 Paola Chiani1*
Paola Chiani1* Valeria Michelacci1
Valeria Michelacci1 Elisabetta Delibato1
Elisabetta Delibato1 Eleonora Ventola1Samra Mannan1
Eleonora Ventola1Samra Mannan1 Manuela Marra2
Manuela Marra2 Valentina Libri2
Valentina Libri2 Rubén L. Rodríguez-Expósito3,4,5Maria Reyes-Batlle3,4,5Isabel de Fuentes6
Rubén L. Rodríguez-Expósito3,4,5Maria Reyes-Batlle3,4,5Isabel de Fuentes6 Jacob Lorenzo-Morales3,4,5
Jacob Lorenzo-Morales3,4,5 Stefano Morabito1
Stefano Morabito1 Margherita Montalbano Di Filippo1
Margherita Montalbano Di Filippo1- 1Department of Food Safety, Nutrition and Veterinary Public Health, Istituto Superiore di Sanità, Rome, Italy
- 2Core Facilities Technical-Scientific Service, Istituto Superiore di Sanità, Rome, Italy
- 3Instituto Universitario de Enfermedades Tropicales y Salud Pública de Canarias (IUETSPC), Universidad de La Laguna (ULL), San Cristóbal de La Laguna, Spain
- 4CIBER de Enfermedades Infecciosas (CIBERINFEC), Instituto de Salud Carlos III, Madrid, Spain
- 5Departamento de Obstetricia y Ginecología, Pediatría, Medicina Preventiva y Salud Pública, Toxicología, Medicina Legal y Forense y Parasitología, Universidad de La Laguna (ULL), San Cristóbal de La Laguna, Spain
- 6Servicio de Parasitología, Centro Nacional de Microbiología, Instituto de Ciencias de la Salud Carlos III, Madrid, Spain
Introduction: Free-living amoebae (FLA) are widespread protozoa that can host bacterial pathogens, promoting their persistence in the environment. Yersinia enterocolitica, a foodborne zoonotic pathogen, has been detected within amoebae, but its intracellular dynamics remain unclear.
Methods: In this study, we explored the interaction between three Y. enterocolitica strains—differing in biotype and virulence gene profile—and two Acanthamoeba spp.—a reference strain and a wild environmental isolate.
Results and discussion: All strains were internalized and survived up to 8 days in the collection strain and 16 days in the wild isolate. Intracellular persistence did not affect amoebal integrity or bacterial virulence profiles. Whole genome sequencing (WGS) revealed high genomic stability across strains, though specific mutations—such as in the igaA gene, involved in stress response—emerged after persistence in the collection strain. These findings suggest that Acanthamoeba spp. not only shields Y. enterocolitica from environmental stress but may also influence its genome and adaptive potential. This work expands the current understanding of Y. enterocolitica biology and highlights the role of FLA as reservoirs and potential drivers of bacterial evolution. Their contribution to the bacteria persistence and gene exchange warrants further investigation, particularly in the context of antimicrobial resistance and food safety.
Introduction
Free-living amoebae (FLA) are widespread protozoa found in a wide range of natural and human-made environments, where they prey on bacteria, fungi, and algae. Among pathogenic genera, Acanthamoeba and Vermamoeba are linked to ocular infections in contact lens users (Lorenzo-Morales et al., 2015; Moran et al., 2022), Naegleria fowleri causes primary amoebic meningoencephalitis (PAM) in immunocompetent individuals (Visvesvara et al., 2007), while Balamuthia and Acanthamoeba are responsible for granulomatous amoebic encephalitis (GAE) in immunocompromised patients (Kiderlen et al., 2014). FLA present two main life stages: the trophozoite, an active feeding form that consumes bacteria, viruses, algae, and yeast, and the cyst, the resistant structure that allows survival under adverse environmental conditions (i.e., nutrient scarcity, extreme temperatures, pH changes, gamma radiation, and water treatments). These amoebae not only are efficient bacterial predators but also act as crucial environmental reservoirs for pathogenic microorganisms. Some bacterial species evade predation by internalizing within amoebal trophozoites, forming endosymbiotic relationships that shield them from environmental stress (Henriquez et al., 2021; Dinda et al., 2024). Inside the vacuolar environment of the amoebae, these bacteria resist degradation, can replicate, and enhance their survival. This mechanism promotes both persistence and dissemination, raising the risk of human exposure. Additionally, amoebal endosymbiosis could drive horizontal gene transfer, contributing to antibiotic resistance and adaptation to hostile conditions (Greub and Raoult, 2004; Henriquez et al., 2021; Montalbano Di Filippo et al., 2022; Dinda et al., 2024).
Notably, several foodborne zoonotic agents, including Salmonella enterica, Listeria monocytogenes, and Shiga toxin-producing Escherichia coli (STEC), have been detected within amoebal hosts, where endosymbiosis enhances their resistance to disinfection and environmental stress (Vaerewijck et al., 2014; Chavatte et al., 2014; Scheid, 2018; Montalbano Di Filippo et al., 2022; Dinda et al., 2024). This relationship underscores the role of FLA as reservoirs that support the persistence and transmission of zoonotic pathogens in food processing and water distribution systems. Among foodborne pathogens, Y. enterocolitica represents a significant threat due to its zoonotic potential and ability to persist in various environments.
Y. enterocolitica is a foodborne zoonotic pathogen responsible for yersiniosis, the fourth most-reported foodborne zoonosis in Europe in 2023, with 8,738 cases (EFSA ECDC, 2024). It belongs to the Yersiniaceae family (28 species https://lpsn.dsmz.de/genus/yersinia—Adeolu et al., 2016), which also includes the pathogenic species Y. pestis and Y. pseudotuberculosis (Adeolu et al., 2016). The main route of human infection is through the consumption of contaminated food, particularly undercooked pork, but Y. enterocolitica has also been detected in milk, dairy products, fish, molluscs, vegetables, fruits, and water (Mancini et al., 2022; Mancusi et al., 2024). Although infections are typically self-limiting with gastrointestinal symptoms, severe complications like mesenteric lymphadenitis and terminal ileitis may occur, especially in children, the elderly, and immunocompromised individuals (Shoaib et al., 2019; Todd, 2014).
Y. enterocolitica displays considerable genetic and phenotypic diversity, classified into six biotypes (BT)—1A, 1B, 2, 3, 4, and 5—and approximately 70 serotypes (Fredriksson-Ahomaa, 2017). Pathogenicity varies significantly among biotypes: BT1A is generally non-pathogenic, BT1B is highly virulent, while BT2 to BT5 exhibit moderate pathogenic potential (Reuter et al., 2014). Pathogenic strains harbor the 70-kb virulence plasmid (pYV), which encodes essential virulence factors such as yadA (adhesin A) and virF (transcriptional regulator), alongside chromosomal genes invA (invasin), ail (attachment and invasion locus), ystA (Yersinia stable toxin A), and myfA (mucoid Yersinia factor A), contributing to its pathogenicity (Rivas et al., 2021). BT1A strains are generally considered non-pathogenic due to the absence of the pYV plasmid and key chromosomal virulence genes like ail. However, they may carry alternative virulence factors such as ystB (Yersinia stable toxin B) (Bhagat and Virdi, 2007).
Despite its relevance in food safety, interactions between Y. enterocolitica and FLA remain largely unexplored (Lambrecht et al., 2013; 2017). Understanding how Y. enterocolitica exploits FLA as environmental reservoirs could reveal mechanisms that enhance its resistance to disinfection and its persistence in adverse conditions. Filling this knowledge gap may lead to improved food safety protocols and reduced zoonotic transmission risks.
In this study, we applied a combined approach to explore the interaction between Y. enterocolitica and a reference and a wild strain of Acanthamoeba spp., used as a model organism. Specifically, we (i) evaluated the internalization capacity of Y. enterocolitica strains with distinct virulence gene profiles; (ii) determined whether the bacterium remains dormant or actively proliferates within the protozoan; and (iii) assessed the integrity and genomic stability of internalized Y. enterocolitica strains.
Materials and methods
Amoebal strains and culture conditions
For the invasion assays, two distinct Acanthamoeba strains were employed: (i) a reference strain (Acanthamoeba sp., CDC: V036, NR-46463) provided by BEI Resources, NIAID, NIH, and (ii) a wild-type isolate of Acanthamoeba spp. collected from a natural water source in Torrijos, Spain, kindly provided by our colleagues (co-authors of this work). The wild-type strain was cultured at 37°C and initially identified at the genus level by light microscopy. Molecular characterization was subsequently performed through conventional polymerase chain reaction (PCR) targeting the 18S rRNA gene, confirming its assignment as Acanthamoeba sp., genotype T4 (Schroeder et al., 2001).
Both strains were cultured axenically as monolayers in 25-cm2 tissue culture flasks using proteose peptone–yeast–glucose (PYG) medium, composed of 0.75% (w/v) proteose peptone, 0.75% (w/v) yeast extract, and 1.5% (w/v) glucose (Montalbano Di Filippo et al., 2022).
Cultures were maintained at 37°C under static conditions. Adherent amoebae were detached by vigorous tapping of the culture flasks, followed by centrifugation at 300 × g for 5 min. Cells were then washed with phosphate-buffered saline (PBS) and resuspended in fresh PYG medium. All experiments were performed with stationary-phase cultures (5–10 days old), containing approximately 95% trophozoites.
Bacterial strains—culture conditions and genetic characterization
Three Y. enterocolitica strains were used in this study, representing both pathogenic and non-pathogenic biotypes. In particular, the reference strain Y. enterocolitica ATCC 23715 (bioserotype 1B/O:8) and strains Ye13P (bioserotype 4/O:3) and Ye15H (bioserotype 1A/O:5) were used. The latter two strains were isolated from human fecal samples of patients. The main characteristics of the strains, including bioserotype, source, country, year of isolation, and virulence markers, are summarized in Table 1.

Table 1. Characteristics of Y. enterocolitica strains used in this study.
The Y. enterocolitica strains were cultured in Tryptone Soy Broth (TSB, Biolife Italiana, Milan, Italy) and subsequently inoculated onto CIN Agar plates (Cefsulodin–Irgasan–Novobiocin, Biolife Italiana, Milan, Italy). Plates were incubated under aerobic conditions at 30°C for 24 ± 2 h. Biotyping was carried out according to the ISO 10273 method (EN ISO 10273:2017, 2017), evaluating specific biochemical reactions, including pyrazinamidase and lipase activity, indole production, xylose and trehalose fermentation, and esculin hydrolysis. For serotyping, O-antisera specific to O:3, O:5, O:8, O:9, and O:27 (Biolife Italiana, Milan, Italy) were employed.
The genetic characterization of the three Y. enterocolitica strains was performed using a SYBR Green-based real-time PCR, targeting the main virulence genes associated with pathogenicity. The amplification conditions and the melting curve analysis were conducted according to the protocols described by Ventola et al. (2023). Specifically, the presence of the ail gene was used as a marker for pathogenic strains, according to the ISO/TS 18867 method (ISO/TS 18867, 2015; Tast Lahti et al., 2023). The strain belonging to biotype 4/O:3 was further characterized by the detection of the virulence plasmid pYV, which encodes virF and yadA. In contrast, the non-pathogenic strain (1A/O:5) was identified by the exclusive presence of ystB, with no detection of ail or pYV.
Antibiotic susceptibility of Yersinia enterocolitica
To optimize the conditions for the invasion assay, different antibiotics were tested to determine their ability to inhibit Y. enterocolitica growth. Pathogenic and non-pathogenic Y. enterocolitica strains were cultured in TSB until reaching an optical density (OD) of 0.5, then distributed into 96-well plates and exposed to different concentrations of tetracycline, gentamicin, penicillin/streptomycin, and kanamycin.
The antibiotic treatments were evaluated at different time points: 1 h, 2 h, 3 h, 4 h, and overnight. Following each interval, the cultures were plated onto CIN Agar to assess bacterial survival.
Intracellular survival assays
The ability of Y. enterocolitica to survive and persist within Acanthamoeba spp. was evaluated using intracellular survival assays performed two technical replicates per condition within a single representative experiment, following the protocol described by Montalbano Di Filippo et al. (2022). Acanthamoeba spp. were seeded into 48-well plates containing PYG medium and incubated at 37°C for 24–48 h to reach at least 70% confluency. After incubation, the medium was removed, and the wells were gently washed with PBS. Y. enterocolitica strains were then added [106 colony-forming units (CFU)/mL is equal to a multiplicity of infection (MOI) of 1], and multi-well plates were incubated at 30°C (optimal Y. enterocolitica growth temperature).
The infection assay was monitored for a total of 55 days, with samples collected at defined time points: 3, 8, 16, 21, and 55 days post-infection. At each time point, the culture medium was carefully inspected, and antibiotic (see details in the Results section) was added to eliminate extracellular bacteria, followed by overnight incubation at 30°C.
To recover intracellular bacteria, Acanthamoeba spp. were washed rigorously three times with PBS to remove any residual of extracellular bacteria and antibiotics and lysed with sodium dodecyl sulfate (SDS; 0.5% final concentration) for 40 min. A portion of the resulting lysate was plated onto CIN Agar and incubated at 30°C for 24 h to quantify CFU, while the remaining lysate was stored at −20°C for subsequent molecular analysis. If Y. enterocolitica colonies were observed on CIN plates, a randomly selected colony was used as template for real-time PCR to confirm the presence of virulence genes, following the protocol described in the Bacterial strains—culture conditions and genetic characterization section. The assays were used for qualitative confirmation purposes only and were not intended to measure gene expression levels.
When no colonies were observed on CIN Agar, the stored lysate was subjected to real-time PCR targeting strain-specific virulence genes to inspect the presence of Y. enterocolitica DNA inside Acanthamoeba spp. (data not shown). This additional analysis aimed to assess whether bacterial DNA persisted within the amoebal lysate, suggesting that the absence of growth might be due to loss of viability during SDS lysis rather than lack of internalization.
As a rule, all experiments included Y. enterocolitica cultures grown in the absence of Acanthamoeba spp. as negative controls, subjected to the same experimental conditions.
Whole genome sequencing of the internalized Yersinia enterocolitica
In order to assess genome integrity and stability, 14 Y. enterocolitica strains were subjected to whole genome sequencing (WGS), including wild-type strains YeREF, Ye13P, and Ye15H strains grown in broth for 24 h, 8 days, and 16 days and five strains that had successfully internalized into both Acanthamoeba spp. strains and were recovered from agar plates. Total DNA was extracted from 2 mL of overnight culture in TSB at 37°C using the GRS Genomic DNA Kit for Bacteria (GRISP Research Solutions, Porto, Portugal). Sequencing was performed using Ion Torrent S5 and S5 Prime platforms (Thermo Fisher Scientific, MA, USA). Libraries of approximately 400 bp were prepared from 100 ng of genomic DNA using the NEBNext® Fast DNA Fragmentation & Library Prep Set for Ion Torrent™ (#E6285L, New England BioLabs, MA, USA). Library quality and quantity were evaluated with the Agilent TapeStation 4200 system using the High Sensitivity D1000 Reagents Kit (#5067-5585, Agilent Technologies). Enrichment was carried out on the Ion Chef System with the Ion 510™, 520™, and 530™ Kits (#A34018, #A27754, and #A27755, Thermo Fisher Scientific), followed by sequencing.
Genomic characterization of the Yersinia enterocolitica strains used in the internalization experiments
The bioinformatic analyses were carried out using the tools present in the Galaxy public server ARIES (https://w3.iss.it/site/aries/) (Knijn et al., 2020). The reads were assembled in contigs using SPAdes v3.14.1 (Bankevich et al., 2012), and the contigs were filtered with the tool “Filter SPAdes repeats” (Iskander, 2022) by using default parameters. The virulence gene content of the genomes was determined using blastn through ARIES with an in-house developed sequence database (accession numbers: ail, CP009846; virF, AF336309; yadA, CP009845; and ystB, D88145), by using 85% of sequence identity and 70% alignment length as thresholds. Phylogenomics analysis was performed by determining the core genome multilocus sequence typing (cgMLST) using the chewBBACA tool (Silva et al., 2018) and the scheme developed by the INNUENDO project, which comprises 2,406 loci in total (Llarena et al., 2018; Mirko et al., 2018). The distances between strains were calculated by pairwise comparison of the allelic profiles through the chewTree tool (https://www.iss.it/site/aries). The pairwise comparison was considered reliable when >90% of loci for each sample were assigned to an allele. For each pair of samples, the alleles that were not found, only partially found, or not correctly assigned to any locus were excluded from the analysis, as previously described (Gigliucci et al., 2021). Genome annotation was performed with the Prokka tool (Seemann, 2014) on the ARIES server.
Results
Yersinia enterocolitica survival within Acanthamoeba spp.
Before presenting our results, it is important to note that among the antibiotics tested, only gentamicin fully suppressed bacterial growth after overnight incubation (optimal concentration of 350 µg/mL).
Our findings showed that all tested Y. enterocolitica strains were internalized by both Acanthamoeba species and remained viable for different timeframes, depending on the strain and the amoebal host. Viability was maintained for up to 8 days in Acanthamoeba spp. CDC: V036 and for up to 16 days in the wild-type isolate. Further details are provided in Table 2. Microscopic examination confirmed that Acanthamoeba spp. strains remained structurally intact throughout the entire incubation period at 30°C, even in the absence of nutrient supplementation. Furthermore, all internalized Y. enterocolitica strains retained their original virulence gene profiles, as confirmed by real-time PCR (data not shown), indicating preservation of their pathogenic potential. In detail, all the tested recovered colonies of the Ye13P strain were positive for ail, yadA, and virF genes; all those of the YeREF strain were positive for the ail gene; and all those of the Ye15H strain were positive for the ystB gene only.
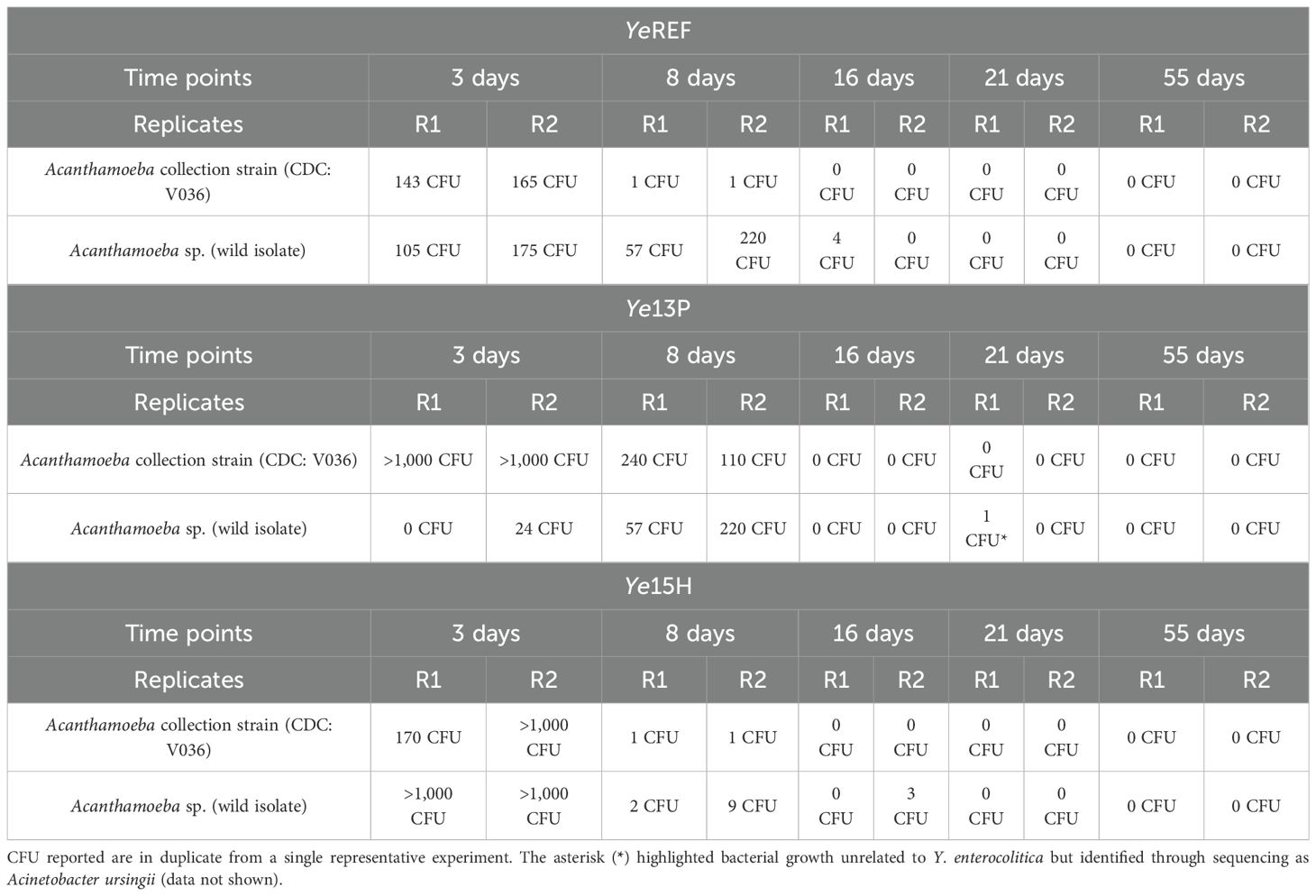
Table 2. Intracellular survival of Y. enterocolitica within Acanthamoeba spp.
Genomic characterization of the internalized Yersinia enterocolitica strains
The median number of contigs obtained from the genome assemblies was 64, with a median N50 value of 140,608 bp, indicating high-quality assemblies across all samples. The expected virulence genes could be retrieved in the genomes for all the strains analyzed. In detail, blast analysis confirmed that the YeREF strain was positive for the ail gene either after growth in broth for 24 h, 8 days, or 16 days or when recovered after internalization in the Acanthamoeba collection strain for 8 days and in the wild Acanthamoeba strain for 16 days; similarly, the Ye13P strain was positive for ail, yadA, and virF either after growth in broth for 24 h, 8 days, or 16 days or when recovered after internalization in the Acanthamoeba collection strain for 8 days; finally, Ye15H was positive for ystB either after growth in broth for 24 h, 8 days, or 16 days or when recovered after internalization in the Acanthamoeba collection strain for 8 days and in the wild Acanthamoeba strain for 16 days.
To evaluate genome integrity and stability following intracellular persistence, a comparative phylogenetic analysis was performed using cgMLST. A minimum of 2,348 allelic exact matches were detected in all the genomes tested among the 2,406 loci part of the scheme used.
For comparison, the same Y. enterocolitica strains cultured in broth under standard laboratory conditions for 16 days were included in the analysis. The resulting allelic distance-based dendrogram was visualized and modified using iTOL (https://itol.embl.de/; Letunic and Bork, 2007) and is shown in Figure 1.

Figure 1. Phylogenomic analysis of Y. enterocolitica strains analyzed in this study—both those recovered under standard laboratory conditions (broth culture—highlighted by a star mark) and those following internalization within amoebal hosts, through cgMLST. Clade colors denote bacterial pathogenicity (red: pathogenic, orange: moderately pathogenic, and blue: non-pathogenic); pathogenicity was determined based on the detection of known virulence genes and serotype information as reported in the Materials and Methods section.
The analysis revealed three distinct clades corresponding to the strains under study: YeREF (Y. enterocolitica ATCC 23715 1B/O:8), Ye13P (Y. enterocolitica 4/O:3), and Ye15H (Y. enterocolitica 1A/O:5). Each clade exhibited minimal variability, suggesting a relatively high level of genomic stability during intracellular persistence within Acanthamoeba spp. Specifically, the maximum number of allelic differences (ADs) detected among isolates internalized for 8 days in Acanthamoeba spp. CDC: V036 was 10, detected in Ye13P.
Notably, no allelic variation was detected in strains recovered from wild Acanthamoeba spp., whereas isolates internalized within the reference strain (Acanthamoeba spp. CDC: V036) accumulated a moderate number of mutations, exceeding those observed under standard laboratory (broth) conditions. In detail, (i) Ye13P accumulated 6 ADs after 8 days in broth and 10 ADs after 8 days within the reference strain of amoeba. Six loci were shared between both conditions, while four additional loci were uniquely mutated during the intracellular persistence: in detail, dusB (tRNA-dihydrouridine synthase B), ravA (ATPase RavA), igaA (intracellular growth attenuator protein IgaA), and rnb (Exoribonuclease 2); (ii) Ye15H showed no ADs after 8 days in broth, but displayed 2 ADs following internalization in the CDC: V036 host, specifically in manA (Mannose-6-phosphate isomerase) and tap (Methyl-accepting chemotaxis protein IV).
Interestingly, a mutation in the igaA gene was identified in strain Ye13P exclusively following interaction with the Acanthamoeba CDC: V036 host. This gene encodes an intracellular growth attenuator described in S. enterica, where it is involved in the negative regulation of the Rcs phosphorelay system, a key pathway controlling bacterial stress responses and virulence (Dominguez-Bernal et al., 2004). However, the igaA locus was not detected at all in the Ye13P genome after growing in broth for 8 days.
Discussion
FLA often act as environmental shelters that enhance the survival of foodborne zoonotic pathogens under adverse conditions. Y. enterocolitica has been shown to benefit from this interaction, with early work by King et al. (1988) demonstrating increased chlorine resistance following internalization by Acanthamoeba castellanii. Later studies proposed a possible mutualistic relationship under nutrient-rich conditions, allowing bacterial survival without damaging the host (Greub and Raoult, 2004; Siddiqui and Khan, 2012).
Experimental studies confirmed that Y. enterocolitica can persist in amoebal hosts under multiple stress conditions, including long-term intracellular survival (up to 14 days) and viability at temperatures simulating refrigeration (Anacarso et al., 2011; Lambrecht et al., 2013). Internalization has also been associated with increased resistance to disinfectants like chlorine, raising concerns for sanitation protocols also in industrial settings (Thomas et al., 2010; Vaerewijck et al., 2014; Lambrecht et al., 2017).
Together, these findings highlight the ecological and public health relevance of Y. enterocolitica and FLA interactions. However, important gaps remain regarding the effect of internalization on strains with different virulence profiles, their intracellular fate, and the consequences for structural and genetic stability. To address these aspects, we explored the intracellular dynamics of different Y. enterocolitica isolates using two Acanthamoeba spp. strains: one collection-derived and one field-isolated.
Our results confirmed the ability of all tested Y. enterocolitica strains to remain viable after internalization, with intracellular persistence observed up to 8 days in the collection-derived Acanthamoeba spp. strain and extending to 16 days in the wild-type isolate; this extends the findings of Lambrecht et al. (2013), who reported viability within A. castellanii for up to 14 days, and suggests that host origin may influence the duration of bacterial survival. Our morphological analysis revealed that both amoebal strains remained structurally intact at 30°C throughout the incubation period, even in the absence of nutrient supplementation, corroborating earlier reports on the stability and resilience of Acanthamoeba spp. as a protozoan host (Greub and Raoult, 2004; Siddiqui and Khan, 2012).
Importantly, real-time PCR and genome analysis demonstrated the conservation of virulence gene profiles in all internalized Y. enterocolitica strains. This observation supports the idea that intracellular persistence does not compromise the pathogenic potential of the Y. enterocolitica, in contrast with scenarios in which intracellular niches lead to attenuation or gene loss. While this aspect had not been extensively addressed in previous studies on Y. enterocolitica, similar findings have been reported in S. enterica and in STEC, where intracellular maintenance in protozoa did not alter key virulence traits (Gaze et al., 2003; Montalbano Di Filippo et al., 2022).
However, WGS revealed that internalization may induce subtle genetic changes depending on the amoebal host. Indeed, no allelic variation was observed in bacteria recovered from the wild-type Acanthamoeba spp. strain, while ADs were detected in isolates internalized within the culture-derived strain, suggesting that wild amoebal strains could serve as reservoirs where genetic stability of Y. enterocolitica strains is preserved. The detected differences included a mutation in the igaA gene of strain Ye13P, detected exclusively after interaction with Acanthamoeba CDC: V036. This gene has been described in S. enterica to encode an inner membrane protein involved in the negative regulation of the Rcs phosphorelay system, a key regulator of envelope stress responses and virulence factor expression in enterobacteria (Dominguez-Bernal et al., 2004). This system has been implicated in bacterial adaptation to intracellular environments and biofilm formation (Clarke, 2010; Wall et al., 2020). Notably, the igaA locus became undetectable in Ye13P cultured in broth under standard laboratory conditions for the same time of interaction with the amoebal strain, suggesting that this locus could be particularly unstable and subjected to growing conditions. Further analysis would be necessary to investigate the role of this gene in the intracellular survival of Y. enterocolitica and in virulence regulation.
Taken together, our findings, in line with previous literature (Montalbano Di Filippo et al., 2022), support the hypothesis that Acanthamoeba spp. does not merely provide physical protection for bacterial pathogens, including Y. enterocolitica, but may also act as a selective microenvironment capable of modulating bacterial genome stability. Moreover, survival in Acanthamoeba species could allow Y. enterocolitica to escape detection in animal samples or in the vehicles of infection, while maintaining intact its virulence gene asset, as observed for the Ye13P pathogenic strain. While these results contribute to a broader understanding of amoebae as environmental reservoirs, they primarily expand current knowledge on Y. enterocolitica biology, particularly in relation to its genomic and functional plasticity in protozoan hosts.
In this context, amoebae should be increasingly considered as dynamic hosts with the potential to influence bacterial evolution. Their capacity to internalize diverse microbial taxa, maintain structural integrity under stress, and potentially mediate gene exchange through mechanisms such as outer membrane vesicles or microvesicles warrants further investigation.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found below: https://www.ncbi.nlm.nih.gov/, PRJEB89775.
Author contributions
PC: Methodology, Writing – original draft, Conceptualization. VM: Methodology, Writing – original draft, Data curation. ED: Methodology, Writing – original draft. EV: Writing – original draft, Methodology. SMa: Writing – original draft, Methodology. MM: Methodology, Writing – original draft. VL: Methodology, Writing – original draft. RL-E: Methodology, Writing – original draft. MR-B: Methodology, Writing – original draft. IF: Methodology, Writing – original draft. JL: Methodology, Funding acquisition, Writing – original draft. SMo: Funding acquisition, Writing – original draft. MMF: Writing – original draft, Conceptualization, Methodology.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. The study was supported by intramural funds of the Istituto Superiore di Sanità and by Consorcio Centro De Investigación Biomédica En Red M.P. (CIBERINFEC) de Enfermedades Infecciosas (CB21/13/00100), Instituto de Salud Carlos III, Madrid, España; el Ministerio de Sanidad, Consumo y Bienestar Social; and Fundación La Caixa-Caja Canarias 2022CLISA26. RL-E was funded by Cabildo Insular de Tenerife 2023–2028 PROYECTO CC20230222, CABILDO 23.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Adeolu, M., Alnajar, S., Naushad, S., and Gupta, R. S. (2016). Genome-based phylogeny and taxonomy of the ‘Enterobacteriales’: proposal for Enterobacterales ord. nov. divided into the families Enterobacteriaceae, Erwiniaceae fam. nov., Pectobacteriaceae fam. nov., Yersiniaceae fam. nov., Hafniaceae fam. nov., Morganellaceae fam. nov., and Budviciaceae fam. nov. Int. J. Syst. Evol. Microbiol. 66, 5575–5599. doi: 10.1099/ijsem.0.001485
Anacarso, I., de Niederhäusern, S., Messi, P., Guerrieri, E., Iseppi, R., Sabia, C., et al. (2011). Acanthamoeba polyphaga, a potential environmental vector for the transmission of food-borne and opportunistic pathogens. J. Basic Microbiol. 52, 261–268. doi: 10.1002/jobm.201100097
Bankevich, A., Nurk, S., Antipov, D., Gurevich, A. A., Dvorkin, M., Kulikov, A. S., et al. (2012). SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 19, 455–477. doi: 10.1089/cmb.2012.0021
Bhagat, N. and Virdi, J. S. (2007). Distribution of virulence-associated genes in Yersinia enterocolitica biovar 1A correlates with clonal groups and not the source of isolation. FEMS Microbiol. Lett. 266, 177–183. doi: 10.1111/j.1574-6968.2006.00524.x
Chavatte, N., Baré, J., Lambrecht, E., Van Damme, I., Vaerewijck, M., Sabbe, K., et al. (2014). Co-occurrence of free-living protozoa and foodborne pathogens on dishcloths: implications for food safety. Int. J. Food Microbiol. 191, 89–96. doi: 10.1016/j.ijfoodmicro.2014.08.030
Clarke, D. J. (2010). The Rcs phosphorelay: more than just a two-component pathway. Future Microbiol. 5, 1173–1184. doi: 10.2217/fmb.10.83
Dinda, S. K., Hazra, S., De, A., Datta, A., Das, L., Pattanayak, S., et al. (2024). Amoebae: beyond pathogens- exploring their benefits and future potential. Front. Cell Infect. Microbiol. 14. doi: 10.3389/fcimb.2024.1518925
Domínguez-Bernal, G., Pucciarelli, M. G., Ramos-Morales, F., García-Quintanilla, M., Cano, D. A., Casadesús, J., et al. (2004). Repression of the RcsC-YojN-RcsB phosphorelay by the IgaA protein is a requisite for Salmonella virulence. Mol. Microbiol. 53, 1437–1449. doi: 10.1111/j.1365-2958.2004.04213.x
EFSA and ECDC (2024). The European Union One Health 2023 zoonoses report. EFSA J. 22, e09106. doi: 10.2903/j.efsa.2024.9106
EN ISO 10273:2017 (2017). Microbiology of the food chain - Horizontal method for the detection of pathogenic Yersinia enterocolitica (Geneva: International Organization for Standardization).
Fredriksson-Ahomaa, M. (2017). Yersinia enterocolitica. In: Foodborne diseases. 3rd ed (London: Elsevier, Academic Press), 223–233. doi: 10.1016/B978-0-12-385007-2.00009-7
Gaze, W. H., Burroughs, N., Gallagher, M. P., and Wellington, E. M. (2003). Interactions between Salmonella typhimurium and Acanthamoeba polyphaga, and observation of a new mode of intracellular growth within contractile vacuoles. Microb. Ecol. 46, 358–369. doi: 10.1007/s00248-003-1001-3
Gigliucci, F., van Hoek, A. H. A. M., Chiani, P., Knijn, A., Minelli, F., Scavia, G., et al. (2021). Genomic characterization of hlyF-positive shiga toxin-producing Escherichia coli, Italy and the Netherlands 2000–2019. Emerg. Infect. Dis. 27, 853–861. doi: 10.3201/eid2703.203110
Greub, G. and Raoult, D. (2004). Microorganisms resistant to free-living amoebae. Clin. Microbiol. Rev. 17, 413–433. doi: 10.1128/cmr.17.2.413-433.2004
Henriquez, F. L., Mooney, R., Bandel, T., Giammarini, E., Zeroual, M., Fiori, P. L., et al. (2021). Paradigms of protist/bacteria symbioses affecting human health: Acanthamoeba species and Trichomonas vaginalis. Front. Microbiol. 11. doi: 10.3389/fmicb.2020.616213
Iskander, M. (2022). Galaxy tools repository. Available online at: https://github.com/phac-nml/galaxy_tools/. (Accessed May 9, 2025)
ISO/TS 18867 (2015). Microbiology of the food chain – Polymerase chain reaction (PCR) for the detection of foodborne pathogens – Detection of pathogenic Yersinia enterocolitica and Yersinia pseudotuberculosis (Geneva: International Organization for Standardization).
Kiderlen, A. F., Radam, E., Laube, U., and Martinez, A. J. (2014). Resistance to intranasal infection with Balamuthia mandrillaris amoebae is T-cell dependent. J. Eukaryot Microbiol. 62, 26–33. doi: 10.1111/jeu.12189
King, C. H., Shotts, E. B., Wooley, R. E., and Porter, K. G. (1988). Survival of coliforms and bacterial pathogens within protozoa during chlorination. Appl. Environ. Microbiol. 54, 3023–3033. doi: 10.1128/aem.54.12.3023-3033.1988
Knijn, A., Michelacci, V., Orsini, M., and Morabito, S. (2020). Advanced research infrastructure for experimentation in genomics (ARIES): a lustrum of Galaxy experience. bioRxiv, 2020.05.14.095901. doi: 10.1101/2020.05.14.095901
Lambrecht, E., Baré, J., Sabbe, K., and Houf, K. (2017). Impact of Acanthamoeba cysts on stress resistance of Salmonella enterica serovar Typhimurium, Yersinia enterocolitica 4/O:3, Listeria monocytogenes 1/2a, and Escherichia coli O:26. Appl. Environ. Microbiol. 83, e00754–e00717. doi: 10.1128/AEM.00754-17
Lambrecht, E., Baré, J., Van Damme, I., Bert, W., Sabbe, K., and Houf, K. (2013). Behavior of Yersinia enterocolitica in the presence of the bacterivorous Acanthamoeba castellanii. Appl. Environ. Microbiol. 79, 6407–6413. doi: 10.1128/AEM.01915-13
Letunic, I. and Bork, P. (2007). Interactive tree of life (iTOL): An online tool for phylogenetic tree display and annotation. Bioinformatics 23, 127–128. doi: 10.1093/bioinformatics/btl529
Llarena, A. K., Ribeiro-Gonçalves, B. F., Silva, D. N., Halkilahti, J., MaChado, M. P., da Silva, M. S., et al. (2018). INNUENDO: A cross-sectoral platform for the integration of genomics in the surveillance of food-borne pathogens. EFSA Supporting Publ. 15, EN–1498. doi: 10.2903/sp.efsa.2018.EN-1498
Lorenzo-Morales, J., Khan, N. A., and Walochnik, J. (2015). An update on Acanthamoeba keratitis: diagnosis, pathogenesis and treatment. Parasite 22, 10. doi: 10.1051/parasite/2015010
Mancini, M. E., Beverelli, M., Donatiello, A., Didonna, A., Dattoli, L., Faleo, S., et al. (2022). Isolation and characterization of Yersinia enterocolitica from foods in Apulia and Basilicata regions (Italy) by conventional and modern methods. PloS One 17, e0268706. doi: 10.1371/journal.pone.0268706
Mancusi, A., Delibato, E., Peruzy, M. F., Girardi, S., Di Maro, O., Cristiano, D., et al. (2024). Evaluation of two real-time PCR methods to detect Yersinia enterocolitica in bivalve molluscs collected in Campania region. Food Res. Int. 192, 114789. doi: 10.1016/j.foodres.2024.114789
Mirko, R., Silva, M. S., Ribeiro-Gonçalves, B. F., Silva, D. N., MaChado, M. P., Oleastro, M., et al. (2018). INNUENDO Whole Genome and Core Genome MLST Schemas and Datasets for Yersinia Enterocolitica (Version 1.0) (Zenodo). doi: 10.5281/zenodo.1421262
Montalbano Di Filippo, M., Boni, A., Chiani, P., Marra, M., Carollo, M., Cristofari, L., et al. (2022). Exploring the nature of interaction between shiga toxin producing Escherichia coli (STEC) and free-living amoeba - Acanthamoeba sp. Front. Cell Infect. Microbiol. 12. doi: 10.3389/fcimb.2022.926127
Moran, S., Mooney, R., and Henriquez, F. L. (2022). Diagnostic considerations for non-Acanthamoeba amoebic keratitis and clinical outcomes. Pathogens 11, 219. doi: 10.3390/pathogens11020219
Reuter, S., Connor, T. R., Barquist, L., Walker, D., Feltwell, T., Harris, D., et al. (2014). Parallel independent evolution of pathogenicity within the genus Yersinia. Proc. Natl. Acad. Sci. U.S.A. 111, 6768–6773. doi: 10.1073/pnas.1317161111
Rivas, L., Strydom, H., Paine, S., Wang, J., and Wright, J. (2021). Yersiniosis in New Zealand. Pathogens 10, 191. doi: 10.3390/pathogens10020191
Scheid, P. (2018). Free-living amoebae as human parasites and hosts for pathogenic microorganisms. Proceedings 2, 692. doi: 10.3390/proceedings2110692
Schroeder, J. M., Booton, G. C., Hay JNiszl, I. A., Seal DVMarkus, M. B., Fuerst, P. A., and Byers, T. J. (2001). Use of subgenic 18S ribosomal DNA PCR and sequencing for genus and genotype identification of acanthamoebae from humans with keratitis and from sewage sludge. J Clin Microbiol 39, 1903–1911. doi: 10.1128/jcm.39.5
Seemann, T. (2014). Prokka: rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069. doi: 10.1093/bioinformatics/btu153
Shoaib, M., Shehzad, A., Raza, H., Niazi, S., Khan, I. M., Akhtar, W., et al. (2019). A comprehensive review on the prevalence, pathogenesis and detection of Yersinia enterocolitica. RSC Adv. 9, 41010–41021. doi: 10.1039/c9ra06988g
Siddiqui, R. and Khan, N. A. (2012). Biology and pathogenesis of acanthamoeba. Parasit Vectors 5, 6. doi: 10.1186/1756-3305-5-6
Silva, M., MaChado, M. P., Silva, D. N., Rossi, M., Moran-Gilad, J., Santos, S., et al. (2018). chewBBACA: A complete suite for gene-by-gene schema creation and strain identification. Microb. Genom 4, e000166. doi: 10.1099/mgen.0.000166
Tast Lahti, E., Karamehmedovic, N., Riedel, H., Blom, L., Boel, J., Delibato, E., et al. (2023). One Health surveillance – A cross-sectoral detection, characterization, and notification of foodborne pathogens. Front. Public Health 11. doi: 10.3389/fpubh.2023.1129083
Thomas, V., McDonnell, G., Denyer, S. P., and Maillard, J. Y. (2010). Free-living amoebae and their 480 intracellular pathogenic microorganisms: risks for water quality. FEMS Microbiol. Rev. 34, 231–259. doi: 10.1111/j.1574-6976.2009.00190.x
Todd, E. C. D. (2014). Bacteria: Yersinia enterocolitica and Yersinia pseudotuberculosis. Encyclopedia Food Saf., 574–580. doi: 10.1016/B978-0-12-378612-8.00121-9
Vaerewijck, M., Baré, J., Lambrecht, E., Sabbe, K., and Houf, K. (2014). Interactions of foodborne pathogens with free-living protozoa: potential consequences for food safety. Compr. Rev. Food Sci. Food Saf. 13, 924–944. doi: 10.1111/1541-4337.12100
Ventola, E., Lovari, S., Farneti, S., Finazzi, G., Bilei, S., Owczarek, S., et al. (2023). Molecular characterization of Yersinia enterocolitica strains to evaluate virulence associated genes. Ann. Ist Super Sanita 59, 280–285. doi: 10.4415/ann_23_04_07
Visvesvara, G. S., Moura, H., and Schuster, F. L. (2007). Pathogenic and opportunistic free-living amoebae: Acanthamoeba spp., Balamuthia mandrillaris, Naegleria fowleri, and Sappinia diploidea. FEMS Immunol. Med. Microbiol. 50, 1–26. doi: 10.1111/j.1574-695X.2007.00232.x
Keywords: Y. enterocolitica, free living amoebae (FLA), food safety, zoonosis, Y. enterocolitica biotypes, Acanthamoeba spp.
Citation: Chiani P, Michelacci V, Delibato E, Ventola E, Mannan S, Marra M, Libri V, Rodríguez-Expósito RL, Reyes-Batlle M, de Fuentes I, Lorenzo-Morales J, Morabito S and Montalbano Di Filippo M (2025) Survival and genomic stability of Yersinia enterocolitica in environmental Acanthamoeba spp. Front. Cell. Infect. Microbiol. 15:1642352. doi: 10.3389/fcimb.2025.1642352
Received: 06 June 2025; Accepted: 08 October 2025;
Published: 30 October 2025.
Edited by:
Li Zhang, University of New South Wales, AustraliaReviewed by:
Fauzy Nasher, University of London, United KingdomDipak Manna, Ramakrishna Mission Vivekananda Educational and Research Institute, India
Copyright © 2025 Chiani, Michelacci, Delibato, Ventola, Mannan, Marra, Libri, Rodríguez-Expósito, Reyes-Batlle, de Fuentes, Lorenzo-Morales, Morabito and Montalbano Di Filippo. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paola Chiani, cGFvbGEuY2hpYW5pQGlzcy5pdA==