 Romy Kronstein-Wiedemann1,2*Madeleine Teichert2Elisa Michel3
Romy Kronstein-Wiedemann1,2*Madeleine Teichert2Elisa Michel3 Janina Berg3George Robinson1Kristin Tausche4,5Martin Kolditz4,5Johannes Bergleiter6Jessica Thiel1,2Dirk Koschel4,5,7Stephan R. Künzel1,2Kristina Hölig1,2,6
Janina Berg3George Robinson1Kristin Tausche4,5Martin Kolditz4,5Johannes Bergleiter6Jessica Thiel1,2Dirk Koschel4,5,7Stephan R. Künzel1,2Kristina Hölig1,2,6 Torsten Tonn1,2,8
Torsten Tonn1,2,8 Manuela Rossol3,9*
Manuela Rossol3,9*- 1Laboratory for Experimental Transfusion Medicine, Transfusion Medicine, Faculty of Medicine Carl Gustav Carus, Technische Universität Dresden, Dresden, Germany
- 2Institute for Transfusion Medicine, German Red Cross Blood Donation Service North-East, Dresden, Germany
- 3Molecular Immunology, Faculty of Health Sciences, Brandenburgische Technische Universität (BTU) Cottbus-Senftenberg, Senftenberg, Germany
- 4Division of Pneumology, Medical Department I, University Hospital Carl Gustav Carus and Technische Universität (TU) Dresden, Dresden, Germany
- 5East German Lung Center/Ostdeutsches Lungenzentrum, Dresden-Coswig, Germany
- 6Department of Transfusion Medicine, Medical Clinic and Polyclinic I, University Hospital Carl Gustav Carus, Dresden, Germany
- 7Department of Internal Medicine and Pneumology, Fachkrankenhaus Coswig, Lung Center, Coswig, Germany
- 8Institute for Transfusion Medicine and Immunohematology, Goethe University Hospital Medical School, German Red Cross Blood Donor Service, Frankfurt, Germany
- 9Faculty of Environment and Natural Sciences, Brandenburgische Technische Universität (BTU) Cottbus-Senftenberg, Senftenberg, Germany
Introduction: 1-5% of all patients with COVID-19, a disease caused by infection with Severe Acute Respiratory Syndrome Virus 2 (SARS-Cov-2), even those with mild COVID-19 symptoms, continue to have symptoms after initial recovery. Symptoms associated with the post-acute sequelae of COVID-19 (PACS) include, among others, fatigue, shortness of breath, cough, and cognitive dysfunction. Since the dysregulated immune response appears to be caused by the sustained activation of certain immune cells, including monocytes, and the release of specific cytokines, the aim of our study was to investigate the effect of PACS disease on monocyte subpopulations.
Methods: Twenty-two healthy and thirty-two patients with PACS were included into this study. We performed blood gas analysis and measured hematological parameters from peripheral blood of PACS patients and compared them with healthy donors. Surface markers to identify monocyte subpopulations were analyzed by flow cytometry.
Results: PACS patients had higher numbers of intermediate and CD56+ monocytes, whereas the numbers of total monocytes, classical and non-classical monocytes were normal compared to healthy donors. Comparison of patients with and without fatigue, cough, and dyspnea showed no difference in monocyte subset frequencies. However, patients with cognitive dysfunction had increased numbers of non-classical monocytes compared to patients without this symptom.
Discussion: This suggests a disturbed homeostasis of the monocyte subsets in the peripheral blood of patients with PACS.
Introduction
1-5% of all people infected with severe acute respiratory syndrome coronavirus 2 (SARS-CoV-2) develop long-lasting symptoms, collectively referred to as post-acute sequelae of SARS-CoV-2 infection (PACS) or Long-COVID (1–3). Symptoms include fatigue, cognitive dysfunction, and pulmonary complications, such as shortness of breath (dyspnea) and chronic cough (4). In addition, PACS patients show a persistent low grade inflammation (5–8). Overall, the symptoms are similar to other post-infectious conditions (9, 10). As patients with PACS vary considerably in terms of symptoms, severity and recovery profile, attempts have been made to distinguish different clinical phenotypes, but no consensus has yet been reached (11, 12) and the underlying cause of PACS symptoms has remained a mystery.
Human monocytes can be divided into three major populations: classical (CD14++CD16-), intermediate (CD14++CD16+), and non-classical (CD14+CD16+) monocytes (13), of which classical monocytes in particular express high levels of the ACE-2 receptor (14), which is one of the cellular entry receptor for SARS-CoV-2 (15). Monocytes and macrophages are involved in all phases of the response to viral infections (16–18). Infection with SARS-CoV-2 leads to an altered monocyte phenotype in the acute phase (19–23), and the alterations can predict the severity of the disease (24, 25). Monocytes are also implicated in the pathogenesis of PACS. PACS patients have elevated monocyte-platelet aggregates (7), dysregulated monocyte subpopulations (5, 26–28), show signs of activation (8, 28), and might be a reservoir for persistent infection (27).
In this study, we analyzed peripheral blood monocyte subpopulations in PACS patients. We found increased absolute numbers of intermediate monocytes and CD56+ monocytes. In addition, non-classical monocytes were increased in PACS patients with cognitive dysfunction and decreased in PACS patients without cognitive dysfunction compared to healthy controls. Moreover, D-dimer concentrations were higher in PACS patients with cognitive dysfunction than in patients without this symptom. The monocyte compartment alterations in PACS patients suggest a dysregulation of monocyte fate which might contribute to the persistence of Long-COVID symptoms.
Materials and methods
Patients and controls
Blood samples from 32 PACS patients were provided by the Post-COVID center of the Department of Pulmonology, Medical Department I (University Hospital Carl Gustav Carus, Dresden, Germany). The patients were referred because they showed persisting symptoms after SRAS-CoV-2 infection, and the diagnosis of PACS was made according to the German national guideline for Long-COVID and included documentation of previous SARS-CoV-2 infection, standardized symptom and functional assessment, and standardized exclusion of other causes (29). All patients suffered previously from mild COVID-19 infection according to World Health Organization (WHO) criteria for the clinical management of COVID-19. Fatigue was assessed using the Fatigue Severity Scale (FSS) and the Fatigue Assessment Scale (FAS). The FSS has been shown to demonstrate high internal consistency, validity and sensitivity to changing clinical conditions (30). The FAS questionnaire consists of 10 items answered on a 5-point Likert scale ranging from 1 (never) to 5 (always) (31). Sleepiness was measured using the Epworth Sleepiness Score (ESS). The ESS is a widely used, validated questionnaire and has been shown to be a reliable measure of persistent daytime sleepiness in adults (32). The PACS symptom cognitive dysfunction was self-reported by the patients and was included in the analysis with present or not present.
Age- and sex-matched blood samples from healthy donors (n=22) were provided by the German Red Cross Blood Donation Service North-East, Institute for Transfusion Medicine Dresden and by the Department of Pulmonology, Medical Department I (University Hospital Carl Gustav Carus, Dresden, Germany).
Blood collection and sampling
Six ml whole blood from healthy donors and from Long-COVID patients were collected in S-Monovettes EDTA K3 (Sarstedt, Nümbrecht, Germany) by venipuncture using a sterile disposable Safety-Multifly-Needle 21 G (Sarstedt). Blood cell numbers were determined using a Sysmex XN 1000 (Sysmex Deutschland GmbH, Norderstedt, Germany). The normal reference range for monocyte numbers is 200–1000 monocytes per µl blood. Measurement of COHb was carried out as part of the blood gas analyzing with an ABL800 Flex (Radiometer Medical ApS, Brønshø, Denmark). D-dimer was measured using the STA R Max® (Stago, Düsseldorf, Germany) and blood was collected in citrate monovettes (Sarstedt).
Ethics approval
Blood samples from all donors and Long-COVID patients were used in anonymized form and in accordance with the guidelines approved by the Ethics Committee of the Technical University of Dresden [BO-EK-49012022]. Informed consent was obtained from all donors and patients.
Flow cytometry
50µL whole blood was incubated with PE-labeled anti-CD14 (clone MφP9, BD Bioscience, Heidelberg, Germany), APC-labeled anti-CD56 (clone NCAM16.2, BD Bioscience) and BV421-labeled anti-CD16 (clone 3G8, BD Bioscience) for 15 minutes at room temperature. Samples were then incubated in 450µL BD FACS™ Lysing Solution for 15 min, vortexed and analyzed by flow cytometry using a FACSLyric flow cytometer (BD Bioscience) with FlowJo software. The gating strategy to identify the monocyte subsets within all CD14+ monocytes has been described previously (33). Absolute numbers of monocyte subsets were calculated using the absolute monocyte count of the blood.
Graphs and statistics
Graphs and statistics were prepared with GraphPad Prism 10.2.2. Bar charts represent mean + s.e.m. and individual values of each experiment are represented as symbols in bars. Statistical significance was determined accordingly using the two-tailed non-parametric, unpaired Mann-Whitney U tests, confidence interval of 95%. For multiple comparisons, the Kruskall-Wallis test with Dunn’s test was performed and adjusted p-values were used.
Results
The clinical characteristics of patients with post-acute sequelae of COVID-19 (PACS) and healthy donors are presented in Table 1.

Table 1. Clinical characteristics of the study participants.
Absolute monocyte numbers in the peripheral blood were indistinguishable between healthy controls and PACS patients (Figure 1a). Comparisons of patients with and without fatigue (Figure 1b), cough (Figure 1c), dyspnea (Figure 1d), and cognitive dysfunction (Figure 1e) revealed that total monocytes were not different between the two groups.
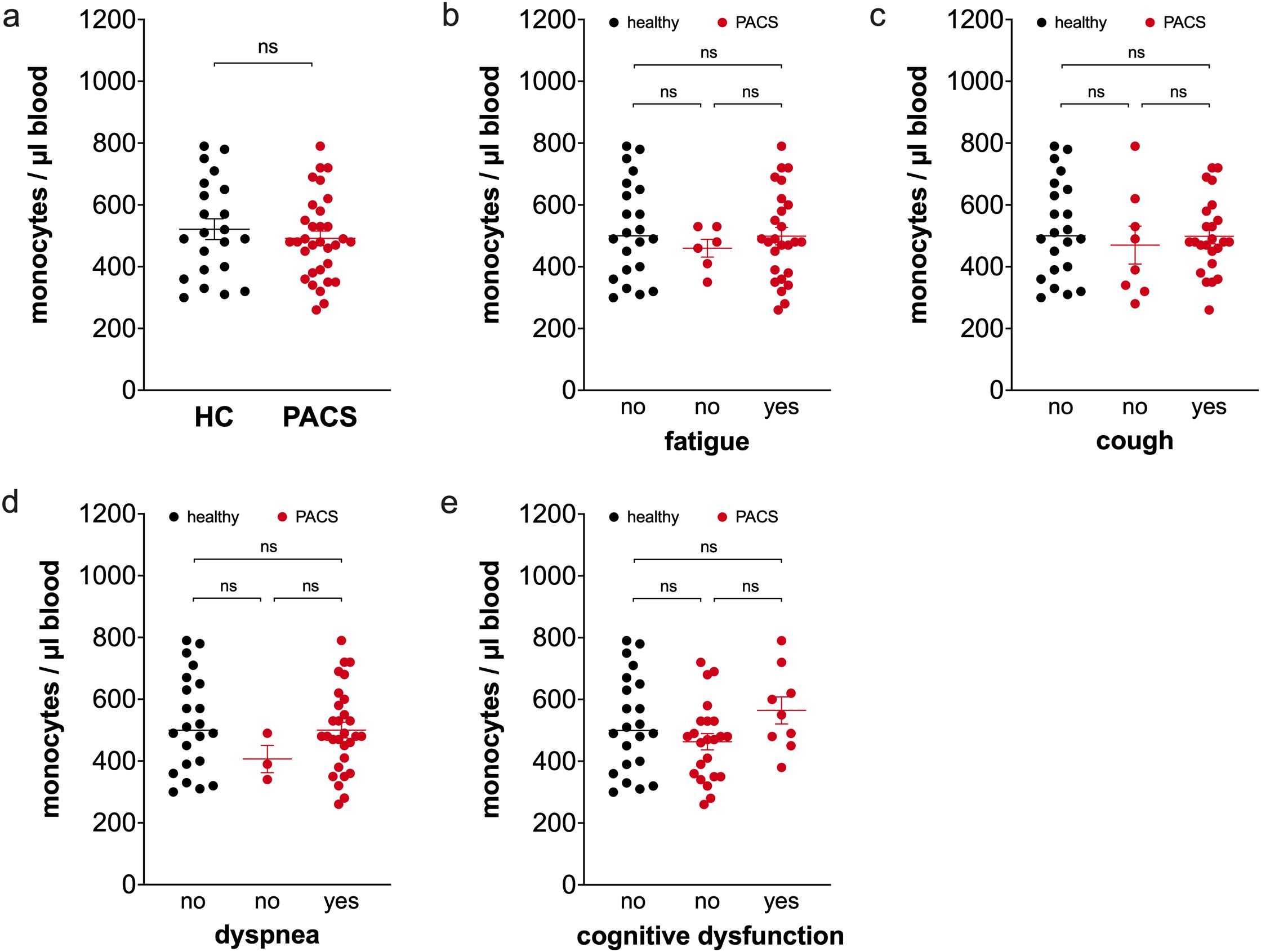
Figure 1. Comparison of total monocyte numbers of healthy controls and PACS patients. (a) Absolute numbers of total monocytes of healthy controls (HC, n = 22) and PACS patients (PACS, n = 32). Scatter plots show mean ± SEM. (b) Absolute numbers of total monocytes of healthy controls (HC, n = 22) and PACS patients without fatigue (n = 6) and with fatigue (n = 26). (c) Absolute numbers of total monocytes of healthy controls (HC, n = 22) and PACS patients without cough (n = 8) and with cough (n = 24). (d) Absolute numbers of total monocytes of healthy controls (HC, n = 22) and PACS patients without dyspnea (n = 3) and with dyspnea (n = 29). (e) Absolute numbers of total monocytes of healthy controls (HC, n = 22) and PACS patients without cognitive dysfunction (n = 23) and with cognitive dysfunction (n = 9). Scatter plots show mean ± SEM. Statistical analysis was performed using two-tailed Mann–Whitney U test (a) or Kruskall-Wallis test with Dunn’s test (b-e). ns, not significant.
PACS patients and healthy controls had equal percentages of classical monocytes (86.6% ± 0.9 vs. 86.9% ± 0.9, ns) in total monocytes. Calculations of absolute numbers of classical monocytes also showed no difference between PACS patients and controls (Figure 2a). PACS patients with fatigue (Figure 2b), with cough (Figure 2c), with dyspnea (Figure 2d), or with cognitive dysfunction (Figure 2e) had the same classical monocyte numbers than patients without the respective symptoms.

Figure 2. Comparison of classical monocyte numbers of healthy controls and PACS patients. (a) Absolute numbers of classical monocytes of healthy controls (HC, n = 22) and PACS patients (PACS, n = 32). Scatter plots show mean ± SEM. (b) Absolute numbers of classical monocytes of healthy controls (HC, n = 22) and PACS patients without fatigue (n = 6) and with fatigue (n = 26). (c) Absolute numbers of classical monocytes of healthy controls (HC, n = 22) and PACS patients without cough (n = 8) and with cough (n = 24). (d) Absolute numbers of classical monocytes of healthy controls (HC, n = 22) and PACS patients without dyspnea (n = 3) and with dyspnea (n = 29). (e) Absolute numbers of classical monocytes of healthy controls (HC, n = 22) and PACS patients without cognitive dysfunction (n = 23) and with cognitive dysfunction (n = 9). Scatter plots show mean ± SEM. Statistical analysis was performed using two-tailed Mann–Whitney U test (a) or Kruskall-Wallis test with Dunn’s test (b-e). ns, not significant.
PACS patients had a higher percentage of intermediate monocytes in total monocytes than healthy controls (6.0% ± 0.4 vs. 3.2% ± 0.3, p<0.0001), and as shown in Figure 3a the absolute number of intermediate monocytes was also increased. This expansion of intermediate monocytes is independent of PACS symptoms fatigue (Figure 3b), cough (Figure 3c), dyspnea (Figure 3d), and cognitive dysfunction (Figure 3e). We observed no difference in the number of intermediate monocytes between PACS patients with a disease duration >2 years and <2 years (median 26 cells/µl blood, n=19 vs. median 26 cells/µl blood, n=13, ns).

Figure 3. Comparison of intermediate monocyte numbers of healthy controls and PACS patients. (a) Absolute numbers of intermediate monocytes of healthy controls (HC, n = 22) and PACS patients (PACS, n = 32). Scatter plots show mean ± SEM. (b) Absolute numbers of intermediate monocytes of healthy controls (HC, n = 22) and PACS patients without fatigue (n = 6) and with fatigue (n = 26). (c) Absolute numbers of intermediate monocytes of healthy controls (HC, n = 22) and PACS patients without cough (n = 8) and with cough (n = 24). (d) Absolute numbers of intermediate monocytes of healthy controls (HC, n = 22) and PACS patients without dyspnea (n = 3) and with dyspnea (n = 29). (e) Absolute numbers of intermediate monocytes of healthy controls (HC, n = 22) and PACS patients without cognitive dysfunction (n = 23) and with cognitive dysfunction (n = 9). Scatter plots show mean ± SEM. Statistical analysis was performed using two-tailed Mann–Whitney U test (a) or Kruskall-Wallis test with Dunn’s test (b-e). ns, not significant.
The percentage of non-classical monocytes in total monocytes was not different between PACS patients and healthy controls (5.0% ± 0.7 vs. 4.9 ± 0.8, ns). The absolute numbers of non-classical monocytes also did not differ between PACS patients and healthy controls (Figure 4a). We also observed equal non-classical monocyte numbers in PACS patients with and without fatigue (Figure 4b), cough (Figure 4c), and dyspnea (Figure 4d). However, we observed a striking difference between PACS patients with cognitive dysfunction and without (Figure 4e). PACS patients with cognitive dysfunction had more non-classical monocytes than PACS patients without cognitive dysfunction and healthy controls.

Figure 4. Comparison of non-classical monocyte numbers of healthy controls and PACS patients. (a) Absolute numbers of non-classical monocytes (NCM) of healthy controls (HC, n = 22) and PACS patients (PACS, n = 32). Scatter plots show mean ± SEM. (b) Absolute numbers of non-classical monocytes of healthy controls (HC, n = 22) and PACS patients without fatigue (n = 6) and with fatigue (n = 26). (c) Absolute numbers of non-classical monocytes of healthy controls (HC, n = 22) and PACS patients without cough (n = 8) and with cough (n = 24). (d) Absolute numbers of non-classical monocytes of healthy controls (HC, n = 22) and PACS patients without dyspnea (n = 3) and with dyspnea (n = 29). (e) Absolute numbers of non-classical monocytes of healthy controls (HC, n = 22) and PACS patients without cognitive dysfunction (n = 23) and with cognitive dysfunction (n = 9). (f) D-dimer concentration in the serum of PACS patients without cognitive dysfunction (n = 23) and with cognitive dysfunction (n = 9). (a-f) Scatter plots show mean ± SEM. Statistical analysis was performed using two-tailed Mann–Whitney U test (a, f) or Kruskall-Wallis test with Dunn’s test (b-e). (g) Correlation of absolute numbers of non-classical monocytes of PACS patients without cognitive dysfunction with carboxylated hemoglobin (COHb) (n = 23). Spearman correlation coefficient and level of significance as indicated. ns, not significant.
It has been previously reported that D-dimer levels in acute COVID-19 predict the development if cognitive dysfunction in PACS patients (34). We analyzed D-dimer levels in our cohort and found increased concentrations in the blood of PACS patients with cognitive dysfunction in comparison to patients without (Figure 4f). However, only two patients with cognitive dysfunction and one patient without cognitive dysfunction had clinical relevant D-dimer concentrations above the cutoff of 500 µg/ml. Carbon monoxide binding to hemoglobin, previously described by us to be elevated in PACS patients (35), negatively correlated strongly with non-classical monocyte numbers in PACS patients without cognitive dysfunction (Figure 4g), whereas no correlation was observed in PACS patients with cognitive dysfunction (Supplementary Figure 1).
We also included the CD56+ monocyte subpopulation in our analysis. This subpopulation is part of the classical monocyte subpopulation and expands during aging, obesity, and in autoimmune diseases (33, 36, 37). PACS patients had a higher percentage of CD56+ monocytes (10.5% ± 1.4 vs. 5.2% ± 0.7, p<0.0001) in total monocytes than healthy controls. Calculations of absolute numbers of CD56+ monocytes revealed that PACS patients had more CD56+ monocytes than healthy controls (Figure 5a). We reported previously that the CD56+ monocyte subpopulation expands with age (36). We observed this again in the healthy controls (Supplementary Figure 2a), but there was no correlation of CD56+ monocytes and age within the PACS cohort (Supplementary Figure 2b). The CD56+ monocytes were already expanded in young PACS patients compared to age-matched controls (median 34 cells/µl blood, n=11 vs. 16 cell/µl blood, n=7, p = 0.0121, age <40 years). The number of CD56+ monocytes was equal between PACS patients with a disease duration >2 years and <2 years (median 37 cells/µl blood, n=19 vs. median 38 cells/µl blood, n=12, ns). The increase in CD56+ monocyte numbers is independent of PACS symptoms fatigue (Figure 5b), cough (Figure 5c), dyspnea (Figure 5d), and cognitive dysfunction (Figure 5e).
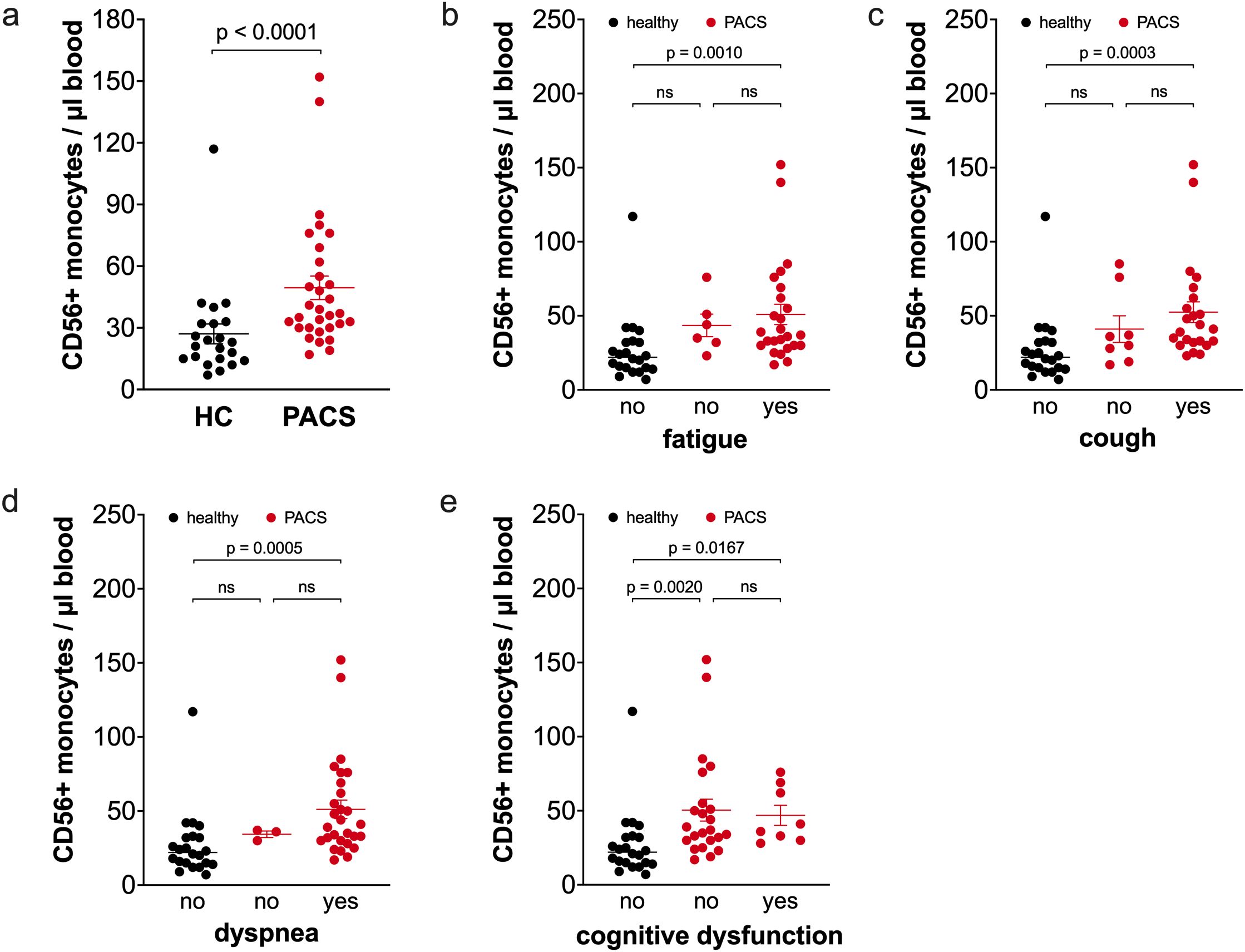
Figure 5. Comparison of CD56+ monocyte numbers of healthy controls and PACS patients. (a) Absolute numbers of CD56+ monocytes of healthy controls (HC, n = 22) and PACS patients (PACS, n = 32). Scatter plots show mean ± SEM. (b) Absolute numbers of CD56+ monocytes of healthy controls (HC, n = 22) and PACS patients without fatigue (n = 6) and with fatigue (n = 26). (c) Absolute numbers of CD56+ monocytes of healthy controls (HC, n = 22) and PACS patients without cough (n = 8) and with cough (n = 24). (d) Absolute numbers of CD56+ monocytes of healthy controls (HC, n = 22) and PACS patients without dyspnea (n = 3) and with dyspnea (n = 29). (e) Absolute numbers of CD56+ monocytes of healthy controls (HC, n = 22) and PACS patients without cognitive dysfunction (n = 23) and with cognitive dysfunction (n = 9). Scatter plots show mean ± SEM. Statistical analysis was performed using two-tailed Mann–Whitney U test (a) or Kruskall-Wallis test with Dunn’s test (b-e). ns, not significant.
Discussion
Our findings indicate that intermediate monocytes and CD56+ monocytes are expanded in PACS patients. These monocyte compartment perturbations were also found in PACS patients with a disease duration greater than two years, suggesting a persistent and long-term dysregulation of the monocyte subsets. Peripheral blood monocyte subpopulations circulate in a dynamic equilibrium (38). Classical monocytes emerge from the bone marrow and circulate for one day. 99% of classical monocytes leave the circulation, the other 1% then transition to intermediate monocytes which circulate for four days. Finally, all intermediate monocytes mature to non-classical monocytes which circulate for seven days. This results in approximately 87-92% classical monocytes, 3-5% intermediate monocytes, and 5-8% non-classical monocytes in the peripheral blood monocyte pool. In a state of inflammation, the differentiation from classical monocytes into intermediate and non-classical monocytes can occur over a shorter timespan (38).
Increased numbers of intermediate monocytes in PACS patients have been reported previously. Park et al. reported about increased total monocyte numbers and increased numbers of all three monocyte subpopulations (28), whereas Patterson et al. observed an increased percentage of intermediate monocytes (27). The increase in intermediate monocytes in our PACS cohort was not associated with any specific PACS symptom but was present in all PACS patients. Intermediate monocytes are found in increased proportions during acute Sars-CoV-2 (20–22) infection, other viral infections (39, 40), and many inflammatory diseases like rheumatoid arthritis (41), obesity (33), and sepsis (42). We among others have reported that intermediate monocytes are the main producers of pro-inflammatory cytokines (41, 43). In addition, intermediate monocytes are the main producers of reactive oxygen species (44), they express high levels of HLA-DR, CD80, and CD86 (41, 45), indicating their role in antigen presentation, and CCR5 (41, 43, 45), a chemokine receptor.
With the dynamic equilibrium of the monocyte subsets in the peripheral blood and the subsequent differentiation of all intermediate monocytes into non-classical monocytes (38), we expected to see also an increased proportion of non-classical monocytes. This was only true for PACS patients with cognitive dysfunction, all other patients showed normal non-classical monocyte numbers. While other studies observed increased percentages or numbers of non-classical monocytes in PACS patients (26, 27), it is interesting to note that COVID-19 patients show a depletion of non-classical monocytes in the peripheral blood as well as in the airways (19, 20, 22, 23, 46). Previously, we reported that PACS patients had impaired oxygen-hemoglobin binding and enhanced carbon monoxide binding (35). In the current study, non-classical monocyte numbers in PACS patients without cognitive dysfunction negatively correlated with carboxyhemoglobin (COHb). COHb is also increased in COVID-19 (47), respiratory infections (48), and sepsis (49), and endogenously derived from the metabolism of heme by heme oxygenase.
We observed a striking increase in non-classical monocyte numbers in PACS patients with a cognitive dysfunction. However, it is important to note, as a limitation of our study, that the cognitive dysfunction was a self-reported symptom and no additional tests were performed. In general, there is no objective cognitive marker for PACS. Recently, cognitive slowing was described by Zhao et al. as such a marker that is easy to evaluate using two short web-based cognitive tasks (50). A systematic study on non-classical monocytes and cognitive dysfunction in a larger cohort of PACS patients needs to be done to gain more insight into these findings.
Non-classical monocytes in general are known to be involved in the innate surveillance of tissues and in the response to viruses (43). The murine counterpart of human non-classical monocytes was found to be patrolling the endothelium of blood vessels and be involved in the resolution of inflammation (43, 51). The non-classical monocyte subset has been implicated to play a role in the cognitive impairment of aging people with HIV (52) and in the steady-state immune surveillance of the central nervous system (53). In COVID-19, some patients develop neurological sequelae and those patients have monocytes in the cerebrospinal fluid (54), and PACS patients with cognitive dysfunction also show recruitment of monocytes into the cerebrospinal fluid (55).
D-dimer is a fibrin degradation product and we found increased D-dimer concentrations in PACS patients with cognitive dysfunction in comparison to PACS patients without this symptom. The presence of D-dimers are associated with a cognitive decline in older people (56, 57), and Taquet et al. linked elevated D-dimer relative to C-reactive protein in acute COVID-19 to subjective cognitive deficits and occupational impact in PACS patients (34). There is some evidence for thromboinflammatory dysregulation in PACS patients. Increased D-dimer concentrations have been reported in PACS patients (58–61) as well as coagulation factor 11 (7). PACS patients have circulating fibrinolysis-resistant microclots (62), platelet-monocyte aggregates (7), and persistent complement dysregulation (7).
The combination of unchanged total monocyte numbers, the increase in intermediate monocytes and normal non-classical monocyte numbers in the majority of patients suggest a disturbed homeostasis of the monocyte subsets in the peripheral blood or an increased extravasation of non-classical monocytes in PACS patients. Cervia-Hasler et al. reported about the down-regulation of the transcription factor NR4A1 in classical monocytes of PACS patients (7). NR4A1 is a transcription factor that is necessary for the maturation of non-classical monocytes (63) and a down-regulation could point to a disturbed homeostasis of the monocyte subsets. There is also some evidence for an increased recruitment of monocytes into tissues in PACS patients. Scott et al. describe a monocyte migration profile in PACS patients that would promote migration of monocytes into the lung (64). They found an increased expression of the chemokine receptor CXCR6 on monocytes of PACS patients with unresolved lung injury and the ligand CXCL16 is abundantly expressed in the lung. Cheong et al. showed that monocytes and hematopoietic stem and progenitor cells (HSPC) showed epigenomic reprogramming up until one year following severe COVID-19 (65). The monocytes had a more pronounced pro-inflammatory response, and they found an increased recruitment of monocytes into the lung and brain in a mouse model. The focus of this study was not on PACS patients, however, Cheong et al. re-analyzed data from post-mortem lung tissue from post-acute COVID-19 patients from a study by Rendeiro et al. (66), and found an increased accumulation of monocytes in the lung. Interestingly, Rendeiro et al. observed a persistent presence of SARS-CoV-2 epitopes in the lung of the patients up to 359 days after the acute phase.
We also observed a marked increase of CD56+ monocytes in patients with PACS. The increase of CD56+ monocytes was found in all PACS patients except patients without cough. CD56+ monocytes, a subpopulation within classical monocytes, are expanded during healthy aging (36), in autoimmune diseases such as rheumatoid arthritis (36) and Crohn’s disease (37), in obesity (33), and in cancer (67). They produce more reactive oxygen intermediates and pro-inflammatory cytokines, and are more efficient antigen-presenting cells (36, 37, 68). CD56+ monocytes have also been described in acute COVID-19. Campana et al. observed an increased frequency of CD56+ monocytes in intensive care unit (ICU) patients but not in non-ICU COVID-19 patients (46). They also described them as hyperinflammatory, because the CD56+ monocytes produced more pro-inflammatory cytokines. Dutt et al. also reported an increase of CD56+ monocytes in COVID-19 patients (69). There is not much known about the CD56+ monocyte subset in the convalescence phase. Ravkov et al. observed no difference between convalescent COVID-19 patients and healthy controls, however, they only included convalescent patients who had a mild form of the disease (70).
We have described previously that the CD56+ monocyte subset expands with age (36). We could replicate this finding again in the healthy controls in this study. However, the CD56+ monocyte subset in PACS patients did not show any dependence on age, and more importantly the expansion of the subset was also found in young PACS patients. This could point to a premature aging of the monocytes (71), we observed this also in patients with rheumatoid arthritis (36). Lo Tartaro et al. reported about an increase in CD56+ monocytes in aged patients (>70 years old) with severe COVID-19 pneumonia in comparison to younger patients (<60 years old) and healthy controls (72). However, they also observed increased CD56+ monocyte numbers in the younger patients compared to healthy controls, suggesting that the expansion of the subset is also not age-dependent in acute COVID-19.
The appearance of CD56+ monocytes during aging is most likely caused by the low-grade inflammation, and we have reported about the expanded CD56+ subset in obese patients, another low-grade inflammatory disease (33). A low-grade inflammation has also been described in PACS patients (5, 7). In addition to the effect of low-grade inflammation, the highly inflammatory events during acute COVID-19 can have long-lasting effects to lead to epigenetic re-programming of the monocytes (65, 73).
In conclusion, this study shows that PACS patients have increased numbers of intermediate monocytes and CD56+ monocytes, while non-classical monocyte numbers were found to be increased in PACS patients with cognitive dysfunction. This suggests a disturbed homeostasis of the monocyte subsets in the peripheral blood of patients with PACS.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The studies involving humans were approved by Ethics Committee of the Technical University of Dresden [BO-EK-49012022]. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
RK-W: Writing – review & editing, Conceptualization, Writing – original draft, Formal Analysis. MT: Writing – review & editing, Data curation. EM: Formal Analysis, Writing – review & editing. JaB: Writing – review & editing, Formal Analysis. GR: Writing – review & editing, Formal Analysis. KT: Resources, Writing – review & editing. MK: Writing – review & editing, Resources. JoB: Resources, Writing – review & editing. JT: Writing – review & editing, Formal Analysis. DK: Resources, Writing – review & editing. SK: Writing – review & editing, Resources. KH: Resources, Writing – review & editing. TT: Writing – review & editing, Conceptualization. MR: Visualization, Formal Analysis, Writing – original draft, Conceptualization, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. We thank the German Red Cross Blood Donation Service North-East and the Roland Ernst Foundation (Grant 3/22) for funding this project.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fimmu.2025.1613034/full#supplementary-material
References
1. Tesch F, Ehm F, Loser F, Bechmann L, Vivirito A, Wende D, et al. Post-viral symptoms and conditions are more frequent in COVID-19 than influenza, but not more persistent. BMC Infect Diseases. (2024) 24:1126. doi: 10.1186/s12879-024-10059-y
2. Xie Y, Choi T, and Al-Aly Z. Postacute sequelae of SARS-coV-2 infection in the pre-delta, delta, and omicron eras. N Engl J Med. (2024) 391:515–25. doi: 10.1056/NEJMoa2403211
3. Greenhalgh T, Sivan M, Perlowski A, and Nikolich JZ. Long COVID: a clinical update. Lancet. (2024) 404:707–24. doi: 10.1056/NEJMoa2403211
4. Shah W, Hillman T, Playford ED, and Hishmeh L. Managing the long term effects of covid-19: summary of NICE, SIGN, and RCGP rapid guideline (2021). Available online at: https://www.bmj.com/content/372/bmj.n136.long (Accessed Feb 17, 2025).
5. Berentschot JC, Drexhage HA, Aynekulu Mersha DG, Wijkhuijs AJM, GeurtsvanKessel CH, Koopmans MPG, et al. Immunological profiling in long COVID: overall low grade inflammation and T-lymphocyte senescence and increased monocyte activation correlating with increasing fatigue severity. Front Immunol. (2023) 14:1254899. doi: 10.3389/fimmu.2023.1254899
6. Brambilla M, Fumoso F, Conti M, Becchetti A, Bozzi S, Mencarini T, et al. Low-grade inflammation in long COVID syndrome sustains a persistent platelet activation associated with lung impairment. JACC: Basic to Trans Science. (2025) 10:20–39. doi: 10.1016/j.jacbts.2024.09.007
7. Cervia-Hasler C, Brüningk SC, Hoch T, Fan B, Muzio G, Thompson RC, et al. Persistent complement dysregulation with signs of thromboinflammation in active Long Covid. Science. (2024) 383:eadg7942. doi: 10.1126/science.adg7942
8. Phetsouphanh C, Darley DR, Wilson DB, Howe A, Munier CML, Patel SK, et al. Immunological dysfunction persists for 8 months following initial mild-to-moderate SARS-CoV-2 infection. Nat Immunol. (2022) 23:210–6. doi: 10.1038/s41590-021-01113-x
9. Komaroff AL and Lipkin WI. Insights from myalgic encephalomyelitis/chronic fatigue syndrome may help unravel the pathogenesis of postacute COVID-19 syndrome. Trends Mol Med. (2021) 27:895–906. doi: 10.1016/j.molmed.2021.06.002
10. Kedor C, Freitag H, Meyer-Arndt L, Wittke K, Hanitsch LG, Zoller T, et al. A prospective observational study of post-COVID-19 chronic fatigue syndrome following the first pandemic wave in Germany and biomarkers associated with symptom severity. Nat Commun. (2022) 13:5104. doi: 10.1038/s41467-022-32507-6
11. Deer RR, Rock MA, Vasilevsky N, Carmody L, Rando H, Anzalone AJ, et al. Characterizing long COVID: deep phenotype of a complex condition. EBioMedicine. (2021) 74:103722. doi: 10.1016/j.ebiom.2021.103722
12. Kenny G, McCann K, O’Brien C, Savinelli S, Tinago W, Yousif O, et al. Identification of distinct long COVID clinical phenotypes through cluster analysis of self-reported symptoms. Open Forum Infect Dis. (2022) 9:ofac060. doi: 10.1093/ofid/ofac060
13. Kapellos TS, Bonaguro L, Gemünd I, Reusch N, Saglam A, Hinkley ER, et al. Human monocyte subsets and phenotypes in major chronic inflammatory diseases. Front Immunol. (2019) 10:2035. doi: 10.3389/fimmu.2019.02035
14. Rutkowska-Zapała M, Suski M, Szatanek R, Lenart M, Węglarczyk K, Olszanecki R, et al. Human monocyte subsets exhibit divergent angiotensin I-converting activity. Clin Exp Immunol. (2015) 181:126–32. doi: 10.1111/cei.12612
15. Hoffmann M, Kleine-Weber H, Schroeder S, Krüger N, Herrler T, Erichsen S, et al. SARS-coV-2 cell entry depends on ACE2 and TMPRSS2 and is blocked by a clinically proven protease inhibitor. Cell. (2020) 181:271–280.e8. doi: 10.1016/j.cell.2020.02.052
16. Ruscitti C, Radermecker C, and Marichal T. Journey of monocytes and macrophages upon influenza A virus infection. Curr Opin Virology. (2024) 66:101409. doi: 10.1016/j.coviro.2024.101409
17. Hoeve MA, Nash AA, Jackson D, Randall RE, and Dransfield I. Influenza virus A infection of human monocyte and macrophage subpopulations reveals increased susceptibility associated with cell differentiation. PloS One. (2012) 7:e29443. doi: 10.1371/journal.pone.0029443
18. Movita D, van de Garde MDB, Biesta P, Kreefft K, Haagmans B, Zuniga E, et al. Inflammatory Monocytes Recruited to the Liver within 24 Hours after Virus-Induced Inflammation Resemble Kupffer Cells but Are Functionally Distinct. J Virology. (2015) 89:4809–17. doi: 10.1128/JVI.03733-14
19. Szabo PA, Dogra P, Gray JI, Wells SB, Connors TJ, Weisberg SP, et al. Longitudinal profiling of respiratory and systemic immune responses reveals myeloid cell-driven lung inflammation in severe COVID-19. Immunity. (2021) 54:797–814.e6. doi: 10.1016/j.immuni.2021.03.005
20. Liu N, Jiang C, Cai P, Shen Z, Sun W, Xu H, et al. Single-cell analysis of COVID-19, sepsis, and HIV infection reveals hyperinflammatory and immunosuppressive signatures in monocytes. Cell Rep. (2021) 37:109793. doi: 10.1016/j.celrep.2021.109793
21. Mann ER, Menon M, Knight SB, Konkel JE, Jagger C, Shaw TN, et al. Longitudinal immune profiling reveals key myeloid signatures associated with COVID-19. Sci Immunol. (2020) 5:eabd6197. doi: 10.1126/sciimmunol.abd6197
22. Haschka D, Petzer V, Burkert FR, Fritsche G, Wildner S, Bellmann-Weiler R, et al. Alterations of blood monocyte subset distribution and surface phenotype are linked to infection severity in COVID-19 inpatients. Eur J Immunol. (2022) 52:1285–96. doi: 10.1002/eji.202149680
23. Silvin A, Chapuis N, Dunsmore G, Goubet AG, Dubuisson A, Derosa L, et al. Elevated calprotectin and abnormal myeloid cell subsets discriminate severe from mild COVID-19. Cell. (2020) 182:1401–1418.e18. doi: 10.1016/j.cell.2020.08.002
24. Utrero-Rico A, González-Cuadrado C, Chivite-Lacaba M, Cabrera-Marante O, Laguna-Goya R, Almendro-Vazquez P, et al. Alterations in Circulating Monocytes Predict COVID-19 Severity and Include Chromatin Modifications Still Detectab le Six Months after Recovery. Biomedicines. (2021) 9:1253. doi: 10.3390/biomedicines9091253
25. Marais C, Claude C, Semaan N, Charbel R, Barreault S, Travert B, et al. Myeloid phenotypes in severe COVID-19 predict secondary infection and mortality: a pilot study. Ann Intensive Care. (2021) 11:111. doi: 10.1186/s13613-021-00896-4
26. Klein J, Wood J, Jaycox JR, Dhodapkar RM, Lu P, Gehlhausen JR, et al. Distinguishing features of long COVID identified through immune profiling. Nature. (2023) 623:139–48. doi: 10.1038/s41586-023-06651-y
27. Patterson BK, Francisco EB, Yogendra R, Long E, Pise A, Rodrigues H, et al. Persistence of SARS coV-2 S1 protein in CD16+ Monocytes in post-acute sequelae of COVID-19 (PASC) up to 15 months post-infection. Front Immunol. (2021) 12:746021. doi: 10.3389/fimmu.2021.746021
28. Park J, Dean LS, Jiyarom B, Gangcuangco LM, Shah P, Awamura T, et al. Elevated circulating monocytes and monocyte activation in COVID-19 convalescent individuals. Front Immunol. (2023) 14:1151780. doi: 10.3389/fimmu.2023.1151780
29. Koczulla AR, Ankermann T, Behrends U, Berlit P, Böing S, Brinkmann F, et al. S1 guideline post-COVID/long-COVID. Pneumologie. (2021) 75:869–900. doi: 10.1055/a-1551-9734
30. Krupp LB, LaRocca NG, Muir-Nash J, and Steinberg AD. The fatigue severity scale. Application to patients with multiple sclerosis and systemic lupus erythematosus. Arch Neurol. (1989) 46:1121–3. doi: 10.1001/archneur.1989.00520460115022
31. Michielsen HJ, De Vries J, and Van Heck GL. Psychometric qualities of a brief self-rated fatigue measure: The Fatigue Assessment Scale. J Psychosom Res. (2003) 54:345–52. doi: 10.1016/S0022-3999(02)00392-6
32. Johns MW. A new method for measuring daytime sleepiness: the Epworth sleepiness scale. Sleep. (1991) 14:540–5. doi: 10.1093/sleep/14.6.540
33. Friedrich K, Sommer M, Strobel S, Thrum S, Blüher M, Wagner U, et al. Perturbation of the monocyte compartment in human obesity. Front Immunol. (2019) 10:1874. doi: 10.3389/fimmu.2019.01874
34. Taquet M, Skorniewska Z, Hampshire A, Chalmers JD, Ho LP, Horsley A, et al. Acute blood biomarker profiles predict cognitive deficits 6 and 12 months after COVID-19 hospitalization. Nat Med. (2023) 29:2498–508. doi: 10.1038/s41591-023-02525-y
35. Kronstein-Wiedemann R, Tausche K, Kolditz M, Teichert M, Thiel J, Koschel D, et al. Long-COVID is associated with impaired red blood cell function. Horm Metab Res. (2024) 56:318–23. doi: 10.1055/a-2186-8108
36. Krasselt M, Baerwald C, Wagner U, and Rossol M. CD56+ monocytes have a dysregulated cytokine response to lipopolysaccharide and accumulate in rheumatoid arthritis and immunosenescence. Arthritis Res Ther. (2013) 15:R139. doi: 10.1186/ar4321
37. Grip O, Bredberg A, Lindgren S, and Henriksson G. Increased subpopulations of CD16(+) and CD56(+) blood monocytes in patients with active Crohn’s disease. Inflammation Bowel Dis. (2007) 13:566–72. doi: 10.1002/ibd.20025
38. Patel AA, Zhang Y, Fullerton JN, Boelen L, Rongvaux A, Maini AA, et al. The fate and lifespan of human monocyte subsets in steady state and systemic inflammation. J Exp Medicine. (2017), 20170355. doi: 10.1084/jem.20170355
39. Vangeti S, Falck-Jones S, Yu M, Österberg B, Liu S, Asghar M, et al. Human influenza virus infection elicits distinct patterns of monocyte and dendritic cell mobilization in blood and the nasopharynx. eLife. (2023) 12:e77345. doi: 10.7554/eLife.77345.sa2
40. Prabhu VM, Singh AK, Padwal V, Nagar V, Patil P, and Patel V. Monocyte based correlates of immune activation and viremia in HIV-infected long-term non-progressors. Front Immunol. (2019) 10. doi: 10.3389/fimmu.2019.02849/full
41. Rossol M, Kraus S, Pierer M, Baerwald C, and Wagner U. The CD14brightCD16+ monocyte subset is expanded in rheumatoid arthritis and promotes expansion of the Th17 cell population. Arthritis Rheumatism. (2012) 64:671–7. doi: 10.1002/art.33418
42. Poehlmann H, Schefold JC, Zuckermann-Becker H, Volk HD, and Meisel C. Phenotype changes and impaired function of dendritic cell subsets in patients with sepsis: a prospective observational analysis. Crit Care. (2009) 13:R119. doi: 10.1186/cc7969
43. Cros J, Cagnard N, Woollard K, Patey N, Zhang SY, Senechal B, et al. Human CD14dim monocytes patrol and sense nucleic acids and viruses via TLR7 and TLR8 receptors. Immunity. (2010) 33:375–86. doi: 10.1016/j.immuni.2010.08.012
44. Zawada AM, Rogacev KS, Rotter B, Winter P, Marell RR, Fliser D, et al. SuperSAGE evidence for CD14++CD16+ monocytes as a third monocyte subset. Blood. (2011) 118:e50–61. doi: 10.1182/blood-2011-01-326827
45. Wong KL, Tai JJY, Wong WC, Han H, Sem X, Yeap WH, et al. Gene expression profiling reveals the defining features of the classical, intermediate, and nonclassical human monocyte subsets. Blood. (2011) 118:e16–31. doi: 10.1182/blood-2010-12-326355
46. Campana S, De Pasquale C, Sidoti Migliore G, Pezzino G, Cavaliere R, Venanzi Rullo E, et al. Cutting edge: hyperinflammatory monocytes expressing CD56 abound in severe COVID-19 patients. J Immunol. (2022) 209:655–9. doi: 10.4049/jimmunol.2200021
47. Faisal H, Ali ST, Xu J, Nisar T, Sabawi M, Salazar E, et al. Carboxyhemoglobinemia in critically ill coronavirus disease 2019 patients. J Clin Med. (2021) 10:2731. doi: 10.3390/jcm10122731
48. Yasuda H, Yamaya M, Yanai M, Ohrui T, and Sasaki H. Increased blood carboxyhaemoglobin concentrations in inflammatory pulmonary diseases. Thorax. (2002) 57:779–83. doi: 10.1136/thorax.57.9.779
49. Zegdi R, Perrin D, Burdin M, Boiteau R, and Tenaillon A. Increased endogenous carbon monoxide production in severe sepsis. Intensive Care Med. (2002) 28:793–6. doi: 10.1007/s00134-002-1269-7
50. Zhao S, Martin EM, Reuken PA, Scholcz A, Ganse-Dumrath A, Srowig A, et al. Long COVID is associated with severe cognitive slowing: a multicentre cross-sectional study. eClinicalMedicine. (2024), 68. https://www.thelancet.com/journals/eclinm/article/PIIS2589-5370(24)00013-0/fulltext (Mar 5, 2025).
51. Auffray C, Fogg D, Garfa M, Elain G, Join-Lambert O, Kayal S, et al. Monitoring of blood vessels and tissues by a population of monocytes with patrolling behavior. Science. (2007) 317:666–70. doi: 10.1126/science.1142883
52. Singh MV, Uddin MN, Covacevich Vidalle M, Sutton KR, Boodoo ZD, Peterson AN, et al. Non-classical monocyte levels correlate negatively with HIV-associated cerebral small vessel disease and cognitive performance. Front Cell Infect Microbiol. (2024) 14. doi: 10.3389/fcimb.2024.1405431/full
53. Waschbisch A, Schröder S, Schraudner D, Sammet L, Weksler B, Melms A, et al. Pivotal role for CD16+ Monocytes in immune surveillance of the central nervous system. J Immunol. (2016) 196:1558–67. doi: 10.4049/jimmunol.1501960
54. Heming M, Li X, Räuber S, Mausberg AK, Börsch AL, Hartlehnert M, et al. Neurological manifestations of COVID-19 feature T cell exhaustion and dedifferentiated monocytes in cerebrospinal fluid. Immunity. (2021) 54:164–75. doi: 10.1016/j.immuni.2020.12.011
55. Hu WT, Kaluzova M, Dawson A, Sotelo V, Papas J, Lemenze A, et al. Clinical and CSF single-cell profiling of post-COVID-19 cognitive impairment. Cell Rep Medicine. (2024) 5:101561. doi: 10.1016/j.xcrm.2024.101561
56. Wilson CJ, Cohen HJ, and Pieper CF. Cross-linked fibrin degradation products (D-dimer), plasma cytokines, and cognitive decline in community-dwelling elderly persons. J Am Geriatr Soc. (2003) 51:1374–81. doi: 10.1046/j.1532-5415.2003.51454.x
57. Stott DJ, Robertson M, Rumley A, Welsh P, Sattar N, Packard CJ, et al. Activation of hemostasis and decline in cognitive function in older people. Arteriosclerosis Thrombosis Vasc Biol. (2010) 30:605–11. doi: 10.1161/ATVBAHA.109.199448
58. Moreno-Pérez O, Merino E, Leon-Ramirez JM, Andres M, Ramos JM, Arenas-Jiménez J, et al. Post-acute COVID-19 syndrome. Incidence and risk factors: A Mediterranean cohort study. J Infect. (2021) 82:378–83. doi: 10.1016/j.jinf.2021.01.004
59. Mandal S, Barnett J, Brill SE, Brown JS, Denneny EK, Hare SS, et al. Long-COVID”: a cross-sectional study of persisting symptoms, biomarker and imaging abnormalities following hospitalisation for COVID-19. Thorax. (2021) 76:396–8. doi: 10.1136/thoraxjnl-2020-215818
60. Pasini E, Corsetti G, Romano C, Scarabelli TM, Chen-Scarabelli C, Saravolatz L, et al. Serum metabolic profile in patients with long-covid (PASC) syndrome: clinical implications. Front Med (Lausanne). (2021) 8:714426. doi: 10.3389/fmed.2021.714426
61. Di Gennaro L, Valentini P, Sorrentino S, Ferretti MA, De Candia E, Basso M, et al. Extended coagulation profile of children with Long Covid: a prospective study. Sci Rep. (2022) 12:18392. doi: 10.1038/s41598-022-23168-y
62. Pretorius E, Vlok M, Venter C, Bezuidenhout JA, Laubscher GJ, Steenkamp J, et al. Persistent clotting protein pathology in Long COVID/Post-Acute Sequelae of COVID-19 (PASC) is accompanied by increased levels of antiplasmin. Cardiovasc Diabetol. (2021) 20:172. doi: 10.1186/s12933-021-01359-7
63. Hanna RN, Carlin LM, Hubbeling HG, Nackiewicz D, Green AM, Punt JA, et al. The transcription factor NR4A1 (Nur77) controls bone marrow differentiation and the survival of Ly6C- monocytes. Nat Immunol. (2011) 12:778–85. doi: 10.1038/ni.2063
64. Scott NA, Pearmain L, Knight SB, Brand O, Morgan DJ, Jagger C, et al. Monocyte migration profiles define disease severity in acute COVID-19 and unique features of long COVID. European Respiratory Journal. (2023) 61. Available online at https://erj.ersjournals.com/content/61/5/2202226.
65. Cheong JG, Ravishankar A, Sharma S, Parkhurst CN, Grassmann SA, Wingert CK, et al. Epigenetic memory of coronavirus infection in innate immune cells and their progenitors. Cell. (2023) 186:3882–902. doi: 10.1016/j.cell.2023.07.019
66. Rendeiro AF, Ravichandran H, Kim J, Borczuk AC, Elemento O, and Schwartz RE. Persistent alveolar type 2 dysfunction and lung structural derangement in post-acute COVID-19. medRxiv. (2022) 2022:11.28.22282811. doi: 10.1101/2022.11.28.22282811
67. Papewalis C, Jacobs B, Baran AM, Ehlers M, Stoecklein NH, Willenberg HS, et al. Increased numbers of tumor-lysing monocytes in cancer patients. Mol Cell Endocrinology. (2011) 337:52–61. doi: 10.1016/j.mce.2011.01.020
68. Sconocchia G, Keyvanfar K, El Ouriaghli F, Grube M, Rezvani K, Fujiwara H, et al. Phenotype and function of a CD56+ peripheral blood monocyte. Leukemia. (2005) 19:69–76. doi: 10.1038/sj.leu.2403550
69. Dutt TS, LaVergne SM, Webb TL, Baxter BA, Stromberg S, McFann K, et al. Comprehensive immune profiling reveals CD56+ Monocytes and CD31+ Endothelial cells are increased in severe COVID-19 disease. J Immunol. (2022) 208:685–96. doi: 10.4049/jimmunol.2100830
70. Ravkov EV, Williams ESCP, Elgort M, Barker AP, Planelles V, Spivak AM, et al. Reduced monocyte proportions and responsiveness in convalescent COVID-19 patients. Front Immunol. (2024) 14. doi: 10.3389/fimmu.2023.1329026/full
71. Basu S, Ulbricht Y, and Rossol M. Healthy and premature aging of monocytes and macrophages. Front Immunol. (2025) 16:1506165. doi: 10.3389/fimmu.2025.1506165
72. Lo Tartaro D, Neroni A, Paolini A, Borella R, Mattioli M, Fidanza L, et al. Molecular and cellular immune features of aged patients with severe COVID-19 pneumonia. Commun Biol. (2022) 5:590. doi: 10.1038/s42003-022-03537-z
73. Brauns E, Azouz A, Grimaldi D, Xiao H, Thomas S, Nguyen M, et al. Functional reprogramming of monocytes in patients with acute and convalescent severe COVID-19. JCI Insight. (2022) 7. https://insight.jci.org/articles/view/154183 (Mar 7,2025).
Keywords: monocytes, intermediate monocytes, CD56+ monocytes, COVID-19, PACS, SARS-CoV-2
Citation: Kronstein-Wiedemann R, Teichert M, Michel E, Berg J, Robinson G, Tausche K, Kolditz M, Bergleiter J, Thiel J, Koschel D, Künzel SR, Hölig K, Tonn T and Rossol M (2025) Dysregulated monocyte compartment in PACS patients. Front. Immunol. 16:1613034. doi: 10.3389/fimmu.2025.1613034
Received: 16 April 2025; Accepted: 15 May 2025;
Published: 06 June 2025.
Edited by:
Robert Weissert, University of Regensburg, GermanyReviewed by:
Timothy Hanley, The University of Utah, United StatesEugene Ravkov, ARUP Laboratories, United States
Copyright © 2025 Kronstein-Wiedemann, Teichert, Michel, Berg, Robinson, Tausche, Kolditz, Bergleiter, Thiel, Koschel, Künzel, Hölig, Tonn and Rossol. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Manuela Rossol, bWFudWVsYS5yb3Nzb2xAYi10dS5kZQ==; Romy Kronstein-Wiedemann, ci5rcm9uc3RlaW5AYmx1dHNwZW5kZS5kZQ==