., Sirmarova, J., Elsterova, J., Perner, J., and Ruzek, D. (2017). Tick-borne encephalitis virus infects human brain microvascular endothelial cells without compromising blood-brain barrier integrity. Virology 507, 110–122. doi: 10.1016/j.virol.2017.04.012
Papa, M. P., Meuren, L. M., Coelho, S. V. A., Lucas, C. G., de, O., Mustafá, Y. M., et al. (2017). Zika virus infects, activates, and crosses brain microvascular endothelial cells, without barrier disruption. Front. Microbiol. 8. doi: 10.3389/fmicb.2017.02557
Perera, D. R., Ranadeva, N. D., Sirisena, K., and Wijesinghe, K. J. (2023). Roles of NS1 protein in flavivirus pathogenesis. ACS Infect. Diseases. 10 (1), 20–56. doi: 10.1021/acsinfecdis.3c00566
Petry, M., Palus, M., Leitzen, E., Mitterreiter, J. G., Huang, B., Kröger, A., et al. (2021). Immunity to TBEV Related Flaviviruses with Reduced Pathogenicity Protects Mice from Disease but Not from TBEV Entry into the CNS. Vaccines 9, 196. doi: 10.3390/vaccines9030196
Pivoriūnas, A. and Verkhratsky, A. (2021). Astrocyte–endotheliocyte axis in the regulation of the blood–brain barrier. Neurochem. Res. 46, 2538–2550. doi: 10.1007/s11064-021-03338-6
Postler, T. S., Beer, M., Blitvich, B. J., Bukh, J., de Lamballerie, X., Drexler, J. F., et al. (2023). Renaming of the genus Flavivirus to Orthoflavivirus and extension of binomial species names within the family Flaviviridae. Arch. Virol. 168, 224. doi: 10.1007/s00705-023-05835-1
Pradier, S., Lecollinet, S., and Leblond, A. (2012). West Nile virus epidemiology and factors triggering change in its distribution in Europe. Rev. Sci. Tech 31, 829–844. doi: 10.20506/rst.31.3.2167
Prajeeth, C. K., Dittrich-Breiholz, O., Talbot, S. R., Robert, P. A., Huehn, J., and Stangel, M. (2018). IFN-γ Producing th1 cells induce different transcriptional profiles in microglia and astrocytes. Front. Cell. Neurosci. 12. doi: 10.3389/fncel.2018.00352
Puerta-Guardo, H., Glasner, D. R., Espinosa, D. A., Biering, S. B., Patana, M., Ratnasiri, K., et al. (2019). Flavivirus NS1 triggers tissue-specific vascular endothelial dysfunction reflecting disease tropism. Cell Rep. 26, 1598–1613.e8. doi: 10.1016/j.celrep.2019.01.036
Rathore, A. P. S. and St. John, A. L. (2020). Cross-reactive immunity among flaviviruses. Front. Immunol. 11. doi: 10.3389/fimmu.2020.00334
Reed, L. J. and Muench, H. (1938). A simple method of estimating fifty per cent endpoints. Am. J. Epidemiol. 27, 493–497. doi: 10.1093/oxfordjournals.aje.a118408
Rochfort, K. D., Collins, L. E., Murphy, R. P., and Cummins, P. M. (2014). Downregulation of blood-brain barrier phenotype by proinflammatory cytokines involves NADPH oxidase-dependent ROS generation: consequences for interendothelial adherens and tight junctions. PloS One 9, e101815. doi: 10.1371/journal.pone.0101815
Ruscher, C., Patzina-Mehling, C., Melchert, J., Graff, S. L., McFarland, S. E., Hieke, C., et al. (2023). Ecological and clinical evidence of the establishment of West Nile virus in a large urban area in Europe, Berlin, Germany 2021 to 2022. Eurosurveillance 28, 2300258. doi: 10.2807/1560-7917.ES.2023.28.48.2300258
Ruzek, D., Avšič Županc, T., Borde, J., Chrdle, A., Eyer, L., Karganova, G., et al. (2019). Tick-borne encephalitis in Europe and Russia: Review of pathogenesis, clinical features, therapy, and vaccines. Antiviral Res. 164, 23–51. doi: 10.1016/j.antiviral.2019.01.014
Ruzek, D., Salát, J., Singh, S. K., and Kopecký, J. (2011). Breakdown of the blood-brain barrier during tick-borne encephalitis in mice is not dependent on CD8+ T-cells. PloS One 6, e20472. doi: 10.1371/journal.pone.0020472
Schweitzer, F., Letoha, T., Osterhaus, A., and Prajeeth, C. K. (2025). Impact of tick-borne orthoflaviviruses infection on compact human brain endothelial barrier. Int. J. Mol. Sci. 26, 2342. doi: 10.3390/ijms26052342
Smorodintsev, A. A. and Dubov, A. V. (1986). The correlation between the pathogenesis and biological characteristics of tick-borne encephalitis virus. Tick-borne encephalitis its vaccino-prophylaxis Medicine Moscow 113–124, 190–211.
Störk, T., de le Roi, M., Haverkamp, A.-K., Jesse, S. T., Peters, M., Fast, C., et al. (2021). Analysis of avian Usutu virus infections in Germany from 2011 to 2018 with focus on dsRNA detection to demonstrate viral infections. Sci. Rep. 11, 24191. doi: 10.1038/s41598-021-03638-5
Verma, S., Kumar, M., Gurjav, U., Lum, S., and Nerurkar, V. R. (2010). Reversal of West Nile virus-induced blood–brain barrier disruption and tight junction proteins degradation by matrix metalloproteinases inhibitor. Virology 397, 130–138. doi: 10.1016/j.virol.2009.10.036
Verma, S., Lo, Y., Chapagain, M., Lum, S., Kumar, M., Gurjav, U., et al. (2009). West Nile virus infection modulates human brain microvascular endothelial cells tight junction proteins and cell adhesion molecules: Transmigration across the in vitro blood-brain barrier. Virology 385, 425–433. doi: 10.1016/j.virol.2008.11.047
Keywords: orthoflavivirus, blood-brain barrier, endothelial cells, astrocytes, neuroinvasion, transendothelial electrical resistance
Citation: Schweitzer F, Schröder L-J, Friedrichs A, Gudi V, Skripuletz T, Steffen I, Palus M, Růžek D, Osterhaus A and Prajeeth CK (2025) Mosquito- and tick-borne orthoflaviviruses cross an in vitro endothelial-astrocyte barrier. Front. Cell. Infect. Microbiol. 15:1624636. doi: 10.3389/fcimb.2025.1624636
Received: 07 May 2025; Accepted: 18 June 2025;
Published: 02 July 2025.
Edited by:
Karam Chand, Indian Veterinary Research Institute (ICAR-IVRI), Mukteswar Campus, IndiaReviewed by:
Naveen Kumar, ICAR-National Institute of High Security Animal Diseases (ICAR-NIHSAD), IndiaBasavaraj S. Mathapati, Indian Council of Medical Research (ICMR), India
Copyright © 2025 Schweitzer, Schröder, Friedrichs, Gudi, Skripuletz, Steffen, Palus, Růžek, Osterhaus and Prajeeth. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Chittappen Kandiyil Prajeeth, cHJhamVldGguY2hpdHRhcHBlbi5rYW5kaXlpbEB0aWhvLWhhbm5vdmVyLmRl
†Present address: Imke Steffen, Department of Microbiology, Immunology, and Pathology, Center for Vector-Borne Infectious Diseases, Colorado State University, Fort Collins, CO, United States