 Joana Duarte1,2
Joana Duarte1,2
- 1 Instituto de Medicina Molecular, University of Lisbon, Lisbon, Portugal
- 2 Instituto Gulbenkian de Ciência, Oeiras, Portugal
Monoclonal antibodies (mAb) have been shown effective in inducing immune tolerance in a range of animal models of autoimmunity, allergy, and transplantation. We investigated whether CD4-blockade, effective in inducing transplantation tolerance, could prevent systemic immune responses leading to anaphylaxis. We found that treatment with a non-depleting anti-CD4 mAb could prevent peanut-induced anaphylaxis following subsequent systemic exposure to crude peanut extract (CPE). Furthermore, the effect of CD4-blockade did not interfere with overall immune competence, as anti-CD4 treated mice remained fully competent to respond to unrelated antigens. Protection from anaphylaxis correlated with increased frequency of Foxp3+ regulatory T cells (Treg), and was abrogated following Treg depletion. Taken together our data suggest that activation of T cells by CPE in presence of CD4-blockade leads to Treg expansion that can prevent peanut-induced anaphylaxis.
Introduction
Anaphylaxis is an acute, life-threatening, allergic reaction where a physiologic process that normally acts in a local and limited manner to protect against infection occurs massively and systemically. Peanut allergy is a major cause of food-induced anaphylaxis, affecting around 1% of the population, with increasing prevalence worldwide (Kanny et al., 2001; Sampson, 2004). To date there is no cure for peanut allergy, and unlike many other food allergies, it persists through adulthood. Currently, avoidance is the only treatment advised. There is therefore, a clear need for safe and effective tolerance-inducing therapies for patients who might be exposed to anaphylactic reactions.
Monoclonal antibodies (mAb) that target T cell co-receptor and co-stimulatory molecules have been reported effective in inducing tolerance to non-self antigens. Waldmann and coworkers have shown non-lytic CD4 antibodies (with an isotype that does not directly deplete target cells) can induce long-term transplantation tolerance in mice (Graca et al., 2003; Kendal and Waldmann, 2010). The resulting tolerance state is mediated by Foxp3+ regulatory T cells (Treg), although other mechanisms may also operate (Graca et al., 2002, 2004; Lin et al., 2002). It was reported that a non-depleting anti-CD4 mAb was effective in preventing allergic airways disease in mice sensitized with ovalbumin (OVA; Li et al., 1999a). We have recently extended these data, showing that tolerance can be induced in mice to a clinically relevant aeroallergen – house dust mite (HDM). In this case, tolerant mice were protected from airways hyperreactivity (AHR), eosinophilia, goblet cell hyperplasia, and production of antigen-specific IgG1 and IgE (Agua-Doce and Graca, 2011). These data contrasts with the disappointing results from a clinical trial with a depleting anti-CD4 mAb (keliximab; Kon et al., 1998). In this trial the depleting nature of the mAb precluded the use of a dose sufficient to achieve effective CD4-blockade, as it led to immune suppression.
Surprisingly, the same non-depleting anti-CD4 mAb we successfully used to induce tolerance to HDM (or OVA) was reported to be less effective when tolerance was induced to systemically delivered human factor VIII in a mouse model of hemophilia (Salooja et al., 2002). Therefore, we decided to explore to which extent CD4-blockade is able to prevent a systemic allergic response: anaphylaxis.
We took advantage of a well established model of peanut-induced anaphylaxis, where the antigen crude peanut extract (CPE) is delivered through i.p. injection, allowing the precise control of the dose and time of exposure (Pons et al., 2004). C3H/HeJ mice have high susceptibility to peanut-induced anaphylaxis, being able to produce high peanut-specific antibody titers. Moreover, upon challenge through the i.p. route, these mice develop manifestations of anaphylactic shock, including a sharp drop of body temperature, which facilitates the quantification of clinical manifestations, and resemble anaphylactic reactions in human subjects (Li et al., 2000; Berin et al., 2006).
We confirmed C3H/HeJ mice can be sensitized with CPE, producing high titers of CPE-specific Th2-driven antibodies. We found that CD4-blockade, during the sensitization, prevented the generation of peanut-specific immunoglobulins, even following subsequent sensitization with CPE-alum, rendering the mice protected from anaphylaxis. The protective effect is abrogated following depletion of Treg cells. Importantly, CD4-blockade does not lead to immune deficiency, as mice remain competent to respond to different antigens.
Materials and Methods
Experimental Animals
C3H/HeJ mice were bred and maintained under specific pathogen-free facilities. Animals were sex-matched and used at 6–10 weeks of age. All experiments involving animals were approved by Direccao Geral Veterinaria (approval 018831). Sensitization was achieved by administration of 0.5 mg CPE in 2 mg aluminum hydroxide (alum, Alu-gel-S, Serva, Heidelberg, Germany) i.p. at days 1, 7, and 21. Mice were subsequently challenged with 10 mg CPE in PBS i.p.
Clinical Assessment of Anaphylaxis
Mice were assessed during 45 min following CPE challenge. Body temperature was measured at the indicated times with a rectally inserted thermal probe. The clinical score was evaluated as described elsewhere (Li et al., 2000): 0 – no manifestations; 1 – Scratching/rubbing around the nose and head; 2 – puffiness around eyes and mouth, reduced activity, diarrhea, pilar erecti; 3 – wheezing, labored respiration, cyanosis around mouth and tail; 4 – no activity after prodding, or tremor and convulsion; 5 – death. Scoring was performed blinded by two independent researchers.
CPE Preparation
Peanut flour was extensive defatted with diethyl ether, and the dried defatted peanut flour was extracted in ice-cold 10× PBS overnight at 4°C, centrifuged at 30,000 g for 60′, and filter-sterilized. Protein concentration was measured with the BCA method and aliquots were stored at −20ºC.
Monoclonal Antibodies
Non-depleting anti-CD4 (YTS177), the isotype control (YKIX302), and anti-CD25 (PC61) mAbs were produced in our laboratory using Integra CL1000 flasks (IBS, Chur, Switzerland), purified by 50% ammonium sulfate precipitation, dialyzed against PBS, and purity checked by native and SDS gel electrophoresis. The hybridomas were generously provided by Professor Herman Waldmann (Oxford, UK).
In vitro Cultures
Splenocytes (1 × 106) were cultured for 3 days in 96 well plates, with complete culture medium (RPMI-1640 with Glutamax, supplemented with 10% FBS, 1% hepes, 1% penicillin/streptomycin, 1% sodium pyruvate, 0.1% β-mercaptoethanol; Invitrogen), with addition of 20 μg OVA or CPE. At day 3, cells were centrifuged and supernatants recovered and kept at −80º C until cytokine quantification.
ELISA
The serum IgE and CPE- or OVA-specific IgG1 was measured in microtiter plates coated with 50 μg/ml CPE or OVA. IgE was quantified with an Opteia kit (BD Pharmingen) and IgG1 with a kit from Southern Biotech. Quantification of cytokines in cell-culture supernatants was performed using IL-10 and IL-13 kits (Peprotech, London, UK), and IL-5 Opteia kits (BD Pharmingen). All assays were performed according to the manufacturer’s instructions.
Flow Cytometry
Single cell suspensions were analyzed with the following fluorochrome-labeled mAb: CD3 Percp–Cy5.5 (145–2C11), CD4 PE (GK1.5), CD8 APC–Cy7 (53–6.7), CD25 Pe–Cy7 (PC61.5), and Foxp3 (FJK165; eBiosciences). Samples were run in a FACS Canto and analyzed with FlowJo.
Statistical Analysis
Statistical significance was determined using the two-tailed non-parametric Mann–Whitney U test and P values < 0.05 were deemed significant (*P < 0.05; ** P < 0.01; *** P < 0.001).
Results
CPE-Induced an Anaphylactic Response in Sensitized C3H/HeJ Mice
C3H/HeJ mice have been widely used as an experimental murine model of allergic diseases, being prone for the development of Th2-like immune responses, namely peanut-induced anaphylaxis (Li et al., 2000; Morafo et al., 2003). We found that upon i.p. sensitization with CPE-alum, an anaphylactic response was observed following challenge with CPE administered i.p. (Figure 1A). Female mice appeared to display greater disease severity, however, both males and females developed clinical manifestations of anaphylaxis, reaching a state where they did not respond to external stimuli (score 4). In addition, these mice displayed a sharp drop in body temperature, during the initial 45′ following challenge with CPE (Figures 1B,C). Serum concentration of Th2-driven immunoglobulins, such as IgE and CPE-specific IgG1, were markedly increased in CPE-sensitized mice, compared to non-exposed controls (Figures 1D,E). These data are similar to prior reports (Li et al., 2000; Pons et al., 2004), supporting the use of C3H/HeJ mice as an adequate model for the study of anaphylaxis.
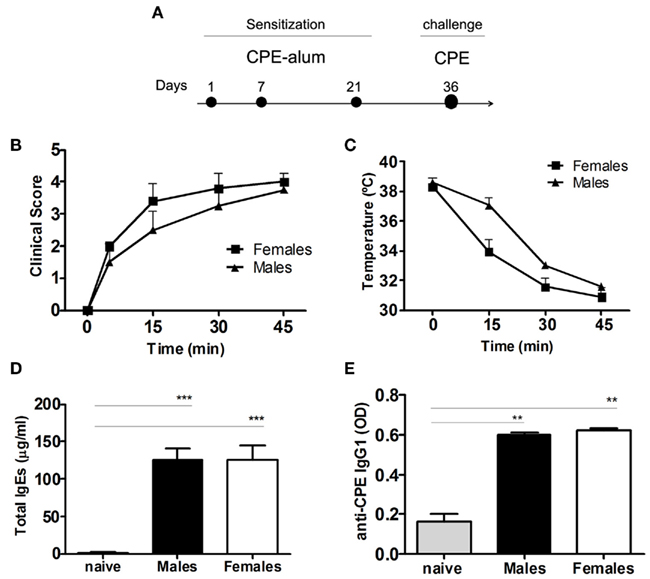
Figure 1. Crude peanut extract induces anaphylaxis in C3H/HeJ mice. (A) C3H/HeJ mice were sensitized i.p. with 0.5 mg CPE in 2 mg alum on days 1, 7, and 21. The animals were assessed at day 36 following challenge with 10 mg CPE i.p., with clinical score and temperature monitored during 45 min after challenge. (B) Clinical score was evaluated as described in the Section “Materials and Methods,” and (C) body temperature was measured. Females show more severe clinical manifestations of anaphylaxis, compared to male mice, but without reaching statistical significance. (D) Total IgE and (E) CPE-specific IgG1 were measured in the serum of individual mice. Both males and females showed an increase in the immunoglobulin levels compared to naïve mice (n = 5, ** P < 0.01). Data (B–E) are representative of two independent experiments.
Non-Depleting Anti-CD4 MAb Prevented CPE-Induced Anaphylaxis
We have recently shown non-depleting anti-CD4 mAb can prevent allergic airways disease in mice in an antigen-specific way (Agua-Doce and Graca, 2011). To assess if this monoclonal antibody was equally efficient in the prevention of a systemic Th2-driven response, we investigated whether anti-CD4 treatment could prevent anaphylaxis in C3H/HeJ mice.
We found anti-CD4 treatment at the time of sensitization was effective in preventing the development of anaphylactic manifestations observed during the 45 min following challenge with CPE (Figures 2A,B). Anti-CD4 treated mice also maintained their body temperature, unlike control animals where the temperature dropped rapidly (Figure 2C). The production of IgE and CPE-specific IgG1 was also impaired in mice treated with anti-CD4 (Figures 2D,E).
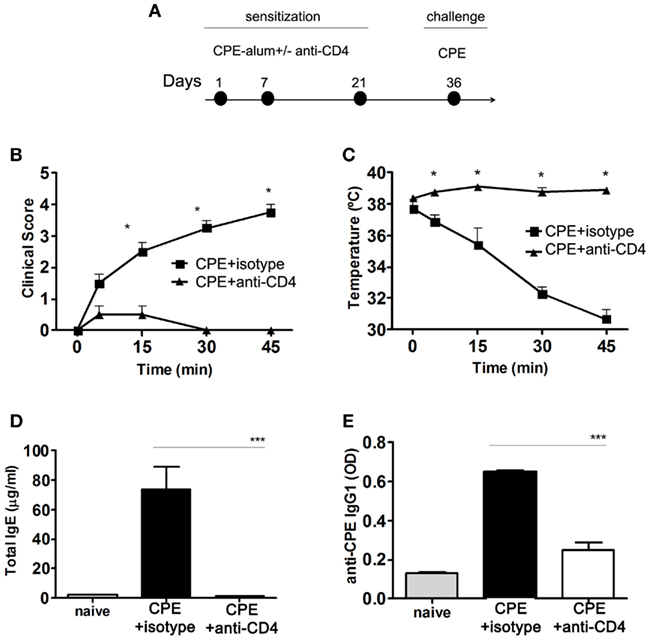
Figure 2. Anti-CD4 prevents peanut-induced anaphylaxis in C3H/HeJ mice. (A) Female C3H/HeJ mice were sensitized to CPE at the indicated days, with a group of animals being treated with 1 mg anti-CD4 or an isotype control i.p. at the time of each sensitization. All mice were assessed at day 36 following challenge with 10 mg CPE i.p. (B) Mice were assessed for their clinical score and (C) body temperature during 45 min following CPE challenge. Anti-CD4 treated mice did not develop any significant manifestations of disease (n = 4, * P < 0.05). In addition, CPE-sensitized mice showed a sharp drop of body temperature (n = 4, * P < 0.05). (D) Serum concentration of total IgE and (E) CPE-specific IgG1. Anti-CD4 treated mice maintained IgE levels at a concentration similar to naïve non-immunized mice, while mice treated with an isotype control significantly increased serum IgE and IgG1 (n = 8, *** P < 0.001). Data (B–E) are representative of three independent experiments.
Anti-CD4 Did not Prevent Anaphylaxis in Pre-Sensitized Mice with High IgE
We next assessed the impact of anti-CD4 treatment in pre-sensitized mice. Note that the exposure to CPE at days 36–58 (Figure 3A), probably due to the use of alum, only induced mild clinical manifestations of disease even in the absence of anti-CD4 (not shown). Furthermore, in spite of such mild clinical manifestations of anaphylaxis, all mice strongly developed anaphylaxis when exposed to CPE without alum at day 73 (Figures 3B,C).
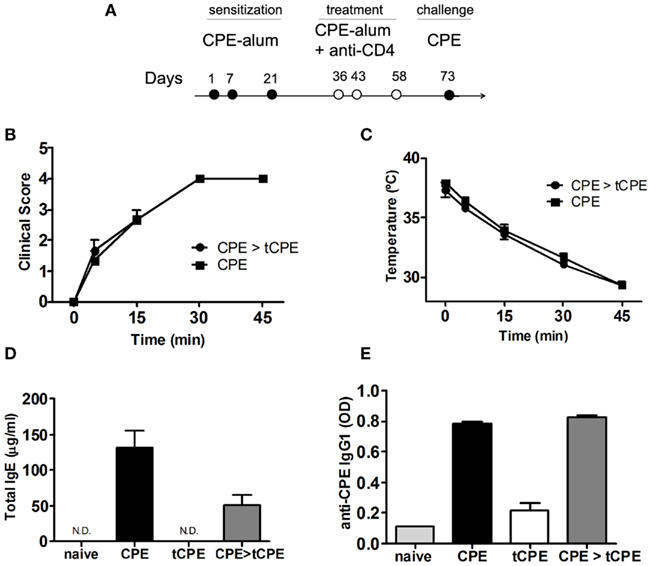
Figure 3. Anti-CD4 does not prevent anaphylaxis in pre-sensitized mice with high titers of IgE. (A) C3H/HeJ mice were sensitized with CPE-alum on days 1, 7, and 21, and treated with anti-CD4 administered together with CPE at the indicated days. All mice were assessed at day 73 after challenge with 10 mg CPE i.p. (B) Sensitized mice subsequently treated with anti-CD4 (CPE > tCPE) displayed anaphylactic manifestations identical to the control group, as well as (C) sharp reduction of body temperature. (D) Total IgE and (E) CPE-specific IgG1 levels were similar in sensitized mice even when treated with anti-CD4 after sensitization (CPE and CPE > tCPE). These high levels of immunoglobulins contrast with mice treated with anti-CD4 at the time of initial sensitization (tCPE) that maintain low titers of IgE and IgG1 (n = 4, *** P < 0.001). Data (B–E) are representative of two independent experiments.
We found that mice sensitized with CPE in advance of anti-CD4 treatment were not protected from the development of clinical manifestations of anaphylaxis, namely reduction of body temperature (Figures 3B,C). Mice treated with anti-CD4 had increased levels of total IgE and CPE-specific IgG1, similar to CPE-sensitized controls (Figures 3D,E). These results suggest that in pre-sensitized individuals pre-existing IgE, produced at the time of initial sensitization, will drive clinical manifestations of anaphylaxis in a T-cell independent manner.
Non-Depleting Anti-CD4 Monoclonal Antibody-Induced Protection from Subsequent Exposure to CPE
Non-depleting anti-CD4 mAb was shown to induce long-term tolerance in transplantation (Waldmann and Cobbold, 2001). We also found long-term protection to HDM could be induced with this mAb in mice (Agua-Doce and Graca, 2011). Therefore, we evaluated whether treatment with non-depleting anti-CD4 at the time of CPE sensitization could protect mice from peanut-induced anaphylaxis following a subsequent sensitization. As a consequence, 2 weeks after treatment, mice were re-sensitized with CPE-alum as shown in Figure 4A. The 2 weeks is sufficient to allow for clearance of the mAb below therapeutic levels, as we found by treating mice with anti-CD4 2 weeks in advance of providing the antigen (in previous experiments using OVA as the antigen, data not shown).
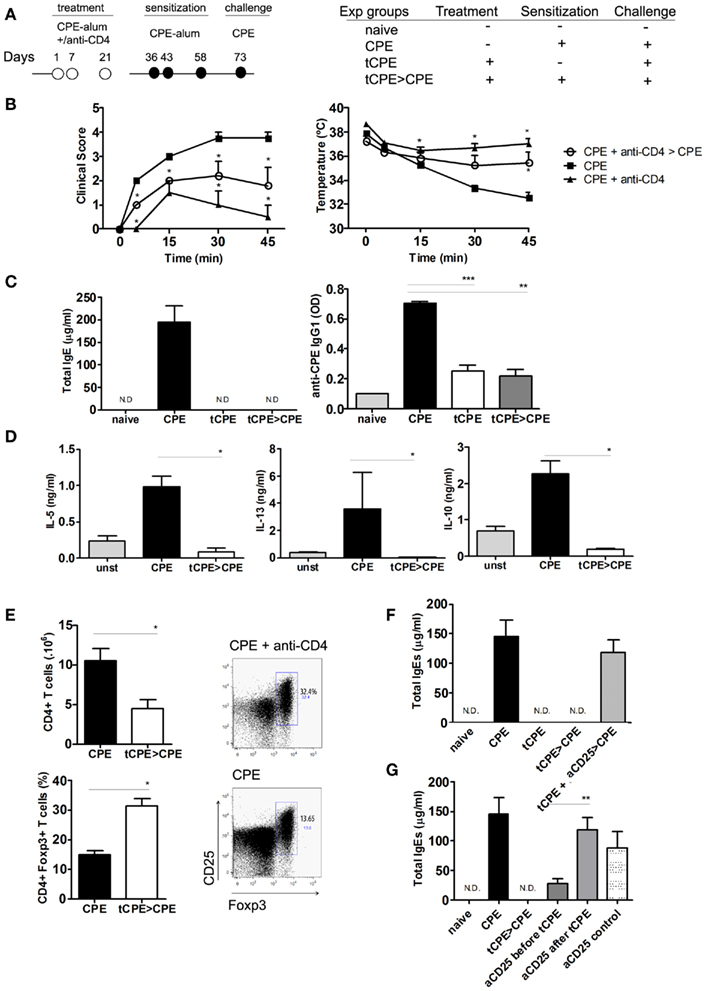
Figure 4. Anti-CD4 treated mice are protected from subsequent sensitization with CPE. (A) C3H/HeJ mice were treated with anti-CD4 (tCPE) at the time of initial sensitization. Following 2 weeks, the mice were sensitized with CPE-alum (tCPE > CPE) and finally assessed at day 73 following challenge with 10 mg CPE i.p. (B) Mice treated with anti-CD4 (tCPE), even when re-sensitized with CPE-alum (tCPE > CPE), did not develop anaphylactic manifestations as severe as CPE-sensitized group (CPE), showing clinical scores and body temperature significantly different (n = 5, * P < 0.05). (C) All groups of mice treated with anti-CD4, showed no significant increase in IgE and CPE-specific IgG1 levels when compared to naïve mice. The Th2-driven immunoglobulins were significantly increased in CPE-sensitized mice (n = 5, * P < 0.05, ** P < 0.001, *** P < 0.001). (D) Cytokines were measured in the supernatants of CPE-stimulated spleen cells. IL-5, IL-13, and IL-10 were almost undetectable in anti-CD4 treated mice following re-sensitization (tCPE > CPE) and significantly reduced compared with untreated mice (n = 5, * P < 0.05). (E) The number of splenic CD4+ T cells was reduced in anti-CD4 treated mice, while the frequency of CD4+Foxp3+ T cells was significantly higher (n = 5, * P < 0.05). (F) C3H/HeJ mice were treated as described above, while some animals were depleted of CD25 cells at the time of anti-CD4 treatment (tCPE + aCD25 > CPE). CD25 depletion abrogated protection, leading to production of IgE to levels similar to mice not treated with anti-CD4. (G) We compared the effect of CD25 depletion before and after treatment with anti-CD4 (2 × 1 mg, 10 and 8 days before, or after tolerance induction with CPE+ anti-CD4). Control mice were CD25-depleted before challenge with CPE, in the absence of treatment with anti-CD4 (n = 5, * P < 0.05, ** P < 0.001, *** P < 0.001). Data (B–F) are representative of two independent experiments.
Manifestations of anaphylaxis were evident after challenge in CPE-sensitized mice, whereas mice treated with anti-CD4 were protected from severe clinical manifestations (Figure 4B). In addition, serum titers of IgE and CPE-specific IgG1 remained low in mice treated with anti-CD4 (Figure 4C). Injection of alum alone (in the absence of antigen and anti-CD4) did not prevent subsequent response elicited following sensitization (not shown).
We next compared the production of Th2-type cytokines by T cells from anti-CD4 treated mice. We cultured spleen T cells for 3 days in presence of CPE stimulation, and quantified the cytokines in cultures supernatants. T cells from mice treated with anti-CD4 mAb, unlike splenocytes from control CPE-sensitized mice, did not produce detectable levels of IL-5 and IL-13 (Figure 4D). Several reports have implicated IL-10 in protection from allergy (Akdis et al., 1998, 2004). However, production of IL-10 following in vitro stimulation with CPE was also reduced in cells from anti-CD4 treated mice (Figure 4D). In fact, IL-10 production was higher in cells from animals sensitized with CPE in the absence of anti-CD4 treatment.
Several studies in transplantation have shown that long-term tolerance induced with CD4-blockade is associated with Foxp3+ Treg expansion (Cobbold et al., 2004; Graca et al., 2005; Oliveira et al., 2011). We found that although the anti-CD4 mAb has a non-depleting isotype, and does not directly lyse CD4+ T cells (Figure A1 in Appendix), the absolute number of CD4+ T cells in the spleen of anti-CD4 treated mice were lower than in controls (Figure 4E). However, the frequency of Foxp3+ Treg cells within the T cell population was significantly increased in anti-CD4 treated mice (Figure 4E).
To further confirm the participation of Treg cells in the protection induced following anti-CD4 treatment, we evaluated the efficacy of CD4-blockade in CD25-depleted mice. We found that mice depleted of CD25 T cells at the time of CD4-blockade were not protected from peanut-induced anaphylaxis, induced following subsequent immunization with CPE-alum as described in Figure 4A. In fact, CD25-depleted mice exhibited high levels of total IgE, similar to what was observed in mice not treated with anti-CD4 (Figure 4F). These data suggest Foxp3+ Treg cells participate in protection from peanut-induced anaphylaxis induced following CD4-blockade. In addition, we compared the effect of CD25 depletion when applied before or after tolerance induction with anti-CD4. We found that treatment with anti-CD25 in advance of tolerance induction was not as effective in abrogating tolerance induction as when CD25 depletion was performed after anti-CD4 treatment (Figure 4G). These data suggest the participation of adaptive Treg cells, induced at the time of anti-CD4 treatment, in tolerance induction.
Anti-CD4 Treatment Induced Antigen-Specific Protection
We finally assessed whether anti-CD4 treatment was affecting the global immunocompetence, by studying the ability of mAb-treated mice to respond to different antigens. Therefore, following treatment of C3H/HeJ mice with CPE in presence of anti-CD4, some mice were re-sensitized with the same (CPE) or a different (OVA) antigen (Figure 5A). Mice treated with anti-CD4 remained fully competent to respond to sensitization with OVA-alum, developing a Th2-immune response leading to production of high IgE titers (Figure 5B). In fact, the levels of IgE were comparable to what was observed in CPE-sensitized control mice.
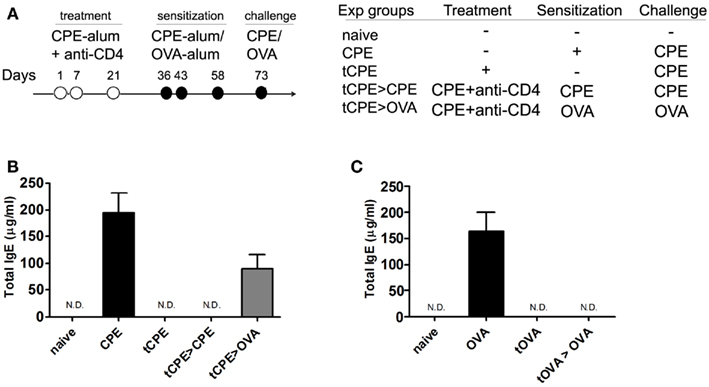
Figure 5. Anti-CD4 treatment induces antigen-specific tolerance. (A) C3H/HeJ mice were sensitized with CPE or OVA, and treated with anti-CD4 as described. The animals were assessed at day 73. (B) Mice treated with anti-CD4 mAb (tCPE) even following subsequent sensitization with the same antigens (tCPE > CPE) maintained IgE titers significantly lower than control animals (CPE; *P < 0.05). However, anti-CD4 treatment did not prevent the production of IgE following subsequent sensitization with OVA, a different antigen (tCPE > OVA), that led to IgE titers similar to the levels of control mice (CPE). (C) Equivalent experiment to (B) where OVA was used as the antigen to sensitize C3H/HeJ mice. OVA sensitization led to high titers of IgE (OVA). Exposure to OVA in presence of CD4-blockade (tOVA) prevented IgE production even following subsequent sensitization with OVA (tOVA > OVA) (n = 5, * P < 0.05). Data (B,C) are representative of two independent experiments.
Conversely, when we treated mice with anti-CD4 together with OVA, we prevented production of IgE following subsequent sensitization with OVA (Figure 5C).
Taken together these data show anti-CD4 treatment induces unresponsiveness to the antigens administered at the time of CD4-blockade, without preventing subsequent immune responses to different antigens. Therefore, anti-CD4 treated mice remain immune competent.
Discussion
Our data show CD4-blockade is effective in inducing antigen-specific tolerance to CPE, thus preventing subsequent sensitization with the same antigens. This conclusion contrasts with the apparent difficulty to induce tolerance in mice to systemically delivered human factor VIII or factor IX using the same mAb (Salooja et al., 2002). We are currently investigating the mechanisms that may explain the reason distinct proteins are tolerated with different ability.
Anti-CD4 is a non-depleting monoclonal antibody which has been extensively studied in several animal models of immune pathology. In transplantation and autoimmune arthritis, non-depleting anti-CD4 is effective in preventing transplant rejection and inducing long-term tolerance in mice (Waldmann and Cobbold, 2001; Duarte et al., 2010). Experiments with TCR-transgenic mice have shown the tolerant state is maintained by Foxp3+ Treg cells that can be induced from Foxp3− precursors (Cobbold et al., 2004; Oliveira et al., 2011). Such T cells can actively maintain tolerance by recruiting non-tolerant cells into the regulatory pool – a phenomenon termed infectious tolerance (Qin et al., 1993; Graca et al., 2000; Waldmann et al., 2006). In fact, mice transplanted under the cover of non-depleting anti-CD4 mAb do not become tolerant (i.e., they reject the transplant) when treated with anti-CD25 (Cobbold et al., 2006). We followed the same approach to show mice treated with anti-CD25 remained competent to produce CPE-specific antibodies, suggesting a similar mechanism of action. We cannot exclude that part of the effect of CD4-blockade is due to activation-induced cell death of T cells specific for the antigen present at the time of tolerance induction – a mechanism well described in transplantation tolerance induced with co-stimulation blockade (Li et al., 1999b; Wells et al., 1999). In any case, Treg cells appear to play the key role in our experiments, something suggested not only by the Treg-depletion experiment, but also by the marked increase in the frequency of Foxp3+ T cells in anti-CD4 treated mice. In addition we found that Treg depletion in advance of anti-CD4 treatment did not have a major impact on tolerance induction. On the contrary, CD25 depletion after anti-CD4 treatment abrogated tolerance induction. Although these data are not conclusive, as we cannot follow the fate of antigen-specific cells, they suggest that induced Treg cells are probably key participants in tolerance induction. Such tolerance mechanism is also distinct from other immune tolerance induction protocols that rely on the production of IL-10 or shifting the response toward Th1 (Ebner et al., 1997; Francis et al., 2003; Akdis et al., 2004; Valenta et al., 2004).
When we treated with anti-CD4 mAb mice previously sensitized with CPE, we could not prevent manifestations of anaphylaxis given the presence of high IgE titers. It remains, however, to be shown whether therapeutic neutralization of IgE, could allow tolerance to be imposed in a way preventing further production of IgE. In fact, our results with a mouse model of allergic airways disease show that tolerance can be achieved in pre-sensitized mice, even following one episode of airways inflammation (Agua-Doce and Graca, 2011). Taken together, these findings suggest IgE is not preventing tolerance from being achieved within the T cell compartment, but high levels of IgE are sufficient to lead to acute manifestations of disease. In addition, memory cells have been recognized as a major barrier for therapeutic induction of tolerance (Adams et al., 2003). In any case, immune tolerance can be induced in some experimental models with anti-CD4, in presence of memory T cells (Agua-Doce and Graca, 2011). Concerning tolerance induction to CPE, we cannot conclude whether, besides high IgE levels, the presence of memory T cells in pre-sensitized mice constitute a barrier to tolerance induction.
Interestingly, we found sensitized mice only developed mild manifestations of anaphylaxis when injected with CPE-alum, while CPE alone triggered a severe anaphylactic response with a sharp drop in core body temperature. However, when we injected alum alone, or alum together with a different antigen (OVA), we could not observe a protective effect toward subsequent sensitization with CPE.
To find an efficient and specific therapy for peanut-induced anaphylaxis has been a major challenge (Burks et al., 2008). Once the diagnosis is established, the course of action is restricted to strict elimination of the causative allergen from the patient’s diet, and it was estimated that about 50% of the affected individuals develop an accidental reaction over a 2-year period (Sicherer et al., 2003). Distinct approaches to induce tolerance to food allergens have been or are being pursued (Canonica and Passalacqua, 2003; Clark et al., 2009; Jones et al., 2009; Blumchen et al., 2010; Vickery et al., 2011). It is however yet unclear whether the evidence will support clinical adoption of any of those tolerogenic strategies.
We believe our data provide evidence that targeting CD4 T cells may provide a way to achieve or facilitate tolerance induction to systemic antigens, such as the ones that may trigger anaphylaxis.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
We are grateful to A. Wesley Burks (Duke University) for helpful advice and for providing samples of CPE to validate the CPE we have produced in-house. We thank Herman Waldmann (University of Oxford) for hybridomas of anti-CD4 and anti-CD25. Funded by Fundacao para a Ciencia e Tecnologia Portugal – PTDC/SAU-MII/64279/2006. The authors have no competing financial interests.
References
Adams, A. B., Pearson, T. C., and Larsen, C. P. (2003). Heterologous immunity: an overlooked barrier to tolerance. Immunol. Rev. 196, 147–160.
Agua-Doce, A., and Graca, L. (2011). Prevention of house dust mite induced allergic airways disease in mice through immune tolerance. PLoS ONE 6, e22320. doi: 10.1371/journal.pone.0022320
Akdis, C. A., Blesken, T., Akdis, M., Wuthrich, B., and Blaser, K. (1998). Role of interleukin 10 in specific immunotherapy. J. Clin. Invest. 102, 98–106.
Akdis, M., Verhagen, J., Taylor, A., Karamloo, F., Karagiannidis, C., Crameri, R., Thunberg, S., Deniz, G., Valenta, R., Fiebig, H., Kegel, C., Disch, R., Schmidt-Weber, C. B., Blaser, K., and Akdis, C. A. (2004). Immune responses in healthy and allergic individuals are characterized by a fine balance between allergen-specific T regulatory 1 and T helper 2 cells. J. Exp. Med. 199, 1567–1575.
Berin, M. C., Zheng, Y., Domaradzki, M., Li, X. M., and Sampson, H. A. (2006). Role of TLR4 in allergic sensitization to food proteins in mice. Allergy 61, 64–71.
Blumchen, K., Ulbricht, H., Staden, U., Dobberstein, K., Beschorner, J., De Oliveira, L. C., Shreffler, W. G., Sampson, H. A., Niggemann, B., Wahn, U., and Beyer, K. (2010). Oral peanut immunotherapy in children with peanut anaphylaxis. J. Allergy Clin. Immunol. 126, 83–91 e81.
Burks, A. W., Laubach, S., and Jones, S. M. (2008). Oral tolerance, food allergy, and immunotherapy: implications for future treatment. J. Allergy Clin. Immunol. 121, 1344–1350.
Canonica, G. W., and Passalacqua, G. (2003). Noninjection routes for immunotherapy. J. Allergy Clin. Immunol. 111, 437–448; quiz 449.
Clark, A. T., Islam, S., King, Y., Deighton, J., Anagnostou, K., and Ewan, P. W. (2009). Successful oral tolerance induction in severe peanut allergy. Allergy 64, 1218–1220.
Cobbold, S. P., Adams, E., Graca, L., Daley, S., Yates, S., Paterson, A., Robertson, N. J., Nolan, K. F., Fairchild, P. J., and Waldmann, H. (2006). Immune privilege induced by regulatory T cells in transplantation tolerance. Immunol. Rev. 213, 239–255.
Cobbold, S. P., Castejon, R., Adams, E., Zelenika, D., Graca, L., Humm, S., and Waldmann, H. (2004). Induction of foxP3+ regulatory T cells in the periphery of T cell receptor transgenic mice tolerized to transplants. J. Immunol. 172, 6003–6010.
Duarte, J., Agua-Doce, A., Oliveira, V. G., Fonseca, J. E., and Graca, L. (2010). Modulation of IL-17 and Foxp3 expression in the prevention of autoimmune arthritis in mice. PLoS ONE 5, e10558. doi: 10.1371/journal.pone.0010558
Ebner, C., Siemann, U., Bohle, B., Willheim, M., Wiedermann, U., Schenk, S., Klotz, F., Ebner, H., Kraft, D., and Scheiner, O. (1997). Immunological changes during specific immunotherapy of grass pollen allergy: reduced lymphoproliferative responses to allergen and shift from TH2 to TH1 in T-cell clones specific for Phl p 1, a major grass pollen allergen. Clin. Exp. Allergy 27, 1007–1015.
Francis, J. N., Till, S. J., and Durham, S. R. (2003). Induction of IL-10+CD4+CD25+ T cells by grass pollen immunotherapy. J. Allergy Clin. Immunol. 111, 1255–1261.
Graca, L., Chen, T. C., Le Moine, A., Cobbold, S. P., Howie, D., and Waldmann, H. (2005). Dominant tolerance: activation thresholds for peripheral generation of regulatory T cells. Trends Immunol. 26, 130–135.
Graca, L., Honey, K., Adams, E., Cobbold, S. P., and Waldmann, H. (2000). Cutting edge: anti-CD154 therapeutic antibodies induce infectious transplantation tolerance. J. Immunol. 165, 4783–4786.
Graca, L., Le Moine, A., Cobbold, S. P., and Waldmann, H. (2003). Antibody-induced transplantation tolerance: the role of dominant regulation. Immunol. Res. 28, 181–191.
Graca, L., Le Moine, A., Lin, C. Y., Fairchild, P. J., Cobbold, S. P., and Waldmann, H. (2004). Donor-specific transplantation tolerance: the paradoxical behavior of CD4+CD25+ T cells. Proc. Natl. Acad. Sci. U.S.A. 101, 10122–10126.
Graca, L., Thompson, S., Lin, C. Y., Adams, E., Cobbold, S. P., and Waldmann, H. (2002). Both CD4(+)CD25(+) and CD4(+)CD25(-) regulatory cells mediate dominant transplantation tolerance. J. Immunol. 168, 5558–5565.
Jones, S. M., Pons, L., Roberts, J. L., Scurlock, A. M., Perry, T. T., Kulis, M., Shreffler, W. G., Steele, P., Henry, K. A., Adair, M., Francis, J. M., Durham, S., Vickery, B. P., Zhong, X., and Burks, A. W. (2009). Clinical efficacy and immune regulation with peanut oral immunotherapy. J. Allergy Clin. Immunol. 124, 292–300, 300 e291–297.
Kanny, G., Moneret-Vautrin, D. A., Flabbee, J., Beaudouin, E., Morisset, M., and Thevenin, F. (2001). Population study of food allergy in France. J. Allergy Clin. Immunol. 108, 133–140.
Kendal, A. R., and Waldmann, H. (2010). Infectious tolerance: therapeutic potential. Curr. Opin. Immunol. 22, 560–565.
Kon, O. M., Sihra, B. S., Compton, C. H., Leonard, T. B., Kay, A. B., and Barnes, N. C. (1998). Randomised, dose-ranging, placebo-controlled study of chimeric antibody to CD4 (keliximab) in chronic severe asthma. Lancet 352, 1109–1113.
Li, L., Crowley, M., Nguyen, A., and Lo, D. (1999a). Ability of a nondepleting anti-CD4 antibody to inhibit Th2 responses and allergic lung inflammation is independent of coreceptor function. J. Immunol. 163, 6557–6566.
Li, Y., Li, X. C., Zheng, X. X., Wells, A. D., Turka, L. A., and Strom, T. B. (1999b). Blocking both signal 1 and signal 2 of T-cell activation prevents apoptosis of alloreactive T cells and induction of peripheral allograft tolerance. Nat. Med. 5, 1298–1302.
Li, X. M., Serebrisky, D., Lee, S. Y., Huang, C. K., Bardina, L., Schofield, B. H., Stanley, J. S., Burks, A. W., Bannon, G. A., and Sampson, H. A. (2000). A murine model of peanut anaphylaxis: T- and B-cell responses to a major peanut allergen mimic human responses. J. Allergy Clin. Immunol. 106, 150–158.
Lin, C. Y., Graca, L., Cobbold, S. P., and Waldmann, H. (2002). Dominant transplantation tolerance impairs CD8+ T cell function but not expansion. Nat. Immunol. 3, 1208–1213.
Morafo, V., Srivastava, K., Huang, C. K., Kleiner, G., Lee, S. Y., Sampson, H. A., and Li, A. M. (2003). Genetic susceptibility to food allergy is linked to differential TH2-TH1 responses in C3H/HeJ and BALB/c mice. J. Allergy Clin. Immunol. 111, 1122–1128.
Oliveira, V. G., Caridade, M., Paiva, R. S., Demengeot, J., and Graca, L. (2011). Sub-optimal CD4(+) T-cell activation triggers autonomous TGF-beta-dependent conversion to Foxp3(+) regulatory T cells. Eur. J. Immunol. 41, 1249–1255.
Pons, L., Ponnappan, U., Hall, R. A., Simpson, P., Cockrell, G., West, C. M., Sampson, H. A., Helm, R. M., and Burks, A. W. (2004). Soy immunotherapy for peanut-allergic mice: modulation of the peanut-allergic response. J. Allergy Clin. Immunol. 114, 915–921.
Qin, S., Cobbold, S. P., Pope, H., Elliott, J., Kioussis, D., Davies, J., and Waldmann, H. (1993). “Infectious” transplantation tolerance. Science 259, 974–977.
Salooja, N., Kemball-Cook, G., Tuddenham, E. G., and Dyson, J. (2002). Use of a non-depleting anti-CD4 antibody to modulate the immune response to coagulation factors VIII and IX. Br. J. Haematol. 118, 839–842.
Sicherer, S. H., Munoz-Furlong, A., and Sampson, H. A. (2003). Prevalence of peanut and tree nut allergy in the United States determined by means of a random digit dial telephone survey: a 5-year follow-up study. J. Allergy Clin. Immunol. 112, 1203–1207.
Valenta, R., Ball, T., Focke, M., Linhart, B., Mothes, N., Niederberger, V., Spitzauer, S., Swoboda, I., Vrtala, S., Westritschnig, K., and Kraft, D. (2004). Immunotherapy of allergic disease. Adv. Immunol. 82, 105–153.
Vickery, B. P., Scurlock, A. M., Jones, S. M., and Burks, A. W. (2011). Mechanisms of immune tolerance relevant to food allergy. J. Allergy Clin. Immunol. 127, 576–584; quiz 585–576.
Waldmann, H., Adams, E., Fairchild, P., and Cobbold, S. (2006). Infectious tolerance and the long-term acceptance of transplanted tissue. Immunol. Rev. 212, 301–313.
Waldmann, H., and Cobbold, S. (2001). Regulating the immune response to transplants. a role for CD4+ regulatory cells? Immunity 14, 399–406.
Wells, A. D., Li, X. C., Li, Y., Walsh, M. C., Zheng, X. X., Wu, Z., Nunez, G., Tang, A., Sayegh, M., Hancock, W. W., Strom, T. B., and Turka, L. A. (1999). Requirement for T-cell apoptosis in the induction of peripheral transplantation tolerance. Nat. Med. 5, 1303–1307.
Appendix
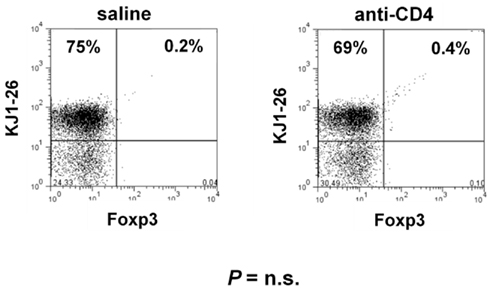
Figure A1. Anti-CD4 treatment does not directly deplete T cells in the absence of antigen. DO11.10.Rag−/− mice were treated with two shots of 1 mg anti-CD4 in alternate days. Spleen and LN were collected at day 7 and stained with the TCR-specific mAb (KJ1-26) and Foxp3. No significant difference was observed in the frequency or number of T cells from mAb-treated animals.
Keywords: immune tolerance, anaphylaxis, peanut, regulatory T cells, Foxp3, anti-CD4 monoclonal antibody
Citation: Duarte J, Caridade M and Graca L (2011) CD4-blockade can induce protection from peanut-induced anaphylaxis. Front. Immun. 2:56. doi: 10.3389/fimmu.2011.00056
Received: 24 June 2011; Paper pending published: 12 July 2011;
Accepted: 04 October 2011; Published online: 19 October 2011.
Edited by:
Stephen Paul Cobbold, University of Oxford, UKReviewed by:
Alain Le Moine, Université Libre de Bruxelles, BelgiumHans-dieter Volk, Charité – University Medicine Berlin, Germany
Valerie Julia, INSERM, France
Copyright: © 2011 Duarte, Caridade and Graca. This is an open-access article subject to a non-exclusive license between the authors and Frontiers Media SA, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and other Frontiers conditions are complied with.
*Correspondence: Luis Graca, Instituto de Medicina Molecular, University of Lisbon, Avenida Professor Egas Moniz, 1649-025 Lisbon, Portugal. e-mail:bGdyYWNhQGZtLnVsLnB0