 Li Zhou
Li Zhou Tiantian Zhao5†
Tiantian Zhao5† Jiwei Li
Jiwei Li- 1Department of Pulmonary and Critical Care Medicine, The Second Xiangya Hospital, Central South University, Changsha, Hunan, China
- 2Research Unit of Respiratory Disease, Central South University, Changsha, Hunan, China
- 3Diagnosis and Treatment Center of Respiratory Disease, Central South University, Changsha, Hunan, China
- 4Clinical Medical Research Center for Pulmonary and Critical Care Medicine in Hunan Province, Changsha, China
- 5Department of Oncology, The Second Xiangya Hospital, Central South University, Changsha, China
- 6Department of Pathology, Fudan University Shanghai Cancer Center, Shanghai, China
Macrophages are the main component of the tumor microenvironment, which are differentiated from monocytes in the blood and play an important role in cancer development. Tumor-associated macrophages (TAMs) can promote tumor growth, invasion, metastasis, and resistance to anti–programmed death receptor 1 therapy by regulating programmed cell death ligand 1 expression and interacting with other immune cells in the tumor microenvironment. However, when activated properly, macrophages can also play an anti-tumor role by enhancing the phagocytosis and cytotoxicity of tumor cells. TAM is associated with poor prognosis and drug resistance in patients treated with immunotherapy, indicating that macrophages are attractive targets for combined therapy in cancer treatment. Combination of targeting TAMs and immunotherapy overcomes the drug resistance and achieved excellent results in some cancers, which may be a promising strategy for cancer treatment in the future. Herein, we review the recent findings on the role of macrophages in tumor development, metastasis, and immunotherapy. We focus mainly on macrophage-centered therapy, including strategies to deplete and reprogram TAMs, which represent the potential targets for improving tumor immunotherapy efficacy.
1 Introduction
Cancer is one of the major public health issues worldwide and is the leading cause of death in many countries. According to the latest data published in 2023, approximately 1,958,310 new cancer cases were present in the United States (1). Moreover, due to the high mortality rate and low cure rate of cancer, it has brought heavy economic burden to individuals, families, and society. Therefore, the prevention and treatment of tumors were urgent to further reduce the morbidity and mortality rates. Surgery, radiotherapy, and chemotherapy are three traditional treatment strategies for cancer, but the treatment outcome was still dismal in some patients (1, 2). In recent years, emerging treatment methods have been developed, such as Chimeric antigen receptor (CAR)-T cell therapy and immune-checkpoint inhibitors, which were considered the fourth treatment mode following traditional therapy. At present, immunotherapy has been approved for clinical use, mainly including programmed death receptor 1 (PD-1) inhibitors and CAR-T cell therapy, both of which have achieved excellent results in some advanced stage malignant tumors (3–6). However, the efficacy of PD-1 inhibitor was limited in some patients with cancer (7), and the efficacy needs to be further improved.
The tumor microenvironment was considered to be a key factor affecting tumor progression, metastasis, and treatment results (8, 9). Exploring the tumor microenvironment is the cornerstone of improving the response rate and developing new cancer immunotherapy strategies. In addition, macrophages were reported to be one of the most important immune cells in the tumor microenvironment (9). Based on the function of phagocytosis, macrophages can eliminate tumor cells at an early stage, but, under the stimulation of the stimulating factors in the tumor microenvironment, they gradually transform into tumor-related macrophages with the M2 phenotype and promote tumor growth and metastasis by inhibiting immunity, inducing angiogenesis and supporting cancer stem cells (10). To sum up, it is of great significance to explore in great depth the role of macrophages in the tumor microenvironment, and targeting macrophages may be a promising anti-tumor strategy in the future.
2 Origin, polarization, and function of macrophages
Macrophages originate from the monocytes in the circulation, and substantial heterogeneity was observed among each macrophage population (11). According to phenotype and function, macrophages can be divided into two types: classically activated macrophages (M1 macrophages) and alternatively activated macrophages (M2 macrophages) (12). M0 macrophages could differentiate into M1 macrophages under the stimulation of lipopolysaccharide and interferon-γ (IFN-γ), whereas they differentiate into M2 macrophages with the stimulation of interleukin (IL)-4, IL-10, and IL-13 (13). M1 macrophages could produce multiple cytotoxic substances, such as nitric oxide and reactive oxygen species, and thereby activate the function of multiple immune cells and reduce microbial activity, ultimately eliminating microbial infection (14). Meanwhile, a variety of cytokines were produced by M1 macrophages, including tumor necrosis factor–α (TNF-α), growth inhibitors, and anti-angiogenic factors, which could inhibit cancer progression (14). On the contrary, M2 macrophages often function as anti-inflammatory factors by reducing the inflammation response, promoting tissue repair and remodeling the immune system (10, 14). Tumor-associated macrophages (TAMs) were mainly thought to be M2 type in the tumor microenvironment, which could promote tumor growth, invasion, and metastasis.
3 Macrophages in the TME promote tumor progression
Macrophages are involved in different stages of tumor development. In the early stage, tumor cells release cytokines and exosomes and attract macrophages and other immune cells into the tumor stroma, where macrophages promote tumor growth, migration, and metastasis (10). As a key component of the tumor microenvironment, macrophages can produce an anti-tumor effect and cause tumor necrosis with powerful swallowing phagocytosis (15), but some studies have shown that TAM is an important driving factor of tumor progression. In the tumors formed, TAM promotes the growth and proliferation of cancer cells, angiogenesis, and lymphangiogenesis and inhibits the immune response of effector T cells (16).
TAM is considered a proinflammatory and anti-tumor phenotype (M1 type) in the early stage of lung cancer and gradually displays an anti-inflammatory and tumor-promoting phenotype in the process of cancer progression (10). TAM could promote tumor development through immune regulation and non-immune processes (17–19). For example, TAM secretes a large number of pro-angiogenic factors such as vascular endothelial growth factor (VEGF) to promote tumor angiogenesis and metastasis (20).
In the tumor microenvironment, macrophages account for half of the total number of tumor cells and are mainly M2 phenotypes. The quantity of macrophages in the tumor microenvironment is associated with tumor micro-vessels and is negatively correlated with the survival outcome in patients with non-small cell lung cancer (NSCLC) (21, 22). In recent years, a growing body of research has revealed the TAM multifaceted regulation of the co-evolving cancer ecosystem based on next-generation technologies and single-cell sequencing technology (12, 22). Therefore, this section mainly introduces the function and mechanism of TAM in tumors.
3.1 Anti-tumor effect of M1 type TAM
Inhibition of anti-tumor immunity was reported to be the main pathogenic mechanism of TAM. TAM could downregulate the release of the immunostimulatory factor IL-12, which can trigger the tumor-killing effect of natural killer (NK) cells and cytotoxic CD4+ T cells (23). In addition, many immunosuppressive factors produced by TAM could also mediate cancer development, such as IL-10, transforming growth factor–β, and prostaglandin E2 (10, 24, 25).
TAM can also directly inhibit the function of T cells through specific enzyme activities, such as arginase 1 (ARG1), which is a hydrolase that controls the catabolism of L-arginine. ARG1 is induced by multiple signaling pathways mediated by IL-4, IL-10, and hypoxia and affects T-cell function by limiting the activity of the semi-essential amino acid L-arginine (25). TAM can also promote T-cell apoptosis by inhibiting the expression of programmed cell death ligand 1 (PD-L1) and B7 homolog 1 on T cells (12, 25).
3.2 The function of M2 type TAM in promoting tumor development
The function of M2 macrophages in promoting tumor development depends on the proinflammatory cytokines, such as TNF-α, IL-6, and IL-11, which can activate the nuclear factor–κB (NF-κB) and signal transduction and activator of transcription 3 (STAT3) pathway in cancer cells (10, 12, 13, 18, 25). In addition, M2 TAM promoted tumor progression by promoting angiogenesis and lymphangiogenesis by increasing the expression of VEGF-A and VEGF-C (18, 20, 25).
4 Macrophages and anti–PD-1/PD-L1 immunotherapy
4.1 Effect of TAMs on PD-1/PD-L1 expression
The PD-1/PD-L1 pathway was abnormally activated in various cancers (6, 26), and anti–PD-1/PD-L1 immunotherapy has been widely used or tried in clinical trials in many solid tumors, such as lung cancer, advanced metastatic melanoma, esophagus cancer, and colorectal cancer (27, 28). However, the efficacy of PD-1 inhibitors was still dismal in some patients with high expression of PD-L1, and the concrete mechanisms remain largely unknown.
Previous studies have demonstrated that TAMs can regulate the expression of PD-1/PD-L1 through the activation of different signaling pathways (Figure 1), which, in turn, affects the efficacy of PD-1/PD-L1 inhibitors. CD163+ TAMs in the tumor microenvironment are reported to be positively correlated to PD-L1 expression in various cancers, including pancreatic cancer and liver cancer. Multiple cytokines released by TAM, including IL-6 and TNF-α, can upregulate PD-L1 expression by activating Janus kinase (JAK)/STAT3, phosphoinositide 3-kinase (PI3K)/AKT, NF-κB, or Extracellular signal-regulated kinase (ERK) 1 and 2 signaling pathways (29, 30). In addition, PD-L1 protein expression could also be upregulated by TNF-α through post-translational regulation (29).
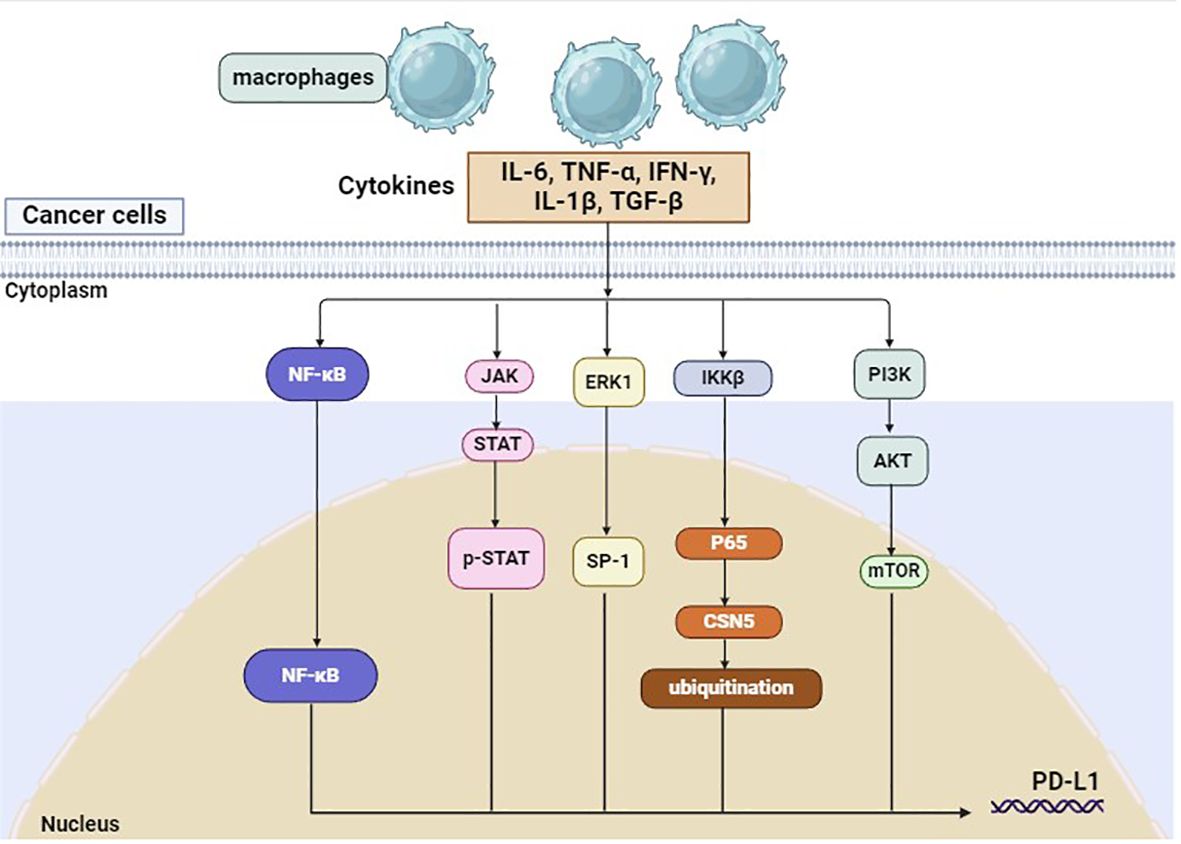
Figure 1 PD-L1 on tumor cells can be regulated by macrophages.
4.2 TAMs and anti–PD-1 resistance
In addition to the PD-L1 expression on tumor cells, the tumor microenvironment was also a key factor associated with anti–PD-1 resistance. As mentioned above, cytokines released by TAMs could regulate PD-L1 protein expression, which was reported to be an important predictor for anti–PD≥1/PD≥L1 therapy. In recent years, multiple immune cells have been identified in TME, and the cancer ecosystem has evolved over time, which plays a complex role in cancer development (31, 32). The interaction between macrophages and other immune cells was explored and demonstrated to be correlated to the response to immunotherapy (31). Single-cell and spatial analysis showed that interaction between FAP+ fibroblasts and SPP1+ macrophages could promote the formation of immune-excluded desmoplasic structures and restrict T-cell which reduces the efficacy of immunotherapy (31). In triple-negative breast cancer, high levels of CXCL13+ T cells are associated with the proinflammatory features of macrophages and can predict the clinical benefit of checkpoint inhibitors (32).
Exosomes are small extracellular vesicles that play a crucial role in various cell activities in cancer. Recent studies have reported that macrophage-derived exosomes may promote the formation of a pre-metastatic niche that facilitates tumor growth and metastasis. M2 macrophage–derived EVs can drive anti–PD≥1/PD≥L1 therapy resistance, promote the expression of drug-resistant genes in tumor cells, or affect the immune cell spectrum in TME (33, 34). Therefore, the interaction between TAMs and TME may contribute to anti–PD≥1 therapy resistance in cancer, providing a theoretical basis for the combination use of targeting macrophages and anti–PD≥1/PD≥L1 therapy.
4.3 Effect of anti–PD-1/PD-L1 therapy on macrophages
Previous studies have shown that PD-1 inhibitors have an impact on TME in various cancers (35). In non–small cell lung cancer, single-cell RNA sequencing demonstrated that the tumor microenvironment was remodeled after neoadjuvant PD-1 blockade combined with chemotherapy, and TAMs were transformed into a neutral type instead of an anti-tumor phenotype (36). Furthermore, anti–PD-L1 therapy can inhibit tumor growth by reducing PD-L1 expression and promoting the expression of the co-stimulatory molecules CD86 and major histocompatibility complex class II (MHC-II) (37). In addition, the phagocytic ability and immune function of macrophages were also enhanced by anti–PD-L1 therapy, which activates T cells in the TME and eradicates cancer cells (37). Therefore, anti–PD-L1 therapy may repolarize macrophages, enhance the phagocytic ability of macrophages, and ameliorate the tumor microenvironment in some patients.
5 Targeting macrophages in the tumor microenvironment
As TAM is involved in tumor immunity and tumor development, it may become a promising target in the future. Current treatment strategies targeting macrophages can be roughly divided into two categories: TAM depletion and TAM reprogramming (Supplementary Figure 1). In order to ensure treatment efficacy, targeting TAMs was frequently combined with other treatments in clinical studies, such as immunotherapy, chemotherapy, and radiotherapy (Table 1) (38–48).
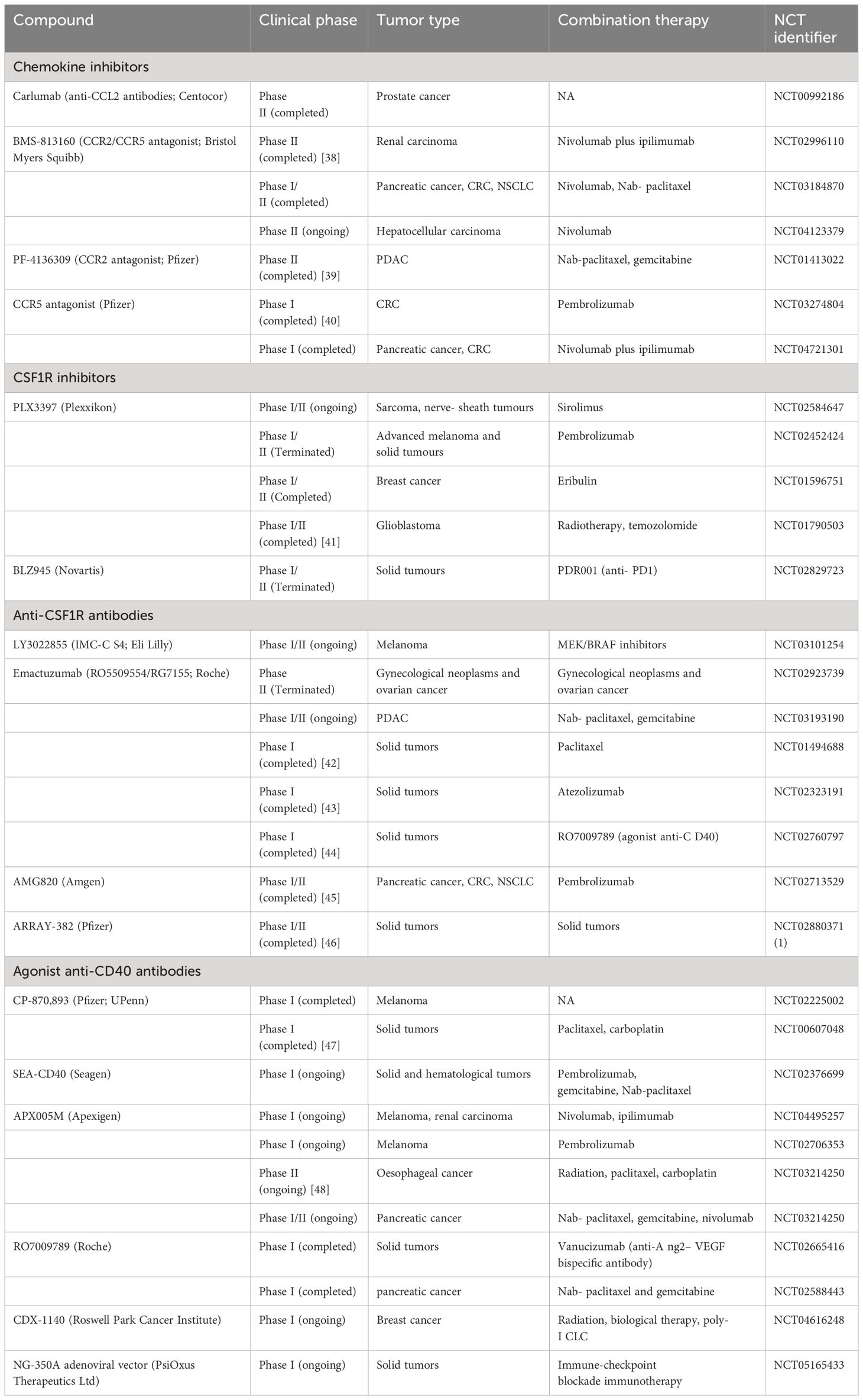
Table 1 Selected clinical trials of agents targeting tumor-associated macrophages.
5.1 Depletion of TAM
Depletion of macrophages in the tumor microenvironment may be an effective treatment strategy for cancer, either alone or in combination with chemotherapy. Inhibition of the signal transduction axis of colony-stimulating factor-1/colony-stimulating factor-1 receptor (CSF1/CSF1R), which is necessary for macrophage survival, can induce apoptosis of macrophages. On the one hand, inhibition of CSF-1R combined with radiotherapy or chemotherapy can improve T-cell responses. Blockade of CSF1R signaling can effectively deplete the immunosuppressive TAM and then stimulate the CD8+ T-cell response, resulting in prolonged survival in glioblastoma brain tumors (49). At present, CSF1R inhibitors in combination with chemotherapy are being tested in clinical trials in some cancers, such as localized prostate cancer and orthotopic glioblastoma (49, 50). In addition, blocking CSF1/CSF1R can improve the efficacy of a variety of immunotherapies, including CD-40 agonists (51) and PD-1 inhibitors (52).
As TAM was transformed from monocytes, blocking the recruitment of monocytes in the circulation to the tumor site was another method to reduce TAM in the tumor microenvironment. Recruitment of monocytes from bone marrow to the tumor site is dependent on C-C motif ligand 2 (CCL2)-CC chemokine receptor 2 (CCR2) signal transduction (53). Inhibition of CCR2 causes monocyte retention in bone marrow and leads to depletion of monocytes in the peripheral circulation, reduction of monocyte recruitment to the primary tumor sites and metastatic foci, and consequent reduction of TAM number, resulting in tumor shrinkage and survival improvement (54–56).
Other pathways involved in macrophage recruitment include CXCL12-CXCR4 and the angiopoietin 2 (ANG2)–TIE2 axis (57–59). Therefore, depletion of TEM may cause vascular destruction, neutralization of ANG2 may improve the response to vascular VEGFA blockade, and inhibition of TEM recruitment may inhibit tumor growth (60).
5.2 Reprogramming of TAM
As macrophages were the main phagocyte and antigen-presenting cell in the tumor, the immune stimulation function of macrophages was lost after the removal of TAMs. Therefore, reprogramming or repolarization of TAM to enhance its anti-tumor function and limit tumor-promoting properties is a more attractive strategy for cancer treatment. For example, in the mouse model of breast cancer, TAM represents the main source of IL-10 and inhibition of IL-10 signal transduction can significantly improve the efficacy of chemotherapy. The IL-10 secreted by TAM inhibits the IL-12 produced by APCs, thereby inhibiting the anti-tumor response of CD8+ T cells induced by paclitaxel and carboplatin (23). In addition, the repolarization of TAM makes it specifically express the proinflammatory cytokine IFN-α, which could activate NK cells and T cells in the tumor environment and significantly slow tumor growth in the mouse model (61). The epigenetic reprogramming of macrophages by inhibiting histone deacetylase (HDAC) can also trigger an immune response in T cells (62, 63). In the breast cancer model, selective class IIa HDAC inhibitor induces the anti-tumor macrophage phenotype, promotes the T-cell immune response, and increases the response to chemotherapy and immune checkpoint inhibitors (62). In addition, the activation of the PI3K signaling in macrophages can drive the immunosuppressive activity in TAM, whereas inhibition of the PI3K pathway can reprogram macrophages enhance T-cell responses (64, 65).
5.3 Macrophage cell therapy
CAR-T cells are reported to be effective in hematological malignancies, whereas the efficacy of CAR-T therapy remains dismal in solid tumors, as the entry of T cells into tumors is restrained (66, 67). However, CAR-macrophages (CAR-M) overcome this disadvantage as the macrophages in the TME could be replenished by circulating monocytes. CAR expression could enhance the antigen-dependent functions of macrophages, such as the secretion of cytokines, polarization, enhanced phagocytic ability, and anti-cancer activity (68). CAR-M cells mediate phagocytosis, exhibit M1 functions in a relatively stable way, and exert anti-tumor effects in primary and metastatic tumors (69). Currently, several clinical trials are underway or being developed to evaluate the anti-cancer efficacy of CAR-M in different tumors.
5.4 Combination of targeting macrophages and anti–PD-1 therapy in cancer
The combination of targeting macrophages and anti–PD-1 therapy in cancer has been investigated in vitro and in vivo (37, 70–72). As we have noted above, repolarization of TAM was considered a promising strategy for cancer treatment, and this approach can potentiate anti–PD-1 therapy efficacy in hepatocellular carcinoma (72). Chemotherapy and radiotherapy may reset macrophages toward an M1 phenotype and improving the efficacy of immunotherapy in cancer (71). Vinblastine can drive the polarization of TAMs to the M1 phenotype by activating NF-κB, increasing CD8+ T-cell populations, and improving the survival outcome of malignant tumor immunotherapy (71). Bi-target treatment such as PD-1–IL-2 cytokine variant (IL2v), which employs anti–PD-1 as a target moiety that is fused into an immuno-stimulatory IL2v, can improve the therapeutic efficacy by reprogramming immunosuppressive TAMs (70). In conclusion, targeting macrophages combined with anti–PD–1 therapy may be a promising strategy to overcome drug resistance in patients with cancer.
6 Conclusion
Macrophages are involved in various cell activities in cancer, and the interaction between macrophages and cancer cells or other immune cells is associated with tumor development. As an important part of the tumor microenvironment, TAMs may be a promosing target for cancer treatment. Targeting macrophages alone or combined with radiotherapy, chemotherapy, and immune-checkpoint inhibitors may produce excellent anti-tumor activity. In addition, the upstream and downstream pathways that may regulate the function of macrophages may also serve as therapeutic targets. In particular, the use of genetic engineering to reprogram macrophages to transform tumor-promoting TAM into anti-tumor macrophages is of great clinical application. Although the combination of targeting macrophages and anti–PD-1 therapy in cancer has been tried in clinical trials or preclinical experiments, this treatment approach is still in its infancy and needs further investigation. Stumbling blocks in the transformation and application of TAM-targeted therapy include the diversity and plasticity of mononuclear phagocytes in the TME (73). The dissection of the TME at the single-cell level confirmed the diversity of macrophages and their relationship with other immune cells (22, 31), which provides a rationale to selectively deplete tumor-promoting macrophages and eliminate tumors. The application of macrophage-targeted therapy in cancer is still in its infancy, and the efficacy and tolerance need to be confirmed in more experiments and clinical trials in the future.
Author contributions
LZ: Writing – original draft. TZ: Writing – original draft, Formal analysis, Data curation, Conceptualization. RZ: Resources, Project administration, Writing – original draft. CC: Methodology, Investigation, Writing – original draft. JL: Writing – review & editing, Visualization, Validation, Supervision, Funding acquisition.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was supported by the Scientific Research Project of Hunan Provincial Health Commission (No. 20231607), the Scientific Research Launch Project for new employees of the Second Xiangya Hospital of Central South University, the Beijing Xisike Clinical Oncology Research Foundation (Grant No. Y-Young2023-0175), the funding of the National Natural Science Foundation of China (No. 82300116), the Natural Science Foundation of Changsha City (CN) (No. kq2208304), and Natural Science Foundation of Hunan Province (No. 2023JJ40878).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2024.1381225/full#supplementary-material
Supplementary Figure 1 | Targeting macrophages in the tumor microenvironment.
References
1. Siegel RL, Miller KD, Wagle NS, Jemal A. Cancer statistics, 2023. CA Cancer J Clin. (2023) 73:17–48. doi: 10.3322/caac.21763
2. Shi D, Gao L, Wan XC, Li J, Tian T, Hu J, et al. Clinicopathologic features and abnormal signaling pathways in plasmablastic lymphoma: a multicenter study in China. BMC Med. (2022) 20:483. doi: 10.1186/s12916-022-02683-9
3. Kamdar M, Solomon SR, Arnason J, Johnston PB, Glass B, Bachanova V, et al. Lisocabtagene maraleucel versus standard of care with salvage chemotherapy followed by autologous stem cell transplantation as second-line treatment in patients with relapsed or refractory large B-cell lymphoma (TRANSFORM): results from an interim analysis of an open-label, randomised, phase 3 trial. Lancet. (2022) 399:2294–308. doi: 10.1016/S0140-6736(22)00662-6
4. Del Bufalo F, De Angelis B, Caruana I, Del Baldo G, De Ioris MA, Serra A, et al. GD2-CART01 for relapsed or refractory high-risk neuroblastoma. N Engl J Med. (2023) 388:1284–95. doi: 10.1056/NEJMoa2210859
5. Janjigian YY, Shitara K, Moehler M, Garrido M, Salman P, Shen L, et al. First-line nivolumab plus chemotherapy versus chemotherapy alone for advanced gastric, gastro-oesophageal junction, and oesophageal adenocarcinoma (CheckMate 649): a randomised, open-label, phase 3 trial. Lancet. (2021) 398:27–40. doi: 10.1016/S0140-6736(21)00797-2
6. Li JW, Deng C, Zhou XY, Deng R. The biology and treatment of Epstein-Barr virus-positive diffuse large B cell lymphoma, NOS. Heliyon. (2024) 10:e23921. doi: 10.1016/j.heliyon.2023.e23921
7. Goc J, Lv M, Bessman NJ, Flamar AL, Sahota S, Suzuki H, et al. Dysregulation of ILC3s unleashes progression and immunotherapy resistance in colon cancer. Cell. (2021) 184:5015–30 e16. doi: 10.1016/j.cell.2021.07.029
8. Scholler N, Perbost R, Locke FL, Jain MD, Turcan S, Danan C, et al. Tumor immune contexture is a determinant of anti-CD19 CAR T cell efficacy in large B cell lymphoma. Nat Med. (2022) 28:1872–82. doi: 10.1038/s41591-022-01916-x
9. Hirz T, Mei S, Sarkar H, Kfoury Y, Wu S, Verhoeven BM, et al. Dissecting the immune suppressive human prostate tumor microenvironment via integrated single-cell and spatial transcriptomic analyses. Nat Commun. (2023) 14:663. doi: 10.1038/s41467-023-36325-2
10. Cassetta L, Pollard JW. A timeline of tumour-associated macrophage biology. Nat Rev Cancer. (2023) 23:238–57. doi: 10.1038/s41568-022-00547-1
11. Chan JM, Quintanal-Villalonga A, Gao VR, Xie Y, Allaj V, Chaudhary O, et al. Signatures of plasticity, metastasis, and immunosuppression in an atlas of human small cell lung cancer. Cancer Cell. (2021) 39:1479–96 e18. doi: 10.1016/j.ccell.2021.09.008
12. Kloosterman DJ, Akkari L. Macrophages at the interface of the co-evolving cancer ecosystem. Cell. (2023) 186:1627–51. doi: 10.1016/j.cell.2023.02.020
13. Kerneur C, Cano CE, Olive D. Major pathways involved in macrophage polarization in cancer. Front Immunol. (2022) 13:1026954. doi: 10.3389/fimmu.2022.1026954
14. Boutilier AJ, Elsawa SF. Macrophage polarization states in the tumor microenvironment. Int J Mol Sci. (2021) 22:6995. doi: 10.3390/ijms22136995
15. Tang Z, Davidson D, Li R, Zhong MC, Qian J, Chen J, et al. Inflammatory macrophages exploit unconventional pro-phagocytic integrins for phagocytosis and anti-tumor immunity. Cell Rep. (2021) 37:110111. doi: 10.1016/j.celrep.2021.110111
16. Zheng X, Jiang Q, Han M, Ye F, Wang M, Qiu Y, et al. FBXO38 regulates macrophage polarization to control the development of cancer and colitis. Cell Mol Immunol. (2023) 20:1367–78. doi: 10.1038/s41423-023-01081-2
17. Zhang G, Gao Z, Guo X, Ma R, Wang X, Zhou P, et al. CAP2 promotes gastric cancer metastasis by mediating the interaction between tumor cells and tumor-associated macrophages. J Clin Invest. (2023) 133:e166224. doi: 10.1172/JCI166224
18. Chen S, Saeed A, Liu Q, Jiang Q, Xu H, Xiao GG, et al. Macrophages in immunoregulation and therapeutics. Signal Transduct Target Ther. (2023) 8:207. doi: 10.1038/s41392-023-01452-1
19. Park MD, Reyes-Torres I, LeBerichel J, Hamon P, LaMarche NM, Hegde S, et al. TREM2 macrophages drive NK cell paucity and dysfunction in lung cancer. Nat Immunol. (2023) 24:792–801. doi: 10.1038/s41590-023-01475-4
20. Yu Y, Dai K, Gao Z, Tang W, Shen T, Yuan Y, et al. Sulfated polysaccharide directs therapeutic angiogenesis via endogenous VEGF secretion of macrophages. Sci Adv. (2021) 7:eabd8217. doi: 10.1126/sciadv.abd8217
21. Casanova-Acebes M, Dalla E, Leader AM, LeBerichel J, Nikolic J, Morales BM, et al. Tissue-resident macrophages provide a pro-tumorigenic niche to early NSCLC cells. Nature. (2021) 595:578–84. doi: 10.1038/s41586-021-03651-8
22. Leader AM, Grout JA, Maier BB, Nabet BY, Park MD, Tabachnikova A, et al. Single-cell analysis of human non-small cell lung cancer lesions refines tumor classification and patient stratification. Cancer Cell. (2021) 39:1594–609 e12. doi: 10.1016/j.ccell.2021.10.009
23. Ruffell B, Chang-Strachan D, Chan V, Rosenbusch A, Ho CM, Pryer N, et al. Macrophage IL-10 blocks CD8+ T cell-dependent responses to chemotherapy by suppressing IL-12 expression in intratumoral dendritic cells. Cancer Cell. (2014) 26:623–37. doi: 10.1016/j.ccell.2014.09.006
24. Pittet MJ, Michielin O, Migliorini D. Clinical relevance of tumour-associated macrophages. Nat Rev Clin Oncol. (2022) 19:402–21. doi: 10.1038/s41571-022-00620-6
25. Aegerter H, Lambrecht BN, Jakubzick CV. Biology of lung macrophages in health and disease. Immunity. (2022) 55:1564–80. doi: 10.1016/j.immuni.2022.08.010
26. Li JW, Shi D, Wan XC, Hu J, Su YF, Zeng YP, et al. Universal extracellular vesicles and PD-L1+ extracellular vesicles detected by single molecule array technology as circulating biomarkers for diffuse large B cell lymphoma. Oncoimmunology. (2021) 10:1995166. doi: 10.1080/2162402X.2021.1995166
27. Janjigian YY, Kawazoe A, Bai Y, Xu J, Lonardi S, Metges JP, et al. Pembrolizumab plus trastuzumab and chemotherapy for HER2-positive gastric or gastro-oesophageal junction adenocarcinoma: interim analyses from the phase 3 KEYNOTE-811 randomised placebo-controlled trial. Lancet. (2023) 402:2197–208. doi: 10.1016/S0140-6736(23)02033-0
28. Brahmer JR, Drake CG, Wollner I, Powderly JD, Picus J, Sharfman WH, et al. Phase I study of single-agent anti-programmed death-1 (MDX-1106) in refractory solid tumors: safety, clinical activity, pharmacodynamics, and immunologic correlates. J Clin Oncol. (2023) 41:715–23. doi: 10.1200/JCO.22.02270
29. Pu Y, Ji Q. Tumor-associated macrophages regulate PD-1/PD-L1 immunosuppression. Front Immunol. (2022) 13:874589. doi: 10.3389/fimmu.2022.874589
30. Zhang H, Liu L, Liu J, Dang P, Hu S, Yuan W, et al. Roles of tumor-associated macrophages in anti-PD-1/PD-L1 immunotherapy for solid cancers. Mol Cancer. (2023) 22:58. doi: 10.1186/s12943-023-01725-x
31. Qi J, Sun H, Zhang Y, Wang Z, Xun Z, Li Z, et al. Single-cell and spatial analysis reveal interaction of FAP(+) fibroblasts and SPP1(+) macrophages in colorectal cancer. Nat Commun. (2022) 13:1742. doi: 10.1038/s41467-022-29366-6
32. Zhang Y, Chen H, Mo H, Hu X, Gao R, Zhao Y, et al. Single-cell analyses reveal key immune cell subsets associated with response to PD-L1 blockade in triple-negative breast cancer. Cancer Cell. (2021) 39:1578–93 e8. doi: 10.1016/j.ccell.2021.09.010
33. Ning J, Hou X, Hao J, Zhang W, Shi Y, Huang Y, et al. METTL3 inhibition induced by M2 macrophage-derived extracellular vesicles drives anti-PD-1 therapy resistance via M6A-CD70-mediated immune suppression in thyroid cancer. Cell Death Differ. (2023) 30:2265–79. doi: 10.1038/s41418-023-01217-x
34. You Q, Wang F, Du R, Pi J, Wang H, Huo Y, et al. m(6) A reader YTHDF1-targeting engineered small extracellular vesicles for gastric cancer therapy via epigenetic and immune regulation. Adv Mater. (2023) 35:e2204910. doi: 10.1002/adma.202204910
35. Li J, Wu C, Hu H, Qin G, Wu X, Bai F, et al. Remodeling of the immune and stromal cell compartment by PD-1 blockade in mismatch repair-deficient colorectal cancer. Cancer Cell. (2023) 41:1152–69 e7. doi: 10.1016/j.ccell.2023.04.011
36. Hu J, Zhang L, Xia H, Yan Y, Zhu X, Sun F, et al. Tumor microenvironment remodeling after neoadjuvant immunotherapy in non-small cell lung cancer revealed by single-cell RNA sequencing. Genome Med. (2023) 15:14. doi: 10.1186/s13073-023-01164-9
37. Song CH, Kim N, Nam RH, Choi SI, Jang JY, Kim JW, et al. Combination treatment with 17beta-estradiol and anti-PD-L1 suppresses MC38 tumor growth by reducing PD-L1 expression and enhancing M1 macrophage population in MC38 colon tumor model. Cancer Lett. (2022) 543:215780. doi: 10.1016/j.canlet.2022.215780
38. Choueiri TK, Kluger H, George S, Tykodi SS, Kuzel TM, Perets R, et al. FRACTION-RCC: nivolumab plus ipilimumab for advanced renal cell carcinoma after progression on immuno-oncology therapy. J Immunother Cancer. (2022) 10:e005780. doi: 10.1136/jitc-2022-005780
39. Nywening TM, Wang-Gillam A, Sanford DE, Belt BA, Panni RZ, Cusworth BM, et al. Targeting tumour-associated macrophages with CCR2 inhibition in combination with FOLFIRINOX in patients with borderline resectable and locally advanced pancreatic cancer: a single-centre, open-label, dose-finding, non-randomised, phase 1b trial. Lancet Oncol. (2016) 17:651–62. doi: 10.1016/S1470-2045(16)00078-4
40. Haag GM, Springfeld C, Grun B, Apostolidis L, Zschabitz S, Dietrich M, et al. Pembrolizumab and maraviroc in refractory mismatch repair proficient/microsatellite-stable metastatic colorectal cancer - The PICCASSO phase I trial. Eur J Cancer. (2022) 167:112–22. doi: 10.1016/j.ejca.2022.03.017
41. Gilbert MR, Dignam JJ, Armstrong TS, Wefel JS, Blumenthal DT, Vogelbaum MA, et al. A randomized trial of bevacizumab for newly diagnosed glioblastoma. N Engl J Med. (2014) 370:699–708. doi: 10.1056/NEJMoa1308573
42. Cassier PA, Italiano A, Gomez-Roca C, Le Tourneau C, Toulmonde M, D'Angelo SP, et al. Long-term clinical activity, safety and patient-reported quality of life for emactuzumab-treated patients with diffuse-type tenosynovial giant-cell tumour. Eur J Cancer. (2020) 141:162–70. doi: 10.1016/j.ejca.2020.09.038
43. Gomez-Roca C, Cassier P, Zamarin D, Machiels JP, Perez Gracia JL, Stephen Hodi F, et al. Anti-CSF-1R emactuzumab in combination with anti-PD-L1 atezolizumab in advanced solid tumor patients naive or experienced for immune checkpoint blockade. J Immunother Cancer. (2022) 10:e004076. doi: 10.1136/jitc-2021-004076
44. Machiels JP, Gomez-Roca C, Michot JM, Zamarin D, Mitchell T, Catala G, et al. Phase Ib study of anti-CSF-1R antibody emactuzumab in combination with CD40 agonist selicrelumab in advanced solid tumor patients. J Immunother Cancer. (2020) 8:e001153. doi: 10.1136/jitc-2020-001153
45. Razak AR, Cleary JM, Moreno V, Boyer M, Calvo Aller E, Edenfield W, et al. Safety and efficacy of AMG 820, an anti-colony-stimulating factor 1 receptor antibody, in combination with pembrolizumab in adults with advanced solid tumors. J Immunother Cancer. (2020) 8:e001006. doi: 10.1136/jitc-2020-001006
46. Johnson M, Dudek AZ, Sukari A, Call J, Kunk PR, Lewis K, et al. ARRY-382 in combination with pembrolizumab in patients with advanced solid tumors: results from a phase 1b/2 study. Clin Cancer Res. (2022) 28:2517–26. doi: 10.1158/1078-0432.CCR-21-3009
47. Vonderheide RH, Burg JM, Mick R, Trosko JA, Li D, Shaik MN, et al. Phase I study of the CD40 agonist antibody CP-870,893 combined with carboplatin and paclitaxel in patients with advanced solid tumors. Oncoimmunology. (2013) 2:e23033. doi: 10.4161/onci.23033
48. Bauer C, Kuhnemuth B, Duewell P, Ormanns S, Gress T, Schnurr M. Prevailing over T cell exhaustion: New developments in the immunotherapy of pancreatic cancer. Cancer Lett. (2016) 381:259–68. doi: 10.1016/j.canlet.2016.02.057
49. Liaw K, Reddy R, Sharma A, Li J, Chang M, Sharma R, et al. Targeted systemic dendrimer delivery of CSF-1R inhibitor to tumor-associated macrophages improves outcomes in orthotopic glioblastoma. Bioeng Transl Med. (2021) 6:e10205. doi: 10.1002/btm2.10205
50. Siddiqui BA, Chapin BF, Jindal S, Duan F, Basu S, Yadav SS, et al. Immune and pathologic responses in patients with localized prostate cancer who received daratumumab (anti-CD38) or edicotinib (CSF-1R inhibitor). J Immunother Cancer. (2023) 11:e006262. doi: 10.1136/jitc-2022-006262
51. Wiehagen KR, Girgis NM, Yamada DH, Smith AA, Chan SR, Grewal IS, et al. Combination of CD40 agonism and CSF-1R blockade reconditions tumor-associated macrophages and drives potent antitumor immunity. Cancer Immunol Res. (2017) 5:1109–21. doi: 10.1158/2326-6066.CIR-17-0258
52. Omstead AN, Paskewicz M, Gorbunova A, Zheng P, Salvitti MS, Mansoor R, et al. CSF-1R inhibitor, pexidartinib, sensitizes esophageal adenocarcinoma to PD-1 immune checkpoint blockade in a rat model. Carcinogenesis. (2022) 43:842–50. doi: 10.1093/carcin/bgac043
53. Yang H, Zhang Q, Xu M, Wang L, Chen X, Feng Y, et al. CCL2-CCR2 axis recruits tumor associated macrophages to induce immune evasion through PD-1 signaling in esophageal carcinogenesis. Mol Cancer. (2020) 19:41. doi: 10.1186/s12943-020-01165-x
54. Kim D, An L, Moon J, Maymi VI, McGurk AI, Rudd BD, et al. Ccr2+ Monocyte-derived macrophages influence trajectories of acquired therapy resistance in braf-mutant melanoma. Cancer Res. (2023) 83:2328–44. doi: 10.1158/0008-5472.CAN-22-2841
55. Miyamoto T, Murakami R, Hamanishi J, Tanigaki K, Hosoe Y, Mise N, et al. B7-H3 suppresses antitumor immunity via the CCL2-CCR2-M2 macrophage axis and contributes to ovarian cancer progression. Cancer Immunol Res. (2022) 10:56–69. doi: 10.1158/2326-6066.CIR-21-0407
56. Trac N, Chen LY, Zhang A, Liao CP, Poon C, Wang J, et al. CCR2-targeted micelles for anti-cancer peptide delivery and immune stimulation. J Control Release. (2021) 329:614–23. doi: 10.1016/j.jconrel.2020.09.054
57. Wang D, Wang X, Si M, Yang J, Sun S, Wu H, et al. Exosome-encapsulated miRNAs contribute to CXCL12/CXCR4-induced liver metastasis of colorectal cancer by enhancing M2 polarization of macrophages. Cancer Lett. (2020) 474:36–52. doi: 10.1016/j.canlet.2020.01.005
58. Kocher F, Puccini A, Untergasser G, Martowicz A, Zimmer K, Pircher A, et al. Multi-omic characterization of pancreatic ductal adenocarcinoma relates CXCR4 mRNA expression levels to potential clinical targets. Clin Cancer Res. (2022) 28:4957–67. doi: 10.1158/1078-0432.CCR-22-0275
59. Qiao L, Dong C, Jia W, Ma B. Exosomal miR-655-3p inhibits growth, and invasion and macrophage M2 polarization through targeting CXCR4 in papillary thyroid carcinoma. Acta Biochim Pol. (2022) 69:773–9. doi: 10.18388/abp.2020_6027
60. Thapa K, Khan H, Kaur G, Kumar P, Singh TG. Therapeutic targeting of angiopoietins in tumor angiogenesis and cancer development. Biochem Biophys Res Commun. (2023) 687:149130. doi: 10.1016/j.bbrc.2023.149130
61. Yang X, Liu X, Li J, Zhang P, Li H, Chen G, et al. Caerin 1.1/1.9 enhances antitumour immunity by activating the IFN-alpha response signalling pathway of tumour macrophages. Cancers (Basel). (2022) 14:5785. doi: 10.3390/cancers14235785
62. Guerriero JL, Sotayo A, Ponichtera HE, Castrillon JA, Pourzia AL, SChad S, et al. Class IIa HDAC inhibition reduces breast tumours and metastases through anti-tumour macrophages. Nature. (2017) 543:428–32. doi: 10.1038/nature21409
63. Cassetta L, Pollard JW. Repolarizing macrophages improves breast cancer therapy. Cell Res. (2017) 27:963–4. doi: 10.1038/cr.2017.63
64. Goulielmaki E, Bermudez-Brito M, Andreou M, Tzenaki N, Tzardi M, de Bree E, et al. Pharmacological inactivation of the PI3K p110delta prevents breast tumour progression by targeting cancer cells and macrophages. Cell Death Dis. (2018) 9:678. doi: 10.1038/s41419-018-0717-4
65. Cheng Y, Bai F, Ren X, Sun R, Guo X, Liu W, et al. Phosphoinositide-binding protein TIPE1 promotes alternative activation of macrophages and tumor progression via PIP3/akt/TGFbeta axis. Cancer Res. (2022) 82:1603–16. doi: 10.1158/0008-5472.CAN-21-0003
66. Zhang X, Zhu L, Zhang H, Chen S, Xiao Y. CAR-T cell therapy in hematological Malignancies: current opportunities and challenges. Front Immunol. (2022) 13:927153. doi: 10.3389/fimmu.2022.927153
67. Jogalekar MP, Rajendran RL, Khan F, Dmello C, Gangadaran P, Ahn BC. CAR T-Cell-Based gene therapy for cancers: new perspectives, challenges, and clinical developments. Front Immunol. (2022) 13:925985. doi: 10.3389/fimmu.2022.925985
68. Zhang L, Tian L, Dai X, Yu H, Wang J, Lei A, et al. Pluripotent stem cell-derived CAR-macrophage cells with antigen-dependent anti-cancer cell functions. J Hematol Oncol. (2020) 13:153. doi: 10.1186/s13045-020-00983-2
69. Klichinsky M, Ruella M, Shestova O, Lu XM, Best A, Zeeman M, et al. Human chimeric antigen receptor macrophages for cancer immunotherapy. Nat Biotechnol. (2020) 38:947–53. doi: 10.1038/s41587-020-0462-y
70. Tichet M, Wullschleger S, Chryplewicz A, Fournier N, Marcone R, Kauzlaric A, et al. Bispecific PD1-IL2v and anti-PD-L1 break tumor immunity resistance by enhancing stem-like tumor-reactive CD8(+) T cells and reprogramming macrophages. Immunity. (2023) 56:162–79 e6. doi: 10.1016/j.immuni.2022.12.006
71. Wang YN, Wang YY, Wang J, Bai WJ, Miao NJ, Wang J. Vinblastine resets tumor-associated macrophages toward M1 phenotype and promotes antitumor immune response. J Immunother Cancer. (2023) 11:e007253. doi: 10.1136/jitc-2023-007253
72. Zhou C, Weng J, Liu C, Liu S, Hu Z, Xie X, et al. Disruption of SLFN11 deficiency-induced CCL2 signaling and macrophage M2 polarization potentiates anti-PD-1 therapy efficacy in hepatocellular carcinoma. Gastroenterology. (2023) 164:1261–78. doi: 10.1053/j.gastro.2023.02.005
Keywords: macrophages, tumor microenvironment, cancer, immunotherapy, PD-L1
Citation: Zhou L, Zhao T, Zhang R, Chen C and Li J (2024) New insights into the role of macrophages in cancer immunotherapy. Front. Immunol. 15:1381225. doi: 10.3389/fimmu.2024.1381225
Received: 03 February 2024; Accepted: 18 March 2024;
Published: 28 March 2024.
Edited by:
David Entenberg, Albert Einstein College of Medicine, United StatesReviewed by:
Remya Raja, Mayo Clinic Arizona, United StatesCopyright © 2024 Zhou, Zhao, Zhang, Chen and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Jiwei Li, bGlqaXdlaTIwMjFAY3N1LmVkdS5jbg==
†These authors have contributed equally to this work