 Bettina Bongiovanni1,2,3
Bettina Bongiovanni1,2,3 Ariana Díaz1,2
Ariana Díaz1,2 Natalia Santucci1,2
Natalia Santucci1,2 Luciano David D’Attilio1,2
Luciano David D’Attilio1,2 Oscar Bottasso1,2
Oscar Bottasso1,2 Rogelio Hernández Pando4*
Rogelio Hernández Pando4* María Luisa Bay1,2*
María Luisa Bay1,2*- 1Instituto de Inmunología Clínica y Experimental de Rosario (IDICER CONICET-UNR), Rosario, Argentina
- 2Facultad de Cs. Médicas, UNR, Rosario, Argentina
- 3Facultad de Cs. Bioquímicas y Farmacéuticas, Universidad Nacional de Rosario (UNR), Rosario, Argentina
- 4Sección de Patología Experimental, Departamento de Patología, Instituto Nacional de Ciencias Médicas y Nutrición Salvador Zubirán, México, Mexico
Dehydroepiandrosterone (DHEA) is an androgen synthesized by the adrenal cortex, which is an intermediary in the biosynthesis of sex hormones, such as testosterone and estradiol. DHEA mostly circulates as a conjugated ester, in the form of sulfate (DHEA-S). There exist several endogenous factors able to influence its synthesis, the most common ones being the corticotrophin-releasing hormone (CRH), adrenocorticotrophin (ACTH), growth factors, and proinflammatory cytokines, among others. Like other steroid hormones, DHEA, can alter the functioning of immune cells and therefore the course of diseases exhibiting an immune-inflammatory component, mostly from autoimmune or infectious nature. We herein review the role played by DHEA during a major infectious disease like tuberculosis (TB). Data recorded from TB patients, mouse models, or in vitro studies show that DHEA is likely to be implied in better disease control. This provides a stimulating background for carrying out clinical studies aimed at assessing the usefulness of DHEA as an adjuvant in TB patients.
A Brief Overview of DHEA
The bidirectional communication between the neuroendocrine and immune systems has been widely evidenced, both in humans and in experimental models. Immune compounds are likely to modify the functioning of the endocrine system. Conversely, adrenal steroids can alter the functioning of immune cells and therefore the course of diseases exhibiting an immune-inflammatory component, mostly autoimmune or infectious. This interconnection between the immune and the neuroendocrine systems is partly due to the stimulatory action of inflammatory cytokines on the Hypothalamic-Pituitary-Adrenal axis. Cytokines such as IL-6, IL-1β, and TNF-α stimulate the production of CRH by the hypothalamus with the subsequent release of ACTH in the pituitary gland further promoting the secretion of steroid hormones at the level of the adrenal cortex: glucocorticoids, in humans mainly cortisol, and DHEA (1, 2).
DHEA is an androgen synthesized by the adrenal cortex. Many endogenous factors can influence its synthesis, i.e., CRH, ACTH, growth factors, and cytokines. While sulfated steroids were considered metabolic end-products, additional research showed that sulfated steroids, such as DHEA-S, can act as circulating reservoirs for the peripheral formation of bioactive. Therefore, DHEA synthesized in the adrenal gland, is transformed into DHEA-S to be released into the bloodstream (3). In turn, desulfation, through steroid sulfatase, predominates in the mammary gland, ovary, prostate, testicles, placenta, and uterus wherein produced DHEA serves as a precursor of sex hormones. However, in the liver, a so-called “futile-loop” of DHEA/DHEA-S is present, as well as for other steroids, challenging the view that the pathway can only go one way (3).
As well as being the physiological precursor for the synthesis of androgens and estrogens DHEA can also be metabolized into oxygenated derivatives in non-steroidogenic tissues, especially in the human liver (4). There is evidence that DHEA can be converted into oxygenated derivatives such as 3β, 17β-androstenediol (AED) and 3β, 16β, 17β-androstentriol by monocyte-derived macrophages (5).
Concerning their metabolic functions, both DHEA and its metabolites carry out biological actions through the activation of various receptors, such as estrogen receptors, the receptor for activated peroxisome proliferators (PPAR), and the pregnane X receptor, as well as other membrane-associated receptors (6). The full range of biologically active forms of DHEA is a pending issue. Since DHEA is metabolized intracellularly to other steroids, including estradiol, androstenediol (7, 8), among others, its multiple actions may be mediated by different metabolites and receptors, including differential effects on alpha and beta estrogen receptors. Although the mechanism of action of DHEA is not fully elucidated, the hormone would exert its action indirectly on the peripheral target tissues after its conversion to androgens and/or estrogens; or directly, as a neurosteroid (9). The same holds for macrophages (10–12).
DHEA and Immune Response in the Anti-Infective Context
Animal studies showed that DHEA increases immune function protecting animals from viral, bacterial, and parasitic infections (13, 14). This hormone also has antiglucocorticoid effects both at the level of macrophage (10) and lymphocyte functions (15).
Studies in human cells indicate that DHEA favors a Th1 cytokine profile by acting as a transcriptional activator of the CD4+ cell IL-2 gene (16, 17). DHEA-S, in turn, has immunomodulatory functions with low levels of this hormone being associated with changes in the repertoire of immune cells, such as a decrease in the main T cell subsets, together with increased numbers of NK cells, and T cells exhibiting an activated phenotype (18, 19). Furthermore, during septic shock or multiple traumas, low levels of DHEA-S seem to be a marker of poor outcomes (20, 21).
The immunoregulatory actions of DHEA are also tissue-dependent. As stated above, after biosynthesis, DHEA circulates primarily in its sulfated form. At the tissue level, target cells express transmembrane organic anion transporting polypeptides (OATPs) that facilitate cellular uptake of the sulfated steroid. Once inside the cell, sulfatases hydrolyze this steroid sulfate ester to active, unconjugated DHEA. This and its metabolites must bind to the corresponding receptors to carry out their biological functions. Sulfation and desulfation are thus fundamental pathways, for which tissue distribution and regulation of these processes are essential for steroid functions (3, 22).
In this sense, at the lymphoid level, Daynes et al. observed that, although recirculating T lymphocytes have the inherent potential to produce both IL-2 and IL-4, in DHEA challenged mice, lymphocyte isolates from peripheral lymph nodes, spleen, and Peyer’s patches showed an increased ability to produce IL-2 and IFNγ after activation. At variance with these results, the challenge with DHEA sulfate was only active on lymphocytes from peripheral lymph nodes and spleen; with lymphocytes obtained from Peyer’s patches continuing to produce IL-4 and IL-5 (23, 24).
As well as lymph nodes contain one of the highest levels of sulfatase activity, macrophages exhibit the main DHEAS sulfatase activity within secondary lymphoid tissue. In this context, macrophages play a fundamental role, since when exposed to microbial stimulants produce mediators with paracrine or autocrine action (TNF-α and IFN-α) capable of inhibiting their ability to metabolize DHEAS. Chronic infections, including the one induced by Mycobacterium tuberculosis, result in elevated circulating levels of several cytokines, including TNF-α and type 1 interferon, as well as low levels of DHEA that correlate negatively with the severity of lung disease (25).
A decrease in DHEA plasma concentrations also occurs during severe infections, such as African trypanosomiasis, cysticercosis, AIDS, TB (26–29), as well as autoimmune disturbances or other chronic diseases (30). This dysregulated adrenal response may be partly due to the presence of inadequate concentrations of TNF-α (31, 32); reinforced by the fact that increased levels of TNF-α and IL-6 during chronic inflammation are also accompanied by a drop in circulating levels of DHEA-S and DHEA (33).
Turning to the infectious context, immune-mediated protective effects of both DHEA and AED were observed in mice inoculated with lethal doses of Pseudomonas aeruginosa or Enterococcus faecalis (14). It has also been shown that DHEA supplementation in Trypanosoma cruzi infected mice enhanced the immune response, resulting in lower parasitemia (34). In dexamethasone-immunosuppressed mice undergoing an experimental infection by Cryptosporidium parvum, treatment with DHEA reduced both the elimination of fecal oocysts and the parasitic colonization of the ileum along with higher amounts of splenic CD4+ and CD8+ T cells (35). Another study in mice infected with avirulent C. parvum or Toxoplasma gondii also showed that DHEA administration reduced mortality along with a significant reduction in the cryptosporidium oocyst count in feces and intestinal villi or brain toxoplasma cysts (36).
In a cross-sectional study about Plasmodium falciparum parasitemia in 12-18-year-old schoolgirls from an area of intense transmission in Kenya, DHEA levels were associated with a decrease in parasitemia, even after age adjustment. People with low levels of DHEA had significantly higher parasite densities than those with higher amounts of DHEA (37).
DHEA and TB
Studies at the Systemic Level
Infection in man with Mycobacterium tuberculosis (Mtb) can result in a varying degree of organic compromise, ranging from an asymptomatic process to frank pulmonary pathology, depending on the interaction between Mtb, and the host immune response (38). The first line of defense against Mtb is made up of macrophages, which acquire their maximum bactericidal potential when activated by IFN-γ (37). This cytokine is mainly secreted by CD4+ Th1 lymphocytes (39). The cytokines released during the immune response, TNF-α and IL-6, among others, stimulate the neuroendocrine system to provoke an important hormonal response (40), which in turn impacts immunocompetent cells. As anticipated, glucocorticoids can inhibit Th1 responses, while their natural antagonist DHEA can promote them (28).
When analyzing the circulating levels of cytokines and hormones in TB patients with different degrees of pulmonary involvement, we found that they presented increased levels of IFN-γ, IL-6, and cortisol, while those of DHEA situated well below the normal values, the lowest values corresponding to the advanced forms (39). Plasma levels of DHEA were positively correlated with the amounts of IFN-γ present in the culture supernatants from their Mtb-stimulated peripheral blood mononuclear cells (PBMCs). Conversely, the cortisol/DHEA ratio was inversely associated with the in vitro production of this cytokine (28, 40). However, after the second month of TB-specific treatment, the cortisol/DHEA ratio shows a reduction (41, 42); suggesting that this relationship may constitute a useful tool for the monitoring of TB patients undergoing therapy.
We further analyzed the relationship between cortisol, DHEA, and the antimicrobial peptides cathelicidin LL-37, β-defensin-2 (HBD- 2), and β-defensin-3 (HBD-3) that play a fundamental role in the antimycobacterial response (43). We observed a positive correlation between plasma levels of cortisol and HBD-3, as well as DHEA and LL-37 in patients with severe TB that disappeared during successful treatment (43), indicating that peptides are partly regulated by adrenal steroids.
In the case of TB and HIV comorbidity, in active TB patients, the plasma levels of DHEA were also below the ones recorded in HIV-positive subjects without Mtb coinfection or healthy donors. Concomitantly, in HIV-TB patients with immune reconstitution syndrome, plasma levels of DHEA-S were three-times lower than in non-TB groups (44). A phenomenon that points out the inverse relationship between the inflammatory response and the presence of DHEA in circulation. HIV-TB patients also displayed a positive correlation between plasma DHEA levels and the frequency of a terminally differentiated population of CD8+ T cells thought to be implied in mycobacterial containment (45).
In Vitro and Experimental Studies
We also demonstrated that the culture supernatants from the Mtb-stimulated PBMCs of TB patients restrained the secretion of DHEA by a human adrenal cell line NCI-H295-R (46). To ascertain the possibility of inhibiting the activity of some products from the anti-tuberculous immune response on adrenal steroidogenesis, treatment with anti-TGF-β antibodies abolished the inhibitory effects of such supernatants on DHEA production by NCI-H295-R cells (46), adding another piece of evidence to the complex network of immune endocrine influences.
Recent experiments employing the same cell line led us to investigate the possible involvement of pro-inflammatory cytokines dealing with the anti-TB immune response, like IL- 1β or IFN-γ in in situ steroidogenesis. Whereas IFN-γ treatment produced no effect in this regard, IL-1β showed a subtle increase in steroidogenesis when added in combination with Forskolin, as judged by the increased production of cortisol and DHEA. When evaluating the possible mechanisms involved in this phenomenon, we provided evidence indicating that the NR4A family of nuclear receptors was likely to play a role compatible with the post-transcriptionally influence of IL-1β on steroidogenesis (47).
Work on dendritic cells showed that DHEA increases IL-12 production while reducing IL-10 secretion when these cells were exposed to Mtb favoring the expression of MHC I, MHC II, and CD86 and the phosphorylation of ERK1/2, which led to a better specific T cell performance in terms of proliferation and IFN-γ production (48).
Regarding macrophage function, we analyzed the effects of cortisol and DHEA on human THP-1 cells differentiated into macrophages infected with Mtb. Cortisol-exposed cultures showed a lower production of IL-1β and TNF-α, no matter DHEA was added or not. Nevertheless, intra-macrophage bacillary growth was reduced after DHEA treatment, in coincidence with increased autophagy (49); which is quite relevant since autophagy is not only influential for mycobacteria clearance but also for ameliorating tissue damage.
When exposing these cells to gamma-irradiated Mtb, DHEA increased the levels of reactive oxygen species (ROS)e. DHEA is likely to play an important role in the elimination of Mtb (49), both by autophagy and the production of oxidant species.
We also showed that phagocytic cells infected with Mtb and exposed overnight to DHEA in presence of cortisol, led to an increased expression of HBD-2 and HBD-3 transcripts if compared to unexposed counterparts (unpublished data). This effect correlated with increased amounts of HBD-2 and HBD-3 in 24 h supernatants along with a decrease in colony-forming units.
Concerning adaptive immunity, in vitro DHEA treatment increased the proportion of Mtb-specific CD8+ T cells and their terminal differentiation in HIV-TB coinfected patients. When studying Mtb-stimulated dendritic cells, DHEA improved their production of IL-12 and the ensuing antigen-specific T-cell responses (48, 50).
While evidence on the effects of DHEA in clinical TB is lacking, studies employing DHEA in animal models of Mtb infection are encouraging (51, 52).
Studies in an experimental model of progressive TB in BALB/c mice showed that at the time of maximal protective activity mediated by high production of IFN-γ, TNF-α, and IL-1β (day 21 after infection), there is a strong activation of the HPA axis mediated by high production of CRH in the hypothalamus and adrenal hyperplasia with high serum concentrations of corticosterone. Afterward, during the chronic or late phase of infection, there is progressive adrenal atrophy and a decrease in circulating corticosterone associated with extensive pneumonia (53).
Interestingly, during this late phase of pulmonary TB, there is a high conversion of inactive cortisone to active corticosterone by high activity of the enzyme 11-β-hydroxysteroid dehydrogenase type 1 (11-βHSD1), particularly overexpressed by highly infected macrophages (foamy macrophages). Thus, there is high pulmonary production of corticosterone, apparently to avoid tissue damage produced by excessive inflammation, but this high concentration of active glucocorticoid decreases Th1 cytokine production and promotes a Th2 response that impairs granuloma formation favoring disease progression (54). Studies in this BALB/c mouse pulmonary TB model showed a Th1➔Th2 cytokine pattern as the disease progresses, and such anomaly was partially reversed when treating mice with corticosterone and DHEA or AED in proper concentrations, along with a better Mtb-driven lymphoproliferation (55, 56). Because DHEA is metabolized to sex steroids, synthetic analogs of DHEA were generated, including 16α-bromoepiandrosterone (HE2000) which modulates immune and metabolic responses but does not display anabolic activities, rendering a more feasible and safer candidate drug for human use. Further work in the same murine model revealed a good therapeutic response mediated by the synthetic derivative of DHEA (57). In these studies, HE2000 reduced the bacterial load during progressive TB along with a decreased IL-4 expression. BALB/c mice with active TB given HE2000 also showed less bacterial proliferation and a better balance between Th1/Th2 cytokines. In the same sense, HE2000 administration lowered the amount of pneumonic involvement together with greater and rapid bacterial clearance, when given as an adjunct to conventional chemotherapy, suggesting that HE2000 could be useful to shorten antibiotic treatment (58).
Tuberculous mice treated with a new water-miscible formulation of 16α-bromoepiandrosterone showed similar results, besides a significant reduction of 11-βHSD1 activity and corticosterone production that efficiently reactivates the protective anti-TB immunity. Thus, 16α-bromoepiandrosterone can induce CD4 Th1 cells and macrophages activation by a direct activity, and by the suppression of the local production of corticosterone in the lungs (54).
A similar increase of the 11-βHSD1 activity with high cortisol production has been observed in the liver, skeletal muscles, and adipose tissue of patients with obesity and type 2 diabetes mellitus. Indeed, cortisol is a potent diabetogenic hormone, due to direct inhibition of the pancreatic secretion of insulin in addition to enhancing the glucose secretion by inhibiting gluconeogenesis in the liver as well as opposing insulin signaling and glucose uptake through a translocation inhibition of the glucose transporter GLUT4 to the plasma membrane (59). Like diabetic patients, diabetic BALB/c mice show more severe pulmonary TB with higher activity of 11-βHSD1 and corticosterone production in both lung and liver. Treatment with water-miscible 16α-bromoepiandrosterone decreased the expression of both enzymes and hormones in the lung and liver, correcting blood glucose concentration and decreasing pulmonary bacilli loads. At the experimental level, administration of 16α-bromoepiandrosterone seems to be an effective novel treatment for TB, particularly in the setting of the quite common diabetes comorbidity, providing a rationale for the implementation of clinical trials assessing its usefulness (54).
Table 1 summarizes DHEA’s beneficial effects on TB and Figure 1 shows DHEA’s effects on the anti-infectious immune response, particularly against Mtb.

Table 1 Beneficial Effects of DHEA in Tuberculosis.
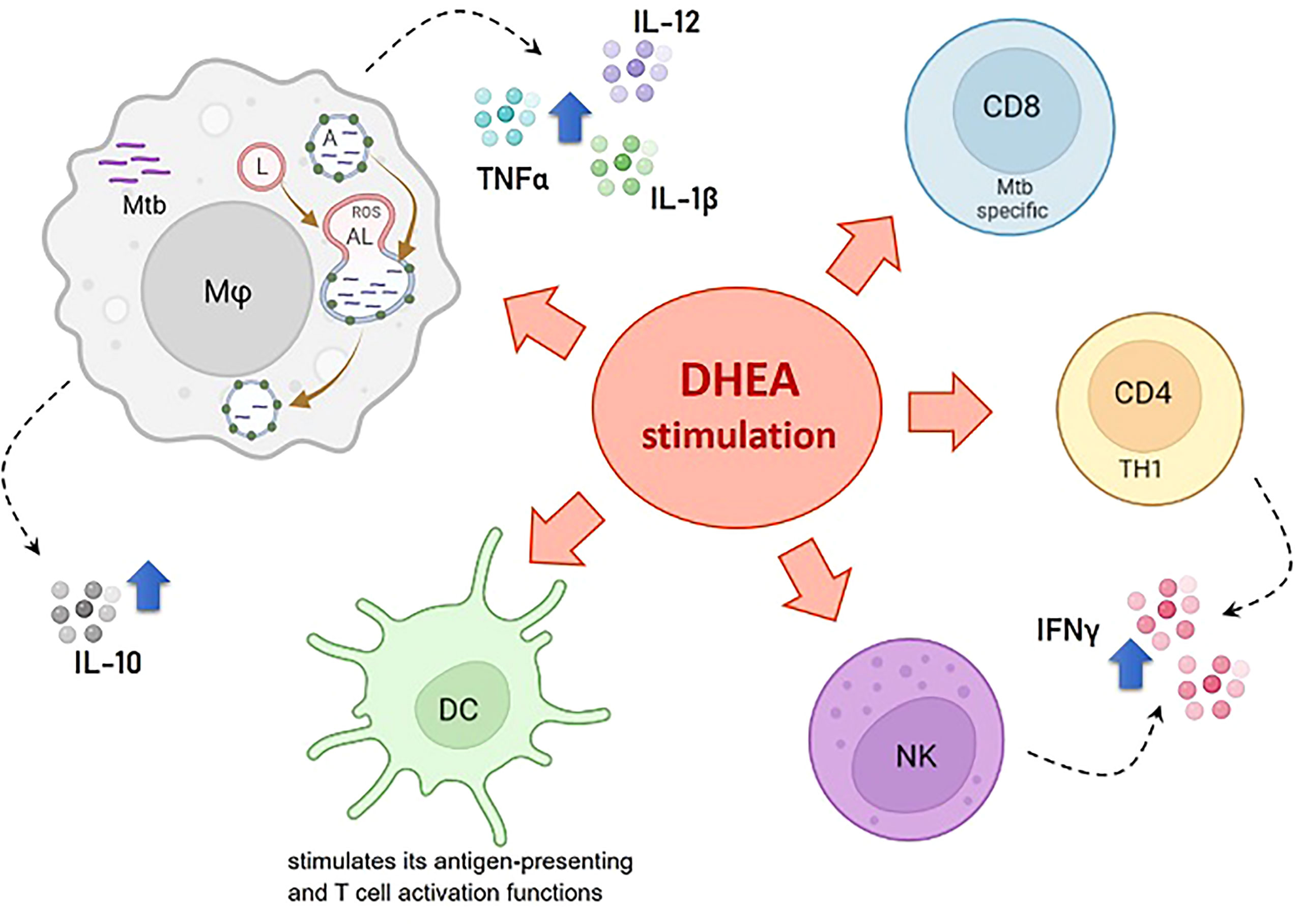
Figure 1 Effects of DHEA on the anti-infectious immune response particularly against M. tuberculosis. References: Mφ, macrophage; DC, dendritic cell; NK, natural killer; CD4 TH1, CD4+ cells with the TH1 cell profile; CD8, Mtb specific CD8+ T cells; Mtb, M. tuberculosis; ROS, reactive oxygen species; A, autophagosome; L, lysosome; AL, autophagolysosome. The blue arrow denotes stimulating effects.
The Potential of DHEA as an Adjuvant Therapy at the Clinical Level
DHEA exhibited favorable effects in autoimmune diseases such as systemic lupus erythematosus, rheumatoid arthritis, and inflammatory bowel disease (60–63).
Replacement DHEA studies also pointed out an encouraging role in the elderly (17, 56, 58, 64) wherein DHEA therapy may be an effective approach for preserving bone and muscle mass in women. In turn, DHEA treatment (50 mg/day for 3 months) improves self-reported well-being: sleep quality, general mood, vital energy, and stress management (65). At the immunological level, DHEA improved the plasma levels of monocytes (CD14), and NK cells (CD16, CD57), doubled the number of T gamma/delta cells, and the levels of the soluble IL2 receptor in plasma (66).
Women with Addison’s disease in whom DHEA was added to standard treatment showed a reduction in the number of circulating regulatory T cells as well as NK and NKT cells in presence of an increased lymphoproliferation and levels of IFN-γ, IL-5, IL-10, and TGF-β, along with a significant clinical improvement (67).
DHEA may be also beneficial in women’s infertility by augmenting fecundity and fertility (68–70), although some metanalysis failed to demonstrate a difference in pregnancy and miscarriage rates among those pretreated with DHEA and those who did not receive such treatment (71, 72).
Data from two randomized, double-blind, placebo-controlled studies on the effect of DHEA supplementation in women with systemic lupus erythematosus were consistent with the beneficial effects of DHEA (56, 61, 68, 73). One study reported that DHEA treatment (200 mg/d) improved the patient’s global assessment of disease activity (61); whereas in the other one the same DHEA dose served to reduce the dose of corticosteroids without affecting disease stability or even reducing its activity (73).
In a randomized, double-blind, placebo-controlled, small-scale study of 12-week duration, DHEA treatment improved the oxidative imbalance induced by hyperglycemia, downregulated the TNF-α/TNF-α receptor system, and prevented advanced glycation end-product formation, suggesting a beneficial effect on the onset and/or progression of chronic complications in type 2 diabetic patients (74).
As in many pharmacological developments, the issue of DHEA implementation in the clinical field should be carried out paying attention to its safety profile. Partly because of the possibility of DHEA conversion to testosterone, or estradiol, which might favor the development of prostate, ovary, or uterine neoplasms, particularly in elderly people (75–77). Although there are encouraging studies regarding the use of DHEA in the treatment of inflammatory pathologies and even neoplasms (78, 79).
The use of the above-mentioned synthetic DHEA analog may serve to circumvent some of these constraints. It is worth commenting that studies carried out a few years ago indicate that HE2000, exerted beneficial effects on HIV infection, at the level of the specific immune response, or a decreased TB coinfection in parallel with a fewer occurrence of opportunistic infections (54, 56, 80). In essence, the bulk of reviewed data set the basis for carrying out exploratory clinical studies aimed at assessing the potential benefit of adjuvant DHEA therapy in TB patients. Since two months of specific treatment in TB patients improves DHEA values to normality (41, 42), the length of adjuvant DHEA therapy would not be longer than at doses well established in other pathological settings [i.e., fertility stimulation studies in women 25-75 mg/day] (68, 69). Given our results in M. tuberculosis-infected macrophages, wherein the effect of DHEA seems to be related to mycobacterial elimination (49) it may be assumes that beneficial effects of DHEA will be apparent in the initial stages of therapeutical intervention.
Adjuvant therapy may offer a better way of treating tuberculosis not only in the field of drug resistance but also in non-adherence to chemotherapy and reducing the length of modern short-course chemotherapy, provided this approach turns out to be useful.
Author Contributions
All authors have written and edited the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by Fondo para la Investigación Científica y Tecnológica –FONCyT (PICT 2018- 02375), Argentina; Facultad de Ciencias Médicas –Universidad Nacional de Rosario, Argentina and Consejo Nacional de Ciencia y Tecnología, México, CONACyT, Grant/Award Number: 433346 and FC2015-1/115.
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Steinman L. Elaborate Interactions Between the Immune and Nervous Systems. Nat Immunol (2004) 5(6):575–81. doi: 10.1038/ni1078
2. Turnbull AV, Rivier CL. Regulation of the Hypothalamic-Pituitary-Adrenal Axis by Cytokines: Actions and Mechanisms of Action. Physiol Rev (1999) 79(1):1–71. doi: 10.1152/physrev.1999.79.1.1
3. Mueller JW, Gilligan LC, Idkowiak J, Arlt W, Foster PA. The Regulation of Steroid Action by Sulfation and Desulfation. Endocr Rev (2015) 36(5):526–63. doi: 10.1210/er.2015-1036
4. Kihel LE. Oxidative Metabolism of Dehydroepiandrosterone (DHEA) and Biologically Active Oxygenated Metabolites of DHEA and Epiandrosterone (EpiA)–recent Reports. Steroids (2012) 77(1–2):10–26. doi: 10.1016/j.steroids.2011.09.008
5. Schmidt M, Kreutz M, Löffler G, Schölmerich J, Sträub RH. Conversion of Dehydroepiandrosterone to Downstream Steroid Hormones in Macrophages. J Endocrinol (2000) 164(2):161–9. doi: 10.1677/joe.0.1640161
6. Webb SJ, Geoghegan TE, Prough RA, Miller KKM. The Biological Actions of Dehydroepiandrosterone Involves Multiple Receptors. Drug Metab Rev (2006) 38(1–2):89–116. doi: 10.1080/03602530600569877
7. Geraldes P, Sirois MG, Bernatchez PN, Tanguay JF. Estrogen Regulation of Endothelial and Smooth Muscle Cell Migration and Proliferation: Role of P38 and P42/44 Mitogen-Activated Protein Kinase. Arterioscler Thromb Vasc Biol (2002) 22(10):1585–90. doi: 10.1161/01.ATV.0000035393.11854.6A
8. Padgett DA, Loria RM, Sheridan JF. Endocrine Regulation of the Immune Response to Influenza Virus Infection With a Metabolite of DHEA-Androstenediol. J Neuroimmunol (1997) 78(1–2):203–11. doi: 10.1016/S0165-5728(97)00102-1
9. Straub RH, Schuld A, Mullington J, Haack M, Schölmerich J, Pollmächer T. The Endotoxin-Induced Increase of Cytokines is Followed by an Increase of Cortisol Relative to Dehydroepiandrosterone (DHEA) in Healthy Male Subjects. J Endocrinol (2002) 175(2):467–74. doi: 10.1677/joe.0.1750467
10. Araneo B, Daynes R. Dehydroepiandrosterone Functions as More Than an Antiglucocorticoid in Preserving Immunocompetence After Thermal Injury. Endocrinology (1995) 136(2):393–401. doi: 10.1210/endo.136.2.7835270
11. Cutolo M, Wilder RL. Different Roles for Androgens and Estrogens in the Susceptibility to Autoimmune Rheumatic Diseases. Rheum Dis Clin North Am (2000) 26(4):825–39. doi: 10.1016/S0889-857X(05)70171-X
12. Cutolo M, Straub RH. Sex Steroids and Autoimmune Rheumatic Diseases: State of the Art. Nat Rev Rheumatol (2020) 16(11):628–44. doi: 10.1038/s41584-020-0503-4
13. Loria RM, Inge TH, Cook SS, Szakal AK, Regelson W. Protection Against Acute Lethal Viral Infections With the Native Steroid Dehydroepiandrosterone (DHEA). J Med Virol (1988) 26(3):301–14. doi: 10.1002/jmv.1890260310
14. Ben-Nathan D, Padgett DA, Loria RM. Androstenediol and Dehydroepiandrosterone Protect Mice Against Lethal Bacterial Infections and Lipopolysaccharide Toxicity. J Med Microbiol (1999) 48(5):425–31. doi: 10.1099/00222615-48-5-425
15. Blauer KL, Poth M, Rogers WM, Bernton EW. Dehydroepiandrosterone Antagonizes the Suppressive Effects of Dexamethasone on Lymphocyte Proliferation. Endocrinology (1991) 129(6):3174–9. doi: 10.1210/endo-129-6-3174
16. Suzuki T, Suzuki N, Daynes RA, Engleman EG. Dehydroepiandrosterone Enhances IL2 Production and Cytotoxic Effector Function of Human T Cells. Clin Immunol Immunopathol (1991) 61(2 Pt 1):202–11. doi: 10.1016/S0090-1229(05)80024-8
17. Kidd P. Th1/Th2 Balance: The Hypothesis, Its Limitations, and Implications for Health and Disease. Altern Med Rev (2003) 8(3):223–46.
18. Martıínez-Taboada V, Bartolomé MJ, Amado JA, Blanco R, Garcıía-Unzueta MT, Rodrıíguez-Valverde V, et al. Changes in Peripheral Blood Lymphocyte Subsets in Elderly Subjects Are Associated With an Impaired Function of the Hypothalamic-Pituitary-Adrenal Axis. Mech Ageing Dev (2002) 123(11):1477–86. doi: 10.1016/S0047-6374(02)00116-1
19. Hammer F, Drescher DG, Schneider SB, Quinkler M, Stewart PM, Allolio B, et al. Sex Steroid Metabolism in Human Peripheral Blood Mononuclear Cells Changes With Aging. J Clin Endocrinol Metab (2005) 90(11):6283–9. doi: 10.1210/jc.2005-0915
20. Straub RH, Paimela L, Peltomaa R, Schölmerich J, Leirisalo-Repo M. Inadequately Low Serum Levels of Steroid Hormones in Relation to Interleukin-6 and Tumor Necrosis Factor in Untreated Patients With Early Rheumatoid Arthritis and Reactive Arthritis. Arthritis Rheumatol (2002) 46(3):654–62. doi: 10.1002/art.10177
21. Bentley C, Hazeldine J, Greig C, Lord J, Foster M. Dehydroepiandrosterone: A Potential Therapeutic Agent in the Treatment and Rehabilitation of the Traumatically Injured Patient. Burns Trauma (2019) 2:7–26. doi: 10.1186/s41038-019-0158-z
22. Reed MJ, Purohit A, Woo LWL, Newman SP, Potter BVL. Steroid Sulfatase: Molecular Biology, Regulation, and Inhibition. Endocr Rev (2005) 26(2):171–202. doi: 10.1210/er.2004-0003
23. Daynes RA, Dudley DJ, Araneo BA. Regulation of Murine Lymphokine Production In Vivo II. Dehydroepiandrosterone Is a Natural Enhancer of Interleukin 2 Synthesis by Helper T Cells. Eur J Immunol (1990) 20(4):793–802. doi: 10.1002/eji.1830200413
24. Huang KUN, Dudley D, Daynes RA., Araneo BA., Dowell TA. Regulation of Murine Lymphokine Production In Vivo III . The Lymphoid Tissue Microenvironment Exerts Regulatory Influences Over T Helper Cell Function. Cytokines (1990) 171:979–96. doi: 10.1084/jem.171.4.979
25. Hennebold JD. Regulation of Macrophage Dehydroepiandrosterone Sulfate Metabolism by Inflammatory Cytokines. Endocrinology (1994) 135(1):67–75. doi: 10.1210/endo.135.1.8013393
26. Dluhy RG. The Growing Spectrum of HIV-Related Endocrine Abnormalities. J Clin Endocrinol Metab (1990) 70(3):563–5. doi: 10.1210/jcem-70-3-563
27. Reincke M, Heppner C, Petzke F, Allolio B, Arlt W, Mbulamberi D, et al. Impairment of Adrenocortical Function Associated With Increased Plasma Tumor Necrosis Factor-Alpha and Interleukin-6 Concentrations in African Trypanosomiasis. Neuroimmunomodulation (1994) 1(1):14–22. doi: 10.1159/000095930
28. Bozza VV, D’Attilio L, Mahuad CV, Giri AA, del Rey A, Besedovsky H, et al. Altered Cortisol/DHEA Ratio in Tuberculosis Patients and its Relationship With Abnormalities in the Mycobacterial-Driven Cytokine Production by Peripheral Blood Mononuclear Cells. Scand J Immunol (2007) 66(1):97–103. doi: 10.1111/j.1365-3083.2007.01952.x
29. Cárdenas G, Valdez R, Sáenz B, Bottasso O, Fragoso G, Sciutto E, et al. Impact of Taenia Solium Neurocysticercosis Upon Endocrine Status and its Relation With Immuno-Inflammatory Parameters. Int J Parasitol (2012) 42(2):171–6. doi: 10.1016/j.ijpara.2011.11.009
30. Rook GAW. Glucocorticoids and Immune Function. Baillieres Best Pract Res Clin Endocrinol Metab (1999) 13(4):567–81. doi: 10.1053/beem.1999.0044
31. Weidler C, Struharova S, Schmidt M, Ugele B, Schölmerich J, Straub RH. Tumor Necrosis Factor Inhibits Conversion of Dehydroepiandrosterone Sulfate (DHEAS) to DHEA in Rheumatoid Arthritis Synovial Cells: A Prerequisite for Local Androgen Deficiency. Arthritis Rheumatol (2005) 52(6):1721–9. doi: 10.1002/art.21112
32. Straub RH, Pongratz G, Schölmerich J, Kees F, Schaible TF, Antoni C, et al. Long-Term Anti-Tumor Necrosis Factor Antibody Therapy in Rheumatoid Arthritis Patients Sensitizes the Pituitary Gland and Favors Adrenal Androgen Secretion. Arthritis Rheumatol (2003) 48(6):1504–12. doi: 10.1002/art.11036
33. Straub RH, Lehle K, Herfarth H, Weber M, Falk W, Preuner J, et al. Dehydroepiandrosterone in Relation to Other Adrenal Hormones During an Acute Inflammatory Stressful Disease State Compared With Chronic Inflammatory Disease: Role of Interleukin-6 and Tumour Necrosis Factor. Eur J Endocrinol (2002) 146(3):365–274. doi: 10.1530/eje.0.1460365
34. Brazão V, Santello FH, Caetano LC, Del Vecchio Filipin M, Paula Alonso Toldo M, do Prado JC. Immunomodulatory Effects of Zinc and DHEA on the Th-1 Immune Response in Rats Infected With Trypanosoma Cruzi. Immunobiology (2010) 215(5):427–34. doi: 10.1016/j.imbio.2009.05.005
35. Rasmussen KR, Martin EG, Healey MC. Effects of Dehydroepiandrosterone in Immunosuppressed Rats Infected With Cryptosporidium Parvum. J Parasitol (1993) 79(3):364–70. doi: 10.2307/3283571
36. Khalifa AM, Ibrahim IR, El-Kerdany ED. Coccidial Infection in Immunosuppressed Mice: Prophylaxis and Treatment With Dehydroepiandrosterone. East Mediterr Health J (2000) 6(5):908–18. doi: 10.26719/2000.6.5-6.908
37. Leenstra T, ter Kuile FO, Kariuki SK, Nixon CP, Oloo AJ, Kager PA, et al. Dehydroepiandrosterone Sulfate Levels Associated With Decreased Malaria Parasite Density and Increased Hemoglobin Concentration in Pubertal Girls From Western Kenya. J Infect Dis (2003) 188(2):297–304. doi: 10.1086/376508
38. Bottasso O, Bay ML, Besedovsky H, del Rey A. Adverse Neuro-Immune-Endocrine Interactions in Patients With Active Tuberculosis. Mol Cell Neurosci (2013) 53:77–85. doi: 10.1016/j.mcn.2012.11.002
39. Rey Ad, Mahuad CV, Bozza VV, Bogue C, Farroni MA, Bay ML, et al. Endocrine and Cytokine Responses in Humans With Pulmonary Tuberculosis. Brain Behav Immun (2007) 21(2):171–9. doi: 10.1016/j.bbi.2006.06.005
40. Mahuad C, Bay ML, Farroni MA, Bozza V, Del Rey A, Besedovsky H, et al. Cortisol and Dehydroepiandrosterone Affect the Response of Peripheral Blood Mononuclear Cells to Mycobacterial Antigens During Tuberculosis. Scand J Immunol (2004) 60(6):639–46. doi: 10.1111/j.0300-9475.2004.01514.x
41. Bongiovanni B, Díaz A, D'Attilio L, Santucci N, Dídoli G, Lioi S, et al. Changes in the Immune and Endocrine Responses of Patients With Pulmonary Tuberculosis Undergoing Specific Treatment. Ann N Y Acad Sci (2012) 1262:10–5. doi: 10.1111/j.1749-6632.2012.06643.x
42. Díaz A, Bongiovanni B, D’Attilio L, Santucci N, Dídoli G, Fernández R del V, et al. The Clinical Recovery of Tuberculosis Patients Undergoing Specific Treatment is Associated With Changes in the Immune and Neuroendocrine Responses. Pathog Dis (2017) 75(7):1–11. doi: 10.1093/femspd/ftx087
43. Bongiovanni B, Marín-Luevano S, D’Attilio L, Díaz A, Fernández R del V, Santucci N, et al. Evidence That Changes in Antimicrobial Peptides During Tuberculosis are Related to Disease Severity, Clinical Presentation, Specific Therapy and Levels of Immune-Endocrine Mediators. Cytokine (2020) 126:154913. doi: 10.1016/j.cyto.2019.154913
44. Quiroga MF, Angerami MT, Santucci N, Ameri D, Francos JL, Wallach J, et al. Dynamics of Adrenal Steroids are Related to Variations in Th1 and Treg Populations During Mycobacterium Tuberculosis Infection in HIV Positive Persons. PloS One (2012) 7(3):e33061. doi: 10.1371/journal.pone.0033061
45. Bruns H, Meinken C, Schauenberg P, Härter G, Kern P, Modlin RL, et al. Anti-TNF Immunotherapy Reduces CD8+ T Cell-Mediated Antimicrobial Activity Against Mycobacterium Tuberculosis in Humans. J Clin Invest (2009) 119(5):1167–77. doi: 10.1172/JCI38482
46. D'Attilio L, Bozza VV, Santucci N, Bongiovanni B, Dídoli G, Radcliffe S, et al. TGF-β Neutralization Abrogates the Inhibited DHEA Production Mediated by Factors Released From M. Tuberculosis-Stimulated PBMC. Ann N Y Acad Sci (2012) 1262(1):1–9. doi: 10.1111/j.1749-6632.2012.06644.x
47. Santucci N, Stampone R, Brandão Ferreira Da Silva E, Villar S, Spinelli S, Linhares-Lacerda L, et al. Local Regulation of Adrenal Steroidogenesis: Subtle In Vitro Effects of IL-1β on the Human Cell Line NCI-H295R Steroid Production Along With Changes in MicroRNA Profile and Orphan Nuclear Receptors NR4As. Neuroimmunomodulation (2020) 27(3):131–41. doi: 10.1159/000512325
48. Angerami M, Suarez G, Pascutti MF, Salomon H, Bottasso O, Quiroga MF. Modulation of the Phenotype and Function of Mycobacterium Tuberculosis-Stimulated Dendritic Cells by Adrenal Steroids. Int Immunol (2013) 25(7):405–11. doi: 10.1093/intimm/dxt004
49. Bongiovanni B, Mata-Espinosa D, D’Attilio L, Leon-Contreras JC, Marquez-Velasco R, Bottasso O, et al. Effect of Cortisol and/or DHEA on THP1-Derived Macrophages Infected With Mycobacterium Tuberculosis. Tuberculosis (2015) 95(5):562–9. doi: 10.1016/j.tube.2015.05.011
50. Suarez GV, Angerami MT, Vecchione MB, Laufer N, Turk G, Ruiz MJ, et al. HIV-TB Coinfection Impairs CD8(+) T-Cell Differentiation and Function While Dehydroepiandrosterone Improves Cytotoxic Antitubercular Immune Responses. Eur J Immunol (2015) 45(9):2529–41. doi: 10.1002/eji.201545545
51. Hernandez-Pando R, de la Luz Streber M, Orozco H, Arriaga K, Pavon L, Marti O, et al. Emergent Immunoregulatory Properties of Combined Glucocorticoid and Anti-Glucocorticoid Steroids in a Model of Tuberculosis. QJM (1998) 91(11):755–66. doi: 10.1093/qjmed/91.11.755
52. Hernandez-Pando R, de la Luz Streber M, Orozco H, Arriaga K, Pavon L, Al-Nakhli SA, et al. The Effects of Androstenediol and Dehydroepiandrosterone on the Course and Cytokine Profile of Tuberculosis in BALB/c Mice. Immunology (1998) 95(2):234–41. doi: 10.1046/j.1365-2567.1998.00601.x
53. Hernandez-Pando R, Orozco H, Honour J, Silva P, Leyva R, Rook GA. Adrenal Changes in Murine Pulmonary Tuberculosis; a Clue to Pathogenesis? FEMS Immunol Med Microbiol (1995) 12(1):63–72. doi: 10.1111/j.1574-695X.1995.tb00176.x
54. López-Torres MO, Marquina-Castillo B, Ramos-Espinosa O, Mata-Espinosa D, Barrios-Payan JA, Baay-Guzman G, et al. 16α-Bromoepiandrosterone as a New Candidate for Experimental Diabetes-Tuberculosis Co-Morbidity Treatment. Clin Exp Immunol (2021) 205(2):232–45. doi: 10.1111/cei.13603
55. Binello E, Gordon CM. Clinical Uses and Misuses of Dehydroepiandrosterone. Curr Opin Pharmacol (2003) 3(6):635–41. doi: 10.1016/j.coph.2003.07.006
56. Dillon JS. Dehydroepiandrosterone, Dehydroepiandrosterone Sulfate and Related Steroids: Their Role in Inflammatory, Allergic and Immunological Disorders. Curr Drug Targets Inflamm Allergy (2005) 4(3):377–85. doi: 10.2174/1568010054022079
57. Ribeiro F, Lopes R, Nunes C, Maito F, Bonorino C, Bauer M. Dehydroepiandrosterone Sulphate Enhances IgG and Interferon-Gamma Production During Immunization to Tuberculosis in Young But Not Aged Mice Fla. Biogerontology (2007) 8(2):209–20. doi: 10.1007/s10522-006-9069-z
58. Hernández-Pando R, Aguilar-Leon D, Orozco H, Serrano A, Ahlem C, Trauger R, et al. 16alpha-Bromoepiandrosterone Restores T Helper Cell Type 1 Activity and Accelerates Chemotherapy-Induced Bacterial Clearance in a Model of Progressive Pulmonary Tuberculosis. J Infect Dis (2005) 191(2):299–306. doi: 10.1086/426453
59. Shukla R, Basu AK, Mandal B, Mukhopadhyay P, Maity A, Chakraborty S, et al. 11β Hydroxysteroid Dehydrogenase - 1 Activity in Type 2 Diabetes Mellitus: A Comparative Study. BMC Endocr Disord (2019) 19(1):15. doi: 10.1186/s12902-019-0344-9
60. Stickney DR, Noveljic Z, Garsd A, Destiche DA, Frincke JM. Safety and Activity of the Immune Modulator HE2000 on the Incidence of Tuberculosis and Other Opportunistic Infections in AIDS Patients. Antimicrob Agents Chemother (2007) 51(7):2639–41. doi: 10.1128/AAC.01446-06
61. Chang DM, Lanv JL, Lin HY, Luo SF. Dehydroepiandrosterone Treatment of Women With Mild-To-Moderate Systemic Lupus Erythematosus: A Multicenter Randomized, Double-Blind, Placebo-Controlled Trial. Arthritis Rheumatol (2002) 46(11):2924–7. doi: 10.1002/art.10615
62. Andus T, Klebl F, Rogler G, Bregenzer N, Schölmerich J, Straub RH. Patients With Refractory Crohn’s Disease or Ulcerative Colitis Respond to Dehydroepiandrosterone: A Pilot Study. Aliment Pharmacol Ther (2003) 17(3):409–14. doi: 10.1046/j.1365-2036.2003.01433.x
63. Rutkowski K, Sowa P, Rutkowska-Talipska J, Kuryliszyn-Moskal A, Rutkowski R. Dehydroepiandrosterone (DHEA): Hypes and Hopes. Drugs (2014) 74(11):1195–207. doi: 10.1007/s40265-014-0259-8
64. de la Torre B, Hedman M. Blood and Tissue Dehydroepiandrosterone Sulphate Levels and Their Relationship to Chronic Inflammatory Bowel Disease. Clin Exp Rheumatol (1998) 16(5):579–82.
65. Morales AJ, Nolan JJ, Nelson JC, Yen SSC. Effects of Replacement Dose of Dehydroepiandrosterone in Men and Women of Advancing Age. J Clin Endocrinol Metab (1994) 78(6):1360–7. doi: 10.1210/jcem.78.6.7515387
66. Yen SSC, Morales AJ, Khorram O. Replacement of DHEA in Aging Men and Women. Potential Remedial Effects. Ann N Y Acad Sci (1995) 774(1):128–42. doi: 10.1111/j.1749-6632.1995.tb17377.x
67. Coles AJ, Thompson S, Cox AL, Curran S, Gurnell EM, Chatterjee VK. Dehydroepiandrosterone Replacement in Patients With Addison’s Disease has a Bimodal Effect on Regulatory (CD4+CD25hi and CD4+FoxP3+) T Cells. Eur J Immunol (2005) 35(12):3694–703. doi: 10.1002/eji.200526128
68. Fusi FM, Ferrario M, Bosisio C, Arnoldi M, Zanga L. DHEA Supplementation Positively Affects Spontaneous Pregnancies in Women With Diminished Ovarian Function. Gynecol Endocrinol (2013) 29(10):940–3. doi: 10.3109/09513590.2013.819087
69. Kara M, Aydin T, Aran T, Turktekin N, Ozdemir B. Does Dehydroepiandrosterone Supplementation Really Affect IVF-ICSI Outcome in Women With Poor Ovarian Reserve? Eur J Obstet Gynecol Reprod Biol (2014) 173(1):63–5. doi: 10.1016/j.ejogrb.2013.11.008
70. Strauss S, Greve T, Ernst E, Fraidakis M, Grudzinskas JG, Andersen CY. Administration of DHEA Augments Progesterone Production in a Woman With Low Ovarian Reserve Being Transplanted With Cryopreserved Ovarian Tissue. J Assist Reprod Genet (2014) 31(6):645–9. doi: 10.1007/s10815-014-0214-3
71. Sunkara SK, Pundir J, Khalaf Y. Effect of Androgen Supplementation or Modulation on Ovarian Stimulation Outcome in Poor Responders: A Meta-Analysis. Reprod BioMed Online (2011) 22(6):545–55. doi: 10.1016/j.rbmo.2011.01.015
72. Narkwichean A, Maalouf W, Campbell BK, Jayaprakasan K. Efficacy of Dehydroepiandrosterone to Improve Ovarian Response in Women With Diminished Ovarian Reserve: A Meta-Analysis. Reprod Biol Endocrinol (2013) 11:44. doi: 10.1186/1477-7827-11-44
73. Petri MA, Lahita RG, Van Vollenhoven RF, Merrill JT, Schiff M, Ginzler EM, et al. GL601 Study Group. Effects of Prasterone on Corticosteroid Requirements of Women With Systemic Lupus Erythematosus: A Double-Blind, Randomized, Placebo-Controlled Trial. Arthritis Rheumatol (2002) 46(7):1820–9. doi: 10.1002/art.10364
74. Brignardello E, Runzo C, Aragno M, Catalano MG, Cassader M, Perin PC, et al. Dehydroepiandrosterone Administration Counteracts Oxidative Imbalance and Advanced Glycation End Product Formation in Type 2 Diabetic Patients. Diabetes Care (2007) 30(11):2922–7. doi: 10.2337/dc07-1110
75. Nippoldt TB, Nair KS. Is There a Case for DHEA Replacement? Baillieres Clin Endocrinol Metab (1998) 12(3):507–20. doi: 10.1016/S0950-351X(98)80286-3
76. Nair KS, Rizza RA, O’Brien P, Dhatariya K, Short KR, Nehra A, et al. DHEA in Elderly Women and DHEA or Testosterone in Elderly Men. N Engl J Med (2006) 355(16):1647–59. doi: 10.1056/NEJMoa054629
77. Arnold JT, Blackman MR. Does DHEA Exert Direct Effects on Androgen and Estrogen Receptors, and Does it Promote or Prevent Prostate Cancer? Endocrinology (2005) 146(11):4565–7. doi: 10.1210/en.2005-0901
78. Aoki K, Terauchi Y. Effect of Dehydroepiandrosterone (DHEA) on Diabetes Mellitus and Obesity. Vitam Horm (2018) 108:355–65. doi: 10.1016/bs.vh.2018.01.008
79. Colín-Val Z, López-Díazguerrero NE, López-Marure R. DHEA Inhibits Proliferation, Migration and Alters Mesenchymal-Epithelial Transition Proteins Through the PI3K/Akt Pathway in MDA-MB-231 Cells. J Steroid Biochem Mol Biol (2021) 208:105818. doi: 10.1016/j.jsbmb.2021.105818
80. Reading C, Dowding C, Schramm B, Garsd A, Onizuka-Handa N, Stickney D, et al. Improvement in Immune Parameters and Human Immunodeficiency Virus-1 Viral Response in Individuals Treated With 16alpha-Bromoepiandrosterone (He2000). Clin Microbiol Infect (2006) 12(11):1082–8. doi: 10.1111/j.1469-0691.2006.01520.x
Keywords: immunoendocrinology, adrenal hormones, infection disease, Dehydroepiandrosterone (DH EA), Tuberculosis
Citation: Bongiovanni B, Díaz A, Santucci N, D’Attilio LD, Bottasso O, Hernández Pando R and Bay ML (2022) The Immunoregulatory Actions of DHEA in Tuberculosis, A Tool for Therapeutic Intervention? Front. Endocrinol. 13:892270. doi: 10.3389/fendo.2022.892270
Received: 08 March 2022; Accepted: 03 May 2022;
Published: 06 June 2022.
Edited by:
Leandro Kasuki, Instituto Estadual do Cérebro Paulo Niemeyer (IECPN), BrazilReviewed by:
Pandurangan Ramaraj, A.T. Still University, United StatesMoisés Evandro Bauer, Pontifical Catholic University of Rio Grande do Sul, Brazil
Copyright © 2022 Bongiovanni, Díaz, Santucci, D’Attilio, Bottasso, Hernández Pando and Bay. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: María Luisa Bay, YmF5QGlkaWNlci1jb25pY2V0LmdvYi5hcg==; Rogelio Hernández Pando, cmhkZXpwYW5kb0Bob3RtYWlsLmNvbQ==