 Gopi Patel1
Gopi Patel1 Robert A. Bonomo2,3,4,5,6*
Robert A. Bonomo2,3,4,5,6*- 1Department of Medicine, Mount Sinai School of Medicine, New York, NY, USA
- 2Research Service, Louis Stokes Cleveland Department of Veterans Affairs Medical Center, Cleveland, OH, USA
- 3Division of Infectious Diseases and HIV Medicine, University Hospitals Case Medical Center, Cleveland, OH, USA
- 4Department of Medicine, Case Western Reserve School of Medicine, Cleveland, OH, USA
- 5Department of Molecular Biology and Microbiology, Case Western Reserve School of Medicine, Cleveland, OH, USA
- 6Department of Pharmacology, Case Western Reserve School of Medicine, Cleveland, OH, USA
Carbapenems, once considered the last line of defense against of serious infections with Enterobacteriaceae, are threatened with extinction. The increasing isolation of carbapenem-resistant Gram-negative pathogens is forcing practitioners to rely on uncertain alternatives. As little as 5 years ago, reports of carbapenem resistance in Enterobacteriaceae, common causes of both community and healthcare-associated infections, were sporadic and primarily limited to case reports, tertiary care centers, intensive care units, and outbreak settings. Carbapenem resistance mediated by β-lactamases, or carbapenemases, has become widespread and with the paucity of reliable antimicrobials available or in development, international focus has shifted to early detection and infection control. However, as reports of Klebsiella pneumoniae carbapenemases, New Delhi metallo-β-lactamase-1, and more recently OXA-48 (oxacillinase-48) become more common and with the conveniences of travel, the assumption that infections with highly resistant Gram-negative pathogens are limited to the infirmed and the heavily antibiotic and healthcare exposed are quickly being dispelled. Herein, we provide a status report describing the increasing challenges clinicians are facing and forecast the “stormy waters” ahead.
Carbapenems are potent and broad-spectrum β-lactam antibiotics traditionally reserved for the treatment of the most serious infections (El-Gamal and Oh, 2010). The emergence and dissemination of carbapenem-resistant Gram-negative pathogens including Pseudomonas aeruginosa, Acinetobacter baumannii, and Enterobacteriaceae is a significant contributor to patient morbidity and mortality (Patel et al., 2008; Schwaber et al., 2008; Lautenbach et al., 2009, 2010; Marchaim et al., 2011). Despite radical efforts in infection control (Schwaber et al., 2011) and improvements in rapid molecular diagnostics (Centers for Disease Control and Prevention, 2009; Nordmann et al., 2012c), carbapenem-resistant Gram-negative bacilli remain a formidable threat as few antimicrobial agents are reliably active and very little is expected to be available in the near future.
Clinicians hold that the increasing prevalence of extended-spectrum β-lactamases (ESBLs) among Klebsiella pneumoniae and Escherichia coli in the 1980s and 1990s contributed to the increased consumption of carbapenems. Experience implied that delayed administration of carbapenems in at-risk patients led to poor clinical outcomes (Paterson and Bonomo, 2005; Endimiani and Paterson, 2007). Thus, carbapenems (i.e., imipenem, meropenem, ertapenem, and doripenem) became vital tools in the treatment of healthcare-associated and severe community-acquired infections. Despite heavy reliance on these agents, carbapenem resistance in Enterobacteriaceae, common causes of both community and healthcare-associated infections, remained rare until the past decade.
Carbapenem resistance among Gram-negative bacteria results from one or more of the following mechanisms: (i) hyperproduction or derepression of Ambler class C β-lactamases (AmpC β-lactamases) or ESBLs (e.g., sulfhydryl variable (SHV), temoneira (TEM), cefotaxime (CTX-M) type β-lactamases) with loss or alteration in outer membrane porins; (ii) augmented drug efflux; (iii) alterations in penicillin binding proteins (PBPs); (iv) carbapenemase production (Patel and Bonomo, 2011). Carbapenemases belong to three molecular classes of β-lactamases, Ambler class A, B, and D (Ambler, 1980; Bush and Jacoby, 2010). Our aim is to provide a status report of the molecular diversity and epidemiology of carbapenemases as well as current and future therapeutics. The increasing public safety concerns associated with organisms harboring these enzymes has created significant turmoil. Regrettably, the situation is critical and our patients are in peril.
Ambler Class a Carbapenemases
Few Ambler class A β-lactamases demonstrate carbapenem-hydrolyzing activity and, up until a decade ago, these were rarely recovered. Class A carbapenemases include: K. pneumoniae carbapenemase (KPC), Guiana extended-spectrum (GES), non-metallo-carbapenemase-A (Nmc-A)/imipenem-resistant (IMI), Serratia marcescens enzyme (SME), serratia fonticola carbapenemase (SFC), and BIC β-lactamases (Table 1; Walther-Rasmussen and Høiby, 2007). With the notable exception of KPCs, the clinical isolation of these types of carbapenemases is relatively limited.
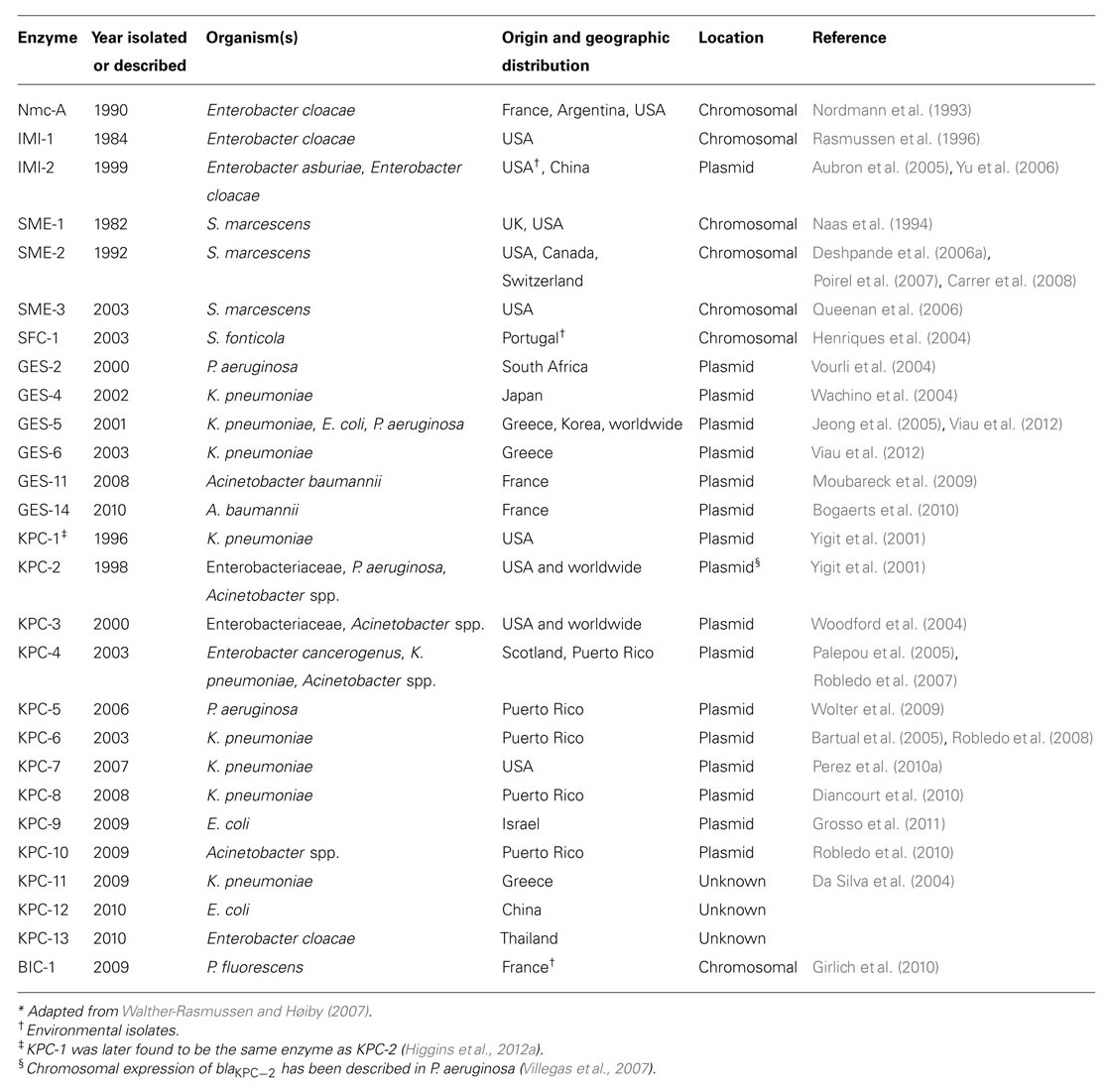
TABLE 1. Class A carbapenemases*.
Non-metallo-carbapenemase-A is a chromosomal carbapenemase originally isolated from Enterobacter cloacae in France (Nordmann et al., 1993). Currently, reports of this particular β-lactamase are still rare (Pottumarthy et al., 2003; Castanheira et al., 2008; Osterblad et al., 2012). IMI-1 was initially recovered from the chromosome of an Enterobacter cloacae isolate in the southwestern USA (Rasmussen et al., 1996). A variant of IMI-1, IMI-2, has been identified on plasmids isolated from environmental strains of Enterobacter asburiae in USA rivers (Aubron et al., 2005).
SME-1 (S. marcescens enzyme) was originally identified in an isolate of S. marcescens from a patient in London in 1982 (Yang et al., 1990). SME-2 and SME-3 were subsequently isolated in the USA, Canada, and Switzerland (Naas et al., 1994; Queenan et al., 2000, 2006; Deshpande et al., 2006b; Poirel et al., 2007; Carrer et al., 2008). Chromosomally encoded SME-type carbapenemases continue to be isolated at a low frequency in North America (Deshpande et al., 2006a,b; Fairfax et al., 2011; Mataseje et al., 2012). Both SFC-1 and BIC-1 are chromosomal serine carbapenemases recovered from environmental isolates. The former from a S. fonticola isolate in Portugal (Henriques et al., 2004) and the latter from Pseudomonas fluorescens isolates recovered from the Seine River (Girlich et al., 2010).
The GES-type β-lactamases are acquired β-lactamases recovered from P. aeruginosa, Enterobacteriaceae, and A. baumannii (Poirel et al., 2000a; Castanheira et al., 2004a). The genes encoding these β-lactamase have often, but not exclusively, been identified within class 1 integrons residing on transferable plasmids (Bonnin et al., 2013; Walther-Rasmussen and Høiby, 2007). GES-1 has a similar hydrolysis profile to other ESBLs, although they essentially spare monobactams. Several GES β-lactamases are described with six (i.e., GES-2, GES-4, GES-5, GES-6, GES-11, and GES-14), demonstrating detectable carbapenemase activity in the setting of amino acid substitutions at their active sites (specifically at residue 104 and 170; Walther-Rasmussen and Høiby, 2007; Kotsakis et al., 2010). These GES-type carbapenemases have been described in Europe, South Africa, Asia, and the Middle East (Poirel et al., 2002; Jeong et al., 2005; da Fonseca et al., 2007; Moubareck et al., 2009; Bonnin et al., 2011, 2013).
Currently, most carbapenem resistance among Enterobacteriaceae in the USA and Israel is attributed to plasmid-mediated expression of a KPC-type carbapenemase (Endimiani et al., 2009b; Nordmann et al., 2009; Gupta et al., 2011; Schwaber et al., 2011). KPC-producing Enterobacteriaceae are considered endemic to Greece along with other carbapenemases, specifically VIM-type [Verona integron-encoded metallo-β-lactamases (MBLs); Canton et al., 2012]. KPCs efficiently hydrolyze carbapenems as well as penicillins, cephalosporins, and aztreonam and are not overcome in vitro by clinically available β-lactamase inhibitors (i.e., clavulanic acid, sulbactam, tazobactam – in fact these are hydrolyzed). These enzymes have been identified in several genera of Enterobacteriaceae as well as Pseudomonas spp. and A. baumannii (Miriagou et al., 2003; Yigit et al., 2003; Bratu et al., 2005; Villegas et al., 2007; Cai et al., 2008; Rasheed et al., 2008; Tibbetts et al., 2008; Robledo et al., 2010; Mathers et al., 2011; Geffen et al., 2012).
Carbapenem resistance secondary to KPC production was first described in a K. pneumoniae recovered in North Carolina in 1996 (Yigit et al., 2001). To date 12 KPC subtypes (KPC-2 to KPC-13; Robledo et al., 2008; Kitchel et al., 2009a; Navon-Venezia et al., 2009; Wolter et al., 2009; Gregory et al., 2010) have been reported with the vast majority of analyzed isolates expressing either KPC-2 or KPC-3.
The blaKPC gene has been mapped to a highly conserved Tn3-based transposon, Tn4401 (Figure 1A), and five isoforms of Tn4401 are described (Naas et al., 2008; Cuzon et al., 2010; Kitchel et al., 2010). Plasmids carrying blaKPC are of various sizes and many carry additional genes conferring resistance to fluoroquinolones and aminoglycosides thus limiting the antibiotics available to treat infections with KPC-producing pathogens (Endimiani et al., 2008; Rice et al., 2008). blaKPC has rarely been mapped to a chromosomal location (Villegas et al., 2007; Castanheira et al., 2009).
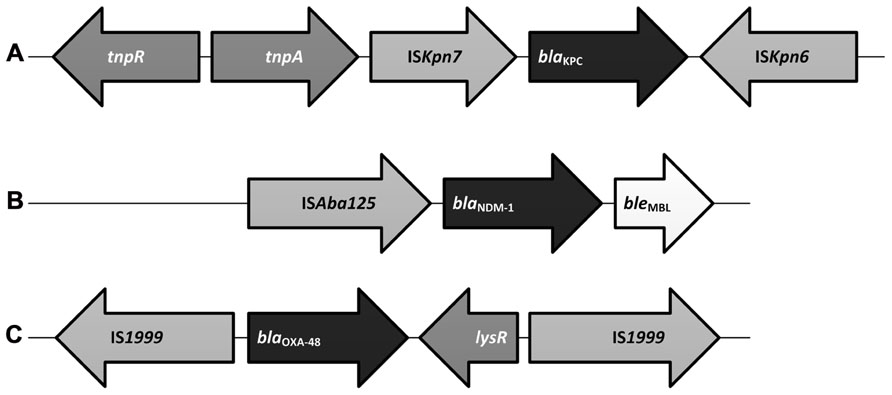
FIGURE 1. Basic genetic construct of select carbapenemase genes. (A) Schematic representation of Tn4401 type of transposon associated with blaKPC which includes a transposase gene (tnpA), a resolvase gene (tnpR), as well as insertion sequences, ISKpn6 and ISKpn7 (Cuzon et al., 2010). (B) The blaNDM-1 construct demonstrates ISAba125 insertion sequence(s) upstream of the blaNDM-1 and a novel bleomycin resistance gene, bleMBL, downstream (Dortet et al., 2012). (C) blaOXA-48 is often mapped to a Tn1999 composite transposon where it is bracketed between two copies of the same insertion sequence, IS1999. Downstream of blaOXA-48 lies a lysR gene which encodes for a regulatory protein (Poirel et al., 2012b).
A predominant strain of K. pneumoniae appears responsible for outbreaks and the international spread of KPC-producing K. pneumoniae (Woodford et al., 2008; Kitchel et al., 2009a; Samuelsen et al., 2009). Congruent pulsed-field gel electrophoresis (PFGE) patterns also suggest a clonal relationship between outbreak-associated strains of KPC-producing K. pneumoniae recovered from different areas that are endemic (Navon-Venezia et al., 2009; Woodford et al., 2011). The Centers for Disease Control and Prevention (CDC) performed PFGE and multilocus sequence typing (MLST) on isolates submitted to their reference laboratory from 1996 to 2008. A dominant PFGE pattern was observed and noted to be of a specific MLST type, ST 258 (Kitchel et al., 2009a). A second sequence type, ST 14, was common in institutions in the Midwest (Kitchel et al., 2009b). These findings implied that certain strains of K. pneumoniae may be more apt to obtain and retain the blaKPC gene. Another study, however, analyzing 16 KPC-2 producing K. pneumoniae isolates from different geographic regions demonstrated diverse PFGE patterns and MLST types. This included four different MLST types in Colombia (ST 14, ST 337, ST 338, and ST 339) and two in Israel (ST 227 and ST 340). Although this study analyzed a smaller number of isolates, these findings suggest that the global propagation of KPC-2 is more complicated than the successful expansion of a fixed number of clones (Cuzon et al., 2010; Qi et al., 2011). More recently, a study evaluating the MLST types associated with widespread KPC-2 production in K. pneumoniae in Greece suggested that although ST 258 predominates at least 10 additional sequence types were found to carry blaKPC-2. Of note three (i.e., ST 147, ST 323, and ST 383) carried both KPC-2 as well as genes encoding VIM-type MBLs (Giakkoupi et al., 2011; Woodford et al., 2011). A retrospective study in Cleveland documented the presence of ST 36 in a long-term care facility for children (Viau et al., 2012).
Klebsiella pneumoniae carbapenemases-production can confer variable levels of carbapenem resistance with reported minimum inhibitory concentrations (MICs) ranging from susceptible to ≥ μg/mL. Analysis of isolates displaying high-level carbapenem resistance demonstrated that increased phenotypic resistance may be due to increased blaKPC gene copy number or the loss of an outer membrane porin, OmpK35 and/or OmpK36. The highest level of imipenem resistance was seen with isolates lacking both porins and with augmented KPC enzyme production (Kitchel et al., 2010).
Ambler Class B Carbapenemases: Metallo-β-Lactamases
Class B β-lactamases (Table 2) are referred to as MBLs and require a metal ion, usually zinc, for β-lactam hydrolysis (Walsh et al., 2005). Due to the dependence on Zn2+, catalysis is inhibited in the presence of metal-chelating agents like ethylenediaminetetraacetic acid (EDTA). MBL expression in Gram-negative bacteria confers resistance to penicillins, cephalosporins, and carbapenems. MBLs are not inhibited by the presence of commercially available β-lactamase inhibitors and susceptibility to monobactams (e.g., aztreonam) appears to be preserved in the absence of concomitant expression of other resistance mechanisms (e.g., ESBL production). The more geographically widespread MBLs include IMP (imipenem-resistant), VIM, and New Delhi metallo-β-lactamase (NDM).
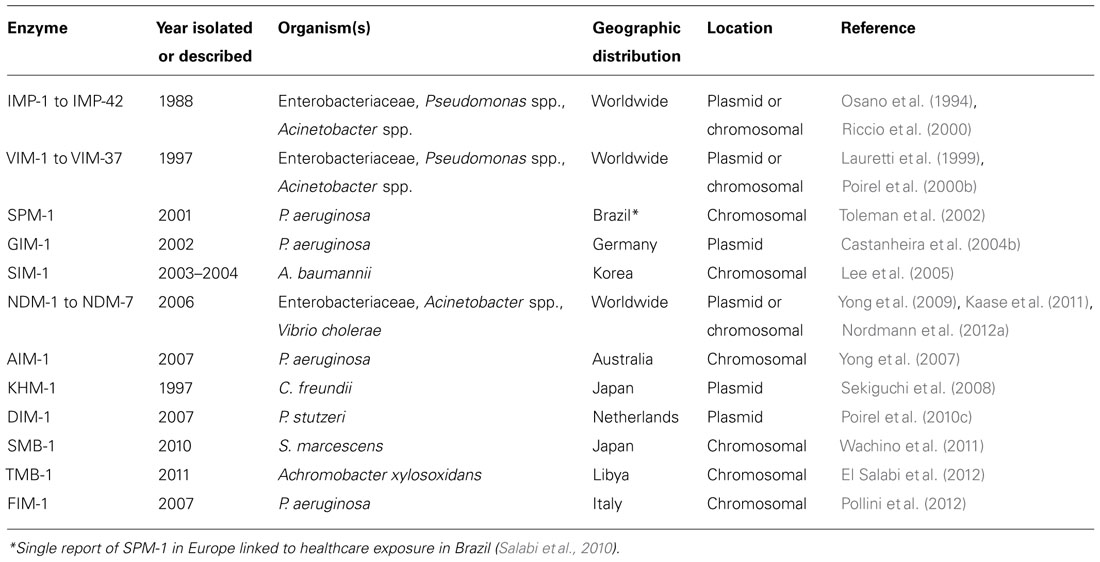
TABLE 2. Metallo-β-lactamases.
Chromosomal MBLs were the first to be identified and are the cause of carbapenem resistance observed in Bacillus cereus, Aeromonas spp., and Stenotrophomonas maltophilia (Walsh et al., 2005). However, of growing concern are the “mobile” MBLs that have been reported since the mid-1990s. Although most frequently found in carbapenem-resistant isolates of P. aeruginosa and occasionally Acinetobacter spp., there is growing isolation of these enzymes in Enterobacteriaceae.
Prior to the description of NDM-1, frequently detected MBLs include IMP-type and VIM-type with VIM-2 being the most prevalent. These MBLs are embedded within a variety of genetic structures, most commonly integrons. When these integrons are associated with transposons or plasmids they can readily be transferred between species.
In 1991, IMP-1, a plasmid-mediated MBL, was identified in an isolates of S. marcescens from Japan (Ito et al., 1995). Since then plasmid-mediated carbapenem resistance secondary to IMP-1 spread widely in Japan, Europe, Brazil, and other parts of Asia and in several species of Gram-negative bacilli including Acinetobacter spp. and Enterobacteriaceae. At the present time, 42 variants of IMP have been identified with most cases of IMP-mediated carbapenem resistance being reported from Asia and among P. aeruginosa (Bush and Jacoby, 2010).
A more commonly recovered MBL is the VIM-type enzyme. VIM-1 was first described in Italy in 1997 in P. aeruginosa (Lauretti et al., 1999). VIM-2 was next discovered in southern France in P. aeruginosa cultured from a neutropenic patient in 1996 (Poirel et al., 2000b). Although originally thought to be limited to non-fermenting Gram-negative bacilli, VIM-type MBLs are being increasingly identified in Enterobacteriaceae as well (Giakkoupi et al., 2003; Kassis-Chikhani et al., 2006; Morfin-Otero et al., 2009; Canton et al., 2012). To date, 37 variants of VIM have been described with VIM-2 being the most common MBL recovered worldwide.
Other more geographically restricted MBLs include SPM-1 (Sao Paulo MBL), which has been associated with hospital outbreaks in Brazil (Toleman et al., 2002; Rossi, 2011); GIM-1 (German imipenemase) isolated in carbapenem-resistant P. aeruginosa isolates in Germany (Castanheira et al., 2004b); SIM-1 (Seoul imipenemase) isolated from A. baumannii isolates in Korea (Lee et al., 2005); KHM-1 (Kyorin Health Science MBL) isolated from a C. freundii isolate in Japan (Sekiguchi et al., 2008); AIM-1 (Australian imipenemase) isolated from P. aeruginosa in Australia (Yong et al., 2007); DIM-1 (Dutch imipenemase) isolated from a clinical P. stutzeri isolate in the Netherlands (Poirel et al., 2010c); SMB-1 (S. marcescens MBL) in S. marcescens in Japan (Wachino et al., 2011); TMB-1 (Tripoli MBL) in Achromobacter xylosoxidans in Libya (El Salabi et al., 2012), and FIM-1 (Florence imipenemase) from a clinical isolate of P. aeruginosa in Italy (Pollini et al., 2012). With the notable exception of SPM-1, these MBLs have remained confined to their countries of origin (Salabi et al., 2010).
NDM-1 was first identified in 2008. Due to its rapid international dissemination and its ability to be expressed by numerous Gram-negative pathogens, NDM is poised to become the most commonly isolated and distributed carbapenemase worldwide. Initial reports frequently demonstrated an epidemiologic link to the Indian subcontinent where these MBLs are endemic (Kumarasamy et al., 2010). Indeed, retrospective analyses of stored isolates suggest that NDM-1 may have been circulating in the subcontinent as early as 2006 (Castanheira et al., 2011). Despite initial controversy, the Balkans may be another area of endemicity for NDM-1 (Struelens et al., 2010; Jovcic et al., 2011; Livermore et al., 2011c; Halaby et al., 2012). Sporadic recovery of NDM-1 in the Middle East suggests that this region may be an additional reservoir (Poirel et al., 2010a, 2011d; Nordmann et al., 2011; Ghazawi et al., 2012).
Like KPCs, the conveniences of international travel and medical tourism have quickly propelled this relatively novel MBL into a formidable public health threat. Gram-negative bacilli harboring blaNDM have been identified worldwide with the exception of Central and South America.
NDM-1 was first identified in Sweden in a patient of Indian descent previously hospitalized in India (Yong et al., 2009). The patient was colonized with a K. pneumoniae and an E. coli carrying blaNDM-1 on transferable plasmids. In the UK, an increase in the number of clinical isolates of carbapenem-resistant Enterobacteriaceae was seen in both 2008 and 2009. A UK reference laboratory reported that at least 17 of 29 patients found to be harboring NDM-1 expressing Enterobacteriaceae had a history of recent travel to the Indian subcontinent with the majority having been hospitalized in those countries (Kumarasamy et al., 2010).
European reports suggest that horizontal transfer of blaNDM-1 exists within hospitals outside endemic areas. Of overwhelming concern are the reported cases without specific contact with the healthcare system locally or in endemic areas suggesting autochthonous acquisition (Kumarasamy et al., 2010; Kus et al., 2011; Arpin et al., 2012; Borgia et al., 2012; Nordmann et al., 2012b).
Surveillance of public water supplies in India indicates that exposure to NDM-1 may be environmental. Walsh et al. (2011) analyzed samples of public tap water and seepage water from sites around New Delhi. The results were disheartening in that blaNDM-1 was detected by PCR in 4% of drinking water samples and 30% of seepage samples. In this survey, carriage of blaNDM-1 was noted in 11 species of bacteria not previously described, including virulent ones like Shigella boydii and Vibrio cholerae.
The rapid spread of NDM-1 highlights the fluidity and rapidity of gene transfer between bacterial species. Although blaNDM-1 was initially and repeatedly mapped to plasmids isolated from carbapenem-resistant E. coli and K. pneumoniae, reports of both plasmid and chromosomal expression of blaNDM-1 has been noted in other species of Enterobacteriaceae as well as Acinetobacter spp. and P. aeruginosa (Moubareck et al., 2009; Bogaerts et al., 2010; Bonnin et al., 2011; Nordmann et al., 2011; Patel and Bonomo, 2011). Recently, bacteremia with a NDM-1 expressing V. cholerae has been described in a patient previously hospitalized in India colonized with a variety of Enterobacteriaceae previously known to be capable of carrying plasmids with blaNDM-1 (Darley et al., 2012).
In contrast to KPCs, the presence of a dominant clone among blaNDM-1 carrying isolates remains elusive (Poirel et al., 2011c). NDM-1 expression in E. coli has been noted among sequence types previously associated with the successful dissemination of other β-lactamases including ST 101 and ST 131 (Mushtaq et al., 2011). Mushtaq et al. (2011) analyzed a relatively large group of blaNDM-1 expressing E. coli from the UK, Pakistan, and India in order to potentially identify a predominant strain responsible for the rapid and successful spread of NDM-1. The most frequent sequence type identified was ST 101. Another study examining a collection of carbapenem-resistant Enterobacteriaceae from India demonstrates the diversity of strains capable of harboring blaNDM-1. Carriage of blaNDM-1 was confirmed in 10 different sequence types of K. pneumoniae and 5 sequence types of E. coli (Lascols et al., 2011). This multiplicity was confirmed in a study looking at a collection of blaNDM-1 expressing Enterobacteriaceae from around the world (Poirel et al., 2011c). Of most concern is that NDM-1 has been identified in E. coli ST 131, the strain of E. coli credited with the global propagation of CTX-M-15 ESBLs (Mushtaq et al., 2011; Peirano et al., 2011; Pfeifer et al., 2011b; Woodford et al., 2011). Similar to KPCs, NDM-1 expression portends variable levels of carbapenem resistance and there is often concomitant carriage of a myriad of resistance determinants including other β-lactamases and carbapenemases as well as genes associated with resistance to fluoroquinolones and aminoglycosides (Nordmann et al., 2011).
NDM-1 shares the most homology with VIM-1 and VIM-2. It is a 28-kDa monomeric protein that demonstrates tight binding to both penicillins and cephalosporins (Zhang and Hao, 2011). Binding to carbapenems does not appear to be as strong as other MBLs, but hydrolysis rates appear to be similar. Using ampicillin as a substrate, allowed for detailed characterization of the interactions between NDM's active site and β-lactams as well as improved evaluation of MBLs unique mechanism of β-lactam hydrolysis. More recent crystal structures of NDM-1 reveal the molecular details of how carbapenem antibiotics are recognized by dizinc-containing MBLs (King et al., 2012).
To date, NDM-1 remains the most common NDM variant isolated. Seven variants (NDM-1 to NDM-7) exist (Kaase et al., 2011; Nordmann et al., 2012a). It is currently held that blaNDM-1 is a chimeric gene that may have evolved from A. baumannii (Toleman et al., 2012). Contributing to this theory is the presence of complete or variations of the insertion sequence, ISAba125, upstream to the blaNDM-1 gene in both Enterobacteriaceae and A. baumannii (Pfeifer et al., 2011a; Poirel et al., 2011a; Dortet et al., 2012; Toleman et al., 2012). This insertion sequence has primarily been found in A. baumannii.
A recent evaluation of the genetic construct associated with blaNDM-1 (Figure 1B) has lead to the discovery of a new bleomycin resistance protein, BRPMBL. Evaluation of 23 isolates of blaNDM-1/2 harboring Enterobacteriaceae and A. baumannii noted that the overwhelming majority of them possessed a novel bleomycin resistance gene, bleMBL (Dortet et al., 2012). Co-expression of blaNDM-1 and bleMBL appear to be mediated by a common promoter (PNDM-1) which includes portions of ISAba125. It is postulated that BRPMBL expression may contribute some sort of selective advantage allowing NDM-1 to persist in the environment.
A contemporary evaluation of recently recovered NDM-1 producing A. baumannii isolates from Europe demonstrates that blaNDM-1 and blaNDM-2 genes are situated on the same chromosomally located transposon, Tn125 (Bonnin et al., 2012). Dissemination of blaNDM in A. baumannii seems be due to different strains carrying Tn125 or derivatives of Tn125 rather than plasmid-mediated or clonal (Bonnin et al., 2013; Poirel et al., 2012a).
Carbapenem-Hydrolyzing Class D β-Lactamases
Oxacillinases comprise a heterogeneous group of class D β-lactamases which are able to hydrolyze amino- and carboxypenicillins (Poirel et al., 2010b). The majority of class D β-lactamases are not inhibited by commercially available β-lactamase inhibitors but are inhibited in vitro by NaCl. Over 250 types of oxacillinases are reported with a minority demonstrating low levels of carbapenem-hydrolyzing activity. This select group of enzymes is also referred to as the carbapenem-hydrolyzing class D β-lactamases (CHDLs; Table 3). CHDLs have been identified most frequently in Acinetobacter spp., however, there has been increasing isolation among Enterobacteriaceae, specifically OXA-48 (oxacillinase-48; Lascols et al., 2012; Mathers et al., 2012).
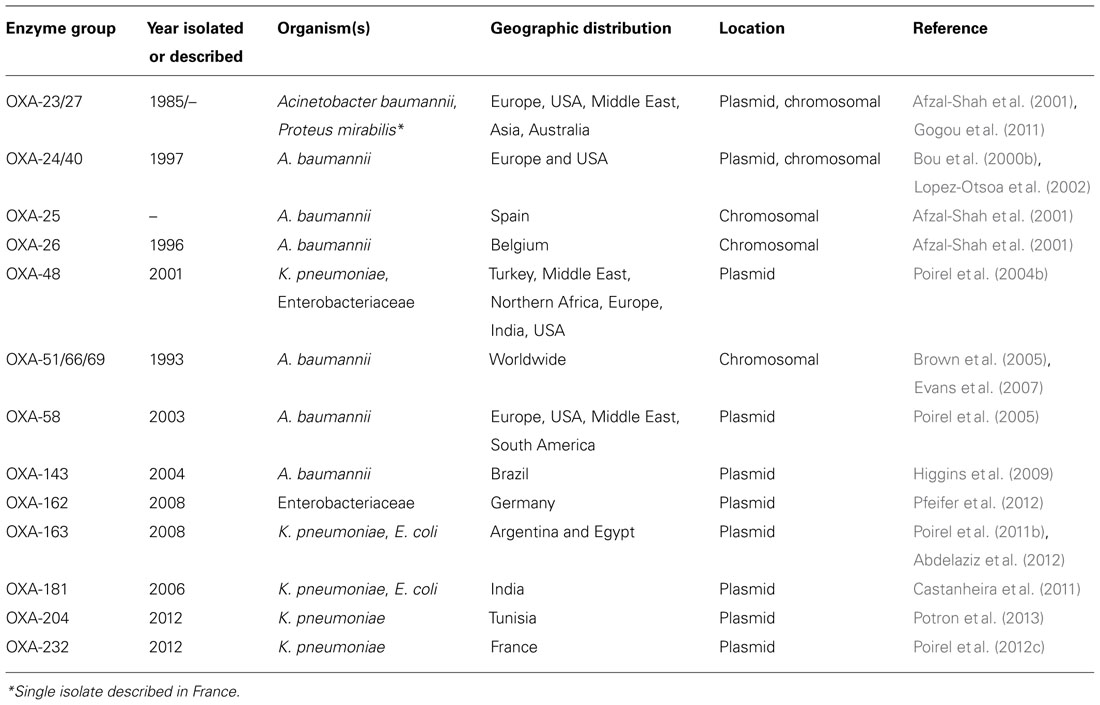
TABLE 3. Carbapenem-hydrolyzing class D β-lactamases.
With the exception of OXA-163 (Poirel et al., 2011b), CHDLs efficiently inactivate penicillins, first generations cephalosporins, and β-lactam/β-lactamase inhibitor combinations, but spare extended-spectrum cephalosporins. Carbapenem hydrolysis efficiency is lower than that of other carbapenemases, including the MBLs, and often additional resistance mechanisms are expressed in organisms demonstrating higher levels of phenotypic carbapenem resistance. These include expression of other carbapenemases, alterations in outer membrane proteins (e.g., CarO, OmpK36; Perez et al., 2007; Gülmez et al., 2008; Pfeifer et al., 2012), increased transcription mediated by IS elements functioning as promoters, increased gene copy number, and amplified drug efflux (Poirel and Nordmann, 2006; Perez et al., 2007). Many subgroups of CHDLs have been described. We will focus on those found in A. baumannii and Enterobacteriaceae: OXA-23 and OXA-27; OXA-24/40, OXA-25, and OXA-26; OXA-48 variants; OXA-51, OXA-66, OXA-69; OXA-58, and OXA-143.
CHDLs can be intrinsic or acquired. A. baumannii does have naturally occurring but variably expressed chromosomal CHDLs, OXA-51, OXA-66, and OXA-69 (Brown et al., 2005; Héritier et al., 2005b). For the most part, in isolation the phenotypic carbapenem resistance associated with these oxacillinases is low. However, levels of carbapenem resistance appear to be increased in the presence of specific insertion sequences promoting gene expression (Figueiredo et al., 2009; Culebras et al., 2010). Additional resistance to extended-spectrum cephalosporins can be seen in the setting of co-expression of ESBLs and/or other carbapenemases (Castanheira et al., 2011; Mathers et al., 2012; Pfeifer et al., 2012; Voulgari et al., 2012; Potron et al., 2013).
The first reported “acquired” oxacillinase with appreciable carbapenem-hydrolyzing activity was OXA-23. OXA-23, or ARI-1, was identified from an A. baumannii isolate in Scotland in 1993 (the isolate was first recovered in 1985; Paton et al., 1993). Subsequently, OXA-23 expression has been reported worldwide (Mugnier et al., 2010) and both plasmid and chromosomal carriage of blaOXA-23 are described. The OXA-23 group includes OXA-27, found in a single A. baumannii isolate from Singapore (Afzal-Shah et al., 2001). With the exception of an isolate of Proteus mirabilis identified in France in 2002, this group of β-lactamases has been exclusively recovered from Acinetobacter species (Bonnet et al., 2002). Increased expression of OXA-23 has been associated with the presence of upstream insertion sequences (e.g., ISAba1 and ISAba4) acting as strong promoters (Corvec et al., 2007).
Another group of CHDLs include OXA-24/40, OXA-25, and OXA-26 (Bou et al., 2000b; Afzal-Shah et al., 2001). OXA-24 and OXA-40 differ by a few amino acid substitutions and OXA-25 and OXA-26 are point mutation derivatives of OXA-40 (Afzal-Shah et al., 2001). Although primarily linked with clonal outbreaks in Spain and Portugal (Bou et al., 2000a; Lopez-Otsoa et al., 2002; Da Silva et al., 2004; Acosta et al., 2011), OXA-24/40 β-lactamases has been isolated in other European countries and the USA (Lolans et al., 2006). OXA-40 was in fact the first CHDL documented in the USA (Lolans et al., 2006).
OXA-58 has also only been detected in Acinetobacter spp. initially identified in France (Héritier et al., 2005a; Poirel et al., 2005), OXA-58 has been associated with institutional outbreaks and has been recovered from clinical isolates of A. baumannii worldwide (Coelho et al., 2006; Mendes et al., 2009; Gales et al., 2012).
As civilian and military personnel began returning from Afghanistan and the Middle East, practitioners noted increasing recovery of A. baumannii from skin and soft tissue infections. Drug resistance was associated with expression of both OXA-23 and OXA-58 (Hujer et al., 2006; Scott et al., 2007; Perez et al., 2010b). Many isolates carrying the blaOXA-58 gene concurrently carry insertion sequences (e.g., ISaba1, ISAba2, or ISAba3) associated with increased carbapenemase production and thus higher levels of carbapenem resistance. In one report increased gene copy number was also associated with a higher level of enzyme production and increased phenotypic carbapenem resistance (Bertini et al., 2007).
Spread of OXA-type carbapenemases among A. baumannii appears to be clonal and in depth reviews of the molecular epidemiology and successful dissemination of these clones have been published (Woodford et al., 2011; Zarrilli et al., 2013). Two MLST schemes with three loci in common exist for A. baumannii – the PubMLST scheme (Bartual et al., 2005) and the Pasteur scheme (Diancourt et al., 2010). Both schemes assign different sequence types into clonal complexes (CC). Sequence types and CC from both schemes can be further categorized into the international (European) clones I, II, and III. It should be noted, however, that the molecular taxonomy of A. baumannii continues to evolve (Higgins et al., 2012a). OXA-23 producing A. baumannii predominantly belong to international clones I and II with a notable proportion being part of CC92 (PubMLST; Mugnier et al., 2010; Adams-Haduch et al., 2011). Similarly, A. baumannii isolates associated with epidemic spread of OXA-24/40 in Portugal and Spain appear are incorporated in international clone II (Da Silva et al., 2004; Grosso et al., 2011) and ST 56 (PubMLST; Acosta et al., 2011). OXA-58 expressing A. baumannii have been associated with international clones I, II, and II and a variety of unrelated sequence types (Di Popolo et al., 2011; Gogou et al., 2011).
OXA-48 was originally identified in a carbapenem-resistant isolate of K. pneumoniae in Turkey (Poirel et al., 2004c). Early reports suggested that this enzyme was geographically restricted to Turkey. In the past few years, however, the enzyme has been recovered from variety of Enterobacteriaceae and has successfully circulated outside of Turkey with reports of isolation in the Middle East, North Africa, Europe (Carrer et al., 2010), and most recently the USA (Lascols et al., 2012; Mathers et al., 2012). The Middle East and North Africa may be secondary reservoirs for these CHDLs (Hays et al., 2012; Poirel et al., 2012c). Indeed, the introduction of OXA-48 expressing Enterobacteriaceae in some countries has been from patients from the Middle East or Northern Africa (Decre et al., 2010; Adler et al., 2011; Poirel et al., 2011e; Canton et al., 2012). In the USA, the first clinical cases were associated with ST 199 and ST 43 (Mathers et al., 2012).
At least six OXA-48 variants (e.g., OXA-48, OXA-162, OXA-163, OXA-181, OXA-204, and OXA-232) have been identified. OXA-48 is by far the most globally dispersed and its epidemiology has been recently reviewed (Poirel et al., 2012c). Unlike KPCs and NDM-1 which have been associated with a variety of plasmids, a single 62 kb self-conjugative IncL/M-type plasmid has contributed to a large proportion of the distribution of blaOXA-48 in Europe (Potron et al., 2011a). Sequencing of this plasmid (pOXA-48a) notes that blaOXA-48 had been integrated through the acquisition of a Tn1999 composite transposon (Figure 1C; Poirel et al., 2012b) blaOXA-48 appears to be associated with a specific insertion sequence, IS1999 (Poirel et al., 2004c, 2012b). A variant of Tn1999, Tn1999.2, has been identified among isolates from Turkey and in Europe (Carrer et al., 2010; Potron et al., 2011a). Tn1999.2 harbors an IS1R element within the IS1999. OXA-48 appears to have the highest affinity for imipenem of the CHDLs specifically those harboring blaOXA-48 within a Tn1999.2 composite transposon (Docquier et al., 2009). Three isoforms of the Tn1999 transposon have been described (Giani et al., 2012).
Although much of the spread of OXA-48 is attributed to a specific plasmid, outbreak evaluations demonstrate that a variety of strains have contributed to dissemination of this emerging carbapenemase in K. pneumoniae. The same K. pneumoniae sequence type, ST 395, harboring blaOXA-48 was identified in Morocco, France, and the Netherlands (Cuzon et al., 2011; Potron et al., 2011a). ST 353 was associated with an outbreak of OXA-48 producing K. pneumoniae in London (Woodford et al., 2011) and ST 221 with an outbreak of OXA-48 in Ireland (Canton et al., 2012). OXA-48 production in K. pneumoniae, like KPC-expressing K. pneumoniae, has also been associated with ST 14 (Poirel et al., 2004c) and a recent outbreak in Greece was associated with ST 11 (Voulgari et al., 2012).
blaOXA-48 is remarkably similar to blaOXA-54, a β-lactamase gene intrinsic to Shewanella oneidensis (Poirel et al., 2004a). Shewanella spp. are relatively ubiquitous waterborne Gram-negative bacilli and are proving to be a potential environmental reservoir for OXA-48 like carbapenemases as well as other resistance determinants (Héritier et al., 2004; Poirel et al., 2004b; Potron et al., 2011b).
OXA-163, a single amino acid variant of OXA-48, was identified in isolates of K. pneumoniae and Enterobacter cloacae from Argentina and is unique in that it has activity against extended-spectrum cephalosporins (Poirel et al., 2011b). OXA-163 also has been identified in Egypt, which has a relatively prevalence of OXA-48, in patients without epidemiologic links to Argentina (Abdelaziz et al., 2012).
OXA-181 was initially identified among carbapenem-resistant Enterobacteriaceae collected from India (Castanheira et al., 2011). OXA-181 differs from OXA-48 by four amino acids, however, appears to be nestled in an entirely different genetic platform. The blaOXA-181 gene has been mapped to a different group of plasmids, the ColE family, and has been associated with an alternative insertion sequence, ISEcp1. The latter insertion sequence has been associated with the acquisition of other β-lactamases including CTX-M-like ESBLs. Like, OXA-48, it appears that OXA-181 may have evolved from a waterborne environmental species Shewanella xiamenensis (Potron et al., 2011b).
OXA-204 differs from OXA-48 by a two amino acid substitution. It was recently identified in a clinical K. pneumoniae isolate from Tunisia (Potron et al., 2013). Its genetic construct appears to be similar to that of OXA-181. OXA-232 was recently identified among K. pneumoniae isolates in France (Poirel et al., 2012c).
OXA-143 is a novel plasmid-borne carbapenem-hydrolyzing oxacillinase recovered from clinical A. baumannii isolates in Brazil (Higgins et al., 2009). Information regarding its significance and prevalence continues to evolve (Antonio et al., 2010; Werneck et al., 2011; Mostachio et al., 2012).
Available Agents and Drugs in Development
Few antimicrobials are currently available to treat infections with carbapenemase-producing Gram-negative bacteria. Carriage of concurrent resistance determinants can result in decreased susceptibility non-β-lactams including the fluoroquinolones and aminoglycosides thus further compromising an already limited antimicrobial arsenal. What frequently remains available are the polymyxins (including colistin), tigecycline, and fosfomycin but susceptibilities to these agents are unpredictable (Falagas et al., 2011).
The reintroduction of polymyxins, both polymyxin B and colistin overlaps with the evolution of carbapenem resistance among Gram-negative bacilli. The clinical “resurgence” of these agents is well documented (Falagas and Kasiakou, 2005; Li et al., 2006a; Landman et al., 2008). Some experts advocate for the use of polymyxins in combination with other agents like rifampicin (Hirsch and Tam, 2010; Urban et al., 2010). In vitro evaluations of different combinations including carbapenems, rifamycins, and/or tigecycline demonstrate variable results (Bercot et al., 2011; Biswas et al., 2012; Deris et al., 2012; Jernigan et al., 2012). Most evaluations of the clinical outcomes or “effectiveness” of combination therapies have been retrospective (Qureshi et al., 2012; Tumbarello et al., 2012). Prospective clinical trials evaluating the superiority of colistin-based combination therapy over monotherapy are in their infancy. A real interest in combination therapy persists due to the concern of hetero-resistance (Li et al., 2006b; Poudyal et al., 2008; Lee et al., 2009; Yau et al., 2009; Meletis et al., 2011).
Early evaluations of the glycylcycline, tigecycline, demonstrated favorable in vitro activity against ESBL-producing Enterobacteriaceae and specific isolates of carbapenem-resistant A. baumannii and Enterobacteriaceae (Bratu et al., 2005; Fritsche et al., 2005; Noskin, 2005; Castanheira et al., 2008; Wang and Dowzicky, 2010). Tigecycline remains untested in prospective trials and reports of resistance are increasing (Navon-Venezia et al., 2007; Anthony et al., 2008; Wang and Dowzicky, 2010; Sun et al., 2012). The role of tigecycline in treating primary bloodstream infections or urinary tract infections remains undefined due less than therapeutic concentrations of drug achieved in the serum (Rodvold et al., 2006) and urine (Satlin et al., 2011). We also note that meta-analyses of pooled data from trials evaluating the use of tigecycline for a variety of indications suggest there is a excess mortality associated with the use of tigecycline over comparator regimens (Cai et al., 2011; Tasina et al., 2011; Yahav et al., 2011; Verde and Curcio, 2012). However, in the absence of other tested regimens tigecycline may be an appropriate or perhaps the only therapeutic option.
Growing resistance to both the polymyxins and tigecycline has resulted the revisiting of older drugs including chloramphenicol, nitrofurantoin, and temocillin (Livermore et al., 2011d). Fosfomycin is also one of these earlier antibiotics being reassessed (Falagas et al., 2008). In an in vitro evaluation of 68 KPC-expressing K. pneumoniae isolates, fosfomycin demonstrated in vitro activity against 87% of tigecycline and/or polymyxin non-susceptible isolates and 83% of isolates that were resistant to both (Endimiani et al., 2010b). Fosfomycin may be a potential therapeutic option for patients infected with carbapenemase-producing Enterobacteriaceae if the infection is localized to the genitourinary tract. Unfortunately, fosfomycin does not demonstrate reliable activity against non-urinary pathogens. Fosfomycin demonstrated activity against only 30.2% of 1693 multidrug-resistant (MDR) P. aeruginosa isolates and 3.5% of 85 MDR A. baumannii isolates (Falagas et al., 2009). The individual studies included in this review did not employ uniform MDR definitions or consistent susceptibility breakpoints. Moreover, access to the parenteral fosfomycin is limited and the threshold for resistance is low (Rodriguez-Rojas et al., 2010; Karageorgopoulos et al., 2012). Concerns regarding the emergence of resistance have lead to an increasing interest in the utility of combination therapy (Michalopoulos et al., 2010; Bercot et al., 2011; Souli et al., 2011).
Few agents are in the advanced stages of development with demonstrable in vitro activity against carbapenemase-producing organisms. These include β-lactamase inhibitors, aminoglycoside derivatives, polymyxin derivatives, and novel monobactams and monobactams-β-lactamase inhibitor combinations.
Avibactam, or NXL104, is a β-lactamase inhibitor which has been tested in combination with ceftazidime, ceftaroline, and aztreonam against several carbapenemase-producing Enterobacteriaceae with impressive decreases in MICs (Livermore et al., 2008, 2011b; Endimiani et al., 2009a; Mushtaq et al., 2010c). Cephalosporin–avibactam combinations do not inhibit MBLs. Avibactam in combination with aztreonam, however, does seem to demonstrate activity against isolates harboring a variety of carbapenem resistance mechanisms including MBLs (Livermore et al., 2011b). Regrettably, the avibactam and aztreonam combination is not currently in clinical trials. The combination of ceftazidime–avibactam has been evaluated against collections of non-fermenting Gram-negative pathogens and its role remains undefined (Mushtaq et al., 2010b). In some evaluations of ceftazidime non-susceptible isolates of P. aeruginosa decrease MICs were noted with the addition of avibactam (Mushtaq et al., 2010b; Walkty et al., 2011; Crandon et al., 2012; Levasseur et al., 2012). The combinations of ceftaroline–avibactam and ceftazidime–avibactam are currently in clinical trials.
Methylidene penems (penem-1 and penem-2) are β-lactamase inhibitors and appear to be potent inhibitors of KPC-2 (Papp-Wallace et al., 2010). The combination of cefepime with penem-1 demonstrated lower cefepime MICs in 88.1% of the 42 KPC-producing K. pneumoniae isolates evaluated (Endimiani et al., 2010a). MK-7655 is a novel β-lactamase being evaluated in combination with imipenem against carbapenem-resistant Gram-negative bacilli (Hirsch et al., 2012).
ME1071, formerly CP3242 (Bassetti et al., 2011), is a maleic acid derivative that competitively inhibits MBLs. Earlier studies demonstrated concentration-dependent decreases in carbapenem MICs in MBL-producing P. aeruginosa (Ishii et al., 2010), A. baumannii, and select Enterobacteriaceae (Shahid et al., 2009) A contemporary pre-clinical evaluation of ME1071 in combination with various type 2 carbapenems (i.e., biapenem, doripenem, meropenem, imipenem) confirms remarkable decreases in the carbapenem MICs for Enterobacteriaceae and A. baumannii harboring IMP, VIM, and NDM-type MBLs (Livermore et al., 2013). Irrespective of the candidate carbapenem, ME1071 activity against NDM MBLs was less than that of VIM-type and IMP-type MBLs. Of note, biapenem was the carbapenem with the lowest baseline MICs to the MBLs, but it is commercially unavailable in many countries including the USA. Other MBL-specific inhibitors are in pre-clinical development (Chen et al., 2012).
Plazomicin (ACHN-490) is an aminoglycoside derivative with potent activity against some carbapenem-resistant Gram-negative bacilli (Zhanel et al., 2012). Studies have noted that susceptibilities to aminoglycosides vary among KPC-producing K. pneumoniae. In one evaluation, 48% of 25 tested isolates were susceptible to amikacin, 44% to gentamicin, and 8% to tobramycin. Plazomicin demonstrated an MIC90 significantly lower than that of amikacin (Endimiani et al., 2009c). In vitro studies also indicate that depending on the aminoglycoside resistance mechanisms present, Plazomicin may have activity against select isolates of P. aeruginosa and A. baumannii (Aggen et al., 2010; Landman et al., 2011). Susceptibility to plazomicin in the setting of resistance to other aminoglycosides appears to be dependent on the mechanism of aminoglycoside resistance (Livermore et al., 2011a).
NAB739 and NAB7061 are polymyxin derivatives that may be less nephrotoxic than commercially available polymyxins. In a small in vitro study, NAB739 displayed activity against nine carbapenemase-producing polymyxin-susceptible isolates of Enterobacteriaceae (Vaara et al., 2010). A contemporary evaluation of NAB739 demonstrated higher MICs compared to those of polymyxin B in a collection of polymyxin-susceptible and non-susceptible Enterobacteriaceae, P. aeruginosa, and A. baumannii (Vaara et al., 2012). NAB7061 when used in combination with rifampicin or clarithromycin demonstrated synergistic activity against seven strains of carbapenemase-producing Gram-negative bacilli including one polymyxin-resistant strain (Vaara et al., 2010). It remains unclear what role these agents will play in the setting the increasing burden of infections with carbapenemase-producing Enterobacteriaceae.
The activity of the siderophore monosulfactam, BAL30072, has been against non-fermenting carbapenemase-producing Gram-negative bacilli (Page et al., 2010). In one study, susceptibility to BAL30072 was noted in 73% of 200 isolates of carbapenemase-producing A. baumannii, the majority of which were of the same OXA-23 producing clone (Mushtaq et al., 2010a). In that same study, smaller percentages of susceptibility were noted in a selection of carbapenem-resistant Burkholderia cepacia and P. aeruginosa isolates. Recent evaluations of BAL30072 confirm that there may be a role for this agent in the treatment of resistant A. baumannii infections (Russo et al., 2011; Higgins et al., 2012b). BAL 30376 is a combination of a siderophore monobactam with clavulanic acid. In two studies, this combination demonstrated reasonable in vitro activity against CHDL, including OXA-48, and MBLs but not KPCs (Livermore et al., 2010; Page et al., 2011).
Concluding Remarks
In the last 5 years, we have witnessed the global spread of carbapenem resistance among Gram-negative organisms. The notion that multidrug resistance among these pathogens is limited to isolated outbreaks among the critically ill has met the ultimate challenge with NDM-1 (Kumarasamy et al., 2010). The conveniences of travel and medical tourism have introduced resistance mechanisms across states, countries, and even continents at an alarming rate (Rogers et al., 2011; van der Bij and Pitout, 2012). Rates of resistance in some countries may be underestimated due to the lack of organized reporting structures and limited resources. Long-term healthcare facilities are now recognized reservoirs for the continued propagation of MDR organisms (Urban et al., 2008; Aschbacher et al., 2010; Perez et al., 2010a; Ben-David et al., 2011; Prabaker et al., 2012; Viau et al., 2012).
Until the introduction of accurate, affordable, and readily accessible diagnostics and reliably effective antimicrobials a major focus remains containment and eradication of these organisms within the healthcare environment. Many cite a “bundle” type approach that includes administrative support, active surveillance, antimicrobial stewardship, and augmented infection control practices (Centers for Disease Control and Prevention, 2009; Schwaber et al., 2011; Snitkin et al., 2012). Just as with drug development (Tillotson, 2010), the future savings of investing in prevention is not as tangible as the immediate capital investment required to allot appropriate resources including advanced laboratory platforms, experienced laboratory personnel, dedicated nursing staff, and infection control personnel (Bilavsky et al., 2010). Expanding these efforts to non-acute healthcare settings is recommended to begin to stem the evolving pandemic of carbapenem resistance (Gupta et al., 2011).
The prudent use of antibiotics is essential in combating the continuing evolution of resistance (Marchaim et al., 2012). This may be even more crucial in areas where non-prescription antimicrobial use is common and continues to be unregulated. In an age where multidrug resistance is so widespread, even the appropriate use of broad-spectrum antibiotics has contributed to our current state.
Research funding and support for the description of resistance mechanisms, validation of current infection control practices, and antimicrobial development must be prioritized. Institutions supporting infection control, state of the art microbiology laboratories, and antimicrobial stewardship programs should receive recognition and incentives for their foresight. Despite these continuing challenges, considerable progress has been made to identify at-risk populations and to describe resistance determinants. Collaborative efforts (Kitchel et al., 2009a; Struelens et al., 2010; Canton et al., 2012) have led to a better understanding and awareness of the epidemiology and the contribution of antimicrobial use and the environment to the propagation of antimicrobial resistance. These joint efforts have proven crucial for the propagation of information about carbapenemases. Continuing to encourage these partnerships is imperative in the ongoing struggle against antimicrobial resistance and to prevent antimicrobials from essentially becoming obsolete.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported in part by the Veterans Affairs Merit Review Program (to Robert A. Bonomo), the National Institutes of Health (grants R01-A1063517 and RO3-A1081036 to Robert A. Bonomo), and the Geriatric Research Education and Clinical Center VISN 10 (to Robert A. Bonomo).
References
Abdelaziz, M. O., Bonura, C., Aleo, A., El-Domany, R. A., Fasciana, T., and Mammina, C. (2012). OXA-163-producing Klebsiella pneumoniae in Cairo, Egypt, in 2009 and 2010. J. Clin. Microbiol. 50, 2489–2491.
Acosta, J., Merino, M., Viedma, E., Poza, M., Sanz, F., Otero, J. R., et al. (2011). Multidrug-resistant Acinetobacter baumannii harboring OXA-24 carbapenemase, Spain. Emerg. Infect. Dis. 17, 1064–1067.
Adams-Haduch, J. M., Onuoha, E. O., Bogdanovich, T., Tian, G. B., Marschall, J., Urban, C. M., et al. (2011). Molecular epidemiology of carbapenem-nonsusceptible Acinetobacter baumannii in the United States. J. Clin. Microbiol. 49, 3849–3854.
Adler, A., Shklyar, M., Schwaber, M. J., Navon-Venezia, S., Dhaher, Y., Edgar, R., et al. (2011). Introduction of OXA-48-producing Enterobacteriaceae to Israeli hospitals by medical tourism. J. Antimicrob. Chemother. 66, 2763–2766.
Afzal-Shah, M., Woodford, N., and Livermore, D. M. (2001). Characterization of OXA-25, OXA-26, and OXA-27, molecular class D beta-lactamases associated with carbapenem resistance in clinical isolates of Acinetobacter baumannii. Antimicrob. Agents Chemother. 45, 583–588.
Aggen, J. B., Armstrong, E. S., Goldblum, A. A., Dozzo, P., Linsell, M. S., Gliedt, M. J., et al. (2010). Synthesis and spectrum of the neoglycoside ACHN-490. Antimicrob. Agents Chemother. 54, 4636–4642.
Ambler, R. P. (1980). The structure of beta-lactamases. Philos. Trans. R. Soc. Lond. B Biol. Sci. 289, 321–331.
Anthony, K. B., Fishman, N. O., Linkin, D. R., Gasink, L. B., Edelstein, P. H., and Lautenbach, E. (2008). Clinical and microbiological outcomes of serious infections with multidrug-resistant Gram-negative organisms treated with tigecycline. Clin. Infect. Dis. 46, 567–570.
Antonio, C. S., Neves, P. R., Medeiros, M., Mamizuka, E. M., Elmor De Araujo, M. R., and Lincopan, N. (2010). High prevalence of carbapenem-resistant Acinetobacter baumannii carrying the blaOXA-143 gene in Brazilian hospitals. Antimicrob. Agents Chemother. 55, 1322–1323.
Arpin, C., Noury, P., Boraud, D., Coulange, L., Manetti, A., Andre, C., et al. (2012). NDM-1-producing Klebsiella pneumoniae resistant to colistin in a French community patient without history of foreign travel. Antimicrob. Agents Chemother. 56, 3432–3434.
Aschbacher, R., Pagani, L., Doumith, M., Pike, R., Woodford, N., Spoladore, G., et al. (2010). Metallo-β-lactamases among Enterobacteriaceae from routine samples in an Italian tertiary-care hospital and long-term care facilities during 2008. Clin. Microbiol. Infect. 17, 181–189.
Aubron, C., Poirel, L., Ash, R. J., and Nordmann, P. (2005). Carbapenemase-producing Enterobacteriaceae, U.S. rivers. Emerg. Infect. Dis. 11, 260–264.
Bartual, S. G., Seifert, H., Hippler, C., Luzon, M. A., Wisplinghoff, H., and Rodriguez-Valera, F. (2005). Development of a multilocus sequence typing scheme for characterization of clinical isolates of Acinetobacter baumannii. J. Clin. Microbiol. 43, 4382–4390.
Bassetti, M., Ginocchio, F., and Mikulska, M. (2011). New treatment options against Gram-negative organisms. Crit. Care 15, 215.
Ben-David, D. M. D., Masarwa, S. M. A., Navon-Venezia, S. P., Mishali, H. M. A., Fridental, I. M. D., Rubinovitch, B. M. D., et al. (2011). Carbapenem-resistant Klebsiella pneumoniae in post-acute-care facilities in Israel. Infect. Control Hosp. Epidemiol. 32, 845–853.
Bercot, B., Poirel, L., Dortet, L., and Nordmann, P. (2011). In vitro evaluation of antibiotic synergy for NDM-1-producing Enterobacteriaceae. J. Antimicrob. Chemother. 66, 2295–2297.
Bertini, A., Poirel, L., Bernabeu, S., Fortini, D., Villa, L., Nordmann, P., et al. (2007). Multicopy blaOXA-58 gene as a source of high-level resistance to carbapenems in Acinetobacter baumannii. Antimicrob. Agents Chemother. 51, 2324–2328.
Bilavsky, E., Schwaber, M. J., and Carmeli, Y. (2010). How to stem the tide of carbapenemase-producing Enterobacteriaceae?: proactive versus reactive strategies. Curr. Opin. Infect. Dis. 23, 327–331.
Biswas, S., Brunel, J.-M., Dubus, J.-C., Reynaud-Gaubert, M., and Rolain, J.-M. (2012). Colistin: an update on the antibiotic of the 21st century. Expert Rev. Anti Infect. Ther. 10, 917–934.
Bogaerts, P., Naas, T., El Garch, F., Cuzon, G., Deplano, A., Delaire, T., et al. (2010). GES extended-spectrum beta-lactamases in Acinetobacter baumannii isolates in Belgium. Antimicrob. Agents Chemother. 54, 4872–4878.
Bonnet, R., Marchandin, H., Chanal, C., Sirot, D., Labia, R., De Champs, C., et al. (2002). Chromosome-encoded class D beta-lactamase OXA-23 in Proteus mirabilis. Antimicrob. Agents Chemother. 46, 2004–2006.
Bonnin, R. A., Nordmann, P., Potron, A., Lecuyer, H., Zahar, J. R., and Poirel, L. (2011). Carbapenem-hydrolyzing GES-type extended-spectrum beta-lactamase in Acinetobacter baumannii. Antimicrob. Agents Chemother. 55, 349–354.
Bonnin, R. A., Poirel, L., Naas, T., Pirs, M., Seme, K., Schrenzel, J., et al. (2012). Dissemination of New Delhi metallo-beta-lactamase-1-producing Acinetobacter baumannii in Europe. Clin. Microbiol. Infect. 18, E362–E365.
Bonnin, R. A., Rotimi, V. O., Al Hubail, M., Gasiorowski, E., Al Sweih, N., Nordmann, P., et al. (2013). Wide dissemination of GES-type carbapenemases in Acinetobacter baumannii isolates in Kuwait. Antimicrob. Agents Chemother. 57, 183–188.
Borgia, S., Lastovetska, O., Richardson, D., Eshaghi, A., Xiong, J., Chung, C., et al. (2012). Outbreak of carbapenem-resistant Enterobacteriaceae containing blaNDM-1, Ontario, Canada. Clin. Infect. Dis. 55, e109–e117.
Bou, G., Cervero, G., Dominguez, M. A., Quereda, C., and Martinez-Beltran, J. (2000a). Characterization of a nosocomial outbreak caused by a multiresistant Acinetobacter baumannii strain with a carbapenem-hydrolyzing enzyme: high-level carbapenem resistance in A. baumannii is not due solely to the presence of beta-lactamases. J. Clin. Microbiol. 38, 3299–3305.
Bou, G., Oliver, A., and Martinez-Beltran, J. (2000b). OXA-24, a novel class D beta-lactamase with carbapenemase activity in an Acinetobacter baumannii clinical strain. Antimicrob. Agents Chemother. 44, 1556–1561.
Bratu, S., Landman, D., Alam, M., Tolentino, E., and Quale, J. (2005). Detection of KPC carbapenem-hydrolyzing enzymes in Enterobacter spp. from Brooklyn, New York. Antimicrob. Agents Chemother. 49, 776–778.
Brown, S., Young, H. K., and Amyes, S. G. B. (2005). Characterisation of OXA-51, a novel class D carbapenemase found in genetically unrelated clinical strains of Acinetobacter baumannii from Argentina. Clin. Microbiol. Infect. 11, 15–23.
Bush, K., and Jacoby, G. A. (2010). Updated functional classification of beta-lactamases. Antimicrob. Agents Chemother. 54, 969–976.
Cai, J. C., Zhou, H. W., Zhang, R., and Chen, G.-X. (2008). Emergence of Serratia marcescens, Klebsiella pneumoniae, and Escherichia coli isolates possessing the plasmid-mediated carbapenem-hydrolyzing beta-lactamase KPC-2 in intensive care units of a Chinese hospital. Antimicrob. Agents Chemother. 52, 2014–2018.
Cai, Y., Wang, R., Liang, B., Bai, N., and Liu, Y. (2011). Systematic review and meta-analysis of the effectiveness and safety of tigecycline for treatment of infectious disease. Antimicrob. Agents Chemother. 55, 1162–1172.
Canton, R., Akova, M., Carmeli, Y., Giske, C. G., Glupczynski, Y., Gniadkowski, M., et al. (2012). Rapid evolution and spread of carbapenemases among Enterobacteriaceae in Europe. Clin. Microbiol. Infect. 18, 413–431.
Carrer, A., Poirel, L., Pitout, J. D., Church, D., and Nordmann, P. (2008). Occurrence of an SME-2-producing Serratia marcescens isolate in Canada. Int. J. Antimicrob. Agents 31, 181–182.
Carrer, A., Poirel, L., Yilmaz, M., Akan, O. A., Feriha, C., Cuzon, G., et al. (2010). Spread of OXA-48-encoding plasmid in Turkey and Beyond. Antimicrob. Agents Chemother. 54, 1369–1373.
Castanheira, M., Deshpande, L. M., Dipersio, J. R., Kang, J., Weinstein, M. P., and Jones, R. N. (2009). First descriptions of blaKPC in Raoultella spp. (R. planticola and R. ornithinolytica): report from the SENTRY antimicrobial surveillance program. J. Clin. Microbiol. 47, 4129–4130.
Castanheira, M., Deshpande, L. M., Mathai, D., Bell, J. M., Jones, R. N., and Mendes, R. E. (2011). Early dissemination of NDM-1- and OXA-181-producing Enterobacteriaceae in Indian hospitals: report from the SENTRY antimicrobial surveillance program, 2006–2007. Antimicrob. Agents Chemother. 55, 1274–1278.
Castanheira, M., Mendes, R. E., Walsh, T. R., Gales, A. C., and Jones, R. N. (2004a). Emergence of the extended-spectrum beta-lactamase GES-1 in a Pseudomonas aeruginosa strain from Brazil: report from the SENTRY antimicrobial surveillance program. Antimicrob. Agents Chemother. 48, 2344–2345.
Castanheira, M., Toleman, M. A., Jones, R. N., Schmidt, F. J., and Walsh, T. R. (2004b). Molecular characterization of a beta-lactamase gene, blaGIM-1, encoding a new subclass of metallo-beta-lactamase. Antimicrob. Agents Chemother. 48, 4654–4661.
Castanheira, M., Sader, H. S., Deshpande, L. M., Fritsche, T. R., and Jones, R. N. (2008). Antimicrobial activities of tigecycline and other broad-spectrum antimicrobials tested against serine carbapenemase- and metallo-beta-lactamase-producing Enterobacteriaceae: report from the SENTRY antimicrobial surveillance program. Antimicrob. Agents Chemother. 52, 570–573.
Centers for Disease Control and Prevention. (2009). Guidance for control of infections with carbapenem-resistant or carbapenemase-producing Enterobacteriaceae in acute care facilities. MMWR Morb. Mortal. Wkly. Rep. 58, 256–260.
Chen, P., Horton, L. B., Mikulski, R. L., Deng, L., Sundriyal, S., Palzkill, T., et al. (2012). 2-Substituted 4,5-dihydrothiazole-4-carboxylic acids are novel inhibitors of metallo-β-lactamases. Bioorg. Med. Chem. Lett. 22, 6229–6232.
Coelho, J., Woodford, N., Afzal-Shah, M., and Livermore, D. (2006). Occurrence of OXA-58-like carbapenemases in Acinetobacter spp. collected over 10 years in three continents. Antimicrob. Agents Chemother. 50, 756–758.
Corvec, S., Poirel, L., Naas, T., Drugeon, H., and Nordmann, P. (2007). Genetics and expression of the carbapenem-hydrolyzing oxacillinase gene blaOXA-23 in Acinetobacter baumannii. Antimicrob. Agents Chemother. 51, 1530–1533.
Crandon, J. L., Schuck, V. J., Banevicius, M. A., Beaudoin, M. E., Nichols, W. W., Tanudra, M. A., et al. (2012). Comparative in vitro and in vivo efficacies of human simulated doses of ceftazidime and ceftazidime–avibactam against Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 56, 6137–6146.
Culebras, E., Gonzalez-Romo, F., Head, J., Gomez, M., Morales, G., and Picazo, J. J. (2010). Outbreak of Acinetobacter baumannii producing OXA-66 in a Spanish hospital: epidemiology and study of patient movements. Microb. Drug Resist. 16, 309–315.
Cuzon, G., Naas, T., Truong, H., Villegas, M. V., Wisell, K. T., Carmeli, Y., et al. (2010). Worldwide diversity of Klebsiella pneumoniae that produce beta-lactamase blaKPC-2 gene. Emerg. Infect. Dis. 16, 1349–1356.
Cuzon, G., Ouanich, J., Gondret, R., Naas, T., and Nordmann, P. (2011). Outbreak of OXA-48-positive carbapenem-resistant Klebsiella pneumoniae isolates in France. Antimicrob. Agents Chemother. 55, 2420–2423.
da Fonseca, é. L., Vieira, V. V., Cipriano, R., and Vicente, A. C. P. (2007). Emergence of blaGES-5 in clinical colistin-only-sensitive (COS) Pseudomonas aeruginosa strain in Brazil. J. Antimicrob. Chemother. 59, 576–577.
Darley, E., Weeks, J., Jones, L., Daniels, V., Wootton, M., Macgowan, A., et al. (2012). NDM-1 polymicrobial infections including Vibrio cholerae. Lancet 380, 1358.
Da Silva, G. J., Quinteira, S., Bértolo, E., Sousa, J. C., Gallego, L., Duarte, A., et al. (2004). Long-term dissemination of an OXA-40 carbapenemase-producing Acinetobacter baumannii clone in the Iberian Peninsula. J. Antimicrob. Chemother. 54, 255–258.
Decre, D., Birgand, G., Geneste, D., Maury, E., Petit, J. C., Barbut, F., et al. (2010). Possible importation and subsequent cross-transmission of OXA-48-producing Klebsiella pneumoniae, France, 2010. Euro Surveill. 15, pii: 19718.
Deris, Z. Z., Yu, H. H., Davis, K., Soon, R. L., Jacob, J., Ku, C. K., et al. (2012). The combination of colistin and doripenem is synergistic against Klebsiella pneumoniae at multiple inocula and suppresses colistin resistance in an in vitro pharmacokinetic/pharmacodynamic model. Antimicrob. Agents Chemother. 56, 5103–5112.
Deshpande, L. M., Jones, R. N., Fritsche, T. R., and Sader, H. S. (2006a). Occurrence and characterization of carbapenemase-producing Enterobacteriaceae: report from the SENTRY antimicrobial surveillance program (2000–2004). Microb. Drug Resist. 12, 223–230.
Deshpande, L. M., Rhomberg, P. R., Sader, H. S., and Jones, R. N. (2006b). Emergence of serine carbapenemases (KPC and SME) among clinical strains of Enterobacteriaceae isolated in the United States Medical Centers: report from the MYSTIC Program (1999–2005). Diagn. Microbiol. Infect. Dis. 56, 367–372.
Diancourt, L., Passet, V., Nemec, A., Dijkshoorn, L., and Brisse, S. (2010). The population structure of Acinetobacter baumannii: expanding multiresistant clones from an ancestral susceptible genetic pool. PLoS ONE 5:e10034. doi: 10.1371/journal.pone.0010034
Di Popolo, A., Giannouli, M., Triassi, M., Brisse, S., and Zarrilli, R. (2011). Molecular epidemiological investigation of multidrug-resistant Acinetobacter baumannii strains in four Mediterranean countries with a multilocus sequence typing scheme. Clin. Microbiol. Infect. 17, 197–201.
Docquier, J.-D., Calderone, V., De Luca, F., Benvenuti, M., Giuliani, F., Bellucci, L., et al. (2009). Crystal structure of the OXA-48 β-lactamase reveals mechanistic diversity among class D carbapenemases. Chem. Biol. 16, 540–547.
Dortet, L., Nordmann, P., and Poirel, L. (2012). Association of the emerging carbapenemase NDM-1 with a bleomycin resistance protein in Enterobacteriaceae and Acinetobacter baumannii. Antimicrob. Agents Chemother. 56, 1693–1697.
El-Gamal, M. I., and Oh, C. H. (2010). Current status of carbapenem antibiotics. Curr. Top. Med. Chem. 10, 1882–1897.
El Salabi, A., Borra, P. S., Toleman, M. A., Samuelsen, O., and Walsh, T. R. (2012). Genetic and biochemical characterization of a novel metallo-beta-lactamase, TMB-1, from an Achromobacter xylosoxidans strain isolated in Tripoli, Libya. Antimicrob. Agents Chemother. 56, 2241–2245.
Endimiani, A., Bethel, C., Choudhary, Y., and Bonomo, R. A. (2010a). In vitro activity of penem-1 in combination with beta-lactams against blaKPC-possessing Klebsiella pneumoniae isolates. Antimicrob. Agents Chemother. 54, 1650–1651.
Endimiani, A., Patel, G., Hujer, K. M., Swaminathan, M., Perez, F., Rice, L. B., et al. (2010b). In vitro activity of fosfomycin against blaKPC-containing Klebsiella pneumoniae isolates, including those nonsusceptible to tigecycline and/or colistin. Antimicrob. Agents Chemother. 54, 526–529.
Endimiani, A., Carias, L. L., Hujer, A. M., Bethel, C. R., Hujer, K. M., Perez, F., et al. (2008). Presence of plasmid-mediated quinolone resistance in Klebsiella pneumoniae isolates possessing blaKPC in the United States. Antimicrob. Agents Chemother. 52, 2680–2682.
Endimiani, A., Choudhary, Y., and Bonomo, R. A. (2009a). In vitro activity of NXL104 in combination with beta-lactams against Klebsiella pneumoniae isolates producing KPC carbapenemases. Antimicrob. Agents Chemother. 53, 3599–3601.
Endimiani, A., Hujer, A. M., Perez, F., Bethel, C. R., Hujer, K. M., Kroeger, J., et al. (2009b). Characterization of blaKPC-containing Klebsiella pneumoniae isolates detected in different institutions in the Eastern USA. J. Antimicrob. Chemother. 63, 427–437.
Endimiani, A., Hujer, K. M., Hujer, A. M., Armstrong, E. S., Choudhary, Y., Aggen, J. B., et al. (2009c). ACHN-490, a neoglycoside with potent in vitro activity against multidrug-resistant Klebsiella pneumoniae isolates. Antimicrob. Agents Chemother. 53, 4504–4507.
Endimiani, A., and Paterson, D. L. (2007). Optimizing therapy for infections caused by enterobacteriaceae producing extended-spectrum beta-lactamases. Semin. Respir. Crit. Care Med. 28, 646–655.
Evans, B. A., Brown, S., Hamouda, A., Findlay, J., and Amyes, S. G. B. (2007). Eleven novel OXA-51-like enzymes from clinical isolates of Acinetobacter baumannii. Clin. Microbiol. Infect. 13, 1137–1138.
Fairfax, M. R., Queenan, A. M., Lephart, P. R., Kaye, K. S., Dror, M., Arnous, N., et al. (2011). Detection of 2 SME-1 carbapenemase-producing Serratia marcescens in Detroit. Diagn. Microbiol. Infect. Dis. 71, 325–326.
Falagas, M. E., Kanellopoulou, M. D., Karageorgopoulos, D. E., Dimopoulos, G., Rafailidis, P. I., Skarmoutsou, N. D., et al. (2008). Antimicrobial susceptibility of multidrug-resistant Gram negative bacteria to fosfomycin. Eur. J. Clin. Microbiol. Infect. Dis. 27, 439–443.
Falagas, M. E., Karageorgopoulos, D. E., and Nordmann, P. (2011). Therapeutic options for infections with Enterobacteriaceae producing carbapenem-hydrolyzing enzymes. Future Microbiol. 6, 653–666.
Falagas, M. E., and Kasiakou, S. K. (2005). Colistin: the revival of polymyxins for the management of multidrug-resistant Gram-negative bacterial infections. Clin. Infect. Dis. 40, 1333–1341.
Falagas, M. E., Kastoris, A. C., Karageorgopoulos, D. E., and Rafailidis, P. I. (2009). Fosfomycin for the treatment of infections caused by multidrug-resistant non-fermenting Gram-negative bacilli: a systematic review of microbiological, animal and clinical studies. Int. J. Antimicrob. Agents 34, 111–120.
Figueiredo, S., Poirel, L., Croize, J., Recule, C., and Nordmann, P. (2009). In vivo selection of reduced susceptibility to carbapenems in Acinetobacter baumannii related to ISAba1-mediated overexpression of the natural blaOXA-66 oxacillinase gene. Antimicrob. Agents Chemother. 53, 2657–2659.
Fritsche, T. R., Strabala, P. A., Sader, H. S., Dowzicky, M. J., and Jones, R. N. (2005). Activity of tigecycline tested against a global collection of Enterobacteriaceae, including tetracycline-resistant isolates. Diagn. Microbiol. Infect. Dis. 52, 209–213.
Gales, A. C., Castanheira, M., Jones, R. N., and Sader, H. S. (2012). Antimicrobial resistance among Gram-negative bacilli isolated from Latin America: results from SENTRY Antimicrobial Surveillance Program (Latin America, 2008–2010). Diagn. Microbiol. Infect. Dis. 73, 354–360.
Geffen, Y., Adler, A., Paikin, S., Khabra, E., Gorenshtein, S., Aronov, R., et al. (2012). Detection of the plasmid-mediated KPC-2 carbapenem-hydrolysing enzyme in three unusual species of the Enterobacteriaceae family in Israel. J. Antimicrob. Chemother. 68, 719–720.
Ghazawi, A., Sonnevend, A., Bonnin, R. A., Poirel, L., Nordmann, P., Hashmey, R., et al. (2012). NDM-2 carbapenemase-producing Acinetobacter baumannii in the United Arab Emirates. Clin. Microbiol. Infect. 18, E34–E36.
Giakkoupi, P., Papagiannitsis, C. C., Miriagou, V., Pappa, O., Polemis, M., Tryfinopoulou, K., et al. (2011). An update of the evolving epidemic of blaKPC-2-carrying Klebsiella pneumoniae in Greece (2009–10). J. Antimicrob. Chemother. 66, 1510–1513.
Giakkoupi, P., Xanthaki, A., Kanelopoulou, M., Vlahaki, A., Miriagou, V., Kontou, S., et al. (2003). VIM-1 metallo-beta-lactamase-producing Klebsiella pneumoniae strains in Greek hospitals. J. Clin. Microbiol. 41, 3893–3896.
Giani, T., Conte, V., Di Pilato, V., Aschbacher, R., Weber, C., Larcher, C., et al. (2012). Escherichia coli from Italy producing OXA-48 carbapenemase encoded by a novel Tn1999 transposon derivative. Antimicrob. Agents Chemother. 56, 2211–2213.
Girlich, D., Poirel, L., and Nordmann, P. (2010). Novel ambler class A carbapenem-hydrolyzing beta-lactamase from a Pseudomonas fluorescens isolate from the Seine River, Paris, France. Antimicrob. Agents Chemother. 54, 328–332.
Gogou, V., Pournaras, S., Giannouli, M., Voulgari, E., Piperaki, E.-T., Zarrilli, R., et al. (2011). Evolution of multidrug-resistant Acinetobacter baumannii clonal lineages: a 10 year study in Greece (2000–09). J. Antimicrob. Chemother. 66, 2767–2772.
Gregory, C. J., Llata, E., Stine, N., Gould, C., Santiago, L. M., Vazquez, G. J., et al. (2010). Outbreak of carbapenem-resistant Klebsiella pneumoniae in Puerto Rico associated with a novel carbapenemase variant. Infect. Control Hosp. Epidemiol. 31, 476–484.
Grosso, F., Quinteira, S., and Peixe, L. (2011). Understanding the dynamics of imipenem-resistant Acinetobacter baumannii lineages within Portugal. Clin. Microbiol. Infect. 17, 1275–1279.
Gülmez, D., Woodford, N., Palepou, M.-F. I., Mushtaq, S., Metan, G., Yakupogullari, Y., et al. (2008). Carbapenem-resistant Escherichia coli and Klebsiella pneumoniae isolates from Turkey with OXA-48-like carbapenemases and outer membrane protein loss. Int. J. Antimicrob. Agents 31, 523–526.
Gupta, N., Limbago, B. M., Patel, J. B., and Kallen, A. J. (2011). Carbapenem-resistant Enterobacteriaceae: epidemiology and prevention. Clin. Infect. Dis. 53, 60–67.
Halaby, T., Reuland, A. E., Al Naiemi, N., Potron, A., Savelkoul, P. H., Vandenbroucke-Grauls, C. M., et al. (2012). A case of New Delhi metallo-beta-lactamase 1 (NDM-1)-producing Klebsiella pneumoniae with putative secondary transmission from the Balkan region in the Netherlands. Antimicrob. Agents Chemother. 56, 2790–2791.
Hays, C., Benouda, A., Poirel, L., Elouennass, M., and Nordmann, P. (2012). Nosocomial occurrence of OXA-48-producing enterobacterial isolates in a Moroccan hospital. Int. J. Antimicrob. Agents 39, 545–547.
Henriques, I., Moura, A., Alves, A., Saavedra, M. J., and Correia, A. (2004). Molecular characterization of a carbapenem-hydrolyzing class A beta-lactamase, SFC-1, from Serratia fonticola UTAD54. Antimicrob. Agents Chemother. 48, 2321–2324.
Héritier, C., Dubouix, A., Poirel, L., Marty, N., and Nordmann, P. (2005a). A nosocomial outbreak of Acinetobacter baumannii isolates expressing the carbapenem-hydrolysing oxacillinase OXA-58. J. Antimicrob. Chemother. 55, 115–118.
Héritier, C., Poirel, L., Fournier, P.-E., Claverie, J.-M., Raoult, D., and Nordmann, P. (2005b). Characterization of the naturally occurring oxacillinase of Acinetobacter baumannii. Antimicrob. Agents Chemother. 49, 4174–4179.
Héritier, C., Poirel, L., and Nordmann, P. (2004). Genetic and biochemical characterization of a chromosome-encoded carbapenem-hydrolyzing ambler class D β-lactamase from Shewanella algae. Antimicrob. Agents Chemother. 48, 1670–1675.
Higgins, P. G., Janssen, K., Fresen, M. M., Wisplinghoff, H., and Seifert, H. (2012a). Molecular epidemiology of Acinetobacter baumannii bloodstream isolates obtained in the United States from 1995 to 2004 using rep-PCR and multilocus sequence typing. J. Clin. Microbiol. 50, 3493–3500.
Higgins, P. G., Stefanik, D., Page, M. G. P., Hackel, M., and Seifert, H. (2012b). In vitro activity of the siderophore monosulfactam BAL30072 against meropenem-non-susceptible Acinetobacter baumannii. J. Antimicrob. Chemother. 67, 1167–1169.
Higgins, P. G., Poirel, L., Lehmann, M., Nordmann, P., and Seifert, H. (2009). OXA-143, a novel carbapenem-hydrolyzing class D beta-lactamase in Acinetobacter baumannii. Antimicrob. Agents Chemother. 53, 5035–5038.
Hirsch, E. B., Ledesma, K. R., Chang, K. T., Schwartz, M. S., Motyl, M. R., and Tam, V. H. (2012). In vitro activity of MK-7655, a novel beta-lactamase inhibitor, in combination with imipenem against carbapenem-resistant Gram-negative bacteria. Antimicrob. Agents Chemother. 56, 3753–3757.
Hirsch, E. B., and Tam, V. H. (2010). Detection and treatment options for Klebsiella pneumoniae carbapenemases (KPCs): an emerging cause of multidrug-resistant infection. J. Antimicrob. Chemother. 65, 1119–1125.
Hujer, K. M., Hujer, A. M., Hulten, E. A., Bajaksouzian, S., Adams, J. M., Donskey, C. J., et al. (2006). Analysis of antibiotic resistance genes in multidrug-resistant Acinetobacter sp. isolates from military and civilian patients treated at the Walter Reed Army Medical Center. Antimicrob. Agents Chemother. 50, 4114–4123.
Ishii, Y., Eto, M., Mano, Y., Tateda, K., and Yamaguchi, K. (2010). In vitro potentiation of carbapenems with ME1071, a novel metallo-beta-lactamase inhibitor, against metallo-beta-lactamase-producing Pseudomonas aeruginosa clinical isolates. Antimicrob. Agents Chemother. 54, 3625–3629.
Ito, H., Arakawa, Y., Ohsuka, S., Wacharotayankun, R., Kato, N., and Ohta, M. (1995). Plasmid-mediated dissemination of the metallo-beta-lactamase gene blaIMP among clinically isolated strains of Serratia marcescens. Antimicrob. Agents Chemother. 39, 824–829.
Jeong, S. H., Bae, I. K., Kim, D., Hong, S. G., Song, J. S., Lee, J. H., et al. (2005). First outbreak of Klebsiella pneumoniae clinical isolates producing GES-5 and SHV-12 extended-spectrum beta-lactamases in Korea. Antimicrob. Agents Chemother. 49, 4809–4810.
Jernigan, M. G., Press, E. G., Nguyen, M. H., Clancy, C. J., and Shields, R. K. (2012). The combination of doripenem and colistin is bactericidal and synergistic against colistin-resistant, carbapenemase-producing Klebsiella pneumoniae. Antimicrob. Agents Chemother. 56, 3395–3398.
Jovcic, B., Lepsanovic, Z., Suljagic, V., Rackov, G., Begovic, J., Topisirovic, L., et al. (2011). Emergence of NDM-1 metallo-beta-lactamase in Pseudomonas aeruginosa clinical isolates from Serbia. Antimicrob. Agents Chemother. 55, 3929–3931.
Kaase, M., Nordmann, P., Wichelhaus, T. A., Gatermann, S. G., Bonnin, R. A., and Poirel, L. (2011). NDM-2 carbapenemase in Acinetobacter baumannii from Egypt. J. Antimicrob. Chemother. 66, 1260–1262.
Karageorgopoulos, D. E., Wang, R., Yu, X.-H., and Falagas, M. E. (2012). Fosfomycin: evaluation of the published evidence on the emergence of antimicrobial resistance in Gram-negative pathogens. J. Antimicrob. Chemother. 67, 255–268.
Kassis-Chikhani, N., Decre, D., Gautier, V., Burghoffer, B., Saliba, F., Mathieu, D., et al. (2006). First outbreak of multidrug-resistant Klebsiella pneumoniae carrying blaVIM-1 and blaSHV-5 in a French university hospital. J. Antimicrob. Chemother. 57, 142–145.
King, D. T., Worrall, L. J., Gruninger, R., and Strynadka, N. C. (2012). New Delhi metallo-beta-lactamase: structural insights into beta-lactam recognition and inhibition. J. Am. Chem. Soc. 134, 11362–11365.
Kitchel, B., Rasheed, J. K., Endimiani, A., Hujer, A. M., Anderson, K. F., Bonomo, R. A., et al. (2010). Genetic factors associated with elevated carbapenem resistance in KPC-producing Klebsiella pneumoniae. Antimicrob. Agents Chemother. 54, 4201–4207.
Kitchel, B., Rasheed, J. K., Patel, J. B., Srinivasan, A., Navon-Venezia, S., Carmeli, Y., et al. (2009a). Molecular epidemiology of KPC-producing Klebsiella pneumoniae isolates in the United States: clonal expansion of multilocus sequence type 258. Antimicrob. Agents Chemother. 53, 3365–3370.
Kitchel, B., Sundin, D. R., and Patel, J. B. (2009b). Regional dissemination of KPC-producing Klebsiella pneumoniae. Antimicrob. Agents Chemother. 53, 4511–4513.
Kotsakis, S. D., Miriagou, V., Tzelepi, E., and Tzouvelekis, L. S. (2010). Comparative biochemical and computational study of the role of naturally occurring mutations at Ambler positions 104 and 170 in GES beta-lactamases. Antimicrob. Agents Chemother. 54, 4864–4871.
Kumarasamy, K. K., Toleman, M. A., Walsh, T. R., Bagaria, J., Butt, F., Balakrishnan, R., et al. (2010). Emergence of a new antibiotic resistance mechanism in India, Pakistan, and the UK: a molecular, biological, and epidemiological study. Lancet Infect. Dis. 10, 597–602.
Kus, J. V., Tadros, M., Simor, A., Low, D. E., Mcgeer, A. J., Willey, B. M., et al. (2011). New Delhi metallo-beta-lactamase-1: local acquisition in Ontario, Canada, and challenges in detection. CMAJ 183, 1257–1261.
Landman, D., Georgescu, C., Martin, D.A., and Quale, J. (2008). Polymyxins revisited. Clin. Microbiol. Rev. 21, 449–465.
Landman, D., Kelly, P., Bäcker, M., Babu, E., Shah, N., Bratu, S., et al. (2011). Antimicrobial activity of a novel aminoglycoside, ACHN-490, against Acinetobacter baumannii and Pseudomonas aeruginosa from New York City. J. Antimicrob. Chemother. 66, 332–334.
Lascols, C., Hackel, M., Marshall, S. H., Hujer, A. M., Bouchillon, S., Badal, R., et al. (2011). Increasing prevalence and dissemination of NDM-1 metallo-β-lactamase in India: data from the SMART study (2009). J. Antimicrob. Chemother. 66, 1992–1997.
Lascols, C., Peirano, G., Hackel, M., Laupland, K. B., and Pitout, J. D. (2012). Surveillance and molecular epidemiology of Klebsiella pneumoniae that produce carbapenemases; the first report of OXA-48-like enzymes in North America. Antimicrob Agents Chemother. 57, 130–136.
Lauretti, L., Riccio, M. L., Mazzariol, A., Cornaglia, G., Amicosante, G., Fontana, R., et al. (1999). Cloning and characterization of blaVIM, a new integron-borne metallo-beta-lactamase gene from a Pseudomonas aeruginosa clinical isolate. Antimicrob. Agents Chemother. 43, 1584–1590.
Lautenbach, E., Synnestvedt, M., Weiner, M. G., Bilker, W. B., Vo, L., Schein, J., et al. (2009). Epidemiology and impact of imipenem resistance in Acinetobacter baumannii. Infect. Control Hosp. Epidemiol. 30, 1186–1192.
Lautenbach, E., Synnestvedt, M., Weiner, M. G., Bilker, W. B., Vo, L., Schein, J., et al. (2010). Imipenem resistance in Pseudomonas aeruginosa: emergence, epidemiology, and impact on clinical and economic outcomes. Infect. Control Hosp. Epidemiol. 31, 47–53.
Lee, J., Patel, G., Huprikar, S., Calfee, D. P., and Jenkins, S. G. (2009). Decreased susceptibility to polymyxin B during treatment for carbapenem-resistant Klebsiella pneumoniae infection. J. Clin. Microbiol. 47, 1611–1612.
Lee, K., Yum, J. H., Yong, D., Lee, H. M., Kim, H. D., Docquier, J.-D., et al. (2005). Novel acquired metallo-beta-lactamase gene, blaSIM-1, in a class 1 integron from Acinetobacter baumannii clinical isolates from Korea. Antimicrob. Agents Chemother. 49, 4485–4491.
Levasseur, P., Girard, A. M., Claudon, M., Goossens, H., Black, M. T., Coleman, K., et al. (2012). In vitro antibacterial activity of the ceftazidime–avibactam (NXL104) combination against Pseudomonas aeruginosa clinical isolates. Antimicrob. Agents Chemother. 56, 1606–1608.
Li, J., Nation, R. L., Turnidge, J. D., Milne, R. W., Coulthard, K., Rayner, C. R., et al. (2006a). Colistin: the re-emerging antibiotic for multidrug-resistant Gram-negative bacterial infections. Lancet Infect. Dis. 6, 589–601.
Li, J., Rayner, C. R., Nation, R. L., Owen, R. J., Spelman, D., Tan, K. E., et al. (2006b). Heteroresistance to colistin in multidrug-resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 50, 2946–2950.
Livermore, D. M., Mushtaq, S., Morinaka, A., Ida, T., Maebashi, K., and Hope, R. (2013). Activity of carbapenems with ME1071 (disodium 2,3-diethylmaleate) against Enterobacteriaceae and Acinetobacter spp. with carbapenemases, including NDM enzymes. J. Antimicrob. Chemother. 68, 153–158.
Livermore, D. M., Mushtaq, S., and Warner, M. (2010). Activity of BAL30376 (monobactam BAL19764 + BAL29880 + clavulanate) versus Gram-negative bacteria with characterized resistance mechanisms. J. Antimicrob. Chemother. 65, 2382–2395.
Livermore, D. M., Mushtaq, S., Warner, M., Miossec, C., and Woodford, N. (2008). NXL104 combinations versus Enterobacteriaceae with CTX-M extended-spectrum beta-lactamases and carbapenemases. J. Antimicrob. Chemother. 62, 1053–1056.
Livermore, D. M., Mushtaq, S., Warner, M., Zhang, J.-C., Maharjan, S., Doumith, M., et al. (2011a). Activity of aminoglycosides, including ACHN-490, against carbapenem-resistant Enterobacteriaceae isolates. J. Antimicrob. Chemother. 66, 48–53.
Livermore, D. M., Mushtaq, S., Warner, M., Zhang, J., Maharjan, S., Doumith, M., et al. (2011b). Activities of NXL104 combinations with ceftazidime and aztreonam against carbapenemase-producing Enterobacteriaceae. Antimicrob. Agents Chemother. 55, 390–394.
Livermore, D. M., Walsh, T. R., Toleman, M., and Woodford, N. (2011c). Balkan NDM-1: escape or transplant? Lancet Infect. Dis. 11, 164.
Livermore, D. M., Warner, M., Mushtaq, S., Doumith, M., Zhang, J., and Woodford, N. (2011d). What remains against carbapenem-resistant Enterobacteriaceae? Evaluation of chloramphenicol, ciprofloxacin, colistin, fosfomycin, minocycline, nitrofurantoin, temocillin and tigecycline. Int. J. Antimicrob. Agents 37, 415–419.
Lolans, K., Rice, T. W., Munoz-Price, L. S., and Quinn, J. P. (2006). Multicity outbreak of carbapenem-resistant Acinetobacter baumannii isolates producing the carbapenemase OXA-40. Antimicrob. Agents Chemother. 50, 2941–2945.
Lopez-Otsoa, F., Gallego, L., Towner, K. J., Tysall, L., Woodford, N., and Livermore, D. M. (2002). Endemic carbapenem resistance associated with OXA-40 carbapenemase among Acinetobacter baumannii isolates from a hospital in Northern Spain. J. Clin. Microbiol. 40, 4741–4743.
Marchaim, D., Chopra, T., Bhargava, A., Bogan, C., Dhar, S., Hayakawa, K., et al. (2012). Recent exposure to antimicrobials and carbapenem-resistant Enterobacteriaceae: the role of antimicrobial stewardship. Infect. Control Hosp. Epidemiol. 33, 817–830.
Marchaim, D., Chopra, T., Perez, F., Hayakawa, K., Lephart, P. R., Bheemreddy, S., et al. (2011). Outcomes and genetic relatedness of carbapenem-resistant enterobacteriaceae at Detroit medical center. Infect. Control Hosp. Epidemiol. 32, 861–871.
Mataseje, L. F., Bryce, E., Roscoe, D., Boyd, D. A., Embree, J., Gravel, D., et al. (2012). Carbapenem-resistant Gram-negative bacilli in Canada 2009–10: results from the Canadian Nosocomial Infection Surveillance Program (CNISP). J. Antimicrob. Chemother. 67, 1359–1367.
Mathers, A. J., Cox, H. L., Kitchel, B., Bonatti, H., Brassinga, A. K., Carroll, J., et al. (2011). Molecular dissection of an outbreak of carbapenem-resistant enterobacteriaceae reveals intergenus KPC carbapenemase transmission through a promiscuous plasmid. MBio 2, e00204–e00211.
Mathers, A. J., Hazen, K. C., Carroll, J., Yeh, A. J., Cox, H. L., Bonomo, R. A., et al. (2012). First clinical cases of OXA-48 producing carbapenem resistant Klebsiella pneumoniae in the United States: the “menace” arrives in the New World. J. Clin. Microbiol. 51, 680–683.
Meletis, G., Tzampaz, E., Sianou, E., Tzavaras, I., and Sofianou, D. (2011). Colistin heteroresistance in carbapenemase-producing Klebsiella pneumoniae. J. Antimicrob. Chemother. 66, 946–947.
Mendes, R. E., Bell, J. M., Turnidge, J. D., Castanheira, M., and Jones, R. N. (2009). Emergence and widespread dissemination of OXA-23, -24/40 and -58 carbapenemases among Acinetobacter spp. in Asia-Pacific nations: report from the SENTRY Surveillance Program. J. Antimicrob. Chemother. 63, 55–59.
Michalopoulos, A., Virtzili, S., Rafailidis, P., Chalevelakis, G., Damala, M., and Falagas, M. E. (2010). Intravenous fosfomycin for the treatment of nosocomial infections caused by carbapenem-resistant Klebsiella pneumoniae in critically ill patients: a prospective evaluation. Clin. Microbiol. Infect. 16, 184–186.
Miriagou, V., Tzouvelekis, L. S., Rossiter, S., Tzelepi, E., Angulo, F. J., and Whichard, J. M. (2003). Imipenem resistance in a Salmonella clinical strain due to plasmid-mediated class A carbapenemase KPC-2. Antimicrob. Agents Chemother. 47, 1297–1300.
Morfin-Otero, R., Rodriguez-Noriega, E., Deshpande, L. M., Sader, H. S., and Castanheira, M. (2009). Dissemination of a bla(VIM-2)-carrying integron among Enterobacteriaceae species in Mexico: report from the SENTRY Antimicrobial Surveillance Program. Microb. Drug Resist. 15, 33–35.
Mostachio, A. K., Levin, A. S., Rizek, C., Rossi, F., Zerbini, J., and Costa, S. F. (2012). High prevalence of OXA-143 and alteration of outer membrane proteins in carbapenem-resistant Acinetobacter spp. isolates in Brazil. Int. J. Antimicrob. Agents 39, 396–401.
Moubareck, C., Bremont, S., Conroy, M. C., Courvalin, P., and Lambert, T. (2009). GES-11, a novel integron-associated GES variant in Acinetobacter baumannii. Antimicrob. Agents Chemother. 53, 3579–3581.
Mugnier, P. D., Poirel, L., Naas, T., and Nordmann, P. (2010). Worldwide dissemination of the blaOXA-23 carbapenemase gene of Acinetobacter baumannii. Emerg. Infect. Dis. 16, 35–40.
Mushtaq, S., Irfan, S., Sarma, J. B., Doumith, M., Pike, R., Pitout, J., et al. (2011). Phylogenetic diversity of Escherichia coli strains producing NDM-type carbapenemases. J. Antimicrob. Chemother. 66, 2002–2005.
Mushtaq, S., Warner, M., and Livermore, D. (2010a). Activity of the siderophore monobactam BAL30072 against multiresistant non-fermenters. J. Antimicrob. Chemother. 65, 266–270.
Mushtaq, S., Warner, M., and Livermore, D. M. (2010b). In vitro activity of ceftazidime + NXL104 against Pseudomonas aeruginosa and other non-fermenters. J. Antimicrob. Chemother. 65, 2376–2381.
Mushtaq, S., Warner, M., Williams, G., Critchley, I., and Livermore, D. M. (2010c). Activity of chequerboard combinations of ceftaroline and NXL104 versus β-lactamase-producing Enterobacteriaceae. J. Antimicrob. Chemother. 65, 1428–1432.
Naas, T., Cuzon, G., Villegas, M.-V., Lartigue, M.-F., Quinn, J. P., and Nordmann, P. (2008). Genetic structures at the origin of acquisition of the beta-lactamase blaKPC gene. Antimicrob. Agents Chemother. 52, 1257–1263.
Naas, T., Vandel, L., Sougakoff, W., Livermore, D. M., and Nordmann, P. (1994). Cloning and sequence analysis of the gene for a carbapenem-hydrolyzing class A beta-lactamase, Sme-1, from Serratia marcescens S6. Antimicrob. Agents Chemother. 38, 1262–1270.
Navon-Venezia, S., Leavitt, A., and Carmeli, Y. (2007). High tigecycline resistance in multidrug-resistant Acinetobacter baumannii. J. Antimicrob. Chemother. 59, 772–774.
Navon-Venezia, S., Leavitt, A., Schwaber, M. J., Rasheed, J. K., Srinivasan, A., Patel, J. B., et al. (2009). First report on a hyperepidemic clone of KPC-3-producing Klebsiella pneumoniae in Israel genetically related to a strain causing outbreaks in the United States. Antimicrob. Agents Chemother. 53, 818–820.
Nordmann, P., Boulanger, A. E., and Poirel, L. (2012a). NDM-4 metallo-beta-lactamase with increased carbapenemase activity from Escherichia coli. Antimicrob. Agents Chemother. 56, 2184–2186.
Nordmann, P., Couard, J.-P., Sansot, D., and Poirel, L. (2012b). Emergence of an autochthonous and community-acquired NDM-1–producing Klebsiella pneumoniae in Europe. Clin. Infect. Dis. 54, 150–151.
Nordmann, P., Gniadkowski, M., Giske, C. G., Poirel, L., Woodford, N., and Miriagou, V. (2012c). Identification and screening of carbapenemase-producing Enterobacteriaceae. Clin. Microbiol. Infect. 18, 432–438.
Nordmann, P., Cuzon, G., and Naas, T. (2009). The real threat of Klebsiella pneumoniae carbapenemase-producing bacteria. Lancet Infect. Dis. 9, 228–236.
Nordmann, P., Mariotte, S., Naas, T., Labia, R., and Nicolas, M. H. (1993). Biochemical properties of a carbapenem-hydrolyzing beta-lactamase from Enterobacter cloacae and cloning of the gene into Escherichia coli. Antimicrob. Agents Chemother. 37, 939–946.
Nordmann, P., Poirel, L., Walsh, T. R., and Livermore, D. M. (2011). The emerging NDM carbapenemases. Trends Microbiol. 19, 588–595.
Noskin, G. A. (2005). Tigecycline: a new glycylcycline for treatment of serious infections. Clin. Infect. Dis. 41, S303–S314.
Osano, E., Arakawa, Y., Wacharotayankun, R., Ohta, M., Horii, T., Ito, H., et al. (1994). Molecular characterization of an enterobacterial metallo beta-lactamase found in a clinical isolate of Serratia marcescens that shows imipenem resistance. Antimicrob. Agents Chemother. 38, 71–78.
Osterblad, M., Kirveskari, J., Hakanen, A. J., Tissari, P., Vaara, M., and Jalava, J. (2012). Carbapenemase-producing Enterobacteriaceae in Finland: the first years (2008–11). J. Antimicrob. Chemother. 67, 2860–2864.
Page, M. G., Dantier, C., and Desarbre, E. (2010). In vitro properties of BAL30072, a novel siderophore sulfactam with activity against multiresistant Gram-negative bacilli. Antimicrob. Agents Chemother. 54, 2291–2302.
Page, M. G., Dantier, C., Desarbre, E., Gaucher, B., Gebhardt, K., and Schmitt-Hoffmann, A. (2011). In vitro and in vivo properties of BAL30376, a beta-lactam and dual beta-lactamase inhibitor combination with enhanced activity against Gram-negative bacilli that express multiple beta-lactamases. Antimicrob. Agents Chemother. 55, 1510–1519.
Palepou, M. F., Woodford, N., Hope, R., Colman, M., Glover, J., Kaufmann, M., et al. (eds). (2005). “Novel class A carbapenemase, KPC-4, in an Enterobacter isolate from Scotland, abstr. 1134_01_20,” in Program and Abstracts of the 15th European Congress of Clinical Microbiology and Infectious Diseases, Copenhagen.
Papp-Wallace, K. M., Bethel, C. R., Distler, A. M., Kasuboski, C., Taracila, M., and Bonomo, R. A. (2010). Inhibitor resistance in the KPC-2 beta-lactamase, a preeminent property of this class A beta-lactamase. Antimicrob. Agents Chemother. 54, 890–897.
Patel, G., and Bonomo, R. A. (2011). Status report on carbapenemases: challenges and prospects. Expert Rev. Anti Infect. Ther. 9, 555–570.
Patel, G., Huprikar, S., Factor, S. H., Jenkins, S. G., and Calfee, D. P. (2008). Outcomes of carbapenem-resistant Klebsiella pneumoniae infection and the impact of antimicrobial and adjunctive therapies. Infect. Control Hosp. Epidemiol. 29, 1099–1106.
Paterson, D. L., and Bonomo, R. A. (2005). Extended-spectrum beta-lactamases: a clinical update. Clin. Microbiol. Rev. 18, 657–686.
Paton, R., Miles, R. S., Hood, J., and Amyes, S. G. (1993). ARI 1: beta-lactamase-mediated imipenem resistance in Acinetobacter baumannii. Int. J. Antimicrob. Agents 2, 81–87.
Peirano, G., Schreckenberger, P. C., and Pitout, J. D. (2011). Characteristics of NDM-1-producing Escherichia coli isolates that belong to the successful and virulent clone ST131. Antimicrob. Agents Chemother. 55, 2986–2988.
Perez, F., Endimiani, A., Ray, A. J., Decker, B. K., Wallace, C. J., Hujer, K. M., et al. (2010a). Carbapenem-resistant Acinetobacter baumannii and Klebsiella pneumoniae across a hospital system: impact of post-acute care facilities on dissemination. J. Antimicrob. Chemother. 65, 1807–1818.
Perez, F., Hujer, A. M., Hulten, E. A., Fishbain, J., Hujer, K. M., Aron, D., et al. (2010b). Antibiotic resistance determinants in Acinetobacter spp and clinical outcomes in patients from a major military treatment facility. Am. J. Infect. Control 38, 63–65.
Perez, F., Hujer, A. M., Hujer, K. M., Decker, B. K., Rather, P. N., and Bonomo, R. A. (2007). Global challenge of multidrug-resistant Acinetobacter baumannii. Antimicrob. Agents Chemother. 51, 3471–3484.
Pfeifer, Y., Schlatterer, K., Engelmann, E., Schiller, R. A., Frangenberg, H. R., Stiewe, D., et al. (2012). Emergence of OXA-48-type carbapenemase-producing Enterobacteriaceae in German hospitals. Antimicrob. Agents Chemother. 56, 2125–2128.
Pfeifer, Y., Wilharm, G., Zander, E., Wichelhaus, T. A., Göttig, S., Hunfeld, K.-P., et al. (2011a). Molecular characterization of blaNDM-1 in an Acinetobacter baumannii strain isolated in Germany in 2007. J. Antimicrob. Chemother. 66, 1998–2001.
Pfeifer, Y., Witte, W., Holfelder, M., Busch, J., Nordmann, P., and Poirel, L. (2011b). NDM-1-producing Escherichia coli in Germany. Antimicrob. Agents Chemother. 55, 1318–1319.
Poirel, L., Al Maskari, Z., Al Rashdi, F., Bernabeu, S., and Nordmann, P. (2010a). NDM-1-producing Klebsiella pneumoniae isolated in the Sultanate of Oman. J. Antimicrob. Chemother. 66, 304–306.
Poirel, L., Naas, T., and Nordmann, P. (2010b). Diversity, epidemiology, and genetics of class D beta-lactamases. Antimicrob. Agents Chemother. 54, 24–38.
Poirel, L., Rodriguez-Martinez, J.-M., Al Naiemi, N., Debets-Ossenkopp, Y. J., and Nordmann, P. (2010c). Characterization of DIM-1, an integron-encoded metallo-beta-lactamase from a Pseudomonas stutzeri clinical isolate in the Netherlands. Antimicrob. Agents Chemother. 54, 2420–2424.
Poirel, L., Bonnin, R. A., Boulanger, A., Schrenzel, J., Kaase, M., and Nordmann, P. (2012a). Tn125-related acquisition of blaNDM-like genes in Acinetobacter baumannii. Antimicrob. Agents Chemother. 56, 1087–1089.
Poirel, L., Bonnin, R. A., and Nordmann, P. (2012b). Genetic features of the widespread plasmid coding for the carbapenemase OXA-48. Antimicrob. Agents Chemother. 56, 559–562.
Poirel, L., Potron, A., and Nordmann, P. (2012c). OXA-48-like carbapenemases: the phantom menace. J. Antimicrob. Chemother. 67, 1597–1606.
Poirel, L., Bonnin, R. A., and Nordmann, P. (2011a). Analysis of the resistome of a multidrug-resistant NDM-1-producing Escherichia coli strain by high-throughput genome sequencing. Antimicrob. Agents Chemother. 55, 4224–4229.
Poirel, L., Castanheira, M., Carrer, A., Rodriguez, C. P., Jones, R. N., Smayevsky, J., et al. (2011b). OXA-163, an OXA-48-related class D beta-lactamase with extended activity toward expanded-spectrum cephalosporins. Antimicrob. Agents Chemother. 55, 2546–2551.
Poirel, L., Dortet, L., Bernabeu, S., and Nordmann, P. (2011c). Genetic features of blaNDM-1-positive Enterobacteriaceae. Antimicrob. Agents Chemother. 55, 5403–5407.
Poirel, L., Fortineau, N., and Nordmann, P. (2011d). International transfer of NDM-1-producing Klebsiella pneumoniae from Iraq to France. Antimicrob. Agents Chemother. 55, 1821–1822.
Poirel, L., Ros, A., Carrer, A., Fortineau, N., Carricajo, A., Berthelot, P., et al. (2011e). Cross-border transmission of OXA-48-producing Enterobacter cloacae from Morocco to France. J. Antimicrob. Chemother. 66, 1181–1182.
Poirel, L., Héritier, C., and Nordmann, P. (2004a). Chromosome-encoded ambler class D beta-lactamase of Shewanella oneidensis as a progenitor of carbapenem-hydrolyzing oxacillinase. Antimicrob. Agents Chemother. 48, 348–351.
Poirel, L., Héritier, C., and Nordmann, P. (2004b). Chromosome-encoded ambler class D β-lactamase of Shewanella oneidensis as a progenitor of carbapenem-hydrolyzing oxacillinase. Antimicrob. Agents Chemother. 48, 348–351.
Poirel, L., Héritier, C., Tolun, V., and Nordmann, P. (2004c). Emergence of oxacillinase-mediated resistance to imipenem in Klebsiella pneumoniae. Antimicrob. Agents Chemother. 48, 15–22.
Poirel, L., Le Thomas, I., Naas, T., Karim, A., and Nordmann, P. (2000a). Biochemical sequence analyses of GES-1, a novel class A extended-spectrum beta-lactamase, and the class 1 integron In52 from Klebsiella pneumoniae. Antimicrob. Agents Chemother. 44, 622–632.
Poirel, L., Naas, T., Nicolas, D., Collet, L., Bellais, S., Cavallo, J.-D., et al. (2000b). Characterization of VIM-2, a carbapenem-hydrolyzing metallo-beta-lactamase and its plasmid- and integron-borne gene from a Pseudomonas aeruginosa clinical isolate in France. Antimicrob. Agents Chemother. 44, 891–897.
Poirel, L., Marqué, S., Héritier, C., Segonds, C., Chabanon, G., and Nordmann, P. (2005). OXA-58, a novel class D beta-lactamase involved in resistance to carbapenems in Acinetobacter baumannii. Antimicrob. Agents Chemother. 49, 202–208.
Poirel, L., and Nordmann, P. (2006). Carbapenem resistance in Acinetobacter baumannii: mechanisms and epidemiology. Clin. Microbiol. Infect. 12, 826–836.
Poirel, L., Weldhagen, G. F., De Champs, C., and Nordmann, P. (2002). A nosocomial outbreak of Pseudomonas aeruginosa isolates expressing the extended-spectrum β-lactamase GES-2 in South Africa. J. Antimicrob. Chemother. 49, 561–565.
Poirel, L., Wenger, A., Bille, J., Bernabeu, S., Naas, T., and Nordmann, P. (2007). SME-2-producing Serratia marcescens isolate from Switzerland. Antimicrob. Agents Chemother. 51, 2282–2283.
Pollini, S., Maradei, S., Pecile, P., Olivo, G., Luzzaro, F., Docquier, J. D., et al. (2012). FIM-1, a new acquired metallo-beta-lactamase from a Pseudomonas aeruginosa clinical isolate from Italy. Antimicrob. Agents Chemother. 57, 410–416.
Potron, A., Kalpoe, J., Poirel, L., and Nordmann, P. (2011a). European dissemination of a single OXA-48-producing Klebsiella pneumoniae clone. Clin. Microbiol. Infect. 17, E24–E26.
Potron, A., Poirel, L., and Nordmann, P. (2011b). Origin of OXA-181, an emerging carbapenem-hydrolyzing oxacillinase, as a chromosomal gene in Shewanella xiamenensis. Antimicrob. Agents Chemother. 55, 4405–4407.
Potron, A., Nordmann, P., and Poirel, L. (2013). Characterization of OXA-204, a carbapenem-hydrolyzing class D β-lactamase from Klebsiella pneumoniae. Antimicrob. Agents Chemother. 57, 633–636.
Pottumarthy, S., Moland, E. S., Juretschko, S., Swanzy, S. R., Thomson, K. S., and Fritsche, T. R. (2003). NmcA carbapenem-hydrolyzing enzyme in Enterobacter cloacae in North America. Emerg. Infect. Dis. 9, 999–1002.
Poudyal, A., Howden, B. P., Bell, J. M., Gao, W., Owen, R. J., Turnidge, J. D., et al. (2008). In vitro pharmacodynamics of colistin against multidrug-resistant Klebsiella pneumoniae. J. Antimicrob. Chemother. 62, 1311–1318.
Prabaker, K., Lin, M. Y., McNally, M., Cherabuddi, K., Ahmed, S., Norris, A., et al. (2012). Transfer from high-acuity long-term care facilities is associated with carriage of Klebsiella pneumoniae carbapenemase-producing Enterobacteriaceae: a multihospital study. Infect. Control Hosp. Epidemiol. 33, 1193–1199.
Qi, Y., Wei, Z., Ji, S., Du, X., Shen, P., and Yu, Y. (2011). ST11, the dominant clone of KPC-producing Klebsiella pneumoniae in China. J. Antimicrob. Chemother. 66, 307–312.
Queenan, A. M., Shang, W., Schreckenberger, P., Lolans, K., Bush, K., and Quinn, J. (2006). SME-3, a novel member of the Serratia marcescens SME family of carbapenem-hydrolyzing beta-lactamases. Antimicrob. Agents Chemother. 50, 3485–3487.
Queenan, A. M., Torres-Viera, C., Gold, H. S., Carmeli, Y., Eliopoulos, G. M., Moellering, R. C. Jr., et al. (2000). SME-type carbapenem-hydrolyzing class A beta-lactamases from geographically diverse Serratia marcescens strains. Antimicrob. Agents Chemother. 44, 3035–3039.
Qureshi, Z. A., Paterson, D. L., Potoski, B. A., Kilayko, M. C., Sandovsky, G., Sordillo, E., et al. (2012). Treatment outcome of bacteremia due to KPC-producing Klebsiella pneumoniae: superiority of combination antimicrobial regimens. Antimicrob. Agents Chemother. 56, 2108–2113.
Rasheed, J. K., Biddle, J. W., Anderson, K. F., Washer, L., Chenoweth, C., Perrin, J., et al. (2008). Detection of the Klebsiella pneumoniae carbapenemase type 2 carbapenem-hydrolyzing enzyme in clinical isolates of Citrobacter freundii and K. oxytoca carrying a common plasmid. J. Clin. Microbiol. 46, 2066–2069.
Rasmussen, B., Bush, K., Keeney, D., Yang, Y., Hare, R., O'Gara, C., et al. (1996). Characterization of IMI-1 beta-lactamase, a class A carbapenem- hydrolyzing enzyme from Enterobacter cloacae. Antimicrob. Agents Chemother. 40, 2080–2086.
Riccio, M. L., Franceschini, N., Boschi, L., Caravelli, B., Cornaglia, G., Fontana, R., et al. (2000). Characterization of the metallo-beta-lactamase determinant of Acinetobacter baumannii AC-54/97 reveals the existence of bla(IMP) allelic variants carried by gene cassettes of different phylogeny. Antimicrob. Agents Chemother. 44, 1229–1235.
Rice, L. B., Carias, L. L., Hutton, R. A., Rudin, S. D., Endimiani, A., and Bonomo, R. A. (2008). The KQ element, a complex genetic region conferring transferable resistance to carbapenems, aminoglycosides, and fluoroquinolones in Klebsiella pneumoniae. Antimicrob. Agents Chemother. 52, 3427–3429.
Robledo, I. E., Aquino, E. E., Sante, M. I., Santana, J. L., Otero, D. M., Leon, C. F., et al. (2010). Detection of KPC in Acinetobacter spp. in Puerto Rico. Antimicrob. Agents Chemother. 54, 1354–1357.
Robledo, I. E., Moland, E. S., Aquino, E. A., Vazquez, G. J., Sante, M. I., Bertran, J., et al. (2007). “First report of a KPC-4 and CTX-M producing K. pneumoniae (KP) isolated from Puerto Rico, abstr. C2-1933,” in 47th Annual Interscience Conference on Antimicrobial Agents and Chemotherapy (Washington, DC: American Society for Microbiology).
Robledo, I. E., Vázquez, G. J., Aquino, E. A., Moland, E. S., Santé, M. I., and Hanson, N. D. (2008). “A novel KPC variant, KPC-6, in a Klebsiella pneumoniae (Kp) isolated in Puerto Rico (PR), abstr. C2-3738,” in 48th Annual Interscience Conference on Antimicrobial Agents and Chemotherapy and the 46th Annual Meeting of Infectious Diseases Society of America (Washington, DC: American Society for Microbiology).
Rodriguez-Rojas, A., Couce, A., and Blazquez, J. (2010). Frequency of spontaneous resistance to fosfomycin combined with different antibiotics in Pseudomonas aeruginosa. Antimicrob. Agents Chemother. 54, 4948–4949.
Rodvold, K. A., Gotfried, M. H., Cwik, M., Korth-Bradley, J. M., Dukart, G., and Ellis-Grosse, E. J. (2006). Serum, tissue and body fluid concentrations of tigecycline after a single 100 mg dose. J. Antimicrob. Chemother. 58, 1221–1229.
Rogers, B. A., Aminzadeh, Z., Hayashi, Y., and Paterson, D. L. (2011). Country-to-country transfer of patients and the risk of multi-resistant bacterial infection. Clin. Infect. Dis. 53, 49–56.
Rossi, F. (2011). The challenges of antimicrobial resistance in Brazil. Clin. Infect. Dis. 52, 1138–1143.
Russo, T. A., Page, M. G. P., Beanan, J. M., Olson, R., Hujer, A. M., Hujer, K. M., et al. (2011). In vivo and in vitro activity of the siderophore monosulfactam BAL30072 against Acinetobacter baumannii. J. Antimicrob. Chemother. 66, 867–873.
Salabi, A. E., Toleman, M. A., Weeks, J., Bruderer, T., Frei, R., and Walsh, T. R. (2010). First report of the metallo-beta-lactamase SPM-1 in Europe. Antimicrob. Agents Chemother. 54, 582.
Samuelsen, O., Naseer, U., Tofteland, S., Skutlaberg, D. H., Onken, A., Hjetland, R., et al. (2009). Emergence of clonally related Klebsiella pneumoniae isolates of sequence type 258 producing plasmid-mediated KPC carbapenemase in Norway and Sweden. J. Antimicrob. Chemother. 63, 654–658.
Satlin, M. J., Kubin, C. J., Blumenthal, J. S., Cohen, A. B., Furuya, E. Y., Wilson, S. J., et al. (2011). Comparative effectiveness of aminoglycosides, polymyxin B, and tigecycline for clearance of carbapenem-resistant Klebsiella pneumoniae from urine. Antimicrob. Agents Chemother. 55, 5893–5899.
Schwaber, M. J., Klarfeld-Lidji, S., Navon-Venezia, S., Schwartz, D., Leavitt, A., and Carmeli, Y. (2008). Predictors of carbapenem-resistant Klebsiella pneumoniae acquisition among hospitalized adults and effect of acquisition on mortality. Antimicrob. Agents Chemother. 52, 1028–1033.
Schwaber, M. J., Lev, B., Israeli, A., Solter, E., Smollan, G., Rubinovitch, B., et al. (2011). Containment of a country-wide outbreak of carbapenem-resistant Klebsiella pneumoniae in Israeli hospitals via a nationally implemented intervention. Clin. Infect. Dis. 52, 848–855.
Scott, P., Deye, G., Srinivasan, A., Murray, C., Moran, K., Hulten, E., et al. (2007). An outbreak of multidrug-resistant Acinetobacter baumannii–calcoaceticus complex infection in the US military health care system associated with military operations in Iraq. Clin. Infect. Dis. 44, 1577–1584.
Sekiguchi, J.-I., Morita, K., Kitao, T., Watanabe, N., Okazaki, M., Miyoshi-Akiyama, T., et al. (2008). KHM-1, a novel plasmid-mediated metallo-beta-lactamase from a Citrobacter freundii clinical isolate. Antimicrob. Agents Chemother. 52, 4194–4197.
Shahid, M., Sobia, F., Singh, A., Malik, A., Khan, H. M., Jonas, D., et al. (2009). Beta-lactams and beta-lactamase-inhibitors in current- or potential-clinical practice: a comprehensive update. Crit. Rev. Microbiol. 35, 81–108.
Snitkin, E. S., Zelazny, A. M., Thomas, P. J., Stock, F., Program, N. C. S., Henderson, D. K., et al. (2012). Tracking a hospital outbreak of carbapenem-resistant Klebsiella pneumoniae with whole-genome sequencing. Sci. Transl. Med. 4, 148ra116.
Souli, M., Galani, I., Boukovalas, S., Gourgoulis, M. G., Chryssouli, Z., Kanellakopoulou, K., et al. (2011). In vitro interactions of antimicrobial combinations with fosfomycin against KPC-2-producing Klebsiella pneumoniae and protection of resistance development. Antimicrob. Agents Chemother. 55, 2395–2397.
Struelens, M. J., Monnet, D. L., Magiorakos, A. P., Santos O'Connor, F., and Giesecke, J. (2010). New Delhi metallo-beta-lactamase 1-producing Enterobacteriaceae: emergence and response in Europe. Euro Surveill. 15. pii: 19716.
Sun, Y., Cai, Y., Liu, X., Bai, N., Liang, B., and Wang, R. (2012). The emergence of clinical resistance to tigecycline. Int. J. Antimicrob. Agents 41, 110–116.
Tasina, E., Haidich, A.-B., Kokkali, S., and Arvanitidou, M. (2011). Efficacy and safety of tigecycline for the treatment of infectious diseases: a meta-analysis. Lancet Infect. Dis. 11, 834–844.
Tibbetts, R., Frye, J. G., Marschall, J., Warren, D., and Dunne, W. (2008). Detection of KPC-2 in a clinical isolate of Proteus mirabilis and first reported description of carbapenemase resistance caused by a KPC beta-lactamase in P. mirabilis. J. Clin. Microbiol. 46, 3080–3083.
Toleman, M. A., Simm, A. M., Murphy, T. A., Gales, A. C., Biedenbach, D. J., Jones, R. N., et al. (2002). Molecular characterization of SPM-1, a novel metallo-β-lactamase isolated in Latin America: report from the SENTRY antimicrobial surveillance programme. J. Antimicrob. Chemother. 50, 673–679.
Toleman, M. A., Spencer, J., Jones, L., and Walsh, T. R. (2012). blaNDM-1 is a chimera likely constructed in Acinetobacter baumannii. Antimicrob. Agents Chemother. 56, 2773–2776.
Tumbarello, M., Viale, P., Viscoli, C., Trecarichi, E. M., Tumietto, F., Marchese, A., et al. (2012). Predictors of mortality in bloodstream infections caused by Klebsiella pneumoniae carbapenemase-producing K. pneumoniae: importance of combination therapy. Clin. Infect. Dis. 55, 943–950.
Urban, C., Bradford, P. A., Tuckman, M., Segal-Maurer, S., Wehbeh, W., Grenner, L., et al. (2008). Carbapenem-resistant Escherichia coli harboring Klebsiella pneumoniae carbapenemase β-lactamases associated with long-term care facilities. Clin. Infect. Dis. 46, e127–e130.
Urban, C., Mariano, N., and Rahal, J. J. (2010). In vitro double and triple bactericidal activities of doripenem, polymyxin B, and rifampin against multidrug-resistant Acinetobacter baumannii, Pseudomonas aeruginosa, Klebsiella pneumoniae, and Escherichia coli. Antimicrob. Agents Chemother. 54, 2732–2734.
Vaara, M., Sader, H. S., Rhomberg, P. R., Jones, R. N., and Vaara, T. (2012). Antimicrobial activity of the novel polymyxin derivative NAB739 tested against Gram-negative pathogens. J. Antimicrob. Chemother. 68, 636–639.
Vaara, M., Siikanen, O., Apajalahti, J., Frimodt-Moller, N., and Vaara, T. (2010). Susceptibility of carbapenemase-producing strains of Klebsiella pneumoniae and Escherichia coli to the direct antibacterial activity of NAB739 and to the synergistic activity of NAB7061 with rifampicin and clarithromycin. J. Antimicrob. Chemother. 65, 942–945.
van der Bij, A. K., and Pitout, J. D. (2012). The role of international travel in the worldwide spread of multiresistant Enterobacteriaceae. J. Antimicrob. Chemother. 67, 2090–2100.
Verde, P. E., and Curcio, D. (2012). Imbalanced mortality evidence for tigecycline: 2011, the year of the meta-analysis. Clin. Infect. Dis. 55, 471–472.
Viau, R. A., Hujer, A. M., Marshall, S. H., Perez, F., Hujer, K. M., Briceño, D. F., et al. (2012). “Silent” dissemination of Klebsiella pneumoniae isolates bearing K. pneumoniae carbapenemase in a long-term care facility for children and young adults in Northeast Ohio. Clin. Infect. Dis. 54, 1314–1321.
Villegas, M. V., Lolans, K., Correa, A., Kattan, J. N., Lopez, J. A., and Quinn, J. P. (2007). First identification of Pseudomonas aeruginosa isolates producing a KPC-type carbapenem-hydrolyzing beta-lactamase. Antimicrob. Agents Chemother. 51, 1553–1555.
Voulgari, E., Zarkotou, O., Ranellou, K., Karageorgopoulos, D. E., Vrioni, G., Mamali, V., et al. (2012). Outbreak of OXA-48 carbapenemase-producing Klebsiella pneumoniae in Greece involving an ST11 clone. J. Antimicrob. Chemother. 68, 84–88.
Vourli, S., Giakkoupi, P., Miriagou, V., Tzelepi, E., Vatopoulos, A. C., and Tzouvelekis, L. S. (2004). Novel GES/IBC extended-spectrum β-lactamase variants with carbapenemase activity in clinical enterobacteria. FEMS Microbiol. Lett. 234, 209–213.
Wachino, J., Doi, Y., Yamane, K., Shibata, N., Yagi, T., Kubota, T., et al. (2004). Molecular characterization of a cephamycin-hydrolyzing and inhibitor-resistant class A beta-lactamase, GES-4, possessing a single G170S substitution in the omega-loop. Antimicrob. Agents Chemother. 48, 2905–2910.
Wachino, J., Yoshida, H., Yamane, K., Suzuki, S., Matsui, M., Yamagishi, T., et al. (2011). SMB-1, a novel subclass B3 metallo-beta-lactamase, associated with ISCR1 and a class 1 integron, from a carbapenem-resistant Serratia marcescens clinical isolate. Antimicrob. Agents Chemother. 55, 5143–5149.
Walkty, A., Decorby, M., Lagace-Wiens, P. R., Karlowsky, J. A., Hoban, D. J., and Zhanel, G. G. (2011). In vitro activity of ceftazidime combined with NXL104 versus Pseudomonas aeruginosa isolates obtained from patients in Canadian hospitals (CANWARD 2009 study). Antimicrob. Agents Chemother. 55, 2992–2994.
Walsh, T. R., Toleman, M. A., Poirel, L., and Nordmann, P. (2005). Metallo-beta-lactamases: the quiet before the storm? Clin. Microbiol. Rev. 18, 306–325.
Walsh, T. R., Weeks, J., Livermore, D. M., and Toleman, M. A. (2011). Dissemination of NDM-1 positive bacteria in the New Delhi environment and its implications for human health: an environmental point prevalence study. Lancet Infect. Dis. 11, 355–362.
Walther-Rasmussen, J., and Høiby, N. (2007). Class A carbapenemases. J. Antimicrob. Chemother. 60, 470–482.
Wang, Y. F., and Dowzicky, M. J. (2010). In vitro activity of tigecycline and comparators on Acinetobacter spp. isolates collected from patients with bacteremia and MIC change during the Tigecycline Evaluation and Surveillance Trial, 2004 to 2008. Diagn. Microbiol. Infect. Dis. 68, 73–79.
Werneck, J. S., Picao, R. C., Girardello, R., Cayo, R., Marguti, V., Dalla-Costa, L., et al. (2011). Low prevalence of blaOXA-143 in private hospitals in Brazil. Antimicrob. Agents Chemother. 55, 4494–4495; author reply 4495.
Wolter, D. J., Kurpiel, P. M., Woodford, N., Palepou, M. F., Goering, R. V., and Hanson, N. D. (2009). Phenotypic and enzymatic comparative analysis of the novel KPC variant KPC-5 and its evolutionary variants, KPC-2 and KPC-4. Antimicrob. Agents Chemother. 53, 557–562.
Woodford, N., Tierno, P. M. Jr., Young, K., Tysall, L., Palepou, M. F., Ward, E., et al. (2004). Outbreak of Klebsiella pneumoniae producing a new carbapenem-hydrolyzing class A beta-lactamase, KPC-3, in a New York Medical Center. Antimicrob. Agents Chemother. 48, 4793–4799.
Woodford, N., Turton, J. F., and Livermore, D. M. (2011). Multiresistant Gram-negative bacteria: the role of high-risk clones in the dissemination of antibiotic resistance. FEMS Microbiol. Rev. 35, 736–755.
Woodford, N., Zhang, J., Warner, M., Kaufmann, M. E., Matos, J., Macdonald, A., et al. (2008). Arrival of Klebsiella pneumoniae producing KPC carbapenemase in the United Kingdom. J. Antimicrob. Chemother. 62, 1261–1264.
Yahav, D., Lador, A., Paul, M., and Leibovici, L. (2011). Efficacy and safety of tigecycline: a systematic review and meta-analysis. J. Antimicrob. Chemother. 66, 1963–1971.
Yang, Y. J., Wu, P. J., and Livermore, D. M. (1990). Biochemical characterization of a beta-lactamase that hydrolyzes penems and carbapenems from two Serratia marcescens isolates. Antimicrob. Agents Chemother. 34, 755–758.
Yau, W., Owen, R. J., Poudyal, A., Bell, J. M., Turnidge, J. D., Yu, H. H., et al. (2009). Colistin hetero-resistance in multidrug-resistant Acinetobacter baumannii clinical isolates from the Western Pacific region in the SENTRY antimicrobial surveillance programme. J. Infect. 58, 138–144.
Yigit, H., Queenan, A. M., Anderson, G. J., Domenech-Sanchez, A., Biddle, J. W., Steward, C. D., et al. (2001). Novel carbapenem-hydrolyzing beta-lactamase, KPC-1, from a carbapenem-resistant strain of Klebsiella pneumoniae. Antimicrob. Agents Chemother. 45, 1151–1161.
Yigit, H., Queenan, A. M., Rasheed, J. K., Biddle, J. W., Domenech-Sanchez, A., Alberti, S., et al. (2003). Carbapenem-resistant strain of Klebsiella oxytoca harboring carbapenem-hydrolyzing beta-lactamase KPC-2. Antimicrob. Agents Chemother. 47, 3881–3889.
Yong, D., Toleman, M. A., Giske, C. G., Cho, H. S., Sundman, K., Lee, K., et al. (2009). Characterization of a new metallo-beta-lactamase gene, bla(NDM-1), and a novel erythromycin esterase gene carried on a unique genetic structure in Klebsiella pneumoniae sequence type 14 from India. Antimicrob. Agents Chemother. 53, 5046–5054.
Yong, D., Walsh, T. R., Bell, J., Ritchie, B., Pratt, R., and Toleman, M. A. (2007). “A novel subgroup metallo β-lactamase, AIM-1, emerges in Pseudomonas aeruginosa from Australia abstr. C1-593,” in 47th Annual Interscience Conference on Antimicrobial Agents and Chemotherapy (Washington, DC: American Society for Microbiology).
Yu, Y.-S., Du, X.-X., Zhou, Z.-H., Chen, Y.-G., and Li, L.-J. (2006). First isolation of blaIMI-2 in an Enterobacter cloacae clinical isolate from China. Antimicrob. Agents Chemother. 50, 1610–1611.
Zarrilli, R., Pournaras, S., Giannouli, M., and Tsakris, A. (2013). Global evolution of multidrug-resistant Acinetobacter baumannii clonal lineages. Int. J. Antimicrob. Agents 41, 11–19.
Zhanel, G. G., Lawson, C. D., Zelenitsky, S., Findlay, B., Schweizer, F., Adam, H., et al. (2012). Comparison of the next-generation aminoglycoside plazomicin to gentamicin, tobramycin and amikacin. Expert Rev. Anti Infect. Ther. 10, 459–473.
Keywords: carbapenemases, NDM-1, KPC, OXA-48, metallo-β-lactamases, CHDL
Citation: Patel G and Bonomo RA (2013) “Stormy waters ahead”: global emergence of carbapenemases. Front. Microbiol. 4:48. doi: 10.3389/fmicb.2013.00048
Received: 19 December 2012; Paper pending published: 07 January 2013;
Accepted: 20 February 2013; Published online: 14 March 2013.
Edited by:
Fiona Walsh, Agroscope Changins-Wädenswil, SwitzerlandReviewed by:
Charles W. Knapp, University of Strathclyde, UKYoshikazu Ishii, Toho University School of Medicine, Japan
Copyright: © 2013 Patel and Bonomo. This is an open-access article distributed under the terms of the Creative Commons Attribution License, which permits use, distribution and reproduction in other forums, provided the original authors and source are credited and subject to any copyright notices concerning any third-party graphics etc.
*Correspondence: Robert A. Bonomo, Research Service, Louis Stokes Cleveland Department of Veteran Affairs Medical Center, 10701 East Boulevard, Cleveland, OH 44106, USA. e-mail:cm9iZXJ0LmJvbm9tb0BtZWQudmEuZ292