 Rocío Colello
Rocío Colello María E. Cáceres
María E. Cáceres Analía I. Etcheverría
Analía I. Etcheverría Nora L. Padola
Nora L. Padola- Laboratorio de Inmunoquímica y Biotecnología, Centro de Investigación Veterinaria de Tandil – Consejo Nacional de Investigaciones Científicas y Técnicas – Comisión de Investigaciones Científicas de la Provincia de Buenos Aires, Facultad de Ciencias Veterinarias, Universidad Nacional del Centro de la Provincia de Buenos Aires, Tandil, Argentina
Pigs are important reservoirs of Shiga toxin-producing Escherichia coli (STEC). The entrance of these strains into the food chain implies a risk to consumers because of the severity of hemolytic uremic syndrome. This study reports the prevalence and characterization of STEC throughout the pork production chain. From 764 samples, 31 (4.05%) were stx positive by PCR screening. At farms, 2.86% of samples were stx positive; at slaughter, 4.08% of carcasses were stx positive and at boning rooms, 6% of samples were stx positive. These percentages decreased in pork meat ready for sale at sales markets (4.59%). From positive samples, 50 isolates could be characterized. At farms 37.5% of the isolates carried stx1/stx2 genes, 37.5% possessed stx2e and 25%, carried only stx2. At slaughter we detected 50% of isolates positive for stx2, 33% for stx2e, and 16% for stx1/stx2. At boning rooms 59% of the isolates carried stx1/stx2, 14% stx2e, and 5% stx1/stx2/stx2e. At retail markets 66% of isolates were positive for stx2, 17% stx2e, and 17% stx1/stx2. For the other virulence factors, ehxA and saa were not detected and eae gene was detected in 12% of the isolates. Concerning putative adhesins, agn43 was detected in 72%, ehaA in 26%, aida in 8%, and iha in 6% of isolates. The strains were typed into 14 E. coli O groups (O1, O2, O8, O15, O20, O35, O69, O78, O91, O121, O138, O142, O157, O180) and 10 H groups (H9, H10, H16, H21, H26, H29, H30, H32, H45, H46). This study reports the prevalence and characterization of STEC strains through the chain pork suggesting the vertical transmission. STEC contamination originates in the farms and is transferred from pigs to carcasses in the slaughter process and increase in meat pork at boning rooms and sales markets. These results highlight the need to implement an integrated STEC control system based on good management practices on the farm and critical control point systems in the food chain.
Introduction
Shiga toxin-producing Escherichia coli (STEC) are important foodborne pathogens that can cause severe disease, including a life-threatening complication such as bloody diarrhea and hemolytic uremic syndrome (HUS; Paton and Paton, 1998). HUS is one of the most common etiologies for acute kidney injury and an important cause of acquired chronic kidney disease in children (Grisaru, 2014). This damage is produced by the action of cytotoxins Stx1 and Stx2, being Stx2 and their subtypes associated more frequently with HUS (Beutin et al., 2007). The ability to adhere to epithelial cells is an important virulence trait, because adherence presumably enables to deliver toxins efficiently to host organs (Tarr et al., 2000). Intimin, encoded by eae gene, is required for intimate bacterial adhesion to epithelial cells inducing a characteristic histopathological lesion defined as “attaching and effacing” (A/E) and has been considered as a risk factor for disease in human (Ethelberg et al., 2004). However, the presence of eae would not be essential for pathogenesis, considering that some eae negative STEC have been associated with severe disease in human (Paton et al., 2001). Some studies reported adherence factors other than intimin, such as Saa (Paton et al., 2001), AIDA and Agn43 (Restieri et al., 2007), EhaA (Wu et al., 2010), Iha (Tarr et al., 2000; Szalo et al., 2002). AIDA was identified in diffusely adhering diarrheagenic E. coli strain and is associated with edema disease and diarrhea in pigs (Niewerth et al., 2001), contributing to bacterial intercellular aggregation and biofilm formation (Restieri et al., 2007); iha encode for an outer membrane protein identified as a bacterial adherence conferring iron regulated gene (Tarr et al., 2000) and Agn43 and EhaA are autotransporter proteins of O157:H7 involved in adhesion and biofilm formation (Wells et al., 2008). Other factors are also involved in human pathogenicity such as a plasmid that encoded enterohemolysin (EhxA), among others (Feng and Reddy, 2013).
Argentina, where the HUS is endemic, hold the highest record worldwide of this syndrome with an incidence of 17/100,000 children less than 5 years old (Rivas et al., 2010). Although STEC O157:H7 is recognized as the most important serotype associated with human infection, there are more than 400 non-O157 serotypes that have been involved in human disease and isolated from different reservoirs including cattle, pigs, goats, sheep, cats, and dogs (Parma et al., 2000; Padola et al., 2004; Amézquita-López et al., 2014). STEC usually do not produce disease in animals, however, the Stx2e subtype is involved in edema disease in pigs, a peracute toxemia characterized by vascular necrosis, edema, neurological signs and that in some cases can be fatal (Niewerth et al., 2001). STEC strains have been isolated from pork products and have been associated with human infections as diarrhea and HUS, including strains harboring stx2e subtype (Sonntag et al., 2005; Kaufmann et al., 2006; Trotz-Williams et al., 2012); however, it is unknown if the contamination of pork- derivate food occurs during the processing or by cross contamination (Tseng et al., 2014).
There is an increase in worldwide demand for fast-growing species with efficient feed conversion rates, such as pigs, because they represent a major share in the growth in the livestock subsector (Food and Agriculture Organization [FAO], 2014). Because of the limited epidemiologic data of STEC in pork and the increasing role of non-O157 STEC in human illnesses, it is very important to study the role of pigs as reservoirs of STEC and the transmission to the swine production chain (Ercoli et al., 2015). Taking into account the data mentioned above, the aim of this study was to determine the prevalence and to characterize STEC throughout the pork production chain in Argentina.
Materials and Methods
Management of Farms and Animals
The study was conducted in two pig production farm systems. Both farms are intensively organized in total confinement. The production stages are: gestation, farrowing, weaning, and growing/finishing (fattening), which are geographically separated from each other within the same farm. The usual group size varies between 10 and 30 pigs. Pigs and employees move from one building to others by corridors that are isolated from external traffic.
Management of Carcasses Until Retails Markets
Pigs at finishing stage are transported to the slaughterhouses. After slaughtered, the pork carcasses are chilled during 24 to 48 h and sent to boning rooms in refrigerated trucks.
At the boning rooms the carcasses were boning to products such as meat and minced meat. Finally, the products are transferred to the retails markets.
Collection of Samples
Seven hundred and sixty four samples were collected from May, 2012 to November, 2014 from two pig production systems.
This study was carried out in accordance with the recommendations of the Animal Welfare Committee from the Veterinary Science Faculty, UNCPBA, Resolution 087/02.
Samples at Farms
Three hundred and forty eight samples were taken at farms. From these, 277 corresponded to rectal swabs, and 71 come from the environment obtained from water drink, feed and feces on the floor by swabbing.
Samples at Slaughterhouses
One hundred and forty seven samples were by swabbing. Off these, 22 were from rectal swabs after slaughter, 85 from carcasses, and 40 from the slaughterhouses environment (pre-washing, scalding, deharing, dressing, cooling, and knives).
Carcasses swabs were taken in concordance with circular No 3496/02 of Servicio Nacional de Sanidad y Calidad Agroalimentaria (SENASA, 2002). Five quarters areas of 100 cm2 each one were taken and processed separately, they are named heads (H), external rectum (ER), internal rectum (IR), external thoracic (ET), and internal thoracic (IT; Figure 1).
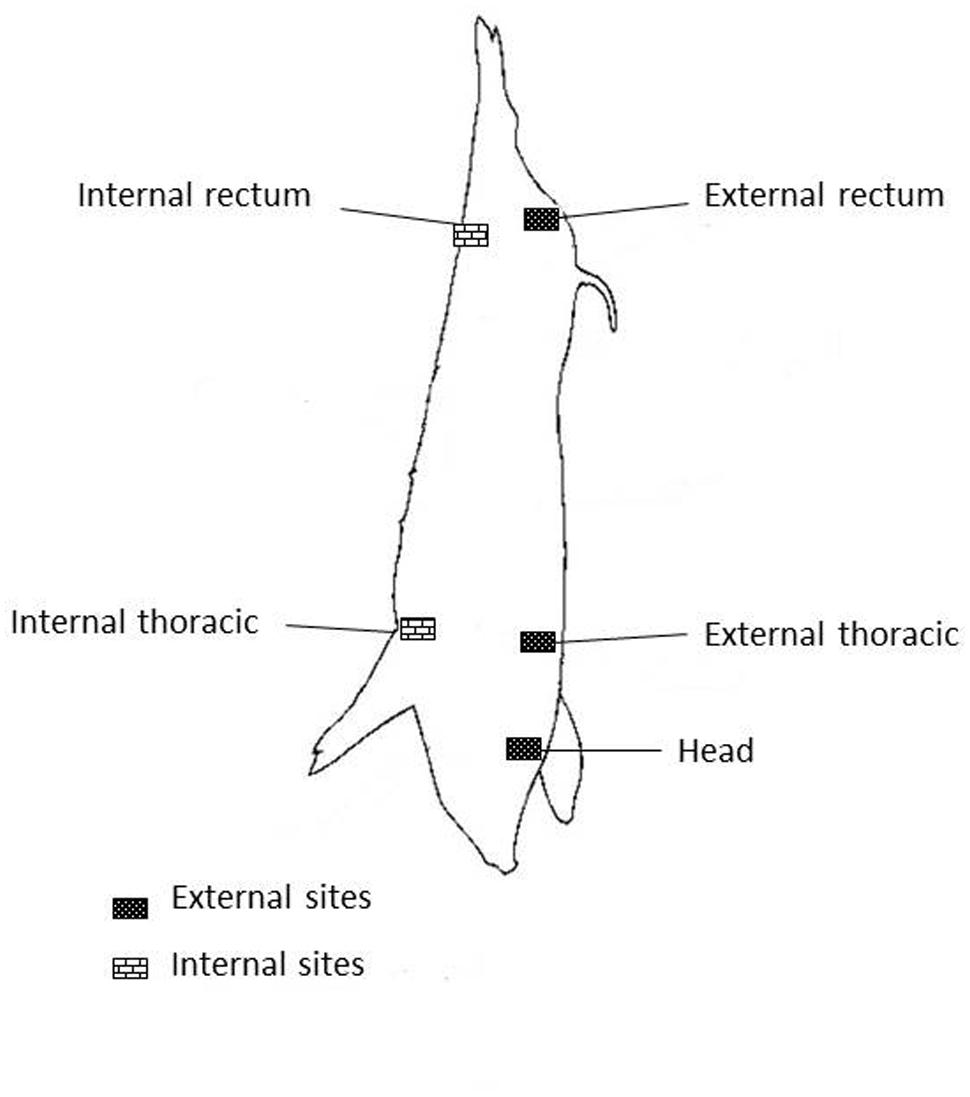
FIGURE 1. Sites on pig carcass.
Samples at Boning Rooms
One hundred and eighty one samples were taken. From these, 94 come from carcasses, 24 from meat, 23 from minced meat, and 40 from environmental samples (refrigerated trucks and meat contact surfaces such as meat tables, knives, meat mincing machine, and vertical band saw machine).
Sampling at Retail Markets
Eighty seven samples were taken from retail markets (43 samples come from meat, 13 from minced meat, and 31 from the environment). The environmental samples were obtained from meat tables, knives, vertical band saw machine, and refrigerators.
Sample Preparation and Isolation of STEC
Swabs were processed according to Etcheverría et al. (2010). Briefly, the swabs were cultured in Luria Bertani broth (LB) with shaking at 37°C for 18 h, and then an aliquot was grown on MacConkey agar plates by incubating at 37°C for 24 h.
Ten to fifty individual colonies were processed for amplification of Shiga toxin genes (stx1, stx2, and stx2e; Table 1). Each positive colony for either stx was tested for the presence of the eae, ehxA, and saa by multiplex polymerase chain reaction (PCR; Paton and Paton, 2002). Genes encoding adhesins (ehaA, agn43, iha, aida) were amplified using monoplex PCR. STEC strains used as positive control were E. coli O157:H7 EDL933 (stx1, stx2, eae, ehxA, ehaA, agn43, iha), E. coli O8 (stx2e), E. coli O91:H21 (stx1, stx2, ehxA, saa), and E. coli O157:H19 (aida).

TABLE 1. Genes, primers sequences, and size of amplified product of Shiga toxin-producing Escherichia coli (STEC).
Amplification products were separated by electrophoresis on 2% agarose gel containing 0.8 μg/ml of ethidium bromide in running buffer and were visualized by UV transillumination.
Determination of Serotype
O and H types were determined by microagglutination technique in plates and tubes as described by Guinée et al. (1981) and modified by Blanco et al. (1996) using all available O (O1–O175) antisera plus six putative new O antigens (OX176 through OX181) and H (H1–H56) antisera (Pradel et al., 2000).
Results
The results indicate that STEC occurrence is widespread throughout pork production chain. Among the 764 samples, 31 (4.05%) were positive for stx. In rectal swabs from the different pig categories, 2.86% were STEC positive, distributed 5.88% at fattening, 4.3% at growing, 2.38% at gestation and 1.51% at farrowing. STEC were not detected in feed, water, and fecal samples taken from farms. At slaughter, 4.08% of carcasses sampled were stx positive. The distribution in the different quarters of the carcasses was: 50% from ER, 16.6% from ET, 16.6% from IT, 16.6 % from heads. At boning rooms, 6% of samples were STEC positive, belonging 82% to carcasses, and 18% to pork meat. The distribution in the different quarters of the carcasses was: 33.3% from ER, 22.2% from IR, 22.2% from IT, 11.2% from ET and 11% from head. At sale markets 4.59% of STEC positive samples were detected in pork meat ready for sale.
Characterization of STEC
From positive samples, 50 isolates could be characterized by PCR. In samples from farms 6/16 (37.5%) of the isolates carried stx1/stx2, 6/16 (37.5%) possessed stx2e, and 4/16 (25%) carried stx2. At slaughter 3/6 (50%) of isolates were positive for stx2, 2/6 (33%) for stx2e, and 1/6 (16%) for stx1/stx2. At boning rooms 13/22 (59%) of the isolates carried stx1/stx2, 3/22 (14%) stx2e, and 1/22 (5%) stx1/stx2/stx2e. At retail markets 4/6 (66%) of isolates were positive for stx2, 1/6 (17%) for stx2e, and 1/6 (17%) for stx1/stx2. Other virulence factors such as ehxA and saa were not detected and eae was detected in 6/50 (12%) of samples. Concerning putatives adhesins, agn43 was detected in 36/50 (72%), ehaA in 13/50 (26%), aida in 4/50 (8%), and iha in 3/50 (6%) of isolates. The most frequent virulence profiles found were stx1, stx2, or stx2e combined with agn43 in 36 (72%) strains. The 50 isolates were typed into 14 E. coli O groups (O1, O2, O8, O15, O20, O35, O69, O78, O91, O121, O138, O142, O157, O180) and 15 were considered O non-typable (NT). Ten H antigens (H9, H10, H16, H21, H26, H29, H30, H32, H45, H46) were distributed among the 50 strains, while one isolate were non-motile (H–). Table 2 indicates the relationships between virulence profiles, sites of samples and serotypes in isolated STEC strains.
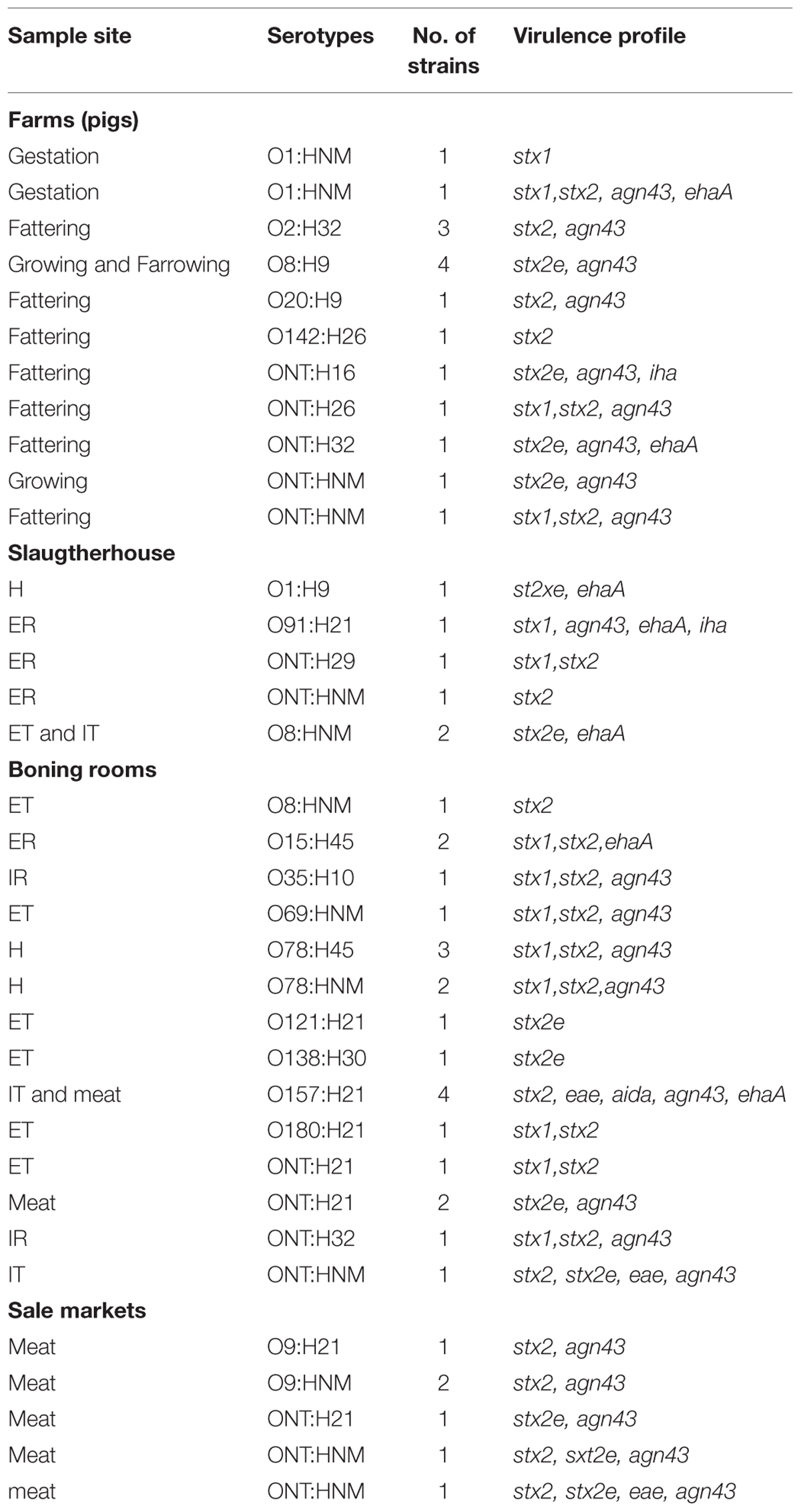
TABLE 2. Relationships between virulence profiles, sites of samples, and serotypes in STEC strains.
Discussion
To our knowledge, this study is the first that reports the prevalence and characterization of STEC strains through the chain pork suggesting the vertical transmission of these pathogens. However, there are studies that demonstrate the prevalence in farms, finishing pigs, slaughter, and pork meat in sales markets, separately. The prevalence of STEC in pigs, carcasses, and pork meat at different stage of production from other countries is variable and it is necessary to take caution when comparing prevalence since the variation may be due to several factors, such as sampling method, samples processing, and season in which the study was performed. Different prevalence of STEC in pigs was reported previously, ranged from 2 to 31%, in agreement with our results (Parma et al., 2000; Kaufmann et al., 2006; Meng et al., 2014). In carcasses at slaughter and boning rooms, the prevalence found is in concordance with other studies whose prevalence ranged from 0.2 to 26% (Leung et al., 2001; Bouvet et al., 2002; Bohaychuk et al., 2011; Koláčková et al., 2014).
At slaughter some operations such as skinning, evisceration and handling are more likely than others to contaminate carcass and meat (Koohmaraie et al., 2005; Etcheverría et al., 2010). For this, some areas of carcasses are more prone than others to be exposure to potential or cross contamination, thus the suggestion of sampling at three or four sites on carcass, because contamination appears to vary considerably among different sites (Roberts et al., 1984). In addition, the ER is the area that involves a particular risk of contamination during early stages of dressing as our result in that the ER was the more contaminated area, in concordance with Bouvet et al. (2002). However, other areas sampled as IT and IR shown more contamination probably due to handling at the boning room.
In our study, the prevalence at sale market was less than that informed by Magwedere et al. (2013) in USA (50%), Martin and Beutin (2011) in Germany (14%) and Lee et al. (2009) Korea (15%). This could be due because these studies were performed in sale markets where meat from different origins were sold and cross contamination during handling can occur. In the present study the samples were obtained from sale markets where only meat pork was sold.
From 50 STEC isolates, stx1/stx2 and stx2 occurred more frequently than isolates carrying stx1. Epidemiologically, Stx2-producing strains are more often related with HUS than strains that produce Stx1 (Paton and Paton, 2002).
Regarding stx2e, its prevalence decreased from pigs at farms to pork meat. Although some authors have reported the presence of stx2e in STEC strains in human patients on a few occasions, STEC harboring stx2e are more likely to cause edema disease in pigs causing economic losses in pig production (Kaufmann et al., 2006). It is necessary to determine a rol of these strains in human infection.
The presence of eae detected in isolates that harbor too stx2, stx2e, agn43 from boning rooms and sale markets implies a high risk for human health (Tseng et al., 2014). The most prevalent adhesin identified among all isolates and involved in adhesion and biofilm formation was Agn43, followed by EhaA in agreement with Biscola et al. (2011) and Tseng et al. (2014) which detected them in swine and different sources, respectively. In this study, 8% of strain harbored AIDA, similar to that found in South Africa (Mohlatlole et al., 2013) and China (Zhao et al., 2009). The iha was present in few isolates, but this gene has been detected over 70% of the eae-negative STEC strains associated with human clinical cases examined in studies in Germany (Hauser et al., 2013) and Argentina (Galli et al., 2010). The high prevalence of LEE negative STEC isolated from pigs in our study emphasizes the need of further work to better define the role that the attachment proteins outside the LEE may play in the adherence to both pork and human epithelial cells.
Although many serotypes isolated in this study have been detected with low incidence in human disease and rarely associated with outbreaks (Friedrich et al., 2002), they have been isolated from pigs, sheep, cattle and food in other countries (Beutin et al., 1993; Kaufmann et al., 2006; Beutin et al., 2007). At slaughter and boning rooms serogroups associated with human illnesses such as E. coli O91, O121, and O157 were detected in agreement with other studies that recovered these serogroups from pig fecal samples (Desrosiers et al., 2001; Friedrich et al., 2002; Karmali et al., 2003; Bielaszewska et al., 2009; Trotz-Williams et al., 2012; Yoon et al., 2013; Tseng et al., 2014).
Conclusion
The presented study investigated the tracking of STEC from the farm to table and indicates that the production of meat pork harbored STEC strains. STEC contamination originated in the farms is transferred from pigs to carcasses in the slaughter process and increase in meat pork at boning rooms and sales markets. Besides, the entrance of these strains into the food chain implies a risk to consumers because of severity of the illness they can cause. If STEC is present in any food product, it has the possibility of causing foodborne illness. In addition to public health problem, the presence of strains carrying the stx2e gene is a problem for the pig production because they can cause the edema disease causing important economic losses. In spite of the wealth of data available on this important disease, it is necessary to effectively prevent this contamination by educating employees, retailers and consumers on the appropriate handling and storage of meat. Further studies are needed to provide more systematic data in order to fuel the development of novel approaches for control of STEC in foods, including pork meat.
Author Contributions
RC conceived, designed, analyzed the experiments, and wrote the manuscript. MR, MS, and MC did some of the experiments. NP and AE designed some of the experiments, analyzed the data, and revised the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
Authors thank Guillermo Arroyo for their collaboration during sampling collection, María Rosa Ortiz for her technical assistance. This work was supported by PICT 2010-1655, CIC, and SECAT from Argentina.
References
Amézquita-López, B. A., Quiñones, B., Lee, B. G., and Chaidez, C. (2014). Virulence profiling of Shiga toxin-producing Escherichia coli recovered from domestic farm animals in Northwestern Mexico. Front. Cell. Infect. Microbiol. 4:7. doi: 10.3389/fcimb.2014.00007
Beutin, L., Geier, D., Steinrück, H., Zimmermann, S., and Scheutz, F. (1993). Prevalence and some properties of verotoxin (Shiga-like toxin)-producing Escherichia coli in seven different species of healthy domestic animals. J. Clin. Microbiol. 31, 2483–2488.
Beutin, L., Miko, A., Krause, G., Pries, K., Haby, S., Steege, K., et al. (2007). Identification of human-pathogenic strains of Shiga toxin-producing Escherichia coli from food by a combination of serotyping and molecular typing of Shiga toxin genes. Appl. Environ. Microbiol. 73, 4769–4775. doi: 10.1128/AEM.00873-07
Bielaszewska, M., Stoewe, F., Fruth, A., Zhang, W., Prager, R., Brockmeyer, J., et al. (2009). Shiga toxin, cytolethal distending toxin, and hemolysin repertoires in clinical Escherichia coli O91 isolates. J. Clin. Microbiol. 47, 2061–2066. doi: 10.1128/JCM.00201-09
Biscola, F. T., Abe, C. M., and Guth, B. E. C. (2011). Determination of adhesin gene sequences in, and biofilm formation by, O157 and non-O157 Shiga toxin-producing Escherichia coli strains isolated from different sources. Appl. Environ. Microbiol. 77, 2201–2208. doi: 10.1128/AEM.01920-10
Blanco, J. E., Blanco, M., Blanco, J., Mora, A., Balaguer, L., Mourino, M., et al. (1996). O serogroups, biotypes, and eae genes in Escherichia coli strains isolated from diarrheic and healthy rabbits. J. Clin. Microbiol. 34, 3101–3107.
Bohaychuk, V. M., Gensler, G. E., and Barrios, P. R. (2011). Microbiological baseline study of beef and pork carcasses from provincially inspected abattoirs in Alberta. Canada. Can. Vet. J. 52, 1095.
Bouvet, J., Montet, M., Rossel, R., Le Roux, A., Bavai, C., Ray-Gueniot, S., et al. (2002). Prevalence of verotoxin-producing Escherichia coli (VTEC) and E. coli O157: H7 in French pork. J. Appl. Microbiol. 93, 7–14. doi: 10.1046/j.1365-2672.2002.01672.x
Desrosiers, A., Fairbrother, J. M., Johnson, R. P., Desautels, C., Letellier, A., and Quessy, S. (2001). Phenotypic and genotypic characterization of Escherichia coli verotoxin-producing isolates from humans and pigs. J. Food Prot. 64, 1904–1911.
Ercoli, L., Farneti, S., Ranucci, D., Scuota, S., and Branciari, R. (2015). Role of verocytotoxigenic Escherichia coli in the swine production chain. Ital. J. Food Saf. 4, 51–56. doi: 10.4081/ijfs.2015.5156
Etcheverría, A., Padola, N., Sanz, M., Polifroni, R., Krüger, A., Passucci, J., et al. (2010). Occurrence of Shiga toxin-producing E. coli (STEC) on carcasses and retail beef cuts in the marketing chain of beef in Argentina. Meat Sci. 86, 418–421. doi: 10.1016/j.meatsci.2010.05.027
Ethelberg, S., Olsen, K., Scheutz, F., Jensen, C., Schiellerup, P., Enberg, J., et al. (2004). Virulence factors for hemolytic uremic syndrome, Denmark. Emerg. Infect. Dis. 10, 842–847. doi: 10.3201/eid1005.030576
Feng, P. C., and Reddy, S. (2013). Prevalences of Shiga toxin subtypes and selected other virulence factors among Shiga-toxigenic Escherichia coli strains isolated from fresh produce. Appl. Environ. Microbiol. 79, 6917–6923. doi: 10.1128/AEM.02455-13
Food and Agriculture Organization [FAO] (2014). Pigs and Animal Production [Online]. Available at: http://www.fao.org/ag/againfo/themes/en/pigs/production.html [Accessed December 10, 2014]
Friedrich, A. W., Bielaszewska, M., Zhang, W.-L., Pulz, M., Kuczius, T., Ammon, A., et al. (2002). Escherichia coli harboring Shiga toxin 2 gene variants: frequency and association with clinical symptoms. J. Infect. Dis. 185, 74–84. doi: 10.1086/338115
Galli, L., Miliwebsky, E., Irino, K., Leotta, G., and Rivas, M. (2010). Virulence profile comparison between LEE-negative Shiga toxin-producing Escherichia coli (STEC) strains isolated from cattle and humans. Vet. Microbiol. 143, 307–313. doi: 10.1016/j.vetmic.2009.11.028
Grisaru, S. (2014). Management of hemolytic-uremic syndrome in children. Int. J. Nephrol. Renovasc. Dis. 7, 231–239. doi: 10.2147/IJNRD.S41837
Guinée, P., Jansen, W., Wadström, T., and Sellwood, R. (1981). “Escherichia coli associated with neonatal diarrhoea in piglets and calves,” in Laboratory Diagnosis in Neonatal Calf and Pig Diarrhoea, Vol. 13, eds P. W. de Leeuw and P. A. M. Guinée (Dordrecht: Springer), 126–162.
Hauser, E., Mellmann, A., Semmler, T., Stoeber, H., Wieler, L. H., Karch, H., et al. (2013). Phylogenetic and molecular analysis of food-borne shiga toxin-producing Escherichia coli. Appl. Environ. Microbiol. 79, 2731–2740. doi: 10.1128/AEM.03552-12
Karmali, M. A., Mascarenhas, M., Shen, S., Ziebell, K., Johnson, S., Reid-Smith, R., et al. (2003). Association of genomic O island 122 of Escherichia coli EDL 933 with verocytotoxin-producing Escherichia coli seropathotypes that are linked to epidemic and/or serious disease. J. Clin. Microbiol. 41, 4930–4940. doi: 10.1128/JCM.41.11.4930-4940.2003
Kaufmann, M., Zweifel, C., Blanco, M., Blanco, J., Blanco, J., Beutin, L., et al. (2006). Escherichia coli O157 and non-O157 Shiga toxin–producing Escherichia coli in fecal samples of finished pigs at slaughter in Switzerland. J. Food Prot. 69, 260–266. doi: 10.4315/0362-028X.JFP-10-579
Koláčková, I., Házová, K., Skočková, A., and Karpíšková, R. (2014). [Occurrence of Shiga toxigenic Escherichia coli strains in pigs and cattle at slaughterhouses in the Czech Republic in 2013.]. Klin. Mikrobiol. Infekc. Lek. 20, 36–39.
Koohmaraie, M., Arthur, T., Bosilevac, J., Guerini, M., Shackelford, S., and Wheeler, T. (2005). Post-harvest interventions to reduce/eliminate pathogens in beef. Meat Sci. 71, 79–91. doi: 10.1016/j.meatsci.2005.03.012
Lee, G. Y., Jang, H. I., Hwang, I. G., and Rhee, M. S. (2009). Prevalence and classification of pathogenic Escherichia coli isolated from fresh beef, poultry, and pork in Korea. Int. J. Food Microbiol. 134, 196–200. doi: 10.1016/j.ijfoodmicro.2009.06.013
Leung, P., Yam, W., Ng, W., and Peiris, J. (2001). The prevalence and characterization of verotoxin-producing Escherichia coli isolated from cattle and pigs in an abattoir in Hong Kong. Epidemiol. Infect. 126, 173–179. doi: 10.1017/S0950268801005210
Magwedere, K., Dang, H. A., Mills, E. W., Cutter, C. N., Roberts, E. L., and Debroy, C. (2013). Incidence of Shiga toxin–producing Escherichia coli strains in beef, pork, chicken, deer, boar, bison, and rabbit retail meat. J. Vet. Diagn. Invest. 25, 254–258. doi: 10.1177/1040638713477407
Martin, A., and Beutin, L. (2011). Characteristics of Shiga toxin-producing Escherichia coli from meat and milk products of different origins and association with food producing animals as main contamination sources. Int. J. Food Microbiol. 146, 99–104. doi: 10.1016/j.ijfoodmicro.2011.01.041
Meng, Q., Bai, X., Zhao, A., Lan, R., Du, H., Wang, T., et al. (2014). Characterization of Shiga toxin-producing Escherichia coli isolated from healthy pigs in China. BMC Microbiol. 14:5. doi: 10.1186/1471-2180-14-5
Mohlatlole, R. P., Madoroba, E., Muchadeyi, F. C., Chimonyo, M., Kanengoni, A. T., and Dzomba, E. F. (2013). Virulence profiles of enterotoxigenic, shiga toxin and enteroaggregative Escherichia coli in South African pigs. Trop. Anim. Health Prod. 45, 1399–1405. doi: 10.1007/s11250-013-0377-4
Niewerth, U., Frey, A., Voss, T., Le Bouguénec, C., Baljer, G., Franke, S., et al. (2001). The Aida autotransporter system is associated with F18 and stx2e in Escherichia coli isolates from pigs diagnosed with edema disease and postweaning diarrhea. Clin. Diagn. Lab. Immunol. 8, 143–149.
Padola, N. L., Sanz, M. E., Blanco, J. E., Blanco, M., Blanco, J., Etcheverria, A. A. I., et al. (2004). Serotypes and virulence genes of bovine Shigatoxigenic Escherichia coli (STEC) isolated from a feedlot in Argentina. Vet. Microbiol. 100, 3–9. doi: 10.1016/S0378-1135(03)00127-5
Parma, A., Sanz, M., Blanco, J., Blanco, J., Viñas, M., Blanco, M., et al. (2000). Virulence genotypes and serotypes of verotoxigenic Escherichia coli isolated from cattle and foods in Argentina. Eur. J. Epidemiol. 16, 757–762. doi: 10.1023/A:1026746016896
Paton, A. W., and Paton, J. C. (2002). Direct detection and characterization of Shiga toxigenic Escherichia coli by multiplex PCR for stx1, stx2, eae, ehxA, and saa. J. Clin. Microbiol. 40, 271–274. doi: 10.1128/JCM.40.1.271-274.2002
Paton, A. W., Srimanote, P., Woodrow, M. C., and Paton, J. C. (2001). Characterization of Saa, a novel autoagglutinating adhesin produced by locus of enterocyte effacement-negative Shiga-toxigenic Escherichia coli strains that are virulent for humans. Infect. Immun. 69, 6999–7009. doi: 10.1128/IAI.69.11.6999-7009.2001
Paton, J. C., and Paton, A. W. (1998). Pathogenesis and diagnosis of Shiga toxin-producing Escherichia coli infections. Clin. Microbiol. Rev. 11, 450–479.
Pradel, N., Livrelli, V., De Champs, C., Palcoux, J.-B., Reynaud, A., Scheutz, F., et al. (2000). Prevalence and characterization of Shiga toxin-producing Escherichia coli isolated from cattle, food, and children during a one-year prospective study in France. J. Clin. Microbiol. 38, 1023–1031.
Restieri, C., Garriss, G., Locas, M.-C., and Dozois, C. M. (2007). Autotransporter-encoding sequences are phylogenetically distributed among Escherichia coli clinical isolates and reference strains. Appl. Environ. Microbiol. 73, 1553–1562. doi: 10.1128/AEM.01542-06
Rivas, M., Padola, N. L., Lucchesi, P. M., Masana, M., and Torres, A. (2010). “Diarrheagenic Escherichia coli in Argentina,” in Pathogenic Escherichia coli in Latin America, ed. A. G. Torres (Oak Park, IL: Bentham Science Publishers), 142–161.
Roberts, T., Hudson, W., Whelehan, O., Simonsen, B., Olgaard, K., Labots, H., et al. (1984). Number and distribution of bacteria on some beef carcasses at selected abattoirs in some member states of the European Communities. Meat Sci. 11, 191–205. doi: 10.1016/0309-1740(84)90037-8
SENASA (2002). “Circular 3496/02,” in Animal. Argentina: Servicio Nacional de Sanidad y Calidad Agroalimentaria.
Sonntag, A.-K., Bielaszewska, M., Mellmann, A., Dierksen, N., Schierack, P., Wieler, L. H., et al. (2005). Shiga toxin 2e-producing Escherichia coli isolates from humans and pigs differ in their virulence profiles and interactions with intestinal epithelial cells. Appl. Environ. Microbiol. 71, 8855–8863. doi: 10.1128/AEM.71.12.8855-8863.2005
Szalo, I. M., Goffaux, F., Pirson, V., Piérard, D., Ball, H., and Mainil, J. (2002). Presence in bovine enteropathogenic (EPEC) and enterohaemorrhagic (EHEC) Escherichia coli of genes encoding for putative adhesins of human EHEC strains. Res. Microbiol. 153, 653–658. doi: 10.1016/S0923-2508(02)01379-7
Tarr, P. I., Bilge, S. S., Vary, J. C., Jelacic, S., Habeeb, R. L., Ward, T. R., et al. (2000). Iha: a novel Escherichia coli O157: H7 adherence-conferring molecule encoded on a recently acquired chromosomal island of conserved structure. Infect. Immun. 68, 1400–1407. doi: 10.1128/IAI.68.3.1400-1407.2000
Trotz-Williams, L. A., Mercer, N. J., Walters, J. M., Maki, A. M., and Johnson, R. P. (2012). Pork implicated in a Shiga toxin-producing Escherichia coli O157: H7 outbreak in Ontario, Canada. Can. J. Public Health 103, e322–e326.
Tseng, M., Fratamico, P. M., Bagi, L., Delannoy, S., Fach, P., Manning, S. D., et al. (2014). Diverse virulence gene content of Shiga toxin-producing Escherichia coli from finishing swine. Appl. Environ. Microbiol. 80, 6395–6402. doi: 10.1128/AEM.01761-14
Wells, T. J., Sherlock, O., Rivas, L., Mahajan, A., Beatson, S. A., Torpdahl, M., et al. (2008). EhaA is a novel autotransporter protein of enterohemorrhagic Escherichia coli O157: H7 that contributes to adhesion and biofilm formation. Environ. Microbiol. 10, 589–604. doi: 10.1111/j.1462-2920.2007.01479.x
Woodward, M., Carroll, P., and Wray, C. (1992). Detection of entero-and verocyto-toxin genes in Escherichia coli from diarrhoeal disease in animals using the polymerase chain reaction. Vet. Microbiol. 31, 251–261. doi: 10.1016/0378-1135(92)90083-6
Wu, Y., Hinenoya, A., Taguchi, T., Nagita, A., Shima, K., Tsukamoto, T., et al. (2010). Distribution of virulence genes related to adhesions and toxins in Shiga toxin-producing Escherichia coli strains isolated from healthy cattle and diarrheal patients in Japan. J. Vet. Med. Sci. 72, 589–597. doi: 10.1292/jvms.09-0557
Yoon, S. C., Windham, W. R., Ladely, S., Heitschmidt, G. W., Lawrence, K. C., Park, B., et al. (2013). Differentiation of big-six non-O157 Shiga-toxin producing Escherichia coli (STEC) on spread plates of mixed cultures using hyperspectral imaging. J. Food Meas. Charact. 7, 47–59. doi: 10.4315/0362-028X.JFP-12-497
Keywords: STEC, foodborne pathogens, pork production chain, prevalence, characterization
Citation: Colello R, Cáceres ME, Ruiz MJ, Sanz M, Etcheverría AI and Padola NL (2016) From Farm to Table: Follow-Up of Shiga Toxin-Producing Escherichia coli Throughout the Pork Production Chain in Argentina. Front. Microbiol. 7:93. doi: 10.3389/fmicb.2016.00093
Received: 03 November 2015; Accepted: 18 January 2016;
Published: 08 February 2016.
Edited by:
Pina Fratamico, United States Department of Agriculture – Agricultural Research Service, USAReviewed by:
James L. Smith, United States Department of Agriculture, USAYanhong Liu, European Roma Rights Centre – United States Department of Agriculture – Agricultural Research Service, USA
Copyright © 2016 Colello, Cáceres, Ruiz, Sanz, Etcheverría and Padola. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Analía I. Etcheverría, YW5hbGlhaW5AdmV0LnVuaWNlbi5lZHUuYXI=