 Fang-Fang Liu1
Fang-Fang Liu1 Ke Li2*
Ke Li2*- 1Department of Pathology, The Central Hospital of Wuhan, Tongji Medical College, Huazhong University of Science and Technology, Wuhan, China
- 2School of Life and Health Sciences, Hainan Province Key Laboratory of One Health, Collaborative Innovation Center of One Health, Hainan University, Haikou, Hainan, China
Malaria associated anemia is increasingly recognized as a consequence not only of red cell destruction but of profound, parasite driven disruption of erythropoiesis within the bone marrow niche. Here, we synthesize recent in vitro, ex vivo, clinical and postmortem studies to construct a unified mechanistic framework in which four interlocking pathways converge to produce dyserythropoiesis. First, a cytokine storm dominated by IL-6, TNF-α, IFN-γ and MIF suppresses erythropoietin synthesis, upregulates hepcidin and diverts erythroid progenitors toward myeloid fate via destabilization of GATA-1. Second, hemozoin crystals catalyze Fenton chemistry and lipid peroxidation, generating 4-hydroxynonenal adducts that cripple GATA-1 and trigger mitochondrial apoptosis of erythroblasts. Third, Plasmodium parasites preferentially infect orthochromatic erythroblasts, prolonging a 10-day gametocyte maturation cycle beyond the host’s 3–4-day enucleation window and releasing extracellular vesicles that arrest terminal differentiation. Fourth, hemozoin-laden macrophages remodel erythroblastic islands, precipitating local iron restriction and sustained oxidative stress. Together these processes create a “developmental sanctuary” that favors parasite persistence while crippling host erythropoiesis. We also highlight emerging single-cell and spatial-omics technologies, together with 3-D bone-marrow organoids, as platforms for dissecting spatiotemporal parasite–host interactions and for testing niche-targeted therapies aimed at reversing ineffective erythropoiesis.
1 Introduction
Malaria, a major parasitic disease threatening global public health security, exhibits pathological complexity not only in the systemic inflammatory response during acute infection but also in the long-term impact of chronic anemia on patient quality of life. According to the World Health Organization’s 2024 World Malaria Report, there were an estimated 263 million malaria cases globally in 2023, with approximately 597,000 malaria-related deaths (Venkatesan, 2025). Severe anemia, a critical complication resulting from red blood cell destruction, significantly contributes to malaria’s severity and mortality, particularly in high-transmission regions. Children under five years of age and pregnant women represent particularly vulnerable groups. In high-transmission areas, malaria is one of the most common indications for blood transfusion, yet transfusion thresholds in severe malaria management remain uncertain (Ackerman et al., 2020). Although artemisinin-based combination therapies (ACTs) have significantly reduced acute-phase mortality, clinical observations reveal that a subset of patients experience persistent anemia even after parasite clearance (Eastman and Fidock, 2009; Fernandez-Arias et al., 2016; Ndour et al., 2017).
Classical theories attribute malarial anemia to three primary mechanisms: direct parasite lysis of infected red blood cells (RBCs); splenic hyper-removal of uninfected RBCs with impaired deformability and disturbances in iron metabolism coupled with insufficient erythropoietin (EPO) synthesis due to chronic inflammation (Looareesuwan et al., 1991; Dondorp et al., 1999). However, this framework is increasingly challenged by new evidence. For instance, The deposition of malaria pigments (hemozoin) in the bone marrow directly induces apoptosis of erythroid precursors and is significantly associated with ineffective erythropoiesis and dyserythropoiesis in bone marrow (Casals-Pascual et al., 2006; Lamikanra et al., 2009; Aguilar et al., 2014), while iron kinetic studies demonstrate that the decline in hemoglobin synthesis rate precedes RBC destruction (Pathak and Ghosh, 2016). Thus, ineffective erythropoiesis and dyserythropoiesis may be potential core drivers of malaria anemia, while the bone marrow is a “hidden battlefield”. Substantial progress in understanding malaria pathogenesis has stemmed from meticulous characterization of the bone marrow microenvironment. Intravital imaging confirms that immature P. falciparum gametocytes and P. vivax asexual/sexual stages colonize the bone marrow, forming a significant reservoir (Joice et al., 2014; Baro et al., 2017; De Niz et al., 2018). Single-cell transcriptomics in murine models reveal dysregulation of erythroid transcriptional networks (e.g., GATA1) in infected progenitors, contributing to impaired erythropoiesis (Silva-Filho et al., 2020). Additionally, parasite metabolites (e.g., hemozoin) and inflammatory cytokines (TNF-α, IFN-γ) disrupt hematopoietic niche function, triggering microenvironmental remodeling (Silva-Filho et al., 2020).
The regulatory role of host genetic background in anemia susceptibility cannot be overlooked. Polymorphisms in the ABO and Duffy blood group systems significantly influence disease progression: individuals with blood group O exhibit reduced P. falciparum PfEMP1-mediated cytoadhesion due to the absence of A/B antigens (Degarege et al., 2019), while Duffy antigen-negative populations (common in individuals of African descent) display innate resistance to P. vivax infection (Baird, 2022). Since blood group influences malaria susceptibility, the anemia status of individuals with different blood groups after contracting malaria may vary. This may also suggest that blood group molecules could potentially impact erythropoiesis. However, changes in the expression of blood group molecules might merely be a downstream manifestation of altered erythropoiesis. Overall, the pathology of malaria anemia may result from the interplay of host genetics, parasite virulence, and microenvironmental remodeling.
This mini-review aims to elucidate the bone marrow pathogenesis of malarial anemia, focusing on three core questions: 1) How parasites disrupt erythropoiesis through direct infection and paracrine effects; 2) How the inflammatory microenvironment and oxidative stress synergistically disrupt hematopoietic homeostasis; and 3) Prospects for developing novel therapeutic strategies targeting bone marrow pathways. By integrating clinical data, molecular biology evidence, and innovative model systems (e.g., bone marrow organoids), this study will provide a theoretical foundation for reversing ineffective erythropoiesis.
2 Cytokine-mediated suppression of erythropoiesis
To control parasitemia, host immune cells release a cascade of pro-inflammatory and anti-inflammatory cytokines, chemokines, growth factors, and other mediators. While these immune responses confer host protection, in this inflammatory state, multiple factors simultaneously suppress erythropoiesis by interfering with erythroid progenitor differentiation, disrupting iron metabolism homeostasis, and altering the hematopoietic microenvironment, thereby exacerbating anemia.
During the early stages of malaria, the cytokine IL-6 plays a protective role, stimulating the acute phase response and supporting immune cell differentiation (Gowda and Wu, 2018). Peripheral blood mononuclear cells (PBMCs) are likely the primary source of IL-6 during acute malaria (Aubouy et al., 2002). Studies in murine models indicate that interleukin-6 (IL-6) mediates protective immunity against the pre-erythrocytic stage of malaria by inducing interleukin-1β (IL-1β) and tumor necrosis factor-α (TNF-α), and enhances specific immunoglobulin G (IgG) antibodies to control blood-stage parasitemia during the erythrocytic phase (Pied et al., 1992). However, as the disease progresses, persistently elevated levels of IL-6, IL-1β, and others lead to systemic inflammation (Obeagu, 2024). Research in malaria infection models reveals that IL-6 regulates iron metabolism homeostasis through a dual mechanism: on one hand, it induces hepatic hepcidin synthesis via the JAK2-STAT3 pathway, resulting in decreased serum iron concentration (Wrighting and Andrews, 2006); on the other hand, it directly suppresses renal erythropoietin (EPO) expression (Jelkmann, 1998). Furthermore, elevated serum IL-6 levels in malaria patients positively correlate with anemia severity (Lyke et al., 2004).
Beyond IL-6, IL-1β also plays a crucial role in suppressing erythropoietic differentiation. This cytokine promotes PU.1 expression by activating the NLRP3 inflammasome (Pietras et al., 2016), while simultaneously disrupting the erythroid-myeloid differentiation balance through caspase-1-mediated proteolysis of GATA-1 (Tyrkalska et al., 2019). Indeed, similar to TNF-α, IL-1β promotes the acute inflammatory response at the onset of malaria infection, providing a first line of defense against invading pathogens (Penha-Goncalves, 2019). For example, IL-1β can synergize with IL-1α and TNF-α to enhance nitric oxide (NO) and IFN-γ production in murine malaria models (Rockett et al., 1994). However, sustained high-level production of IL-1β can induce anemia, a phenomenon also documented in numerous other disease models beyond malaria (Pascual et al., 2005).
Members of the interferon family, particularly IFN-γ, play a prominent role in inflammation-associated anemia. Plasmodium parasites and their byproducts, such as hemozoin, can amplify a robust inflammatory response by increasing TNF-α and IFN-γ (Giribaldi et al., 2004; Jaramillo et al., 2004). While these inflammatory mediators stimulate monocyte/macrophage activation and help control parasitemia during the early infection phase (Kremsner et al., 1995), substantial evidence confirms that their persistent overproduction can also significantly suppress erythropoiesis (Miller et al., 1989; Felli et al., 2005). IFN-γ promotes myeloid differentiation via the IRF1/PU.1 axis (Libregts et al., 2011). In TLR9-driven models of hemophagocytic lymphohistiocytosis (HLH), this cytokine not only suppresses bone marrow erythropoiesis but also limits compensatory splenic erythropoiesis (Canna et al., 2013).
Macrophage migration inhibitory factor (MIF) plays a distinct role in parasite infection-associated anemia. The malarial metabolite hemozoin induces monocytes to release MIF, which exerts synergistic inhibitory effects with TNF and IFN-γ (Martiney et al., 2000; McDevitt et al., 2015). Furthermore, parasite-secreted MIF homologs may amplify the suppression of uninfected erythroid progenitors (Ghosh et al., 2019). Additionally, studies show that MIF inhibits erythropoietin-dependent erythroid colony formation, as well as colony formation derived from multipotential (CFU-GEMM) and granulocyte-macrophage (CFU-GM) progenitor cells (Martiney et al., 2000; Chaiyaroj et al., 2004). It has been also demonstrated in murine studies that compared to wild-type controls, MIF-knockout mice infected with malaria develop less severe anemia, exhibit improved erythroid progenitor development, and display higher survival rates (McDevitt et al., 2006).
3 Oxidative damage effects of hemozoin
Hemozoin (Hz), a dark brown crystalline substance, is formed when malaria parasites detoxify toxic heme into insoluble β-hematin crystals during hemoglobin digestion within infected erythrocytes. Following erythrocyte invasion, Plasmodium parasites extensively ingest host hemoglobin, transport it to acidic digestive vacuoles for proteolytic degradation, and polymerize liberated heme into Hz via biomineralization. Clinical studies demonstrate that plasma Hz levels correlate significantly with anemia severity and reticulocyte suppression in malaria patients (Aguilar et al., 2014), while in vitro exposure to Hz directly inhibits erythropoiesis (Giribaldi et al., 2004; Skorokhod et al., 2010). Crucially, Hz accumulates in the bone marrow niche where it inflicts direct damage on erythroid hematopoiesis primarily through induction of oxidative stress and lipid peroxidation cascades.
3.1 Iron-mediated ROS generation and cellular consequences
The iron-rich crystalline lattice of Hz catalyzes robust Reactive Oxygen Species (ROS) production via Fenton chemistry (Schwarzer et al., 2003; Dumarchey et al., 2022). Studies have shown bone marrow Hz burden correlates positively with anemia severity in patients (Casals-Pascual et al., 2006; Aguilar et al., 2014). Experimental models confirm that Hz-exposed macrophages and erythroid precursors exhibit markedly elevated ROS levels, resulting in membrane lipid peroxidation, mitochondrial dysfunction, and direct suppression of erythroid progenitor proliferation and differentiation (Jaramillo et al., 2003; Shio et al., 2009; Cambos et al., 2010; Barrera et al., 2011). This oxidative milieu further activates pro-apoptotic pathways (notably caspase-3) to induce erythroblast apoptosis (Lamikanra et al., 2009; Skorokhod et al., 2010). Importantly, Hz-induced oxidative stress may trigger mitochondrial dysfunction, as elevated ROS levels are known to disrupt mitochondrial integrity in other pathological contexts (Zorov et al., 2014). This could potentially initiate a cycle of amplified oxidative damage and depletion of antioxidants such as glutathione (GSH) (Kotepui et al., 2023), though direct evidence linking Hz to mitochondrial electron transport chain disruption remains limited.
3.2 4-Hydroxynonenal as a cytotoxic effector
Hz-triggered peroxidation of polyunsaturated fatty acids (PUFAs) non-enzymatically generates 4-hydroxynonenal (4-HNE), a highly reactive lipid peroxidation end-product (Miller et al., 2005; Skorokhod et al., 2005). In vitro studies reveal that even low concentrations of 4-HNE potently inhibit BFU-E and CFU-E colony formation independently of apoptosis induction (Giribaldi et al., 2004; Skorokhod et al., 2010). Instead, 4-HNE imposes G0/G1 cell cycle arrest in erythroid progenitors and downregulates expression of GATA-1—a master transcriptional regulator of erythropoiesis—through covalent modification of its DNA-binding domain (Skorokhod et al., 2010). This dual action disrupts hemoglobin synthesis and terminal erythroid maturation. Mechanistically, 4-HNE-adducted GATA-1 suffers impaired DNA-binding affinity and accelerated proteasomal degradation, thereby crippling erythroid differentiation programs.
3.3 Immunomodulatory amplification of erythropoietic suppression
Beyond direct cytotoxicity, Hz indirectly exacerbates anemia by stimulating pro-inflammatory cytokine production (Shio et al., 2010). Notably, natural Hz (containing adsorbed parasite DNA and proteins) usually exhibits stronger immunomodulatory activity than synthetic Hz due to its parasite-derived nucleic acids and protein complexes. Evidence shows that natural Hz can synergistically induces robust secretion of IL-6, TNF-α, and IFN-γ from immune cells (Martiney et al., 2000; Jaramillo et al., 2004; Thawani et al., 2014; Banesh et al., 2022). In contrast, synthetic Hz lacking parasite-derived components exhibits minimal immunogenicity (Thawani et al., 2014). Incidentally, it is suggested that the inflammation status induced by natural Hz is likely to be mediated by activating Toll like receptor 9 (TLR9) (Parroche et al., 2007). Recent studies using human iPSC-derived cerebral malaria models further demonstrate that natural Hz enhances secretion of IFN-γ, IL-1β, IL-8, and IL-16, collectively exacerbating inflammatory anemia (Pranty et al., 2024).
4 Direct parasite infection and transcriptional reprogramming
Invasion of bone-marrow erythroid precursors by malaria parasites has emerged as a critical intra-medullary driver of malarial anemia. Using ex-vivo cultures, Tamez and colleagues first showed that the parasite preferentially infects orthochromatic erythroblasts, whereas polychromatic erythroblasts exhibit only sporadic, low-level invasion (Tamez et al., 2009). Microarray profiling revealed that infected orthochromatic cells display altered expression of 609 genes—570 of which are up-regulated—with significant enrichment of NRF2-mediated oxidative-stress pathways and heat-shock proteins (Tamez et al., 2011). Subsequent RNA-seq analyses corroborated the activation of oxidative-stress and mitochondrial-dysfunction pathways, indicating that the parasite re-programmes host metabolism and stress responses to support its own development (Feldman et al., 2023). Co-culture experiments further demonstrated that polychromatic erythroblasts exposed to malaria parasites up-regulate GDF15 (Tamez et al., 2011), a cytokine known to suppress hepcidin and thereby exacerbate ineffective erythropoiesis (Tanno et al., 2007; Guimaraes et al., 2015). Systematic autopsy studies employing dual immunohistochemistry for pLDH and Pfs16 identified the extravascular niche of the bone marrow as the principal site for immature gametocyte sequestration and maturation, with gametocyte fractions reaching 44.9%—far exceeding those in brain (4.8%) or spleen (1.3%) (Joice et al., 2014). Neveu and co-workers (Neveu et al., 2020) subsequently demonstrated that immature gametocytes require approximately ten days to complete stages I–IV within erythroblasts, markedly surpassing the host’s normal enucleation timeline and thus delaying terminal differentiation. Moreover, parasite-infected erythroblasts release extracellular vesicles that elicit oxidative stress and inhibit reticulocyte release (Neveu et al., 2020; Ben Ami Pilo et al., 2022). Collectively, malaria infection establishes a “developmental sanctuary” within the bone marrow by directly infecting terminally committed erythroblasts and deploying paracrine modulators; through transcriptional reprogramming, oxidative-stress signaling is activated and the cell cycle is arrested, thereby disrupting erythropoiesis. Notably, Plasmodium infection also profoundly remodels the expression profile of erythroid blood group antigens (Liu and Li, 2024). The EMP3 antigen (Blood Group System 41), whose normal expression is critical for enucleation during terminal erythropoiesis (Thornton et al., 2020), shows aberrant regulation following infection. This dysregulation likely contributes to observed enucleation defects in malaria patients, evidenced by morphological abnormalities in blood smears such as nuclear budding, inter-nuclear bridges, and multinucleated erythroblasts (Brito et al., 2022). This phenomenon may partially explain the impaired reticulocyte production characteristic of malarial anemia.
5 Host- and context-dependent heterogeneity in malaria-associated dyserythropoiesis
It should be noted that the mechanisms underlying malaria-induced anemia differ according to host age, nutritional status and clinical contexts, with the relative contribution of each factor mentioned above varying markedly.
In malaria-endemic regions, children under five years of age are likely to experience dyserythropoiesis driven chiefly by inflammatory cytokines such as IL-6, TNF-α and IFN-γ. Their still-maturing immune systems mount vigorous yet poorly regulated pro-inflammatory cascades (Simon et al., 2015) that rapidly suppress erythropoietin synthesis and disrupt the stability of the “master” erythroid transcription factor GATA-1. Moreover, compared with adults, these children possess fewer memory lymphocytes, are more susceptible to infection, and rely more heavily on innate immune responses during the initial phase of infection. A recent study of Malian children corroborates this hierarchy: in 1–5-year-olds, gene expression signatures are dominated by type I interferon, TLR and NLR innate pathways, with minimal T-cell memory signatures, and the proportion of neutrophils rises steeply with parasite density (Tebben et al., 2024). By contrast, older children and adults subjected to repeated low-density parasitemias accumulate hemozoin within splenic and marrow macrophages; persistent oxidative stress then fuels the progressive build-up of lipid-peroxidation products such as 4-HNE, which potently inhibit GATA-1. Consequently, anemia can persist long after peripheral parasitemia has been cleared.
When the host is chronically undernourished, deficiencies in iron, folate or vitamin B12 directly limit hemoglobin synthesis and erythroblast proliferation (Kassebaum et al., 2014). Concurrently, the acute inflammatory response in malaria rapidly elevates hepcidin levels, blocking intestinal iron absorption and macrophage iron recycling (Camaschella, 2019), thereby exacerbating functional iron deficiency and preventing effective erythroid expansion even under EPO stimulation. Moreover, hemoglobinopathies such as sickle cell disease (SCD) or α/β-thalassemia not only alter red-cell susceptibility to Plasmodium but also modify the marrow’s “starting burden” and “proliferative ceiling.” (Lelliott et al., 2015) When superimposed on the marrow-suppressive signals of malaria, these conditions can rapidly progress to transfusion-dependent severe anemia (Henrici et al., 2021; Uyoga et al., 2022).
The above factors, such as malnutrition, α/β-thalassemia and SCD, are all distinctly distributed geographically. The Mediterranean-Middle East-South Asia belt is dominated by thalassemia (Tuo et al., 2024), whereas sub-Saharan Africa has the highest prevalence of SCD (Collaborators GBDSCD, 2023). The degree of protection provided by these haemoglobinopathies and their genotypes (homozygous and heterozygous) differs substantially (Taylor et al., 2012). Additionally, the geographic distribution of plasmodium species also differs: Plasmodium falciparum predominates in sub-Saharan Africa and causes earlier and more severe anemia, while Plasmodium vivax is widespread across Asia-Pacific and the Americas and is associated with comparatively milder anemia (Weiss et al., 2025). Therefore, only by integrating the local composite etiologic profile and the prevailing parasite species can precise bone-marrow-targeted interventions be designed.
6 Future perspectives
The mechanisms underlying malaria-induced dyserythropoiesis are summarized in Figure 1. Despite significant progress in elucidating the pathogenesis of malarial anemia, critical scientific questions remain unresolved. Integration of single-cell technologies with spatial transcriptomics, combined with artificial intelligence-driven multi-omics analysis (e.g., deep learning), holds promise to elucidate the spatiotemporal dynamics of erythropoietic blockade. Spatial transcriptomics preserves tissue spatial conformation, overcoming the technical limitations of single-cell RNA sequencing by enabling single-cell-resolution mapping of parasite-induced transcriptional gradients in the bone marrow microenvironment. Furthermore, the integration of three-dimensional bone marrow organoids with intravital imaging techniques may enable real-time observation of parasite colonization dynamics and host-pathogen interactions within the hematopoietic niche. With advancements in biotechnology, the application of these novel methods and experimental approaches enables further exploration of metabolic dormancy mechanisms in the bone marrow microenvironment and bidirectional signaling between parasites and erythroblasts or other stromal cells. This will deepen our understanding of erythropoietic dysfunction and anemia in malaria. Additionally, ligand-trap agents such as luspatercept (TGF-β/activin inhibition, facilitating late-stage erythroid differentiation) (Martinez et al., 2020; Santini and Consagra, 2025), BRAF inhibitors that treat anemia through activation of MAPK signaling (Wu et al., 2024), and low-cost host-defense peptides with anti-inflammatory properties (e.g., IDR-1018) (Achtman et al., 2012) warrant systematic evaluation as adjunctive therapies for malaria-associated anemia, given their established erythropoietic or anti-inflammatory efficacy in other contexts; such studies may yield unforeseen clinical benefits.
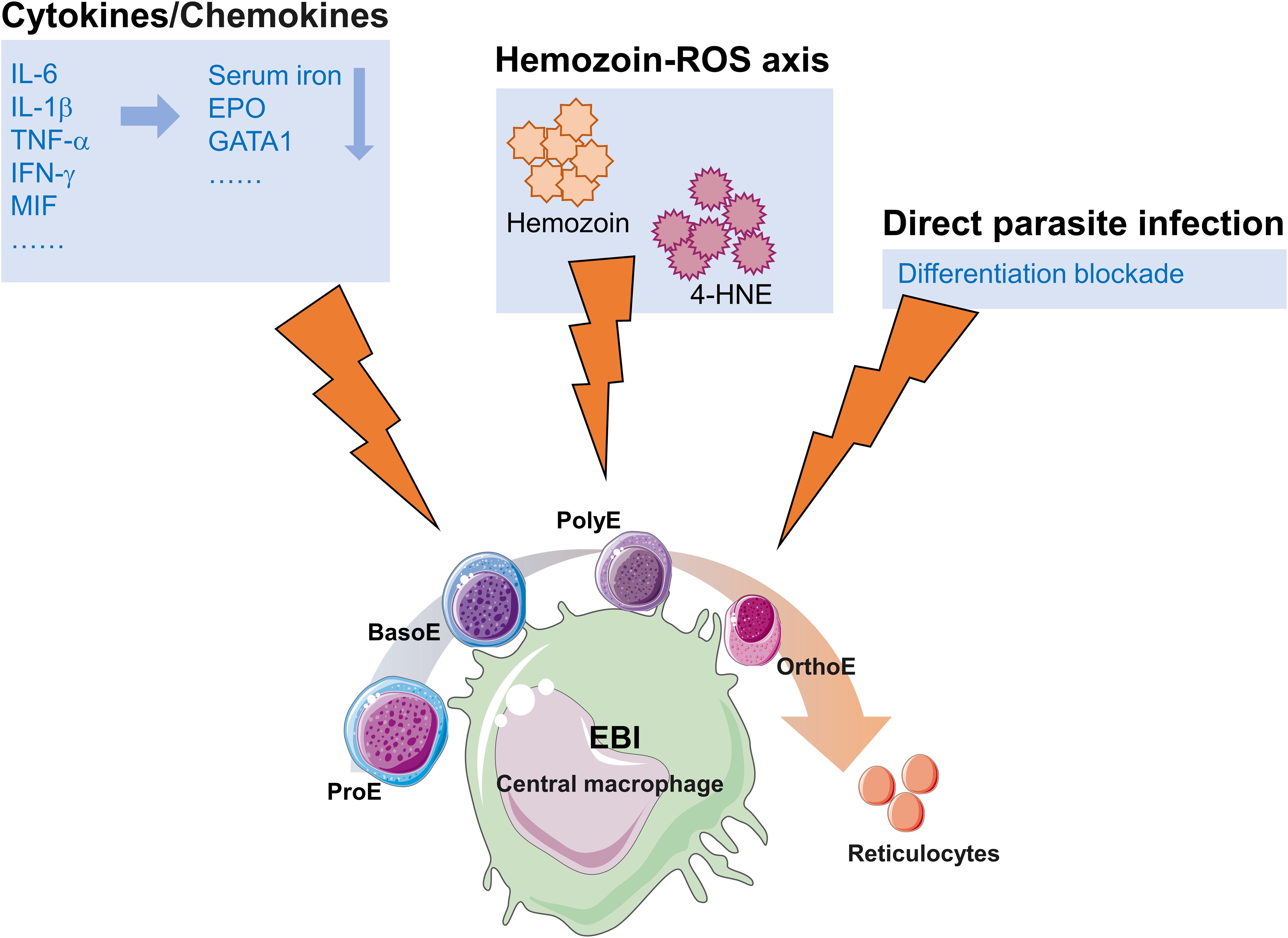
Figure 1. Schematic summary of malaria-induced dyserythropoiesis. EBI, The erythroblastic island; ProE, proerythroblast; BasoE, basophilic erythroblast; PolyE, polychromatophilic erythroblast; OrthoE, orthochromatic erythroblast.
Author contributions
F-FL: Investigation, Writing – original draft. KL: Funding acquisition, Investigation, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This study was partially supported by grants from the National Natural Science Foundation of China (Grant No. 81901103).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Achtman, A. H., Pilat, S., Law, C. W., Lynn, D. J., Janot, L., Mayer, M. L., et al. (2012). Effective adjunctive therapy by an innate defense regulatory peptide in a preclinical model of severe malaria. Sci. Transl. Med. 4, 135ra64. doi: 10.1126/scitranslmed.3003515
Ackerman, H., Ayestaran, A., Olola, C. H. O., Jallow, M., Agbenyega, T., Bojang, K., et al. (2020). The effect of blood transfusion on outcomes among African children admitted to hospital with Plasmodium falciparum malaria: a prospective, multicentre observational study. Lancet Haematol. 7, e789–ee97. doi: 10.1016/S2352-3026(20)30288-X
Aguilar, R., Moraleda, C., Achtman, A. H., Mayor, A., Quinto, L., Cistero, P., et al. (2014). Severity of anaemia is associated with bone marrow haemozoin in children exposed to Plasmodium falciparum. Br. J. Haematol. 164, 877–887. doi: 10.1111/bjh.2014.164.issue-6
Aubouy, A., Deloron, P., and Migot-Nabias, F. (2002). Plasma and in vitro levels of cytokines during and after a Plasmodium falciparum malaria attack in Gabon. Acta Trop. 83, 195–203. doi: 10.1016/S0001-706X(02)00094-3
Baird, J. K. (2022). African Plasmodium vivax malaria improbably rare or benign. Trends Parasitol. 38, 683–696. doi: 10.1016/j.pt.2022.05.006
Banesh, S., Layek, S., and Trivedi, V. (2022). Hemin acts as CD36 ligand to activate down-stream signalling to disturb immune responses and cytokine secretion from macrophages. Immunol. Lett. 243, 1–18. doi: 10.1016/j.imlet.2022.01.004
Baro, B., Deroost, K., Raiol, T., Brito, M., Almeida, A. C., de Menezes-Neto, A., et al. (2017). Plasmodium vivax gametocytes in the bone marrow of an acute malaria patient and changes in the erythroid miRNA profile. PloS Negl. Trop. Dis. 11, e0005365. doi: 10.1371/journal.pntd.0005365
Barrera, V., Skorokhod, O. A., Baci, D., Gremo, G., Arese, P., and Schwarzer, E. (2011). Host fibrinogen stably bound to hemozoin rapidly activates monocytes via TLR-4 and CD11b/CD18-integrin: a new paradigm of hemozoin action. Blood. 117, 5674–5682. doi: 10.1182/blood-2010-10-312413
Ben Ami Pilo, H., Khan Khilji, S., Luhle, J., Biskup, K., Levy Gal, B., Rosenhek Goldian, I., et al. (2022). Sialylated N-glycans mediate monocyte uptake of extracellular vesicles secreted from Plasmodium falciparum-infected red blood cells. J. Extracell Biol. 1, e33. doi: 10.1002/jex2.v1.2
Brito, M. A. M., Baro, B., Raiol, T. C., Ayllon-Hermida, A., Safe, I. P., Deroost, K., et al. (2022). Morphological and transcriptional changes in human bone marrow during natural plasmodium vivax malaria infections. J. Infect. Dis. 225, 1274–1283. doi: 10.1093/infdis/jiaa177
Cambos, M., Bazinet, S., Abed, E., Sanchez-Dardon, J., Bernard, C., Moreau, R., et al. (2010). The IL-12p70/IL-10 interplay is differentially regulated by free heme and hemozoin in murine bone-marrow-derived macrophages. Int. J. Parasitol. 40, 1003–1012. doi: 10.1016/j.ijpara.2010.02.007
Canna, S. W., Wrobel, J., Chu, N., Kreiger, P. A., Paessler, M., and Behrens, E. M. (2013). Interferon-gamma mediates anemia but is dispensable for fulminant toll-like receptor 9-induced macrophage activation syndrome and hemophagocytosis in mice. Arthritis Rheumatol. 65, 1764–1775. doi: 10.1002/art.37958
Casals-Pascual, C., Kai, O., Cheung, J. O., Williams, S., Lowe, B., Nyanoti, M., et al. (2006). Suppression of erythropoiesis in malarial anemia is associated with hemozoin in vitro and in vivo. Blood 108, 2569–2577. doi: 10.1182/blood-2006-05-018697
Chaiyaroj, S. C., Rutta, A. S., Muenthaisong, K., Watkins, P., Na Ubol, M., and Looareesuwan, S. (2004). Reduced levels of transforming growth factor-beta1, interleukin-12 and increased migration inhibitory factor are associated with severe malaria. Acta Trop. 89, 319–327. doi: 10.1016/j.actatropica.2003.10.010
Collaborators GBDSCD (2023). Global, regional, and national prevalence and mortality burden of sickle cell disease, 2000-2021: a systematic analysis from the Global Burden of Disease Study 2021. Lancet Haematol. 10, e585–ee99. doi: 10.1016/S2352-3026(23)00118-7
Degarege, A., Gebrezgi, M. T., Ibanez, G., Wahlgren, M., and Madhivanan, P. (2019). Effect of the ABO blood group on susceptibility to severe malaria: A systematic review and meta-analysis. Blood Rev. 33, 53–62. doi: 10.1016/j.blre.2018.07.002
De Niz, M., Meibalan, E., Mejia, P., Ma, S., Brancucci, N. M. B., Agop-Nersesian, C., et al. (2018). Plasmodium gametocytes display homing and vascular transmigration in the host bone marrow. Sci. Adv. 4, eaat3775. doi: 10.1126/sciadv.aat3775
Dondorp, A. M., Angus, B. J., Chotivanich, K., Silamut, K., Ruangveerayuth, R., Hardeman, M. R., et al. (1999). Red blood cell deformability as a predictor of anemia in severe falciparum malaria. Am. J. Trop. Med. Hyg. 60, 733–737. doi: 10.4269/ajtmh.1999.60.733
Dumarchey, A., Lavazec, C., and Verdier, F. (2022). Erythropoiesis and malaria, a multifaceted interplay. Int. J. Mol. Sci. 23, 12762. doi: 10.3390/ijms232112762
Eastman, R. T. and Fidock, D. A. (2009). Artemisinin-based combination therapies: a vital tool in efforts to eliminate malaria. Nat. Rev. Microbiol. 7, 864–874. doi: 10.1038/nrmicro2239
Feldman, T. P., Ryan, Y., and Egan, E. S. (2023). Plasmodium falciparum infection of human erythroblasts induces transcriptional changes associated with dyserythropoiesis. Blood Adv. 7, 5496–5509. doi: 10.1182/bloodadvances.2023010844
Felli, N., Pedini, F., Zeuner, A., Petrucci, E., Testa, U., Conticello, C., et al. (2005). Multiple members of the TNF superfamily contribute to IFN-gamma-mediated inhibition of erythropoiesis. J. Immunol. 175, 1464–1472. doi: 10.4049/jimmunol.175.3.1464
Fernandez-Arias, C., Rivera-Correa, J., Gallego-Delgado, J., Rudlaff, R., Fernandez, C., Roussel, C., et al. (2016). Anti-self phosphatidylserine antibodies recognize uninfected erythrocytes promoting malarial anemia. Cell Host Microbe 19, 194–203. doi: 10.1016/j.chom.2016.01.009
Ghosh, S., Jiang, N., Farr, L., Ngobeni, R., and Moonah, S. (2019). Parasite-produced MIF cytokine: role in immune evasion, invasion, and pathogenesis. Front. Immunol. 10, 1995. doi: 10.3389/fimmu.2019.01995
Giribaldi, G., Ulliers, D., Schwarzer, E., Roberts, I., Piacibello, W., and Arese, P. (2004). Hemozoin- and 4-hydroxynonenal-mediated inhibition of erythropoiesis. Possible role in malarial dyserythropoiesis and anemia. Haematologica. 89, 492–493.
Gowda, D. C. and Wu, X. (2018). Parasite recognition and signaling mechanisms in innate immune responses to malaria. Front. Immunol. 9, 3006. doi: 10.3389/fimmu.2018.03006
Guimaraes, J. S., Cominal, J. G., Silva-Pinto, A. C., Olbina, G., Ginzburg, Y. Z., Nandi, V., et al. (2015). Altered erythropoiesis and iron metabolism in carriers of thalassemia. Eur. J. Haematol. 94, 511–518. doi: 10.1111/ejh.2015.94.issue-6
Henrici, R. C., Sautter, C. L., Bond, C., Opoka, R. O., Namazzi, R., Datta, D., et al. (2021). Decreased parasite burden and altered host response in children with sickle cell anemia and severe anemia with malaria. Blood Adv. 5, 4710–4720. doi: 10.1182/bloodadvances.2021004704
Jaramillo, M., Gowda, D. C., Radzioch, D., and Olivier, M. (2003). Hemozoin increases IFN-gamma-inducible macrophage nitric oxide generation through extracellular signal-regulated kinase- and NF-kappa B-dependent pathways. J. Immunol. 171, 4243–4253. doi: 10.4049/jimmunol.171.8.4243
Jaramillo, M., Plante, I., Ouellet, N., Vandal, K., Tessier, P. A., and Olivier, M. (2004). Hemozoin-inducible proinflammatory events in vivo: potential role in malaria infection. J. Immunol. 172, 3101–3110. doi: 10.4049/jimmunol.172.5.3101
Jelkmann, W. (1998). Proinflammatory cytokines lowering erythropoietin production. J. Interferon Cytokine Res. 18, 555–559. doi: 10.1089/jir.1998.18.555
Joice, R., Nilsson, S. K., Montgomery, J., Dankwa, S., Egan, E., Morahan, B., et al. (2014). Plasmodium falciparum transmission stages accumulate in the human bone marrow. Sci. Transl. Med. 6, 244re5. doi: 10.1126/scitranslmed.3008882
Kassebaum, N. J., Jasrasaria, R., Naghavi, M., Wulf, S. K., Johns, N., Lozano, R., et al. (2014). A systematic analysis of global anemia burden from 1990 to 2010. Blood. 123, 615–624. doi: 10.1182/blood-2013-06-508325
Kotepui, M., Kotepui, K., Mahittikorn, A., Majima, H. J., Tangpong, J., and Yen, H. C. (2023). Association of reduced glutathione levels with Plasmodium falciparum and Plasmodium vivax malaria: a systematic review and meta-analysis. Sci. Rep. 13, 16483. doi: 10.1038/s41598-023-43583-z
Kremsner, P. G., Winkler, S., Brandts, C., Wildling, E., Jenne, L., Graninger, W., et al. (1995). Prediction of accelerated cure in Plasmodium falciparum malaria by the elevated capacity of tumor necrosis factor production. Am. J. Trop. Med. Hyg. 53, 532–538. doi: 10.4269/ajtmh.1995.53.532
Lamikanra, A. A., Theron, M., Kooij, T. W., and Roberts, D. J. (2009). Hemozoin (malarial pigment) directly promotes apoptosis of erythroid precursors. PloS One 4, e8446. doi: 10.1371/journal.pone.0008446
Lelliott, P. M., McMorran, B. J., Foote, S. J., and Burgio, G. (2015). The influence of host genetics on erythrocytes and malaria infection: is there therapeutic potential? Malar J. 14, 289. doi: 10.1186/s12936-015-0809-x
Libregts, S. F., Gutierrez, L., de Bruin, A. M., Wensveen, F. M., Papadopoulos, P., van Ijcken, W., et al. (2011). Chronic IFN-gamma production in mice induces anemia by reducing erythrocyte life span and inhibiting erythropoiesis through an IRF-1/PU. 1 axis. Blood 118, 2578–2588. doi: 10.1182/blood-2010-10-315218
Liu, F. F. and Li, K. (2024). Expression alterations of blood group genes during plasmodium falciparum infection in orthochromatic erythroblasts. Transfus Med. Rev. 38, 150837. doi: 10.1016/j.tmrv.2024.150837
Looareesuwan, S., Davis, T. M., Pukrittayakamee, S., Supanaranond, W., Desakorn, V., Silamut, K., et al. (1991). Erythrocyte survival in severe falciparum malaria. Acta Trop. 48, 263–270. doi: 10.1016/0001-706X(91)90014-B
Lyke, K. E., Burges, R., Cissoko, Y., Sangare, L., Dao, M., Diarra, I., et al. (2004). Serum levels of the proinflammatory cytokines interleukin-1 beta (IL-1beta), IL-6, IL-8, IL-10, tumor necrosis factor alpha, and IL-12(p70) in Malian children with severe Plasmodium falciparum malaria and matched uncomplicated malaria or healthy controls. Infect. Immun. 72, 5630–5637. doi: 10.1128/IAI.72.10.5630-5637.2004
Martiney, J. A., Sherry, B., Metz, C. N., Espinoza, M., Ferrer, A. S., Calandra, T., et al. (2000). Macrophage migration inhibitory factor release by macrophages after ingestion of Plasmodium chabaudi-infected erythrocytes: possible role in the pathogenesis of malarial anemia. Infect. Immun. 68, 2259–2267. doi: 10.1128/IAI.68.4.2259-2267.2000
Martinez, P. A., Li, R., Ramanathan, H. N., Bhasin, M., Pearsall, R. S., Kumar, R., et al. (2020). Smad2/3-pathway ligand trap luspatercept enhances erythroid differentiation in murine beta-thalassaemia by increasing GATA-1 availability. J. Cell. Mol. Med. 24, 6162–6177. doi: 10.1111/jcmm.v24.11
McDevitt, M. A., Xie, J., Ganapathy-Kanniappan, S., Griffith, J., Liu, A., McDonald, C., et al. (2006). A critical role for the host mediator macrophage migration inhibitory factor in the pathogenesis of malarial anemia. J. Exp. Med. 203, 1185–1196. doi: 10.1084/jem.20052398
McDevitt, M. A., Xie, J., Ganapathy-Kanniappan, S., Griffith, J., Liu, A., McDonald, C., et al. (2015). A critical role for the host mediator macrophage migration inhibitory factor in the pathogenesis of malarial anemia. J. Exp. Med. 212, 825. doi: 10.1084/jem.2005239804172015c
Miller, C. M., Carney, C. K., Schrimpe, A. C., and Wright, D. W. (2005). beta-Hematin (hemozoin) mediated decompostion of polyunsaturated fatty acids to 4-hydroxy-2-nonenal. Inorg Chem. 44, 2134–2136. doi: 10.1021/ic048821i
Miller, K. L., Schooley, J. C., Smith, K. L., Kullgren, B., Mahlmann, L. J., and Silverman, P. H. (1989). Inhibition of erythropoiesis by a soluble factor in murine malaria. Exp. Hematol. 17, 379–385.
Ndour, P. A., Larreche, S., Mouri, O., Argy, N., Gay, F., Roussel, C., et al. (2017). Measuring the Plasmodium falciparum HRP2 protein in blood from artesunate-treated malaria patients predicts post-artesunate delayed hemolysis. Sci. Transl. Med. 9, eaaf9377. doi: 10.1126/scitranslmed.aaf9377
Neveu, G., Richard, C., Dupuy, F., Behera, P., Volpe, F., Subramani, P. A., et al. (2020). Plasmodium falciparum sexual parasites develop in human erythroblasts and affect erythropoiesis. Blood. 136, 1381–1393. doi: 10.1182/blood.2019004746
Obeagu, E. I. (2024). Role of cytokines in immunomodulation during malaria clearance. Ann. Med. Surg. (Lond). 86, 2873–2882. doi: 10.1097/MS9.0000000000002019
Parroche, P., Lauw, F. N., Goutagny, N., Latz, E., Monks, B. G., Visintin, A., et al. (2007). Malaria hemozoin is immunologically inert but radically enhances innate responses by presenting malaria DNA to Toll-like receptor 9. Proc. Natl. Acad. Sci. U S A. 104, 1919–1924. doi: 10.1073/pnas.0608745104
Pascual, V., Allantaz, F., Arce, E., Punaro, M., and Banchereau, J. (2005). Role of interleukin-1 (IL-1) in the pathogenesis of systemic onset juvenile idiopathic arthritis and clinical response to IL-1 blockade. J. Exp. Med. 201, 1479–1486. doi: 10.1084/jem.20050473
Pathak, V. A. and Ghosh, K. (2016). Erythropoiesis in malaria infections and factors modifying the erythropoietic response. Anemia. 2016, 9310905. doi: 10.1155/2016/9310905
Penha-Goncalves, C. (2019). Genetics of malaria inflammatory responses: A pathogenesis perspective. Front. Immunol. 10, 1771. doi: 10.3389/fimmu.2019.01771
Pied, S., Civas, A., Berlot-Picard, F., Renia, L., Miltgen, F., Gentilini, M., et al. (1992). IL-6 induced by IL-1 inhibits malaria pre-erythrocytic stages but its secretion is down-regulated by the parasite. J. Immunol. 148, 197–201. doi: 10.4049/jimmunol.148.1.197
Pietras, E. M., Mirantes-Barbeito, C., Fong, S., Loeffler, D., Kovtonyuk, L. V., Zhang, S., et al. (2016). Chronic interleukin-1 exposure drives haematopoietic stem cells towards precocious myeloid differentiation at the expense of self-renewal. Nat. Cell Biol. 18, 607–618. doi: 10.1038/ncb3346
Pranty, A. I., Szepanowski, L. P., Wruck, W., Karikari, A. A., and Adjaye, J. (2024). Hemozoin induces malaria via activation of DNA damage, p38 MAPK and neurodegenerative pathways in a human iPSC-derived neuronal model of cerebral malaria. Sci. Rep. 14, 24959. doi: 10.1038/s41598-024-76259-3
Rockett, K. A., Awburn, M. M., Rockett, E. J., and Clark, I. A. (1994). Tumor necrosis factor and interleukin-1 synergy in the context of malaria pathology. Am. J. Trop. Med. Hyg. 50, 735–742. doi: 10.4269/ajtmh.1994.50.735
Santini, V. and Consagra, A. (2025). How to use luspatercept and erythropoiesis-stimulating agents in low-risk myelodysplastic syndrome. Br. J. Haematol. 207, 15–26. doi: 10.1111/bjh.v207.1
Schwarzer, E., Kuhn, H., Valente, E., and Arese, P. (2003). Malaria-parasitized erythrocytes and hemozoin nonenzymatically generate large amounts of hydroxy fatty acids that inhibit monocyte functions. Blood. 101, 722–728. doi: 10.1182/blood-2002-03-0979
Shio, M. T., Eisenbarth, S. C., Savaria, M., Vinet, A. F., Bellemare, M. J., Harder, K. W., et al. (2009). Malarial hemozoin activates the NLRP3 inflammasome through Lyn and Syk kinases. PloS Pathog. 5, e1000559. doi: 10.1371/journal.ppat.1000559
Shio, M. T., Kassa, F. A., Bellemare, M. J., and Olivier, M. (2010). Innate inflammatory response to the malarial pigment hemozoin. Microbes Infect. 12, 889–899. doi: 10.1016/j.micinf.2010.07.001
Silva-Filho, J. L., Lacerda, M. V. G., Recker, M., Wassmer, S. C., Marti, M., and Costa, F. T. M. (2020). Plasmodium vivax in hematopoietic niches: hidden and dangerous. Trends Parasitol. 36, 447–458. doi: 10.1016/j.pt.2020.03.002
Simon, A. K., Hollander, G. A., and McMichael, A. (2015). Evolution of the immune system in humans from infancy to old age. Proc. Biol. Sci. 282, 20143085. doi: 10.1098/rspb.2014.3085
Skorokhod, O. A., Caione, L., Marrocco, T., Migliardi, G., Barrera, V., Arese, P., et al. (2010). Inhibition of erythropoiesis in malaria anemia: role of hemozoin and hemozoin-generated 4-hydroxynonenal. Blood. 116, 4328–4337. doi: 10.1182/blood-2010-03-272781
Skorokhod, O., Schwarzer, E., Grune, T., and Arese, P. (2005). Role of 4-hydroxynonenal in the hemozoin-mediated inhibition of differentiation of human monocytes to dendritic cells induced by GM-CSF/IL-4. Biofactors 24, 283–289. doi: 10.1002/biof.5520240133
Tamez, P. A., Liu, H., Fernandez-Pol, S., Haldar, K., and Wickrema, A. (2009). Stage-specific susceptibility of human erythroblasts to Plasmodium falciparum malaria infection. Blood. 114, 3652–3655. doi: 10.1182/blood-2009-07-231894
Tamez, P. A., Liu, H., Wickrema, A., and Haldar, K. P. (2011). falciparum modulates erythroblast cell gene expression in signaling and erythrocyte production pathways. PloS One 6, e19307. doi: 10.1371/journal.pone.0019307
Tanno, T., Bhanu, N. V., Oneal, P. A., Goh, S. H., Staker, P., Lee, Y. T., et al. (2007). High levels of GDF15 in thalassemia suppress expression of the iron regulatory protein hepcidin. Nat. Med. 13, 1096–1101. doi: 10.1038/nm1629
Taylor, S. M., Parobek, C. M., and Fairhurst, R. M. (2012). Haemoglobinopathies and the clinical epidemiology of malaria: a systematic review and meta-analysis. Lancet Infect. Dis. 12, 457–468. doi: 10.1016/S1473-3099(12)70055-5
Tebben, K., Yirampo, S., Coulibaly, D., Kone, A. K., Laurens, M. B., Stucke, E. M., et al. (2024). Gene expression analyses reveal differences in children's response to malaria according to their age. Nat. Commun. 15, 2021. doi: 10.1038/s41467-024-46416-3
Thawani, N., Tam, M., Bellemare, M. J., Bohle, D. S., Olivier, M., de Souza, J. B., et al. (2014). Plasmodium products contribute to severe malarial anemia by inhibiting erythropoietin-induced proliferation of erythroid precursors. J. Infect. Dis. 209, 140–149. doi: 10.1093/infdis/jit417
Thornton, N., Karamatic Crew, V., Tilley, L., Green, C. A., Tay, C. L., Griffiths, R. E., et al. (2020). Disruption of the tumour-associated EMP3 enhances erythroid proliferation and causes the MAM-negative phenotype. Nat. Commun. 11, 3569. doi: 10.1038/s41467-020-17060-4
Tuo, Y., Li, Y., Li, Y., Ma, J., Yang, X., Wu, S., et al. (2024). Global, regional, and national burden of thalassemia, 1990-2021: a systematic analysis for the global burden of disease study 2021. EClinicalMedicine. 72, 102619. doi: 10.1016/j.eclinm.2024.102619
Tyrkalska, S. D., Perez-Oliva, A. B., Rodriguez-Ruiz, L., Martinez-Morcillo, F. J., Alcaraz-Perez, F., Martinez-Navarro, F. J., et al. (2019). Inflammasome regulates hematopoiesis through cleavage of the master erythroid transcription factor GATA1. Immunity. 51, 50–63 e5. doi: 10.1016/j.immuni.2019.05.005
Uyoga, S., Olupot-Olupot, P., Connon, R., Kiguli, S., Opoka, R. O., Alaroker, F., et al. (2022). Sickle cell anaemia and severe Plasmodium falciparum malaria: a secondary analysis of the Transfusion and Treatment of African Children Trial (TRACT). Lancet Child Adolesc. Health 6, 606–613. doi: 10.1016/S2352-4642(22)00153-5
Venkatesan, P. (2025). WHO world malaria report 2024. Lancet Microbe 101073. doi: 10.1016/j.lanmic.2025.101073
Weiss, D. J., Dzianach, P. A., Saddler, A., Lubinda, J., Browne, A., McPhail, M., et al. (2025). Mapping the global prevalence, incidence, and mortality of Plasmodium falciparum and Plasmodium vivax malaria, 2000-22: a spatial and temporal modelling study. Lancet. 405, 979–990. doi: 10.1016/S0140-6736(25)00038-8
Wrighting, D. M. and Andrews, N. C. (2006). Interleukin-6 induces hepcidin expression through STAT3. Blood. 108, 3204–3209. doi: 10.1182/blood-2006-06-027631
Wu, S., Deng, Y., Sun, H., Liu, X., Zhou, S., Zhao, H., et al. (2024). BRAF inhibitors enhance erythropoiesis and treat anemia through paradoxical activation of MAPK signaling. Signal Transduct Target Ther. 9, 338. doi: 10.1038/s41392-024-02033-6
Keywords: malaria, anemia, dyserythropoiesis, ineffective erythropoiesis, bone marrow niche, hemozoin, inflammatory cytokines
Citation: Liu F-F and Li K (2025) Malaria and dyserythropoiesis: a mini review. Front. Cell. Infect. Microbiol. 15:1679337. doi: 10.3389/fcimb.2025.1679337
Received: 04 August 2025; Accepted: 22 August 2025;
Published: 05 September 2025.
Edited by:
Myat Htut Nyunt, Department of Medical Research, MyanmarReviewed by:
Juan David Ospina-Villa, Colombian Institute of Tropical Medicine (ICMT), ColombiaCopyright © 2025 Liu and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ke Li, bGlrZWxmZkBoYWluYW51LmVkdS5jbg==