 Yuding Weng1
Yuding Weng1 Fei Chen1
Fei Chen1 Yiwei Liu1Qiang Zhao1
Yiwei Liu1Qiang Zhao1 Ronghao Chen1
Ronghao Chen1 Xiaolei Pan1
Xiaolei Pan1 Chang Liu1
Chang Liu1 Zhihui Cheng1
Zhihui Cheng1 Shouguang Jin1,2
Shouguang Jin1,2 Yongxin Jin1*
Yongxin Jin1* Weihui Wu1*
Weihui Wu1*- 1State Key Laboratory of Medicinal Chemical Biology, Key Laboratory of Molecular Microbiology and Technology of the Ministry of Education, Department of Microbiology, College of Life Sciences, Nankai University, Tianjin, China
- 2Department of Molecular Genetics and Microbiology, College of Medicine, University of Florida, Gainesville, FL, USA
Pseudomonas aeruginosa is a Gram negative opportunistic pathogenic bacterium, which causes acute and chronic infections. Upon entering the host, bacteria alter global gene expression to adapt to host environment and avoid clearance by the host. Enolase is a glycolytic enzyme involved in carbon metabolism. It is also a component of RNA degradosome, which is involved in RNA processing and gene regulation. Here, we report that enolase is required for the virulence of P. aeruginosa in a murine acute pneumonia model. Mutation of enolase coding gene (eno) increased bacterial susceptibility to neutrophil mediated killing, which is due to reduced tolerance to oxidative stress. Catalases and alkyl hydroperoxide reductases play a major role in protecting the cell from oxidative damages. In the eno mutant, the expression levels of catalases (KatA and KatB) were similar as those in the wild type strain in the presence of H2O2, however, the expression levels of alkyl hydroperoxide reductases (AhpB and AhpC) were significantly reduced. Overexpression of ahpB but not ahpC in the eno mutant fully restored the bacterial resistance to H2O2 as well as neutrophil mediated killing, and partially restored bacterial virulence in the murine acute pneumonia model. Therefore, we have identified a novel role of enolase in the virulence of P. aeruginosa.
Introduction
Pseudomonas aeruginosa is a ubiquitous Gram negative bacterium. It is a major cause of nosocomial infections, including severe burn infections, sepsis, urinary tract infection, and pneumonia. Of note, P. aeruginosa is the leading cause of chronic lung infection in cystic fibrosis (CF) patients and ventilator-associated pneumonia (VAP; Diaz et al., 2005; Mcbride, 2005).
During infection, P. aeruginosa orchestrates expression of multiple virulence factors to counteract host immune clearance and increase tolerance to antibiotics (Rahme et al., 2000; Poole, 2011; Balasubramanian et al., 2012; Morita et al., 2014). In a murine acute pneumonia model, it has been demonstrated that neutrophils are rapidly recruited to the lung in response to invading bacteria (Shaver and Hauser, 2004). Neutrophils are phagocytes that kill bacteria by generation of reactive oxygen species (ROS), phagocytosis, and degranulation (Ziltener et al., 2016).
Among the virulence factors of P. aeruginosa, the type III secretion system (T3SS) plays an essential role in killing phagocytes or inhibiting phagocytosis (Brannon et al., 2009; Plano and Schesser, 2013). The T3SS is a needle like structure conserved in various Gram negative animal and plant pathogenic bacteria, through which effector proteins are directly injected into host cell cytosol, altering cell signaling, or killing host cells (Luo and Jin, 2008; Bleves et al., 2010; Pha and Navarro, 2016). Four effector proteins, namely ExoU, ExoS, ExoT, and ExoY have been identified in P. aeruginosa (Hornef et al., 2000). Most clinic isolates express three of the four effectors, including ExoT, ExoY and either ExoU, or ExoS (Feltman et al., 2001; Shaver and Hauser, 2004). Injection of the ExoS or ExoU into phagocytes is critical for the pathogenesis of P. aeruginosa in a murine acute pneumonia model (Shaver and Hauser, 2004).
Meanwhile, P. aeruginosa expresses catalases (KatA and KatB) and alkyl hydroperoxide reductases (AhpB and AhpC) to defend against host produced ROS (Lee et al., 2005). Expression of these antioxidant genes is activated by a transcriptional regulator OxyR in response to oxidative stresses, such as H2O2 (Ochsner et al., 2000). OxyR contains two conserved cysteine residues, oxidation of which results in formation of an intramolecular disulfide bond, promoting the binding between OxyR and target promoters (Jo et al., 2015).
Multiple regulatory proteins and RNAs are involved in the regulation of virulence factors. The bacterial RNA degradosome, which is composed of polynucleotide phosphorylase (PNPase), enolase, RNA helicase (RhlB) and ribonuclease E (RNase E), plays an important role in RNA processing and gene regulation (Favaro and Deho, 2003; Burger et al., 2011; Matos et al., 2011; Saramago et al., 2014). Previously, we demonstrated that PNPase is required for the expression of T3SS genes and pathogenesis of P. aeruginosa in a murine acute pneumonia model (Chen et al., 2016). These results prompted us to explore the functions of other RNA degradosome components in bacterial pathogenesis. Enolase is another key component of RNA degradosome. It is highly conserved in bacteria (Canback et al., 2002). Studies on the Escherichia coli RNA degradosome revealed that enolase binds to a small region in the degradosome-scaffolding domain of RNase E (Chandran and Luisi, 2006). A crystal structure analysis suggested that enolase may facilitate the organization of a RNA-binding motif in RNase E (Nurmohamed et al., 2010). In E. coli, it has been shown that enolase, but not PNPase or RhlB, is required for the RNase E mediated degradation of the glucose transporter PtsG mRNA in response to metabolic stress (Morita et al., 2004). The function of enolase in P. aeruginosa is not well known. Here in this study, we found that enolase is essential for the virulence of P. aeruginosa in a murine acute pneumonia model. Instead of affecting T3SS gene expression, enolase is required for bacterial oxidative stress response. Thus, our results revealed a novel role of enolase in bacterial pathogenesis.
Materials and Methods
Bacterial Strains, Plasmids, and Growth Conditions
Strains and plasmid used in this study are listed in Table 1. For the construction of an eno deletion mutant, a 934-bp upstream fragment and a 1207-bp downstream fragment of the eno conding region were amplified by PCR with PAK chromosome as the template and primers shown in Supplementary Table S1. The fragments were cloned into the plasmid pEX18TC (Hoang et al., 1998). Deletion of the eno gene in P. aeruginosa was performed as previously described (Hoang et al., 1998). For the complementation of eno, the eno gene and its native promoter were amplified with primers shown in Supplementary Table S1. The fragments were ligated into pUC18T-mini-Tn7T-Gm. The plasmid was transferred into the eno mutant strain along with the helper plasmid pTNS3 by electroporation as previously described (Choi and Schweizer, 2006). The ahpB and ahpC coding regions were amplified with primers shown in Supplementary Table S1 and ligated into pUCP20, respectively. The plasmid was transferred into the eno mutant by electroporation. To construct the ahpB- and ahpC-lacZ transcriptional fusions, the promoter regions of ahpB and ahpC were amplified with primers shown in Supplementary Table S1 by PCR. The fragments were ligated into the vector pDN19lacZΩ (Li et al., 2013).

TABLE 1. Strains and plasmids.
All bacterial strains were cultured in Luria broth (LB, 1% Bacto-tryptone, 0.5% yeast extract, 1% NaCl; Oxoid Ltd, USA) at 37°C. Antibiotics were used at the following concentrations: for E. coli, kanamycin 50 μg/ml, gentamicin 15 μg/ml; for P. aeruginosa, carbenicillin 150 μg/ml, gentamicin 50 μg/ml, tetracycline 50 μg/ml. All antibiotics are from BBI Life Science, Shanghai, China.
β-Galactosidase Assay
β-Galactosidase assay was performed as previously described (Miller, 1972) with minor modifications. Briefly, bacteria were cultured overnight and diluted 1:100 in fresh LB medium and grown at 37°C with agitation. When the optical density at 600 nm (OD600) reached 2.0, 0.5 ml bacteria were collected by centrifugation and resuspended in 1.5 ml Z buffer (60 mM Na2HPO4, 60 mM NaH2PO4, 10 mM KCl, 1 mM MgSO4, 50 mM β-mercaptoethanol, pH 7.0; BBI Life Science, Shanghai, China). One milliliter of the suspension was allocated for OD600 measurement. The other 0.5 ml suspension was added with 10 μl chloroform (BBI Life Science, Shanghai, China) and 10 μl 0.1% SDS (BBI Life Science, Shanghai, China), followed by vortex for 10 s. Then 100 μl ONPG (40 mg/ml; Sigma, USA) was added to the mixture and incubated at 37°C. The reaction was stopped by addition of 0.5 ml 1M Na2CO3. The time was recorded and OD420 was measured. β-Galactosidase activity (Miller units) was calculated as (1000 × OD420)/(T × V × OD600). T, reaction time (minute); V, bacteria volume (ml).
Murine Acute Pneumonia Model
Infection of mouse was performed as previously described (Sun et al., 2014). Briefly, overnight bacterial culture was diluted 1:100 in fresh LB medium and grown at 37°C with agitation. When the optical density at 600 nm (OD600) reached 1.0, bacteria were collected and resuspended in phosphate-buffered saline (PBS) at a concentration of 1 × 109 CFU/ml. Six to eight weeks old female BALB/c mice (Vital River, Beijing, China) were anesthetized by the injection of 100 μl 7.5% chloral hydrate (Sigma, USA) intraperitoneally. Twenty microliter bacterial suspension was then inoculated intranasally to each mouse, resulting in 2 × 107 CFU bacteria per mouse. Twelve hours post-infection (hpi), the mice were sacrificed and lungs were isolated and homogenized in 1% proteose peptone (Sigma, USA), followed by determination of bacterial loads by serial dilution and plating. In the mortality assay, each mouse was infected with 4 × 107 CFU bacteria, and monitored for 6 days. The statistical analysis was performed with the Prism software (Version 6, Graphpad Software, La jolla, USA).
RNA Extraction and Real Time PCR (qRT-PCR)
Total RNA was isolated with the RNA prep Pure cell/Bacteria Kit (Tiangen Biotec, Beijing, China). Random primers and the Prime Script Reverse Transcriptase (Takara, Dalian, China) were used to synthesize cDNA. The cDNA was used as the template to detect the relative mRNA levels of indicated genes with specific primers and Fast Start Essential DNA Green Master (Roche, Switzerland). Gene PA1805 was used as the internal control (Son et al., 2007).
Histology
Twelve hours after infection with indicated P. aeruginosa strains, lungs of the mice were removed and fixed with 10% paraformaldehyde (Sigma, USA), then dehydrated with ethanol (Tian Jin chemical reagent company, Tianjin, China), and embedded in paraffin (BBI Life Science, Shanghai, China). The tissue sections were cut into slices and stained with hematoxylin and eosin (BBI Life Science, Shanghai, China). Images were taken with an Olympus microscope (Version IX71, Tokyo, Japan).
Cytotoxicity Assay
Bacterial cytotoxicity was determined by the lactate dehydrogenase (LDH) release assay. Briefly, HeLa cells (ATCC, USA) were cultured in Dulbecco’s modified Eagle medium (DMEM, Hyclone, USA) with and 2% (vol/vol) heat-inactivated fetal bovine serum (hiFBS, Gibco, Australia) at 37°C with 5% CO2. Eighteen hours before infection, 1.2 × 105 HeLa cells were seeded into each well of a 24-well plate. Bacteria were grown to an OD600 of 1.0, collected by centrifugation, then washed twice and resuspended in PBS. After addition of bacteria to each well, the plate was centrifuged at 1,700 g for 10 min to synchronize the infection. Three hours after the infection, LDH released from the dead cells was measured by the LDH cytotoxicity assay kit (Beyotime, Haimen, China). Cells treated with the cell lysis buffer provided by the kit were used as the control of 100% LDH release. The culture medium without cell was used to set the background LDH level. The cytotoxicity percentage was calculated following the manufacturer’s instruction.
Western Blotting
Over night bacterial culture was diluted 1:100 in LB or 1:50 in LB with 5 mM EGTA (BBI Life Science, Shanghai, China) and incubated at 37°C with agitation. After 4 h, the supernatant of each culture was collected by centrifugation. Supernatants collected from equal numbers of bacteria were loaded to a 12% sodium dodecyl sulfate-polyacrylamide gel (SDS-PAGE). Then the proteins were transferred to a polyvinylidene difluoride (PVDF, Millipore, USA) membrane, and probed with a rabbit polyclonal antibody against ExoS (Li et al., 2013) at room temperature for 1 h. The membrane was washed three times with PBS containing 0.2% tween-20 (Tian Jin chemical reagent company, Tianjin, China), followed by incubation with a horseradish peroxidase-conjugated goat anti-rabbit IgG (Millipore, USA) at room temperature for 1 h. The signal was detected with the ECL-plus kit (Millipore, USA).
Cell Culture and HL-60 Cell Differentiation
HL-60 cells (ATCC, USA) were cultured in RPMI 1640 medium (Hyclone, USA) with 10% (vol/vol) heat-inactivated fetal bovine serum (Gibco, Australia) and penicillin G (100 U/ml) and streptomycin (100 μg/ml; Hyclone, USA). The cells were cultured at 37°C with 5% CO2. Differentiation of the HL-60 cells was conducted as previously described (Chen and Seifert, 2011). Briefly, HL-60 cells were diluted to ∼4.5 × 105 cells/ml and 1.3% dimethylsulfoxide (Sigma, USA) was added to the medium. The cells were then cultured for 6–7 days before use.
Measurement of Reactive Oxygen Species (ROS) Levels
The ROS production levels were determined as previously described with slight modifications (Wu and Hsu, 2009). Briefly, differentiated HL-60 cells were washed once with warm Hank’s balanced salt solution (HBSS; Hyclone, USA) and diluted to 7.5 × 104 cells/ml in HBSS containing 100 μM luminol (Sigma, USA) and 5 units per ml horseradish peroxidase (Sigma, USA). Two hundreds microliter cell suspension was added to each well of a 96-well plate, followed by incubation at 37°C for 10 min. Then the cells were infected with wild-type PAK or the Δeno mutant at a multiplicity of infection (MOI) of 30. The ROS levels were measured every 3 min for 4 h with a Luminoskan Ascent Luminometer (Varioskan Flash, Thermo Scientific, USA).
Growth Inhibitory Effect of Differentiated HL-60 Cells
Bacteria were grown to an OD600 of 1.0, collected by centrifugation and washed three times with sterile PBS. Then 1 × 107 bacteria of each strain were incubated with 1 × 106 undifferentiated or differentiated HL-60 cells in 200 μl RPMI 1640 medium at 37°C. At indicated time points, the live bacterial numbers were determined by serial dilution and plating. The growth inhibitory rate of each strain was calculated by dividing the live bacterial number incubated with differentiated HL-60 cells by the live bacterial number incubated with undifferentiated HL-60 cells.
H2O2 Susceptibility Assay
Bacteria at an OD600 of 1.0 were collected and washed for three times with sterile PBS. Then the bacteria were diluted to 2 × 107 CFU/ml in PBS and incubated with or without 10 mM H2O2 at 37°C for 15 min. The live bacterial numbers were determined by serial dilution and plating. The survival rate was calculated by dividing the live bacterial number with H2O2 treatment by the live bacterial number without H2O2 treatment.
Ethical Statement
All animal experiments complied with Chinese national guidelines on the use of animals in research. The protocol was approved by the institutional animal care and use committee of the college of life sciences of Nankai University with a permit number: NK-04-2012.
Results
Enolase Is Required for P. aeruginosa Lung Colonization
To evaluate the role of enolase in P. aeruginosa pathogenesis, we utilized a murine acute pneumonia model as previously described (Sun et al., 2014). Six weeks old female BALB/c mice were infected intranasally with 2 × 107 wild type PAK or an enolase deletion mutant (Δeno). Twelve hours post-infection, lungs were isolated and homogenized. Bacterial loads were determined by serial dilution and plating. Compared to the wild type strain, the number of the Δeno mutant was significantly lower (Figure 1A). For the complementation, an eno gene driven by its native promoter was cloned into pUC18T-mini-Tn7T-Gm and inserted into the chromosome (Choi and Schweizer, 2006). As shown in Figure 1A, complementation with an eno gene fully restored the bacterial load in the lung, indicating a role of enolase in bacterial growth in the lung. When, we grew the bacteria in LB, we noticed that the Δeno mutant grows more slowly than wild type PAK (Supplementary Figure S1A). After 12 h in vitro growth, the bacterial number of the Δeno mutant was ∼70% of that of the wild type PAK or the complemented strain (Supplementary Figure S1B). Given that there was ∼104-fold difference in the bacterial load between wild type PAK and the Δeno mutant infected mice, it is likely that factors other than slow growth contribute to the reduced bacterial number in vivo. To examine the role of enolase in virulence, we monitored the mortality rate in the acute pneumonia model. Infection with wild type PAK or the complemented strain resulted in 82.5% mortality rate, whereas no mouse died after infection with the Δeno mutant (Figure 1B). Furthermore, lungs from mice at 12 hpi were subjected to H&E staining. Infection with wild type PAK resulted in severe occlusion with neutrophil infiltration, which was significantly milder in the Δeno mutant infected lungs (Figure 1C). Consistently, lower mRNA levels of inflammatory cytokines, including IL-1β, IL-6, and TNF-α were detected in the lungs infected with the Δeno mutant compared to those in the wild type PAK infected lungs (Figure 1D). Therefore, enolase is required for bacterial virulence in the acute pneumonia model.
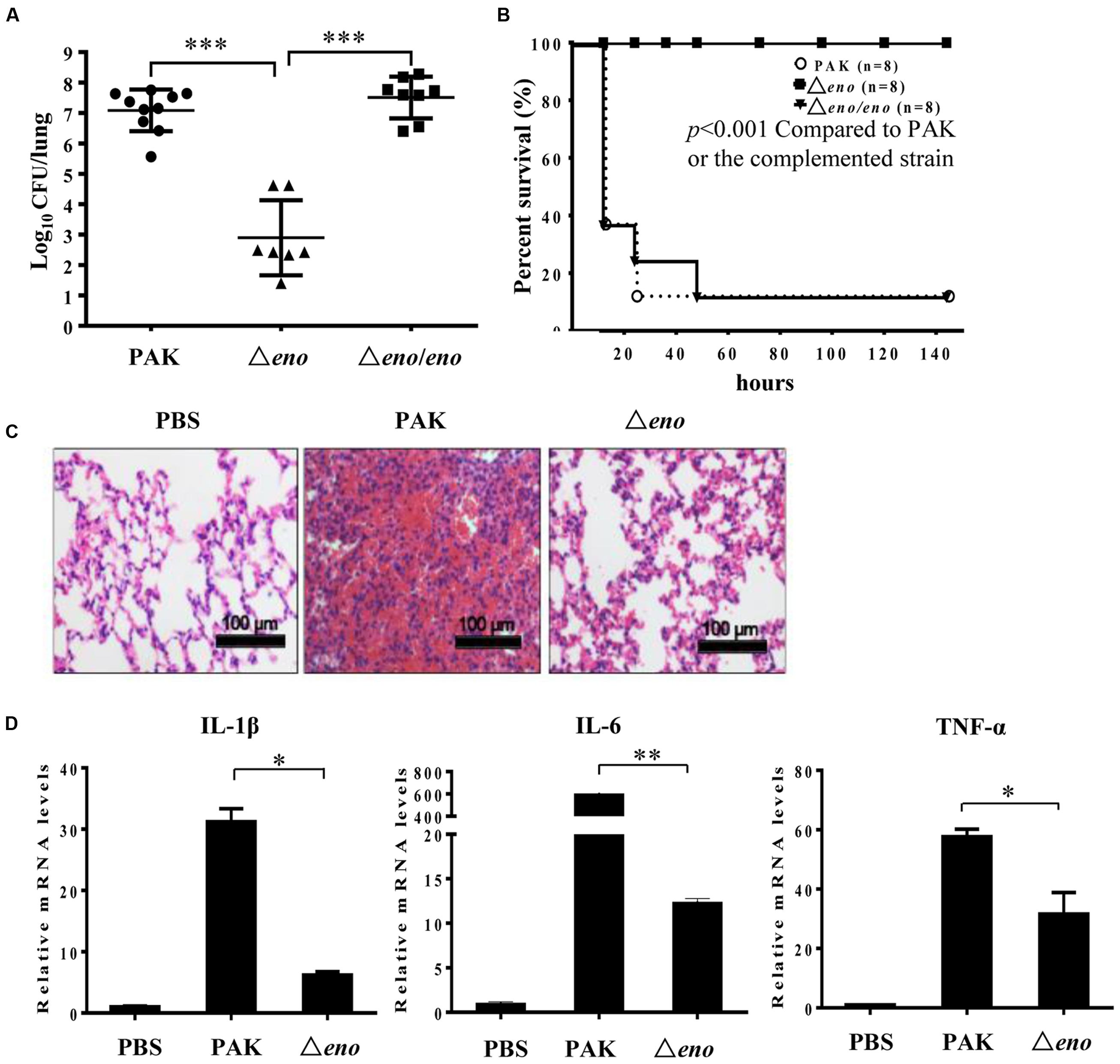
FIGURE 1. Role of enolase in bacterial pathogenesis in a murine acute pneumonia model. (A) Mice were infected with 2 × 107 wild type PAK or the Δeno mutant or the complemented strain intranasally. At 12 hpi, lungs from mice infected with no bacteria (PBS), or the indicated strains were recovered. Bacterial loads were determined by serial dilution and plating. ∗∗∗P < 0.001 by the Mann–Whitney test. (B) Each mouse was infected with 4 × 107 bacteria intranasally. Survival of the mice was monitored at least twice per day for 6 days. The P-value was calculated by Kaplan–Meier survival analysis with a log rank test with the Prism software. (C) Each mouse was infected with 2 × 107 bacteria intranasally. At 12 hpi, the lungs were fixed with paraformaldehyde (PFA), sectioned, and stained with hematoxylin and eosin. Images were taken with a 20× objective lens. (D) Total RNA was isolated from lungs of the infected mice. mRNA levels of IL1-β, IL-6, and TNF-α were determined by qRT-PCR. Bars represent means, and error bars represent standard deviation (SD). A representative of three independent experiments with similar results is shown. ∗P < 0.05; ∗∗P < 0.01 by student’s t-test.
Mutation of eno Increases Bacterial Susceptibility to Oxidative Stresses
In the mouse acute pneumonia model, neutrophils are rapidly recruited to the lung after infection and play a major role in the defense against bacteria (Wu et al., 2012; Ziltener et al., 2016). Induction and delivery of T3SS effector into neutrophils inhibit the bactericidal effects of those cells, enabling the bacterial colonization and dissemination (Diaz and Hauser, 2010; Howell et al., 2012; Rangel et al., 2015). Previously, we found that PNPase is required for the expression of the T3SS genes in the mouse acute pneumonia model (Chen et al., 2016). Since both enolase and PNPase are components of the RNA degradosome, they may share common regulatory targets. Thus, we examined the effect of eno mutation on the activity of the T3SS. Surprisingly, the expression and secretion of ExoS were similar between the Δeno mutant and wild type PAK upon growth in calcium depleted LB medium, which is a typical in vitro T3SS inducing condition (Figure 2A), and the bacterial cytotoxicity were similar between wild type PAK and the Δeno mutant (Figure 2B). We further examined the expression levels of T3SS genes during infection. Bacteria were isolated from bronchoalveolar lavage fluid (BALF) of infected mice. The mRNA levels of T3SS genes exsC and pcrV were determined by qRT-PCR with previously reported PA1805, PA1769, rpsL, and the 16S rRNA PA0668.1 as internal controls for normalization (Savli et al., 2003; Ruzin et al., 2007; Son et al., 2007; Sun et al., 2014). Similar mRNA fold of changes (within 1.2-fold difference) were observed between these internal controls. Therefore, we used the PA1805 as the internal control in this study. As shown in Figure 2C, the expression levels of exsC and pcrV were similar between wild type PAK and the Δeno mutant. In combination, these results suggest that mutation of the eno does not affect the expression of T3SS genes.
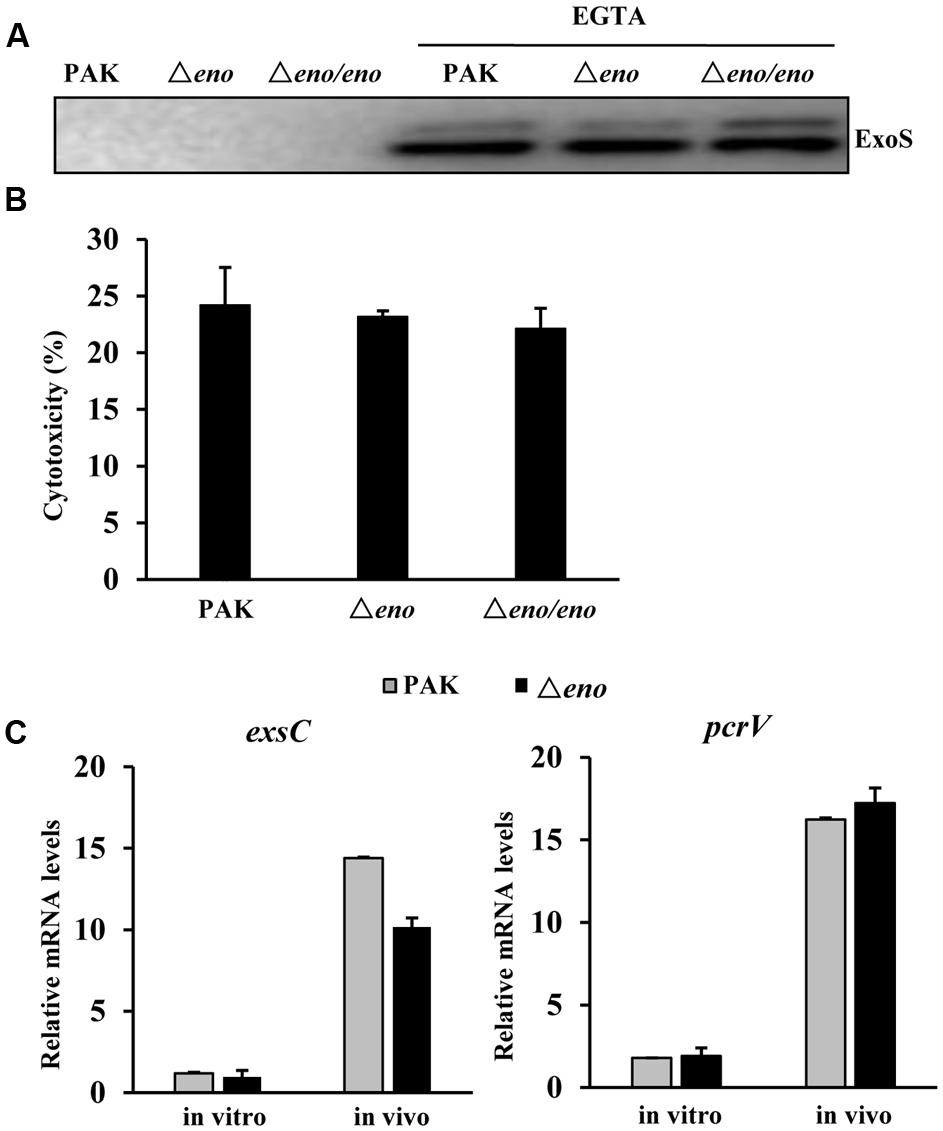
FIGURE 2. Role of enolase in the regulation of T3SS. (A) Wild type PAK, the Δeno mutant and complemented strain were grown with or without 5 mM EGTA for 3 h. The supernatants from equal amount of bacteria were collected by centrifugation and the levels of ExoS were determined by western blot analysis. The data is representative of three independent experiments. (B) Cytotoxicity of wild type PAK, the Δeno mutant and complemented strains to Hela cells. HeLa cells were infected with indicated strain at an MOI of 30 for 3 h. The relative cytotoxicity was determined by the LDH release assay. The error bars represente the standard errors. (C) Mice were infected with wild type PAK or the Δeno mutant for 6 h. Bacteria were harvested from BALFs of the infected mice. Bacteria grown in LB were used as in vitro samples. RNAs were extracted from the bacteria and the relative levels of mRNA were determined by qRT-PCR. Results represent means ± SD.
Next, we compared the impact of neutrophils on the Δeno mutant and wild type PAK. The bacteria were incubated with differentiated HL-60 (designated as dHL-60 hereafter) and undifferentiated HL-60 in RPMI-1640 medium. Compared to wild type PAK, the Δeno mutant was more susceptible to the dHL-60 mediated growth inhibition (Figure 3A). A major bactericidal mechanism of neutrophils is production of ROS (Arai et al., 2001; Alalwani et al., 2009). As shown in Figure 3B, dHL-60 generated large amount of ROS upon encountering PAK or the Δeno mutant. Therefore, we suspected that the Δeno mutant is more susceptible to oxidative stresses. Indeed, in a disk diffusion assay, H2O2 caused bigger inhibition zone on the Δeno mutant than that on the wild type PAK (Figure 3C). And treatment with H2O2 resulted in significant lower survival rate of the Δeno mutant (Figure 3D). Complementation with an eno gene restored the bacteria tolerance to H2O2 (Figure 3D). These results suggest that enolase is involved in the bacterial tolerance to oxidative stresses.
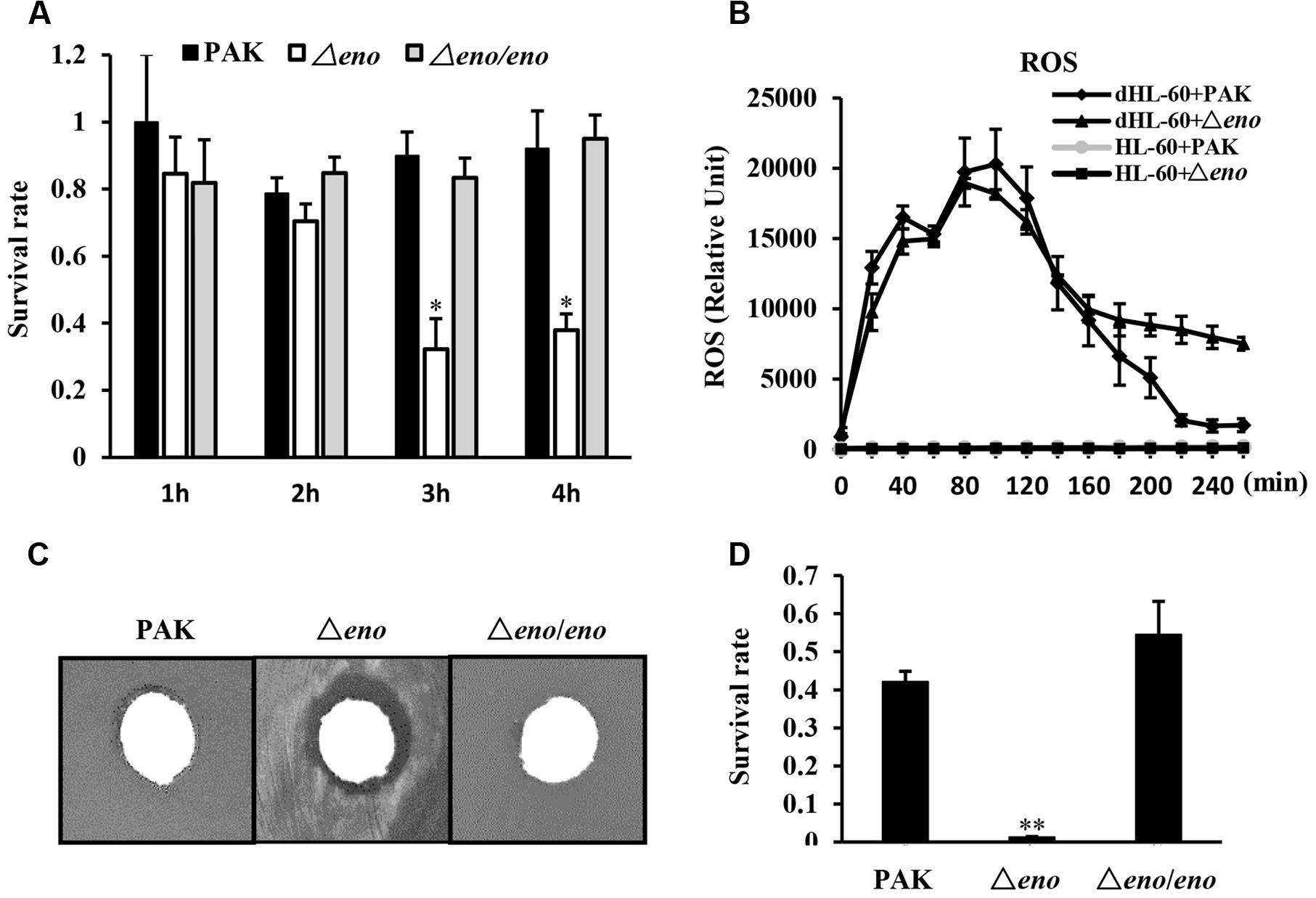
FIGURE 3. Role of enolase in bacterial tolerance to oxidative stresses. (A) Bacteria of indicated strains were incubated with differentiated HL-60 (dHL-60) cells or undifferentiated HL-60 cells. The live bacteria number was determined by plating. The relative growth rate of each strain was calculated as the ratio of live bacterial number in the presence of dHL-60 relative to that in the presence of undifferentiated HL-60. ∗P < 0.05 compared to PAK or the complemented strain by student’s t-test. (B) Differentiated or undifferentiated HL-60 cells were incubated with PAK or the Δeno mutant. ROS levels were determined by a fluorescence spectrophotometer at different time points. (C) Circular filter paper was immersed in H2O2 (250 mM) for 5 s and placed in the center of plates that were spread with PAK, the Δeno mutant or complemented strain. (D) Indicated strains were treated with H2O2 (10 mM) for 10 min and the numbers of live bacteria were determined by serial dilution and plating. ∗∗P < 0.01 compared to PAK or the complemented strain by student’s t-test.
Mutation of eno Resulted in Down Regulation of ahpB and ahpC
In P. aeruginosa, the chromosomally encoded catalases (KatA and KatB), and alkyl hydroperoxide reductases (AhpB and AhpC) play important roles in the bacterial tolerance to oxidative stresses (Hassett et al., 1992; Ma et al., 1999). Thus, we examined whether enolase affects the expression of those genes. In wild type PAK, treatment with H2O2 induced the expression of katA, katB, ahpB, and ahpC. In the eno mutant, similar expression levels of katA and katB were observed (Figures 4A,B), except for the level of katA in the absence of H2O2, which was higher than that in wild type PAK (Figure 4A). However, the mRNA levels of ahpB and ahpC in the Δeno mutant were 20- and 2-fold lower, respectively, in the presence of H2O2 (Figures 4C,D). To further confirm the expression levels of ahpB and ahpC, we constructed transcriptional fusions of ahpB promoter (PahpB) or ahpC promoter (PahpC) with lacZ reporter gene. In the presence of H2O2, the expression levels of ahpB-lacZ and ahpC-lacZ were reduced by ∼45 and 35% in the Δeno mutant, respectively (Figures 4E,F).
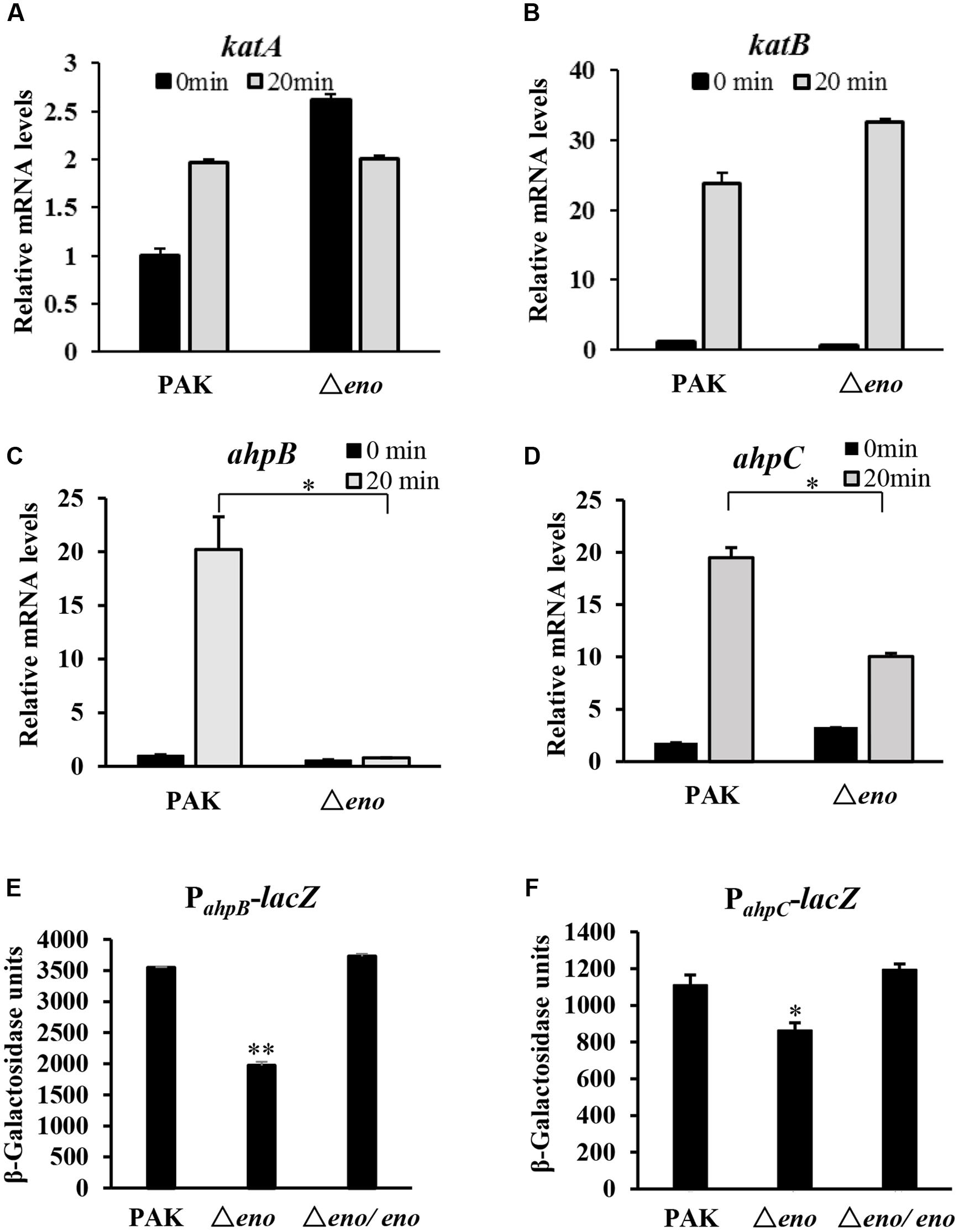
FIGURE 4. Expression of oxidative stress response genes. Wild type PAK and Δeno mutant strain were treated with 10 mM H2O2 for 20 min, followed by RNA extraction. The relative mRNA levels of katA (A), katB (B), ahpB (C), and ahpC (D) were determined by qRT-PCR. ∗P < 0.05 compared to the wild type PAK by Student’s t-test. Results represent means ± SD, and data are representative of three independent experiments. Indicated strains containing PahpB- (E) or PahpC-lacZ (F) transcriptional fusion were treated with 10 mM H2O2 for 30 min. The bacteria were collected, followed by β-Galactosidase assay. ∗P < 0.05, ∗∗P < 0.01 compared to the wild type PAK or the eno complement strain by Student’s t-test. Results represent means ± SD, and data are representative of three independent experiments.
In P. aeruginosa, OxyR activates the expression of katA, katB, ahpB, and ahpC in response to oxidative stresses (Heo et al., 2009). Since the expression levels of katA and katB were similar between wild type PAK and the Δeno mutant in the presence of H2O2, we suspect that the expression and function of OxyR are normal in the eno mutant. Indeed, the mRNA levels of OxyR were similar between wild type PAK and the eno mutant with or without H2O2 treatment (Supplementary Figure S2A). In addition, expression of prpL, toxA, and rgsA, under the control of OxyR, was not affected by the mutation of eno (Supplementary Figures S2B–D). These results suggest that enolase affects the expression of ahpB and ahpC independent of the OxyR.
Next, we examined the expression levels of katA, katB, ahpB, and ahpC in bacteria during mouse lung infection. At six hpi, bacteria were collected from BALF, followed by RNA extraction and qRT-PCR. The mRNA levels of katA and katB in the Δeno mutant were slightly higher than those in wild type PAK (Figures 5C,D), however, the ahpB and ahpC mRNA levels were lower in the Δeno mutant (Figures 5A,B).
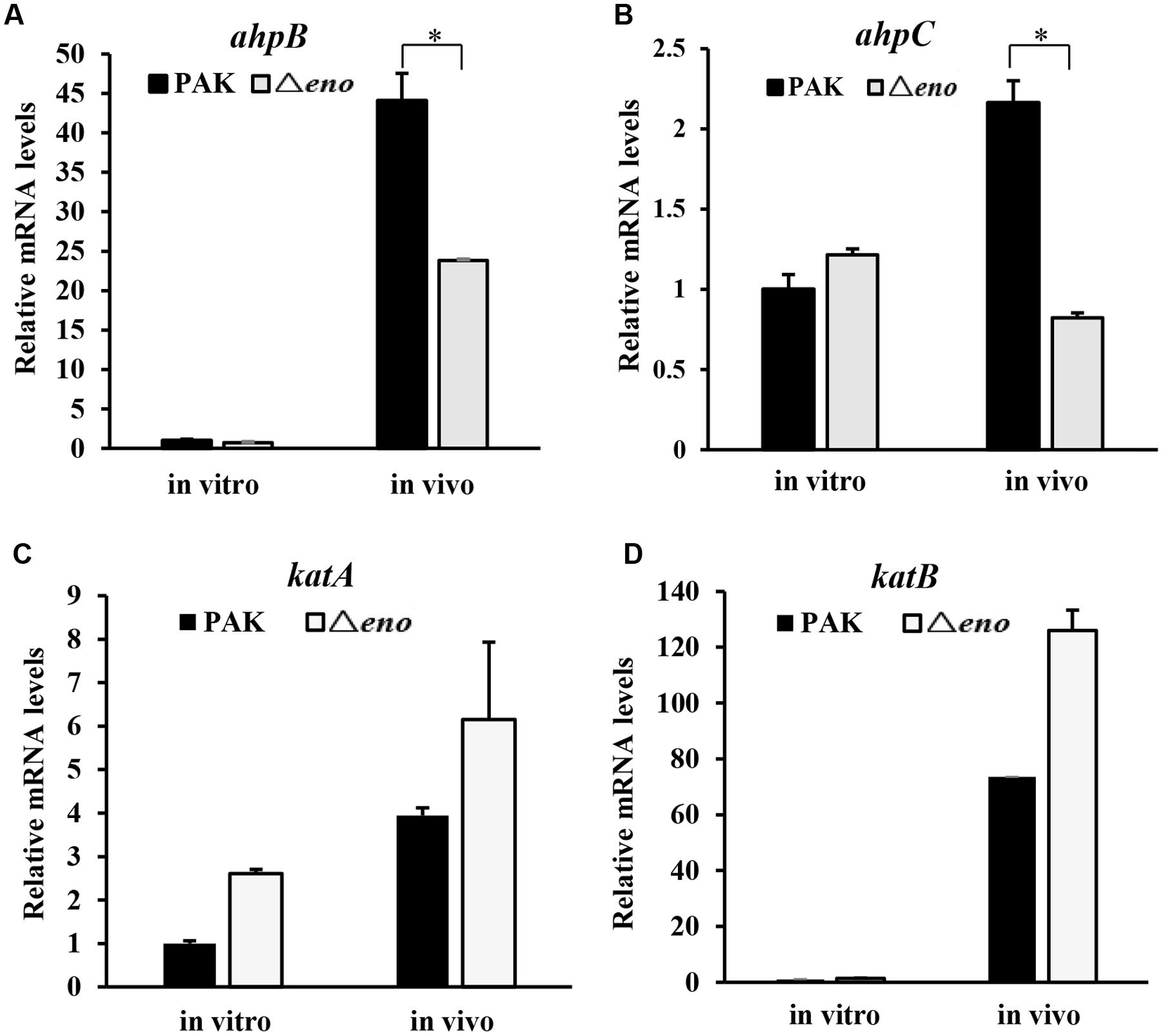
FIGURE 5. Role of enolase in the expression of oxidative stress response genes in vivo. Mice were infected with wild type PAK or the Δeno mutant for 6 h. Bacteria were harvested from BALFs of the infected mice. Bacteria grown in LB were used as in vitro samples. RNA was extracted from the bacteria and the relative mRNA levels of ahpB (A), ahpC (B), katA (C), and katB (D) were determined by qRT-PCR. Results represent means ± SD. ∗P < 0.05 compared to wild type PAK by student’s t-test.
Overexpression of ahpB in the Δeno Mutant Restores the Bacterial Tolerance to H2O2 and Virulence
The in vitro and in vivo results shown above demonstrate defective expression of ahpB and ahpC in the Δeno mutant, which might be the cause of reduced tolerance to oxidative stresses. To test this further, we overexpressed the two genes individually or together in the Δeno mutant and examined the bacterial survival rates after H2O2 treatment. As shown in Figure 6A, overexpression of ahpB but not ahpC in the Δeno mutant restored the survival rate. Compared to ahpB alone, co-overexpression of ahpB and ahpC only slightly increased the bacterial survival rate. Consistently, overexpression of ahpB but not ahpC in the Δeno mutant restored the bacterial growth in the presence of dHL60 (Figure 6B).
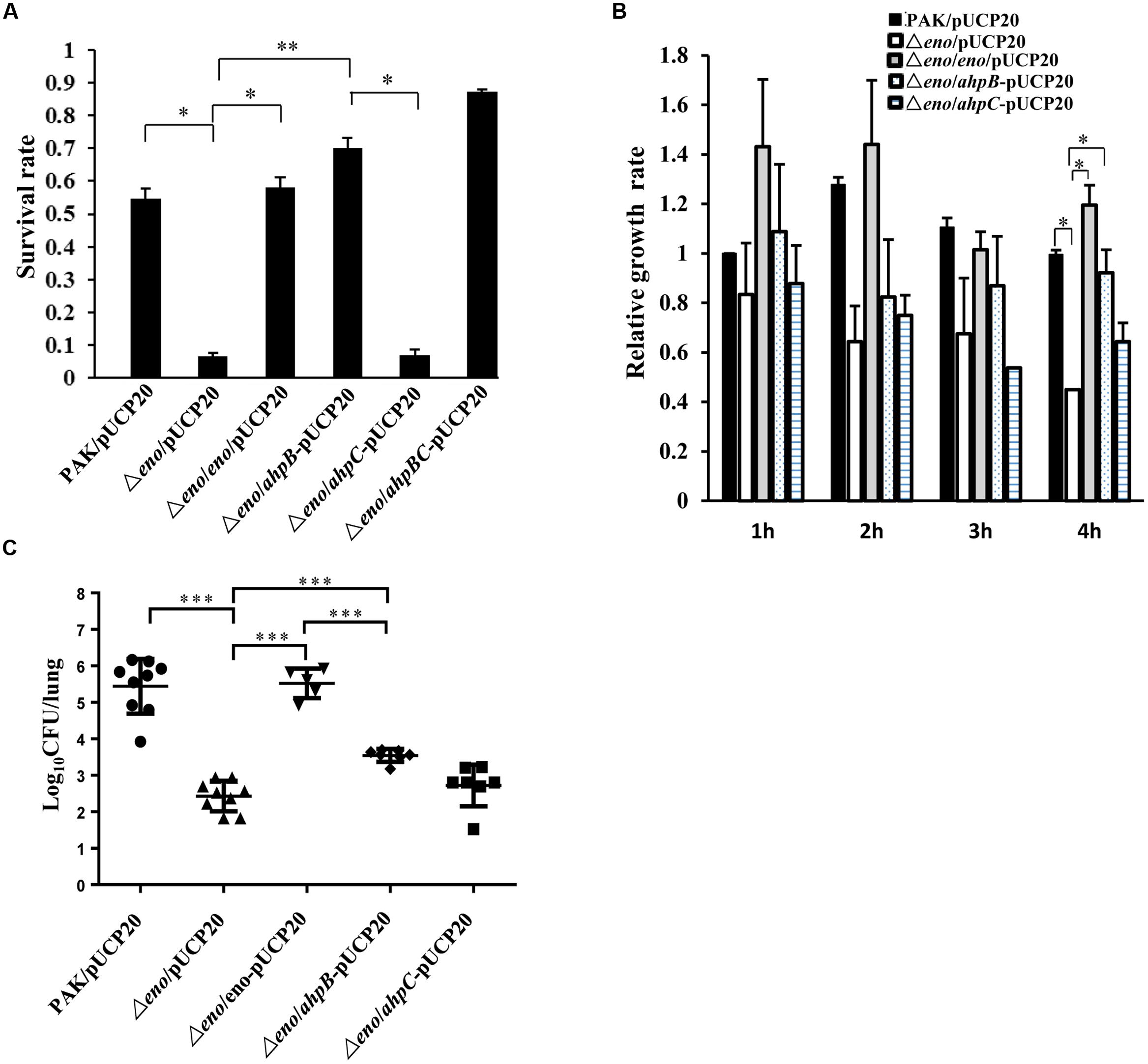
FIGURE 6. Effect of overexpression of ahpB or ahpC on bacterial tolerance to H2O2 and virulence in the Δeno mutant. Empty vector pUCP20 was transferred into wild type PAK, the Δeno mutant and the complemented strain, and plasmid overexpressing ahpB, ahpC, or both of the genes was transferred into the Δeno mutant. (A) The indicated strains were treated with H2O2 (10 mM) for 10 min and the live bacteria numbers were determined by serial dilution and plating. Results represent means ± SD, and data are representative of three independent experiments. ∗P < 0.05, ∗∗P < 0.01 by Student’s t-test. (B) The bacteria were incubated with dHL-60 cells and undifferentiated HL-60 cells. The live bacteria number was determined by plating. The relative growth rate of each strain was calculated as the ratio of live bacterial number in the presence of dHL-60 relative to that in the presence of undifferentiated HL-60. (C) Mice were infected with the indicated strains intranasally. At 16 hpi, lungs from the infected mice were isolated. Bacterial loads were determined by serial dilution and plating. ∗∗∗P < 0.001 by the Mann–Whitney test.
In the mouse acute pneumonia model, overexpression of ahpB in the Δeno mutant increased the average bacterial load by ∼10-fold. However, overexpression of ahpC had no effect on the bacterial load (Figure 6C). In addition, overexpression of ahpB in the Δeno mutant did not alter the bacterial growth rate in LB medium (Supplementary Figure S1). Therefore, these results suggest that down regulation of ahpB is the major cause of decreased tolerance to H2O2 and the reduced bacterial load of the Δeno mutant.
Discussion
In this study, we show that enolase is required for the virulence of P. aeruginosa in a murine acute pneumonia model. Further experimental results demonstrated that enolase affects the expression of two of the oxidative stress responsive genes, ahpB and ahpC. Mutation of eno abolished H2O2 induced expression of ahpB, but only partially affected the expression of ahpC. By overexpressing ahpB or ahpC in the Δeno mutant, we demonstrate that ahpB plays a major role in the reduced bacterial tolerance to oxidative stresses and virulence.
OxyR plays a major role in the regulation of oxidative stress responsive genes (Wei et al., 2012; Jo et al., 2015). In the Δeno mutant, the mRNA level of oxyR is similar to that in the wild type strain. Expression levels of known OxyR regulated genes, including katA, katB, toxA, prpL, rgsA are similar between the Δeno mutant and wild type strain in the presence of H2O2. These results suggest that enolase is unlikely to affect the protein level and function of OxyR. We thus suspect that enolase might affect the expression of an unknown regulatory gene for the ahpB and/or ahpC. The N-terminus coding region of ahpB overlaps with PA0847, which is transcribed in the opposite direction. Thus, the promoters of ahpB and PA0847 should be inside the coding region of each other. We suspected that the transcription initiation or elongation of ahpB might be interfered by the RNA polymerase complex transcribing PA0847. However, the promoter activity of PA0847 in the Δeno mutant was only 20% higher than that in the wild type strain in the presence of H2O2, as revealed by a β-galactosidase assay with a PA0847 promoter lacZ transcriptional fusion. Therefore, it is likely that other regulatory genes are involved in the regulation of ahpB.
Enolase belongs to a glycolytic enzyme, catalyzing the reversible dehydration of 2-phosphoglycerate to phosphoenolpyruvate (Sekowska et al., 2004). Besides, enolase also forms a complex with PNPase, a RNA helicase RhlB and ribonuclease E (RNase E), namely the RNA degradosome (Callaghan et al., 2004; Carpousis, 2007). The RNA degradosome plays an important role in RNA processing (Carpousis, 2007; Mildenhall et al., 2016). In Salmonella enterica, RNase E is involved in the regulation of genes required for intracellular replication (Yang et al., 2008), while in Yersinia pseudotuberculosis, RNase E regulates the expression of T3SS genes (Yang et al., 2008). PNPase plays important role in bacterial responses to various environmental stresses (Goverde et al., 1998; Clements et al., 2002; Len et al., 2004; Rosenzweig et al., 2005; Anderson and Dunman, 2009; Lawal et al., 2011). In Yersinia, it has been demonstrated that PNPase is required for the expression of T3SS and bacterial virulence (Rosenzweig et al., 2005). Our previous study in P. aeruginosa demonstrated that deletion of the RNA binding domains of PNPase leads to defective T3SS and attenuated virulence (Chen et al., 2016). However, the in vitro and in vivo experiments shown in this study demonstrated a normal T3SS function in the Δeno mutant. These results suggest that enolase and other components of the RNA degradosome might affect the expression of distinctive subsets of genes.
In addition, enolase is a major type of moonlighting proteins, which are a group of proteins that have more than one unique biological functions (Henderson and Martin, 2011; Henderson, 2014). Enolase has been found on the cell surface of a growing number of bacteria and play roles in bacterial virulence (Henderson and Martin, 2011; Henderson, 2014). For example, the surface exposed enolase functions as an adhesion by binding to host plasminogen in various Streptococci, including Streptococcus pneumoniae, S. pyogenes, S. gordonii, S. mutans, S. suis, and S. canis (Pancholi and Fischetti, 1998; Ge et al., 2004; Esgleas et al., 2008; Kesimer et al., 2008; Fulde et al., 2013; Figueiredo et al., 2015). Mutation of the plasminogen bringing site of enolase reduced the bacterial virulence of S. pneumoniae in a murine intranasal infection model (Bergmann et al., 2003). Immunization with the enolase of S. suis conferred protection to mice against infection by the bacteria (Feng et al., 2009). In Gram negative bacteria, Borrelia burgdorferi and Aeromonas hydrophila, enolase has been found on cell surface (Carlson et al., 2007; Sierra et al., 2010). The plasminogen binding motif of enolase contributes to bacterial virulence of A. hydrophila, and immunization with enolase conferred protection (Sierra et al., 2010). Here in this study, we demonstrated that overexpression of ahpB in the Δeno mutant fully restored the bacterial tolerance to H2O2, but only partially restored the virulence in the mouse pneumonia model. These results suggest the enolase might play additional roles in the virulence of P. aeruginosa. Further studies on the subcellular location and regulatory functions of the enolase are needed to fully elucidate the role of enolase in bacterial pathogenesis.
Author Contributions
Conceived and designed the experiments: WW, YW, YJ, and SJ. Performed the experiments: YW, FC, YL, RC, CL, XP, and YJ. Analyzed the data: YW, QZ, WW, ZC, SJ, and YJ. Wrote the paper: YW, WW, and SJ.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgments
This work was supported by National Science Foundation of China (31670130, 31370168, and 31370167); Program of international S&T cooperation (2015DFG32500) and Science and Technology Committee of Tianjin (15JCYBJC53900 and 15JCZDJC33000). The funders had no role in study design, data collection and interpretation, or the decision to submit the work for publication.
Supplementary Material
The Supplementary Material for this article can be found online at: http://journal.frontiersin.org/article/10.3389/fmicb.2016.01999/full#supplementary-material
FIGURE S1 | Growth of strains in LB medium. Same number of bacteria of each indicated strain were inoculated in LB and cultured at 37°C with agitation. (A) Growth curve of indicated strain in LB. The OD600 of each culture was monitored every hour for 12 h. (B) After 12-h growth, the bacterial number of each strain was determined by serial dilution and plating.
FIGURE S2 | Expression of oxyR and genes regulated by it. Wild type PAK and the Δeno mutant were treated with 10 mM H2O2 for 20 min. Total RNA was isolated and the relative mRNA levels of oxyR (A), prpL (B), toxA (C), and rgsA (D) were determined by qRT-PCR. Results represent means ± SD, and data are representative of three independent experiments.
References
Alalwani, S. M., Sierigk, J., Herr, C., Pinkenburg, O., Gallo, R., Vogelmeier, C., et al. (2009). The antimicrobial peptide LL-37 modulates the inflammatory and host defense response of human neutrophils. Eur. J. Immunol. 8, 1118–1126. doi: 10.1002/eji.200939275
Anderson, K. L., and Dunman, P. M. (2009). Messenger RNA turnover processes in Escherichia coli, Bacillus subtilis, and emerging studies in Staphylococcus aureus. Int. J. Microbiol. 2009:525491. doi: 10.1155/2009/525491
Arai, T., Yoshikai, Y., Kamiya, J., Nagino, M., Uesaka, K., Yuasa, N., et al. (2001). Bilirubin impairs bactericidal activity of neutrophils through an antioxidant mechanism in vitro. J. Surg. Res. 96, 107–113. doi: 10.1006/jsre.2000.6061
Balasubramanian, D., Schneper, L., Kumari, H., and Mathee, K. (2012). A dynamic and intricate regulatory network determines Pseudomonas aeruginosa virulence. Nucleic Acids Res. 41, 1–20. doi: 10.1093/nar/gks1039
Bergmann, S., Wild, D., Diekmann, O., Frank, R., Bracht, D., Chhatwal, G. S., et al. (2003). Identification of a novel plasmin(ogen)-binding motif in surface displayed α-enolase of Streptococcus pneumoniae. Mol. Microbiol. 49, 411–423. doi: 10.1046/j.1365-2958.2003.03557.x
Bleves, S., Viarre, V., Salacha, R., Michel, G. P. F., Filloux, A., and Voulhoux, R. (2010). Protein secretion systems in Pseudomonas aeruginosa: a wealth of pathogenic weapons. Int. J. Med. Microbiol. 300, 534–543. doi: 10.1016/j.ijmm.2010.08.005
Brannon, M. K., Davis, J. M., Mathias, J. R., Hall, C. J., Emerson, J. C., Crosier, P. S., et al. (2009). Pseudomonas aeruginosa type III secretion system interacts with phagocytes to modulate systemic infection of zebrafish embryos. Cell Microbiol. 11, 755–768. doi: 10.1111/j.1462-5822.2009.01288.x
Burger, A., Whiteley, C., and Boshoff, A. (2011). Current perspectives of the Escherichia coli RNA degradosome. Biotechnol. Lett. 33, 2337–2350. doi: 10.1007/s10529-011-0713-6
Callaghan, A. J., Aurikko, J. P., Ilag, L. L., Grossmann, J. G., Chandran, V., Kühnel, K., et al. (2004). Studies of the RNA degradosome-organizing domain of the Escherichia coli ribonuclease RNase E. J. Mol. Biol. 340, 965–979. doi: 10.1016/j.jmb.2004.05.046
Canback, B., Andersson, S. G., and Kurland, C. G. (2002). The global phylogeny of glycolytic enzymes. Proc. Natl. Acad. Sci. U.S.A. 99, 6097–6102. doi: 10.1073/pnas.082112499
Carlson, P. E., Carroll, J. A., O’dee, D. M., and Nau, G. J. (2007). Modulation of virulence factors in Francisella tularensis determines human macrophage responses. Microb. Pathog. 42, 204–214. doi: 10.1016/j.micpath.2007.02.001
Carpousis, A. J. (2007). The RNA degradosome of Escherichia coli: an mRNA-degrading machine assembled on RNase E. Annu. Rev. Microbiol. 61, 71–87. doi: 10.1146/annurev.micro.61.080706.093440
Chandran, V., and Luisi, B. F. (2006). Recognition of enolase in the Escherichia coli RNA degradosome. J. Mol. Biol. 358, 8–15. doi: 10.1016/j.jmb.2006.02.012
Chen, A., and Seifert, H. S. (2011). Neisseria gonorrhoeae-mediated inhibition of apoptotic signalling in polymorphonuclear leukocytes. Infect Immun. 79, 4447–4458. doi: 10.1128/IAI.01267-10
Chen, R., Weng, Y., Zhu, F., Jin, Y., Liu, C., Pan, X., et al. (2016). Polynucleotide Phosphorylase regulates multiple virulence factors and the stabilities of small RNAs RsmY/Z in Pseudomonas aeruginosa. Front. Microbiol. 7:247. doi: 10.3389/fmicb.2016.00247
Choi, K.-H., and Schweizer, H. P. (2006). Mini-Tn7 insertion in bacteria with single attTn7 sites: example Pseudomonas aeruginosa. Nat. Protoc. 1, 153–161. doi: 10.1038/nprot.2006.26
Clements, M. O., Eriksson, S., Thompson, A., Lucchini, S., Hinton, J. C., Normark, S., et al. (2002). Polynucleotide phosphorylase is a global regulator of virulence and persistency in Salmonella enterica. Proc. Natl. Acad. Sci. U.S.A. 99, 8784–8789. doi: 10.1073/pnas.132047099
Diaz, E., Ulldemolins, M., Lisboa, T., Rello, J., Khardori, N., and Khardori, R. (2005). Etiology of ventilator-associated pneumonia. Clin. Chest Med. 26, 87–95. doi: 10.1016/j.ccm.2004.10.017
Diaz, M. H., and Hauser, A. R. (2010). Pseudomonas aeruginosa cytotoxin ExoU is injected into phagocytic cells during acute pneumonia. Infect. Immun. 78, 1447–1456. doi: 10.1128/IAI.01134-09
Esgleas, M., Li, Y., Hancock, M. A., Harel, J., Dubreuil, J. D., and Gottschalk, M. (2008). Isolation and characterization of alpha-enolase, a novel fibronectin-binding protein from Streptococcus suis. Microbiology 154, 2668–2679. doi: 10.1099/mic.0.2008/017145-0
Favaro, R., and Deho, G. (2003). Polynucleotide phosphorylase-deficient mutants of Pseudomonas putida. J. Bacteriol. 185, 5279–5286. doi: 10.1128/JB.185.17.5279-5286.2003
Feltman, H., Schulert, G., Khan, S., Jain, M., Peterson, L., and Hauser, A. R. (2001). Prevalence of type III secretion genes in clinical and environmental isolates of Pseudomonas aeruginosa. Microbiology 147, 2659–2669. doi: 10.1099/00221287-147-10-2659
Feng, Y. J., Pan, X. Z., Wen, S., Wang, C. J., Zhang, H. M., Li, X. F., et al. (2009). Streptococcus suis enolase functions as a protective antigen displayed on the bacterial cell surface. J. Infect. Dis. 200, 1583–1592. doi: 10.1086/644602
Figueiredo, B. C., Da’dara, A. A., Oliveira, S. C., and Skelly, P. J. (2015). Schistosomes enhance plasminogen activation: the role of tegumental enolase. PLoS Pathog. 11:e1005335. doi: 10.1371/journal.ppat.1005335
Fulde, M., Rohde, M., Polok, A., Preissner, K. T., Chhatwal, G. S., and Bergmann, S. (2013). Cooperative plasminogen recruitment to the surface of Streptococcus canis via M protein and enolase enhances bacterial survival. mBio 4, 49–52. doi: 10.1128/mBio.00629-12
Ge, J., Catt, D. M., and Gregory, R. L. (2004). Streptococcus mutans surface alpha-enolase binds salivary mucin MG2 and human plasminogen. Infect. Immun. 72, 6748–6752. doi: 10.1128/IAI.72.11.6748-6752.2004
Goverde, R. L., Huis in’t Veld, J. H., Kusters, J. G., and Mooi, F. R. (1998). The psychrotrophic bacterium Yersinia enterocolitica requires expression of pnp, the gene for polynucleotide phosphorylase, for growth at low temperature (5°C). Mol. Microbiol. 28, 555–569. doi: 10.1046/j.1365-2958.1998.00816.x
Hassett, D. J., Charniga, L., Bean, K., Ohman, D. E., and Cohen, M. S. (1992). Response of Pseudomonas aeruginosa to pyocyanin: mechanisms of resistance, antioxidant defenses, and demonstration of a manganese-cofactored superoxide dismutase. Infect. Immun. 60, 328–336.
Henderson, B. (2014). An overview of protein moonlighting in bacterial infection. Biochem. Soc. Trans. 42, 1720–1727. doi: 10.1042/BST20140236
Henderson, B., and Martin, A. (2011). Bacterial moonlighting proteins and bacterial virulence. Curr. Top. Microbiol. Immunol. 358, 155–213.
Heo, Y. J., Chung, I. Y., Cho, W. J., Lee, B. Y., Kim, J. H., Choi, K. H., et al. (2009). The major catalase gene (katA) of PA14 is under both positive and negative control of the global transactivator OxyR in response to hydrogen peroxide. J. Bacteriol. 192, 381–390. doi: 10.1128/JB.00980-09
Hoang, T. T., Karkhoff-Schweizer, R. R., Kutchma, A. J., and Schweizer, H. P. (1998). A broad-host-range Flp-FRT recombination system for site-specific excision of chromosomally-located DNA sequences: application for isolation of unmarked Pseudomonas aeruginosa mutants. Gene 212, 77–86. doi: 10.1016/S0378-1119(98)00130-9
Hornef, M. W., Roggenkamp, A., Geiger, A. M., Hogardt, M., Jacobi, C. A., and Heesemann, J. (2000). Triggering the ExoS regulon of Pseudomonas aeruginosa: a GFP-reporter analysis of exoenzyme (Exo) S, ExoT and ExoU synthesis. Microb. Pathog. 29, 329–343. doi: 10.1006/mpat.2000.0398
Howell, H. A., Logan, L. K., and Hauser, A. R. (2012). Type III secretion of ExoU is critical during early Pseudomonas aeruginosa pneumonia. mBio 4, 49–52. doi: 10.1128/mBio.00032-13
Jo, I., Chung, I. Y., Bae, H. W., Kim, J. S., Song, S., Cho, Y. H., et al. (2015). Structural details o the OxyR peroxide-sensing mechanism. Proc. Natl. Acad. Sci. U.S.A. 112, 6443–6448. doi: 10.1073/pnas.1424495112
Kesimer, M., Kiliç, N., Mehrotra, R., Thornton, D. J., and Sheehan, J. K. (2008). Identification of Salivary Mucin MUC7 Binding Proteins from Streptococcus gordonii. BMC Microbiol. 9:163. doi: 10.1186/1471-2180-9-163
Lawal, A., Jejelowo, O., Chopra, A. K., and Rosenzweig, J. A. (2011). Ribonucleases and bacterial virulence. Microb. Biotechnol. 4, 558–571. doi: 10.1111/j.1751-7915.2010.00212.x
Lee, J. S., Heo, Y. J., Lee, J. K., and Cho, Y. H. (2005). KatA, the major catalase, is critical for osmoprotection and virulence in Pseudomonas aeruginosa PA14. Infect. Immun. 73, 4399–4403. doi: 10.1128/IAI.73.7.4399-4403.2005
Len, A. C. L., Harty, D. W. S., and Jacques, N. A. (2004). Stress-responsive proteins are upregulated in Streptococcus mutans during acid tolerance. Microbiology 150, 1339–1351. doi: 10.1099/mic.0.27008-0
Li, K., Xu, C., Jin, Y., Sun, Z., Liu, C., Shi, J., et al. (2013). SuhB is a regulator of multiple virulence genes and essential for pathogenesis of Pseudomonas aeruginosa. mBio 4:e00419-13. doi: 10.1128/mBio.00419-13
Luo, Q., and Jin, S. (2008). Molecular determinants in regulating Pseudomonas aeruginosa type III secretion system–a review. Acta Microbiol. Sin. 48, 1413–1417.
Ma, J., Ochsner, U., Klotz, M. G., Nanayakkara, V., Howell, M., Johnson, Z., et al. (1999). Bacterioferritin A modulates catalase A (KatA) activity and resistance to hydrogen peroxide in Pseudomonas aeruginosa. J Bacteriol. 181, 3730–3742.
Matos, R. G., Bárria, C., Pobre, V., Andrade, J. M., and Arraiano, C. M. (2011). Exoribonucleases as modulators of virulence in pathogenic bacteria. Front. Cell Infect. Microbiol. 2:65. doi: 10.3389/fcimb.2012.00065
Mcbride, R. B. W. J. H. (2005). Mandell, Douglas, and Bennett’s Principles and Practice of Infectious Diseases, 7th Edn. Amsterdam: Elsevier/Churchill Livingstone.
Mildenhall, K. B., Wiese, N., Chung, D., Maples, V. F., Mohanty, B. K., and Kushner, S. R. (2016). RNase E-based degradosome modulates polyadenylation of mRNAs after Rho-independent transcription terminators in Escherichia coli. Mol. Microbiol. 101, 645–655. doi: 10.1111/mmi.13413
Miller, J. H. (1972). Experiments in Molecular Genetics. Cold Spring Harbor, NY: Cold Spring Harbor Laboratory, 352–359.
Morita, T., Kawamoto, H., Mizota, T., Inada, T., and Aiba, H. (2004). Enolase in the RNA degradosome plays a crucial role in the rapid decay of glucose transporter mRNA in the response to phosphosugar stress in Escherichia coli. Mol. Microbiol. 54, 1063–1075. doi: 10.1111/j.1365-2958.2004.04329.x
Morita, Y., Tomida, J., and Kawamura, Y. (2014). Responses of Pseudomonas aeruginosa to antimicrobials. Front. Microbiol. 4:422. doi: 10.3389/fmicb.2013.00422
Nurmohamed, S., McKay, A. R., Robinson, C. V., and Luisi, B. F. (2010). Molecular recognition between Escherichia coli enolase and ribonuclease E. Acta Crystallogr. D Biol. Crystallogr. 66, 1036–1040. doi: 10.1107/S0907444910030015
Ochsner, U. A., Vasil, M. L., Alsabbagh, E., Parvatiyar, K., and Hassett, D. J. (2000). Role of the Pseudomonas aeruginosa oxyR-recG operon in oxidative stress defense and DNA repair: OxyR-dependent regulation of katB-ankB, ahpB, and ahpC-ahpF. J. Bacteriol. 182, 4533–4544. doi: 10.1128/JB.182.16.4533-4544.2000
Pancholi, V., and Fischetti, V. A. (1998). alpha-enolase, a novel strong plasmin(ogen) binding protein on the surface of pathogenic streptococci. J. Biol. Chem. 273, 14503–14515. doi: 10.1074/jbc.273.23.14503
Pha, K., and Navarro, L. (2016). Yersinia type III effectors perturb host innate immune responses. World J. Biol. Chem. 7, 1–13. doi: 10.4331/wjbc.v7.i1.1
Plano, G. V., and Schesser, K. (2013). The Yersinia pestis type III secretion system: expression, assembly and role in the evasion of host defenses. Immunol. Res. 57, 237–245. doi: 10.1007/s12026-013-8454-3
Poole, K. (2011). Pseudomonas aeruginosa: resistance to the max. Front. Microbiol. 2:65. doi: 10.3389/fmicb.2011.00065
Rahme, L., Ausubel, F., Cao, H., Drenkard, E., Goumnerov, B., Lau, G., et al. (2000). Plants and animals share functionally common bacterial virulence factors. Proc. Natl. Acad. Sci. U.S.A. 97, 8815–8821. doi: 10.1073/pnas.97.16.8815
Rangel, S. M., Diaz, M. H., Knoten, C. A., Zhang, A., and Hauser, A. R. (2015). The role of ExoS in dissemination of Pseudomonas aeruginosa during pneumonia. PLoS Pathog. 11:e1004945. doi: 10.1371/journal.ppat.1004945
Rosenzweig, J. A., Weltman, G., Plano, G. V., and Schesser, K. (2005). Modulation of Yersinia type three secretion system by the S1 domain of polynucleotide phosphorylase. J. Biol. Chem. 280, 156–163. doi: 10.1074/jbc.M405662200
Ruzin, A., Keeney, D., and Bradford, P. A. (2007). AdeABC multidrug efflux pump is associated with decreased susceptibility to tigecycline in Acinetobacter calcoaceticus–Acinetobacter baumannii complex. J. Antimicrob. Chemother. 59, 1001–1004. doi: 10.1093/jac/dkm058
Saramago, M., Bárria, C., Santos, R. F. D., Silva, I. J., Pobre, V., Domingues, S., et al. (2014). The role of RNases in the regulation of small RNAs. Curr. Opin. Microbiol. 18, 105–115. doi: 10.1016/j.mib.2014.02.009
Savli, H., Karadenizli, A., Kolayli, F., Gundes, S., Ozbek, U., and Vahaboglu, H. (2003). Expression stability of six housekeeping genes: a proposal for resistance gene quantification studies of Pseudomonas aeruginosa by real-time quantitative RT-PCR. J. Med. Microbiol. 52, 403–408. doi: 10.1099/jmm.0.05132-0
Sekowska, A., Dénervaud, V., Ashida, H., Michoud, K., Haas, D., Yokota, A., et al. (2004). Bacterial variations on the methionine salvage pathway. BMC Microbiol. 4:9. doi: 10.1186/1471-2180-4-9
Shaver, C. M., and Hauser, A. R. (2004). Relative contributions of Pseudomonas aeruginosa ExoU, ExoS, and ExoT to virulence in the lung. Infect. Immun. 72, 6969–6977. doi: 10.1128/IAI.72.12.6969-6977.2004
Sierra, J. C., Suarez, G., Jian, S., Baze, W. B., Foltz, S. M., and Chopra, A. K. (2010). Unraveling the mechanism of action of a new type III secretion system effector AexU from Aeromonas hydrophila. Microb. Pathog. 49, 122–134. doi: 10.1016/j.micpath.2010.05.011
Simon, R., Priefer, U., and Puhler, A. (1983). A broad host range mobilization system for in vivo genetic engineering: transposon mutagenesis in Gram-negative bacteria. Nat. Biotechnol. 1, 784–791. doi: 10.1038/nbt1183-784
Son, M. S., Matthews, W. Jr., Kang, Y., Nguyen, D. T., and Hoang, T. T. (2007). In vivo evidence of Pseudomonas aeruginosa nutrient acquisition and pathogenesis in the lungs of cystic fibrosis patients. Infect Immun. 75, 5313–5324. doi: 10.1128/IAI.01807-06
Sun, Z., Shi, J., Liu, C., Jin, Y., Li, K., Chen, R., et al. (2014). PrtR homeostasis contributes to Pseudomonas aeruginosa pathogenesis and resistance against ciprofloxacin. Infect Immun. 82, 1638–1647. doi: 10.1128/IAI.01388-13
Wei, Q., Minh, P. N. L., Dötsch, A., Hildebrand, F., Panmanee, W., Elfarash, A., et al. (2012). Global regulation of gene expression by OxyR in an important human opportunistic pathogen. Nucleic Acids Res. 40, 4320–4333. doi: 10.1093/nar/gks017
Wu, W., and Hsu, Y. L. (2009). CARD9 facilitates microbe-elicited production of reactive oxygen species by regulating the LyGDI-Rac1 complex. Nat. Immunol. 10, 1208–1214. doi: 10.1038/ni.1788
Wu, W., Huang, J., Duan, B., Traficante, D. C., Hong, H., Risech, M., et al. (2012). Th17-stimulating protein vaccines confer protection against Pseudomonas aeruginosa pneumonia. Am. J. Respir. Crit. Care Med. 186, 420–427. doi: 10.1164/rccm.201202-0182OC
Yang, J., Jain, C., and Schesser, K. (2008). RNase E regulates the Yersinia type 3 secretion system. J Bacteriol. 190, 3774–3778. doi: 10.1128/JB.00147-08
Keywords: Pseudomonas aeruginosa, enolase, oxidative stress response, bacterial virulence, gene regulation
Citation: Weng Y, Chen F, Liu Y, Zhao Q, Chen R, Pan X, Liu C, Cheng Z, Jin S, Jin Y and Wu W (2016) Pseudomonas aeruginosa Enolase Influences Bacterial Tolerance to Oxidative Stresses and Virulence. Front. Microbiol. 7:1999. doi: 10.3389/fmicb.2016.01999
Received: 11 September 2016; Accepted: 29 November 2016;
Published: 15 December 2016.
Edited by:
Yuji Morita, Aichi Gakuin University, JapanReviewed by:
Rolf Kümmerli, University of Zurich, SwitzerlandDouglas Gordon Storey, University of Calgary, Canada
James John Lazenby, University of Technology Sydney, Australia
Copyright © 2016 Weng, Chen, Liu, Zhao, Chen, Pan, Liu, Cheng, Jin, Jin and Wu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Weihui Wu, d3V3ZWlodWlAbmFua2FpLmVkdS5jbg== Yongxin Jin, eXhqaW5AbmFua2FpLmVkdS5jbg==