 Georgina I. López-Cortés
Georgina I. López-Cortés Laura Díaz-Alvarez
Laura Díaz-Alvarez Enrique Ortega
Enrique Ortega- Department of Immunology, Instituto de Investigaciones Biomédicas, Universidad Nacional Autónoma de México, Mexico City, Mexico
For a long time, proteins with enzymatic activity have not been usually considered to carry out other functions different from catalyzing chemical reactions within or outside the cell. Nevertheless, in the last few years several reports have uncovered the participation of numerous enzymes in other processes, placing them in the category of moonlighting proteins. Some moonlighting enzymes have been shown to participate in complex processes such as cell adhesion. Cell adhesion plays a physiological role in multiple processes: it enables cells to establish close contact with one another, allowing communication; it is a key step during cell migration; it is also involved in tightly binding neighboring cells in tissues, etc. Importantly, cell adhesion is also of great importance in pathophysiological scenarios like migration and metastasis establishment of cancer cells. Cell adhesion is strictly regulated through numerous switches: proteins, glycoproteins and other components of the cell membrane. Recently, several cell membrane enzymes have been reported to participate in distinct steps of the cell adhesion process. Here, we review a variety of examples of membrane bound enzymes participating in adhesion of immune cells.
Introduction
Enzymes play crucial roles in all life processes and its perpetuation because they facilitate most biochemical reactions, both within and outside the cells. Enzyme function is determined not only by substrate specificity and enzymatic properties, but, notably, by its cell compartmentalization. Ectoenzymes are membrane-bound enzymes which the catalytic site is found outside the cell, they mainly reach its position by means of vesicles or carrier proteins (1). Ectoenzymes are a widely heterogeneous class of enzymes essential to homeostasis maintenance. They regulate the concentration and activity of certain molecules in the extracellular milieu, such as hormones, nucleotides, bioactive peptides (2), etc. Several ectoenzymes have been reported to perform other functions aside of catalyzing chemical reactions (3), reason for which they have been included in the list of moonlighting proteins (http://www.moonlightingproteins.org/). A moonlighting protein has multiple functions that are not the result of gene fusion, distinct RNA splice variants, or proteolytic fragments. It has been postulated that moonlighting proteins originally had a unique function but have acquired others, in many cases by virtue of post-translational modifications. The origin and the enzymatic functions of moonlighting proteins are thoroughly discussed elsewhere (3, 4).
Some moonlighting proteins are enzymes participating in cell adhesion processes (5–7). Cell adhesion enables cellular organization and communication in multicellular organisms. At the molecular level, it is a finely orchestrated process that includes the activation of canonical adhesion molecules, which trigger signal transduction cascades (8) and ultimately drive cell attachment to other cells or to the extracellular matrix (ECM). In leukocytes, cell adhesion is a critical process not only for cellular distribution in tissues, but also for establishing immunological synapses, mobilization, and migration. Such phenomena entail the coordination of several proteins. For example, leukocyte transendothelial migration requires molecules involved in protein-protein interactions that cause the deceleration of cells in circulation, proteins mediating the firm attachment of leukocytes to the endothelium and their squeezing through the endothelial junctions, and finally other proteins that mediate the active movement of cells through the tissues. Throughout these processes, membrane receptors initiate signal transduction pathways that regulate the cell’s adhesion properties and its changes in morphology. Therefore, it becomes evident that membrane adhesion molecules are activated at different time points to mediate either the approaching of cells’ membranes or strengthen their interaction, while in the intracellular compartment the cytoskeleton is rearranged (9). Here, we review the role of several ectoenzymes expressed by leukocytes involved in different steps of cell adhesion, and thus are considered as moonlighting proteins (Table 1 and Figure 1).
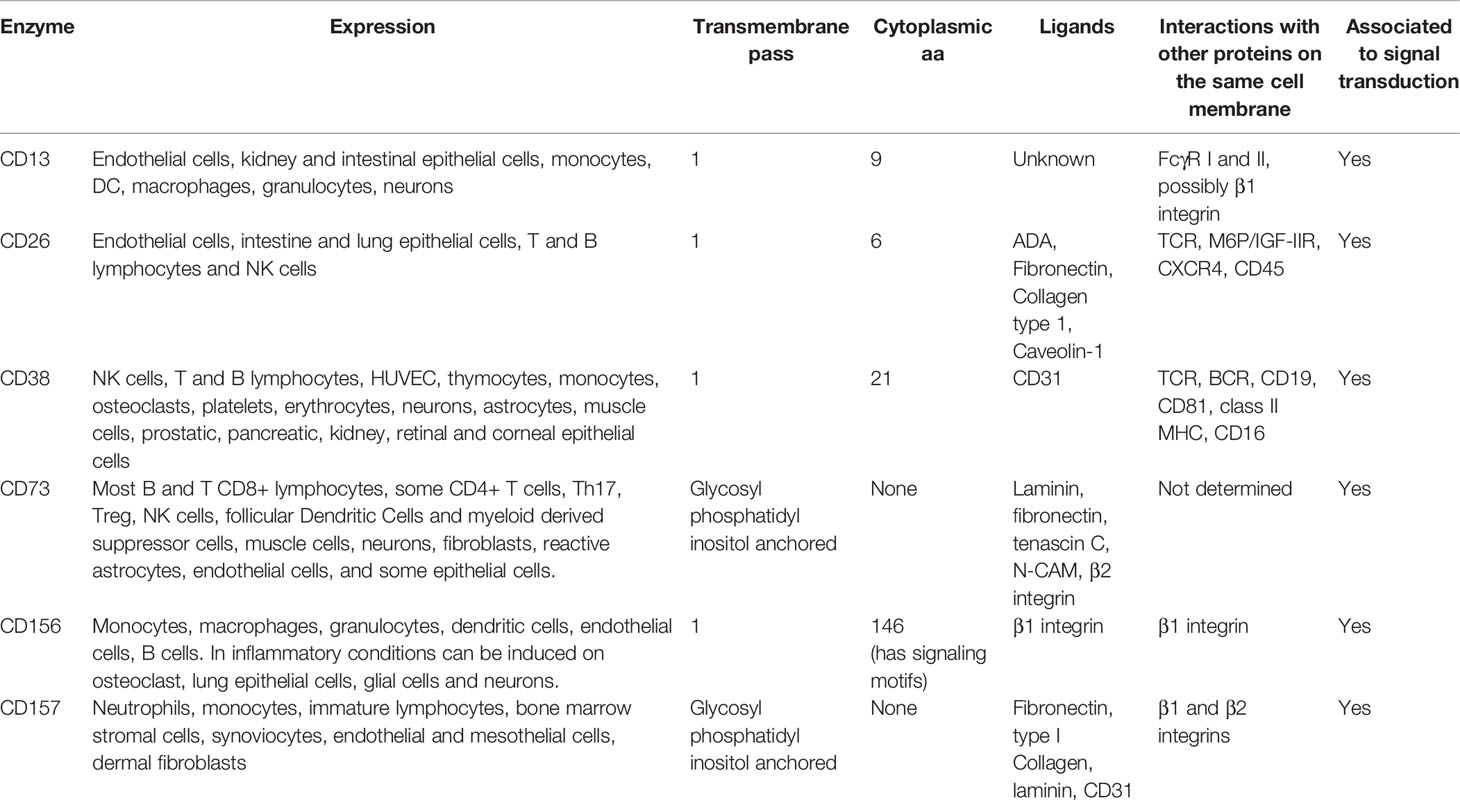
Table 1 Expression and properties of the membrane enzymes that participate in cell adhesion in leukocytes.
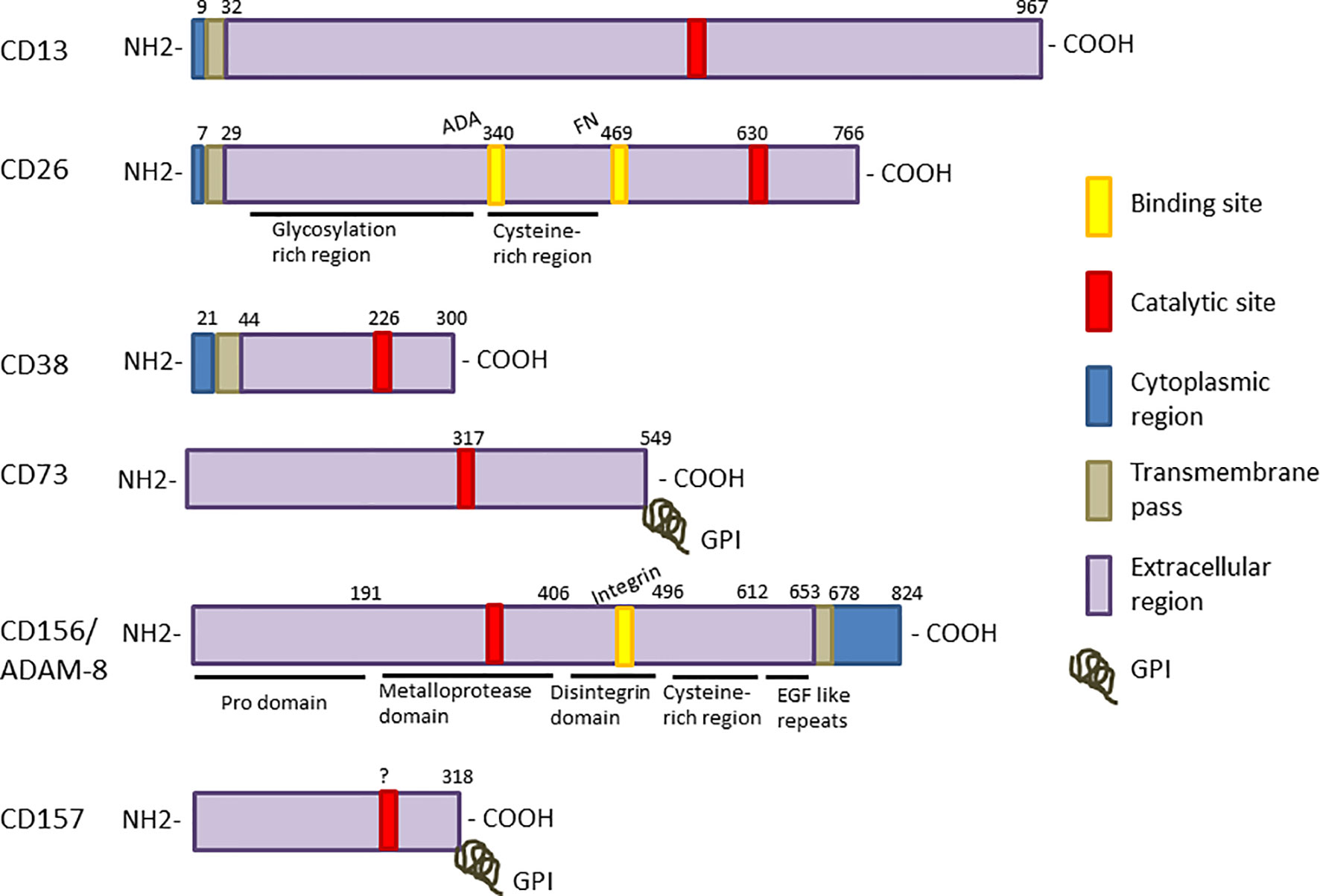
Figure 1 Linear representation each enzyme sequence. The different regions are displayed as color boxes: catalytic sites are red and the known binding sites for specific substrates, yellow. Intracellular regions are shown in blue, trans-membrane regions are represented as grey boxes, and extracellular regions are shown in purple. CD73 and CD157 are bound to the membrane through a glycosylphosphatidyl inositol molecule (GPI), symbolized by a grey coil. CD156/ADAM-8 is the only enzyme with signal transduction motifs (SH3 binding domains) in the cytoplasmic region.
CD13
CD13, also Aminopeptidase N (ANPEP, gp150) (EC: 3.4.11.2) is a Zn2+-dependent metalloproteinase that catalyzes the cleavage of neutral amino acids at the N-terminal portion of peptides. CD13 is highly glycosylated and has a large extracellular region, a single transmembrane pass and a short cytoplasmic tail (10, 11). Through its enzymatic activity, CD13 modulates different processes, as it cleaves a wide variety of bioactive peptides, including Angiotensin III, Enkephalin, some cytokines, and others (10). CD13 is expressed by myeloid cells, endothelium, renal and intestinal epithelial cells, neurons and, importantly, it is overexpressed in various cancer types (12–15).
CD13 is particularly highly expressed in myeloid cells and leukemias, hence, it was used as a marker of myeloid cells even before it was known to be the same as the membrane enzyme Aminopeptidase N (16). CD13 expressed in myeloid cells has since been shown to participate in phagocytosis (17–19), cell adhesion (20, 21) and migration (22, 23). Evidence shows that CD13 participation in these processes is dependent on the activation of signal transduction cascades, and, at the same time, independent of the enzymatic activity, leading to the idea that CD13 could also be a receptor. Moreover, CD13 crosslinking with monoclonal antibodies triggers heterotypic and homotypic aggregation, i.e. between cells of different types or between cells of the same type, respectively. This is true for human neutrophils, monocytes and the monocytic cell line U-937 (21, 24). Cell aggregation induced by CD13 crosslinking with the anti-CD13 antibody mAb452 is dependent on signal transduction and independent of integrins (24). It is noteworthy that, while some anti-CD13 antibodies induce cellular aggregation (mAb 452, mAb MY7 and mAb WM15), others do not (mAb WM47) (21, 24), and there are two monoclonal antibodies (mAb E and mAb C) that disaggregate cell aggregates previously induced by mAb 452 (25). Little is known about the mechanisms involved, or about why different mAbs show different abilities to induce aggregation (24), but it is hypothesized that this could be related to the epitopes recognized by the mAbs.
Alongside well-studied adhesion molecules such as integrins and selectins, other molecules contribute or regulate transendothelial migration of leukocytes. Among them, CD13 participates by promoting monocyte adhesion to the endothelium (20), which is one of the initial stages of leukocyte exit from the blood vessels into the tissues. In a mouse model of peritoneal inflammation, the absence of CD13 reduced the amount of leukocyte infiltration, specifically of inflammatory monocytes, dendritic cells (DC) and neutrophils. The conclusion was that optimal monocyte infiltration is achieved when both monocytes and endothelial cells express CD13 (26).
Although there are reports identifying putative ligands (27), CD13 is still considered an orphan receptor because the natural ligand under physiological conditions remains undiscovered (10) and the short cytoplasmic tail opens the question of how it is able to transduce signals. On one hand, Riemann et al. observed, using Fluorescence Resonance Energy Transfer (FRET), that after crosslinking, CD13 co-localizes with FcγR I and II (CD64 and CD32, respectively) (28), suggesting that CD13 signaling activity stems from its association with molecules with canonical domains for signal transduction. On the other hand, it has been difficult to show the association of CD13 with other membrane proteins by immunoprecipitation. Nevertheless, it is clear that the signal transduction cascade involves CD13 phosphorylation in a Src-dependent manner (29), which causes the activation of the Focal Adhesion Kinase (FAK) and the approach of the scaffold protein IQGAP1, which mediates the remodeling of α-Actin (20, 22, 29, 30). Then, Ca2+ mobilization and PI3K activation ensue, leading to MAPK activation (31).
It was recently reported that CD13 participates in the recycling of β1 integrin (CD29). After the integrin activation by ECM-ligation, CD13 is phosphorylated and forms a complex with IQGAP1 and the small GTPase ARF6, which directs the β1 integrin to early endosomes (Rab5+) and then back to the membrane (22), this would result in an increase of the cell’s adhesive properties. Another report shows evidence that CD13 expression regulates the expression of Syndecan 1 (SNC1) and β4 integrin (CD104) by downregulating the activation of PKCδ (32), which would enhance adhesiveness. However, it is not clear whether CD13 directly modulates PKCδ activity, or it is an indirect effect. Additionally, it has been suggested that CD13 could also promote adhesion by interacting with the ECM, via molecules like Galectin-3 or fibronectin (33), or with other membrane molecules (24).
CD26
The moonlighting protein CD26 is an enzyme of great relevance in T cell biology. CD26 participates in interactions with several proteins during T cell activation, establishing physical contact with the Antigen Presenting Cells (APC) so it reaches an accurate activation level, and enabling cell-cell communication. As an enzyme, CD26 [dipeptidyl peptidase IV (DPPIV) or Adenosine Deaminase Complexing protein 2 (ADCP2)] (EC: 3.4.14.5) is central to the regulation of multiple processes via its serine exopeptidase activity. It cleaves dipeptides from the N-terminus of peptides with proline or alanine at the penultimate position (34). Some of its substrates include Glucagon-like protein 1 (GLP-1) and the Gastric Inhibitory Protein (GIP) (35) (both important in glucose metabolism), as well as Substance P, and various chemokines. CD26 is a type II membrane protein with a short 6-aa cytoplasmic tail, a single transmembrane pass and a large 738-aa extracellular region that includes the catalytic site (35). The extracellular portion of CD26 contains 2 domains: a C-terminal serine protease domain homologous to α/β-hydrolases, and a propeller domain comprising two subdomains: a cysteine-rich region and a highly glycosylated region. CD26 forms homodimers on the cell membrane (36).
CD26 is expressed on the endothelium, epithelial cells of the kidney, liver, lung, intestine and on T cells, some B and NK cells, as well as on myeloid cells (37). In activated T cells, CD26 participates in lymphocyte adhesion to endothelial cells through interaction with another membrane enzyme, adenosine deaminase (ADA), expressed on endothelial cells (38). It was shown that CD26-ADA interaction increases lymphocyte adhesion to the endothelium, and it was suggested that the increased adhesion is mediated by the integrin lymphocyte function-associated antigen-1 (LFA-1, CD11a/CD18), because CD26-ADA interaction promotes the high affinity conformation of the integrin (38). The mechanism through which LFA-1 is activated is not clear, but it certainly involves an inside-out signaling pathway. ADA is implicated in the immunomodulation of adenosine, and both CD26 and ADA are expressed at higher levels on T effector cells than on Treg cells.
It has been suggested that CD26 regulates multiple T-cell processes, including maturation, migration, activation and cytokine secretion (39). Several proteins involved in T-cell activation interact with CD26 in an enzymatic activity-independent manner, including Mannose 6-Phosphate/Insulin-like Growth Factor II receptor (M6P/IGF IIR), CD45 (40), Caveolin- 1, Fibronectin (FN), Collagen type 1, Streptokinase, CXCR4, Plasminogen type 2, HIV gp120 protein, human coronavirus MERS-CoV Spike protein and the extracellular Adenosine Deaminase (ADA) (23, 39, 41–44). Moreno et al. demonstrated a physical interaction between CD26-bound ADA with Adenosine Receptor 2 (A2R), possibly through a molecular bridge between lymphocytes expressing CD26 and DCs expressing A2R (45). Also, CD26 on T cells is presumed to interact with Caveolin-1 on APCs, which could contribute to the overall interaction between these cells (40). In fact, ADA and Caveolin-1 bind to CD26’s highly glycosylated region while FN, Collagen type 1, Plasminogen and Streptokinase bind to the cysteine-rich region (35).
Multiple reports establish the ability of CD26 to directly bind to FN and Collagen type I in the ECM (46). Metastatic cancer cells and blood-born cancer cells move toward different tissues via interactions between CD26 and a polymeric form of FN expressed in lung epithelial cells (34). Similarly, Sato et al. demonstrated in a T-anaplastic large cell lymphoma cell line (T-ALCL), Karpas 299, that CD26 modulates the phosphorylation of β1 integrin by inducing the activation of p38 Mitogen Activated Protein Kinase (MAPK) (47). This suggests that CD26 mediates cell adhesion to the ECM through MAPK-dependent phosphorylation and β1 integrin activation.
CD26 is considered as a potential target for cancer therapy, because it is a marker of multiple types of cancer cells (44, 48, 49). In a model using mice inoculated with Karpas 299 lymphoma cells expressing or not CD26, CD26 was shown to be necessary for tumor development (47). As aforementioned, CD26 participates in cell adhesion to the ECM and the endothelium through its binding sites for FN, Collagen type I and ADA. The anti-CD26 antibody 6A3, which blocks the FN binding site on CD26, decreased adhesion of cancer cells (50).
CD73
The enzyme ecto-5’ nucleotidase (Ecto-5’-NT) (EC: 3.1.3.5), CD73, or Lymphocyte-Vascular Adhesion Protein 2 (LVAP-2) is a glycosyl phosphatidyl inositol-anchored membrane protein involved in the metabolism of nucleotides. Ecto-5’-NT catalyzes the dephosphorylation of nucleotide monophosphates into their corresponding nucleosides (51, 52). It is expressed on the membrane as an homodimer, although it can be also found intracellularly and as a soluble form in circulation (52).
Minimal concentrations of nucleotides are released physiologically into the extracellular space as messengers (53), but injuries or mechanical damage may result in an increased release of nucleotides into the extracellular space, leading to an inflammatory response (54). Conversely, high concentrations of extracellular adenosine downregulate pain, inflammation, proliferation and cytokine secretion (53). Hence, CD73 has been proposed as a key regulator of inflammation and a prominent target in chronic pain (54, 55). CD73 and CD39 work together on the cell membrane regulating of the adenosine metabolism; for example, Treg cells and myeloid- derived suppressor cells (MDSCs) use the nucleotidase CD39 to cleave ATP into AMP, which then is dephosphorylated into adenosine by CD73 (56, 57). The nucleosides can subsequently be transported into the cell through purinergic receptors such as A2A (53), and once inside the cell, be phosphorylated again for different purposes, including cell proliferation (58, 59). The increased expression of CD73 and CD39 produce increased extracellular concentrations of adenosine, which contributes to the immunosuppressive microenvironment of tumors and it has been demonstrated that they both are regulated by the hypoxia- inducible factor- α (HIF-1α) (57). Thus, inhibition of the enzymatic activity of these enzymes has been proposed as a potential therapeutic strategy to combat cancer (60).
CD73 is widely expressed. In the hematopoietic lineage, it is present on mature B lymphocytes, some CD8+ T cells, Tregs, and Follicular Dendritic Cells (FDC) (61). Other non-hematopoietic cells also express CD73, such as fibroblasts, some epithelial and endothelial cells, skeletal muscle cells and neurons and, significantly, a variety of solid tumors (52, 62). In addition to its enzymatic activity, the role of CD73 in different immunological processes has also been studied, including lymphocyte activation, proliferation, cell adhesion, and the formation of germinal centers in secondary lymphoid organs (59, 63, 64).
As an adhesion molecule, CD73 participates in cell interactions with sulphated proteins of the ECM, such interactions potentially contribute to regulate invasiveness and metastasis of cancer cells (65). Chicken’s gizzard CD73 interacts with columns containing Laminin or Fibronectin bound to sepharose beads, which reduces its enzymatic activity (65–68). Human CD73 also interacts with the ECM protein Tenascin C, important for adhesion of MDA-MB-231 breast cancer cells; this interaction inhibits 75% of CD73’s enzymatic activity (69). These observations make it highly likely that these ligands bind near the catalytic site.
In addition to ECM proteins, there is evidence that CD73 interacts with CD56, Neural-Cell Adhesion Molecule (N-CAM) (64) and CD18, the β chain of the integrin Lymphocyte Function-Associated Antigen-1 (LFA-1, integrin αL/β2, CD11a/CD18), enabling cellular adhesion between endothelial cells and lymphocytes (70, 71). Moreover, CD73 is known to be chopped off upon crosslinking with mAbs on lymphocytes but not on endothelial cells, suggesting a mechanism for regulation of lymphocyte-endothelial cell adhesion (59). CD73 interaction with LFA-1 may be the major mechanism by which CD73 contributes to enhance cell adhesion of lymphocytes to endothelial cells and along with its enzymatic activity, it could represent a major function of this moonlighting protein (64).
CD38
The ADP-ribosyl cyclase 1 or CD38 (EC: 2.4.99.20, EC: 3.2.2.6) is a conserved enzyme with 10 cysteines that are essential for maintaining its tertiary structure and for its catalytic activity (72). CD38 catalyzes the reaction of breaking up nicotinamide adenine dinucleotide (NAD+) into adenine dinucleotide phosphate ribose (ADPR), cyclic ADP ribose cADPR and nicotinamide (73–75). cADPR is a second messenger that induces Ca2+ mobilization independently from 3,4,5-inositol triphosphate (IP3) (74, 76). CD38 can also cleave NAD+ into its precursors, releasing nicotinamide mononucleotide and nicotinamide riboside (77). The enzyme can be found on the cell membrane with its catalytic domain either on the extracellular space or facing the cytoplasm, or on intracellular membranes (77). CD38 is a 300-aa long, type II membrane glycoprotein with a short cytoplasmic tail, a single transmembrane pass and the majority of the protein on the outer proportion of the plasma membrane (78, 79).
CD38 is considered a moonlighting enzyme because, aside its enzymatic function, it has been proposed to act as a co-receptor for various cell activation molecules. CD38 is expressed by naïve T cells, a subset of regulatory T cells, chronic infection-related T CD8+ cells, and thymocytes at the double positive stage (80, 81). When T lymphocytes are activated, CD38 synthesis increases, via Protein Kinase A and C (PKA PKC)-dependent pathway (82). Expression of CD38 in human B lymphocytes starts from early differentiation stages in the bone marrow, and it is maintained in mature B cells. In murine B lymphocytes CD38 expression is downregulated in cells entering germinal centers (83). Circulating monocytes express low levels of CD38, but its expression is increased by inflammatory stimuli and, interestingly, this is associated with differentiation into DCs (84). Other cells that express CD38 are NK cells, granulocytes and non-immune cells such as osteoclasts, erythrocytes, platelets, pancreatic and prostatic epithelial cells, neurons, astrocytes, muscle cells, renal tubular cells, retinal ganglion and cornea cells (85, 86).
Since CD38 has a short cytoplasmic tail, it was hypothesized that this molecule should interact with other membrane proteins to initiate intracellular signaling after activation upon antibody ligation. CD38 crosslinking in fact induces several cellular responses. Firstly, CD38 ligation on NK, T and B lymphocytes leads to proliferation and upregulation of activation markers (81). This molecule also participates during cell activation, potentiating signal transduction due to its close proximity to receptors mediating cell activation, such as the CD3/TCR complex in T cells, the surface Immunoglobulin (sIg), CD19, and CD81 in B cells, MHC class II in monocytes and CD16 in NK cells (77, 87). In naïve T cells and monocytes, CD38 crosslinking induces the secretion of both Th1 (IFNγ, M-CSF, IL-1β, IL-6) and Th2 cytokines (IL-4, IL-10 and IL-5) (73, 88). Activated neutrophils use CD38 to produce cyclic ADPR, which induces the release of intracellular Ca2+ that is required for responding to fMLP (75). In B lymphocytes, opposite consequences of CD38 activation have been reported: while its stimulation prevents apoptosis of B cells during clonal expansion in germinal centers, it also induces apoptosis of B cells precursors in the bone marrow, as well as of thymocytes in the thymus (80, 89, 90).
Regarding the role of lymphocytic CD38 in cell adhesion and migration, Dianzani, et al. first proposed that it mediates weak interactions between lymphocytes and the endothelium during the first steps of adhesion, before the stronger interactions occur (e.g. through integrins). They described two mechanisms for the participation of CD38 in adhesion: a direct interaction with an endothelial molecule, or by enabling the interaction of different lymphocytic membrane molecules with an endothelial ligand (91). A few years later, the same group reported that CD38 physically interacts with CD31 on endothelial cells and that this interaction leads to Ca2+ mobilization, which in turn leads to cytokine release, cell activation and cell-cell interaction with endothelial cells (92). Also, ligation induces CD38 molecules to aggregate in cholesterol and sphingolipids-rich membrane microdomains, which facilitates their internalization (93). It is plausible that such mechanism occurs during the first steps of adhesion, decelerating leukocytes in the bloodstream.
CD38–CD31 interactions are also established between human monocyte-derived dendritic cells (MDDCs) and endothelial cells. Migration induced by CCL-21 is inhibited either by blocking CD38 on MDDCs or CD31 on endothelial cells with specific antibodies (84). However, interaction with CD31 is not the only way that CD38 has for enabling cell migration; it was described that the reduced migration of DCs and neutrophils lacking CD38 is related to the lack of cADPR, which impairs Ca2+ mobilization (75, 92, 94). Thus, CD38 participates in migration in at least two ways: by promoting adhesion via CD31, and by the enzymatic production of cADPR, which induces Ca2+ mobilization required for migration.
CD38 ligation in T cells, either with monoclonal antibodies or with CD31, initiates a signal transduction cascade including the phosphorylation of PLC-γ, ZAP-70, MAPK and ERK2; whereas in monocytes the signaling pathway comprises PI3K and c-Cbl (73, 95, 96). This is noteworthy due to the fact that CD38 has only a short cytoplasmic tail with no known signaling motifs. Consequently, it is hypothesized that CD38 uses the signaling machinery of other molecules, such as the BCR, TCR, CD16 and MHC class II. The study of CD38 as a receptor started more than three decades ago and, as is true for several other cell membrane enzymes, more recently it has been refocused toward its expression and function in cancer cells, leading to the proposal of CD38 as a target for anti-cancer drugs (97–99).
CD157
The ectoenzyme ADP-Ribosyl cyclase 2 (EC: 3.2.2.6), also known as CD157 or Bone Marrow Stromal Cell Antigen-1 (BST-1), is a glycosylphosphatidyl inositol-anchored glycoprotein with NAD glycol-hydrolase activity and ADP-ribosyl cyclase activity, like CD38, which belongs to the same gene family. They share 36% of their sequence and have similar pleiotropic functions both as receptors and as enzymes. Although CD38 has a transmembrane pass while CD157 does not, their structures are supported by 10 cysteines essential for disulphide-bond formation and enzymatic activity (78).
CD157 is expressed on myeloid cells, immature lymphocytes, bone marrow stromal cells, synoviocytes, endothelial and mesothelial cells, and dermal fibroblasts. Invasiveness of ovarian cancer epithelial cells is CD157-dependent, which increased expression promotes mesenchymal differentiation (100). The expression of CD157 is upregulated on neutrophils and basophils upon fMLP stimulation and on monocytes treated with MCP-1 (101–103), expression levels do not change on activated endothelial cells (104). CD157 expression on human umbilical vein endothelial cells (HUVEC) is restricted mainly to the intercellular region of membranes (104). CD157 plays a key role in transendothelial migration of neutrophils, as treating neutrophils with an anti-CD157 blocking antibody or treatment of HUVEC cells with an anti-CD31 blocking antibody, hinders their transendothelial migration (104). In humans, paroxysmal nocturnal hemoglobinuria is a genetic defect characterized by the attachment of glycolipids to proteins resulting in the absence of CD157 and other GPI-anchored proteins on the cell membrane, neutrophils from patients with this condition show impaired transmigration even though neutrophils adhere to the vessel wall (104).
CD157, as other GPI-anchored proteins, interacts with different transmembrane molecules to transduce signals. The antibody-induced clustering of CD157 on the cell membrane initiates a Ca2+-dependent cascade, F-actin reorganization toward the opposite pole of the cell, and β2 integrins activation in neutrophils (102). Moreover, CD157 co-localizes and functionally collaborates with β1 and β2 integrins for adhesion to Fibrinogen and Fibronectin (102, 103), inducing a signal transduction pathway that involves FAK, Src, AKT and ERK 1/2 (103). As long as β2 integrin CD18 is associated with CD11b to form MAC-1 (integrin αM/β2), CD157 co-immunoprecipitates with this heterodimer (105, 106), suggesting that CD157 can trigger intracellular signals using the MAC-1 (CD11b/CD18) signal transduction machinery.
Soluble recombinant CD157 binds to various ECM proteins such as Fibronectin, Fibrinogen, Collagen type I and Laminin, and these interactions can be inhibited by an anti-CD157 mAb or by heparin (107). Thus, CD157 participation in cellular adhesion of leukocytes may involve i) its association with integrins on the same cell membrane, ii) the interaction with components of the ECM and iii) direct interaction of CD157 with CD31 on endothelial cells. It is thus conceivable that CD157 interaction with CD31 is employed for transendothelial migration, while the interaction with Laminin, Collagen type I and Fibronectin could be involved in the cell´s migration through the tissues, enhancing the integrin-mediated adhesion to ECM. Finally, even though CD157 does not have signal transduction domains per se, its association with the CD18/CD11b heterodimer suggests that it may use the signal transduction cascade of this integrin.
CD156
Human CD156, or ADAM-8 (A Disintegrin and Metalloproteinase domain-containing protein-8 (EC: 3.4.24.-)) is a type I transmembrane glycoprotein. ADAMs are conserved proteins that were initially related to the hemorrhagic snake venom protein (HSVP), which has a cysteine-rich region preceding the metalloproteinase domain. Structurally, ADAMs have a pro-domain that inhibits the active form of the enzyme, the catalytic domain, the disintegrin domain, a cysteine-rich region, a single transmembrane pass and a cytoplasmatic tail with signal transduction function (108). The intracellular tail of ADAM-8 has a proline-rich region similar to the SH3 (Src homology 3) binding sequence (109). ADAM-8 has a long list of substrates, including cell adhesion molecules, cytokine receptors and ECM proteins (110). ADAM family are metalloproteinases that bind to integrins through a RGD motif (Arg-Gly-Asp), inhibiting platelet aggregation (111).
Under physiological conditions, protein levels of ADAM-8 are low in monocytes, macrophages, granulocytes, dendritic cells, and endothelial cells, and even lower in B cells and neurons. But it has been reported that CD156 is upregulated by different inflammatory stimuli, such as Tumor Necrosis Factor-α (TNF-α) (112), Interleukins (IL) 4 and 13 (113), Lipopolysaccharide (LPS), Interferon-γ (IFN-γ), and by Peroxisome Proliferator-Activated Receptor-γ (PPAR-γ) (114, 115). ADAM-8 expression is also induced under inflammatory conditions in lung epithelium (113), osteoclasts (116), and glial cells in the central nervous system (CNS) (112).
ADAM-8 and ADAM-28 are the only members of the family that do not have a canonical sequence for Furin-like convertases between the pro-domain and the metalloproteinase domain (117). Thus, activation is achieved after their multimerization on the membrane and auto-catalytical cleavage of the pro-domain (117). The disintegrin domain has an integrin-binding loop with the residues KDM, followed by a cysteine-rich region and an EGF-like sequence. Interestingly, three different forms of ADAM-8 have been detected in cells: the inactive form with its pro- domain, the active form without the pro-domain, and the final form resulting from the removal of the metalloproteinase domain, leaving the disintegrin domain as the N-terminal domain (117). The first two forms are much less abundant than the last one. The remaining peptide (named DCE) consists of the disintegrin domain, and the cysteine-rich and EGF-like sequences; DCE participates in homophilic interactions with other cells (109, 117). Although the precise amino acids implicated have not been identified, the interactions with its ligands may include disulphide bonds, since reducing conditions impede adhesion (117).
CD156 molecules from both the endothelial cell and the leukocyte participate in the infiltration of myeloid cells into inflamed tissue. Migration of the monocytic human THP-1 cell line directed by CCL-2 was reduced when CD156 was downregulated using a shRNA. Similarly, reduced levels of CD156 diminished the migratory response of human neutrophils towards CXCL8. This effect on the migratory capacity was achieved either by chemically inhibiting CD156 enzymatic activity or by blocking the enzyme with monoclonal antibodies (115). Furthermore, expression of the α subunit of the integrin LFA-1 (integrin αL, CD11a) in monocytes is upregulated in response to CCL-2, but silencing of CD156 affected this upregulation, decreasing even more the adhesion of leukocytes to endothelial cells (115). These results were confirmed in an in vivo model where monocyte infiltration to inflamed lungs was less significant in ADAM-8-/- than in wild type animals. Wound healing assays revealed that ADAM-8-/- endothelial cells have significantly reduced migratory capacity (115). Although the mechanism is not yet completely understood, the evidence shows that migration is dependent on the enzymatic activity.
In order to accomplish cell adhesion, signal transduction through ADAM-8/CD156 is possible because unlike the other enzymes previously discussed, this enzyme has signaling motifs in its cytoplasmic tail. Potential candidates that could modulate the cascade are Src protein kinases (109), Cdc42-dependent actin assembly protein 1 (TOCA-1), and Cdc42-interacting protein 4 (CIP4) (110, 118). Additionally, ADAM-8/CD156 disintegrin domain interacts with β1 integrin inducing the FAK, ERK and protein kinase B (AKT) activation (110, 119). Precisely these pathways are implicated in ADAM-8-driven chemo-resistance and enhanced invasiveness of human glioblastoma cells (119). This suggests that ADAM-8 either modulates, or initiates signal transduction cascades and that it may be a potential therapeutic target in cancer.
Conclusions
The hypothesis that ectoenzymes could transmit biochemical signals to the intracellular space was formulated before the term of moonlighting proteins was even coined (1). According to Stanley et al., all ectoenzymes must be integral proteins of the cell membrane with the catalytic site on the extracellular face of the membrane (1). However, ectoenzymes can also be found in other intracellular compartments, making larger the list of possible functions for each enzyme. Currently, several ectoenzymes are included in the category of moonlighting proteins, implying that these enzymes perform different functions depending on the cell type that expresses them, the timing, and the microenvironment, while they preserve their catalytic activity. Furthermore, in the past few years, a growing number of moonlighting enzymes expressed on immune cells have been shown to play a role in complex processes such as cellular adhesion, phagocytosis, and cell activation, among others.
Ectoenzymes constitute a heterogeneous category of proteins that regulate multiple physiological processes through their different enzymatic properties, by cleaving peptides and hormones, processing molecules, clearing injured tissue, etc. (2). The products of their enzymatic activity trigger functional cellular responses that contribute to maintaining the homeostasis of the organism. The membrane enzymes discussed (Figure 2) contribute to the 4% of human leukocytes’ surface covered by ectoenzymes (7).
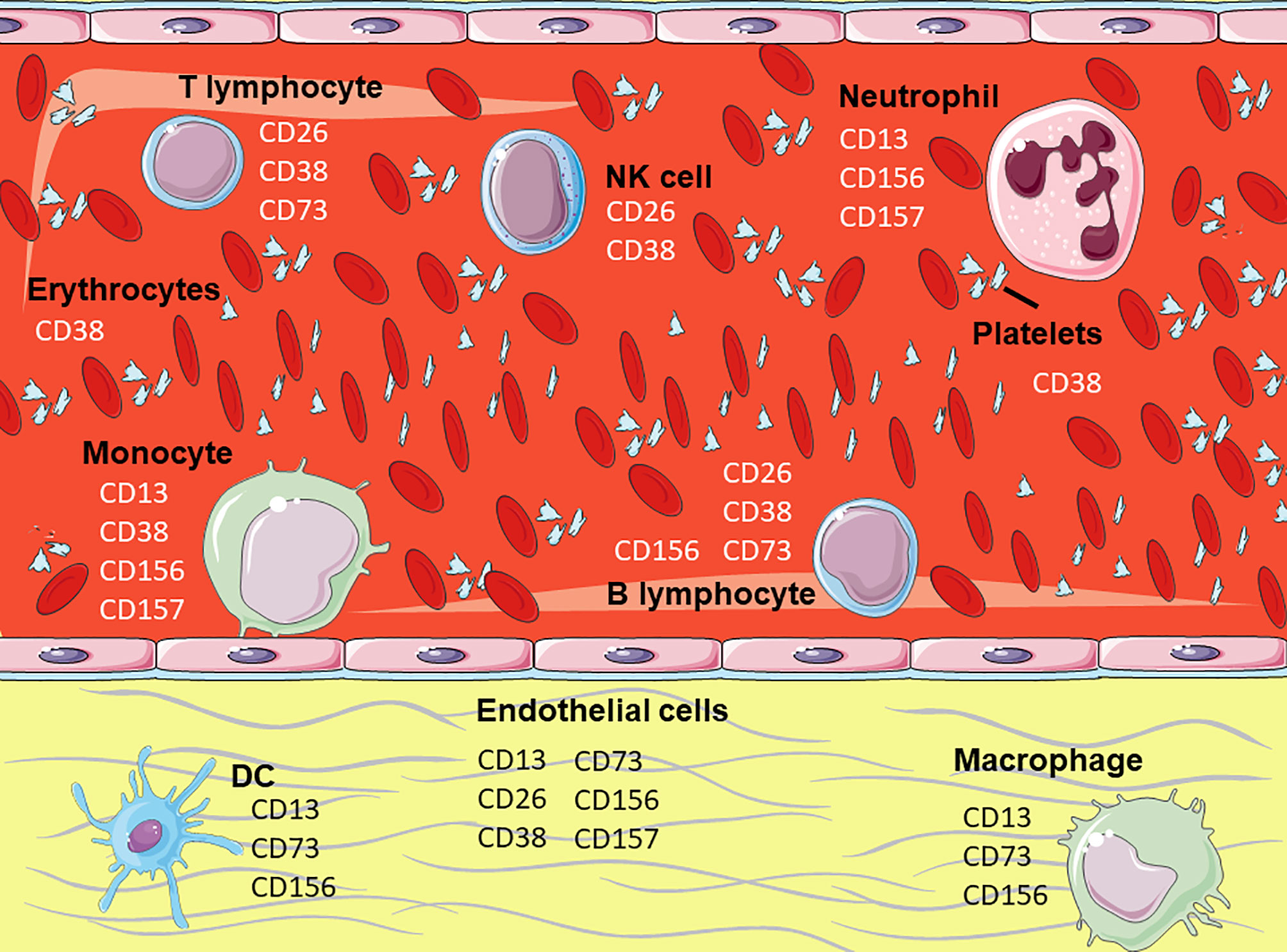
Figure 2 Expression of the membrane enzymes on hematopoietic cells. Representation of hematopoietic and endothelial cells highlighting the membrane enzymes involved in cell adhesion. Figure designed using images from Servier Medical Art (https://smart.servier.com/).
Each subpopulation of immune cells is endowed with a variety of specialized proteins that engage in processes such as phagocytosis, immunological synapses, antigen presentation, chemotaxis, and others. These add up to a variety of highly specialized mechanisms that maintain homeostasis, respond to danger signals, and scrutinize through the entire organism as sentinels. In certain cells, a number of enzymes have evolved to participate in cell adhesion. These ectoenzymes were probably selected not because they had conserved domains relevant for adhesion, but instead because certain chemical features of their structure enabled the cell to establish interactions with molecules on the surface of a different cell. Ultimately, chemical interactions and affinity to specific motifs are the initial motor for cell adhesion. This may have happened with some ectoenzymes on immune cells which suffered a positive selective pressure. In time, they became part of the cell´s array of molecules that mediate cell adhesion. However, the mechanisms by which these enzymes became capable of signal transduction should be part of a different evolutionary history. We hypothesize that the interaction of these enzymes with their ligands induced the aggregation of other adhesion molecules on the membrane that were included in the same microdomains (e.g. lipid rafts), and such molecules were originally responsible for signal transmission.
Since enzymes function in an orchestrated fashion, their expression and co-expression must be considered when formulating hypothesis on their evolution and physiology. For example in Jurkat and Raji cells, CD38 induces a dynamic mobilization of pre-synthesized cytoplasmic CD73 toward the membrane, where both function in the same axis (120). The biological significance of this synergism is still unclear; as both ectoenzymes share substrates, participate in lymphocyte activation and in the adhesion to the endothelium. In the case of CD73 and CD39, another nucleotidase, they take part side by side in the same ATP metabolic pathway, showing critical co-participation in the regulation of metabolism (55, 121). Interestingly, this cascade is used by several cancer cells and Treg cells to downregulate the immune response (55, 122). On the contrary, few cells co-express CD38 and CD157, which function similarly regarding their enzymatic and receptor activities (101).
In addition to ADAM-8, other ADAM proteins may be involved in cell adhesion. Distinct members of this family are overexpressed on cancer cells, and some of them have been shown to provide resistance to chemotherapeutic agents (123). Their deregulation during pathologies like cancer, reveal the importance of not only their catalytic activity, but also of the protein-protein interactions mediated by the disintegrin domain of ADAM proteins. For example, ADAM-12 and ADAM-15 bind to the α9β1 (124) and αvβ3 integrins (125); ADAM-9 to α6β1 integrin (126) and ADAM-23 to αvβ3 integrin (127). Although it is speculated that these interactions contribute to tumor growth and invasion, the fact that these same interactions are formed by ADAM molecules expressed on leukocytes awaits confirmation.
Cell adhesion is an ancient mechanism present since unicellular organisms, as it serves for cell communication. Thus, it is not surprising that many molecules have been evolutionarily selected to participate in such a complex process. Perhaps, selective pressures drove proteins toward the acquisition of post-translational modifications which made them part of this process. Finding other molecules that participate in cellular adhesion would help gaining a deeper understanding of the process per se. Also, as aforesaid, most of the molecules mentioned here are overexpressed in various cancers. However there are other disorders caused by adhesion molecules deregulated that could be treated more efficiently. This is the reason why many pharmacological developments attempt to target specific tissues with certain expression profiles (128). Furthermore, cell markers are especially useful for developing drugs, opening the door for more targeted and, hopefully, virtually side-effects free treatments.
Author Contributions
GL-C: Conceptualization and writing the original draft. LD-A and EO manuscript critical revision. All authors contributed to the article and approved the submitted version.
Funding
Work in the authors´ laboratory is funded by grants from CONACYT (Grant 87163) and PAPIIT-DGAPA-UNAM (IN 208320).
Conflict of Interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s Note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Acknowledgments
GL-C is a doctoral student from the Program Maestría y Doctorado en Ciencias Bioquímicas, Universidad Nacional Autónoma de México (UNAM), and gratefully thanks the fellowship (699886) received from CONACyT to pursue her PhD. LD-A thanks her PhD Program Doctorado en Ciencias Biológicas, Universidad Nacional Autónoma de México (UNAM).
References
1. Stanley KK, Newby AC, Luzio JP. What Do Ectoenzymes Do? Trends Biochem Sci (1982) 7(4):145–7. doi: 10.1016/0968-0004(82)90207-9
2. Konkoy CS, Davis TP. Ectoenzymes as Sites of Peptide Regulation. Trends Pharmacol Sci (1996) 17(8):288–94. doi: 10.1016/0165-6147(96)10036-5
3. Jeffery CJ. Protein Species and Moonlighting Proteins: Very Small Changes in a Protein’s Covalent Structure can Change Its Biochemical Function. J Proteomics (2016) 134:19–24. doi: 10.1016/j.jprot.2015.10.003
4. Perez-Riverol Y, Da F, Leprevost V, Pazos F, Jeffery CJ. Why Study Moonlighting Proteins? Front Genet (2015) 1:211. doi: 10.3389/fgene.2015.00211
5. Salmi M, Jalkanen S. Ectoenzymes Controlling Leukocyte Traffic. Eur J Immunol (2012) 42(2):284–92. doi: 10.1002/eji.201142223
6. Salmi M, Jalkanen S. Ectoenzymes in Leukocyte Migration and Their Therapeutic Potential. Semin Immunopathol (2014) 36(2):163–76. doi: 10.1007/s00281-014-0417-9
7. Salmi M, Jalkanen S. Cell-Surface Enzymes in Control of Leukocyte Trafficking. Nat Rev Immunol (2005) 5(10):760–71. doi: 10.1038/nri1705
8. Parsons JT, Horwitz AR, Schwartz MA. Cell Adhesion: Integrating Cytoskeletal Dynamics and Cellular Tension. Nat Rev Mol Cell Biol (2010) 11(9):633–43. doi: 10.1038/nrm2957
9. Khalili AA, Ahmad MR. A Review of Cell Adhesion Studies for Biomedical and Biological Applications. Int J Mol Sci (2015) 16:18150. doi: 10.3390/ijms160818149
10. Mina-Osorio P. The Moonlighting Enzyme CD13: Old and New Functions to Target. Trends Mol Med (2008) 14(8):361–71. doi: 10.1016/j.molmed.2008.06.003
11. Lu C, Amin MA, Fox DA. CD13/Aminopeptidase N Is a Potential Therapeutic Target for Inflammatory Disorders. J Immunol (2020) 204(1):3–11. doi: 10.4049/jimmunol.1900868
12. Wickström M, Larsson R, Nygren P, Gullbo J. Aminopeptidase N (CD13) as a Target for Cancer Chemotherapy. Cancer Sci (2011) 102(3):501–8. doi: 10.1111/j.1349-7006.2010.01826.x
13. Guo Q, Jing FJ, Xu W, Li X, Li X, Sun JL, et al. Ubenimex Induces Autophagy Inhibition and EMT Suppression to Overcome Cisplatin Resistance in GC Cells by Perturbing the CD13/EMP3/PI3K/AKT/NF-κb Axis. Aging (Albany NY) (2020) 12(1):80–105. doi: 10.18632/aging.102598
14. Haraguchi N, Ishii H, Mimori K, Tanaka F, Ohkuma M, Kim HM, et al. CD13 Is a Therapeutic Target in Human Liver Cancer Stem Cells. J Clin Invest (2010) 120(9):3326–39. doi: 10.1172/JCI42550
15. Zhang J, Fang C, Qu M, Wu H, Wang X, Zhang H, et al. CD13 Inhibition Enhances Cytotoxic Effect of Chemotherapy Agents. Front Pharmacol (2018) 9:1–10. doi: 10.3389/fphar.2018.01042
16. Look AT, Ashmun RA, Shapiro LH, Peiper SC. Human Myeloid Plasma Membrane Glycoprotein CD13 (Gp150) Is Identical to Aminopeptidase N. J Clin Invest (1989) 83(4):1299–307. doi: 10.1172/JCI114015
17. Licona-Limon I, Garay-Canales CA, Munoz-Paleta O, Ortega E. CD13 Mediates Phagocytosis in Human Monocytic Cells. J Leukoc Biol (2015) 98(1):85–98. doi: 10.1189/jlb.2A0914-458R
18. Mendoza-Coronel E, Ortega E. Macrophage Polarization Modulates Fcγr- and CD13-Mediated Phagocytosis and Reactive Oxygen Species Production, Independently of Receptor Membrane Expression. Front Immunol (2017) 8:303. doi: 10.3389/fimmu.2017.00303
19. Villaseñor-Cardoso MI, Frausto-Del-Río DA, Ortega E. Aminopeptidase N (CD13) Is Involved in Phagocytic Processes in Human Dendritic Cells and Macrophages. BioMed Res Int (2013) 2013:12. doi: 10.1155/2013/562984
20. Mina-Osorio P, Winnicka B, O’Conor C, Grant CL, Vogel LK, Rodriguez-Pinto D, et al. CD13 Is a Novel Mediator of Monocytic/Endothelial Cell Adhesion. J Leukoc Biol (2008) 84(2):448–59. doi: 10.1189/jlb.1107802
21. Fiddler CA, Parfrey H, Cowburn AS, Luo D, Nash GB, Murphy G, et al. The Aminopeptidase CD13 Induces Homotypic Aggregation in Neutrophils and Impairs Collagen Invasion. PloS One (2016) 11(7):e0160108. doi: 10.1371/journal.pone.0160108
22. Ghosh M, Lo R, Ivic I, Aguilera B, Qendro V, Devarakonda C, et al. CD13 Tethers the IQGAP1-ARF6-EFA6 Complex to the Plasma Membrane to Promote ARF6 Activation, β1 Integrin Recycling, and Cell Migration. Sci Signal (2019) 12(579):eaav5938. doi: 10.1126/scisignal.aav5938
23. Liang W, Gao B, Xu G, Weng D, Xie M, Qian Y. Possible Contribution of Aminopeptidase N (APN/CD13) to Migration and Invasion of Human Osteosarcoma Cell Lines. Int J Oncol (2014) 45(6):2475–85. doi: 10.3892/ijo.2014.2664
24. Mina-Osorio P, Shapiro LH, Ortega E. CD13 in Cell Adhesion: Aminopeptidase N (CD13) Mediates Homotypic Aggregation of Monocytic Cells. J Leukoc Biol (2006) 79(4):719–30. doi: 10.1189/jlb.0705425
25. Garay-Canales CA, Licona-Limón I, Ortega E. Distinct Epitopes on CD13 Mediate Opposite Consequences for Cell Adhesion. BioMed Res Int (2018) 2018. doi: 10.1155/2018/4093435
26. Ghosh M, Gerber C, Rahman MM, Vernier KM, Pereira FE, Subramani J, et al. Molecular Mechanisms Regulating CD13-Mediated Adhesion. Immunology (2014) 142(4):636–47. doi: 10.1111/imm.12279
27. Nefla M, Sudre L, Denat G, Priam S, Andre-Leroux G, Berenbaum F, et al. The Pro-Inflammatory Cytokine 14-3-3 Is a Ligand of CD13 in Cartilage. J Cell Sci (2015) 128(17):3250–62. doi: 10.1242/jcs.169573
28. Riemann D, Tcherkes A, Hansen GH, Wulfaenger J, Blosz T, Danielsen EM. Functional Co-Localization of Monocytic Aminopeptidase N/CD13 With the Fcγ Receptors CD32 and CD64. Biochem Biophys Res Commun (2005) 331(4):1408–12. doi: 10.1016/j.bbrc.2005.04.061
29. Subramani J, Ghosh M, Rahman MM, Caromile LA, Gerber C, Rezaul K, et al. Tyrosine Phosphorylation of CD13 Regulates Inflammatory Cell-Cell Adhesion and Monocyte Trafficking. J Immunol (2013) 191(7):3905–12. doi: 10.4049/jimmunol.1301348
30. Resheq YJ, Menzner A-K, Bosch J, Tickle J, Li K-K, Wilhelm A, et al. Impaired Transmigration of Myeloid-Derived Suppressor Cells Across Human Sinusoidal Endothelium Is Associated With Decreased Expression of CD13. J Immunol (2017) 199(5):1672–81. doi: 10.4049/jimmunol.1600466
31. Santos N, Langner J, Herrmann M, Riemann D. Aminopeptidase N/CD13 Is Directly Linked to Signal Transduction Pathways in Monocytes. Eur PMC (2000) 201(1):22–32. doi: 10.1006/cimm.2000.1629
32. Cheng Y-C, Ku W-C, Tseng T-T, Wu C-P, Li M, Lee S-C. Anchorage Independence Altered Vasculogenic Phenotype of Melanoma Cells Through Downregulation in Aminopeptidase N /Syndecan-1/Integrin B4 Axis. Aging (Albany NY) (2020) 12(17):16803–19. doi: 10.18632/aging.103425
33. Mina-Osorio P, Soto-Cruz I, Ortega E. A Role for Galectin-3 in CD13-Mediated Homotypic Aggregation of Monocytes. Biochem Biophys Res Commun (2007) 353(3):605–10. doi: 10.1016/j.bbrc.2006.12.081
34. Cheng H-C, Abdel-Ghany M, Pauli BU. A Novel Consensus Motif in Fibronectin Mediates Dipeptidyl Peptidase IV Adhesion and Metastasis. J Biol Chem (2003) 278(27):24600–7. doi: 10.1074/jbc.M303424200
35. Klemann C, Wagner L, Stephan M, von Hörsten S. Cut to the Chase: A Review of CD26/dipeptidyl Peptidase-4’s (DPP4) Entanglement in the Immune System. Clin Exp Immunol (2016) 185:1–21. doi: 10.1111/cei.12781
36. Engel M, Hoffmann T, Wagner L, Wermann M, Heiser U, Kiefersauer R, et al. The Crystal Structure of Dipeptidyl Peptidase IV (CD26) Reveals Its Functional Regulation and Enzymatic Mechanism. PNAS2 (2003) 100(9):5063–8. doi: 10.1073/pnas.0230620100
37. Abbott CA, McCaughan GW, Baker E, Sutherland GR. Genomic Organization, Exact Localization, and Tissue Expression of the Human CD26 (Dipeptidyl Peptidase IV) Gene. Immunogenetics (1994) 40(5):331–8. doi: 10.1007/BF01246674
38. Gine S, Marinm M, Mallol J, Canela EI, Morimoto C, Callebaut C, et al. Regulation of Epithelial and Lymphocyte Cell Adhesion by Adenosine Deaminase-CD26 Interaction. Biochem J (2002) 361:203–9. doi: 10.1042/0264-6021:3610203
39. Morimoto C, Schlossman SF. The Structure and Function of CD26 in the T-Cell Immune Response. Immunol Rev (1998) 161:55–70. doi: 10.1111/j.1600-065X.1998.tb01571.x
40. Fan H, Tansi FL, Weihofen WA, Böttcher C, Hu J, Martinez J, et al. Molecular Mechanism and Structural Basis of Interactions of Dipeptidyl Peptidase IV With Adenosine Deaminase and Human Immunodeficiency Virus Type-1 Transcription Transactivator. Eur J Cell Biol (2012) 91:265–73. doi: 10.1016/j.ejcb.2011.06.001
41. Kameoka J, Tanaka T, Nojima Y, Schlossman SF, Morimoto C. Direct Association of Adenosine Deaminase With a T Cell Activation Antigen, CD26. Sci (80- ) (1993) 261(5120):466–9. doi: 10.1126/science.8101391
42. Lu G, Hu Y, Wang Q, Qi J, Gao F, Li Y, et al. Molecular Basis of Binding Between Novel Human Coronavirus MERS-CoV and Its Receptor CD26. Nature (2013) 500(7461):227–31. doi: 10.1038/nature12328
43. Lambeir AM, Durinx C, Scharpé S, De Meester I. Dipeptidyl-Peptidase IV From Bench to Bedside: An Update on Structural Properties, Functions, and Clinical Aspects of the Enzyme DPP Iv. Crit Rev Clin Lab Sci (2003) 40:209–94. doi: 10.1080/713609354
44. Havre PA, Abe M, Urasaki Y, Ohnuma K, Morimoto C, Dang NH. The Role of CD26/dipeptidyl Peptidase IV in Cancer. Front Bioscience (2008) 13. doi: 10.2741/2787
45. Moreno E, Canet J, Gracia E, Lluís C, Mallol J, Canela EI, et al. Molecular Evidence of Adenosine Deaminase Linking Adenosine A2A Receptor and CD26 Proteins. Front Pharmacol (2018) 9:106. doi: 10.3389/fphar.2018.00106
46. Hanski C, Huhle T, Reutter W. Involvement of Plasma Membrane Dipeptidyl Peptidase IV in Fibronectin-Mediated Adhesion of Cells on Collagen. Biol Chem Hoppe-Seyler (1985) 366:1169–76. doi: 10.1515/bchm3.1985.366.2.1169
47. Sato T, Yamochi T, Yamochi T, Aytac U, Ohnuma K, Mckee KS, et al. CD26 Regulates P38 Mitogen-Activated Protein Kinase-Dependent Phosphorylation of Integrin B 1 , Adhesion to Extracellular Matrix, and Tumorigenicity of T-Anaplastic Large Cell Lymphoma Karpas 299. Cancer Res (2005) 65(15):6950–6. doi: 10.1158/0008-5472.CAN-05-0647
48. Havre PA, Dang LH, Ohnuma K, Iwata S, Morimoto C, Dang NH. CD26 Expression on T-Anaplastic Large Cell Lymphoma (ALCL) Line Karpas 299 Is Associated With Increased Expression of Versican and MT1-MMP and Enhanced Adhesion. BMC Cancer (2013) 13:517. doi: 10.1186/1471-2407-13-517
49. Nishida H, Hayashi M, Morimoto C, Sakamoto M, Yamada T. CD26 Is a Potential Therapeutic Target by Humanized Monoclonal Antibody for the Treatment of Multiple Myeloma. Blood Cancer J (2018) 8(11):99. doi: 10.1038/s41408-018-0127-y
50. Hung T-T, Wu J-Y, Liu J-F, Cheng H-C. Epitope Analysis of the Rat Dipeptidyl Peptidase IV Monoclonal Antibody 6A3 That Blocks Pericellular Fibronectin-Mediated Cancer Cell Adhesion. FEBS J (2009) 276(22):6548–59. doi: 10.1111/j.1742-4658.2009.07352.x
51. Knapp K, Zebisch M, Pippel J, El-Tayeb A, Müller CE, Sträter N. Crystal Structure of the Human Ecto-5′-Nucleotidase (CD73): Insights Into the Regulation of Purinergic Signaling. Structure (2012) 20(12):2161–73. doi: 10.1016/j.str.2012.10.001
52. Rahimova R, Fontanel S, Lionne C, Jordheim LP, Peyrottes S, Chaloin L. Identification of Allosteric Inhibitors of the Ecto-5’-Nucleotidase (CD73) Targeting the Dimer Interface. PloS Comput Biol (2018) 14(1):e1005943. doi: 10.1371/journal.pcbi.1005943
53. Burnstock G. Purinergic Signalling: Therapeutic Developments. Front Pharmacol (2017) 8:661. doi: 10.3389/fphar.2017.00661
54. Regateiro FS, Cobbold SP, Waldmann H. CD73 and Adenosine Generation in the Creation of Regulatory Microenvironments. Clin Exp Immunol (2013) 171(1):1–7. doi: 10.1111/j.1365-2249.2012.04623.x
55. Antonioli L, Pacher P, Vizi ES, Haskó G. CD39 and CD73 in Immunity and Inflammation. Trends Mol Med (2013) 19(6):355–67. doi: 10.1016/j.molmed.2013.03.005
56. Mandapathil M, Szczepanski MJ, Harasymczuk M, Ren J, Cheng D, Jackson EK, et al. CD26 Expression and Adenosine Deaminase Activity in Regulatory T Cells (Treg) and Cd4+ T Effector Cells in Patients With Head and Neck Squamous Cell Carcinoma. Oncoimmunology (2012) 1(5):659–69. doi: 10.4161/onci.20387
57. Li L, Wang L, Li J, Fan Z, Yang L, Zhang Z, et al. Metformin-Induced Reduction of CD39 and CD73 Blocks Myeloid-Derived Suppressor Cell Activity in Patients With Ovarian Cancer. Cancer Res (2018) 78(7):1779. doi: 10.1158/0008-5472.CAN-17-2460
58. Thompson LF. Ecto-5’-Nucleotidase can Provide the Total Purine Requirements of Mitogen-Stimulated Human T Cells and Rapidly Dividing Human B Lymphoblastoid Cells. J Immunol (1985) 134(6):3794–7.
59. Airas L, Niemelä J, Salmi M, Puurunen T, Smith DJ, Jalkanen S. Differential Regulation and Function of CD73, a Glycosyl- Phosphatidylinositol-Linked 70-kD Adhesion Molecule, on Lymphocytes and Endothelial Cells. J Cell Biol (1997) 136(2):421–31. doi: 10.1083/jcb.136.2.421
60. Jeffrey JL, Lawson KV, Powers JP. Targeting Metabolism of Extracellular Nucleotides via Inhibition of Ectonucleotidases CD73 and CD39. J Med Chem (2020) 63(22):13444–65. doi: 10.1021/acs.jmedchem.0c01044
61. Conter LJ, Song E, Shlomchik MJ, Tomayko MM. CD73 Expression Is Dynamically Regulated in the Germinal Center and Bone Marrow Plasma Cells Are Diminished in Its Absence. PloS One (2014) 9(3):e92009. doi: 10.1371/journal.pone.0092009
62. Monteiro I, Vigano S, Faouzi M, Treilleux I, Michielin O, Ménétrier-Caux C, et al. CD73 Expression and Clinical Significance in Human Metastatic Melanoma. Oncotarget (2018) 9(42):26659–69. doi: 10.18632/oncotarget.25426
63. Airas L, Hellman J, Salmi M, Bono P, Puurunen T, Smith DJ, et al. CD73 Is Involved in Lymphocyte Binding to the Endothelium: Characterization of Lymphocyte-Vascular Adhesion Protein 2 Identifies It as CD73. J Exp Med (1995) 182(5):1603–8. doi: 10.1084/jem.182.5.1603
64. Resta R, Yamashita Y, Thompson LF. Ecto-Enzyme and Signaling Functions of Lymphocyte CD73. Immunol Rev (1998) 161(1):95–109. doi: 10.1111/j.1600-065X.1998.tb01574.x
65. Stochaj U, Mannherz HG. Chicken Gizzard 5’-Nucleotidase Functions as a Binding Protein for the Laminin/Nidogen Complex. Eur J Cell Biol (1992) 59(2):364–72.
66. Dieckhoff J, Mollenhauer J, Kuhl U, Niggemeyer B, Von Der Mark K, Mannherz H-G. The Extracellular Matrix Proteins Laminin and Fibronectin Modify the AMPase Activity of S-Nucleotidase From Chicken Gizzard Smooth Muscle. FEBS J (1986) 195(1–2):82–6. doi: 10.1016/0014-5793(86)80135-1
67. Stochaj U, Dieckhoff J, Mollenhauer J, Cramer M, Mannherz HG. Evidence for the Direct Interaction of Chicken Gizzard 5′-Nucleotidase With Laminin and Fibronectin. Biochem Biophys Acta (1989) 992(3):385–92. doi: 10.1016/0304-4165(89)90101-3
68. Stochaj U, Richter H, Mannherz HG. Chicken Gizzard 5’-Nucleotidase Is a Receptor for the Extracellular Matrix Component Fibronectin. Eur J Cell Biol (1990) 51(2):335–8.
69. Sadej R, Inai K, Rajfur Z, Ostapkowicz A, Kohler J, Skladanowski AC, et al. Tenascin C Interacts With Ecto-5′-Nucleotidase (eN) and Regulates Adenosine Generation in Cancer Cells. Biochim Biophys Acta - Mol Basis Dis (2008) 1782(1):35–40. doi: 10.1016/j.bbadis.2007.11.001
70. Airas L, Niemelä J, Jalkanen S. CD73 Engagement Promotes Lymphocyte Binding to Endothelial Cells Via a Lymphocyte Function-Associated Antigen-1-Dependent Mechanism. J Immunol (2000) 165(10):5411–7. doi: 10.4049/jimmunol.165.10.5411
71. Airas L, Salmi M, Jalkanen S. Lymphocyte-Vascular Adhesion Protein-2 Is a Novel 70-kDa Molecule Involved in Lymphocyte Adhesion to Vascular Endothelium. J Immunol (1993) 151:4228–38.
72. States DJ, Walseth TF, Lee HC. Similarities in Amino Acid Sequences of Aplysia ADP-Ribosyl Cyclase and Human Lymphocyte Antigen CD38. Trends Biochem Sci (1992) 17(12):495. doi: 10.1016/0968-0004(92)90337-9
73. Konen JM, Fradette JJ, Gibbons DL. The Good, the Bad and the Unknown of CD38 in the Metabolic Microenvironment and Immune Cell Functionality of Solid Tumors. Cells (2019) 9:52. doi: 10.3390/cells9010052
74. Howard M, Grimaldi JC, Bazan JF, Lund FE, Santos-Argumedo L, Parkhouse RME, et al. Formation and Hydrolysis of Cyclic ADP-Ribose Catalyzed by Lymphocyte Antigen CD38. Sci (80- ) (1993) 262(5136):1056–9. doi: 10.1126/science.8235624
75. Partida-Sánchez S, Cockayne DA, Monard S, Jacobson EL, Oppenheimer N, Garvy B, et al. Cyclic ADP-Ribose Production by CD38 Regulates Intracellular Calcium Release, Extracellular Calcium Influx and Chemotaxis in Neutrophils and Is Required for Bacterial Clearance In Vivo. Nat Med (2001) 7(11):1209–16. doi: 10.1038/nm1101-1209
76. Cheung Lee H, Walsethq TF, Bratt GT, Hayes RN, Clapper DL. Structural Determination of a Cyclic Metabolite of NAD+ With Intracellular Ca2+-Mobilizing Activity. J Biol Chem (1989) 264(3):1608–15. doi: 10.1016/S0021-9258(18)94230-4
77. Glaría E, Valledor AF. Roles of CD38 in the Immune Response to Infection. Cells (2020) 9(1):228. doi: 10.3390/cells9010228
78. Lee HC. Structure and Enzymatic Functions of Human CD38. Mol Med (2006) 12(11–12):317–23. doi: 10.2119/2006-00086.Lee
79. Liu Q, Kriksunov IA, Graeff R, Munshi C, Hon CL, Hao Q. Crystal Structure of Human CD38 Extracellular Domain. Structure (2005) 13(9):1331–9. doi: 10.1016/j.str.2005.05.012
80. Tenca C, Merlo A, Zarcone D, Saverino D, Bruno S, De Santanna A, et al. Death of T Cell Precursors in the Human Thymus: A Role for CD38. Int Immunol (2003) 15:1105–16. doi: 10.1093/intimm/dxg111
81. Funaro A, Spagnoli GC, Ausiello CM, Alessio M, Roggero S, Delia D, et al. Involvement of the Multilineage CD38 Molecule in a Unique Pathway of Cell Activation and Proliferation. J Immunol (1990) 145(8):2390–6.
82. Deterre P, Gelman L, Gary-Gouy H, Arrieumerlou C, Berthelier V, Tixier J-M, et al. Coordinated Regulation in Human T Cells of Nucleotide-Hydrolyzing Ecto-Enzymatic Activities, Including CD38 and PC-1 Possible Role in the Recycling of Nicotinamide Adenine Dinucleotide Metabolites. J Immunol (1996) 157:1381–2.
83. Oliver AM, Martin F, Kearney JF. Mouse CD38 Is Down-Regulated on Germinal Center B Cells and Mature Plasma Cells. J Immunol (1997) 158(3):1108–15.
84. Frasca L, Fedele G, Deaglio S, Capuano C, Palazzo R, Vaisitti T, et al. CD38 Orchestrates Migration, Survival, and Th1 Immune Response of Human Mature Dendritic Cells. Blood (2006) 107(6):2392–9. doi: 10.1182/blood-2005-07-2913
85. Malavasi F, Deaglio S, Funaro A, Ferrero E, Horenstein AL, Ortolan E, et al. Evolution and Function of the ADP Ribosyl Cyclase/CD38 Gene Family in Physiology and Pathology. Physiol Rev (2008) 88(3):841–86. doi: 10.1152/physrev.00035.2007
86. Deaglio S, Zubiaur M, Gregorini A, Bottarel F, Ausiello CM, Dianzani U, et al. Human CD38 and CD16 Are Functionally Dependent and Physically Associated in Natural Killer Cells. Blood (2002) 99(7):2490–8. doi: 10.1182/blood.V99.7.2490
87. Funaro A, De Monte LB, Dianzani U, Forni M, Malavasi F. Human CD38 Is Associated to Distinct Molecules Which Mediate Transmembrane Signaling in Different Lineages. Eur J Immunol (1993) 23(10):2407–11. doi: 10.1002/eji.1830231005
88. Ausiello CM, La Sala A, Ramoni C, Urbani F, Funaro A, Malavasi F. Secretion of IFN-γ, IL-6, Granulocyte-Macrophage Colony-Stimulating Factor and IL-10 Cytokines After Activation of Human Purified T Lymphocytes Upon CD38 Ligation. Cell Immunol (1996) 173(2):192–7. doi: 10.1006/cimm.1996.0267
89. Zupo S, Rugari E, Dono M, Taborelli G, Malavasi F, Ferrarini M. CD38 Signaling by Agonistic Monoclonal Antibody Prevents Apoptosis of Human Germinal Center B Cells. Eur J Immunol (1994) 24(5):1218–22. doi: 10.1002/eji.1830240532
90. Silvennoinen O, Nishigaki H, Kitanaka A, Kumagai M, Ito C, Malavasi F, et al. CD38 Signal Transduction in Human B Cell Precursors. Rapid Induction of Tyrosine Phosphorylation, Activation of Syk Tyrosine Kinase, and Phosphorylation of Phospholipase C-Gamma and Phosphatidylinositol 3-Kinase. J Immunol (1996) 156(1):100–7.
91. Dianzani U, Funaro A, DiFranco D, Garbarino M, Bragardo M, Redoglia V, et al. Interaction Between Endothelium and CD4+CD45RA+ Lymphocytes. Role of the Human CD38 Molecule. J Immunol (1994) 153(3):952–9.
92. Deaglio S, Morra M, Mallone R, Ausiello CM, Prager E, Garbarino G, et al. Human CD38 (ADP-Ribosyl Cyclase) Is a Counter-Receptor of CD31, an Ig Superfamily Member. J Immunol (1998) 160:395–402.
93. Trubiani O, Guarnieri S, Orciani M, Salvolini E, Di Primio R. Sphingolipid Microdomains Mediate CD38 Internalization: Topography of the Endocytosis. Int J Immunopathol Pharmacol (2004) 17(3):293–300. doi: 10.1177/039463200401700309
94. Partida-Sánchez S, Goodrich S, Kusser K, Oppenheimer N, Randall TD, Lund FE. Regulation of Dendritic Cell Trafficking by the ADP-Ribosyl Cyclase CD38: Impact on the Development of Humoral Immunity. Immunity (2004) 20(3):279–91. doi: 10.1016/S1074-7613(04)00048-2
95. Kontani K, Kukimoto I, Nishina H, Hoshino SI, Hazeki O, Kanaho Y, et al. Tyrosine Phosphorylation of the C-Cbl Proto-Oncogene Product Mediated by Cell Surface Antigen CD38 in HL-60 Cells. J Biol Chem (1996) 271(3):1534–7. doi: 10.1074/jbc.271.3.1534
96. Matsuo T, Hazeki K, Tsujimoto N, Inoue S, Kurosu H, Kontani K, et al. Association of Phosphatidylinositol 3-Kinase With the Proto-Oncogene Product Cbl Upon CD38 Ligation by a Specific Monoclonal Antibody in THP-1 Cells. FEBS Lett (1996) 397(1):113–6. doi: 10.1016/S0014-5793(96)01151-9
97. Wong SW, Comenzo RL. CD38 Monoclonal Antibody Therapies for Multiple Myeloma. Clin Lymphoma Myeloma Leuk (2015) 15(11):635–45. doi: 10.1016/j.clml.2015.07.642
98. Martin TG, Corzo K, Chiron M, van de Velde H, Abbadessa G, Campana F, et al. Therapeutic Opportunities With Pharmacological Inhibition of CD38 With Isatuximab. Cells (2019) 8;1522. doi: 10.3390/cells8121522
99. Morandi F, Airoldi I, Marimpietri D, Bracci C, Faini AC, Gramignoli R. CD38, a Receptor With Multifunctional Activities: From Modulatory Functions on Regulatory Cell Subsets and Extracellular Vesicles, to a Target for Therapeutic Strategies. Cells (2019) 8(12):1527. doi: 10.3390/cells8121527
100. Yakymiv Y, Augeri S, Fissolo G, Peola S, Bracci C, Binaschi M, et al. CD157: From Myeloid Cell Differentiation Marker to Therapeutic Target in Acute Myeloid Leukemia. Cells (2019) 8(12):1580. doi: 10.3390/cells8121580
101. Ishihara K, Hirano T. BST-1/CD157 Regulates the Humoral Immune Responses In Vivo. Chem Immunol (2000) 75:235–55. doi: 10.1159/000058772
102. Funaro A, Ortolan E, Ferranti B, Gargiulo L, Notaro R, Luzzatto L, et al. CD157 Is an Important Mediator of Neutrophil Adhesion and Migration. Blood (2004) 104(13):4269–78. doi: 10.1182/blood-2004-06-2129
103. Lo Buono N, Parrotta R, Morone S, Bovino P, Nacci G, Ortolan E, et al. The CD157-Integrin Partnership Controls Transendothelial Migration and Adhesion of Human Monocytes. J Biol Chem (2011) 286(21):18681–91. doi: 10.1074/jbc.M111.227876
104. Ortolan E, Tibaldi EV, Ferranti B, Lavagno L, Garbarino G, Notaro R, et al. CD157 Plays a Pivotal Role in Neutrophil Transendothelial Migration. Blood (2006) 108(13):4214–22. doi: 10.1182/blood-2006-04-017160
105. Lavagno L, Ferrero E, Ortolan E, Malvasi F, Funaro A. CD157 Is Part of a Supramolecular Complex With CD11b/CD18 on the Human Neutrophil Cell Surface. J Biol Regul Homeost Agents (2007) 21(1):5–11.
106. Zen K, Utech M, Liu Y, Soto I, Nusrat A, Parkos CA. Association of BAP31 With CD11b/CD18. Potential Role in Intracellular Trafficking of CD11b/CD18 in Neutrophils. J Biol Chem (2004) 279(43):44924–30. doi: 10.1074/jbc.M402115200
107. Morone S, Augeri S, Cuccioloni M, Mozzicafreddo M, Angeletti M, Lo Buono N, et al. Binding of CD157 Protein to Fibronectin Regulates Cell Adhesion and Spreading. J Biol Chem (2014) 289(22):15588–601. doi: 10.1074/jbc.M113.535070
108. Giebeler N, Zigrino P. A Disintegrin and Metalloprotease (ADAM): Historical Overview of Their Functions. Toxins (2016) 8:122. doi: 10.3390/toxins8040122
109. Yoshiyama K, Higuchi Y, Kataoka M, Matsuura K, Yamamoto S. CD156 (Human ADAM8): Expression, Primary Amino Acid Sequence and Gene Location. Genomics (1997) 41:56–62. doi: 10.1006/geno.1997.4607
110. Conrad C, Benzel J, Dorzweiler K, Cook L, Schlomann U, Zarbock A, et al. ADAM8 in Invasive Cancers: Links to Tumor Progression, Metastasis, and Chemoresistance. Clin Sci (2019) 133(1):83–99. doi: 10.1042/CS20180906
111. Gould RJ, Polokoff MA, Friedman PA, Huang TF, Holt JC, Cook JJ, et al. Disintegrins: A Family of Integrin Inhibitory Proteins From Viper Venoms. Proc Soc Exp Biol Med (1990) 195(2):168–71. doi: 10.3181/00379727-195-43129B
112. Schlomann U, Rathke-Hartlieb S, Yamamoto S, Jockusch H, Bartsch JW. Tumor Necrosis Factor α Induces a Metalloprotease-Disintegrin, ADAM8 (CD 156): Implications for Neuron-Glia Interactions During Neurodegeneration. J Neurosci (2000) 20(21):7964–71. doi: 10.1523/JNEUROSCI.20-21-07964.2000
113. King NE, Zimmermann N, Pope SM, Fulkerson PC, Nikolaidis NM, Mishra A, et al. Expression and Regulation of a Disintegrin and Metalloproteinase (ADAM) 8 in Experimental Asthma. Am J Respir Cell Mol Biol (2004) 31(3):257–65. doi: 10.1165/rcmb.2004-0026OC
114. Koller G, Schlomann U, Golfi P, Ferdous T, Naus S, Bartsch J. ADAM8/MS2/CD156, an Emerging Drug Target in the Treatment of Inflammatory and Invasive Pathologies. Curr Pharm Des (2009) 15(20):2272–81. doi: 10.2174/138161209788682361
115. Dreymueller D, Pruessmeyer J, Schumacher J, Fellendorf S, Hess FM, Seifert A, et al. The Metalloproteinase ADAM8 Promotes Leukocyte Recruitment In Vitro and in Acute Lung Inflammation. Am J Physiol Cell Mol Physiol (2017) 313(3):L602–14. doi: 10.1152/ajplung.00444.2016
116. Choi SJ, Han JH, Roodman GD. ADAM8: A Novel Osteoclast Stimulating Factor. J Bone Miner Res (2001) 16(5):814–22. doi: 10.1359/jbmr.2001.16.5.814
117. Schlomann U, Wildeboer D, Webster A, Antropova O, Zeuschner D, Knight CG, et al. The Metalloprotease Disintegrin ADAM8. Processing by Autocatalysis Is Required for Proteolytic Activity and Cell Adhesion. J Biol Chem (2002) 277(50):48210–9. doi: 10.1074/jbc.M203355200
118. Kleino I, Järviluoma A, Hepojoki J, Huovila AP, Saksela K. Preferred SH3 Domain Partners of ADAM Metalloproteases Include Shared and ADAM-Specific SH3 Interactions. PloS One (2015) 10(3):e0121301. doi: 10.1371/journal.pone.0121301
119. Dong F, Eibach M, Bartsch JW, Dolga AM, Schlomann U, Conrad C, et al. The Metalloprotease-Disintegrin ADAM8 Contributes to Temozolomide Chemoresistance and Enhanced Invasiveness of Human Glioblastoma Cells. Neuro Oncol (2015) 17(11):1474–85. doi: 10.1093/neuonc/nov042
120. Peola S, Borrione P, Matera L, Malavasi F, Pileri A, Massaia M. Selective Induction of CD73 Expression in Human Lymphocytes by CD38 Ligation: A Novel Pathway Linking Signal Transducers With Ecto-Enzyme Activities. J Immunol (1996) 157:4354–62.
121. Allard B, Longhi MS, Robson SC, Stagg J. The Ectonucleotidases CD39 and CD73: Novel Checkpoint Inhibitor Targets. Immunol Rev (2017) 276(1):121–44. doi: 10.1111/imr.12528
122. Horenstein AL, Morandi F, Bracci C, Pistoia V, Malavasi F. Functional Insights Into Nucleotide-Metabolizing Ectoenzymes Expressed by Bone Marrow-Resident Cells in Patients With Multiple Myeloma. Immunol Lett (2019) 205:40–50. doi: 10.1016/j.imlet.2018.11.007
123. Seals DF, Courtneidge SA. The ADAMs Family of Metalloproteases: Multidomain Proteins With Multiple Functions. Genes Dev (2003) 17:7–30. doi: 10.1101/gad.1039703
124. Eto K, Puzon-McLaughlin W, Sheppard D, Sehara-Fujisawa A, Zhang XP, Takada Y. RGD-Independent Binding of Integrin α9β1 to the ADAM-12 and -15 Disintegrin Domains Mediates Cell-Cell Interaction. J Biol Chem (2000) 275(45):34922–30. doi: 10.1074/jbc.M001953200
125. Zhang XP, Kamata T, Yokoyama K, Puzon-McLaughlin W, Takada Y. Specific Interaction of the Recombinant Disintegrin-Like Domain of MDC- 15 (Metargidin, ADAM-15) With Integrin αvβ3. J Biol Chem (1998) 273(13):7345–50. doi: 10.1074/jbc.273.13.7345
126. Nath D, Slocombe PM, Webster A, Stephens PE, Docherty AJ, Murphy G. Meltrin Gamma(ADAM-9) Mediates Cellular Adhesion Through Alpha(6)Beta(1)Integrin, Leading to a Marked Induction of Fibroblast Cell Motility. J Cell Sci (2000) 113(12):2319–28. doi: 10.1242/jcs.113.12.2319
127. Cal S, Freije JMP, López JM, Takada Y, López-Otín C. ADAM 23/MDC3, a Human Disintegrin That Promotes Cell Adhesion via Interaction With the αvβ3 Integrin Through an RGD-Independent Mechanism. Mol Biol Cell (2000) 11(4):1457–69. doi: 10.1091/mbc.11.4.1457
128. Garay-Canales CA, Díaz-Alvarez L, Lopez-Cortes GI. Novel Immunotherapy Strategies Involving Matrix Metalloproteinase (MMP) Family. In: Morales-Montor J, Segovia-Mendoza M, editors. Immunotherapy in Resistant Cancer: From the Lab Bench Work to Its Clinical Perspectives. London, UK: Academic Press Inc (2020). p. 227–47.
Keywords: moonlighting protein, ectoenzyme, cell adhesion, signal transduction, leukocytes
Citation: López-Cortés GI, Díaz-Alvarez L and Ortega E (2021) Leukocyte Membrane Enzymes Play the Cell Adhesion Game. Front. Immunol. 12:742292. doi: 10.3389/fimmu.2021.742292
Received: 19 July 2021; Accepted: 29 October 2021;
Published: 23 November 2021.
Edited by:
Liwu Li, Virginia Tech, United StatesReviewed by:
Valentin P. Yakubenko, East Tennessee State University, United StatesRuoxi Yuan, Hospital for Special Surgery, United States
Copyright © 2021 López-Cortés, Díaz-Alvarez and Ortega. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Enrique Ortega, b3J0c290b0BpaWJpb21lZGljYXMudW5hbS5teA==