 Guillaume Larid1,2
Guillaume Larid1,2 Adriana Delwail3
Adriana Delwail3 Thomas Dalle2Philippe Vasseur2,4Christine Silvain2,4
Thomas Dalle2Philippe Vasseur2,4Christine Silvain2,4 Jean-François Jégou2
Jean-François Jégou2 Franck Morel2Jean-Claude Lecron2,5Elisabeth Gervais1,2*†
Franck Morel2Jean-Claude Lecron2,5Elisabeth Gervais1,2*†- 1Rheumatology Department, CHU de Poitiers, Poitiers, France
- 2University of Poitiers, LITEC, Poitiers, France
- 3UMR 6041, 4CS, Université de Poitiers, Poitiers, France
- 4Gastroenterology Department, CHU de Poitiers, Poitiers, France
- 5Immunology-Inflammation Laboratory, CHU de Poitiers, Poitiers, France
Objectives: Psoriatic arthritis (PsA) and cutaneous psoriasis (PsO) are different phenotypes of psoriatic disease (PsD), whose underlying specific mechanisms remain incompletely understood. As cytokines are key elements to induce and tune up immune responses to drive inflammatory diseases, our objective was to assess whether clinical features, disease phenotype and PsA and PsO activity were associated with a particular ex vivo cytokine production profile.
Methods: Forty-eight patients (37 PsA and 11 PsO) and 11 healthy subjects (HS) were studied. Cytokine production by peripheral blood mononuclear cells (PBMC) that were either unstimulated, or stimulated with LPS or anti-CD3/CD28 antibodies, were analysed by multiplex assay in the culture supernatants.
Results: Cytokine signature of PsD includes a high level of TNFα in supernatants of LPS-stimulated PBMC, higher levels of IL-6 and lower levels of IFN-γ and IL-17A after CD3-CD28 stimulation, as well as higher spontaneous IL-1RA and TNFα production compared to HS. High body mass index (BMI) was associated with lower levels of IL-1β, and metabolic syndrome with lower levels of IFN-γ after LPS stimulation. In PsD, dermatological activity was related with higher IL-17A level, while rheumatic activity was linked with lower levels of IFN-γ and TNFα. Comparing each PsD subtype to HS, IL-1β and IL-6 productions are higher when using LPS stimulation in PsO patients with higher levels of IL-1β and IL-1α in peripheral PsA patients after CD3/CD28 stimulation. LPS stimulation induced high levels of IL-17A in peripheral PsA compared to axial PsA. PsA patients with axial PsA share some features with PsO but shows a distinct cytokine pattern compared to peripheral PsA.
Conclusion: PsO and the different PsA subtypes exhibit distinct ex vivo cytokine production profiles and common features of the so-called PsD. Analysis of IL-1 cytokine family and IL-6 seems to be of particular interest to distinguish PsO and peripheral PsA since it depends on monocytes in PsO and T-lymphocytes in peripheral PsA. Peripheral cytokine profiles are influenced by rheumatic and dermatological activity of the disease, and also by metabolic syndrome features. Our results highlight the crucial role of immune cell interactions with different patterns of interaction depending on clinical phenotype.
1 Introduction
Psoriasis is an inflammatory cutaneous disorder with prevalence up to 3% of people in industrialized countries (1). Originally described as a skin and/or articular pathology, psoriasis appears as a disease involving other tissue damage. In the last ten years, the concept of “psoriatic disease” (PsD) has emerged, which encompasses all the clinical aspects of so-called cutaneous psoriasis (PsO), psoriatic arthritis (PsA), and associated comorbidities such as cardio-metabolic disorders (2–5). As PsD is a systemic disease, patients can present features of PsO, PsA, or both. Numerous studies focusing on PsO patients do not indicate if patients have articular involvement or not. Likewise, among PsO patients, many patients suffering from PsA are undiagnosed (6). This common vagueness in population’s description creates a potential bias both for the clinical diagnosis which determines the follow-up and treatment of patients, and for studies describing the physiopathological mechanisms of PsD, in which the two clinical entities may not be clearly differentiated from one to another.
PsD is a polygenic inflammatory disorder of which the pathophysiology raises numerous questions (7, 8). Interestingly, among the multiple polymorphisms associated with PsO or PsA, a significant number involves genes encoding for cytokines such as IL-12B, IL-23A, IL-23R, IL-13, IL-36G, IL-20, TNFAIP3 or TNFIP1 (7–9), leading to investigations of cytokine profiles in PsO or PsA in order to determine specific cytokine signatures. To date, different biological approaches have been applied to PsD, including serum analysis, transcriptomic analysis, and ex vivo studies, but the majority of these studies do not discriminate between the two clinical entities. While transcriptomic approaches from inflamed skin or synovial tissue samples provide relevant information on the pathophysiology of both diseases, their use in routine practice is not possible for diagnosis, insofar it requires invasive techniques to obtain samples. Therefore, most studies have focused on cytokine assays from patient blood samples, even though it is known that circulating cytokines are the tip of the iceberg for tissue inflammatory diseases. The conflicting data between studies and/or between subjects have confirmed the fact that measurement of serum cytokines such as TNF-α, IL-1α, IL-1β, IL-17, IL-23, IL-18 or IL-33 is not a reliable and reproducible method to determine cytokine status in PsD (10–15).
An alternative approach would consist in analysing the cytokine secretion capacities of circulating immune cells so as to segregate between PsA and PsO. Studies investigating cytokine secretion profiles from ex vivo-stimulated PBMC have been reported in PsO and PsA patients. Nishibu et al. studied spontaneous secretions of a limited number of cytokines (IL-1β, IL-2, IL-6, IL-8 and IFN-γ) and reported higher secretions of IL-1β, IL-6, and IL-8 by peripheral blood mononuclear cells (PBMC) isolated from PsA patients compared to PsO patients and from PsO patients compared to healthy subjects (16). However, this cytokine panel did not include relevant cytokines such as TNF-α or IL-17A. In a second study by Bosè et al., the authors also used a limited panel of T cell cytokines composed of IL-2, IFN-γ, TNF-α, IL-4, IL-5, IL-10, and IL-17 and showed that higher IFN-γ and IL-10 levels were produced by activated PBMC from PsO compared to healthy subjects (17). Finally, cytokine secretion was induced after stimulation using an anti-CD3 monoclonal antibody (mAb). This approach does not take into account non-T cell populations as potential sources of cytokines, and hides possible interactions between activated non-T cells and T cells.
The objective of the present study was to assess the cytokine profiles of PsA and PsO patients by different ex vivo PBMC stimulations in order to highlight potential differences between these two distinct pathological entities. Correlation with the clinical variables of metabolic syndrome (MetS), dermatological activity, and rheumatic activity will likewise be explored, as will the differences before and after DMARD (disease modifying anti-rheumatic drugs) treatment initiation in PsA patients.
2 Materials and methods
2.1 Patients
Patients with PsA were included if they were 18-years-old or older and fulfilled the Classification Criteria for Psoriatic Arthritis (CASPAR) of PsA (18). Patients of 18-years-old or older with PsO diagnosed by dermatologist without any joint complaint were included. Healthy controls were included if they did not suffer from any rheumatic, dermatological or inflammatory condition.
Patients were followed in outpatient clinics and included at any time point of the disease.
Characteristics of patients including dermatological activity, rheumatic activity, CRP sera levels, BMI, HOMA score, PASI, psoriasis duration, age, and abdominal circumference were obtained in the patients’ medical records. Detailed clinical characteristics were available for 31 out of the 37 PsA patients, who were classified as having axial disease, peripheral disease, or both patterns (mixed).
Dermatological activity (i.e. moderate to severe disease) was defined as PASI > 10 (19) and rheumatic activity as DAS28 ≥ 2.6 for peripheral PsA (20), BASDAI ≥ 40/100 for axial PsA (21), and either one of those two criteria for mixed PsA. Overall clinical activity was defined as an active disease that was either dermatological, or rheumatic, or both.
Focusing on metabolic syndrome (MetS) features, high abdominal circumference was defined as above 88 cm for women and 102 cm for men. HOMA score was considered elevated when superior to 3.
High CRP was defined as CRP ≥ 3 mg/L, and MetS was defined according to the ATP III definition (22).
We separately analysed 6 patients for whom we had samples taken before and after DMARD initiation, either csDMARD (methotrexate for 1) or bDMARD (TNF-α inhibitors for 5 of them).
2.2 Samples and cell culture
Ten millilitres of blood were collected from patients and healthy controls in Vacutainer tubes containing heparin for the isolation of PBMCs. Human PBMCs were isolated by density gradient centrifugation using Ficoll-Paque (GE Healthcare) and counted. Analysis of cell viability has been performed using Trypan Blue 0.4% (Merck-Sigma). All samples displayed at least 95% of cell viability. Isolated PBMCs were cultured in RPMI 1640 containing 2 mM glutamine, 100 U/ml penicillin-streptomycin (Invitrogen Life Technologies), and 10% heat-inactivated foetal calf serum (Gibco Life Technologies). Cells were seeded at 1 × 106 cells/ml in 24-well plates and incubated 24 h at 37°C in a humidified atmosphere containing 5% CO2. Cultures were either left unstimulated to reflect the intrinsic state of cell activation or stimulated throughout the 24 h culture period with E. coli LPS (1 µg/ml; Sigma L2654 – Escherichia Coli serotype 026:B6) for monocyte activation or anti-CD3/CD28 mAb coated-beads for T cell activation (2.5 × 105 beads/106 cells/ml; Invitrogen Life Technologies) (23, 24). After incubation, the plates were centrifuged and supernatants were stored at -80°C until analysis. To estimate lymphocyte activity, the secretion of pro- and anti-inflammatory cytokines has been measured with a set of cytokines representative of Th subsets. Even if they were not exclusively secreted by these Th subsets, IFN-γ, IL-4, IL-17, and IL-10 were representative of Th1 cell, Th2 cell, Th17 cell, and Treg cell, respectively (25).
2.3 Cytokine measurements
The concentrations of IL-1α, IL-1β, IL-1RA, IL-4, IL-17, IL-6, TNF-α, IL-10, and γ interferon (IFN-γ) were quantified in cell culture supernatants using the MILLIPLEX MAP Human Cytokine/Chemokine magnetic bead panel kit (Millipore Corporation, Billerica, MA) according to the manufacturer’s instructions. Data were obtained with the Luminex 200™ platform (Luminex Xmap Technology) and analysed with the xPONENT™ software. All samples were assayed in duplicates.
2.4 Ethics
All patients gave informed consent. The study was conducted in accordance with the declaration of HELSINKI and was approved by the Poitiers University Hospital research board and local ethics committee.
2.5 Statistical analysis
All statistical analyses were performed using GraphPad Prism 9 (GraphPad Software, Inc.). Quantitative data were described by means and standard deviation or median. Qualitative data were presented as absolute frequencies and percentages.
The nonparametric Mann-Whitney U test was used to evaluate the difference of quantitative variables between two groups. For comparison of quantitative variables between three groups, Kruskal-Wallis test with post-hoc Dunn’s multiple comparison tests were performed. Paired observations were compared with the Wilcoxon matched-pairs test. Chi-square tests or Fisher’s exact test were conducted to compare qualitative variables between groups. All box plots showed medians with the minimal and maximal values. P values less than 0.05 were considered statistically significant.
3 Results
3.1 Description of the patient population
Thirty-seven PsA patients and 11 patients with PsO were included in the study, as were 11 healthy subjects. The population had a mean age of 53 years and 62.5% were women. MetS was present in 33.3% of PsD patients, with a mean BMI of 28 kg/m², a mean HOMA score of 3.74, and a mean abdominal circumference of 96.4 cm for women and 101.5 cm for men. In PsA patients, 37.8% had a purely peripheral form of the disease, 16.2% a purely axial disease, and 46% a mixed pattern. In PsO and PsA respectively, 63.6% and 67.6% of patients received csDMARDs, while 54.5% and 40.5% received bDMARDs. The mean PASI score was higher in PsO (7.42) than in PsA patients (3.69) (p = 0.030), with 27.2% and 12.9% respectively having a dermatologically active disease. There were no other significant differences between PsO, PsA and PsD groups (Table 1).
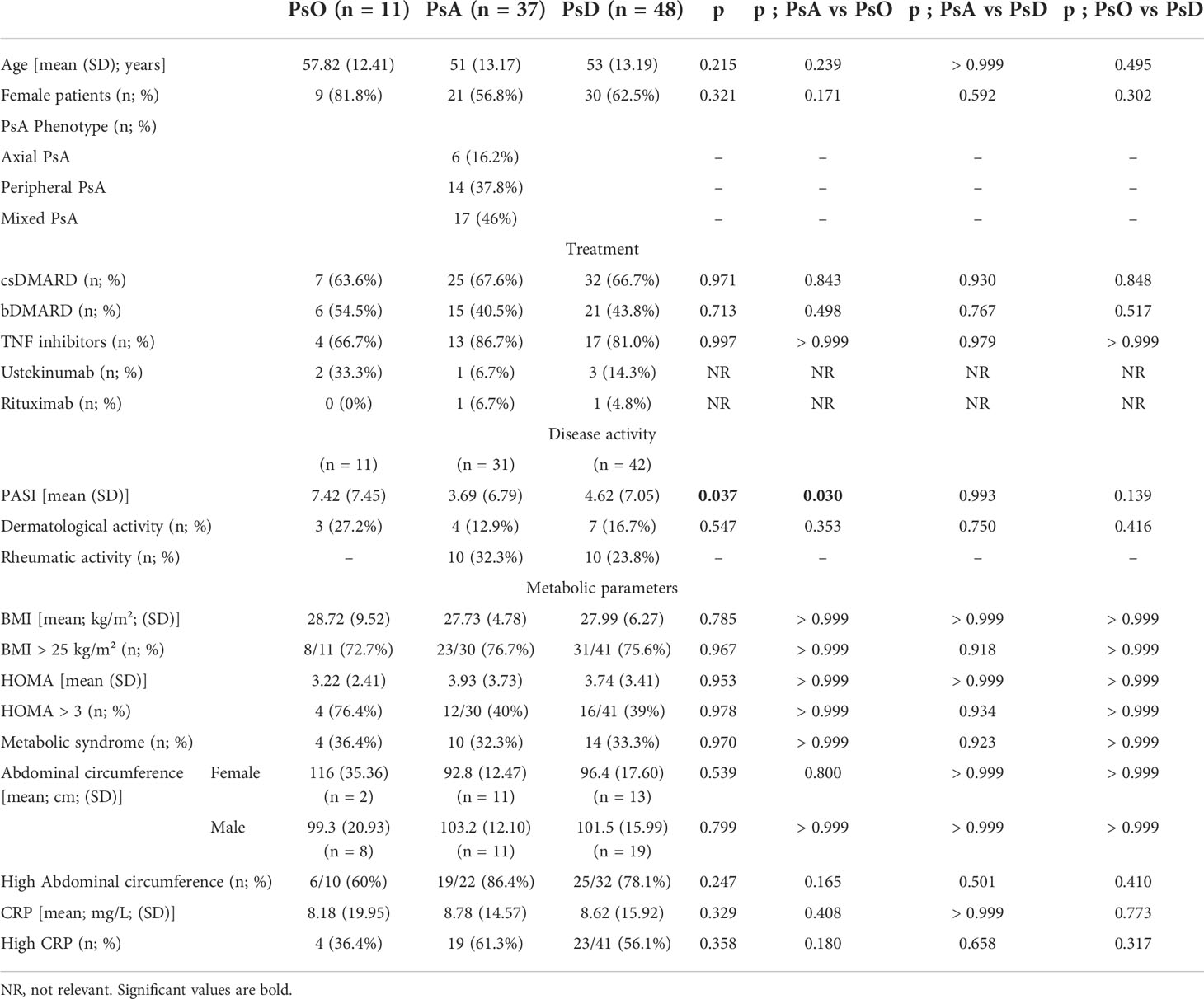
Table 1 Description of the population.
3.2 Cytokine profiles of psoriatic disease
Since PsD is a systemic disorder, analysis of all patients taken together was first performed before deeper analysis depending on clinical phenotype.
3.2.1 Comparison of PsD patients with healthy subjects
In comparison with healthy subjects, unstimulated PBMC of PsD patients exhibited a monocyte-predominant pro-inflammatory pattern with higher spontaneous release of IL-6 and TNF-α and higher release of IL-1RA. Stimulation of the monocytes by LPS induced higher levels of IL-1β and higher secondary activation of Th1 (IFN-γ), Th2 (IL-4), and Th17 (IL-17A) cytokines. In addition, the activation of T lymphocytes by anti CD3/CD28 mAb indirectly stimulated monocyte production of IL-1β at a higher level than in healthy controls. IL-6 higher production could result from either directly stimulated T-cells or indirectly activated monocytes. Surprisingly, these stimulated lymphocytes displayed lower levels of IFN-γ and IL-17A than healthy controls (Figure 1 and Supplementary Table 1).
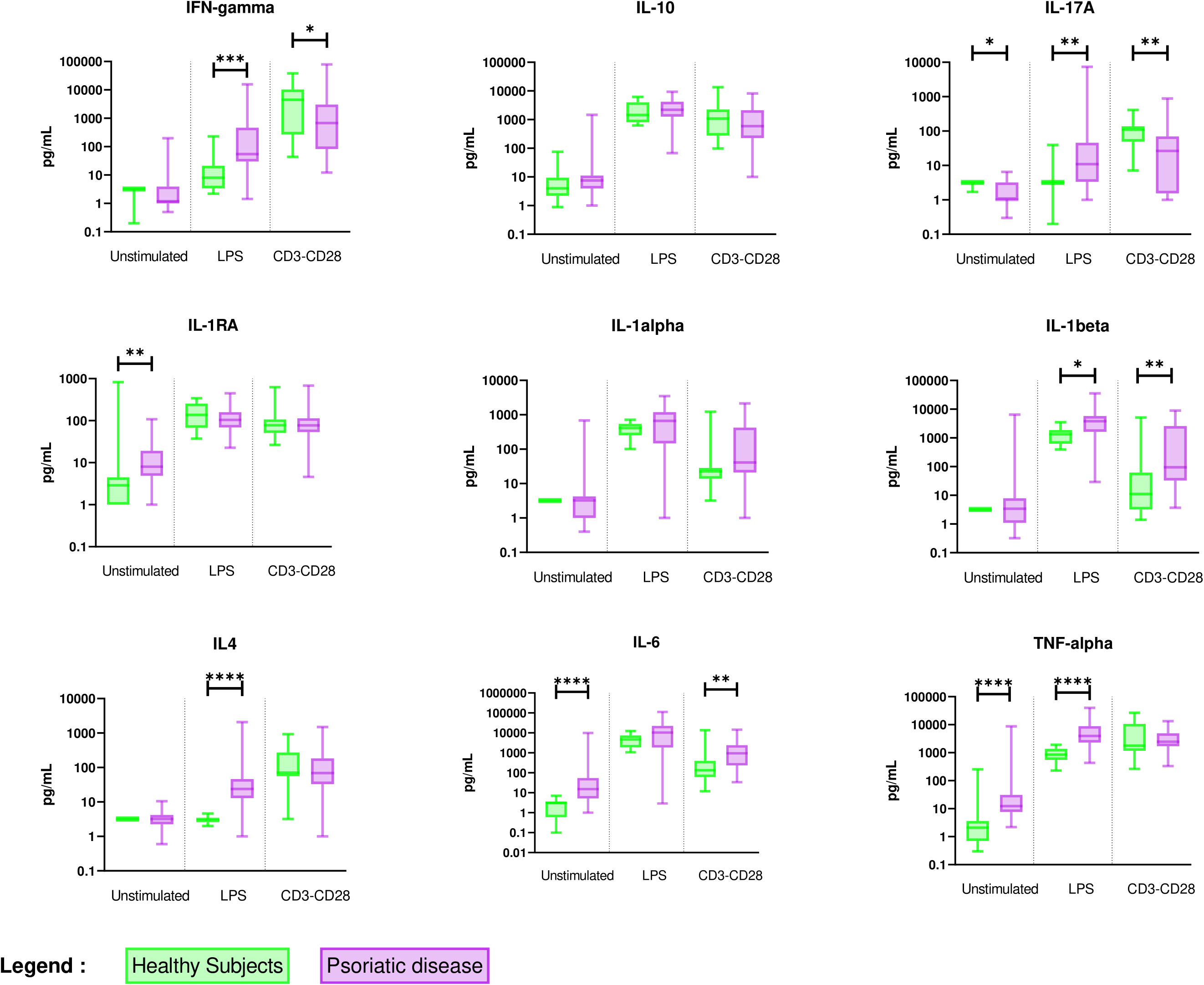
Figure 1 Overall comparison of ex vivo cytokine production in PBMC supernatants between healthy subjects (n = 11) and PsD patients (n = 48). PBMCs from healthy subjects (n = 11) and PsD patients (n= 48) were cultured at 1x106 cells/ml for 24 h at 37°C, 5% CO2 with and without stimulation of monocytes by E. coli LPS (1 µg/mL) or T lymphocytes by anti-CD3/CD28 mAb (2.5 × 104 beads/106 cells/ml). Supernatants were collected and cytokine levels were assayed by Luminex Multiplex ELISA. Results are presented using median values with minimum to maximum box-plot. Difference between the groups using non-parametric Mann-Whitney U test is presented in the top of the figure for each condition. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001.
Those results suggested a pronounced inflammatory state of monocytes able to secondarily activate different T cell subtypes in PsD patients when compared to healthy controls, whereas direct T cell activation in PsD patients was reduced.
3.2.2 Cytokine levels according to biological and clinical parameters of MetS
Differences in cytokine secretion between PsD patients with high circulating CRP (> 3 mg/L), BMI above 25 kg/m², HOMA-score above 3, presence of MetS, or elevated abdominal circumference and PsD patients without these features were analysed (Table 2).
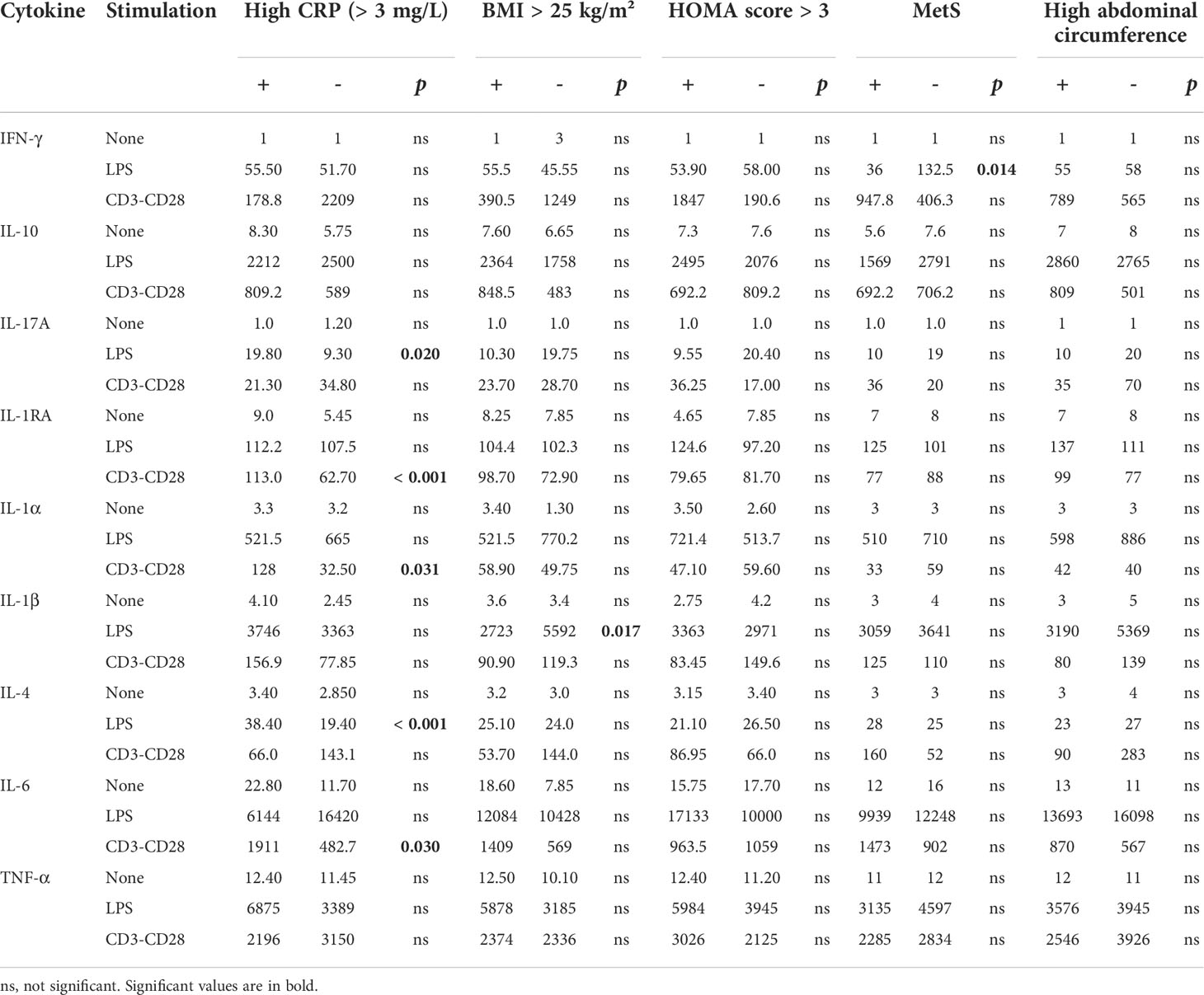
Table 2 Comparison of ex vivo cytokine production depending on MetS and related parameters in PsD patients (median; pg/mL).
In patients with high CRP, LPS-induced monocyte production of IL-17A and IL-4 was higher than in patients with low CRP, while direct stimulation of T cells had an indirectly stronger monocyte-stimulating effect with higher secretion of IL-1α and IL-6. PBMC from patients with high BMI secreted lower levels of IL-1β after LPS stimulation than those from normal BMI patients. When considering patients with MetS, stimulated monocytes were less prone to stimulate Th1 cells (less IFN-γ release) than those without MetS. Regarding these parameters, no difference was observed regarding HOMA-score or abdominal circumference.
3.2.3 Cytokine levels according to cutaneous or rheumatic activity in PsD patients
PBMC from patients with rheumatic activity (10 of 42) showed lower IFN-γ and TNF-α production after LPS stimulation (Figure 2A and Table 3). LPS-stimulated PBMC from patients with dermatological activity (7 of 42 PsD patients) displayed higher IL-17A production (Figure 2B).
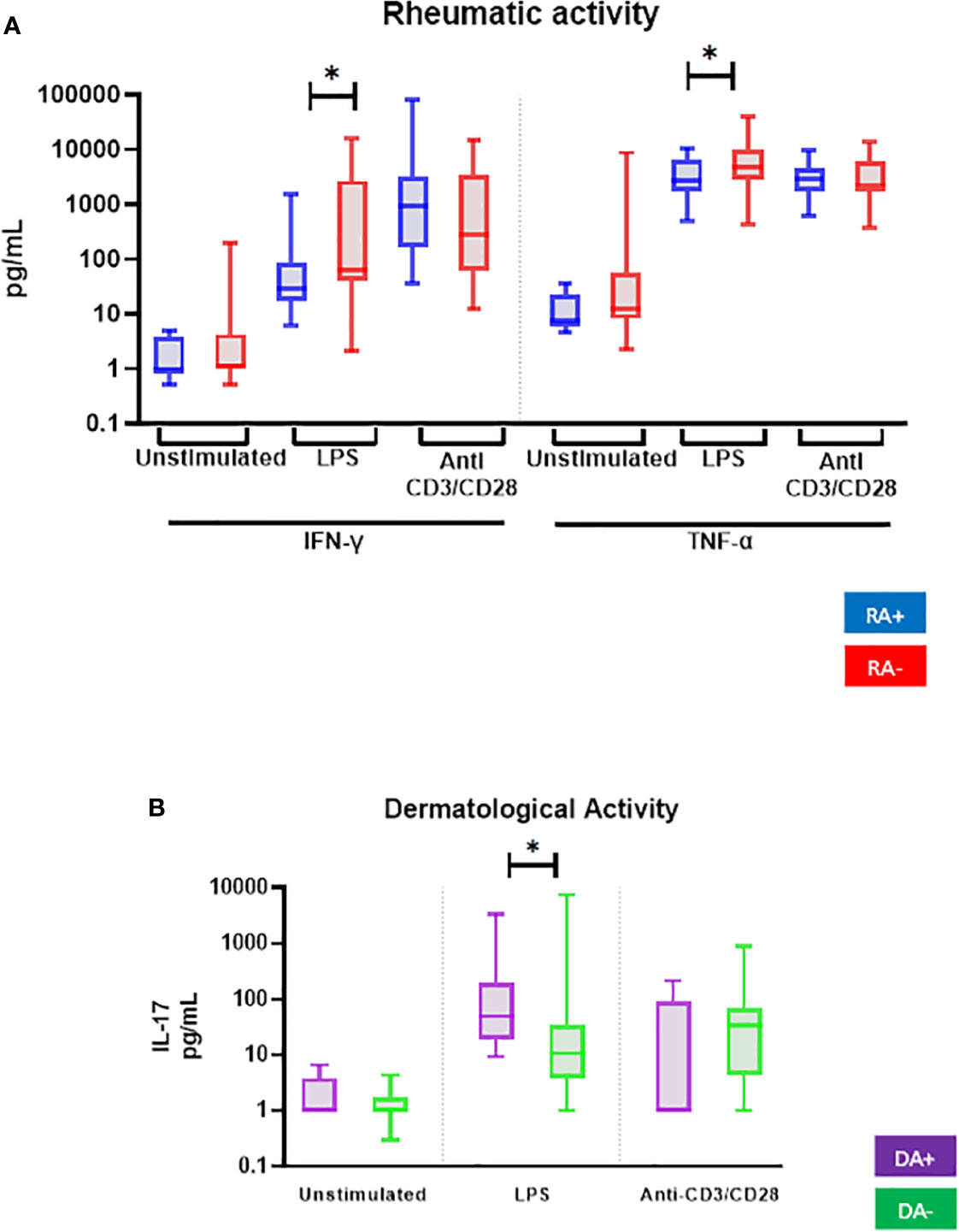
Figure 2 Comparison of ex vivo cytokine production of all PsD patients (n = 48) according to (A) their rheumatic (RA) and (B) dermatological (DA) disease activity. PBMCs from PsD patients (n = 48) were cultured at 1x106 cells/ml for 24 h at 37°C, 5% CO2 with and without stimulation of monocytes by E. coli LPS (1 µg/mL) or T lymphocytes by anti-CD3/CD28 mAb (2.5 × 104 beads/106 cells/ml). Supernatants were collected and cytokine levels were assayed by Luminex Multiplex ELISA. Results are presented using median values with minimum to maximum box-plot. Difference between the groups using non-parametric Mann-Whitney U test is presented in the top of the figure for each condition. *p < 0.05. RA, Rheumatic activity (n RA+ = 10); DA, Dermatological activity (n DA+ = 7).
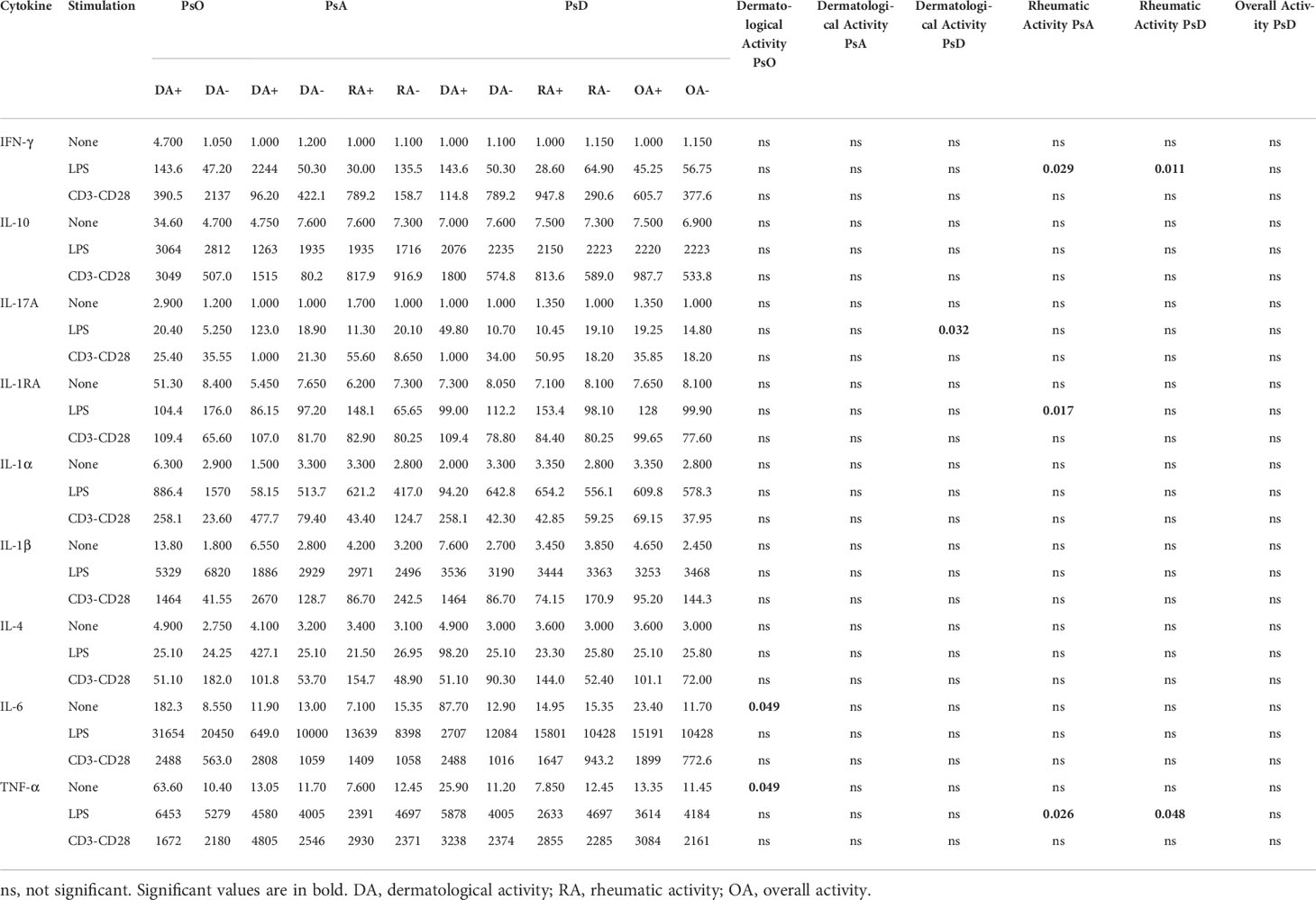
Table 3 Comparison of ex vivo cytokine production depending on disease activity (dermatological or rheumatic) in PsO patients, PsA patients, and PsD (median; pg/mL).
3.2.4 Comparison between PsO and PsA patients
PsO and PsA patients (with all rheumatologic phenotypes encompassed) were compared (Figure 3 and Supplementary Table 2). Unstimulated PBMC from both PsO and PsA displayed higher levels of spontaneous release of IL-6, TNF-α, and IL-1RA without difference between the two. Stimulation of monocytes by LPS induced higher activation of Th1 and Th2 cells, while T cell activation induced higher production of IL-6 in the two groups. A spontaneous inflammatory state (IL-6, TNF-α and IL-1RA) with monocyte-mediated activation of Th1 (IFN-γ, IL-6) cells is a common feature of PsO and PsA patients.
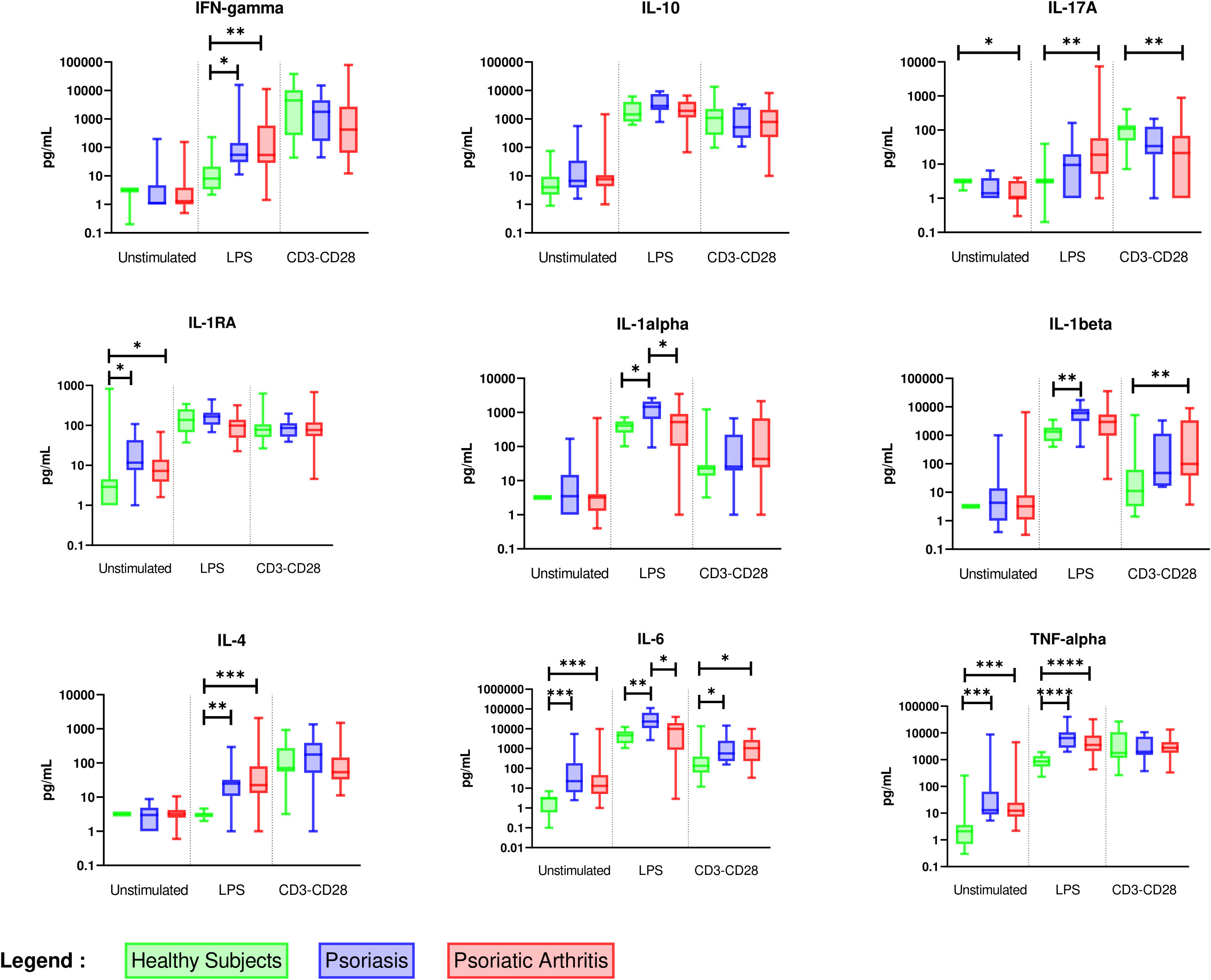
Figure 3 Overall comparison of ex vivo cytokine production between healthy subjects (n = 11), PsO (n = 11), and PsA (n = 37) patients. PBMCs from healthy subjects (n = 11), PsO (n = 11), and PsA (n = 37) patients were cultured at 1x106 cells/ml for 24 h at 37°C, 5% CO2 with and without stimulation of monocytes by E. coli LPS (1 µg/mL) or T lymphocytes by anti-CD3/CD28 mAb (2.5 × 104 beads/106 cells/ml). Supernatants were collected and cytokine levels were assayed by Luminex Multiplex ELISA. Results are presented using median values with minimum to maximum box-plot. Difference between the groups using non- parametric Kruskal-Wallis test with Dunn’s multiple comparison test is presented at the top of the figure for each condition. *p < 0.05; **p < 0.01; ***p < 0.001; ****p < 0.0001.
However, only PsO patients displayed an increased production of IL-1α, IL-1β, and IL-6 by stimulated monocytes, indicating a monocyte-predominant cytokine signature of PsO.
In PsA patients, stimulated monocytes activated Th17 cells, while T lymphocyte-stimulation reduced IL-17A but enhanced IL-1β production when compared to healthy controls. This finding suggests that Th17 cells need interaction with stimulated monocytes to produce IL-17A, while their own stimulation activated only monocytes in PsA.
3.3 Cytokine profiles in psoriatic arthritis
3.3.1 Cytokine profile of each clinical phenotype of PsA
Considering patients with PsA, we further identified the specific signature of each clinical phenotype that would reveal distinctive underlying pathways (Figure 4 and Supplementary Table 3). Peripheral PsA exhibited a cytokine profile distinct from axial PsA, characterized by Th17 and Th2 activation by activated monocytes and higher activation of monocytes by stimulated lymphocytes. Therefore, a specific peripheral PsA cytokine signature is characterised by a stronger monocyte activation with higher IL-6, IL-1α, and IL-1β levels after T lymphocyte stimulation, as well as greater activation of Th17 and Th2 cells after monocyte stimulation.
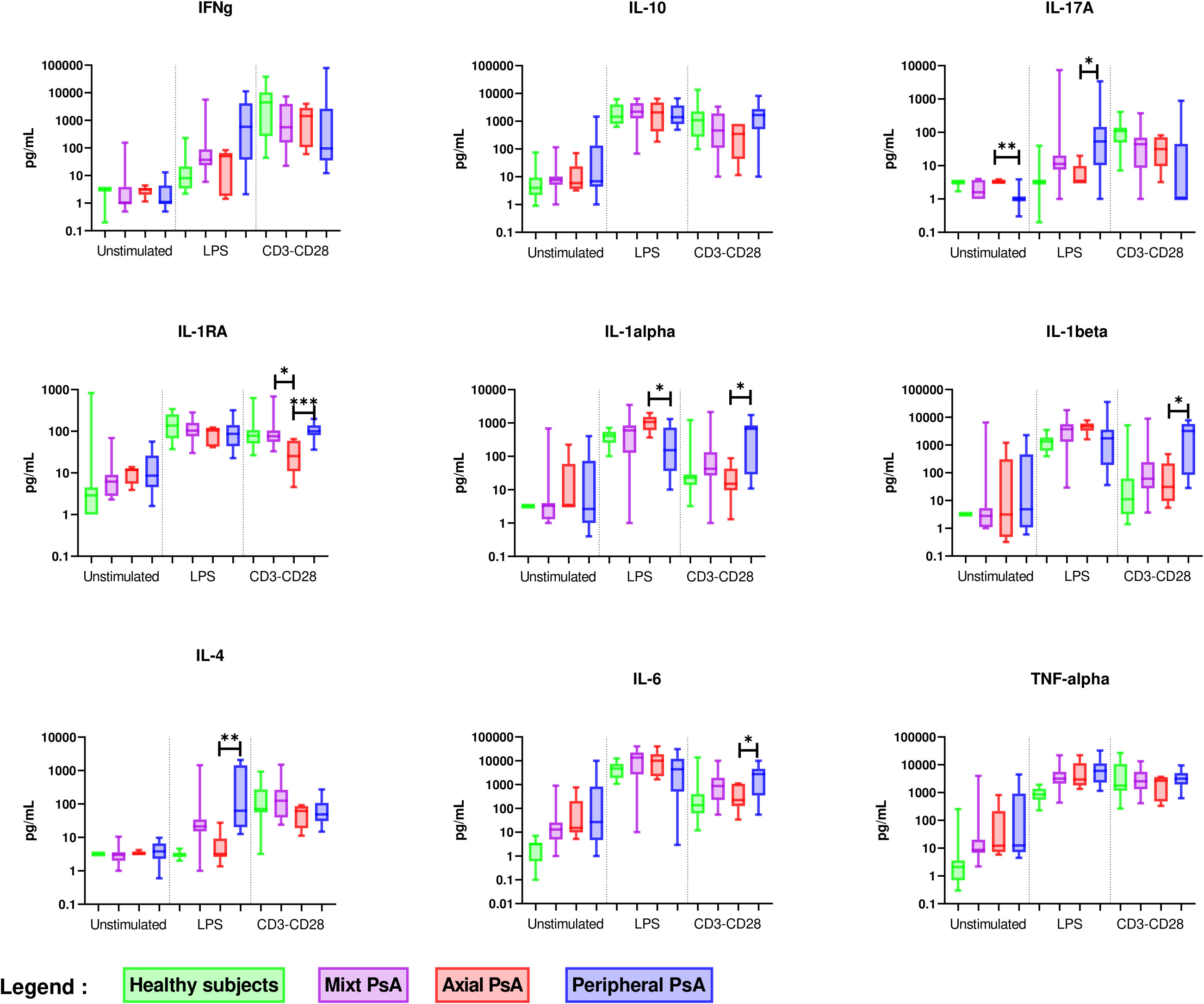
Figure 4 Comparison of ex vivo cytokine production between axial (n = 6), peripheral (n = 14) and mixed PsA (n = 17). PBMCs from axial (n = 6), peripheral (n = 14) and mixed PsA (n = 17) patients were cultured at 1x106 cells/ml for 24 h at 37°C, 5% CO2 with and without stimulation of monocytes by E. coli LPS (1 µg/mL) or T lymphocytes by anti-CD3/CD28 mAb (2.5 × 104 beads/106 cells/ml). Supernatants were collected and cytokine levels were assayed by Luminex Multiplex ELISA. Results are presented using median values with minimum to maximum box-plot. Difference between the groups using non parametric Kruskal-Wallis test with Dunn’s multiple comparison test is presented in the top of the figure for each condition. *p < 0.05; **p < 0.01; ***p < 0.001.
The features distinguishing axial PsA from peripheral PsA were higher production of IL-1α by stimulated monocytes, which is part of the PsO cytokine signature, and a lower production of IL-4 after monocyte stimulation. The singularity of an axial PsA cytokine profile is the absence of Th2 cell activation by stimulated monocytes.
3.3.2 Ex vivo cytokine profiles according to disease activity
As regards rheumatic activity in the 31 PsA patients, a Th1 and monocytic-predominant signature appeared. Higher production of IL-1RA after LPS stimulation was found, as was the case for IFN-γ and TNF-α as previously described in the PsD patients. No difference was observed according to the dermatological activity of PsA (Table 3).
3.3.3 Impact of DMARD initiation on cytokine profiles in psoriatic arthritis
Results of ex vivo cytokine production by PBMC before and after DMARD initiation were available for 6 patients (Figure 5). After treatment, lower production of IL-17A after anti-CD3/CD28 mAb stimulation and higher production of IL-1RA after LPS stimulation were observed, suggesting that the PsA treatment reduced Th17 pro-inflammatory state and reinforced the control of inflammation by monocytes through IL-1RA release.
Figure 5 Comparison of ex vivo cytokine production in PsA patients before and after DMARD initiation (n = 6). PBMCs from 6 PsA patients were cultured at 1x106 cells/ml for 24 h at 37°C, 5% CO2 with and without stimulation of monocytes by E. coli LPS (1 µg/mL) or T lymphocytes by anti-CD3/CD28 mAb (2.5 × 104 beads/106 cells/ml). Supernatants were collected and cytokine levels were assayed by Luminex Multiplex ELISA. Difference between the groups using non-parametric Wilcoxon matched-pairs test is presented in the top of the figure for each condition. *p < 0.05. Each line represents a patient.
4 Discussion
This exploratory study analysed the ex vivo cytokine production by PBMC stimulated with either LPS or anti-CD3/CD28 mAb in clinically well-described populations of PsA and PsO patients (26, 27). PBMCs have the advantage of being easily accessible to perform cytokine measurements after stimulation, although in inflammatory arthritis many other cell types produce cytokines, including fibroblasts, synoviocytes, immune infiltrated cells and osteoblasts. These cells can contribute to the cytokine network of the disease, particularly inside the synovium, and may have different cytokine production patterns (28–31). However, isolating these infiltrated cells is not easy since it requires invasive techniques, making it not suitable for a daily analysis, by contrast to PBMC ex vivo cytokine production (32).
In our study, LPS and anti-CD3/CD28 mAb stimulation of PBMCs were used to stimulate monocytes and T-cells, respectively (23, 24). LPS is a TLR4 agonist traditionally considered as a monocyte activator since TLR4 expression is mainly expressed in myeloid cells in humans although minor expression level of TLR4 has been also described in some lymphoid cell subsets (33). Concerning the activation of T cells following LPS stimulation, the literature is contradictory. Indeed some authors found a direct activation of T-cell by LPS through TLR4 (34) meanwhile some authors hypothesize that it is the result of cell-cell interactions with monocytes. Recent single cell analyses of LPS or anti-CD3/CD28 stimulated-PBMC confirm that LPS only activate monocytes (35). Considering this recent literature, we analysed our results considering that LPS activates monocytes and not T-cells, even if a marginal activation of T-cells could not be ruled out.
Several studies have attempted to determine specific cytokine profiles of PsD. Increased IL-17A production by PBMC after 4 days of anti-CD3/CD28 mAb stimulation has been described in PsO and PsA patients (36). Another study reported high IL-17A in supernatants of CD4+ T cells after 6 days of culture with anti-CD3/CD28 mAb stimulation in very early PsA (37), while we report lower IL-17A levels with similar conditions of stimulation. In our study, 24h of stimulation were used to limit secondary or tertiary production of cytokines that can modify the profile following longer stimulation. Another explanation for this result may be the need for interactions with other cell types and not only T cell stimulation to induce higher IL-17A levels early in PsD. In accordance with Leijten et al. (38), higher IL-1RA levels were observed in unstimulated conditions compared to healthy controls, which reflects an attempt by the immune system to alleviate systemic inflammation. Interestingly, while LPS stimulation induced increased production of all the cytokines studied in PsD patients, no increase of IL-4, IL-17, and IFN-γ was observed in healthy controls. All in all, this suggests a particular ability of circulating T cells to be activated via monocytes in PsD.
Patients suffering from PsD are known to present MetS features, a clinical entity with complex and multiple impacts on the immune system, as reviewed by Andersen et al. (39). There is no literature on ex vivo PBMC-derived cytokine profiles comparing patients according to their BMI status. A recent study showed an exhaustion of T cells inside adipose tissue of obese patients, which is concordant with our findings (40). As reported by Diehl et al., IL-6 exerts an inhibitory effect on Th1 differentiation, thereby lowering IFN-γ production (41). Therefore, since LPS stimulates production of IL-6 by PBMC, we can hypothesize that stimulated monocytes polarize toward a pro-inflammatory phenotype with increased production of IL-6, which hampers IFN-γ production by Th1 cells, especially as IL-6 levels did not differ between patients with and without MetS.
Comparison of the cytokine profiles of the different clinical phenotypes of PsD patients demonstrated features distinctive to each subtype. In PsO patients, our results confirm previous studies highlighting the pivotal role of monocytes and their cytokines (IL-1α, IL-1β, and IL-6) (42–44). Likewise, clinical improvement of PsO with biological therapies has been associated with decreased monocyte activity (45). Since IL-1β induces IL-6 secretion by monocytes (46), we hypothesize that a cascade of cell activation might contribute to IL-6 production. Nevertheless, IL-6 implication in PsO pathogenesis remains unclear, and its inhibition in clinical practice is not efficient (47). Intriguingly, IL-17A is not present in PsO cytokine signature, whereas IL-17A is one of the main cytokines that drives cutaneous inflammation in psoriasis (48). However, activated monocytes may be one of the missing links. In fact, IL-1β enhances the differentiation and expansion of Th17 cells (49). Moreover, the IL-1β/IL-1R pathway regulates dermal IL-17A-producing cells and the stimulation of keratinocytes in PsO (50). Considering the absence of IL-17A expression by PBMCs from PsO patients, we hypothesize that IL-17A might be produced essentially by tissue-resident cells such as type 3 innate lymphoid cells (ILC3) for instance (51).
The specific cytokine profile of peripheral PsA raises the interesting concept of crosstalk between immune cells subtypes as a crucial element of disease pathophysiology. In 1997, Chizzolini et al. demonstrated that activated cells could induce the production of cytokines by monocytes through cell-cell contact (52). In 2009, Evans et al. demonstrated that LPS-activated monocytes induce Th17 responses when co-cultured with CD4+ T cells, thereby highlighting the importance of monocyte/T cell interaction in the shaping of inflammatory T cell responses (53). These interactions seem to be the key to the vicious circle of inflammation found in peripheral but not in axial PsA. It is also well-known that peripheral PsA patients present a more inflammatory phenotype, reflected by higher levels of erythrocyte sedimentation rate or CRP than axial PsA patients (54). We can parallel a study focusing on innate lymphoid cells (ILCs) in different rheumatic diseases with either a synovitis-predominant pattern or not. Differences in ILC populations were found between these two groups, but not between the different rheumatic diseases, implying that the clinical profile may be more important than the disease itself to distinguish particular immune patterns (55). The importance of T cell activation in PsA has also been highlighted in a study reporting a therapeutic strategy based on targeted treatments in which patients with an activated Th17 predominant pattern were treated by an IL-17A inhibitor, while patients harbouring an activated Th1 predominant pattern received an anti-p40 treatment. This study showed better treatment efficacy, highlighting the importance of T cells in the pathophysiology of peripheral PsA (56).
Focusing on disease activity, the spontaneous production of IL-6 and TNF-α was increased in patients with dermatological activity, as previously reported for TNF-α, but not for IL-6 (57). In accordance with our results showing an increase of IL-17A after LPS stimulation in PsD patients with dermatological activity, it has been reported that IL-17+ T cells were increased in active psoriasis (58, 59) and that elevated concentrations of IL-17A in serum were correlated with the dermatological activity (60). In PsA, in agreement with our results, serum IL-1RA was associated with joint severity and correlated with tender and swollen joints (12). Lower levels of IFN-γ associated with rheumatic disease activity was to be expected, given the accumulating evidence of a “yin and yang” effect of IFN-γ on the inflammatory process (61), particularly with negative regulation of IL-17 expression by Th17 cells (62).
After DMARD initiation, we observed decreased IL-17A production and increased IL-1RA production, in accordance with the decreased inflammatory process. Previous studies reported not only decreased serum IL-17A levels (63), but also decreased TNF-α after TNF-α inhibitor treatment, which was not observed in our ex vivo analysis (64, 65). It is noteworthy that the approaches were different, and that, to our knowledge, no ex vivo PBMC stimulation study has focused on this issue.
One of the limits of this study is the inclusion of subjects who, for the most of part, already benefited from DMARDs therapies, which have an impact on cytokines production (66). However, this feature equally concerns both PsO and PsA patients, thereby limiting the bias. The number of patients included is also limited but comparable to other studies in the field (10, 11, 13, 14, 16, 17). In any event, our results should be viewed with all the necessary precautions. While the ex vivo approach is dynamic and easy to implement to study the contribution of circulating cells to the diseases, it excludes the contribution of resident cells. Another limit of this global approach is that the underlying mechanisms of the complex interactions between immune cells and their subsequent cytokine productions were not fully elucidated.
Taken together, our results lead us to propose a core cytokine profile for all PsD patients and specific cytokine patterns distinguishing the different PsD sub-phenotypes, as summarized in Figure 6. The perspective of characterizing distinct particular immune profiles might be of great interest to tailor patient treatment with a potential clinical impact on the effectiveness of biologic DMARDs (56).
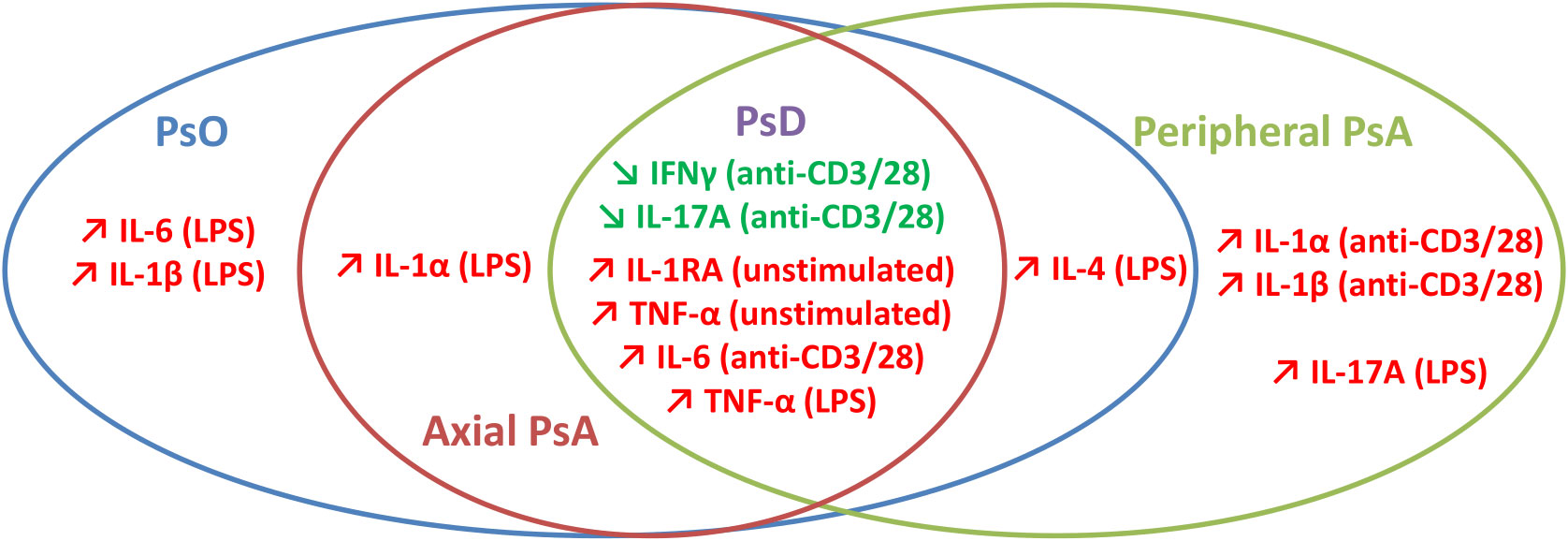
Figure 6 Synthetic view of discriminative ex vivo cytokine profiles in psoriatic disease.
In conclusion, PsO and PsA share not only the common cytokine patterns common to PsD, but a number of specific cytokine patterns, as well. PsO exhibits a monocyte-predominant cytokine signature, whereas peripheral PsA signature is characterized by strong monocyte activation by stimulated T cells and Th17 cell activation by stimulated monocytes. Our study highlights the crucial role of immune cell interaction potentially involving soluble mediators and/or cell-cell contact with different patterns of interaction depending on clinical phenotype. Further studies should be performed to increase knowledge of the immunological mechanisms that underlie every sub-phenotype of PsD.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The studies involving human participants were reviewed and approved by Poitiers University Hospital. Written informed consent for participation was not required for this study in accordance with the national legislation and the institutional requirements.
Author contributions
AD, TD, and PV performed the experiments. CS, EG, and PV acquired data. GL, AD, TD, PV, CS, FM, J-CL, and EG analysed data. GL performed statistical analysis. GL wrote first draft of the manuscript. GL, EG, FM, J-CL, and J-FJ corrected manuscript. All authors contributed to the article and approved the submitted version.
Funding
This research was supported by Poitiers University Hospital and Poitiers University.
Acknowledgments
Pr. Stéphanie RAGOT for her statistical advices and validation. M. Jeffrey Arsham for language editing of the manuscript. The authors thank the European Union for an equipment grant program for research laboratories and platforms.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2022.993363/full#supplementary-material
References
1. Rech J, Sticherling M, Stoessel D, Biermann MHC, Häberle BM, Reinhardt M. Psoriatic arthritis epidemiology, comorbid disease profiles and risk factors: Results from a claims database analysis. Rheumatol Adv Pract (2020) 4(2):rkaa033. doi: 10.1093/rap/rkaa033
2. Scarpa R, Ayala F, Caporaso N, Olivieri I. Psoriasis, psoriatic arthritis, or psoriatic disease? J Rheumatol (2006) 33:210–2.
3. Ritchlin C. Psoriatic disease–from skin to bone. Nat Rev Rheumatol (2007) 3:698–706. doi: 10.1038/ncprheum0670
4. Lubrano E, Scriffignano S, Perrotta FM. Psoriatic arthritis, psoriatic disease, or psoriatic syndrome? J Rheumatol (2019) 46:1428–30. doi: 10.3899/jrheum.190054
5. Sobchak C, Eder L. Cardiometabolic disorders in psoriatic disease. Curr Rheumatol Rep (2017) 19:63. doi: 10.1007/s11926-017-0692-2
6. Villani AP, Rouzaud M, Sevrain M, Barnetche T, Paul C, Richard M-A, et al. Prevalence of undiagnosed psoriatic arthritis among psoriasis patients: Systematic review and meta-analysis. J Am Acad Dermatol (2015) 73:242–8. doi: 10.1016/j.jaad.2015.05.001
7. Wani A, Ganai BA, Akhtar T, Narang T, Kaur R. Association of proinflammatory cytokine IL-20 gene polymorphism with psoriasis in north Indian population. Egyptian J Med Hum Genet (2018) 19:201–5. doi: 10.1016/j.ejmhg.2017.09.002
8. Traks T, Keermann M, Prans E, Karelson M, Loite U, Kõks G, et al. Polymorphisms in IL36G gene are associated with plaque psoriasis. BMC Med Genet (2019) 20:10. doi: 10.1186/s12881-018-0742-2
9. Duffin KC, Woodcock J, Krueger GG. Genetic variations associated with psoriasis and psoriatic arthritis found by genome-wide association. Dermatol Ther (2010) 23:101–13. doi: 10.1111/j.1529-8019.2010.01303.x
10. Szodoray P, Alex P, Chappell-Woodward CM, Madland TM, Knowlton N, Dozmorov I, et al. Circulating cytokines in Norwegian patients with psoriatic arthritis determined by a multiplex cytokine array system. Rheumatology (2007) 46:417–25. doi: 10.1093/rheumatology/kel306
11. Bautista-Herrera L, de la Cruz-Mosso U, Román-Fernández I, Parra-Rojas I, Soñanez-Organis J, Hernández-Bello J, et al. A potential inflammatory role of IL-31 in psoriatic arthritis: A correlation with Th17 cytokine profile. Int J Immunopathol Pharmacol (2020) 34:205873842090718. doi: 10.1177/2058738420907186
12. Elkayam O, Yaron I, Shirazi I, Yaron M, Caspi D. Serum levels of IL-10, IL-6, IL-1ra, and sIL-2R in patients with psoriatic arthritis. Rheumatol Int (2000) 19:101–5. doi: 10.1007/s002960050111
13. Pirowska M, Obtułowicz A, Lipko-Godlewska S, Goździalska A, Podolec K, Wojas-Pelc A. The level of proinflammatory cytokines: Interleukins 12, 23, 17 and tumor necrosis factor α in patients with metabolic syndrome accompanying severe psoriasis and psoriatic arthritis. Adv Dermatol Allergol (2018) 35:360–6. doi: 10.5114/ada.2018.77665
14. Coimbra S, Oliveira H, Reis F, Belo L, Rocha S, Quintanilha A, et al. Interleukin (IL)-22, IL-17, IL-23, IL-8, vascular endothelial growth factor and tumour necrosis factor-α levels in patients with psoriasis before, during and after psoralen-ultraviolet a and narrowband ultraviolet b therapy: Psoriasis vulgaris cytokines and PUVA/NB-UVB therapy. Br J Dermatol (2010) 163:1282–90. doi: 10.1111/j.1365-2133.2010.09992.x
15. Takahashi H, Tsuji H, Hashimoto Y, Ishida-Yamamoto A, Iizuka H. Serum cytokines and growth factor levels in Japanese patients with psoriasis: Serum cytokine levels and disease activity of psoriasis. Clin Exp Dermatol (2009) 35:645–9. doi: 10.1111/j.1365-2230.2009.03704.x
16. Nishibu A, Han G-W, Iwatsuki K, Matsui T, Inoue M, Akiba H, et al. Overexpression of monocyte-derived cytokines in active psoriasis: A relation to coexistent arthropathy. J Dermatol Sci (1999) 21:63–70. doi: 10.1016/S0923-1811(99)00031-6
17. Bosè F, Capsoni F, Molteni S, Raeli L, Diani M, Altomare A, et al. Differential expression of interleukin-2 by anti-CD3-stimulated peripheral blood mononuclear cells in patients with psoriatic arthritis and patients with cutaneous psoriasis. Clin Exp Dermatol (2014) 39:385–90. doi: 10.1111/ced.12251
18. Taylor W, Gladman D, Helliwell P, Marchesoni A, Mease P, Mielants H, et al. Classification criteria for psoriatic arthritis: Development of new criteria from a large international study. Arthritis Rheum (2006) 54:2665–73. doi: 10.1002/art.21972
19. Feldman SR. Psoriasis assessment tools in clinical trials. Ann Rheum Dis (2005) 64:ii65–8. doi: 10.1136/ard.2004.031237
20. Salaffi F, Ciapetti A, Carotti M, Gasparini S, Gutierrez M. Disease activity in psoriatic arthritis: Comparison of the discriminative capacity and construct validity of six composite indices in a real world. BioMed Res Int (2014) 2014:1–12. doi: 10.1155/2014/528105
21. Fernández-Sueiro JL, Willisch A, Pértega-DÃaz S, Tasende JAP, Fernández-LÃ3pez JC, Villar NO, et al. Validity of the bath ankylosing spondylitis disease activity index for the evaluation of disease activity in axial psoriatic arthritis. Arthritis Care Res (2010) 62:78–85. doi: 10.1002/acr.20017
22. Grundy SM, Cleeman JI, Daniels SR, Donato KA, Eckel RH, Franklin BA, et al. Diagnosis and management of the metabolic syndrome: An American heart Association/National heart, lung, and blood institute scientific statement. Circulation (2005) 112:2735–52. doi: 10.1161/CIRCULATIONAHA.105.169404
23. Trickett A, Kwan YL. T Cell stimulation and expansion using anti-CD3/CD28 beads. J Immunol Methods (2003) 275:251–5. doi: 10.1016/S0022-1759(03)00010-3
24. Ramírez-Pérez S, Hernández-Palma LA, Oregon-Romero E, Anaya-Macías BU, García-Arellano S, González-Estevez G, et al. Downregulation of inflammatory cytokine release from IL-1β and LPS-stimulated PBMC orchestrated by ST2825, a MyD88 dimerisation inhibitor. Molecules (2020) 25:4322. doi: 10.3390/molecules25184322
25. Zhou X, Fragala MS, McElhaney JE, Kuchel GA. Conceptual and methodological issues relevant to cytokine and inflammatory marker measurements in clinical research. Curr Opin Clin Nutr Metab Care (2010) 13:541–7. doi: 10.1097/MCO.0b013e32833cf3bc
27. Gladman DD. Psoriatic arthritis: Epidemiology, clinical features, course, and outcome. Ann Rheum Dis (2005) 64:ii14–7. doi: 10.1136/ard.2004.032482
28. Veale DJ, Fearon U. The pathogenesis of psoriatic arthritis. Lancet (2018) 391:2273–84. doi: 10.1016/S0140-6736(18)30830-4
29. Ganesan R, Rasool M. Fibroblast-like synoviocytes-dependent effector molecules as a critical mediator for rheumatoid arthritis: Current status and future directions. Int Rev Immunol (2017) 36:20–30. doi: 10.1080/08830185.2016.1269175
30. Danis J, Janovák L, Gubán B, Göblös A, Szabó K, Kemény L, et al. Differential inflammatory-response kinetics of human keratinocytes upon cytosolic RNA- and DNA-fragment induction. IJMS (2018) 19:774. doi: 10.3390/ijms19030774
31. Mensah KA, Schwarz EM, Ritchlin CT. Altered bone remodeling in psoriatic arthritis. Curr Rheumatol Rep (2008) 10:311–7. doi: 10.1007/s11926-008-0050-5
32. Amos PJ, Cagavi Bozkulak E, Qyang Y. Methods of cell purification: A critical juncture for laboratory research and translational science. Cells Tissues Organs (2012) 195:26–40. doi: 10.1159/000331390
33. Vaure C, Liu Y. A comparative review of toll-like receptor 4 expression and functionality in different animal species. Front Immunol (2014) 5:316. doi: 10.3389/fimmu.2014.00316
34. Zanin-Zhorov A, Tal-Lapidot G, Cahalon L, Cohen-Sfady M, Pevsner-Fischer M, Lider O, et al. Cutting edge: T cells respond to lipopolysaccharide innately via TLR4 signaling. J Immunol (2007) 179:41–4. doi: 10.4049/jimmunol.179.1.41
35. Lawlor N, Nehar-Belaid D, Grassmann JDS, Stoeckius M, Smibert P, Stitzel ML, et al. Single cell analysis of blood mononuclear cells stimulated through either LPS or anti-CD3 and anti-CD28. Front Immunol (2021) 12:636720. doi: 10.3389/fimmu.2021.636720
36. Benham H, Norris P, Goodall J, Wechalekar MD, FitzGerald O, Szentpetery A, et al. Th17 and Th22 cells in psoriatic arthritis and psoriasis. Arthritis Res Ther (2013) 15:R136. doi: 10.1186/ar4317
37. Leipe J, Grunke M, Dechant C, Reindl C, Kerzendorf U, Schulze-Koops H, et al. Role of Th17 cells in human autoimmune arthritis. Arthritis Rheum (2010) 62:2876–85. doi: 10.1002/art.27622
38. Leijten E, Tao W, Pouw J, van Kempen T, Olde Nordkamp M, Balak D, et al. Broad proteomic screen reveals shared serum proteomic signature in patients with psoriatic arthritis and psoriasis without arthritis. Rheumatology (2021) 60:751–61. doi: 10.1093/rheumatology/keaa405
39. Andersen CJ, Murphy KE, Fernandez ML. Impact of obesity and metabolic syndrome on immunity. Adv Nutr (2016) 7:66–75. doi: 10.3945/an.115.010207
40. Porsche CE, Delproposto JB, Geletka L, O’Rourke R, Lumeng CN. Obesity results in adipose tissue T cell exhaustion. JCI Insight (2021) 6:e139793. doi: 10.1172/jci.insight.139793
41. Diehl S, Anguita J, Hoffmeyer A, Zapton T, Ihle JN, Fikrig E, et al. Inhibition of Th1 differentiation by IL-6 is mediated by SOCS1. Immunity (2000) 13:805–15. doi: 10.1016/S1074-7613(00)00078-9
42. Wang Y, Edelmayer R, Wetter J, Salte K, Gauvin D, Leys L, et al. Monocytes/Macrophages play a pathogenic role in IL-23 mediated psoriasis-like skin inflammation. Sci Rep (2019) 9:5310. doi: 10.1038/s41598-019-41655-7
43. Neuner P, Urbanski A, Trautinger F, Möller A, Kirnbauer R, Kapp A, et al. Increased IL-6 production by monocytes and keratinocytes in patients with psoriasis. J Invest Dermatol (1991) 97:27–33. doi: 10.1111/1523-1747.ep12477880
44. Poubelle PE, Pagé N, Longchamps M, Sampaio Moura N, Beck DB, Aksentijevich I, et al. The use of leukocytes’ secretome to individually target biological therapy in autoimmune arthritis: A case report. Clin Trans Med (2019) 8(1):19. doi: 10.1186/s40169-019-0236-7
45. Yamanaka K, Umezawa Y, Yamagiwa A, Saeki H, Kondo M, Gabazza EC, et al. Biologic therapy improves psoriasis by decreasing the activity of monocytes and neutrophils. J Dermatol (2014) 41:679–85. doi: 10.1111/1346-8138.12560
46. Tosato G, Jones K. Interleukin-1 induces interleukin-6 production in peripheral blood monocytes. Blood (1990) 75:1305–10. doi: 10.1182/blood.V75.6.1305.1305
47. Madureira P, Pimenta SS, Bernardo A, Brito JS, Bernardes M, Costa L. Off-label use of tocilizumab in psoriatic arthritis: Case series and review of the literature. Acta Reumatol Port (2016) 41:251–5.
48. Blauvelt A, Chiricozzi A. The immunologic role of IL-17 in psoriasis and psoriatic arthritis pathogenesis. Clin Rev Allergy Immunol (2018) 55:379–90. doi: 10.1007/s12016-018-8702-3
49. Wilson NJ, Boniface K, Chan JR, McKenzie BS, Blumenschein WM, Mattson JD, et al. Development, cytokine profile and function of human interleukin 17–producing helper T cells. Nat Immunol (2007) 8:950–7. doi: 10.1038/ni1497
50. Cai Y, Xue F, Quan C, Qu M, Liu N, Zhang Y, et al. A critical role of the IL-1β-IL-1R signaling pathway in skin inflammation and psoriasis pathogenesis. J Invest Dermatol (2019) 139:146–56. doi: 10.1016/j.jid.2018.07.025
51. Bielecki P, Riesenfeld SJ, Hütter J-C, Torlai Triglia E, Kowalczyk MS, Ricardo-Gonzalez RR, et al. Skin-resident innate lymphoid cells converge on a pathogenic effector state. Nature (2021) 592(7852):128–132. doi: 10.1038/s41586-021-03188-w
52. Chizzolini C, Chicheportiche R, Burger D, Dayer J-M. Human Th1 cells preferentially induce interleukin (IL)-1β while Th2 cells induce IL-1 receptor antagonist production upon cell/cell contact with monocytes. Eur J Immunol (1997) 27:171–7. doi: 10.1002/eji.1830270125
53. Evans HG, Gullick NJ, Kelly S, Pitzalis C, Lord GM, Kirkham BW, et al. In vivo activated monocytes from the site of inflammation in humans specifically promote Th17 responses. Proc Natl Acad Sci USA (2009) 106:6232–7. doi: 10.1073/pnas.0808144106
54. Feld J, Ye JY, Chandran V, Inman RD, Haroon N, Cook R, et al. Is axial psoriatic arthritis distinct from ankylosing spondylitis with and without concomitant psoriasis? Rheumatology (2020) 59:1340–6. doi: 10.1093/rheumatology/kez457
55. Kato M, Ikeda K, Sugiyama T, Tanaka S, Iida K, Suga K, et al. Associations of ultrasound-based inflammation patterns with peripheral innate lymphoid cell populations, serum cytokines/chemokines, and treatment response to methotrexate in rheumatoid arthritis and spondyloarthritis. PloS One (2021) 16:e0252116. doi: 10.1371/journal.pone.0252116
56. Miyagawa I, Nakayamada S, Nakano K, Kubo S, Iwata S, Miyazaki Y, et al. Precision medicine using different biological DMARDs based on characteristic phenotypes of peripheral T helper cells in psoriatic arthritis. Rheumatology (2019) 58:336–44. doi: 10.1093/rheumatology/key069
57. Mizutani H, Ohmoto Y, Mizutani T, Murata M, Shimizu M. Role of increased production of monocytes TNF-α, IL-1β and IL-6 in psoriasis: Relation to focal infection, disease activity and responses to treatments. J Dermatol Sci (1997) 14:145–53. doi: 10.1016/S0923-1811(96)00562-2
58. Colombo E, Galleri G, Erre GL, Piras C, Biondi G, Taras L, et al. Peripheral blood CD8+ T-cell profiles in patients with psoriatic arthritis: A cross-sectional case-control study. Eur Rev Med Pharmacol Sci (2017) 21:5166–71. doi: 10.26355/eurrev_201711_13834
59. Aguilar-Flores C, Castro-Escamilla O, Ortega-Rocha EM, Maldonado-García C, Jurado-Santa Cruz F, Pérez-Montesinos G, et al. Association of pathogenic Th17 cells with the disease severity and its potential implication for biological treatment selection in psoriasis patients. Mediators Inflammation (2020) 2020:1–16. doi: 10.1155/2020/8065147
60. Michalak-Stoma A, Bartosińska J, Kowal M, Raczkiewicz D, Krasowska D, Chodorowska G. IL-17A in the psoriatic patients’ serum and plaque scales as potential marker of the diseases severity and obesity. Mediators Inflammation (2020) 2020:1–9. doi: 10.1155/2020/7420823
61. Kelchtermans H, Billiau A, Matthys P. How interferon-γ keeps autoimmune diseases in check. Trends Immunol (2008) 29:479–86. doi: 10.1016/j.it.2008.07.002
62. Harrington LE, Hatton RD, Mangan PR, Turner H, Murphy TL, Murphy KM, et al. Interleukin 17–producing CD4+ effector T cells develop via a lineage distinct from the T helper type 1 and 2 lineages. Nat Immunol (2005) 6:1123–32. doi: 10.1038/ni1254
63. Sakkas LI, Bogdanos DP. Are psoriasis and psoriatic arthritis the same disease? the IL-23/IL-17 axis data. Autoimmun Rev (2017) 16:10–5. doi: 10.1016/j.autrev.2016.09.015
64. Cordiali-Fei P, Ardigò M, Mastroianni A, Giuliani A, D’Agosto G, Bordignon V, et al. Serum cytokines and bioumoral immunological characterization of psoriatic patients in long term etanercept treatment. Int J Immunopathol Pharmacol (2008) 21:643–9. doi: 10.1177/039463200802100318
65. Mastroianni A, Minutilli E, Mussi A, Bordignon V, Trento E, D’Agosto G, et al. Cytokine profiles during infliximab monotherapy in psoriatic arthritis: Cytokine profiles and infliximab monotherapy in PsA. Br J Dermatol (2005) 153:531–6. doi: 10.1111/j.1365-2133.2005.06648.x
Keywords: psoriasis, psoriatic arthritis (PsA), psoriatic disease, cytokine signature, ex vivo stimulation
Citation: Larid G, Delwail A, Dalle T, Vasseur P, Silvain C, Jégou J-F, Morel F, Lecron J-C and Gervais E (2022) Ex vivo cytokine production in psoriatic disease: Towards specific signatures in cutaneous psoriasis and peripheral psoriatic arthritis. Front. Immunol. 13:993363. doi: 10.3389/fimmu.2022.993363
Received: 13 July 2022; Accepted: 10 October 2022;
Published: 08 November 2022.
Edited by:
Mrinal K. Sarkar, University of Michigan, United StatesReviewed by:
Rajeev Kumar Pandey, Johns Hopkins Medicine, United StatesOlesya Plazyo, Michigan Medicine, University of Michigan, United States
Copyright © 2022 Larid, Delwail, Dalle, Vasseur, Silvain, Jégou, Morel, Lecron and Gervais. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Elisabeth Gervais, ZWxpc2FiZXRoLmdlcnZhaXNAY2h1LXBvaXRpZXJzLmZy
†ORCID: Elisabeth Gervais, 0000-0003-3513-2989