 John T. Phoenix1,2
John T. Phoenix1,2 Audris Budreika1
Audris Budreika1 Raymond J. Kostlan1,2
Raymond J. Kostlan1,2 Justin H. Hwang3
Justin H. Hwang3 Sean W. Fanning1
Sean W. Fanning1 Steven Kregel1*
Steven Kregel1*- 1Department of Cancer Biology, Stritch School of Medicine, Loyola University Chicago, Maywood, IL, United States
- 2Integrated Program in Biomedical Science, Biochemistry, Molecular and Cancer Biology, Loyola University Chicago, Maywood, IL, United States
- 3Department of Medicine, University of Minnesota, Minneapolis, MN, United States
Editorial on the Research Topic
Hormone resistance in cancer
Introduction
Hormone-dependent cancers are the most common non-cutaneous tumors experienced by all sexes. This year, nearly 700,000 combined new cases of breast, ovarian, endometrial, prostate, and thyroid cancers will be diagnosed in the United States (1). Antagonizing hormone signaling pathways is a widely used treatment strategy (2–6). However, due to acquired mutations of hormone receptors, indirect cofactor-mediated changes in cell behavior, and heterogeneity within tumors, the therapeutic durability of these treatments is often short-lived (7–13). In late-stage disease, a lethal, intractable small-blue cell tumor or neuroendocrine-like phenotype may emerge displaying genetic, epigenetic, and hormonal properties promoting cellular plasticity (11, 12, Imamura et al., 14). Specific neuroendocrine tumor features are poorly defined and vary across tissue origin (14, 15). Therefore, understanding these mechanisms of treatment resistance and finding commonalities among neuroendocrine subsets of cancer is vital to uncover new targeted therapies beyond hormone therapy that halt disease progression.
Hormones and cancers
Hormone receptor signaling
Hormones often serve as activating ligands for their respective nuclear hormone receptor (NHR) proteins: Estrogen Receptor (ER) in breast, ovarian, and endometrial tissue, and Androgen Receptor (AR) in prostate tissue (16, 17). Thyroid carcinomas are often fueled by Thyroid–Stimulating Hormone (TSH) produced in the pituitary gland, responsible for the endocrine secretion of many hormones (Mousa et al.). NHRs act as potent activators of oncogenes in transformed cells (4, 17). Ultimately, maintenance of hormone signaling pathways despite attempted blockades is a mechanism of disease progression (18–20).
Hormone therapy in cancer
Targeting hormone signaling pathways are a clinical mainstay for treating hormone–dependent cancers. Exogenous L–Thyroxine (T4) is a common treatment to promote a negative feedback loop of TSH signals in papillary and follicular thyroid cancers (Mousa et al.). Similarly, gonadotropin releasing hormone (GnRH) antagonists, or agonists through feedback, castrate prostate cancer patients by halting testicular production of androgens (21). Androgen and estrogen biosynthesis inhibitors, abiraterone acetate and aromatase inhibitors, respectively, inhibit enzymes that synthesize hormones in patients (22, 23). Second–generation AR antagonists like enzalutamide and darolutamide impede AR interaction with testosterone (24). ER–targeted therapies include the Selective Estrogen Receptor Modulators/Degraders (SERM/Ds). SERMs tamoxifen and raloxifene bind to the ER ligand binding domain and obstruct ER signaling breast tissue (25, 26). Selective Estrogen Receptor Degraders (SERDs) bind and target ER for proteasomal degradation (27, 28).
Hormone therapy resistance
Unfortunately, the response to hormone therapy is temporary. As discussed in Mousa et al., L–Thyroxine may in fact stimulate tumor cell proliferation in patients with therapy–resistant thyroid cancer. Breast and prostate tumors often exhibit NHR mutations that render most treatments ineffective, including activating mutations where enzalutamide and tamoxifen can serve as functional NHR ligands. (7, 8, 20, 29–33). Cancer cells employ growth pathways aside from NHR signaling and utilize alternative cofactors and coregulatory molecules promoting disease progression (2, 9, 10, 34–38). Since NHR family proteins are structurally similar, other NHRs such as Glucocorticoid Receptor (GR) can compensate for loss of AR/ER activity (39, 40). These resistance mechanisms allow for initially hormone–driven tumors to become metastatic, hormone–indifferent disease (18). Imamura et al. found these cancers often display a high degree of lineage–plasticity and sometimes complete loss of NHR–dependence in Neuroendocrine Prostate Cancer (NEPC) and Triple–Negative Breast Cancer (TNBC) (41).
Neuroendocrine differentiation and hormone independence
Neuroendocrine tumor characteristics
Since very few neuroendocrine molecular markers exist across tumor types, specifically defining these populations and targeting these poorly differentiated, aggressive cancer cells remains elusive. Neuroendocrine tumors typically display markers of neuronal differentiation and can originate in various anatomical locations. Nevertheless, they exhibit histological and clinical resemblances (42). Neuroendocrine subsets seen in late–stage cancers are poorly differentiated, with large nuclear–to–cytoplasmic ratios, that aberrantly activate stem cell gene pathways (34, 43, 44).
Some genetic similarities are shared across various small–cell neuroendocrine (SCN) diseases such as small–cell lung cancer (SCLC) and NEPC, regardless of their tissue of origin (44). These can include RB1 deletion, TP53 mutation, and N–MYC overexpression (42, 44–47). In hormone dependent tumors, de novo incidence of SCN/NEPC is rare, and most cases result from therapeutic pressure (48, 49). Further complicating the topic is that neuroendocrine neoplasms occur in many sites: the central nervous system, respiratory tract, gastrointestinal tract, thyroid, breast, and urogenital system, and yet share similar pathologic features (50). Although progress has been made in the management of these lineage plastic neuroendocrine cancers, such as Delta–like–ligand–3 (DLL3) targeted molecules (51, 52), more research is needed to further characterize the exact molecular mechanisms of progression to an SCN phenotype (Figure 1).
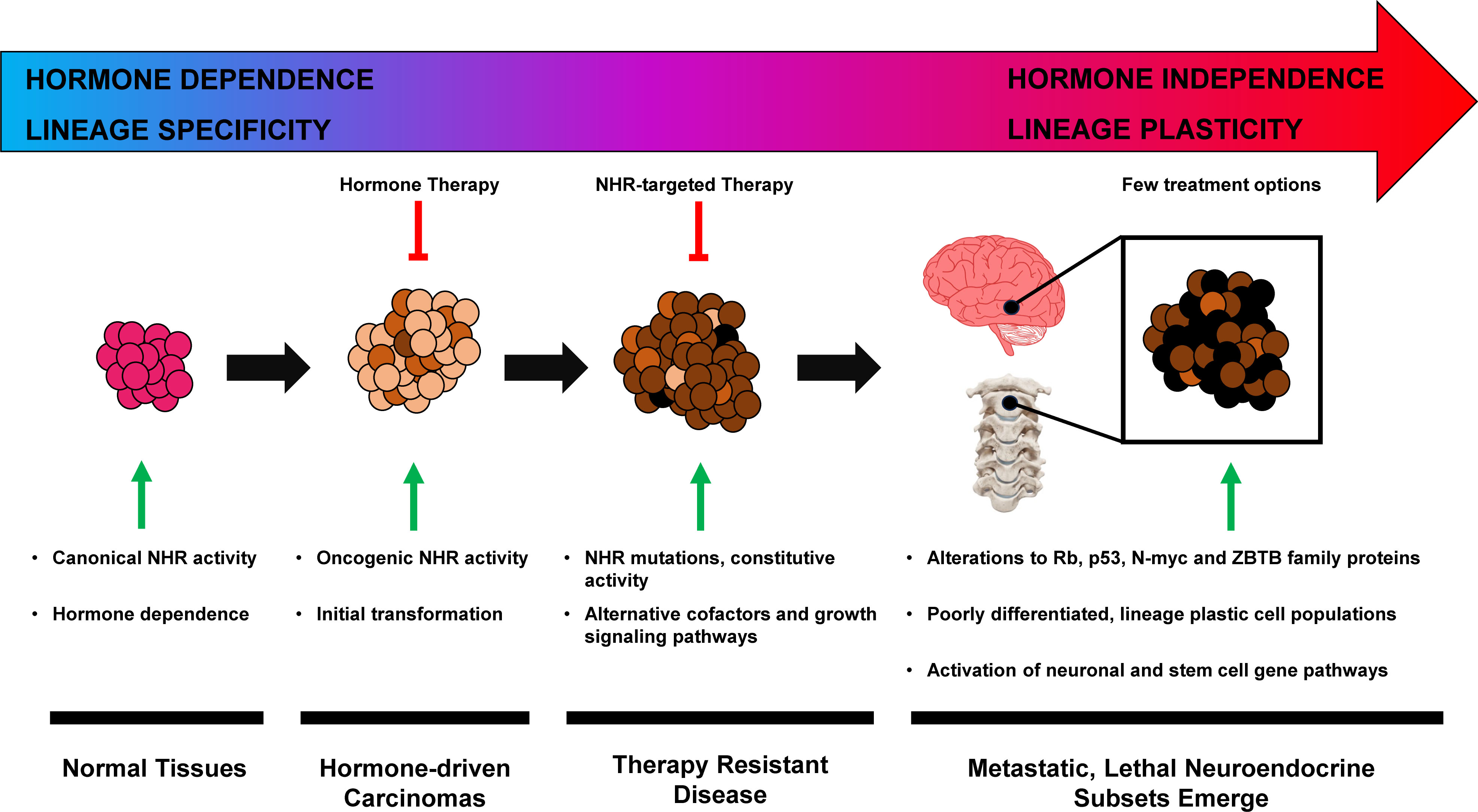
Figure 1 Hormone–driven cancer progression to neuroendocrine phenotype.
Emerging factors in SCN tumors
A recently developed SCN phenotype grading system was used by Bae et al. to demonstrate oncogenic function of the transcription factor ZBTB7A in NEPC (42). ZBTB (Zinc finger and broad–complex, tramtrack and bric–a–brac domain containing) protein family members are multifunctional transcription factors that play significant roles cell proliferation, differentiation, and development. (53, 54). Elevated expression of ZBTB7A has been linked to tumor formation and metastasis in different cancer types, including breast, prostate, lung, ovarian, and colon cancer (55, 56). Interestingly, pituitary neuroendocrine tumors (PitNETs) are characterized by overexpression of prolactin, Studies have demonstrated that ZBTB20 plays a role in controlling prolactin expression in the pituitary gland and contributes to the development of hepatocellular carcinoma (57–59). Lin et al. describe a patient in whom a PitNET had developed resistance to standard of care therapy was treated with hydroxychloroquine and experienced a significant reduction in prolactin secretion. Given the established crosstalk between prolactin and estrogen receptor signaling in breast cancer, ZBTB family transcription factors could play key roles in the emergence and therapy resistance neuroendocrine cancers (37, Bae et al., 60). These factors may drive lineage–plasticity across tumor types and the loss of NHR–dependence that characterizes SNC/NEPC.
Summary
Hormone driven cancers are common malignancies that sometimes differentiate into lethal, aggressive neuroendocrine subtypes. In breast and prostate cancer, neuroendocrine differentiation often emerges after the failure of hormone targeted therapies. Currently, a limited number of dependable neuroendocrine molecular markers exist across tissues. Further research is required to discover genetic similarities that can pave the way for effective targeted therapies capable of eradicating multiple subtypes of neuroendocrine and lineage plastic tumors.
Author contributions
JP: Conceptualization, Writing – original draft, Writing – review & editing. AB: Conceptualization, Writing – original draft, Writing – review & editing. RK: Conceptualization, Writing – original draft, Writing – review & editing. JH: Conceptualization, Writing – original draft, Writing – review & editing. SF: Conceptualization, Writing – original draft, Writing – review & editing. SK: Conceptualization, Project administration, Supervision, Validation, Writing – original draft, Writing – review & editing.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Siegel RL, Miller KD, Wagle NS, Jemal A. Cancer statistics, 2023. CA Cancer J Clin (2023) 73:17–48. doi: 10.3322/caac.21763
2. Chmelar R, Buchanan G, Need EF, Tilley W, Greenberg NM. Androgen receptor coregulators and their involvement in the development and progression of prostate cancer. Int J Cancer (2007) 120:719–33. doi: 10.1002/ijc.22365
3. Scher HI, Fizazi K, Saad F, Taplin ME, Sternberg CN, Miller K, et al. Increased survival with enzalutamide in prostate cancer after chemotherapy. N Engl J Med (2012) 367:1187–97. doi: 10.1056/NEJMoa1207506
4. Yager JD, Davidson NE. Estrogen carcinogenesis in breast cancer. N Engl J Med (2006) 354:270–82. doi: 10.1056/NEJMra050776
5. Early Breast Cancer Trialists’ Collaborative G. Effects of chemotherapy and hormonal therapy for early breast cancer on recurrence and 15–year survival: an overview of the randomised trials. Lancet (2005) 365:1687–717. doi: 10.1016/S0140-6736(05)66544-0
6. Huggins C. Studies on prostate cancer II The effects of castration on advanced carcinoma of the prostate gland. Arch Surg (1941) 43:209–23. doi: 10.1001/archsurg.1941.01210140043004
7. Fanning SW, Mayne CG, Dharmarajan V, Carlson KE, Martin TA, Novick SJ, et al. Estrogen receptor alpha somatic mutations Y537S and D538G confer breast cancer endocrine resistance by stabilizing the activating function–2 binding conformation. Elife (2016) 5:1–2. doi: 10.7554/eLife.12792
8. Prekovic S, van Royen ME, Voet AR, Geverts B, Houtman R, Melchers D, et al. The effect of F877L and T878A mutations on androgen receptor response to enzalutamide. Mol Cancer Ther (2016) 15:1702–12. doi: 10.1158/1535-7163.MCT-15-0892
9. Hwang JH, Arafeh R, Seo JH, Baca SC, Ludwig M, Arnoff TE, et al. CREB5 reprograms FOXA1 nuclear interactions to promote resistance to androgen receptor–targeting therapies. Elife (2022) 11:2. doi: 10.7554/eLife.73223.sa2
10. Bick G, Zhang J, Lower EE, Zhang X. Transcriptional coactivator MED1 in the interface of anti–estrogen and anti–HER2 therapeutic resistance. Cancer Drug Resist (2022) 5:498–510. doi: 10.20517/cdr.2022.33
11. Ge R, Wang Z, Montironi R, Jiang Z, Cheng M, Santoni M, et al. Epigenetic modulations and lineage plasticity in advanced prostate cancer. Ann Oncol (2020) 31:470–9. doi: 10.1016/j.annonc.2020.02.002
12. Luo M, Brooks M, Wicha MS. Epithelial–mesenchymal plasticity of breast cancer stem cells: implications for metastasis and therapeutic resistance. Curr Pharm Des (2015) 21:1301–10. doi: 10.2174/1381612821666141211120604
13. Bhanvadia RR, VanOpstall C, Brechka H, Barashi NS, Gillard M, McAuley EM, et al. MEIS1 and MEIS2 expression and prostate cancer progression: A role for HOXB13 binding partners in metastatic disease. Clin Cancer Res (2018) 24:3668–80. doi: 10.1158/1078-0432.CCR-17-3673
14. Pareja F, D’Alfonso TM. Neuroendocrine neoplasms of the breast: A review focused on the updated World Health Organization (WHO) 5th Edition morphologic classification. Breast J (2020) 26:1160–7. doi: 10.1111/tbj.13863
15. Conteduca V, Oromendia C, Eng KW, Bareja R, Sigouros M, Molina A, et al Clinical features of neuroendocrine prostate cancer. Eur J Cancer (2019) 121:7–18. doi: 10.1016/j.ejca.2019.08.011
16. Eyster KM. The estrogen receptors: an overview from different perspectives. Methods Mol Biol (2016) 1366:1–10. doi: 10.1007/978-1-4939-3127-9_1
17. Lonergan PE, Tindall DJ. Androgen receptor signaling in prostate cancer development and progression. J Carcinog (2011) 10:20. doi: 10.4103/1477-3163.83937
18. Kregel S, Chen JL, Tom W, Krishnan V, Kach J, Brechka H, et al. Acquired resistance to the second–generation androgen receptor antagonist enzalutamide in castration–resistant prostate cancer. Oncotarget (2016) 7:26259–74. doi: 10.18632/oncotarget.8456
19. He MX, Cuoco MS, Crowdis J, Bosma–Moody A, Zhang Z, Bi K, et al. Transcriptional mediators of treatment resistance in lethal prostate cancer. Nat Med (2021) 27:426–33. doi: 10.1038/s41591-021-01244-6
20. Toy W, Weir H, Razavi P, Lawson M, Goeppert AU, Mazzola AM, et al. Activating ESR1 mutations differentially affect the efficacy of ER antagonists. Cancer Discov (2017) 7:277–87. doi: 10.1158/2159-8290.CD-15-1523
21. Wilson AC, Meethal SV, Bowen RL, Atwood CS. Leuprolide acetate: a drug of diverse clinical applications. Expert Opin Investig Drugs (2007) 16:1851–63. doi: 10.1517/13543784.16.11.1851
22. Agarwal N, Hutson TE, Vogelzang NJ, Sonpavde G. Abiraterone acetate: a promising drug for the treatment of castration–resistant prostate cancer. Future Oncol (2010) 6:665–79. doi: 10.2217/fon.10.48
23. Ratre P, Mishra K, Dubey A, Vyas A, Jain A, Thareja S. Aromatase inhibitors for the treatment of breast cancer: A journey from the scratch. Anticancer Agents Med Chem (2020) 20:1994–2004. doi: 10.2174/1871520620666200627204105
24. Teo MY, Rathkopf DE, Kantoff P. Treatment of advanced prostate cancer. Annu Rev Med (2019) 70:479–99. doi: 10.1146/annurev-med-051517-011947
25. Hancock GR, Young KS, Hosfield DJ, Joiner C, Sullivan EA, Yildiz Y, et al. Unconventional isoquinoline–based SERMs elicit fulvestrant–like transcriptional programs in ER+ breast cancer cells. NPJ Breast Cancer (2022) 8:130. doi: 10.1038/s41523-022-00497-9
26. Patel HK, Bihani T. Selective estrogen receptor modulators (SERMs) and selective estrogen receptor degraders (SERDs) in cancer treatment. Pharmacol Ther (2018) 186:1–24. doi: 10.1016/j.pharmthera.2017.12.012
27. Strasser–Weippl K, Goss PE. Advances in adjuvant hormonal therapy for postmenopausal women. J Clin Oncol (2005) 23:1751–9. doi: 10.1200/JCO.2005.11.038
28. Wang L, Sharma A. SERDs: a case study in targeted protein degradation. Chem Soc Rev (2022) 51:8149–59. doi: 10.1039/D2CS00117A
29. Bubley GJ, Balk SP. Association between androgen receptor splice variants and prostate cancer resistance to abiraterone and enzalutamide. J Clin Oncol (2017) 35:2103–5. doi: 10.1200/JCO.2017.72.8808
30. Grinshpun A, Chen V, Sandusky ZM, Fanning SW, Jeselsohn R. ESR1 activating mutations: From structure to clinical application. Biochim Biophys Acta Rev Cancer (2023) 1878:188830. doi: 10.1016/j.bbcan.2022.188830
31. Lallous N, Volik SV, Awrey S, Leblanc E, Tse R, Murillo J, et al. Functional analysis of androgen receptor mutations that confer anti–androgen resistance identified in circulating cell–free DNA from prostate cancer patients. Genome Biol (2016) 17:10. doi: 10.1186/s13059-015-0864-1
32. Fuqua SA, Gu G, Rechoum Y. Estrogen receptor (ER) alpha mutations in breast cancer: hidden in plain sight. Breast Cancer Res Treat (2014) 144:11–9. doi: 10.1007/s10549-014-2847-4
33. Toy W, Shen Y, Won H, Green B, Sakr RA, Will M, et al. ESR1 ligand–binding domain mutations in hormone–resistant breast cancer. Nat Genet (2013) 45:1439–45. doi: 10.1038/ng.2822
34. Kregel S, Kiriluk KJ, Rosen AM, Cai Y, Reyes EE, Otto KB, et al. Sox2 is an androgen receptor–repressed gene that promotes castration–resistant prostate cancer. PloS One (2013) 8:e53701. doi: 10.1371/journal.pone.0053701
35. Hwang JH, Seo JH, Beshiri ML, Wankowicz S, Liu D, Cheung A, et al. CREB5 promotes resistance to androgen–receptor antagonists and androgen deprivation in prostate cancer. Cell Rep (2019) 29:2355–2370 e6. doi: 10.1016/j.celrep.2019.10.068
36. Kregel S, Wang C, Han X, Xiao L, Fernandez–Salas E, Bawa P, et al. Androgen receptor degraders overcome common resistance mechanisms developed during prostate cancer treatment. Neoplasia (2020) 22:111–9. doi: 10.1016/j.neo.2019.12.003
37. Kavarthapu R, Anbazhagan R, Dufau ML, Crosstalk between PRLR. and EGFR/HER2 signaling pathways in breast cancer. Cancers (Basel) (2021) 13:2, 6–9. doi: 10.3390/cancers13184685
38. Kregel S, Malik R, Asangani IA, Wilder–ROmans K, RajendIran T, Xiao L, et al. Functional and mechanistic interrogation of BET bromodomain degraders for the treatment of metastatic castration–resistant prostate cancer. Clin Cancer Res (2019) 25:4038–48. doi: 10.1158/1078-0432.CCR-18-3776
39. Isikbay M, Otto K, Kregel S, Kach J, Cai Y, Vander Griend DJ, et al. Glucocorticoid receptor activity contributes to resistance to androgen–targeted therapy in prostate cancer. Horm Cancer (2014) 5:72–89. doi: 10.1007/s12672-014-0173-2
40. Arora VK, Schenkein E, Murali R, Subudhi SK, Wongvipat J, Balbas MD, et al. Glucocorticoid receptor confers resistance to antiandrogens by bypassing androgen receptor blockade. Cell (2013) 155:1309–22. doi: 10.1016/j.cell.2013.11.012
41. Mohamed GA, Mahmood S, Ognjenovic NB, Lee MK, Wilkins OM, Christensen BC, et al. Lineage plasticity enables low–ER luminal tumors to evolve and gain basal–like traits. Breast Cancer Res (2023) 25:23. doi: 10.1186/s13058-023-01621-8
42. Balanis NG, Sheu KM, Esedebe FN, Patel SJ, Smith BA, Park JW, et al. Pan–cancer convergence to a small–cell neuroendocrine phenotype that shares susceptibilities with hematological malignancies. Cancer Cell (2019) 36:17–34 e7. doi: 10.1016/j.ccell.2019.06.005
43. de Wet L, Williams A, Gillard M, Kregel S, Lamperis S, Gutgesell LC, et al. SOX2 mediates metabolic reprogramming of prostate cancer cells. Oncogene (2022) 41:1190–2. doi: 10.1038/s41388-022-02228-7
44. Lee JK, Phillips JW, Smith BA, Park JW, Stoyanova T, McCaffrey EF, et al. N–myc drives neuroendocrine prostate cancer initiated from human prostate epithelial cells. Cancer Cell (2016) 29:536–47. doi: 10.1016/j.ccell.2016.03.001
45. Park JW, Lee JK, Sheu KM, Wang L, Balanis NG, Nguyen K, et al. Reprogramming normal human epithelial tissues to a common, lethal neuroendocrine cancer lineage. Science (2018) 362:91–5. doi: 10.1126/science.aat5749
46. Cejas P, Xie Y, Font–Tello A, Lim K, Syamala S, Qiu X, et al. Subtype heterogeneity and epigenetic convergence in neuroendocrine prostate cancer. Nat Commun (2021) 12:5775. doi: 10.1038/s41467-021-26042-z
47. Ku SY, Rosario S, Wang Y, Mu P, Seshadri M, Goodrich ZW, et al. Rb1 and Trp53 cooperate to suppress prostate cancer lineage plasticity, metastasis, and antiandrogen resistance. Science (2017) 355:78–83. doi: 10.1126/science.aah4199
48. Oser MG, Niederst MJ, Sequist LV, Engelman JA. Transformation from non–small–cell lung cancer to small–cell lung cancer: molecular drivers and cells of origin. Lancet Oncol (2015) 16:e165–72. doi: 10.1016/S1470-2045(14)71180-5
49. Aggarwal R, Huang J, Alumkal JJ, Zhang L, Feng FY, Thomas GV, et al. Clinical and genomic characterization of treatment–emergent small–cell neuroendocrine prostate cancer: A multi–institutional prospective study. J Clin Oncol (2018) 36:2492–503. doi: 10.1200/JCO.2017.77.6880
50. Modlin IM, Oberg K, Chung DC, Jensen RT, de Herder WW, Thakker RV, et al. Gastroenteropancreatic neuroendocrine tumours. Lancet Oncol (2008) 9:61–72. doi: 10.1016/S1470-2045(07)70410-2
51. Puca L, Gavyert K, Sailer V, Conteduca V, Dardenne E, Sigouros M, et al. Delta–like protein 3 expression and therapeutic targeting in neuroendocrine prostate cancer. Sci Transl Med (2019) 11:7–8. doi: 10.1126/scitranslmed.aav0891
52. Yao J, Bergsland E, Aggarwal R, Aparicio A, Beltran H, Crabtree JS, et al. DLL3 as an emerging target for the treatment of neuroendocrine neoplasms. Oncologist (2022) 27:940–51. doi: 10.1093/oncolo/oyac161
53. Cheng ZY, He TT, Gao XM, Zhao Y, Wang J. ZBTB transcription factors: key regulators of the development, differentiation and effector function of T cells. Front Immunol (2021) 12:713294. doi: 10.3389/fimmu.2021.713294
54. Maeda T. Regulation of hematopoietic development by ZBTB transcription factors. Int J Hematol (2016) 104:310–23. doi: 10.1007/s12185-016-2035-x
55. Singh AK, Verma S, Kushwaha PP, Prajapati KS, Shuaib M, Kumar S, et al. Role of ZBTB7A zinc finger in tumorigenesis and metastasis. Mol Biol Rep (2021) 48:4703–19. doi: 10.1007/s11033-021-06405-x
56. Gupta S, Singh AK, Prajapati KS, Kushwaha PP, Shuaib M, Kumar S. Emerging role of ZBTB7A as an oncogenic driver and transcriptional repressor. Cancer Lett (2020) 483:22–34. doi: 10.1016/j.canlet.2020.04.015
57. Han Q, Yan X, Ye Y, Han L, Ma X, Wang T, et al. ZBTB20 regulates prolactin expression and lactotrope function in adult mice. Endocrinology (2022) 163:5–8. doi: 10.1210/endocr/bqac181
58. To JC, Chiu AP, Tschida BR, Lo LH, Chiu CH, Li XX, et al. ZBTB20 regulates WNT/CTNNB1 signalling pathway by suppressing PPARG during hepatocellular carcinoma tumourigenesis. JHEP Rep (2021) 3:100223. doi: 10.1016/j.jhepr.2020.100223
59. Nishioka H. Aggressive pituitary tumors (PitNETs). Endocr J (2023) 70:241–8. doi: 10.1507/endocrj.EJ23-0007
Keywords: neuroendocrine, nuclear hormone receptors, hormone signaling, breast, prostate, lineage plasticity, pituitary
Citation: Phoenix JT, Budreika A, Kostlan RJ, Hwang JH, Fanning SW and Kregel S (2023) Editorial: Hormone resistance in cancer. Front. Endocrinol. 14:1272932. doi: 10.3389/fendo.2023.1272932
Received: 04 August 2023; Accepted: 17 August 2023;
Published: 24 August 2023.
Edited and Reviewed by:
Antonino Belfiore, University of Catania, ItalyCopyright © 2023 Phoenix, Budreika, Kostlan, Hwang, Fanning and Kregel. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Steven Kregel, c2tyZWdlbEBsdWMuZWR1