 Guangchao Charles Feng
Guangchao Charles Feng- School of Communication, Hong Kong Baptist University, Kowloon, Hong Kong SAR, China
Inoculation theory offers a unique perspective on how beliefs can be fortified against persuasive attacks. However, several aspects of the theory warrant further exploration and reinterpretation, particularly its analogies with immunology. The paper aims to broaden the theory’s boundary conditions by delving into thought-provoking analogies between immunology and persuasion. The author redefines multiple key concepts within inoculation theory and addresses a range of existing issues associated with classic inoculation theory, including the distinction between passive and active defense and addressing “infected” people and “allergy” with therapeutic inoculation. The paper concludes by proposing future research directions that include exploring diverse, especially mRNA forms of “inoculation” and formulating a series of conceptual propositions.
Introduction
The field of immunology has been a rich source offering metaphors, analogies, and conceptual frameworks that have been applied to understand and theorize persuasion. Analogous concepts such as “information contagion,” “social contagion,” “infodemics,” “Memes,” and “information immunity” have emerged to describe information and communication-related phenomena (Christakis and Fowler, 2013; Cinelli et al., 2020), mirroring the language of epidemiology. Several scholars have contributed to this area by examining the relationship between scientific paradigms, analogies, and the use of immunological metaphors (Alexejeva and Mishlanova, 1997; Black, 1962; Hesse, 1970). Communication theories, such as inoculation theory (McGuire, 1961a, 1961b; McGuire and Papageorgis, 1961), have capitalized on this “clever and valid” method of theorizing the connection between immunology and communication phenomena, serving as the springboard for further theoretical investigation (Compton and van der Linden, 2022; Compton and Pfau, 2005).
Inoculation theory posits that when individuals are exposed to attenuated counterarguments, they fortify their cognitive resilience against subsequent, more potent persuasion attempts, reinforcing their deeply ingrained convictions (Niederdeppe et al., 2015). Inoculation theory has garnered widespread recognition as one of the most prominent theories in persuasion research owing to its extensive application across diverse practical contexts (Ivanov et al., 2020, p. 13). However, despite its significant impact and wide-ranging applications, there remains ample room for further exploration to clarify the complex medical analogies upon which it rests. A noteworthy temporal correspondence can be observed between the birth of modern immunology, marked by the discovery of T lymphocytes’ role in cellular immunity in 1961 (Miller, 2002), and the concurrent publication of inoculation theory in the same year (McGuire, 1961a, 1961b; McGuire and Papageorgis, 1961). This synchronicity in timing suggests that the foundational conceptualization of inoculation theory was grounded in a heuristic appreciation of longstanding vaccination principles rather than a nuanced, comprehensive grasp of immunology that would only emerge and mature in subsequent years (Miller, 2002; Plotkin and Plotkin, 2018).
Biological origins and analogies
Mechanisms of immune system
Humans routinely encounter foreign agents—food proteins, environmental microbes, viral particles—each with its own virulence (capacity to cause disease). Whether exposure leads to illness depends on the agent’s virulence and the host’s immune strength (Kasuga et al., 2021). Vaccines exploit this balance by presenting non-pathogenic fragments of a pathogen (antigens) that safely train the immune system. Many formulations include an adjuvant—a compound that heightens the initial immune alert so that the body mounts a stronger, longer-lasting response to the vaccine antigen (Figure 1).
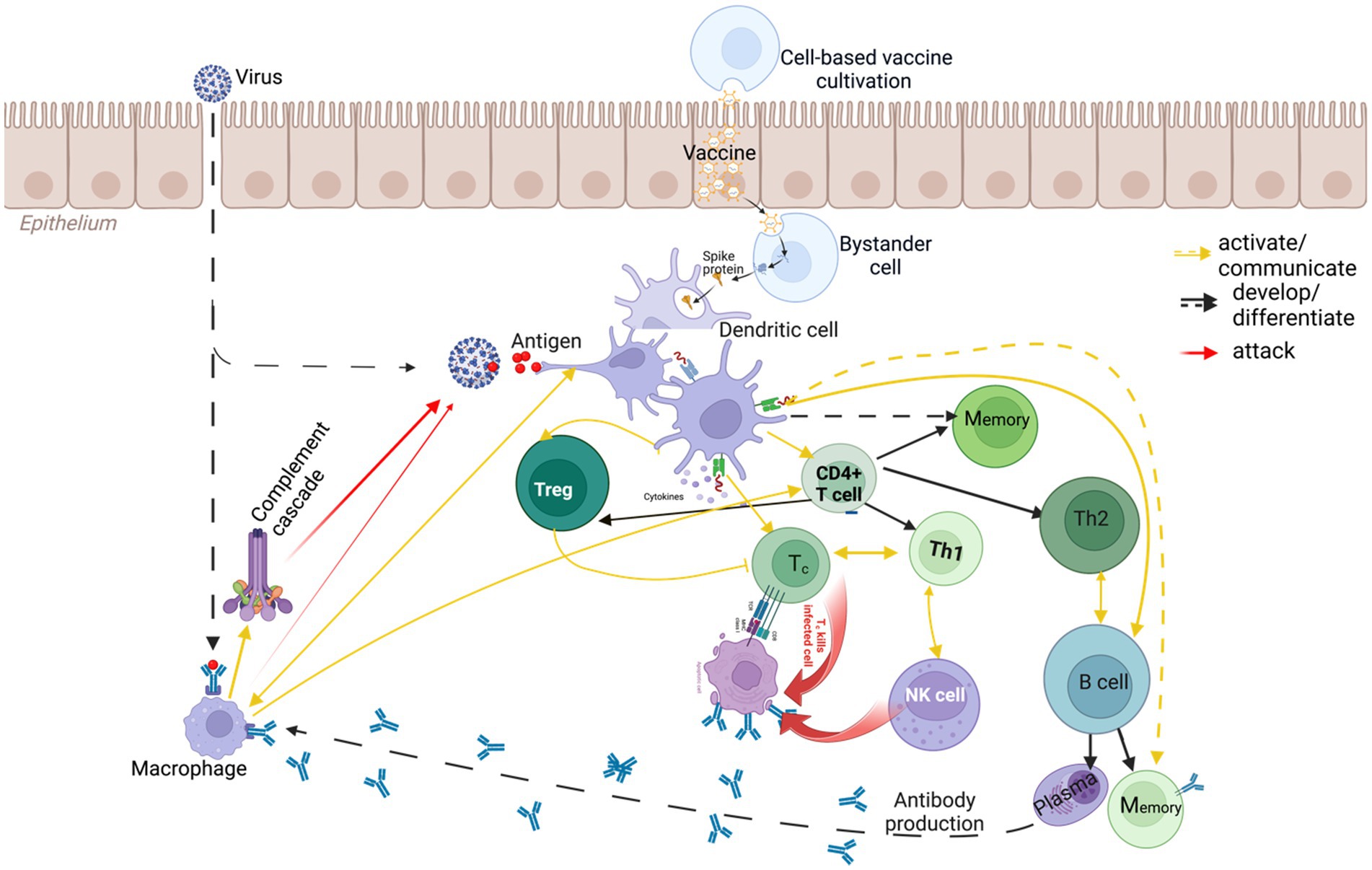
Figure 1. Mechanisms of immune system fighting viral infections.
Innate immunity is the fast, generalized shield. Within minutes or hours, innate cells recognize common danger patterns and release cytokines (short-range messenger proteins) that trigger inflammation and recruit more defenders. Adaptive immunity follows if the threat persists. It is slower but precise, relying on lymphocytes: B cells (which secrete antigen-specific antibodies) and T cells (which coordinate responses or destroy infected cells). First encounters are slow, but they generate memory cells that accelerate any future response (Table 1).
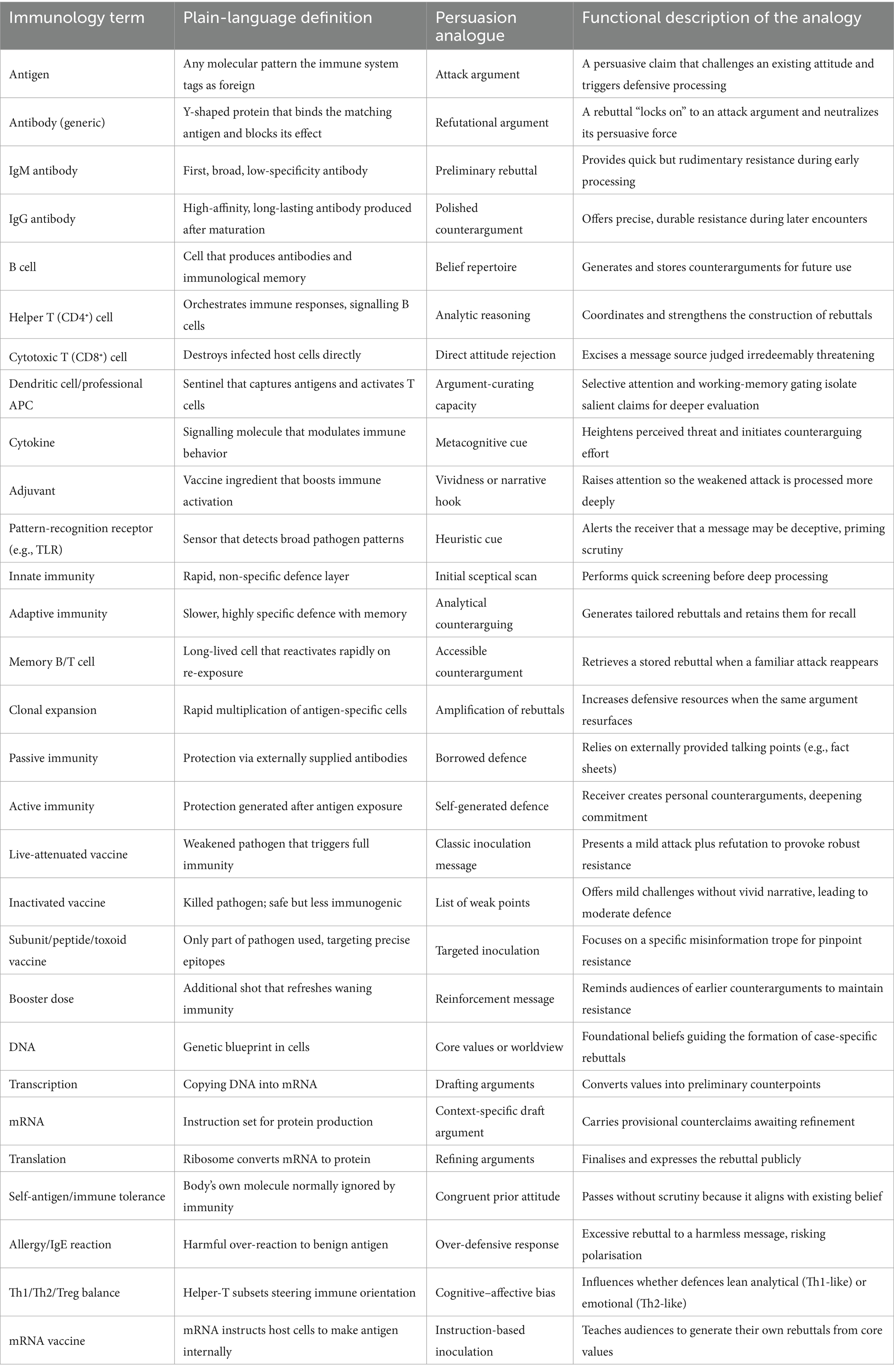
Table 1. Glossary of immunological terms and their persuasion analogues.
Vaccination primarily targets the adaptive branch. Introduced antigens activate helper T cells, which in turn stimulate B cells to produce antibodies and license cytotoxic T cells to eliminate infected targets. Adjuvants ensure that innate sensors, such as Toll-like receptors on dendritic cells, sound an early alarm, supplying cytokine signals that amplify adaptive learning (Banchereau and Steinman, 1998). The result is twofold: immediate neutralization of the harmless antigen and durable immunological memory.
Psychological inoculation theory mirrors this biological sequence. A forewarning parallels the innate alarm by heightening vigilance at the outset. A weakened persuasive attack accompanied by an explicit refutation offers the functional equivalent of antigen practice, allowing recipients to rehearse defensive reasoning in a safe context. Stored counterarguments then play the role of immunological memory; they wait in readiness until a stronger persuasive attack appears. Finally, vivid or credible cues embedded in the message operate like rhetorical adjuvants, ensuring that the initial warning captures attention and receives deeper processing. By emphasizing the temporal pattern—alert, rehearsal, retention—this analogy clarifies how timing and graded challenge cultivate durable resistance without resorting to one-to-one maps between cells and thoughts.
Extended biological analogies
Structure-mapping theory (Gentner, 1983; Gentner and Holyoak, 1997) teaches that an analogy is strongest when it transfers relational structure rather than superficial features. In immunology, a pathogen triggers a cascade: dendritic cells process and present antigens to helper T cells; helper T cells coordinate with B cells; B cells produce antibodies that neutralize the threat. The explanatory power resides in this ordered interaction, not in any single molecule.
A comparable cascade unfolds during psychological inoculation. Core values serve as a stable blueprint, analogous to DNA. When a persuasive threat arises, these values are first “transcribed” into draft reasons that are specific to the context, and then “translated” into fully articulated counterarguments (analogous to antibodies). As in biology, emphasis belongs on the relational dynamics connecting initial activation to eventual defense.
Within this framework, viruses or bacteria correspond to persuasive attacks such as misinformation. A vaccine, which pre-emptively introduces a benign fragment of the pathogen, resembles a preliminary persuasive message designed to stimulate defensive processing. In immunology, an antigen is simply a recognizable molecular signature; in persuasion, the specific argument occupies that role. An antibody, after binding its matching antigen, neutralizes the invader; correspondingly, a refutational argument neutralizes the persuasive claim. Self-antigens illustrate immune tolerance toward benign or beneficial entities, just as congenial information typically escapes critical scrutiny in cognitive processing (Liu et al., 2019; McFall-Ngai et al., 2013; Wood, 2007).
Certain exploratory analogies invite additional theoretical development. Core values and worldviews parallel DNA because they direct how individuals assemble arguments, much as genetic instructions direct protein synthesis. Cultural worldviews, for instance, influence climate-change attitudes more strongly than raw scientific knowledge does (Kahan et al., 2012). Dendritic cells map onto an internal argument-curating capacity composed of selective attention and working-memory gating. These cognitive mechanisms sample salient elements in a message, condense them, and pass them to analytic reasoning—functional behavior that mirrors how dendritic cells deliver processed antigens to T cells. Analytic reasoning itself aligns with helper T cells because it coordinates evaluation and orchestrates the generation of rebuttals. The belief repertoire aligns with B cells because it manufactures specific counterarguments that address the identified claim (Compton and van der Linden, 2022).
Presenting the analogy at this relational level preserves conceptual coherence: each immune component finds its counterpart within a single cognitive system—attention, reasoning, and stored beliefs—rather than oscillating between individual cognition and social-level gatekeeping. The goal remains clarity, not literal equivalence, and the resulting framework highlights measurable stages—alertness, rehearsal quality, memory durability—open to empirical test within communication research.
Review of previous extensions of inoculation theory
Role of affect
Affect—encompassing emotions and moods—has long been a potential but debated factor in inoculation theory. Although much of inoculation research has historically focused on cognitive processes, several studies have explicitly manipulated affective content in inoculation treatments. Notably, factorial experiments have already been conducted to test how affect interacts with inoculation. For example, Pfau et al. (2001) designed inoculation messages to evoke specific emotions (anger-based vs. happiness-based appeals, compared to a standard cognitive message). Intriguingly, their results showed that the inoculation message framed to induce happiness produced the greatest resistance to a later persuasive attack, even more than the anger-framed or purely cognitive messages. This was unexpected—the anger-based inoculation created higher perceived threat (a core cognitive mechanism of inoculation) than the happiness-based message, yet it was the positive-affect (happy) condition that ultimately yielded the strongest persuasive immunity. Pfau et al. (2001) acknowledged they had no obvious theoretical explanation at the time for why a happiness-inducing inoculation was so effective, suggesting that reassurance or a broader cognitive engagement from positive affect might have played a role.
Years later, Pfau et al. (2009) revisited affect in inoculation with a refined design. They compared affective-negative, affective-positive, and cognitive inoculation messages in a large experiment, while keeping other factors (like strength of supporting arguments) constant across conditions. Consistent with inoculation theory, all inoculation conditions conferred more resistance to persuasion than a no-inoculation control. However, when contrasting the treatments, there were some notable differences linked to affect. The affective-negative messages (aimed at inducing anger or fear) successfully elicited more negative emotion in recipients, whereas the affective-positive messages (aimed at inducing happiness) did not significantly elevate positive emotion beyond baseline. Moreover, inoculations that evoked anger/fear or that took a purely rational tone tended to engage participants more deeply: Pfau et al. (2009) found that the negative-emotion and cognitive conditions generated greater involvement, threat appraisal, and attitude certainty immediately after inoculation (and even after the subsequent attack) compared to the positive-emotion condition. In other words, participants receiving an anger-tinged or a strong logical inoculation message felt a bit more warned and involved than those receiving a happy/positive-toned inoculation. Interestingly, despite these process differences, all three message types were effective, and some emotional effects were fleeting. For instance, right after the inoculation message, those who received an affective treatment were slightly angrier and less happy than control participants, though these differences were not statistically significant; immediately post-inoculation, fear actually decreased among those who got an inoculation message (perhaps the refutations provided some reassurance). After the persuasive attack occurred later, inoculated participants again reported being angrier (and less happy) than the non-inoculated group, suggesting the inoculation may have pre-primed some anger in response to the attack. However, not all these affective differences translated into big outcome differences—resistance was observed across the board. Pfau et al. (2009) had predicted that inoculation messages inducing negative affect or focusing on cognition would outperform a positive-affect approach in conferring resistance (in part because Pfau’s earlier work hinted that happiness might actually diminish threat perceptions). Their findings (Pfau et al., 2009) did show affective-negative and cognitive messages elicited more threat than did affective-positive messages, supporting the idea that emotions like anger can amplify the threat mechanism. Yet, importantly, the affective-positive (happiness) message still provided significant resistance, just with slightly lower threat and involvement levels. These mixed results suggested that affect’s role is not straightforward—simply adding a “happy” or “angry” tone to an inoculation message can alter psychological responses (like threat perception and engagement), but the ultimate impact on resisting persuasion can vary and may depend on when and how those emotions come into play during the process.
Further demonstrating the nuanced role of affect, Ivanov et al. (2009a) introduced the idea of attitude base (whether an attitude is primarily affectively or cognitively founded) as a moderator in inoculation. They found that inoculation works best when the message’s content matches the attitude’s base. In practice, this meant that for attitudes rooted in emotion (affective base), an affective inoculation message (one that appeals to emotions or uses emotional arguments) generated stronger resistance than a cognitive message, whereas for attitudes based more on beliefs and reasoning (cognitive base), a cognitive inoculation message (with logical arguments and evidence) was more effective than an affective message. For example, in Ivanov et al. (2009a, 2009b), affective-base attitudes protected by an affective inoculation (emotion-laden language and images) resisted a combined attack more than those protected by a cognitive inoculation; conversely, cognitive-base attitudes protected by a cognitive inoculation resisted more than those protected by an affective inoculation. “More effective,” here, explicitly means more resistant to the subsequent attack as reflected in higher post-attack attitude scores.
This matching effect implies that the same affective treatment can be either beneficial or less so, depending on context—an emotional inoculation is potent in the right context (when tackling an emotion-based attitude), but might underperform if used on a purely cognitive issue. Such findings highlight that affect’s influence is contextual: the impact of emotional content in an inoculation varies with the nature of the attitude and the audience’s frame of mind.
In another creative integration, Miller et al. (2013) embedded a reactance trigger inside the inoculation itself. Psychological reactance refers to the motivational state of anger and negative cognitions that arises when people perceive their freedom to think or choose is being threatened (Brehm, 1966; Dillard and Shen, 2005). In both inoculation conditions, participants first received an explicit forewarning that some upcoming counter-arguments would be “very persuasive, and they might cause [them] to rethink [their] position on this issue.” They then read refutational preemption in which “counterattitudinal arguments [were] introduced in the opening sentences of the paragraphs,” each “successively countered by refutations,” for example: “Opponents of regulation also assert that television programming is protected as ‘free speech’ under the First Amendment.” Refutation followed immediately: “However, this position is questionable. First, children do not enjoy the same First Amendment protections as adults.” The reactance-enhanced version added a second forewarning that opponents threatened participants’ “very freedom to hold” relevant beliefs and preferences—“or even choose how to think for [themselves].” At attack, the study manipulated controlling language; the high-control message included lines such as “we must not resort to government action… Parents must control television content through filtering devices, which have unmistakably proven very effective.” Across sites and issues, this reactance-enhanced inoculation yielded stronger resistance than a traditional inoculation and control, including greater counterarguing and lesser attitude change after the attack.
Taken together, these studies paint a picture of mixed and sometimes contradictory effects of affect in inoculation processes. On one hand, positive emotion during inoculation (as in a reassuring or uplifting message) has been associated with strong resistance outcomes in at least one case (Pfau et al., 2001). On the other hand, negative emotions like anger—especially when tied to a sense of offense or threat to autonomy—can also enhance resistance (Miller et al., 2013), and inducing a bit of anger or fear can heighten the classic mediators of inoculation (threat and involvement, per Pfau et al., 2009). Yet, not all negative affect helps: if fear or anxiety is too strong, it might distract or overwhelm people rather than motivate them. Clearly, affect is not a magic bullet that uniformly increases persuasion immunity; its effect can vary based on the form of affect, its intensity, and the context in which it’s elicited.
Why might affect sometimes help and other times hinder inoculation? Psychoneuroimmunology (PNI), which studies how psychological factors influence the immune system, and research on how affect impacts cognition, suggests that affect operates as a contextual moderator in the inoculation process. Rather than being a separate “third pillar” of inoculation (alongside threat and counterarguing), affect seems to shape the environment in which those cognitive mechanisms function. Using the biological metaphor of inoculation carefully: in the immune system, we know that a person’s emotional state can alter how well a vaccine works. For instance, high stress can trigger the release of cortisol (a stress hormone), which can suppress immune responses. Analogously, if an inoculation message induces a lot of stressful negative affect (say, intense fear or anxiety about the impending attack), that state might actually inhibit the person’s ability to process the message and marshal defenses. The stress response could narrow the individual’s attention or undermine the memory consolidation of the counterarguments provided, thereby weakening the inoculation’s effectiveness. In other words, extreme negative affect during inoculation might backfire, somewhat like how chronic stress makes one biologically more susceptible to illness despite vaccination. This perspective helps explain why purely fear-based messages are not always ideal—a moderate level of threat can motivate, but too much fear can become counterproductive if it tips into paralyzing anxiety.
Conversely, positive affective states tend to have a different effect on the mind. Research in psychology (e.g., the broaden-and-build theory of positive emotions) shows that when people experience a positive mood, their cognitive processing broadens—they become more open-minded, creative, and able to integrate information from diverse sources (Fredrickson and Branigan, 2005). Applying this to inoculation: a positively framed inoculation message might put recipients in a good mood that actually expands their cognitive flexibility. This could help them engage with the counterarguments more thoroughly or see the upcoming attack from a wider perspective, reinforcing their attitude. Such a mechanism could account for why the happiness-inducing inoculation in Pfau et al. (2001) was surprisingly effective—the positive affect may have facilitated deeper or more expansive processing, even though it did not trigger as much perceived “threat” in the moment as the anger-based message did. Positive affect might also reduce defensiveness, allowing people to absorb the inoculation content without feeling overwhelmed.
It is also worth noting that not all negative emotions have the same effect. Emotions like anger are complex—while anger is negative, it is an approach-oriented emotion that often mobilizes people to take action. If the inoculation context makes people angry at the idea of being manipulated or at the source of a potential attack (as in Miller et al., 2013, where participants became annoyed at a threat to their freedom), that anger can channel into motivated resistance, essentially energizing the counterarguing process. However, if the anger or fear is diffuse or directed inward (e.g., feeling helpless or anxious), it may not be useful. The timing of affect also matters: Pfau et al. (2009) observed that some emotional reactions appeared right after the inoculation and then faded, while others only surfaced after the persuasive attack occurred. This indicates that affect can influence both the initial reception of the inoculation and the reaction to the attack, and these influences might differ.
In summary, affect influences inoculation in a moderating, context-dependent manner. Affective states and cues interact with the cognitive processes of resistance rather than act as standalone defenses. The biological “vaccine” metaphor is not a deterministic formula but an interpretive guide: it suggests, for example, that just as stress can weaken an immunization, strong negative emotions might sometimes weaken attitudinal resistance by stifling cognitive functions (e.g., critical thinking or memory). Likewise, just as a healthy internal status can help a vaccine, a positive or appropriately aroused emotional state might enhance the mind’s ability to mount a defense by broadening thinking or increasing motivation. This metaphor is used to illustrate potential mechanisms with limits—psychological resistance is not governed by biology alone, and emotions do not affect persuasion exactly as they do antibodies. Instead, emotions color the psychological context in which inoculation processes unfold.
Rather than treating affect as uniformly good or bad for resistance, it is more accurate to view it as a background factor that can modulate the strength of inoculation’s cognitive defenses. By building on the foundational studies that have begun to untangle these interactions, and by bringing in interdisciplinary insights (from neuroscience, immunology, and psychology), scholars can better understand why inoculation sometimes yields especially robust resistance and other times falters. Such knowledge will ultimately help in designing inoculation messages that are not only logically compelling but also emotionally optimized for maximum persuasive immunity. The continued evolution of inoculation theory will benefit from appreciating these affective nuances while maintaining the clarity of the original biological metaphor as a guiding analogy rather than a literal blueprint.
Motivational threat
Some studies (Clayton et al., 2023; Richards and Banas, 2015; Richards et al., 2017) on inoculating against psychological reactance to persuasive health messages depart from the foundational principles of inoculation theory. In contrast to traditional inoculation theory, the authors use forewarning about potential reactance, which is conceptually analogous to forewarning someone about the side effects of a vaccine rather than introducing the antigen itself. This approach does not present a mild version of the actual persuasive message, which would be necessary to create the specific cognitive antibodies (counterarguments) needed to resist that attack message. Instead, it addresses a meta-cognitive response (reactance) without directly confronting the content of the health message.
Similarly, proposing a so-called motivational threat in the context of psychological inoculation, Banas and Richards (2017) depart from the foundation and principles of biological inoculation. This new operationalization focuses on the motivation to defend attitudes rather than evoking fear about their vulnerability. In the context of biological inoculation, this approach is akin to emphasizing the body’s general preparedness to fight infections without directly engaging with the specific antigens of a pathogen. Traditional inoculation theory, analogous to biological vaccination, relies on introducing a clear and direct “stimulus” (like a weakened virus) to trigger specific, targeted responses (antibodies vis-à-vis counterarguments).
Psychological reactance is a state of the combination of anger and negative cognitions (Dillard and Shen, 2005). What these studies (Clayton et al., 2023; Richards and Banas, 2015; Richards et al., 2017) did is to reduce anger (i.e., psychological reactance), which is analogous to the aim to decrease the cortisol level through stress management techniques, as reviewed above on the role of affect. It is the reduced anger level or a more homeostatic state of mood that might contribute to the inoculation effect.
By shifting the focus to a generalized motivational state, the authors risk undermining the theoretical integrity of inoculation theory, as this new conceptualization does not align with the precise threat-response mechanism fundamental to both biological and psychological inoculation. Their departure from the direct analogy to biological inoculation may undermine the efficacy of building targeted resistance, as the study’s method does not fully engage the cognitive processes that traditional inoculation theory leverages to develop specific and robust defenses against particular persuasive attacks.
Post-inoculation talk
Post-inoculation talk refers to the process where individuals who have received a persuasive “inoculation” (a brief warning and refutation of an argument) go on to discuss or share that experience with others, thereby potentially spreading resistance to a harmful message (Compton and van der Linden, 2022; Ivanov et al., 2012). Compton and van der Linden (2022) likened the effect of PIT to herd immunity in a community. However, this analogy is conceptually misleading. Herd immunity, in the traditional sense, is a passive population-level effect—when enough people are immune to a disease, unimmunized individuals are indirectly protected because the disease has fewer opportunities to spread (John and Samuel, 2000). Importantly, achieving herd immunity does not require any communication or intentional action among those individuals; it simply results from a high proportion of immune people acting as a barrier to transmission.
By contrast, post-inoculation talk is an active, social process. After being “inoculated” against a persuasive message, people actively share their new information or counterarguments with others—for example, a person might warn a friend about a misinformation tactic they learned to resist. This spread of ideas requires conscious communication and social interaction. The herd immunity analogy fails because it confuses a biological, non-communicative process with a social, communicative process. In biology, immune cells coordinate within one organism to fend off infection, but they do not send messages to other organisms. In psychological inoculation, by contrast, individuals do “send messages” to others in their community—they talk about the inoculation content. In other words, PIT operates more like a social contagion of information (Centola, 2010) than a biological immunity effect. Ideas and resistance strategies are passed from person to person through dialogue, much as a beneficial idea might catch on in a social network.
It is also important to note that when we talk about vaccination and immunity, vaccinated people generally do not transmit the vaccine (or virus) to others. Herd immunity stems from immunity halting transmission, not from immune individuals actively immunizing others. (In very rare cases, certain live attenuated vaccines can shed weakened virus to others, but this is an unintended side effect and not how herd immunity works.) This biological reality underscores the difference: the spread of protection in herd immunity is unintentional and passive, whereas the spread of resistance in PIT is intentional and active. Individuals engaging in post-inoculation talk are purposefully sharing their defenses (counterarguments) with peers.
In summary, the structural mismatch between herd immunity and post-inoculation talk makes the analogy weak. Herd immunity happens “in the background” once a threshold of immunity is reached, without people having to do anything additional or communicate. Post-inoculation talk, on the other hand, requires people to take action by talking and persuading. While the herd immunity analogy was a creative suggestion by Compton and van der Linden (2022) to illustrate how inoculation effects might spread, the core mechanisms differ fundamentally. A social transmission framework is more fitting for PIT, because it highlights how resistance to persuasion can spread through conversation and social networks rather than through biological immunity.
Given the controversial extensions of inoculation theory, this critical subject matter requires a more profound understanding, rigorous scrutiny, and suitable practical implementation. The validity of analogic reasoning used in inoculation theory is rooted in shared foundational principles among biology, neuroscience, and psychology. As follows, the author delves into these territories of study and search for innovative approaches to advancing inoculation theory.
Existing issues and new proposals
Passive defense vs. active defense
In immunology, passive immunity is the type of immunity acquired by receiving antibodies produced by another human being or animal (Siegrist, 2018). It can be obtained through the transfer of maternal antibodies to full-term babies and convalescent plasma or by administering antibody-containing blood products, such as biologically engineered monoclonal antibodies (mAbs) (Senefeld et al., 2023; Slifka and Amanna, 2018). Passive immunity provides immediate protection against familiar pathogens or toxins in the received antibodies, but it is ineffective when encountering new pathogens and gradually diminishes within a few weeks (Slifka and Amanna, 2018).
In comparison, active immunity is acquired through the body’s immune response to pathogens or antigens, either naturally or artificially (i.e., through vaccination). Active immune responses occur when the immune system recognizes threats and issues warnings (Siegrist, 2018). Pattern recognition receptor (PRR)-bearing cells, such as macrophages, microglia, and neutrophils, in the innate immune system can identify patterns for certain familiar pathogens. However, it takes approximately 5 days for antigen-presenting cells (APCs) to effectively communicate with the adaptive immune system, such as presenting antigens to T cells, B cells, and other immune cells to trigger more specific cellular responses (e.g., activation of T killer cells) and humoral responses (e.g., antibody production) (Schuijs et al., 2019; Silva-Gomes et al., 2015). In summary, only active immunity can address novel threats, while passive immunity can rapidly respond to familiar threats.
The inoculation theory also considers passive versus active defenses, which differ in how refutation is presented (McGuire, 1961b). The former refutation message (argument) is presented to the subject directly, while the latter refutation is generated internally through thought-provoking means like thought-listing and gaming. A concrete example of a gaming-based active defense is the Bad News prebunking intervention (Roozenbeek and van der Linden, 2019; Roozenbeek et al., 2020). In a short, guided session, players adopt the role of an online misinformation creator and are exposed to attenuated (mild) versions of common manipulation tactics—such as polarization, emotional language, impersonation, conspiratorial framing, and discrediting. Each tactic is presented in a safe, simulated form and is immediately paired with on-screen explanations and feedback that label why the tactic misleads and how to recognize it—functionally providing the refutation. This forewarning + mild attack + immediate explanation/feedback sequence equips players with procedural counterarguments they later apply to novel content, thereby increasing resistance to misinformation; because the weakened attacks are multiple and varied, the exercise also approximates a multivalent active defense in practice (Roozenbeek and van der Linden, 2019; Roozenbeek et al., 2020).
Wood and Quinn (2003) concluded that active defense using thought listing [for utilizing apps, refer to Roozenbeek et al., 2020; Roozenbeek and van der Linden, 2019] generated more resistance to counterarguments compared to passive defense through message-reading in a meta-analysis. McGuire (1961a) demonstrated that passive defense conferred greater immunity against familiar counterarguments than active defense. However, when faced with new counterarguments, active defense proved more effective in enhancing resistance. Therefore, not only did McGuire (1961a) utilize a valid analogy from the immune system, but his findings also align with the immunological mechanism.
Nevertheless, the “active” form of refutation has been operationalized using artificially-controlled instruments, such as writing pads or games, in previous inoculation studies. The external validity of such a practice is hence doubtable. Is there a natural means for people to generate refutational arguments against the attack message like misinformation in their minds? The author will argue below that the mRNA type of inoculation will do the trick.
Therapeutic inoculation
Addressing “infected” people
Inoculation theory does not initially intend to address what if individuals are already “infected” given its limiting application to “cultural truisms” by McGuire (Amazeen et al., 2022; Compton, 2020; Pryor and Steinfatt, 1978). Infected cells are analogous to people who have been exposed and persuaded by attack messages. Prophylactic inoculation can also deal with “infected” people, like the Rabies vaccine, Hepatitis B vaccine, and HPV vaccine, which can be used in certain situations to help the immune system deal with an ongoing infection (Tian et al., 2022). Some studies [e.g., Amazeen and Krishna, 2024; Ivanov et al., 2022; Mason et al., 2024; Wood, 2007] in psychological inoculation merely continued to employ prophylactic strategies for treatment purposes. That is, they channelled the logic of prophylactic vaccination into therapeutic inoculation without acknowledging the unique characteristics of therapeutic vaccination.
Furthermore, unlike prophylactic vaccines reviewed above, which typically induce an immune response primarily through the stimulation of B-cells to produce specific antibodies, truly therapeutic vaccines focus more on activating T-cells, particularly cytotoxic T-lymphocytes, by presenting the antigens of infected cells. For instance, the therapeutic vaccine using oncolytic viruses like talimogene laherparepvec (T-VEC) can treat certain types of cancer by infecting cancer cells, leading to an increased presentation of tumor antigens by dendritic cells (Conry et al., 2018). To wit, a few prophylactic vaccines can treat “infection,” but that does not make them therapeutic vaccines.
Drawing an analogy to the persuasion context, actual therapeutic vaccines would employ a re-packaged (or engineered) message, akin to the antigen of infected cells in the medical context, to treat “persuaded” recipients. This message, designed to be acceptable to those already “infected” by an attack argument, is a foil to reveal the inherent problems and loopholes in the attack. The ultimate goal is to motivate these “infected” individuals to fully leverage their critical abilities (akin to T cells) to refute both the engineered message and the original attack argument, thereby shifting their beliefs. The illustrative example concerns adults who already endorse the claim that “because carbon dioxide is only 0.04% of Earth’s atmosphere, it cannot meaningfully warm the planet” (Reuters Fact Check, 2024). A therapeutic inoculation begins with a congenial narrative delivered by an ingroup peer: “I used to cite that 0.04% figure myself; it sounded like the final word.” The message then introduces an engineered antigen analogue—a concise contradiction embedded in the same first-person voice: “Then I learned that trace chemicals can have outsized effects; ozone is just parts per billion yet blocks UV, and pharmaceutical micro-doses can save lives. The tiny CO₂ fraction works the same way, trapping heat in the infrared band.” By validating the audience’s prior reasoning before surfacing the factual counter, the narrative earns epistemic trust while exposing the flaw.
Immediately after the narrative, viewers receive a self-explanation task: a one-minute animation of the infrared absorption spectrum plays while a prompt asks them to write, in their own words, how a small concentration can yield a large radiative effect. Requiring this generative step is functionally parallel to engaging cytotoxic T-cells in therapeutic vaccines: the audience must produce the disconfirming logic rather than passively accept it. A brief booster text message, sent 1 week later, asks recipients to recall and restate their explanation, mirroring booster dosing that refreshes immunological memory.
Addressing desensitization effect
The immune system only treats non-self external substances as threats through the complicated coordination among T regulatory cells, memory, and other immune cells. This also concerns the immune tolerance and desensitization raised by Compton (2020) in the context of persuasion, a process of training the immune system to tolerate the allergen through the controlled take-in of allergic food or so-called therapeutic vaccine. The therapeutic vaccine aims to shift allergic response (Th2-dominant) to a more balanced response (coordination among Th1, Th2, and T regulatory cells) and to direct the B cells to produce IgG antibodies instead of allergen-specific IgE antibodies (Gutowska-Owsiak and Ogg, 2017).
Time and again, in the context of persuasion that applies the therapeutic vaccine, a congenial message accompanied by a reminder to pause and reflect on the argument, position, and motives inherent in the persuasive message could serve the purpose (cf. Compton, 2020).
Future research directions and possible propositions
Analogies of different types of vaccines
In the medical field, there are many types of vaccines currently available to the market, such as vaccines employing the latest Nobel-prize-winning mRNA technology, inactivated vaccines, live-attenuated vaccines, subunit vaccines, viral vector vaccines, toxoid vaccines, conjugate vaccines, and peptide vaccines (Plotkin et al., 2018). These vaccines differ in their composition, safety profiles, side effects, the level of immune response elicited, the need for booster shots, and target pathogens. Nevertheless, previous psychological inoculation studies have consistently drawn the medical analogy of a live-attenuated vaccine form since the theory was proposed in 1961. However, the advent of mRNA vaccines provides a metaphorical framework that diverges substantially from the traditional live-attenuated paradigm. Unlike traditional vaccines that introduce a weakened or inactivated pathogen into the body, mRNA vaccines administer genetic instructions to cells to manufacture specific viral proteins, such as the SARS-CoV-2 spike protein. These proteins are subsequently recognized by antigen-presenting cells (APCs), which stimulate B cells to produce antibodies and activate other immune responses (Pardi et al., 2018).
One notable innovation that an mRNA model brings to psychological inoculation is the potential obviation of the two classical components: forewarning and refutation. In the context of resisting misinformation, instead of presenting the subject with a weakened counter-argument and then refuting it, the mRNA model could encode certain values or frameworks within the attack messages like misinformation, guiding the individual to critically evaluate and ‘translate’ this misinformation. Upon completing this cognitive ‘translation,’ the individual is prompted to deploy countermeasures akin to antibodies, enhancing their resistance to subsequent misinformation. Some recent studies have found that careful analytical reasoning relative to the use of familiarity and source heuristics is crucial to fighting misinformation (Pennycook and Rand, 2019, 2021) [for other studies on the inoculation effect of information or media literacy in fighting misinformation, see Arechar et al., 2023; Jones-Jang et al., 2019; Machete and Turpin, 2020].
Moreover, some vaccine technologies can provide “broad spectrum” immunity against multiple strains of viruses. Future research should extend by examining diverse argument forms (Banas and Miller, 2013), multiple different attacks comparable to multivalent vaccines (Ivanov et al., 2009b), contexts, combinations of the weakened attack (the vaccine), the “sugar-coated” attack, and the strong attack messages [for the effect of the types of forewarnings, see Benoit, 1998; Papageorgis, 1968].
Conceptual propositions
In advancing our discussion, the author proposes a few conceptual propositions, which are intended to serve as a guiding compass for future empirical investigations, providing a structured and systematic approach to examining these theoretical propositions within the realm of inoculation theory.
P1: The inoculation effect is moderated by the then mood. Specifically, the introduction of positive affect will enhance the inoculation effect, while introducing negative affect will dampen the effect.
P2: The application of therapeutic vaccines in the persuasion context, which involves the use of a congenial message accompanied by a reminder message that encourages reflection on the argument, position, and motives inherent in the persuasive message, will shift the beliefs of “infected” individuals towards a more balanced view. Furthermore, the use of a re-packaged (engineered) message, which can be accepted by “infected” individuals and exposes the problems and loopholes inherent in the attack argument, will motivate these individuals to refute the engineered message and the attack argument altogether, leading to a change in their initial beliefs.
P3: Different types of psychological inoculation—mRNA versus live-attenuated—will exhibit significant differences in their effect on conferring resistance to general attack messages. It is anticipated that the mRNA type of psychological inoculation will be more effective compared to the live-attenuated type, similar to observed efficacy differences in the domain of medical vaccines.
While not exhaustive, these propositions offer a starting point for understanding and exploring the potential intersections between immunology and information processing. They invite researchers to consider how the mechanisms of the immune system might inform our understanding of how individuals process, resist, and change their beliefs in response to persuasive messages. As such, they represent a promising avenue for future research in this area.
Limitations and future research directions
The extensions and analogies discussed in the present paper, while generative, are necessarily bounded by epistemic differences between immunology and persuasion. Immune efficacy, though rooted in molecular pathways, is modulated by host and contextual variables such as age, sex hormones, genotype, stress, sleep, physical activity, microbiome composition, and circadian timing of vaccination (Zimmermann and Curtis, 2019). Persuasion outcomes likewise depend on multiple moderators—culture, identity, emotion, social norms, and both deliberative and automatic cognitive processes (Petty and Cacioppo, 1986). Analogies retain explanatory value when their relational structure, rather than their surface features, is mapped with care (Gentner, 1983; Gentner and Holyoak, 1997).
A productive next step is to position inoculation theory within broader frameworks that explain how and why people process counter-attitudinal information. For instance, motivated reasoning predicts that people defend identity-relevant attitudes even when accuracy cues are present (Kunda, 1990). Future work should examine whether inoculation’s threat-plus-refutation package can redirect this endogenous motive toward systematic counterarguing. One empirical approach is to vary identity centrality (e.g., political versus apolitical topics) and test if inoculation effects weaken when identity stakes rise—unless paired with a self-affirmation preface that lowers defensive arousal (Steele, 1988). This line of inquiry would clarify boundary conditions for inoculation in highly polarized contexts.
The above-mentioned mRNA analogy can be examined through a three-condition randomized experiment. One condition receives a traditional inoculation message: a brief forewarning followed by a weakened persuasive claim and its explicit rebuttal (the communication analogue of a live-attenuated vaccine). A second condition receives an mRNA-style message. Here, the same weakened claim appears, but instead of a ready-made rebuttal the text embeds prompts that nudge recipients to craft their own. A third condition serves as an uninoculated control. After a delay, all participants encounter a strong persuasive attack on the same topic. Primary outcomes should include attitude change after the attack and the number and quality of spontaneous counterarguments, coded from open-ended thought listings. A process check—such as performance on a short analytic-reasoning task immediately after the inoculation message (Pennycook and Rand, 2019)—can verify that the mRNA-style prompts indeed foster reflective thinking. If participants who generated their own rebuttals show equal or greater resistance than those given a prewritten refutation, the mRNA analogy gains empirical support.
A systems-level perspective, grounded in vaccinology, can translate immunological precision into a research agenda for psychological inoculation. Vaccination research shows that protection is probabilistic, governed by the quantitative relation between viral load and antibody concentration, which follows a sigmoidal dose–response curve (Khoury et al., 2021). In persuasion, an analogous curve can be estimated by jointly manipulating the cognitive weight of weakened attacks presented during inoculation and the persuasive load of subsequent challenges, then modelling the probability of attitude change as a function of their ratio. To integrate these elements, resistance can be conceptualized as a continuous index—the summed strength of stored counterarguments, weighted by retrieval speed, divided by the aggregated force of an attack message. Variables contributing to the denominator, such as repetition, credibility, and affective intensity, are well established in dual-process models of persuasion (Petty and Cacioppo, 1986).
These extensions align inoculation theory with the rigor of contemporary vaccine science. Specifying how strong, how often, and how diverse inoculation messages should be invites falsifiable predictions and strengthens the conceptual bridge between biological and psychological resilience.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
GF: Conceptualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Conflict of interest
The author declares that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alexejeva, L. A., and Mishlanova, S. L. (1997). Basic metaphors of immunology. Immunol. Lett. 56:255.
Amazeen, M. A., and Krishna, A. (2024). Refuting misinformation: examining theoretical underpinnings of refutational interventions. Curr. Opin. Psychol. 56:101774. doi: 10.1016/j.copsyc.2023.101774
Amazeen, M. A., Krishna, A., and Eschmann, R. (2022). Cutting the bunk: comparing the solo and aggregate effects of prebunking and debunking Covid-19 vaccine misinformation. Sci. Commun. 44, 387–417. doi: 10.1177/10755470221111558
Arechar, A. A., Allen, J., Berinsky, A. J., Cole, R., Epstein, Z., Garimella, K., et al. (2023). Understanding and combatting misinformation across 16 countries on six continents. Nat. Hum. Behav. 7, 1502–1513. doi: 10.1038/s41562-023-01641-6
Banas, J. A., and Miller, G. (2013). Inducing resistance to conspiracy theory propaganda: testing inoculation and metainoculation strategies. Hum. Commun. Res. 39, 184–207. doi: 10.1111/hcre.12000
Banas, J. A., and Richards, A. S. (2017). Apprehension or motivation to defend attitudes? Exploring the underlying threat mechanism in inoculation-induced resistance to persuasion. Commun. Monogr. 84, 164–178. doi: 10.1080/03637751.2017.1307999
Banchereau, J., and Steinman, R. M. (1998). Dendritic cells and the control of immunity. Nature 392, 245–252. doi: 10.1038/32588
Benoit, W. L. (1998). “Forewarning and persuasion” in Persuasion: Advances through meta-analysis. eds. M. Allen and R. W. Preiss (Cresskill, NJ: Hampton Press).
Black, M. (1962). Models and metaphors: Studies in language and philosophy, vol. 23. Ithaca, NY: Cornell University Press.
Centola, D. (2010). The spread of behavior in an online social network experiment. Science 329, 1194–1197. doi: 10.1126/science.1185231
Christakis, N. A., and Fowler, J. H. (2013). Social contagion theory: examining dynamic social networks and human behavior. Stat. Med. 32, 556–577. doi: 10.1002/sim.5408
Cinelli, M., Quattrociocchi, W., Galeazzi, A., Valensise, C. M., Brugnoli, E., Schmidt, A. L., et al. (2020). The COVID-19 social media infodemic. Sci. Rep. 10:16598. doi: 10.1038/s41598-020-73510-5
Clayton, R. B., Compton, J., Reynolds-Tylus, T., Neumann, D., and Park, J. (2023). Revisiting the effects of an inoculation treatment on psychological reactance: a conceptual replication and extension with self-report and psychophysiological measures. Hum. Commun. Res. 49, 104–111. doi: 10.1093/hcr/hqac026
Compton, J. (2020). Prophylactic versus therapeutic inoculation treatments for resistance to influence. Commun. Theory 30, 330–343. doi: 10.1093/ct/qtz004
Compton, J. A., and Pfau, M. (2005). Inoculation theory of resistance to influence at maturity: recent progress in theory development and application and suggestions for future research. Ann. Int. Commun. Assoc. 29, 97–146. doi: 10.1080/23808985.2005.11679045
Compton, J., and van der Linden, S. (2022). The analogy of/and inoculation theory to mental immunity. Available online at: https://www.prosocial.world/posts/the-analogy-of-and-inoculation-theory-to-mental-immunityfiles/9/the-analogy-of-and-inoculation-theory-to-mental-immunity.html (Accessed March 10, 2024).
Conry, R. M., Westbrook, B., McKee, S., and Norwood, T. G. (2018). Talimogene laherparepvec: first in class oncolytic virotherapy. Hum. Vaccin. Immunother. 14, 839–846. doi: 10.1080/21645515.2017.1412896
Dillard, J. P., and Shen, L. (2005). On the nature of reactance and its role in persuasive health communication. Commun. Monogr. 72, 144–168. doi: 10.1080/03637750500111815
Fredrickson, B. L., and Branigan, C. (2005). Positive emotions broaden the scope of attention and thought-action repertoires. Cognit. Emot. 19, 313–332. doi: 10.1080/02699930441000238
Gentner, D. (1983). Structure-mapping: a theoretical framework for analogy. Cogn. Sci. 7, 155–170. doi: 10.1016/S0364-0213(83)80009-3
Gentner, D., and Holyoak, K. J. (1997). Reasoning and learning by analogy: introduction. Am. Psychol. 52, 32–34. doi: 10.1037/0003-066X.52.1.32
Gutowska-Owsiak, D., and Ogg, G. S. (2017). Therapeutic vaccines for allergic disease. NPJ Vaccin. 2:12. doi: 10.1038/s41541-017-0014-8
Hesse, M. B. (1970). Models and analogies in science. Notre Dame, IN: University of Notre Dame Press.
Ivanov, B., Miller, C. H., Compton, J., Averbeck, J. M., Harrison, K. J., Sims, J. D., et al. (2012). Effects of postinoculation talk on resistance to influence. J. Commun. 62, 701–718. doi: 10.1111/j.1460-2466.2012.01658.x
Ivanov, B., Parker, K. A., and Dillingham, L. (2020). “Inoculation theory as a strategic tool” in The handbook of applied communication research. eds. H. Dan O’Hair, and M. John O’Hair, (New York, NY: Wiley-Blackwell), 11–28.
Ivanov, B., Pfau, M., and Parker, K. A. (2009a). The attitude base as a moderator of the effectiveness of inoculation strategy. Commun. Monogr. 76, 47–72. doi: 10.1080/03637750802682471
Ivanov, B., Pfau, M., and Parker, K. A. (2009b). Can inoculation withstand multiple attacks?: an examination of the effectiveness of the inoculation strategy compared to the supportive and restoration strategies. Commun. Res. 36, 655–676. doi: 10.1177/0093650209338909
Ivanov, B., Rains, S., Dillingham, L., Parker, K., Geegan, S., and Barbati, J. (2022). The role of threat and counterarguing in therapeutic inoculation. South Commun. J. 87, 15–27. doi: 10.1080/1041794X.2021.1983012
John, T. J., and Samuel, R. (2000). Herd immunity and herd effect: new insights and definitions. Eur. J. Epidemiol. 16, 601–606. doi: 10.1023/a:1007626510002
Jones-Jang, S. M., Mortensen, T., and Liu, J. (2019). Does media literacy help identification of fake news? Information literacy helps, but other literacies don’t. Am. Behav. Sci. 65, 371–388. doi: 10.1177/0002764219869406
Kahan, D. M., Peters, E., Wittlin, M., Slovic, P., Ouellette, L. L., Braman, D., et al. (2012). The polarizing impact of science literacy and numeracy on perceived climate change risks. Nat. Clim. Chang. 2, 732–735. doi: 10.1038/nclimate1547
Kasuga, Y., Zhu, B., Jang, K.-J., and Yoo, J.-S. (2021). Innate immune sensing of coronavirus and viral evasion strategies. Exp. Mol. Med. 53, 723–736. doi: 10.1038/s12276-021-00602-1
Khoury, D. S., Cromer, D., Reynaldi, A., Schlub, T. E., Wheatley, A. K., Juno, J. A., et al. (2021). Neutralizing antibody levels are highly predictive of immune protection from symptomatic SARS-CoV-2 infection. Nat. Med. 27, 1205–1211. doi: 10.1038/s41591-021-01377-8
Kunda, Z. (1990). The case for motivated reasoning. Psychol. Bull. 108, 480–498. doi: 10.1037/0033-2909.108.3.480
Liu, L., Gong, T., Tao, W., Lin, B., Li, C., Zheng, X., et al. (2019). Commensal viruses maintain intestinal intraepithelial lymphocytes via noncanonical RIG-I signaling. Nat. Immunol. 20, 1681–1691. doi: 10.1038/s41590-019-0513-z
Machete, P., and Turpin, M. (2020). “The use of critical thinking to identify fake news: a systematic literature review” in Responsible design, implementation and use of information and communication technology. eds. H. Dan O’Hair, and M. John O’Hair (Cham, Switzerland: Springer International Publishing).
Mason, A. M., Compton, J., Tice, E., Peterson, B., Lewis, I., Glenn, T., et al. (2024). Analyzing the prophylactic and therapeutic role of inoculation to facilitate resistance to conspiracy theory beliefs. Commun. Rep. 37, 13–27. doi: 10.1080/08934215.2023.2256803
McFall-Ngai, M., Hadfield, M. G., Bosch, T. C. G., Carey, H. V., Domazet-Lošo, T., Douglas, A. E., et al. (2013). Animals in a bacterial world, a new imperative for the life sciences. Proc. Natl. Acad. Sci. USA 110, 3229–3236. doi: 10.1073/pnas.1218525110
McGuire, W. J. (1961a). The effectiveness of supportive and refutational defenses in immunizing and restoring beliefs against persuasion. Sociometry 24, 184–197. doi: 10.2307/2786067
McGuire, W. J. (1961b). Resistance to persuasion conferred by active and passive prior refutation of the same and alternative counterarguments. J. Abnorm. Soc. Psychol. 63, 326–332. doi: 10.1037/h0048344
McGuire, W. J., and Papageorgis, D. (1961). The relative efficacy of various types of prior belief-defense in producing immunity against persuasion. J. Abnorm. Soc. Psychol. 62, 327–337. doi: 10.1037/h0042026
Miller, J. F. A. P. (2002). The discovery of thymus function and of thymus-derived lymphocytes. Immunol. Rev. 185, 7–14. doi: 10.1034/j.1600-065X.2002.18502.x
Miller, C. H., Ivanov, B., Sims, J., Compton, J., Harrison, K. J., Parker, K. A., et al. (2013). Boosting the potency of resistance: combining the motivational forces of inoculation and psychological reactance. Hum. Commun. Res. 39, 127–155. doi: 10.1111/j.1468-2958.2012.01438.x
Niederdeppe, J., Heley, K., and Barry, C. L. (2015). Inoculation and narrative strategies in competitive framing of three health policy issues. J. Commun. 65, 838–862. doi: 10.1111/jcom.12162
Pardi, N., Hogan, M. J., Porter, F. W., and Weissman, D. (2018). mRNA vaccines — a new era in vaccinology. Nat. Rev. Drug Discov. 17, 261–279. doi: 10.1038/nrd.2017.243
Pennycook, G., and Rand, D. G. (2019). Lazy, not biased: susceptibility to partisan fake news is better explained by lack of reasoning than by motivated reasoning. Cognition 188, 39–50. doi: 10.1016/j.cognition.2018.06.011
Pennycook, G., and Rand, D. G. (2021). The psychology of fake news. Trends Cogn. Sci. 25, 388–402. doi: 10.1016/j.tics.2021.02.007
Petty, R. E., and Cacioppo, J. T. (1986). “The elaboration likelihood model of persuasion” in Advances in experimental social psychology. ed. L. Berkowitz, vol. 19 (New York, NY: Academic Press), 123–205.
Pfau, M., Semmler, S. M., Deatrick, L., Mason, A., Nisbett, G., Lane, L., et al. (2009). Nuances about the role and impact of affect in inoculation. Commun. Monogr. 76, 73–98. doi: 10.1080/03637750802378807
Pfau, M., Szabo, A., Anderson, J., Morrill, J., Zubric, J., and H-Wan, H. H. (2001). The role and impact of affect in the process of resistance to persuasion. Hum. Commun. Res. 27, 216–252. doi: 10.1111/j.1468-2958.2001.tb00781.x
Plotkin, S. A., Orenstein, W. A., Offit, P. A., and Edwards, K. M. (2018). Plotkin's vaccines. 7th Edn. Philadelphia, PA: Elsevier.
Plotkin, S. L., and Plotkin, S. (2018). “A short history of vaccination” in Plotkin's vaccines. eds. S. A. Plotkin, W. A. Orenstein, P. A. Offit, and K. M. Edwards (Philadelphia, PA: Elsevier).
Pryor, B., and Steinfatt, T. M. (1978). The effects of initial belief level on inoculation theory and its proposed mechanisms. Hum. Commun. Res. 4, 217–230. doi: 10.1111/j.1468-2958.1978.tb00611.x
Reuters Fact Check. (2024, 2024/01/04/T16:37:42Z). Fact check: share of CO2 in the atmosphere not a reflection of its climate impact. Reuters. Available online at: https://www.reuters.com/fact-check/share-co2-atmosphere-not-reflection-its-climate-impact-2024-01-04/files/1189/share-co2-atmosphere-not-reflection-its-climate-impact-2024-01-04.html (Accessed January 5, 2025).
Richards, A. S., and Banas, J. A. (2015). Inoculating against reactance to persuasive health messages. Health Commun. 30, 451–460. doi: 10.1080/10410236.2013.867005
Richards, A. S., Banas, J. A., and Magid, Y. (2017). More on inoculating against reactance to persuasive health messages: the paradox of threat. Health Commun. 32, 890–902. doi: 10.1080/10410236.2016.1196410
Roozenbeek, J., Maertens, R., McClanahan, W., and van der Linden, S. (2020). Disentangling item and testing effects in inoculation research on online misinformation: Solomon revisited. Educ. Psychol. Meas. 81, 340–362. doi: 10.1177/0013164420940378
Roozenbeek, J., and van der Linden, S. (2019). Fake news game confers psychological resistance against online misinformation. Palgrave Commun. 5:65. doi: 10.1057/s41599-019-0279-9
Schuijs, M. J., Hammad, H., and Lambrecht, B. N. (2019). Professional and 'amateur' antigen-presenting cells in type 2 immunity. Trends Immunol. 40, 22–34. doi: 10.1016/j.it.2018.11.001
Senefeld, J. W., Franchini, M., Mengoli, C., Cruciani, M., Zani, M., Gorman, E. K., et al. (2023). COVID-19 convalescent plasma for the treatment of immunocompromised patients: a systematic review and Meta-analysis. JAMA Netw. Open 6:e2250647. doi: 10.1001/jamanetworkopen.2022.50647
Siegrist, C.-A. (2018). “Vaccine Immunology” in Plotkin's vaccines. eds. S. A. Plotkin, W. A. Orenstein, P. A. Offit, and K. M. Edwards (Philadelphia, PA: Elsevier), 16–34.e17.
Silva-Gomes, S., Decout, A., and Nigou, J. (2015). “Pathogen-associated molecular patterns (PAMPs)” in Encyclopedia of inflammatory diseases. ed. M. Parnham (Basel, Switzerland: Springer Basel), 1–16.
Slifka, M. K., and Amanna, I. J. (2018). “Passive Immunization” in Plotkin's vaccines. eds. S. A. Plotkin, W. A. Orenstein, P. A. Offit, and K. M. Edwards. 7th ed. (Philadelphia, PA: Elsevier).
Steele, C. M. (1988). “The psychology of self-affirmation: sustaining the integrity of the self” in Advances in experimental social psychology. ed. L. Berkowitz, vol. 21 (New York, NY: Academic Press), 261–302.
Tian, Y., Hu, D., Li, Y., and Yang, L. (2022). Development of therapeutic vaccines for the treatment of diseases. Mol. Biomed. 3:40. doi: 10.1186/s43556-022-00098-9
Wood, M. L. M. (2007). Rethinking the inoculation analogy: effects on subjects with differing preexisting attitudes. Hum. Commun. Res. 33, 357–378. doi: 10.1111/j.1468-2958.2007.00303.x
Wood, W., and Quinn, J. M. (2003). Forewarned and forearmed? Two meta-analysis syntheses of forewarnings of influence appeals. Psychol. Bull. 129, 119–138. doi: 10.1037/0033-2909.129.1.119
Keywords: inoculation theory, immunological analogies, therapeutic inoculation, extensions, new directions
Citation: Feng GC (2025) Inoculation theory revisited: reinterpretations, extensions, and new directions. Front. Commun. 10:1576772. doi: 10.3389/fcomm.2025.1576772
Edited by:
Sara Aboulaghras, Ibn Tofail University, MoroccoReviewed by:
Yerina Ranjit, University of Missouri, United StatesBugingo Pacifique, University of Montreal Hospital Centre (CRCHUM), Canada
Copyright © 2025 Feng. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guangchao Charles Feng, Y2hhcmxlc2ZlbmdAaGtidS5lZHUuaGs=