 Yibo Chen1†
Yibo Chen1† Xin Chen
Xin Chen- 1State Key Laboratory of Quality Research in Chinese Medicine, Institute of Chinese Medical Sciences, University of Macau, Macau, Macau SAR, China
- 2Ministry of Education (MoE) Frontiers Science Center for Precision Oncology, University of Macau, Macau, Macau SAR, China
- 3Department of Pharmaceutical Sciences, Faculty of Health Sciences, University of Macau, Macau, Macau SAR, China
- 4Guangdong-Hong Kong-Macau Joint Lab on Chinese Medicine and Immune Disease Research, Macau, Macau SAR, China
TNFR2 agonists have been investigated as potential therapies for inflammatory diseases due to their ability to activate and expand immunosuppressive CD4+Foxp3+ Treg cells and myeloid-derived suppressor cells (MDSCs). Despite TNFR2 being predominantly expressed in Treg cells at high levels, activated effector T cells also exhibit a certain degree of TNFR2 expression. Consequently, the role of TNFR2 signaling in coordinating immune or inflammatory responses under different pathological conditions is complex. In this review article, we analyze possible factors that may determine the therapeutic outcomes of TNFR2 agonism, including the levels of TNFR2 expression on different cell types, the biological properties of TNFR2 agonists, and disease status. Based on recent progress in the understanding of TNFR2 biology and the study of TNFR2 agonistic agents, we discuss the future direction of developing TNFR2 agonists as a therapeutic agents.
Introduction
Tumor necrosis factor (TNF) is one of the most potent pro-inflammatory cytokines that cause cell death and promote inflammatory responses, and high levels of TNF are attributable to the pathogenesis of autoimmune disease (1, 2). Anti-TNF therapeutics have been used as a first-line biological treatment of a variety of inflammatory diseases, including rheumatoid arthritis (RA), psoriasis, and inflammatory bowel disease (IBD) (3–5). However, paradoxically autoimmune inflammation frequently occurred in a subset of patients who received anti-TNF treatment. For example, anti-TNF therapy can increase the incidence of multiple sclerosis (6). These observations brought intense interest in elucidating the cause and mechanism. TNF receptor type I (TNFR1) and TNFR2 are two different receptors that mediated the biological function of TNF (7). A recent report showed that polymorphisms in TNFR2 frequently occurred in patients with IBD or RA (8, 9), which suggests that TNFR2 signaling plays an essential role in preventing these diseases.
TNFR2 is preferentially expressed by immunosuppressive cells, including Tregs, MDSCs, and some endothelial progenitor cells (EPCs) (10). Now, there is compelling evidence showing that TNF-TNFR2 signaling plays an important role in curbing pro-inflammatory responses and promoting tissue regeneration. It was reported that TNFR2 deficiency aggravated autoimmune inflammatory responses in collagen-induced arthritis (CIA) (11), experimental autoimmune encephalomyelitis (EAE) (12), graft-versus-host diseases (GVHD) (13), and psoriasis (14). Moreover, several studies have shown that TNFR2 agonists protect mice from autoimmune inflammatory diseases and degenerative diseases (15–17). Thus, TNFR2 agonists have been proposed as a novel strategy for the treatment of autoimmune diseases, by mainly activating and expanding TNFR2-expressing Tregs, and MDSCs (18–20). However, some studies indicate that TNFR2 plays an important role in the activation of conventional T cells (Tcon cells) and CD8 T cells (21, 22). Furthermore, antibodies that can trigger the TNFR2 signal in vitro were shown to promote antitumor immune responses by activating CD8 T cells, Tcon cells, or NK cells (23). And consequently, TNFR2 agonists have been developed for the treatment of human cancers (19). Thus, the role of TNFR2 signal in orchestrating the inflammatory responses in autoimmune diseases or immune responses in cancers is complicated, and the mechanism may be involved in the activation of Tregs, MDSCs, CD8+ T cells and ADCC, or inversely, depletion of Tregs in tumor environment (summarized in Table 1). The contradictory pro-inflammatory and anti-inflammatory properties of TNFR2 signaling should be further clarified in future investigations. This contradictory pro-inflammatory and anti-inflammatory property of TNFR2 signaling should be further clarified in future investigation
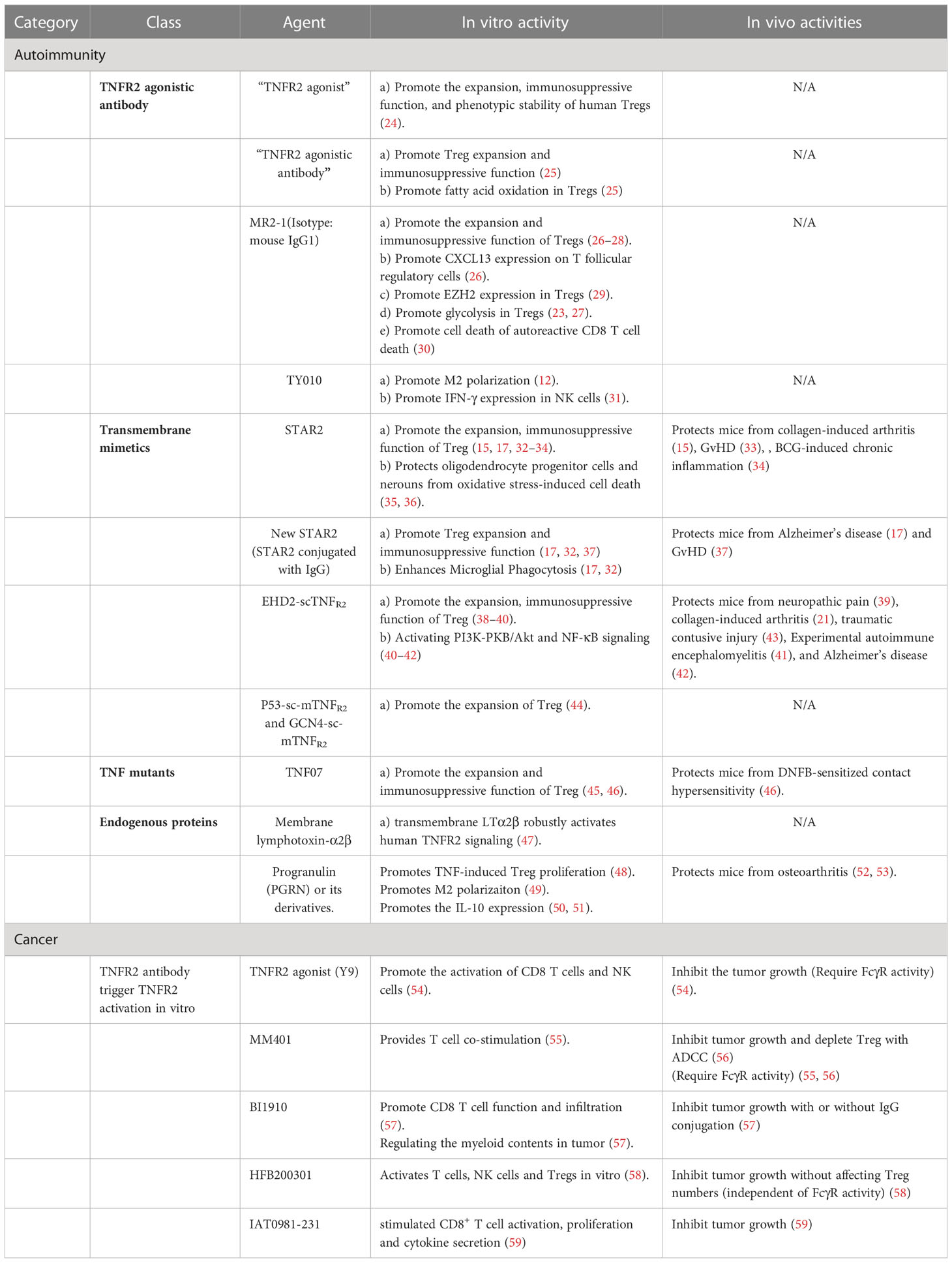
Table 1 The application of TNFR2 agonist in autoimmunity and cancer.
This review will focus on the discussion of the current understanding of the effects of TNFR2 agonists on inflammatory responses and anti-tumor immune responses. The development of TNFR2 agonists is introduced and the effect of these agents on the activation of different subsets of immune cells, and factors that may determine the therapeutic outcome of TNFR2 agonists in the treatment of cancer or autoimmune diseases, are reviewed and analyzed.
Overview of TNFR2 agonists
TNF mutants
Selective mutation of residues in TNF protein significantly altered its affinity to TNF receptors. The TNF mutant (D143N-A145R) is a TNFR2-selective agonist developed for decades. TNF mutant (D143N-A145R) only binds to TNFR2 but not TNFR1 (60). However, such a TNF mutant presents a 5~30 fold lower affinity to TNFR2 in comparison with wild-type (WT) TNF (61, 62) In recent years, several new TNF mutants that selectively bind and activate TNFR2 were developed by the phage display technique (63). The SPR analysis showed that these TNF mutants bind to TNFR2 with lower affinity but had a higher association/dissociation rate in contrast with WT TNF (63), indicating TNF mutants can form a stable complex with TNFR2Moreover, an in-vivo study showed that the TNF mutants fused with IgG could trigger the activation of TNFR2 signaling and induce Treg proliferation in a TNFR2-dependent manner (46). These results suggested that TNFR2-selective TNF mutants exhibit different binding modes for unique biological functions.
Transmembrane TNF mimetics
Compared with TNFR1, TNFR2 can only be fully activated by transmembrane TNF (64). The monomer transmembrane TNF always forms homotrimers on the cell membrane as a consequence of self-assembly before binding with TNFR2 (65, 66). Thus, one of the strategies to enhance the affinity of TNFR2-selective TNF mutants is to construct oligomerized TNFR2-selective TNF mutants (Figure 1). STAR2, a TNFR2 agonist composed of murine TNF mutants (D221N and A223R) and trimerization domain from chicken tenascin C, displays significantly higher affinity to TNFR2 than single chain TNF mutein and can induce TNFR2 activation more effectively (33). Moreover, STAR2 treatment significantly promoted Treg expansion in the mouse GVHD model (33). Based on this idea, Fisher et al. generated several oligomerized TNFR2-selective TNF mutants by using different oligomerization domains. The results showed that dodecavalent ligands by engineering oligomerization domain from GCN4 and TNFR2-selective TNF mutants (GCN4-sc-mTNFR2) displayed superior bioactivity and affinity than other oligomerized TNFR2-selective TNF mutants in vitro (44). Furthermore, GCN4-sc-mTNFR2 could be less immunogenic because the structure of GCN4-sc-mTNFR2 more resembles human protein structure (44). Although oligomerized TNFR2-selective TNF mutants represent a more effective strategy to evoke TNFR2 activation, the risk of immunogenicity cannot be neglected as the sequence of oligomerized TNFR2-selective TNF mutants cannot be found in nature. Thus, the immunogenicity of oligomerized TNFR2-selective TNF mutants should be carefully evaluated.
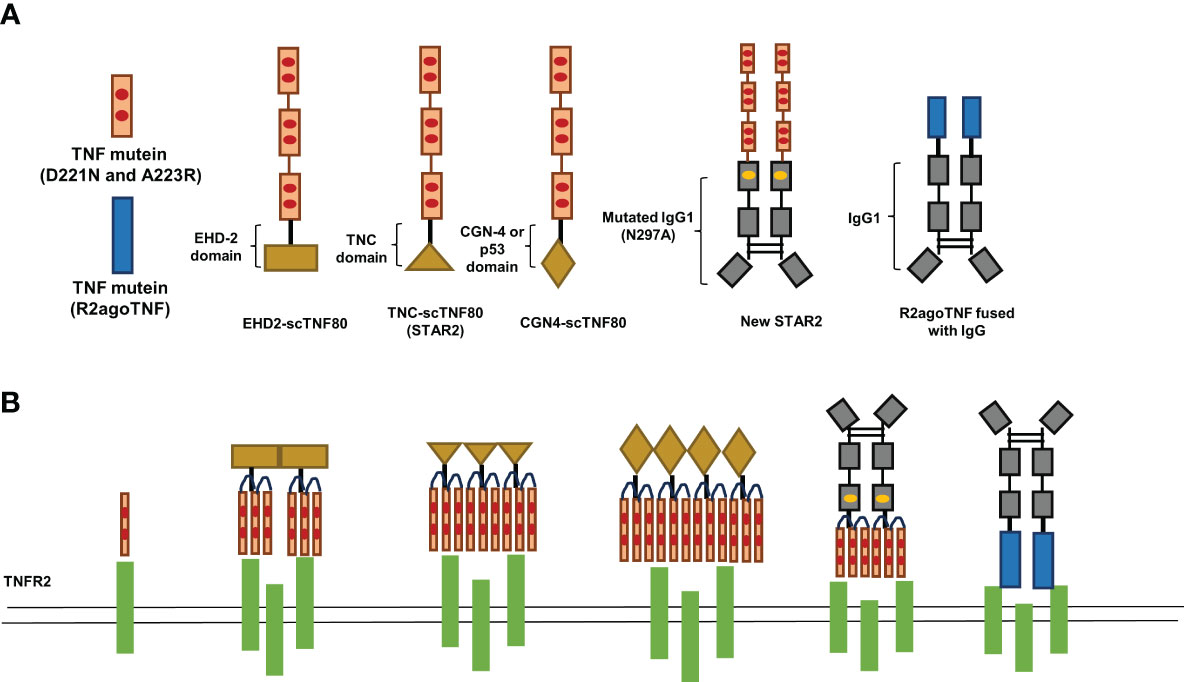
Figure 1 A schematic of TNF mutants and transmembrane TNF mimetics.
Progranulin (Endogenous proteins)
Progranulin (PGRN) is a secreted factor that regulates biological processes including inflammation, wound healing, and tissue repair (67). The initial results showed that progranulin directly interacts with and antagonizes both TNFR1 and TNFR2 (68), but subsequently, studies indicated that progranulin triggers the activation, instead of blocking TNF-TNFR2 signaling (69). Moreover, progranulin exhibits a relatively high affinity to TNFR2 (68). It has been shown that progranulin or its derivatives, Asttrin, alleviated the inflammatory responses in a TNFR2-dependent manner (50, 52, 70). However, there is contradictory evidence that progranulin may not interact with TNFRs, as progranulin failed to block the TNFR1-induced cell death (71). Furthermore, different concentrations of progranulin (2~200 ng/ml) plus IL-2 did not promote Treg proliferation, indicating that progranulin may not directly agonize TNFR2 (48). Thus, further investigation is needed to clarify if progranulin indeed promotes the activation of TNF-TNFR2 signaling.
TNFR2 agonistic antibodies
Recently, several TNFR2 antibodies with the capacity to activate TNFR2 in vitro have been developed for the treatment of autoimmune diseases (25) or cancers (57, 59, 72, 73). These TNFR2-stimulating antibodies were reportedly to possess either immunostimulatory or immunosuppressive in vivo, as its complicated nature, presumably based on different mechanism. For example, Y9, a close of agonistic anti-TNFR2 antibody, was found to be a competitive activator of TNFR2 and bind to CRD2 and CRD3 in TNFR2 (54). Other TNFR2-stimulating antibodies appeared to bind to CRD1 and CRD2 and did not compete with TNF for binding with TNFR2 (25, 57, 58). Whether the antibodies compete to bind to TNFR2 could also be important for their in vivo effect, as competitive activators sparing more TNF in local tissue, which may enhance the TNF-TNFR1 signaling. In contrast, the non-competitive TNFR2-stimulating antibodies did not affect TNF-TNFR2 interaction but may synergize with TNF in activating TNFR2 (74). Therefore, it is important to determine whether the non-competitive activator could elicit different responses with competitive activators.
Fragment crystallizable region (Fc) is another factor that profoundly affects the function of TNFR2-stimulating antibodies. For example, TNFR2 agonists may require FcγR activity for more potent agonistic function, as it confers TNFR2 agonist the transmembrane TNF-like activity (75). This was evidenced by the observation that TNFR2 agonistic antibody (BI-1910) with poor FcγR-binding activity (N297A, IgG1 mutated) exhibited a decreased antitumor effect as compared to IgG1 or IgG2a conjugated BI-1910 (57). Therefore, the engagement of FcγR could be an important factor that determine the therapeutic effect of some TNFR2 agonistic antibodies (76). Moreover, Fc conjugation can induce the antibody-dependent cellular cytotoxicity (ADCC) or antibody-dependent cellular phagocytosis (ADCP). For example, the antitumor effect of Y9 was reportedly dependent on the ADCC and ADCP, as its antitumor effect was diminished in Fcgr2b−/− and Fcer1g−/− mice (54). In this case, Y9 may deplete TNFR2-expressing cells rather than a TNFR2 agonist. Nevertheless, a recent study shows that a TNFR2 agonistic antibody had a more potent function in the presence of antibody that can block crosslinking activities (25), suggesting that neither ADCC nor Fc-mediated crosslink activities were required for the activity of this TNFR2 agonistic antibody.
TNFR2 agonist-induced activation of immune suppressive cells
CD4+Foxp3+ regulatory T cells
There is compelling evidence that TNFR2 plays a pivotal role in Treg activation, function, proliferation, and phenotypic stability (77, 78). Several TNFR2 agonists have been reported to alleviate inflammatory responses by promoting Treg function and expansion (15, 33, 34, 39, 40). TNFR2 agonism also represents an efficient approach to expand the Tregs from low-purity human Tregs for adoptive Treg transfer therapy. Several studies showed that TNFR2 agonistic antibody plus the standard Treg expansion protocol (in the presence of CD3/CD28, IL-2 with or without rapamycin) resulted in the expansion of homogenous stable Tregs with potent immunosuppressive function (24, 25, 33). Moreover, TNFR2 agonistic antibody treated-Treg has lower expression of CD127, IL-17A, and IFN-γ, indicating TNFR2 agonistic antibody help maintain the phenotypic stability of expanded Tregs (79). It is of great interest to examine whether TNFR2-agonist-expanded Tregs are more effective for adoptive Treg transfer therapy.
Myeloid-derived suppressor cells
MDSCs, a subset derives from pathologically activated neutrophils or monocytes, have potent immunosuppressive activity. MDSCs are considered a potential target for the treatment of cancer and autoimmune diseases (80). It was reported that TNF-TNFR2 signaling is important for the recruitment, immunosuppressive function, and survival of MDSCs (81–83). Thus, MDSCs is also a potential target of TNFR2 agonist. However, recent studies showed that the effect of TNFR2 agonists is mainly mediated by CD4 T cells, albeit with a minor effect on MDSCs. Lamontain and colleagues reported that TNFR2 agonist (TNCscTNF80) treatment promoted the expansion of MDSCs in bone marrow, but not in spleen and lymph nodes in the mouse CIA model, indicating the anti-inflammatory effect of TNCscTNF is not dependent on MDSCs (15). This result was further supported by the data from CD4creTNFR2fl/fl and LysMcreTNFR2fl/fl mice. TNFR2 agonist (TNCscTNF80) suppresses T cell proliferation in LysMcreTNFR2fl/fl mice but not in TNFR2-/- mice and CD4creTNFR2fl/fl (34), indicating the anti-inflammatory effect of TNFR2 agonist were mainly dependent on the TNFR2 expression by CD4 T cells. Further evidence is needed to support the claim that TNFR2 agonists can boost MDSC’s activity to suppress inflammatory responses.
Monocytes/macrophages
Monocytes and macrophages express both TNFR1 and TNFR2. These two receptors play complicated roles in the regulation of the viability, function, and recruitment of monocytes/macrophages (84). Moreover, tissue-resident macrophages may also have different responses to TNFR2 activation, as compared with circulating monocytes/macrophages (85). Thus, the effect of TNFR2 agonism on macrophages could be tissue specific. It has been shown that administration of TNFR2 agonist (EHD2-sc-mTNFR2) increased the expression of M2 markers in macrophages and macroglia, and reduced the expression of M1 markers, but without activation of macrophage in mouse central nervous system (16, 39). However, these effects of TNFR2 agonists could be the indirect result of the activated Tregs which may suppress macrophage activity (86). A recent study showed that TNFR2 agonist (TY010) promoted M2 polarization of bone marrow-derived macrophage in TNFR2 dependent manner, indicating TNFR2 agonist may directly activate TNFR2 on macrophage and induced an immunosuppressive phenotype (49). Moreover, TNFR2 agonist (NewStar2, TNCscTNF80 fused with mutated human IgG) has been shown to enhance the phagocytosis of microglia and promote the clearance of Aβ plaques, which contributes to the alleviation of Alzheimer’s diseases in mouse (17). These results indicate that macrophages/monocytes are the targets of TNFR2 agonists in the treatment of inflammatory diseases.
TNFR2 agonist-induced activation of effector immune cells
Conventional T cells
TNFR2 has been shown to promote the activation, function, differentiation, and proliferation of Tcon cells (22, 87, 88). TNFR2+ Tcon cells are more resistant to Treg-mediated immunosuppression (87). However, TNFR2 was expressed much lower by Tcon cells than by Tregs in the resting state (87). Thus, TNFR2 agonists may not effectively activate TNFR2 signaling in unstimulated Tcon cells (89). Previous studies showed that the treatment with TNFR2 agonists (TNF mutants or transmembrane mimetics) inhibited Tcon cell proliferation by promoting Treg expansion (32). However, TNFR2 expression can also be upregulated by TCR stimulation (90) or pro-inflammatory cytokines (91), suggesting the activated Tcon cells could respond to TNFR2 agonists. A recent study showed that stimulation of anti-CD3 and a TNFR2 agonistic antibody (MR2-1) induces a similar alteration of transcriptome profile, albeit the alteration of the Treg cell transcriptomic profile is more obvious (23). This effect of TNFR2 agonism has shown to be pathological-relevant, as TNFR2 is expressed by tumor-infiltrating Tcon cells (89, 91–93) and proinflammatory subsets of CD4 Tcon cells (94, 95), suggesting that TNFR2 agonists could induce the activation of TNFR2 signal in Tcon cells in tumor and inflammatory diseases. In the mouse tumor model, it was reported that TNFR2 agonistic antibodies induced the expansion of CD4+ Tcon cells without affecting the Treg number in vivo (58). Therefore, Tcon cells are also a potential target of TNFR2 agonists albeit with relatively lower TNFR2 expression.
CD8 T cell
As one of the co-costimulatory receptors, TNFR2 promotes the activation, function, proliferation, differentiation, and recruitment of CD8 T cells (96–98). However, TNFR2 signaling can also play a dual role in the modulation of the activation of CD8 T cells. For example, genetic ablation of TNFR2 impairs the production of effector cytokine while can also result in a more persistent activation of CD8 cells in mouse tumor and infection models (99, 100). CD8 T cells at different stages of activation may likely respond to TNFR2 activation differently. This notion is supported by the observation that the activation of TNFR2 promotes the differentiation of naïve CD8 T cells (96), while TNFR2 stimulation also selectively induced the activation-induced cell death (AICD) of the autoreactive CD8 T cells without significantly affecting the other T cell subsets (30). The different responses could be attributable to the alteration of downstream signaling. TNFR2 expression is crucial for the activation of NF-κB signaling in CD8 T cells when stimulated with anti-CD3/CD28. While a persistent activation of TNFR2 can inhibit NF-κB signaling through depleting TRAF2, an important signal component in mediating NF-κB activation (39), thereby sensitizing CD8 T cells to TNF-induced cell death (101).
Although treatment of TNFR2 agonistic antibody can stimulate the activation of tumor-infiltrating CD8 T cells (57), the time frame of TNFR2 agonist treatment in a preclinical mouse tumor model is relatively short. Such studies may not be able to reflect the effect of long-term activation of TNFR2, including activation-induced cell death (AICD) or exhaustion of CD8 T cells (100, 102). More recently, we reported that TNFR2 expression is associated with the exhaustive phenotype of CD8 T cells in human cancers (103). Thus, the role of TNFR2 in tumor-infiltrating CD8 CTLs is complex and needs further investigation. A more thorough understanding of the molecular basis underlying the effect of TNFR2 signal in CD8 CTLs is crucial to device TNFR2 agonists in tumor immunotherapy.
Natural killer cells
TNFR2 has been reported to be expressed by both human and mouse NK cells, albeit the expression pattern could be different (89). It has been shown that genetic ablation of TNFR2 has been shown to significantly decrease the expression of IFN-γ in α-galactosylceramide (α-GalCer)-treated mouse, indicating that TNFR2 signaling is also important for the activation and function of NK cells (31). TNF or TNFR2 agonist (TY010) in concert with IL-12 elevated the expression of IFN-γ in human and mouse NK cells in vitro (31, 104). The antitumor effect of TNFR2-targetingantibody (Y9) can be impaired by the depletion of NK cells, suggesting that TNFR2 agonists may also target the NK cells to elicit antitumor immune responses (54). Now several TNFR2-stimulating antibodies in clinical development have been reported to enhance NK cell activation (58, 73). However, the mechanism that how TNFR2 agonism affects tumor-infiltrating NK cells remains to be investigated.
Factors may determine the therapeutic outcome of TNFR2 agonists in the treatment of cancer or autoimmune diseases
TNFR2 expression: pro-inflammatory vs anti-inflammatory cell subsets
TNFR2 signaling can result in both anti-inflammatory or pro-inflammatory effects, depending on the cell type of TNFR2 expression and the functional status of the cells. High levels of TNFR2 are constitutively expressed by Tregs, and the activation of TNFR2 signaling in Tregs or autoreactive CD8 T cells (and other immunosuppressive cells) can cause immune suppression or elicit an anti-inflammatory effect (30, 78). However, elevated TNFR2 expression can be shown in pathogenic T cell subsets in patients with Crohn’s disease (94) and rheumatoid arthritis (95), and blockade of TNF promoted cell death of pathogenic T cells (94, 95). These results indicated that TNFR2 agonism may also promote pathogenic T-cell responses. The pathogenic T cells with elevated TNFR2 expression can be more resistant to Treg-mediated immune suppression (105). Moreover, activation of TNFR2 also promotes the inflammatory responses of innate immune cells and non-immune cells (106–108). The dual role or bi-phasic effect of TNFR2 signaling is exempted by a study that shows that, in TNFR1 deficient mice, infusion of murine TNF at the initial phase of collagen-induced arthritis increased the disease severity, while the same treatment markedly alleviated the inflammation in the progression phase (109). This study demonstrated the bi-phasic effects of TNF-TNFR2 signaling in an inflammatory response.
Both humanized TNFR2 antagonists and agonists have been developed for the treatment of tumors (19, 110), based on the notion that antagonistic antibodies may eliminate the immunosuppressive Tregs, while antibodies that trigger TNFR2 signaling may activate CD8 CTLs and NK cells (54, 56–58, 72). Despite the assumptions are opposite, the results of the studies appear to support that TNFR2 antagonistic antibodies inhibit the tumor infiltration of Tregs and consequently enhance the antitumor immune responses (111–119), while antibodies that trigger the TNFR2 signal in vitro also elicited antitumor immune responses (54, 56–58, 72). It was shown that TNFR2 activation promotes the differentiation and the production of effector cytokine by CD8 T cells (96, 120). Thus, as a costimulatory molecule, TNFR2 is likely to promote the initiation of antitumor T cell immune responses. In line with this notion, preclinical studies showed that TNFR2 agonistic antibodies with diminished activity to induce ADCC can enhance the antitumor immune responses by activating CD8 T cells and NK cells. However, it should be noted that the preclinical studies about TNFR2 agonistic antibodies were based on the transplanted tumor models and the time of experiment settings is relatively short. Such studies may not be able to reflect the effect of long-term activation of TNFR2, including activation-induced cell death (AICD) or exhaustion of CD8 T cells (100, 102).
Ligand-based agonists vs agonistic antibodies
Different types of TNFR2 agonists may own distinguished features and consequently elicit different effects on immune response in vivo. We summarized the antibodies that can trigger TNFR2 signaling from the published studies (Table 1). Interestingly, some of these TNFR2 antibodies can enhance the anti-tumor immune responses even though they have been shown to significantly promote Treg expansion in vitro. While the TNFR2-selective TNF mutant or transmembrane TNF mimetics elicited anti-inflammatory responses in vitro and vivo. The paradoxical effects of antibodies were also reported in other TNFRSF members. For example, one of the GITR antibody has been shown to promote Treg expansion in vitro (121). However, this GITR antibody has also been shown to enhance the infiltration of non-Foxp3 expressing T cells into tumor tissue and enhance the antitumor immune responses (122). These results suggest the complicated nature of TNFR2-antibodies with stimulating activities. By comparing with transmembrane TNF mimetics, TNFR2-antibodies with agonistic activities also bind with TNFR2 with high affinity. However, the function of TNFR2-stimulating antibodies is significantly affected by the Fc region as aforementioned. Moreover, although there is compelling evidence that TNFR2 agonists promote the expansion of Tregs, excessive or prolonged TNFR2 activation may elicit different responses of Tregs in some circumstances. For example, tumor-infiltrating Tregs are highly activated and express high levels of TNFR2. The activation of TNFR2 may induce the TRAF2 depletion in Tregs present in the tumor environment, thus sensitizing Treg to TNF-induced cell death (123, 124). Moreover, although optimal PI3K/Akt/mTOR signaling could be important for TNF-induced Treg activation and expansion (26, 28), a high dose of TNFR2 agonistic antibody or superclustering of TNFR2 may also induce hyperactivation of PI3K/Akt/mTOR signaling pathway, which can destabilize Foxp3 expression (125). The agonistic antibody-induced Foxp3 instability and Treg cell death have been reported in other TNFRSF members with a similar co-stimulatory capacity (126, 127). Therefore, this evidence may provide an alternative interpretation of the reported anti-tumor effect of TNFR2 agonistic antibodies.
Tissue-specific responses to TNFR2 agonists
The TNFR2 agonist-regulated immune responses could be tissue-specific, as the tissue-resident immune cells and non-immune cells can express TNFR2. TNFR2 activation may induce anti-inflammatory responses or pro-inflammatory responses as well, depending on the cell types targeted. There is compelling evidence that TNFR2 signaling not only suppresses the inflammatory responses in the central nervous system (CNS) but also promotes tissue repair of the CNS system by activating TNFR2 signaling through several different cell types (35, 128, 129). Thus, TNFR2 agonism evokes immunosuppression and tissue repair in the CNS. However, TNFR2 also induced potent inflammatory responses in some organs. For example, activation of TNFR2 signaling in parenchymal cells was important for the development of hepatitis in mice (130). Blockade of TNFR2 has been shown to alleviate anti-PD-1-induced hepatic inflammation in mouse hepatic carcinoma, even infiltration of Tregs was also decreased (131). Moreover, TNFR2 signaling in some tumor cell types also contribute to tumor progression (132, 133), so the potentially undesirable effect of TNFR2 agonistic antibody should be considered. Thus, tissue-specific responses are also an important factor that needs to be considered in TNFR2 agonist therapy.
Conclusion and future perspective
Numerous studies indicated the potential of TNFR2 agonism in the treatment of autoimmune inflammatory diseases and cancer, as TNFR2 has a dual role in modulating immune responses. TNFR2 plays a decisive role in maintaining Treg function and activity, which is important for the suppression of autoimmune inflammatory responses. On the other hand, TNFR2 activation in Tcon cells or CD8 T cells also elicits pro-inflammatory responses. However, the therapeutic outcome of TNFR2 agonism could be affected by the property of agonists, the disease condition, and tissue-specific responses. To minimize the unwanted effect elicited by TNFR2 agonism, one of the strategies is to develop therapeutics that specifically target certain cell types. For example, IL2-EHD2-sc-mTNFR2, a recombinant protein fused with TNFR2 agonist (EHD2-sc-mTNFR2) and IL-2, induced a more potent Treg expansion than IL2 plus EHD2-sc-mTNFR2 (38). Moreover, combining the immunosuppressants with TNFR2 agonists could be another strategy in the treatment of autoimmune inflammatory diseases, as Tregs are more resistant to immunosuppressant-mediated cell death (134). Although several TNFR2 agonistic antibodies were demonstrated in the clinical trial, how the TNFR2 agonistic antibody elicits a different immune response in vivo remains largely unknown. Identifying the factors that affect biological consequences induced by TNFR2 agonists may pave the way to the more effective treatment of cancer or autoimmune diseases.
Data availability statement
The original contributions presented in the study are included in the article/supplementary material. Further inquiries can be directed to the corresponding author.
Author contributions
YC and MJ drafted the manuscript. XC, the corresponding author, was involved in designing the frame of the manuscript and approving the final version to be published. All authors contributed to the article and approved the submitted version.
Funding
This project has been funded by The Science and Technology Development Fund, Macau SAR (FDCT, File No. 0099/2021/A2, 0007/2022/AKP), University of Macau (File No. CPG2023-00025-ICMS, MYRG2022-00260-ICMS, SKL-QRCM(UM)-2020-2022), the 2020 Guangdong Provincial Science and Technology Innovation Strategy Special Fund (EF006/ICMS-CX/2021/GDSTC), and the Research Institute of Tsinghua, Pearl River Delta (EF032/ICMS-CX/2021/RITH).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Brenner D, Blaser H, Mak TW. Regulation of tumour necrosis factor signalling: live or let die. Nat Rev Immunol (2015) 15(6):362–74. doi: 10.1038/nri3834
2. Ramos-Casals M, Roberto Perez A, Diaz-Lagares C, Cuadrado MJ, Khamashta MA, Group BS, et al. Autoimmune diseases induced by biological agents: a double-edged sword? Autoimmun Rev (2010) 9(3):188–93. doi: 10.1016/j.autrev.2009.10.003
3. Cui G, Fan Q, Li Z, Goll R, Florholmen J. Evaluation of anti-TNF therapeutic response in patients with inflammatory bowel disease: Current and novel biomarkers. EBioMedicine (2021) 66:103329. doi: 10.1016/j.ebiom.2021.103329
4. Silva-Fernandez L, Hyrich K. Rheumatoid arthritis: When TNF inhibitors fail in RA–weighing up the options. Nat Rev Rheumatol (2014) 10(5):262–4. doi: 10.1038/nrrheum.2014.34
5. Subedi S, Gong Y, Chen Y, Shi Y. Infliximab and biosimilar infliximab in psoriasis: efficacy, loss of efficacy, and adverse events. Drug Des Devel Ther (2019) 13:2491–502. doi: 10.2147/DDDT.S200147
6. Sicotte NL, Voskuhl RR. Onset of multiple sclerosis associated with anti-TNF therapy. Neurology (2001) 57(10):1885–8. doi: 10.1212/WNL.57.10.1885
7. Aggarwal BB. Signalling pathways of the TNF superfamily: a double-edged sword. Nat Rev Immunol (2003) 3(9):745–56. doi: 10.1038/nri1184
8. Gupta R, Kapuria V, Das SN. Single nucleotide polymorphisms in TNF-alpha, TNFR2 gene and TNF-alpha production in Asian Indians. Immunol Invest (2009) 38(3-4):240–54. doi: 10.1080/08820130902766563
9. Li H, Anderson SK. Association of TNFRSF1B promoter polymorphisms with human disease: further studies examining T-regulatory cells are required. Front Immunol (2018) 9:443. doi: 10.3389/fimmu.2018.00443
10. Naserian S, Abdelgawad ME, Afshar Bakshloo M, Ha G, Arouche N, Cohen JL, et al. The TNF/TNFR2 signaling pathway is a key regulatory factor in endothelial progenitor cell immunosuppressive effect. Cell Commun Signal (2020) 18(1):94. doi: 10.1186/s12964-020-00564-3
11. Tseng WY, Huang YS, Clanchy F, McNamee K, Perocheau D, Ogbechi J, et al. TNF receptor 2 signaling prevents DNA methylation at the Foxp3 promoter and prevents pathogenic conversion of regulatory T cells. Proc Natl Acad Sci U.S.A. (2019) 116(43):21666–72. doi: 10.1073/pnas.1909687116
12. Atretkhany KN, Mufazalov IA, Dunst J, Kuchmiy A, Gogoleva VS, Andruszewski D, et al. Intrinsic TNFR2 signaling in T regulatory cells provides protection in CNS autoimmunity. Proc Natl Acad Sci U.S.A. (2018) 115(51):13051–6. doi: 10.1073/pnas.1807499115
13. Leclerc M, Naserian S, Pilon C, Thiolat A, Martin GH, Pouchy C, et al. Control of GVHD by regulatory T cells depends on TNF produced by T cells and TNFR2 expressed by regulatory T cells. Blood (2016) 128(12):1651–9. doi: 10.1182/blood-2016-02-700849
14. Chen S, Lin Z, Xi L, Zheng Y, Zhou Q, Chen X. Differential role of TNFR1 and TNFR2 in the development of imiquimod-induced mouse psoriasis. J Leukoc Biol (2021) 110(6):1047–55. doi: 10.1002/JLB.2MA0121-082R
15. Lamontain V, Schmid T, Weber-Steffens D, Zeller D, Jenei-Lanzl Z, Wajant H, et al. Stimulation of TNF receptor type 2 expands regulatory T cells and ameliorates established collagen-induced arthritis in mice. Cell Mol Immunol (2019) 16(1):65–74. doi: 10.1038/cmi.2017.138
16. Fischer R, Padutsch T, Bracchi-Ricard V, Murphy KL, Martinez GF, Delguercio N, et al. Exogenous activation of tumor necrosis factor receptor 2 promotes recovery from sensory and motor disease in a model of multiple sclerosis. Brain Behav Immun (2019) 81:247–59. doi: 10.1016/j.bbi.2019.06.021
17. Orti-Casan N, Zuhorn IS, Naude PJW, De Deyn PP, van Schaik PEM, Wajant H, et al. A TNF receptor 2 agonist ameliorates neuropathology and improves cognition in an Alzheimer's disease mouse model. Proc Natl Acad Sci U.S.A. (2022) 119(37):e2201137119. doi: 10.1073/pnas.2201137119
18. Fischer R, Kontermann RE, Pfizenmaier K. Selective targeting of TNF receptors as a novel therapeutic approach. Front Cell Dev Biol (2020) 8:401. doi: 10.3389/fcell.2020.00401
19. Quazi S. TNFR2 antagonist and agonist: a potential therapeutics in cancer immunotherapy. Med Oncol (2022) 39(12):215. doi: 10.1007/s12032-022-01772-2
20. Yang S, Wang J, Brand DD, Zheng SG. Role of TNF-TNF receptor 2 signal in regulatory T cells and its therapeutic implications. Front Immunol (2018) 9:784. doi: 10.3389/fimmu.2018.00784
21. Chen X, Nie Y, Xiao H, Bian Z, Scarzello AJ, Song N-Y, et al. TNFR2 expression by CD4 effector T cells is required to induce full-fledged experimental colitis. Sci Rep (2016) 6(1):32834. doi: 10.1038/srep32834
22. Alam MS, Otsuka S, Wong N, Abbasi A, Gaida MM, Fan Y, et al. TNF plays a crucial role in inflammation by signaling via T cell TNFR2. Proc Natl Acad Sci U.S.A. (2021) 118(50). doi: 10.1073/pnas.2109972118
23. Mensink M, Tran TNM, Zaal EA, Schrama E, Berkers CR, Borst J, et al. TNFR2 costimulation differentially impacts regulatory and conventional CD4+ T-cell metabolism. Front Immunol (2022) 13. doi: 10.3389/fimmu.2022.881166
24. Okubo Y, Mera T, Wang L, Faustman DL. Homogeneous expansion of human T-regulatory cells via tumor necrosis factor receptor 2. Sci Rep (2013) 3:3153. doi: 10.1038/srep03153
25. Torrey H, Kuhtreiber WM, Okubo Y, Tran L, Case K, Zheng H, et al. A novel TNFR2 agonist antibody expands highly potent regulatory T cells. Sci Signal (2020) 13(661). doi: 10.1126/scisignal.aba9600
26. Kawano S, Mitoma H, Inokuchi S, Yamauchi Y, Yokoyama K, Nogami J, et al. TNFR2 signaling enhances suppressive abilities of human circulating T follicular regulatory cells. J Immunol (2022) 208(5):1057–65. doi: 10.4049/jimmunol.2100323
27. de Kivit S, Inoue M, Kamada H, Taki S, Furuya T, Abe Y, et al. Stable human regulatory T cells switch to glycolysis following TNF receptor 2 costimulation. Nat Metab (2020) 2(10):1046–61. doi: 10.1038/s42255-020-00271-w
28. Okuzono Y, Muraki Y, Sato S. TNFR2 pathways are fully active in cancer regulatory T cells. Biosci Biotechnol Biochem (2022) 86(3):351–61. doi: 10.1093/bbb/zbab226
29. Urbano PCM, Koenen HJPM, Joosten I, He X. An autocrine TNFalpha-tumor necrosis factor receptor 2 loop promotes epigenetic effects inducing human Treg stability in vitro. Front Immunol (2018) 9:573. doi: 10.3389/fimmu.2018.00573
30. Ban L, Zhang J, Wang L, Kuhtreiber W, Burger D, Faustman DL, et al. Selective death of autoreactive T cells in human diabetes by TNF or TNF receptor 2 agonism. Proc Natl Acad Sci (2008) 105(36):13644–9. doi: 10.1073/pnas.0803429105
31. Almishri W, Santodomingo-Garzon T, Le T, Stack D, Mody CH, Swain MG, et al. TNFalpha augments cytokine-induced NK cell IFNgamma production through TNFR2. J Innate Immun (2016) 8(6):617–29. doi: 10.1159/000448077
32. Vargas JG, Wagner J, Shaikh H, Lang I, Medler J, Anany M, et al. A TNFR2-specific TNF fusion protein with improved in vivo activity. Front Immunol (2022) 13. doi: 10.3389/fimmu.2022.888274
33. Chopra M, Biehl M, Steinfatt T, Brandl A, Kums J, Amich J, et al. Exogenous TNFR2 activation protects from acute GvHD via host T reg cell expansion. J Exp Med (2016) 213(9):1881–900. doi: 10.1084/jem.20151563
34. Schmid T, Falter L, Weber S, Muller N, Molitor K, Zeller D, et al. Chronic inflammation increases the sensitivity of mouse Treg for TNFR2 costimulation. Front Immunol (2017) 8:1471. doi: 10.3389/fimmu.2017.01471
35. Fischer R, Maier O, Siegemund M, Wajant H, Scheurich P, Pfizenmaier K, et al. A TNF receptor 2 selective agonist rescues human neurons from oxidative stress-induced cell death. PloS One (2011) 6(11):e27621. doi: 10.1371/journal.pone.0027621
36. Maier O, Fischer R, Agresti C, Pfizenmaier K. TNF receptor 2 protects oligodendrocyte progenitor cells against oxidative stress. Biochem Biophys Res Commun (2013) 440(2):336–41. doi: 10.1016/j.bbrc.2013.09.083
37. Vargas JG, Kanzler C, Scheller L, Kreckel J, Shaikh H, Lang I, et al. Harnessing the expression of TNFR2 on Tregs to prevent Agvhd. Blood (2022) 140(Supplement 1):4465–6. doi: 10.1182/blood-2022-168592
38. Padutsch T, Sendetski M, Huber C, Peters N, Pfizenmaier K, Bethea JR, et al. Superior Treg-expanding properties of a novel dual-acting cytokine fusion protein. Front Pharmacol (2019) 10:1490. doi: 10.3389/fphar.2019.01490
39. Fischer R, Sendetski M, Del Rivero T, Martinez GF, Bracchi-Ricard V, Swanson KA, et al. TNFR2 promotes Treg-mediated recovery from neuropathic pain across sexes. Proc Natl Acad Sci U.S.A. (2019) 116(34):17045–50. doi: 10.1073/pnas.1902091116
40. Fischer R, Proske M, Duffey M, Stangl H, Martinez GF, Peters N, et al. Selective activation of tumor necrosis factor receptor II induces antiinflammatory responses and alleviates experimental arthritis. Arthritis Rheumatol (2018) 70(5):722–35. doi: 10.1002/art.40413
41. Fiedler T, Fairless R, Pichi K, Fischer R, Richter F, Kontermann RE, et al. Co-modulation of TNFR1 and TNFR2 in an animal model of multiple sclerosis. J Neuroinflamm (2023) 20(1):100. doi: 10.1186/s12974-023-02784-z
42. Dong Y, Fischer R, Naudé PJW, Maier O, Nyakas C, Duffey M, et al. Essential protective role of tumor necrosis factor receptor 2 in neurodegeneration. Proc Natl Acad Sci U.S.A. (2016) 113(43):12304–9. doi: 10.1073/pnas.1605195113
43. Gerald MJ, Bracchi-Ricard V, Ricard J, Fischer R, Nandakumar B, Blumenthal GH, et al. Continuous infusion of an agonist of the tumor necrosis factor receptor 2 in the spinal cord improves recovery after traumatic contusive injury. CNS Neurosci Ther (2019) 25(8):884–93. doi: 10.1111/cns.13125
44. Fischer R, Marsal J, Gutta C, Eisler SA, Peters N, Bethea JR, et al. Novel strategies to mimic transmembrane tumor necrosis factor-dependent activation of tumor necrosis factor receptor 2. Sci Rep (2017) 7(1):6607. doi: 10.1038/s41598-017-06993-4
45. Ban L, Kuhtreiber W, Butterworth J, Okubo Y, Vanamee ES, Faustman DL. Strategic internal covalent cross-linking of TNF produces a stable TNF trimer with improved TNFR2 signaling. Mol Cell Ther (2015) 3:7. doi: 10.1186/s40591-015-0044-4
46. Inoue M, Yamashita K, Tsuji Y, Miki M, Amano S, Okumura T, et al. Characterization of a TNFR2-selective agonistic TNF-alpha mutant and its derivatives as an optimal regulatory T cell expander. J Immunol (2021) 206(8):1740–51. doi: 10.4049/jimmunol.2000871
47. Kucka K, Lang I, Zhang T, Siegmund D, Medler J, Wajant H, et al. Membrane lymphotoxin-alpha(2)beta is a novel tumor necrosis factor (TNF) receptor 2 (TNFR2) agonist. Cell Death Dis (2021) 12(4):360. doi: 10.1038/s41419-021-03633-8
48. Hu Y, Xiao H, Shi T, Oppenheim JJ, Chen X, et al. Progranulin promotes tumour necrosis factor-induced proliferation of suppressive mouse CD4(+) Foxp3(+) regulatory T cells. Immunology (2014) 142(2):193–201. doi: 10.1111/imm.12241
49. Fu W, Hu W, Yi Y-S, Hettinghouse A, Sun G, Bi Y, et al. TNFR2/14-3-3ϵ signaling complex instructs macrophage plasticity in inflammation and autoimmunity. J Clin Invest (2021) 131(16). doi: 10.1172/JCI144016
50. Wei F, Zhang Y, Jian J, Mundra JJ, Tian Q, Lin J, et al. PGRN protects against colitis progression in mice in an IL-10 and TNFR2 dependent manner. Sci Rep (2014) 4:7023. doi: 10.1038/srep07023
51. Fu W, Hu W, Shi L, Mundra JJ, Xiao G, Dustin ML, et al. Foxo4- and Stat3-dependent IL-10 production by progranulin in regulatory T cells restrains inflammatory arthritis. FASEB J (2017) 31(4):1354–67. doi: 10.1096/fj.201601134R
52. Wei J, Wang K, Hettinghouse A, Liu C. Atsttrin promotes cartilage repair primarily through TNFR2-Akt pathway. Front Cell Dev Biol (2020) 8:577572. doi: 10.3389/fcell.2020.577572
53. Wei JL, Fu W, Ding Y-j, Hettinghouse A, Lendhey M, Schwarzkopf R, et al. Progranulin derivative Atsttrin protects against early osteoarthritis in mouse and rat models. Arthritis Res Ther (2017) 19(1):280. doi: 10.1186/s13075-017-1485-8
54. Tam EM, Fulton B, Sampson JF, Muda M, Camblin A, Richards J, et al. Antibody-mediated targeting of TNFR2 activates CD8+ T cells in mice and promotes antitumor immunity. Sci Trans Med (2019) 11(512):eaax0720. doi: 10.1126/scitranslmed.aax0720
55. Richards J, Wong C, Koshkaryev A, Fulton R, Camblin A, Sampson J, et al. Abstract 4846: MM-401, a novel anti-TNFR2 antibody that induces T cell co-stimulation, robust anti-tumor activity and immune memory. Cancer Res (2019) 79(13_Supplement):4846–6. doi: 10.1158/1538-7445.AM2019-4846
56. Sampson JF, Kurella VB, Paragas V, Kumar S, Lulo JE, Qiu JA, et al. Abstract 555: A novel human TNFR2 antibody (MM-401) modulates T cell responses in anti-cancer immunity. Cancer Res (2019) 79(13_Supplement):555–5. doi: 10.1158/1538-7445.AM2019-555
57. Mårtensson L, Cleary K, Semmrich M, Kovacek M, Holmkvist P, Svensson C, et al. Abstract 936: Targeting TNFR2 for cancer immunotherapy: Ligand blocking depletors versus receptor agonists. Cancer Res (2020) 80(16_Supplement):936–6. doi: 10.1158/1538-7445.AM2020-936
58. Wei S, Fulton R, Lu Y-Y, Zhang Q, Zhou H, Raue A, et al. Abstract 1883: Mechanism of action and biomarker strategy for HFB200301, an anti-TNFR2 agonist antibody for the treatment of cancer. Cancer Res (2021) 81(13_Supplement):1883–3. doi: 10.1158/1538-7445.AM2021-1883
59. Jiang X, Wu C, Yin L. Abstract 5549: First-in-class TNFR2 agonist & antagonist antibodies as Treg-Teff modulator for cancer immunotherapy. Cancer Res (2022) 82(12_Supplement):5549–9. doi: 10.1158/1538-7445.AM2022-5549
60. Loetscher H, Stueber D, Banner D, Mackay F, Lesslauer W. Human tumor necrosis factor alpha (TNF alpha) mutants with exclusive specificity for the 55-kDa or 75-kDa TNF receptors. J Biol Chem (1993) 268(35):26350–7. doi: 10.1016/S0021-9258(19)74322-1
61. Rahman MM, Barrett JW, Brouckaert P, McFadden G. Variation in ligand binding specificities of a novel class of poxvirus-encoded tumor necrosis factor-binding protein. J Biol Chem (2006) 281(32):22517–26. doi: 10.1074/jbc.M604645200
62. Yui J, Hemmings D, Garcia-Lloret M, Guilbert LJ. Expression of the human p55 and p75 tumor necrosis factor receptors in primary villous trophoblasts and their role in cytotoxic signal transduction. Biol Reprod (1996) 55(2):400–9. doi: 10.1095/biolreprod55.2.400
63. Ando D, Inoue M, Kamada H, Taki S, Furuya T, Abe Y, et al. Creation of mouse TNFR2-selective agonistic TNF mutants using a phage display technique. Biochem Biophys Rep (2016) 7:309–15. doi: 10.1016/j.bbrep.2016.06.008
64. Grell M, Douni E, Wajant H, Lohden M, Clauss M, Maxeiner B, et al. The transmembrane form of tumor necrosis factor is the prime activating ligand of the 80 kDa tumor necrosis factor receptor. Cell (1995) 83(5):793–802. doi: 10.1016/0092-8674(95)90192-2
65. Locksley RM, Killeen N, Lenardo MJ. The TNF and TNF receptor superfamilies: integrating mamMalian biology. Cell (2001) 104(4):487–501. doi: 10.1016/S0092-8674(01)00237-9
66. Bodmer JL, Schneider P, Tschopp J. The molecular architecture of the TNF superfamily. Trends Biochem Sci (2002) 27(1):19–26. doi: 10.1016/S0968-0004(01)01995-8
67. Rhinn H, Tatton N, McCaughey S, Kurnellas M, Rosenthal A. Progranulin as a therapeutic target in neurodegenerative diseases. Trends Pharmacol Sci (2022) 43(8):641–52. doi: 10.1016/j.tips.2021.11.015
68. Tang W, Lu Y, Tian QY, Zhang Y, Guo FJ, Liu GY, et al. The growth factor progranulin binds to TNF receptors and is therapeutic against inflammatory arthritis in mice. Science (2011) 332(6028):478–84. doi: 10.1126/science.1199214
69. Liu CJ, Bosch X. Progranulin: a growth factor, a novel TNFR ligand and a drug target. Pharmacol Ther (2012) 133(1):124–32. doi: 10.1016/j.pharmthera.2011.10.003
70. Fu W, Hettinghouse A, Chen Y, Hu W, Ding X, Chen M, et al. 14-3-3 epsilon is an intracellular component of TNFR2 receptor complex and its activation protects against osteoarthritis. Ann Rheum Dis (2021) 80(12):1615–27. doi: 10.1136/annrheumdis-2021-220000
71. Stubert J, Waldmann K, Dieterich M, Richter DU, Briese V. Progranulin shows cytoprotective effects on trophoblast cells in vitro but does not antagonize TNF-alpha-induced apoptosis. Arch Gynecol Obstet (2014) 290(5):867–73. doi: 10.1007/s00404-014-3296-3
72. Lin H, Huang X, Qin Y, Dang Y, Zhang P, Sun J, et al. Abstract 5523: LBL-019, a novel anti-TNFR2 antibody, shows a potent anti-tumor efficacy in a mouse MC38 model. Cancer Res (2022) 82(12_Supplement):5523–3. doi: 10.1158/1538-7445.AM2022-5523
73. Spira AI, Naing A, Babiker HM, Borad MJ, Garralda E, Leventakos K, et al. Phase I study of HFB200301, a first-in-class TNFR2 agonist monoclonal antibody in patients with solid tumors selected via Drug Intelligent Science (DIS). J Clin Oncol (2022) 40(16_suppl):TPS2670–TPS2670. doi: 10.1200/JCO.2022.40.16_suppl.TPS2670
74. Ceglia V, Zurawski S, Montes M, Bouteau A, Wang Z, Ellis J, et al. Anti-CD40 antibodies fused to CD40 ligand have superagonist properties. J Immunol (2021) 207(8):2060–76. doi: 10.4049/jimmunol.2000704
75. Medler J, Nelke J, Weisenberger D, Steinfatt T, Rothaug M, Berr S, et al. TNFRSF receptor-specific antibody fusion proteins with targeting controlled FcgammaR-independent agonistic activity. Cell Death Dis (2019) 10(3):224.
76. Medler J, Wajant H. Tumor necrosis factor receptor-2 (TNFR2): an overview of an emerging drug target. Expert Opin Ther Targets (2019) 23(4):295–307. doi: 10.1080/14728222.2019.1586886
77. Chen X, Baumel M, Mannel DN, Howard OM, Oppenheim JJ. Interaction of TNF with TNF receptor type 2 promotes expansion and function of mouse CD4+CD25+ T regulatory cells. J Immunol (2007) 179(1):154–61. doi: 10.4049/jimmunol.179.1.154
78. Chen X, Wu X, Zhou Q, Howard OM, Netea MG, Oppenheim JJ, et al. TNFR2 is critical for the stabilization of the CD4+Foxp3+ regulatory T. cell phenotype in the inflammatory environment. J Immunol (2013) 190(3):1076–84.
79. He X, Landman S, Bauland SC, van den Dolder J, Koenen HJ, Joosten I, et al. A TNFR2-agonist facilitates high purity expansion of human low purity Treg cells. PloS One (2016) 11(5):e0156311. doi: 10.1371/journal.pone.0156311
80. Veglia F, Sanseviero E, Gabrilovich DI. Myeloid-derived suppressor cells in the era of increasing myeloid cell diversity. Nat Rev Immunol (2021) 21(8):485–98. doi: 10.1038/s41577-020-00490-y
81. Polz J, Remke A, Weber S, Schmidt D, Weber-Steffens D, Pietryga-Krieger A, et al. Myeloid suppressor cells require membrane TNFR2 expression for suppressive activity. Immun Inflammation Dis (2014) 2(2):121–30. doi: 10.1002/iid3.19
82. Ham B, Wang N, D'Costa Z, Fernandez MC, Bourdeau F, Auguste P, et al. TNF receptor-2 facilitates an immunosuppressive microenvironment in the liver to promote the colonization and growth of hepatic metastases. Cancer Res (2015) 75(24):5235–47. doi: 10.1158/0008-5472.CAN-14-3173
83. Zhao X, Rong L, Zhao X, Li X, Liu X, Deng J, et al. TNF signaling drives myeloid-derived suppressor cell accumulation. J Clin Invest (2012) 122(11):4094–104. doi: 10.1172/JCI64115
84. Wajant H, Siegmund D. TNFR1 and TNFR2 in the control of the life and death balance of macrophages. Front Cell Dev Biol (2019) 7. doi: 10.3389/fcell.2019.00091
85. Gao H, Danzi MC, Choi CS, Taherian M, Dalby-Hansen C, Ellman DG, et al. Opposing functions of microglial and macrophagic TNFR2 in the pathogenesis of experimental autoimmune encephalomyelitis. Cell Rep (2017) 18(1):198–212. doi: 10.1016/j.celrep.2016.11.083
86. Okeke EB, Uzonna JE. The pivotal role of regulatory T cells in the regulation of innate immune cells. Front Immunol (2019) 10:680. doi: 10.3389/fimmu.2019.00680
87. Chen X, Hamano R, Subleski JJ, Hurwitz AA, Howard OMZ, Oppenheim JJ, et al. Expression of costimulatory TNFR2 induces resistance of CD4+FoxP3– conventional T cells to suppression by CD4+FoxP3+ Regulatory T cells. J Immunol (2010) 185(1):174–82. doi: 10.4049/jimmunol.0903548
88. Jiang Y, Chen J, Bi E, Zhao Y, Qin T, Wang Y, et al. TNF-α enhances Th9 cell differentiation and antitumor immunity via TNFR2-dependent pathways. J ImmunoTher Cancer (2019) 7(1):28. doi: 10.1186/s40425-018-0494-8
89. Williams GS, Mistry B, Guillard S, Coates Ulrichsen J, Sandercock AM, Wang J, et al. Phenotypic screening reveals TNFR2 as a promising target for cancer immunotherapy. Oncotarget (2016) 7(42). doi: 10.18632/oncotarget.11943
90. Govindaraj C, Scalzo-Inguanti K, Scholzen A, Li S, Plebanski M. TNFR2 expression on CD25(hi)FOXP3(+) T cells induced upon TCR stimulation of CD4 T cells identifies maximal cytokine-producing effectors. Front Immunol (2013) 4:233. doi: 10.3389/fimmu.2013.00233
91. Kampan NC, Madondo MT, McNally OM, Stephens AN, Quinn MA, Plebanski M, et al. Interleukin 6 present in inflammatory ascites from advanced epithelial ovarian cancer patients promotes tumor necrosis factor receptor 2-expressing regulatory T cells. Front Immunol (2017) 8:1482. doi: 10.3389/fimmu.2017.01482
92. Yan F, Du R, Wei F, Zhao H, Yu J, Wang C, et al. Expression of TNFR2 by regulatory T cells in peripheral blood is correlated with clinical pathology of lung cancer patients. Cancer Immunol Immunother (2015) 64(11):1475–85. doi: 10.1007/s00262-015-1751-z
93. Ye L-L, Peng W-B, Niu Y-R, Xiang X, Wei X-S, Wang Z-H, et al. Accumulation of TNFR2-expressing regulatory T cells in Malignant pleural effusion of lung cancer patients is associated with poor prognosis. Ann Trans Med (2020) 8(24):1647. doi: 10.21037/atm-20-7181
94. Schmitt H, Billmeier U, Dieterich W, Rath T, Sonnewald S, Reid S, et al. Expansion of IL-23 receptor bearing TNFR2+ T cells is associated with molecular resistance to anti-TNF therapy in Crohn's disease. Gut (2019) 68(5):814–28. doi: 10.1136/gutjnl-2017-315671
95. Cianciotti BC, Ruggiero E, Campochiaro C, Oliveira G, Magnani ZI, Baldini M, et al. CD4+ Memory stem T cells recognizing citrullinated epitopes are expanded in patients with rheumatoid arthritis and sensitive to tumor necrosis factor blockade. Arthritis Rheumatol (2020) 72(4):565–75. doi: 10.1002/art.41157
96. Yang S, Chen J, Zhang H, Jiang Y, Qin T, Gao S, et al. TNF-a is a potent stimulator of tc9-cell differentiation. J Immunother (2020) 43(9):265–72. doi: 10.1097/CJI.0000000000000335
97. Calzascia T, Pellegrini M, Hall H, Sabbagh L, Ono N, Elford AR, et al. TNF-α is critical for antitumor but not antiviral T cell immunity in mice. J Clin Invest (2007) 117(12):3833–45.
98. Kim EY, Teh HS. TNF type 2 receptor (p75) lowers the threshold of T cell activation. J Immunol (2001) 167(12):6812–20. doi: 10.4049/jimmunol.167.12.6812
99. Wortzman ME, Lin GH, Watts TH. Intrinsic TNF/TNFR2 interactions fine-tune the CD8 T cell response to respiratory influenza virus infection in mice. PloS One (2013) 8(7):e68911. doi: 10.1371/journal.pone.0068911
100. Kim EY, Teh SJ, Yang J, Chow MT, Teh HS. TNFR2-deficient memory CD8 T cells provide superior protection against tumor cell growth. J Immunol (2009) 183(10):6051–7. doi: 10.4049/jimmunol.0803482
101. Twu YC, Gold MR, Teh HS. TNFR1 delivers pro-survival signals that are required for limiting TNFR2-dependent activation-induced cell death (AICD) in CD8+ T cells. Eur J Immunol (2011) 41(2):335–44. doi: 10.1002/eji.201040639
102. Cope AP, Londei M, Chu NR, Cohen SB, Elliott MJ, Brennan FM, et al. Chronic exposure to tumor necrosis factor (TNF) in vitro impairs the activation of T cells through the T cell receptor/CD3 complex; reversal in vivo by anti-TNF antibodies in patients with rheumatoid arthritis. J Clin Invest (1994) 94(2):749–60. doi: 10.1172/JCI117394
103. Liao P, Jiang M, Islam MS, Wang Y, Chen X. TNFR2 expression predicts the responses to immune checkpoint inhibitor treatments. Front Immunol (2023) 14:1097090. doi: 10.3389/fimmu.2023.1097090
104. Tufa DM, Chatterjee D, Low HZ, Schmidt RE, Jacobs R. TNFR2 and IL-12 coactivation enables slanDCs to support NK-cell function via membrane-bound TNF-alpha. Eur J Immunol (2014) 44(12):3717–28. doi: 10.1002/eji.201444676
105. Chen X, Hamano R, Subleski JJ, Hurwitz AA, Howard OM, Oppenheim JJ, et al. Expression of costimulatory TNFR2 induces resistance of CD4+FoxP3- conventional T cells to suppression by CD4+FoxP3+ regulatory T cells. J Immunol (2010) 185(1):174–82. doi: 10.4049/jimmunol.0903548
106. Mysore V, Tahir S, Furuhashi K, Arora J, Rosetti F, Cullere X, et al. Monocytes transition to macrophages within the inflamed vasculature via monocyte CCR2 and endothelial TNFR2. J Exp Med (2022) 219(5). doi: 10.1084/jem.20210562
107. Neuenfeldt F, Schumacher JC, Grieshaber-Bouyer R, Habicht J, Schroder-Braunstein J, Gauss A, et al. Inflammation induces pro-NETotic neutrophils via TNFR2 signaling. Cell Rep (2022) 39(3):110710. doi: 10.1016/j.celrep.2022.110710
108. Suto T, Tosevska A, Dalwigk K, Kugler M, Dellinger M, Stanic I, et al. TNFR2 is critical for TNF-induced rheumatoid arthritis fibroblast-like synoviocyte inflammation. Rheumatol (Oxford) (2022) 61(11):4535–46. doi: 10.1093/rheumatology/keac124
109. Mountz JD. Re: "collagen-induced arthritis in TNF receptor-1-deficient mice: TNF receptor-2 can modulate arthritis in the absence of TNF receptor 1". Clin Immunol (2001) 99(3):305–7. doi: 10.1006/clim.2001.5030
110. Bai J, Ding B, Li H. Targeting TNFR2 in cancer: all roads lead to Rome. Front Immunol (2022) 13:844931. doi: 10.3389/fimmu.2022.844931
111. Chen Y, Jia M, Wang S, Xu S, He N. Antagonistic antibody targeting TNFR2 inhibits regulatory T cell function to promote anti-tumor activity. Front Immunol (2022) 13:835690. doi: 10.3389/fimmu.2022.835690
112. Torrey H, Khodadoust M, Tran L, Baum D, Defusco A, Kim YH, et al. Targeted killing of TNFR2-expressing tumor cells and T(regs) by TNFR2 antagonistic antibodies in advanced Sezary syndrome. Leukemia (2019) 33(5):1206–18. doi: 10.1038/s41375-018-0292-9
113. Torrey H, Butterworth J, Mera T, Okubo Y, Wang L, Baum D, et al. Targeting TNFR2 with antagonistic antibodies inhibits proliferation of ovarian cancer cells and tumor-associated Tregs. Sci Signal (2017) 10(462). doi: 10.1126/scisignal.aaf8608
114. Zhang X, Lao M, Xu J, Duan Y, Yang H, Li M, et al. Combination cancer immunotherapy targeting TNFR2 and PD-1/PD-L1 signaling reduces immunosuppressive effects in the microenvironment of pancreatic tumors. J Immunother Cancer (2022) 10(3). doi: 10.1136/jitc-2021-003982
115. Moatti A, Debesset A, Pilon C, Beldi-Ferchiou A, Leclerc M, Redjoul R, et al. TNFR2 blockade of regulatory T cells unleashes an antitumor immune response after hematopoietic stem-cell transplantation. J Immunother Cancer (2022) 10(4). doi: 10.1136/jitc-2021-003508
116. Fu Q, Shen Q, Tong J, Huang L, Cheng Y, Zhong W, et al. Anti-tumor necrosis factor receptor 2 antibody combined with anti-PD-L1 therapy exerts robust antitumor effects in breast cancer. Front Cell Dev Biol (2021) 9:720472. doi: 10.3389/fcell.2021.720472
117. He T, Chen Y, Yang D, Islam MS, Chou CK, Liu J, et al. TNFR2 antagonistic antibody induces the death of tumor infiltrating CD4(+)Foxp3(+) regulatory T cells. Cell Oncol (Dordr) (2023) 46(1):167–77. doi: 10.1007/s13402-022-00742-0
118. Zhu L, Zhang X, Chen X, Yang D, Nie Y, Pan R, et al. Anti-TNFR2 enhanced the antitumor activity of a new HMGN1/3M-052 stimulated dendritic cell vaccine in a mouse model of colon cancer. Biochem Biophys Res Commun (2023) 653:106–14. doi: 10.1016/j.bbrc.2023.02.039
119. Jiang M, Liu J, Yang D, Tross D, Li P, Chen F, et al. A TNFR2 antibody by countering immunosuppression cooperates with HMGN1 and R848 immune stimulants to inhibit murine colon cancer. Int Immunopharmacol (2021) 101(Pt A):108345. doi: 10.1016/j.intimp.2021.108345
120. Calzascia T, Pellegrini M, Hall H, Sabbagh L, Ono N, Elford AR, et al. TNF-alpha is critical for antitumor but not antiviral T cell immunity in mice. J Clin Invest (2007) 117(12):3833–45.
121. Lubrano di Ricco M, Ronin E, Collares D, Divoux J, Gregoire S, Wajant H, et al. Tumor necrosis factor receptor family costimulation increases regulatory T-cell activation and function via NF-kappaB. Eur J Immunol (2020) 50(7):972–85. doi: 10.1002/eji.201948393
122. Durham NM, Holoweckyj N, MacGill RS, McGlinchey K, Leow CC, Robbins SH, et al. GITR ligand fusion protein agonist enhances the tumor antigen-specific CD8 T-cell response and leads to long-lasting memory. J Immunother Cancer (2017) 5:47. doi: 10.1186/s40425-017-0247-0
123. Rodriguez M, Cabal-Hierro L, Carcedo MT, Iglesias JM, Artime N, Darnay BG, et al. NF-kappaB signal triggering and termination by tumor necrosis factor receptor 2. J Biol Chem (2011) 286(26):22814–24. doi: 10.1074/jbc.M111.225631
124. Siegmund D, Kums J, Ehrenschwender M, Wajant H. Activation of TNFR2 sensitizes macrophages for TNFR1-mediated necroptosis. Cell Death Dis (2016) 7(9):e2375. doi: 10.1038/cddis.2016.285
125. Zeng H, Chi H. mTOR signaling in the differentiation and function of regulatory and effector T cells. Curr Opin Immunol (2017) 46:103–11. doi: 10.1016/j.coi.2017.04.005
126. Hirschhorn-Cymerman D, Rizzuto GA, Merghoub T, Cohen AD, Avogadri F, Lesokhin AM, et al. OX40 engagement and chemotherapy combination provides potent antitumor immunity with concomitant regulatory T cell apoptosis. J Exp Med (2009) 206(5):1103–16. doi: 10.1084/jem.20082205
127. Amoozgar Z, Kloepper J, Ren J, Tay RE, Kazer SW, Kiner E, et al. Targeting Treg cells with GITR activation alleviates resistance to immunotherapy in murine glioblastomas. Nat Commun (2021) 12(1):2582. doi: 10.1038/s41467-021-22885-8
128. Madsen PM, Desu HL, de Rivero Vaccari JP, Florimon Y, Ellman DG, Keane RW, et al. Oligodendrocytes modulate the immune-inflammatory response in EAE via TNFR2 signaling. Brain Behav Immun (2020) 84:132–46. doi: 10.1016/j.bbi.2019.11.017
129. Madsen PM, Motti D, Karmally S, Szymkowski DE, Lambertsen KL, Bethea JR, et al. Oligodendroglial TNFR2 mediates membrane TNF-dependent repair in experimental autoimmune encephalomyelitis by promoting oligodendrocyte differentiation and remyelination. J Neurosci (2016) 36(18):5128–43. doi: 10.1523/JNEUROSCI.0211-16.2016
130. Schumann J, Muhlen K, Kiemer AK, Vollmar AM, Tiegs G. Parenchymal, but not leukocyte, TNF receptor 2 mediates T cell-dependent hepatitis in mice. J Immunol (2003) 170(4):2129–37. doi: 10.4049/jimmunol.170.4.2129
131. Chuah S, Lee J, Song Y, Kim HD, Wasser M, Kaya NA, et al. Uncoupling immune trajectories of response and adverse events from anti-PD-1 immunotherapy in hepatocellular carcinoma. J Hepatol (2022) 77(3):683–94. doi: 10.1016/j.jhep.2022.03.039
132. Li P, Yang XY, Wang YF, Chou CK, Jiang MM, et al. TNFR2 deficiency impairs the growth of mouse colon cancer. Int J Biol Sci (2023) 19(4):1024–35. doi: 10.7150/ijbs.72606
133. Wiedmann L, De Angelis Rigotti F, Vaquero-Siguero N, Donato E, Espinet E, Moll I, et al. HAPLN1 potentiates peritoneal metastasis in pancreatic cancer. Nat Commun (2023) 14(1):2353. doi: 10.1038/s41467-023-38064-w
Keywords: Treg - regulatory T cell, TNFR2 agonism, TNFR2, tumor, autoimmune diseases
Citation: Chen Y, Jiang M and Chen X (2023) Therapeutic potential of TNFR2 agonists: a mechanistic perspective. Front. Immunol. 14:1209188. doi: 10.3389/fimmu.2023.1209188
Received: 20 April 2023; Accepted: 31 July 2023;
Published: 17 August 2023.
Edited by:
Harald Wajant, University Hospital Würzburg, GermanyReviewed by:
Nikolaos Skartsis, Mayo Clinic, United StatesDenise Louise Faustman, Massachusetts General Hospital and Harvard Medical School, United States
Copyright © 2023 Chen, Jiang and Chen. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xin Chen, eGNoZW5AdW0uZWR1Lm1v
†These authors have contributed equally to this work