 Vivek Sharma
Vivek Sharma Richa Salwan2
Richa Salwan2- 1Department of Plant Pathology, Choudhary Sarwan Kumar Himachal Pradesh Agricultural University, Palampur, India
- 2Department of Veterinary Microbiology, Choudhary Sarwan Kumar Himachal Pradesh Agricultural University, Palampur, India
- 3Institute of Himalayan Bioresource Technology, Palampur, India
Genome-wide studies of transcripts expression help in systematic monitoring of genes and allow targeting of candidate genes for future research. In contrast to relatively stable genomic data, the expression of genes is dynamic and regulated both at time and space level at different level in. The variation in the rate of translation is specific for each protein. Both the inherent nature of an mRNA molecule to be translated and the external environmental stimuli can affect the efficiency of the translation process. In biocontrol agents (BCAs), the molecular response at translational level may represents noise-like response of absolute transcript level and an adaptive response to physiological and pathological situations representing subset of mRNAs population actively translated in a cell. The molecular responses of biocontrol are complex and involve multistage regulation of number of genes. The use of high-throughput techniques has led to rapid increase in volume of transcriptomics data of Trichoderma. In general, almost half of the variations of transcriptome and protein level are due to translational control. Thus, studies are required to integrate raw information from different “omics” approaches for accurate depiction of translational response of BCAs in interaction with plants and plant pathogens. The studies on translational status of only active mRNAs bridging with proteome data will help in accurate characterization of only a subset of mRNAs actively engaged in translation. This review highlights the associated bottlenecks and use of state-of-the-art procedures in addressing the gap to accelerate future accomplishment of biocontrol mechanisms.
Introduction
Trichoderma is a cosmopolitan and cardinal representative soil microflora of various climatic conditions (Herrera-Estrella, 2014). The biocontrol role of Trichoderma spp. have emerged as an attractive choice in agriculture sector due to their environmentally friendly nature over synthetic pesticides (Mukherjee et al., 2012, 2013). Among different biocontrol agents (BCAs), the genus Hypocrea/Trichoderma containing Trichoderma harzianum, Trichoderma atroviride, Hypocrea virens are probably the most explored BCAs (Schuster and Schmoll, 2010; Sharma and Shanmugam, 2012; Sharma and Salwan, 2017) and occupies over 60% of all registered biopesticides (Mukherjee et al., 2013). The continuous efforts on the evaluation of biocontrol potential of Trichoderma have led to the identification of several promising species/strains including T. harzianum (Yedidia et al., 1999; Cloyd et al., 2007), Trichoderma virens (Hermosa et al., 2000; Howell, 2006), Trichoderma viride (Papavizas, 1985), T. atroviride (Longa et al., 2009), Trichoderma polysporum (Zhang et al., 2015), and Trichoderma asperellum GDFS1009 (Wu et al., 2017). In recent studies, another potential strains of Trichoderma saturnisporum has been identified for its biocontrol potential (Sharma and Shanmugam, 2012; Diánez Martínez et al., 2016). In addition to primary application in agriculture, Hypocrea jecorina/Trichoderma reesei strains are molecular factory for cellulolytic enzymes (Merino and Cherry, 2007; Singh et al., 2015). The natural potential to secrete lytic enzymes, antibiotics, and defeating opponent for space and nutrition are largely considered responsible for its success against plant pathogenic fungi (Viterbo et al., 2002; Benítez et al., 2004). The root colonization and intimate association of Trichoderma spp. with plant roots are known to promote plant growth and boost immune response against a number of plant pathogens (Contreras-Cornejo et al., 2011; Brotman et al., 2012; Mukherjee, 2012). Biocontrol strains of Trichoderma are used worldwide for the management of various plant pathogens like vascular wilt caused Fusarium (Al-Ani et al., 2013), Botrytis blight or gray mold caused by Botrytis (Elad and Kapat, 1999), anthracnose caused by Colletotrichum spp., and several other plant fungal diseases (Sharma et al., 2016a,b, 2017a). The improvement of Trichoderma species as BCAs for various agricultural applications required, detailed understanding of its active biological repertoire involved in mycoparasitism antibiosis as well as others components (Table 1). Genome sequencing and its annotation in mycoparasitic species have depicted genome sizes of 38.8 and 36.1 Mb for T. viride and of T. atroviride for biocontrol strains, respectively, compared to 34 Mb of T. reesei an industrial strain. Annotation of complete genome depicted a gene pool of 11,800 genes for T. atroviride and 12,400 genes for T. viride, compared to 9,143 genes in saprophytic strain T. reesei. The abundance of gene pool in mycoparasitic strains of Trichoderma genome (Lin et al., 2012; Atanasova et al., 2013) and expression of over 60% of the encoding transcripts during interaction of T. virens and T. atroviride against Rhizoctonia solani have revealed a complex nature of biocontrol mechanisms (Atanasova et al., 2013). Liu and Yang (2005) using simulated mycoparasitic conditions and cDNA libraries identified a total of 3,298 expressed sequence tags (ESTs) which corresponds to 1,740 transcripts. Using inducible conditions for T. harzianum CECT 2413, Vizcaíno et al. (2006) characterized, nearly 8,710 ESTs whereas Yao et al. (2013) identified 1,386 ESTs for T. harzianum 88. Among different ESTs, significant differential expression was observed only for limited transcripts. These EST represents a fragment of mRNA have several biotechnological applications and are being explored for either complementing the sequenced genome projects or cost effective alternatives for identification of genes as well as elucidation of functional genomics of plant–microbe interactions (Vieira et al., 2013). Advancement in molecular tools such as transcriptome profiling using RNA-seq and quantitative real-time PCR (RT-qPCR) technologies also predicted a large number of genes (14,095) for T. harzianum during augmentation on plant pathogen such as Sclerotinia sclerotiorum cell wall and only 297 were found differentially expressed among them (Steindorff et al., 2012, 2014). In addition to plant diseases management potential of biocontrol strains of Trichoderma, its growth promotion abilities in plants have been identified which are significantly enhanced during their antagonistic interactions with pathogens in soil. The molecular action of its biocontrol arsenal is mediated through adaptive recruitment and reprogramming of unique reservoir of several transcripts (Shaw et al., 2016). A comparative account using bioinformatic approaches such as BLAST analysis has revealed a very low overlap for different ESTs libraries (Yao et al., 2013). Therefore, the microarrays set designed based on genome coverage and ESTs may not provide accurate information.

TABLE 1. List of a few selected glycosyl hydrolases, secondary metabolites, and different transcripts of biocontrol strains/species of Trichoderma characterized for their role in biocontrol.
The comprehensive analyses using different molecular approaches including ESTs (Chambergo et al., 2002), subtractive hybridization (Carpenter et al., 2005; Scherm et al., 2009; Vieira et al., 2013), microarray (Chambergo et al., 2002; Breakspear and Momany, 2007; Samolski et al., 2009), and transcriptomes (Atanasova et al., 2013) have established the complex response of Trichoderma species in biocontrol process which induces numerous genes having morphogenetic or other functions as well (Mehrabi-Koushki et al., 2012; Puglisi et al., 2012; Cacciola et al., 2015; Cetz-Chel et al., 2016). The complexity in different attributes may not be related to a particular stress and hence can lead to either imperfect transcriptional representation or a complex response. The continuous development in molecular technologies and advent of cloning free libraries using genome sequencing, deep RNA sequencing and proteomics has played vital role in the accurate identification and enhancing our capabilities of cataloging mRNA and protein populations exclusive to Trichoderma strains in response to changing environmental conditions (Shentu et al., 2014; Xie et al., 2015; Schmoll et al., 2016).
The Trichoderma–plant–pathogen interaction can produce significant amount of noise. Therefore one can speculate that the substantial amount of response at the gene expression level represents noise and that only a few changes are adaptive. Also, the microbes in the environment are continuously subjected to challenges and respond simultaneously to these factors in a complex way. Understanding the regulatory interactions necessitates an approach that can encompasses simultaneous both the transcriptome and proteome to observe and systematically view the adaptive expression at RNA and protein level. The integrated studies based on translatome and proteome level can provide a better state of these adaptive responses during biocontrol interaction. The regulation of mRNA at transcriptional and post-transcriptional levels contributes to reprogramming the behavior of BCAs through protein and secondary bioactive metabolites secretion to counter the pathogen associated challenges.
So far studies on Trichoderma have been conducted extensively using ESTs and transcriptome approach revealed the expression of several genes related to mycoparasitism of BCAs directly (Reithner et al., 2011; Sharma et al., 2017b) or indirectly through the modulation of host transcriptome (Morán-Diez et al., 2012; Perazzolli et al., 2012). In our previous studies, attempts were made to identify the role of different transcripts related to lytic enzymes, transporter system, and other gene related to metabolites of T. harzianum (Sharma et al., 2016a,b, 2017b) and characterization of extracellular proteins from T. saturnisporum (Sharma and Shanmugam, 2012) using autoclaved mycelium of different plant pathogenic fungi. These studies revealed only a limited number of proteins compared to transcripts. The approaches used for cDNA cloning and other array technologies have also created artifacts in accurate identification of candidate transcripts. Therefore, the integrated translatome and proteome based studies can help in a better and accurate depiction of key regulators involved in Trichoderma–plant–pathogen interaction (Figure 1). Recent studies showed that the gene expression of mycoparasitic T. harzianum and T. atroviride strains changes not only to plant-pathogenic fungi (Sharma et al., 2016a, 2017b) but also to itself (Reithner et al., 2011). Thus translational response is a key determinant contributing to adaptation under such interaction stress (Picard et al., 2013). Therefore, present review emphasizes the role of translatome based approach in accurate determination of active mRNA population in a complex dialog coupled to proteome data in a three way interaction of Trichoderma–plant–pathogen.
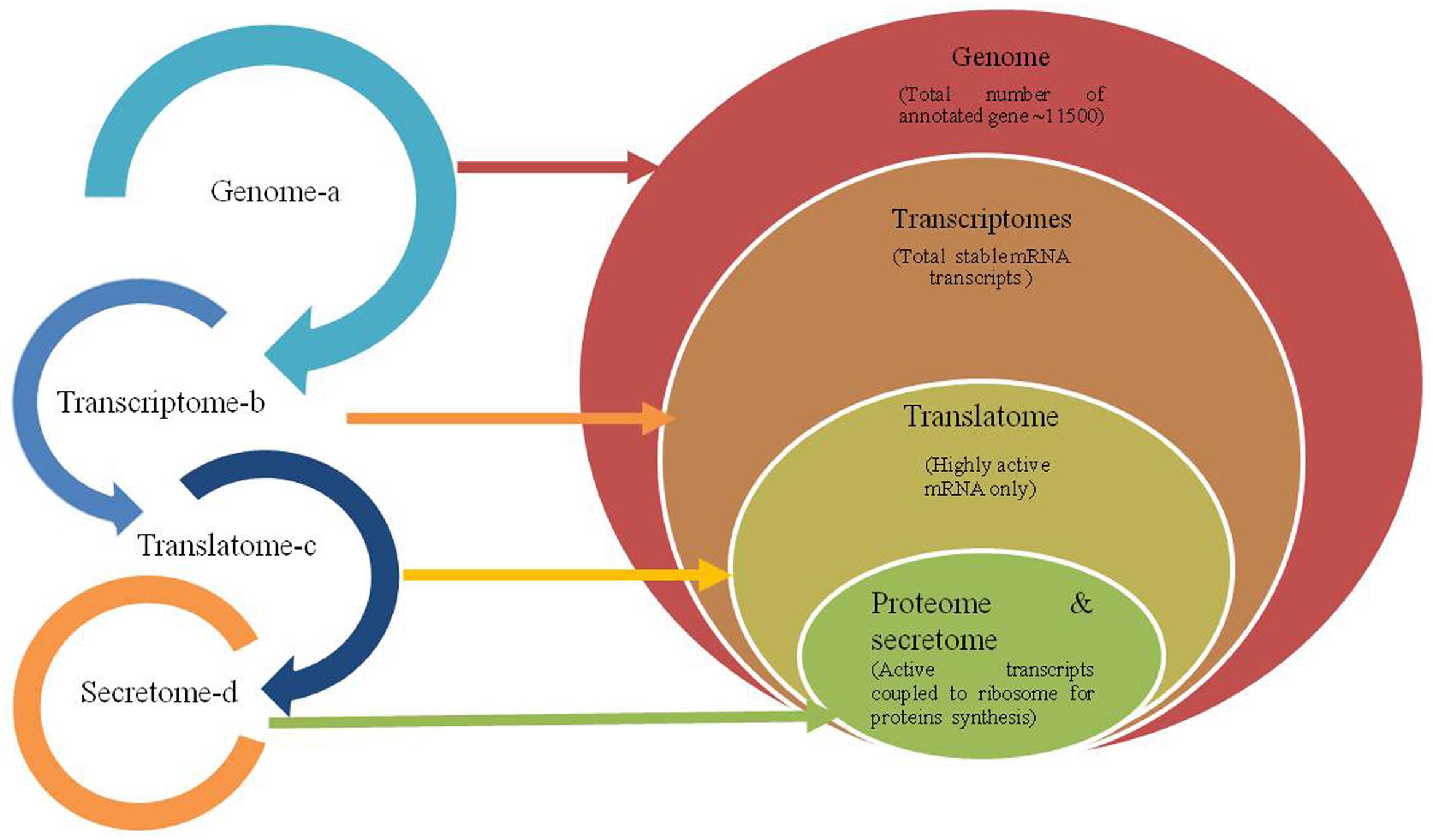
FIGURE 1. Schematic flow of genetic information from genome to proteome level. A- Relationship of different omic approaches in Trichoderma–plant–plant pathogen interaction. B- Hierarchal relationship of total gene predicted using stacked ven diagram in the genome to the total mRNA transcribed (transcriptome) representing both stable and highly active mRNAs population under a stress response. Here depending upon the conditions and alternate splicing a > b > c > d or a > b > c = d or rarely a >b = c=d; a > b = c < d.
Mechanisms of Trichoderma
Trichoderma strains are used as BCAs in agriculture largely due to their abilities to directly antagonize plant-pathogenic fungi through the production of hydrolases (Benítez et al., 2004; Gruber and Seidl-Seiboth, 2012), antibiotics (Rubio et al., 2009; Vinale et al., 2014) and their tolerance to toxin produced by plant pathogens (Sharma et al., 2013) (Table 1). The interaction of Trichoderma with host plants reprograms not only the gene expression of biocontrol strains but also of its associated host plant (Harman, 2011; Figure 2). For example, strains of Trichoderma are explored for growth promotion and boosting immune responses, root development, and activation of seed germination or amelioration of abiotic stresses (Harman et al., 2004; Lorito et al., 2010; Shoresh et al., 2010; Hermosa et al., 2012). The immune responses in host plant are primed through systemic resistance (Tucci et al., 2011), involving a complex signaling of jasmonic acid/ethylene-induced systemic resistance and/or salicylic acid-dependent pathways which may behave differently in plant–Trichoderma interactions (Shoresh et al., 2010). The three way interaction between biocontrol, host plant, and pathogen from initial root colonization is known to change both the transcripts and proteome of host plants (Alfano et al., 2007; Segarra et al., 2007; Shoresh and Harman, 2008; Palmieri et al., 2012; Gomes et al., 2017; Martínez-Medina et al., 2017a,b; Pelagio-Flores et al., 2017). The availability of microarrays, next generation DNA sequencing, RNA-seq, and genome annotation have provided a global insight into the transcriptome response of plant–Trichoderma and Trichoderma–plant pathogen interaction.
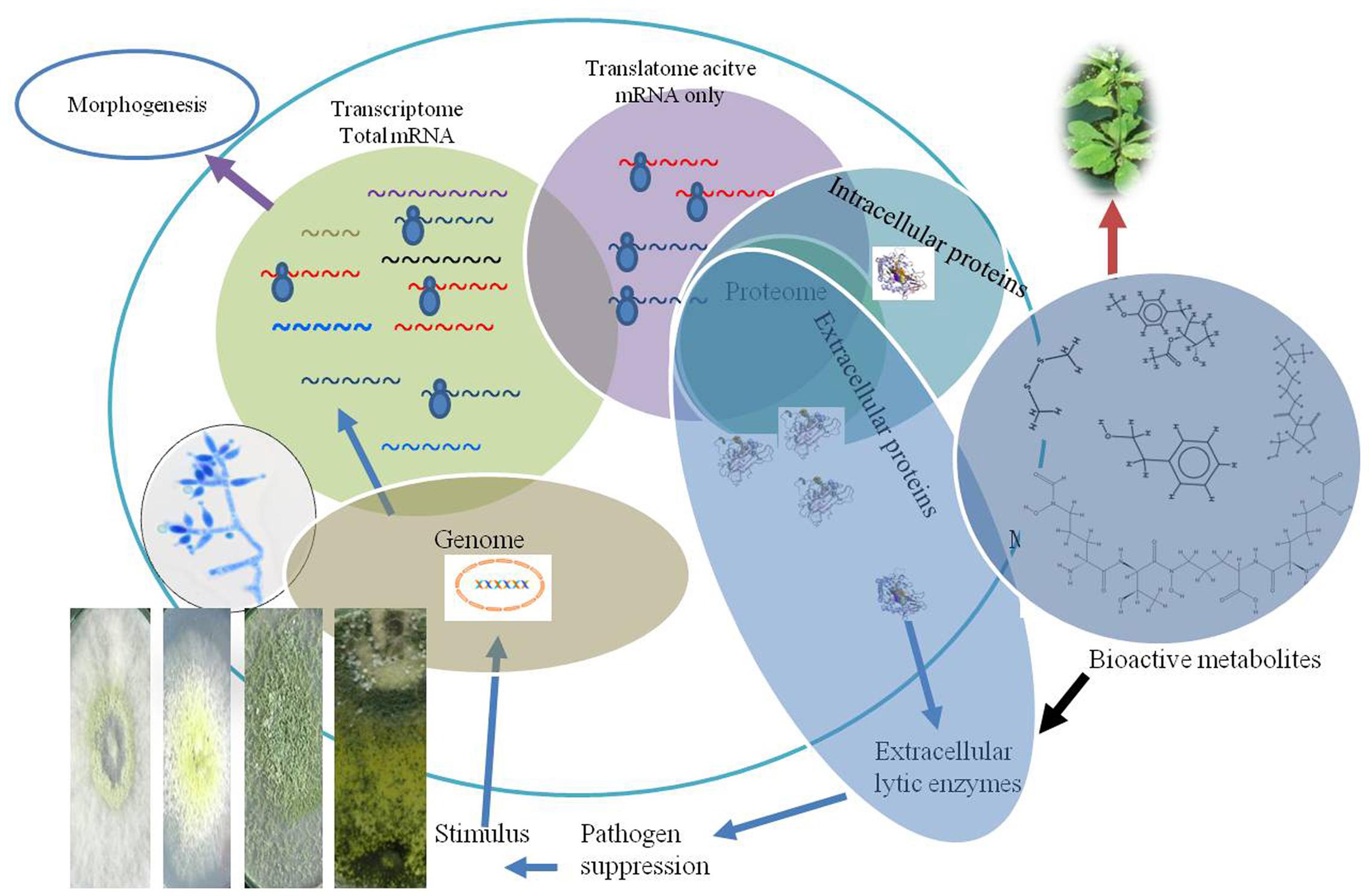
FIGURE 2. Pictorial representation of total mRNA transcripts, active mRNA involved in protein and bioactive metabolites synthesis during interaction with plant or plant pathogens. The figure explains that transcriptome based study in general represents a higher level of mRNA, compared to total translatome which represents only ribosomal loaded active mRNAs during interaction.
Omics Approaches in Uncoupling Genome and Transcriptome Profile
The characterization of candidate transcripts involved in various biological functions using transcriptome is one of the best approach. In comparison to stable nature of the genome, transcriptome is more dynamic and vary in response to different stimuli. The massive transcriptome response to various factors can be tentatively identified, quantified, and correlated to a biological process using ESTs, subtractive libraries, and DNA microarrays (Herrera-Estrella, 2014). A number of studies have been done at genome-wide and transcriptional level to understand the molecular behavior of different Trichoderma strains under contrasting conditions ranging from mycoparasitism of plant pathogens to imparting direct beneficial aspects to plants under stress conditions (Arvas et al., 2006; Vizcaíno et al., 2007; Seidl et al., 2009). The transcriptome analysis of T. atroviride IMI206040 at different stages of interaction with R. solani identified 7,797 out of 11,863 estimated genes which represented over 65% of total gene of the organism genome whereas only 1.47% of total gene (175) transcripts were found significantly differentially expressed in mycoparasitic interactions. The differentially expressed transcripts were also investigated during pathogenic attack on Phytophthora capsici, Botrytis cinerea, and R. solani (Reithner et al., 2011). In comparison to a large number of transcribed genes predicted for T. atroviride based on genomic data, only 38.4% of genes involved in interaction with R. solani, were expressed before contact whereas 52.8% were found responsible for Trichoderma confrontation with itself (Reithner et al., 2011).
The use of EST (Vizcaíno et al., 2006, 2007), subtractive cDNA libraries and DNA array (Rosales-Saavedra et al., 2006; Alfano et al., 2007; Mathys et al., 2012) based studies carried under environmental conditions have helped dramatically to the global-scale identification of active genes of Trichoderma which are not directly linked to plant pathogens but are required for colonization and imparting other beneficial aspects to the host plant. For example, hydrophobins, aspartyl proteases, expansin-like protein of Trichoderma origin have been explored for their involvement in the mycoparasitism mediated biocontrol of these microbes (Brotman et al., 2008; Samolski et al., 2009). Subsequently, the sequencing of complete genome and high-throughput transcriptome using 454 sequencing (Barakat et al., 2009) has enhanced our understanding on investigation of mechanisms at global cellular level under different conditions in better way (Reithner et al., 2011). The transcriptome based approach is far more robust, dynamic, and refined technique compared to genome sequencing which is stable as described below.
Trichoderma Genome Organization
Since the genome sequencing of T. reesei industrial strain nine years back (Martinez et al., 2008), presently the genome of a number of strains representing T. virens, T. harzianum, T. atroviride, T. asperellum, Trichoderma longibrachiatum, and Trichoderma citrinoviride have been sequenced and revised (http://genome.jgi.doe.gov/). A comparative account of genome revealed presence of seven chromosomes in industrial strain T. reesei (Carter et al., 1992; Mantyla et al., 1992; Herrera-Estrella et al., 1993) whereas six chromosomes in biocontrol strains T. harzianum and T. viride (Gómez et al., 1997; Martinez et al., 2008). The genomic annotation of T. virens, T. atroviride, and T. reesei also unveiled lack of transposons and remarkable similarity of genes up to 78–96% among them. In the genome of T. virens and T. atroviride no true orthodox were reported for 2,756 and 2,510 genes, respectively in other species. The genome of T. virens and T. atroviride share 1,273 exclusive orthologs and 26 expanded families which were missing in T. reesei genome that may be a probable answer to mycoparasitic nature of T. atroviride and T. virens (Kubicek et al., 2011; Herrera-Estrella, 2014). A comparative study of genome organization of two Trichoderma species has revealed the expansion of considerable expansion genes involved in mycoparasitic T. virens strain which are missing in T. reesei (Kubicek et al., 2011).
Transcriptome
The development of modern sophisticated omics technologies has played a vital role in developing better system-level understanding of gene expression. In particular, transcriptome based studies have proved a yardstick in the investigation of global cellular mechanisms and identification of several key genes involved in mycoparasitism and imparting other benefits to the host by Trichoderma strains. The measurement of the entire set of RNAs through transcriptome coupled with DNA microarrays or high-throughput RNA sequencing is a reliable and reproducible tool for wide analysis of transcripts. A number of transcriptome studies have been done on Trichoderma–plant–pathogen interaction (Marra et al., 2006; Chacon et al., 2007; Samolski et al., 2009; Mehrabi-Koushki et al., 2012; Rubio et al., 2012).
Stating from initial use of EST for the determination of glucose metabolism in T. reesei (Chambergo et al., 2002) and TrichoEST project (Vizcaíno et al., 2006), ESTs based studies have been done in T. harzianum (Liu and Yang, 2005; Vizcaíno et al., 2006; Suárez et al., 2007; Yao et al., 2013), T. atroviride, T. asperellum (Vizcaíno et al., 2007; Liu et al., 2010), T. virens (Vizcaíno et al., 2007; Morán-Diez et al., 2010), Trichoderma aggressivum, T. viride, and T. longibrachiatum (Vizcaíno et al., 2007) for the identification of transcripts induced during mycoparasitism and other environmental conditions. From a total of unique sequences (3,478), in T. harzianum CECT2413, 23% were found related to secretory chitinases, glucanases, and proteases. A large number of transcripts expressed (9478 ESTs containing 2,734 unique sequences) during the early interaction of T. atroviride with B. cinerea and R. solani were identified (Seidl et al., 2009) whereas 66 genes covering 442 ESTs were induced under mycoparasitic interaction (Herrera-Estrella, 2014).
Similarly, the analysis of transcriptomics changes in T. harzianum, T. virens, and T. hamatum during interactions with tomato plants revealed expression of 1,077 genes and only six of them being common to all three. The majority of genes encoding enzymes belong to chitin degradation during early interactions with tomato plants whereas genes encoding other secreted proteins were likely to involve in the signaling between Trichoderma and plants. Transcriptome based studies have led to the identification of new candidate genes having role in redox reaction, possible elicitors, transporters (Sharma et al., 2017b), lipid metabolism and detoxification (Chacon et al., 2007; Sharma et al., 2013), small secreted proteins (Ruocco et al., 2009; Samolski et al., 2009; Rubio et al., 2014). The de novo sequencing of T. atroviride IMI206040 transcriptome obtained during mycoparasitic interaction in presence of plant-pathogenic fungus R. solani revealed thousands of high-quality reads. An account of transcripts expressed during interaction to the total number of genes predicted in the genome of T. atroviride revealed that almost 45% were induced during interaction with R. solani and only 175 of them were host responsive (Reithner et al., 2011; Gupta et al., 2016).
Microarray analysis of T. harzianum T34 strain interaction with Arabidopsis identified approximately 24,000 transcripts of the host plant which were modulated by the BCA. The significance and global impact of this beneficial microbe in reprogramming the molecular physiology of host plant to stress responses through the regulation of transcription, signal transduction pathways has been reported in different studies (Morán-Diez et al., 2012; Lamdan et al., 2015). Further host specific response of Trichoderma strain with plants representing monocot and dicot hosts under the same conditions have also been explored to identify signature transcriptome repertoires and answer the widely prevalent questions of specificity of responses and role of secreted proteins in mutualistic interaction, root colonization, and induction of immune responses (Morán-Diez et al., 2015; Ho et al., 2016; Sharma et al., 2017b). These studies indicate the limitations of transcriptome based studies in precise estimation of ribosome loaded active mRNA population involved in complex mycoparasitic behavior of Trichoderma species as BCAs.
Translatomes
The mRNA and protein levels do not perfectly correlate in native or engineered systems (Tian et al., 2004; Jayapal et al., 2008; Vogel and Marcotte, 2012; Payne, 2015). The post-transcriptional regulation of transcripts is a complex process and may not be compared with transcription level regulation of genes. Therefore, the post-transcriptional regulation is of great significance for better characterization of functional role of genes (Picard et al., 2013). Although ESTs and transcriptome based experimental studies have provided valuable information in mining genes incited by various stress responses in Trichoderma interaction with plants and plant pathogens, its application is limited because the levels of the proteins and their encoding mRNA are not correlated to each other. Therefore considering the use of the cutoff standards in transcriptome based studies and appearance of artifacts in the differential expression of genes, translatome based studies offers potential choice and a better alternative involving only active mRNA populations (Picard et al., 2013; Yanguez et al., 2013; Piccirillo Ciriaco et al., 2014; King and Gerber, 2016; Meteignier et al., 2017).
Studies involving translational regulation of gene expression are emerging as a prominent tool for the understanding the regulation of protein abundance in adaptive responses of the host (Halbeisen and Gerber, 2009; Spriggs et al., 2010). In the genetic flow of information, the translational regulation reprograms the cell activities by protein synthesis. In last decade due to rapid advancements in technology, efforts on understanding the modulatory role of translation in gene expression have increased significantly. The translatome referring to the active mRNAs population associated with ribosomes has facilitated the removal of background noise and useful for the accurate determination of active mRNA. Originally used in oocytes and embryos (Terman, 1970; Gurdon et al., 1971), translational control has emerged as a key point of eukaryotes. The process is executed by loading of ribosomes on mRNA followed by translation elongation (Groppo and Richter, 2009; Jackson et al., 2010). Since, the translatome based studies are focused only on the pools of genome-wide translated mRNA and therefore have helped in identification of key regulatory factors that are under translational control (Zupanic et al., 2013). This technique offers immense potential in the targeting key regulators which are active during interaction and play important role for the host plant in combating various stress responses. Translatome studies also help in determination of the ribosome number on active mRNA molecule in response to stress in the cellular genes (Koritzinsky and Wouters, 2007; Thomas and Johannes, 2007; Picard et al., 2013; Figure 3).
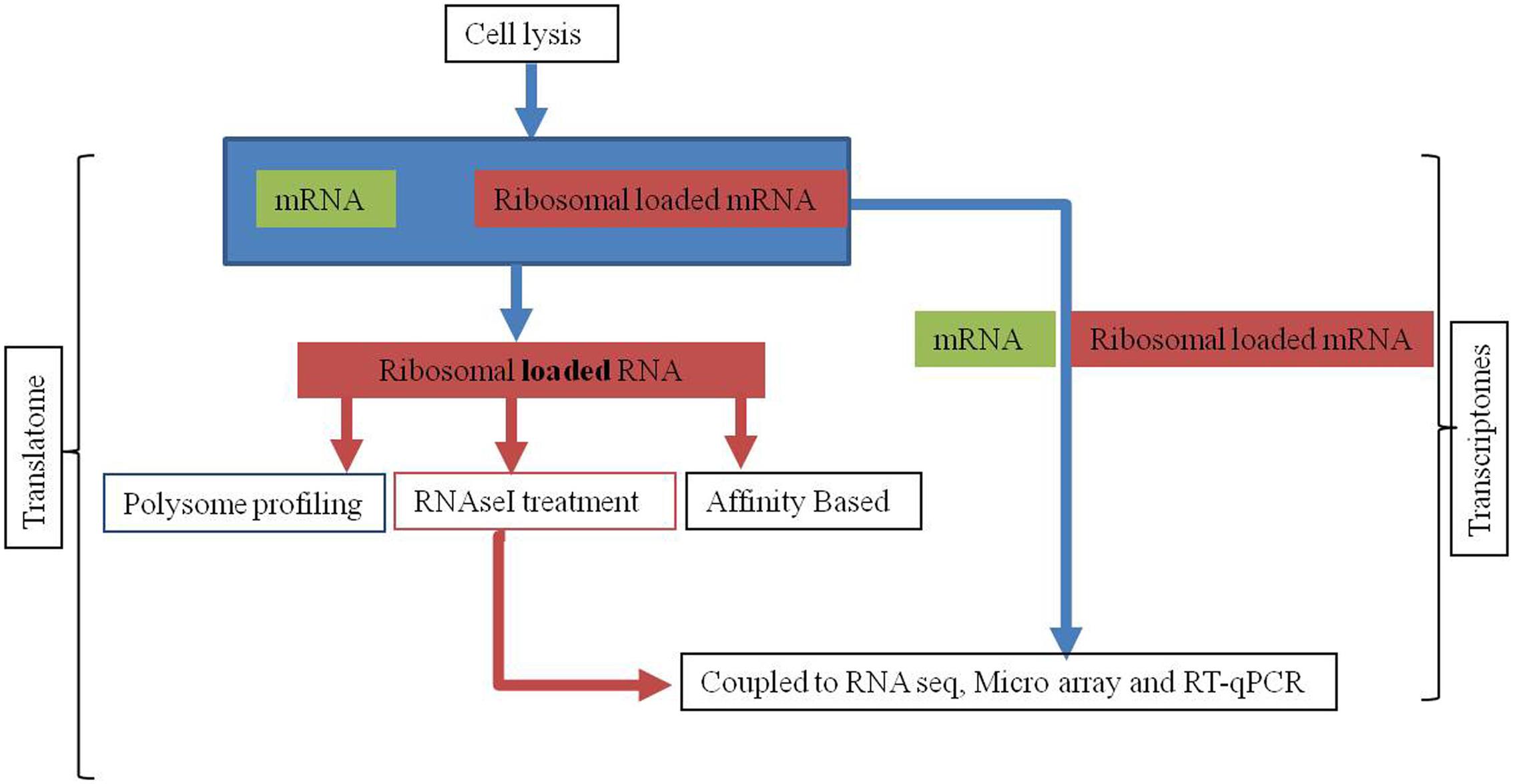
FIGURE 3. The experimental procedure of transcriptome and translatome for genome wide studies. In translatome ribosomal loaded or protected fragments of RNA are size or affinity fractionated, recovered, and ligated to adaptors for reverse transcription, amplification, and high-throughput RNA-seq whereas in transcriptome complete mRNA is used for subsequent analysis.
Presently, there are three methods used for translatome analysis; (a) polysomal profiling, (b) ribosomal profiling, and (c) ribosome affinity purification (RAP) (Figure 3). Polysomal profiling discovered in 1960s involves the separation of actively translated mRNAs bound by several ribosomes from free RNA by sucrose gradient centrifugation and then mRNAs can be coupled to northern blot or RT-qPCR or cDNA microarrays, or RNA-seq on a global level (Karginov and Hannon, 2013; Spangenberg et al., 2013). The second method known as ribosomal profiling was developed by Weissman group in Saccharomyces cerevisiae, determines the location of ribosomes at codon or nucleotide scale (Ingolia et al., 2009). The advantage of this technique is acquisition of information at global scale with respect to the position of the ribosomes on translated mRNA.
The deep nucleotide sequencing of ribosome protected RNA fragments obtained after RNase I treatment of cell lysate helps in accurate determination of ribosome position and its densities along RNA (Ingolia et al., 2012). Both polysome and ribosome based profiling need relatively large sample size to obtain enough RNA for microarray/RNA-seq analysis. The third method known as RAP developed by Inada et al. (2002) in S. cerevisiae capture monosomes and polysomes by using antiFLAG affinity resin. The RAP also known as translating RAP provides a better approximation of the translated mRNA population if coupled with transcriptome analysis (Halbeisen and Gerber, 2009; Jiao and Meyerowitz, 2010).
Integrating Translatome and Proteomic Study
The post-transcriptional events such as translation regulation and protein stability are the principle causes of weak correlations and variations in proteomic, transcriptomic, and genomic data. The associated errors in transcriptome analysis are subjected to arise from the suppression by microarrays which can further impede the identification of active candidate transcripts. On the other side, methods opted for protein staining, limitations associated in visualizing low-abundant and co-migrating proteins seriously hampers proteomic based study. The recent developments in proteomics methods such as use of mass spectrometric (MS) and liquid chromatography (LC) techniques have made quantitative proteomic profiling, currently a driving force for identification of proteins. The highly stable and reproducible performance of mass spectrometers such as Q Exactive hybrid quadrupole-Orbitrap mass spectrometer MS and Triple TOF 5600 MS is capable of identification of both proteomics (Chang et al., 2014) and characterization of bioactive metabolites. Integrated analyses of active mRNAs coupled with protein expression are available for bacteria, yeast, mice, and humans. Similar to transcriptome, the translatome based studies are focused only on transcripts level which are intracellular in nature. The coupling of multiomic approaches based on active mRNA, proteomes, and protein turnover of both intra as well extracellular proteins and biologically active metabolites under different environmental conditions will provide a better answer of reprogramming biocontrol to various plant beneficial attribute and its resiliencies to combat different environmental conditions (Figure 2).
Conclusion
The availability of the fully sequenced genomes of Trichoderma spp. has accelerated our research on understanding of the behavior of different species of this genus and how the information on their gene pool determines their capabilities and limitations. The genomes of Trichoderma which is known to contain thousands of genes encoding different glycosyl hydrolases, secondary metabolites, antibiotics, lectins with insecticidal properties, and transporters with potential in bioremediation involved in antibiotics biosynthesis, and several other candidate genes (Druzhinina et al., 2012; Atanasova et al., 2013). Exploration of genes and their encoding proteins involved in developing tolerance against various stresses such as cold, below-average precipitation, salty conditions, pH, herbicide resistance as well biotic factor are an active field of research. The predicted genome of Trichoderma strains are known to encode a large number genes therefore coupling of translatome studies with proteomics of both extracellular and intracellular proteins offers a wide scope for better understanding the complex behaviors of Trichoderma as BCA.
The genomic comparison of mycoparasitic species of T. harzianum with non-mycoparasitic strains of T. reesei already provides evidences of the expansion of several genes in biocontrol strains. The secretion of a large number of cell wall targeting enzymes and bioactive secondary metabolites require adaptive molecular reprogramming of Trichoderma transcriptome. The variation at genomic, transcriptomics, and proteomic levels is a challenging task and difficult to correlate due to complex and non-systematic post-transcriptional and limitation of proteomic techniques. Further, the translational control is a widespread phenomenon with intense effect; nevertheless it is underestimated for its regulatory roles. In general, extensive uncoupling of both RNA movements and inferred cell activities has been observed for 19 different transcriptome and translatome. Therefore, coupled quantitative transcript and protein abundance studies can serve as a gold standard for proper and accurate depiction of interaction involving Trichoderma–plant–plant pathogens. Although detecting changes in the transcriptome level (total mRNAs), translatome level (ribosome loaded mRNAs) and the proteome is experimentally feasible in a high-throughput way, the integration of these omic technologies is still far away. Systematic global analyses aims at integrating transcriptome, translatome, and proteome level can provide accurate view of widespread adaptive mechanisms of interaction between Trichoderma–plant–pathogen.
In future, integrated efforts will help us to better understand, identify, and then explore the molecular behavior of Trichoderma arsenal involved in its success as BCAs as well as industrial sectors. In such instances, the integration of the translatome using ribosomal profiling and coupling it with proteomic approaches such as liquid chromatography-tandem mass spectrometry (LC-MS/MS) for both extracellular and intracellular proteins offers a lot of scope for accurate characterization of active molecular components involved in biocontrol and then subsequently their utilization of various applications.
Future Directions
A comparative multiomic coupled insights of Trichoderma–plant–plant pathogens in three way interaction will play vital role in accurate characterization of transcripts responsible for cosmopolitan nature of Trichoderma and then targeting the promising one for agricultural based applications. The latest advancements and complete genome sequencing have already provided a platform of gene pool. Further integration with latest functional techniques such as translatome will lead another step close to identification of targets in the form of active transcripts involved in a complex interaction of plant–BCA–plant pathogens.
Author Contributions
VS and RS prepared the manuscript. PS and AG edited the manuscript.
Conflict of Interest Statement
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Acknowledgment
The authors are thankful to SERB, Department of Science and Technology-New Delhi India for providing funding under DST-FAST Track young scientist scheme (award letter NO. SB/FT/LS-365/2012).
References
Al-Ani, L., Salleh, B., and Ghazali, A. H. A. (2013). “Biocontrol of Fusarium wilt of banana by Trichoderma spp,” in Proceedings of the Conference Paper International Symposium on Tropical Fungi (ISTF) IPB International Convention Center, Bogor.
Alfano, G., Ivey, M. L. L., Cakir, C., Bos, J. I. B., Miller, S. A., Madden, L. V., et al. (2007). Systemic modulation of gene expression in tomato by Trichoderma hamatum 382. Phytopathology 97, 429–437. doi: 10.1094/PHYTO-97-4-0429
Anke, H., Kinn, J., Bergquist, K.-E., and Sterner, O. (1991). Production of siderophores by strains of the genus Trichoderma, isolation and characterization of the new lipophilic coprogen derivative, palmitoylcoprogen. Biol. Met. 4, 176–180. doi: 10.1007/BF01141311
Arvas, M., Pakula, T., Lanthaler, K., Saloheimo, M., Valkonen, M., Suortti, T., et al. (2006). Common features and interesting differences in transcriptional responses to secretion stress in the fungi Trichoderma reesei and Saccharomyces cerevisiae. BMC Genomics 7:32. doi: 10.1186/1471-2164-7-32
Atanasova, L., Crom, S. L., Gruber, S., Coulpier, F., Seidl-Seiboth, V., Kubicek, C. P., et al. (2013). Comparative transcriptomics reveals different strategies of Trichoderma mycoparasitism. BMC Genomics 14:121. doi: 10.1186/1471-2164-14-121
Bae, S., Kumar, T., Young, J., Ryu, M., Park, G., Shim, S., et al. (2016). Trichoderma metabolites as biological control agents against Phytophthora pathogens. Biol. Control 92, 128–138. doi: 10.1111/j.1472-765X.2009.02599.x
Barakat, A., DiLoreto, D. S., Zhang, Y., Smith, C., Baier, K., Powell, W. A., et al. (2009). Comparison of the transcriptomes of American chestnut (Castanea dentata) and Chinese chestnut (Castanea mollissima) in response to the chestnut blight infection. BMC Plant Biol. 9:51. doi: 10.1186/1471-2229-9-51
Benítez, T., Rincón, A. M., Limón, M. C., and Codón, A. C. (2004). Biocontrol mechanisms of Trichoderma strains. Int. Microbiol. 7, 249–260.
Breakspear, A., and Momany, M. (2007). The first fifty microarray studies in filamentous fungi. Microbiology 153, 7–15. doi: 10.1099/mic.0.2006/002592-0
Brotman, Y., Briff, E., Viterbo, A., and Chet, I. (2008). Role of Swollenin, an expansin-like protein from Trichoderma, in plant root colonization. Plant Physiol. 147, 779–789. doi: 10.1104/pp.108.116293
Brotman, Y., Lisec, J., Méret, M., Chet, I., Willmitzer, L., and Viterbo, A. (2012). Transcript and metabolite analysis of the Trichoderma-induced systemic resistance response to Pseudomonas syringae in Arabidopsis thaliana. Microbiology 158, 139–46. doi: 10.1099/mic.0.052621-0
Cacciola, S. O., Puglisi, I., Faedda, R., Sanzaro, V., Pane, A., Lo Piero, A. R., et al. (2015). Cadmium induces cadmium-tolerant gene expression in the filamentous fungus Trichoderma harzianum. Mol. Biol. Rep. 42, 1559–1570. doi: 10.1007/s11033-015-3924-4
Carpenter, M. A., Stewart, A., and Ridgway, H. J. (2005). Identification of novel Trichoderma hamatum genes expressed during mycoparasitism using subtractive hybridisation. FEMS Microbiol. Lett. 251, 105–112. doi: 10.1016/j.femsle.2005.07.035
Carter, G. L., Allison, D., Rey, M. W., and Dunn-Coleman, N. S. (1992). Chromosomal and genetic analysis of the electrophoretic karyotype of Trichoderma reesei: mapping of the cellulase and xylanase genes. Mol. Microbiol. 6, 2167–2174. doi: 10.1111/j.1365-2958.1992.tb01390.x
Cetz-Chel, J. E., Balcázar-López, E., Esquivel-Naranjo, E. U., and Herrera-Estrella, A. (2016). The Trichoderma atroviride putative transcription factor Blu7 controls light responsiveness and tolerance. BMC Genomics 17:327. doi: 10.1186/s12864-016-2639-9
Chacon, M. R., Rodriguez-Galan, O., Benitez, T., Sousa, S., Rey, M., Llobell, A., et al. (2007). Microscopic and transcriptome analyses of early colonization of tomato roots by Trichoderma harzianum. Int. Microbiol. 10, 19–27.
Chambergo, F. S., Bonaccorsi, E. D., Ferreira, A. J., Ramos, A. S., Ferreira, J. R. J. J. R., Abrahao-Neto, J., et al. (2002). Elucidation of the metabolic fate of glucose in the filamentous fungus Trichoderma reesei using expressed sequence tag (EST) analysis and cDNA microarrays. J. Biol. Chem. 277, 13983–13988. doi: 10.1074/jbc.M107651200
Chang, C., Li, L., Zhang, C., Wu, S., Guo, K., Zi, J., et al. (2014). Systematic analyses of the transcriptome, translatome, and proteome provide a global view and potential strategy for the C-HPP. J. Proteome Res. 13, 38–49. doi: 10.1021/pr4009018
Chen, J., Sun, S., Miao, C., Wu, K., Chen, Y., Xu, L., et al. (2016). Endophytic Trichoderma gamsii YIM PH30019: a promising biocontrol agent with hyperosmolar, mycoparasitism, and antagonistic activities of induced volatile organic compounds on root-rot pathogenic fungi of Panax notoginseng. J. Ginseng Res. 40, 315–324. doi: 10.1016/j.jgr.2015.09.006
Cloyd, R. A., Dickinson, A., and Kemp, K. E. (2007). Effect of diatomaceous earth and Trichoderma harzianum T-22 (Rifai strain KLR-AG2) on the fungus gnat Bradysia sp. nr. coprophila (Diptera: Sciaridae). J. Econ. Entomol. 100, 1353–1359. doi: 10.1093/jee/100.4.1353
Contreras-Cornejo, H. A., Macías-Rodríguez, L., Beltrán-Peña, E., Herrera-Estrella, A., and López-Bucio, J. (2011). Trichoderma-induced plant immunity likely involves both hormonal- and camalexin-dependent mechanisms in Arabidopsis thaliana and confers resistance against necrotrophic fungus Botrytis cinerea. Plant Signal. Behav. 6, 1554–1563. doi: 10.4161/psb.6.10.17443
de Azevedo, A. M. C., De Marco, J. L., and Felix, C. R. (2000). Characterization of an amylase produced by a Trichoderma harzianum isolate with antagonistic activity against Crinipellis perniciosa, the causal agent of witches’ broom of cocoa. FEMS Microbiol. Lett. 188, 171–175. doi: 10.1016/S0378-1097(00)00231-7
De la Cruz, J., and Llobell, A. (1999). Purification and properties of a basic endo-β-1,6-glucanase (BGN16.1) from the antagonistic fungus Trichoderma harzianum. FEBS J. 265, 145–151. doi: 10.1046/j.1432-1327.1999.00698.x
De la Cruz, J., Pintor-Toro, J. A., Benítez, T., and Llobell, A. (1995). Purification and characterization of an endo- β -1,6-glucanase from Trichoderma harzianum that is related to its mycoparasitism. J. Bacteriol. 177, 1864–1871. doi: 10.1128/jb.177.7.1864-1871.1995
de las Mercedes Dana, M., Pintor-Toro, J. A., and Cubero, B. (2006). Transgenic tobacco plants overexpressing chitinases of fungal origin show enhanced resistance to biotic and abiotic stress agents. Plant Physiol. 142, 722–730. doi: 10.1104/pp.106.086140
De Marco, J. L., and Felix, C. R. (2002). Characterization of a protease produced by a Trichoderma harzianum isolate which controls cocoa plant witches’ broom disease. BMC Biochem. 3:3.
Diánez Martínez, F., Santos, M., Carretero, F., and Marín, F. (2016). Trichoderma saturnisporum, a new biological control agent. J. Sci. Food Agric. 96, 1934–1944. doi: 10.1002/jsfa.7301
Djonović, S., Pozo Maria, J., and Kenerley, C. M. (2006). Tvbgn3, a β-1, 6-Glucanase from the biocontrol fungus Trichoderma virens, is involved in mycoparasitism and control of Pythium ultimum. Appl. Microbiol. Biotechnol. 72, 7661–7670.
Dou, K., Wang, Z., Zhang, R., Wang, N., Fan, H., Diao, G., et al. (2014). Cloning and characteristic analysis of a novel aspartic protease gene Asp55 from Trichoderma asperellum ACCC30536. Microbiol. Res. 169, 915–923. doi: 10.1016/j.micres.2014.04.006
Druzhinina, I. S., Shelest, E., and Kubicek, C. P. (2012). Novel traits of Trichoderma predicted through the analysis of its secretome. FEMS Microbiol. Lett. 337, 1–9. doi: 10.1111/j.1574-6968.2012.02665.x
Elad, Y., and Kapat, A. (1999). The role of Trichoderma harzianum protease in the biocontrol of Botrytis cinerea. Eur. J. Plant Pathol. 105, 177–189. doi: 10.1128/AEM.02486-10
Espino-rammer, L., Ribitsch, D., Przylucka, A., Marold, A., Greimel, K. J., and Acero, H. (2013). Two novel class II hydrophobins from Trichoderma spp. stimulate enzymatic hydrolysis of poly(ethylene terephthalate) when expressed as fusion proteins. Appl. Environ. Microbiol. 79, 4230–4238. doi: 10.1128/AEM.01132-13
Faize, M., Malnoy, M., Dupuis, F., Chevalier, M., Parisi, L., and Chevreau, E. (2003). Chitinases of Trichoderma atroviride Induce scab resistance and some metabolic changes in two cultivars of apple. Genet. Resist. 93, 1496–1504. doi: 10.1094/PHYTO.2003.93.12.1496
Garnica-vergara, A., Barrera-ortiz, S., Mu, E., and Raya-gonz, J. (2015). The volatile 6-pentyl-2H-pyran-2-one from Trichoderma atroviride regulates Arabidopsis thaliana root morphogenesis via auxin signaling and ethylene insensitive 2 functioning. New Phytol. 209, 1469–1512. doi: 10.1111/nph.13725
Garo, E., Starks, C. M., Jensen, P. R., Fenical, W., Lobkovsky, E., and Clardy, J. (2003). Trichodermamides A and B, cytotoxic modified dipeptides from the marine-derived fungus Trichoderma virens. J. Nat. Prod. 66, 423–426. doi: 10.1021/np0204390
Gomes, E. V., Ulhoa, C. J., Cardoza, R. E., Silva, R. N., and Gutiérrez, S. (2017). Involvement of Trichoderma harzianum Epl-1 protein in the regulation of botrytis virulence- and tomato defense-related genes. Front. Plant Sci. 29:880. doi: 10.3389/fpls.2017.00880
Gómez, I., Chet, I., and Herrera-Estrella, A. (1997). Vegetative compatibility and molecular variation among Trichoderma harzianum isolates. Mol. Gen. Genet. 256, 127–135. doi: 10.1007/s004380050554
Groppo, R., and Richter, J. D. (2009). Translational control from head to tail. Curr. Opin. Cell. Biol. 21, 444–451. doi: 10.1016/j.ceb.2009.01.011
Gruber, S., Kubicek, C. P., and Seidl-seiboth, V. (2011). Differential regulation of orthologous chitinase genes in mycoparasitic Trichoderma species. Appl. Environ. Microbiol. 77, 7217–7226. doi: 10.1128/AEM.06027-11
Gruber, S., and Seidl-Seiboth, V. (2012). Self versus non-self: fungal cell wall degradation in Trichoderma. Microbiology 158, 26–34. doi: 10.1099/mic.0.052613-0
Grun, C. H., Dekker, N., Nieuwland, A. A., Klis, F. M., Kamerling, J. P., Vliegenthart, J. F. G., et al. (2006). Mechanism of action of the endo-(1→3)-α-glucanase MutAp from the mycoparasitic fungus Trichoderma harzianum. FEBS Lett. 580, 3780–3786. doi: 10.1016/j.febslet.2006.05.062
Gupta, V. K., Steindorff, A. S., de Paula, R. G., Silva-Rocha, R., Mach-Aigner, A. R., Mach, R. L., et al. (2016). The post-genomic era of Trichoderma reesei: What’s next? Trends Biotechnol. 34, 970–982. doi: 10.1016/j.tibtech.2016.06.003
Gurdon, J. B., Lane, C. D., Woodland, H. R., and Marbaix, G. (1971). Use of frog eggs and oocytes for the study of messenger RNA and its translation in living cells. Nature 233, 177–182. doi: 10.1038/233177a0
Halbeisen, R. E., and Gerber, A. P. (2009). Stress-dependent coordination of transcriptome and translatome in yeast. PLoS Biol. 7:e1000105. doi: 10.1371/journal.pbio.1000105
Harman, G. E. (2011). Multifunctional fungal plant symbionts: new tools to enhance plant growth and productivity. New Phytol. 189, 647–649. doi: 10.1111/j.1469-8137.2010.03614.x
Harman, G. E., Howell, C. R., Viterbo, A., Chet, I., and Lorito, M. (2004). Trichoderma species-opportunistic, avirulent plant symbionts. Nat. Rev. Microbiol. 2, 43–56. doi: 10.1038/nrmicro797
Hermosa, M. R., Grondona, I., Iturriaga, E. A., Diaz-Minguez, J. M., Castro, C., Monte, E., et al. (2000). Molecular characterization and identification of biocontrol isolates of Trichoderma spp. Appl. Environ. Microbiol. 66, 1890–1898. doi: 10.1128/AEM.66.5.1890-1898.2000
Hermosa, R., Viterbo, A., Chet, I., and Monte, E. (2012). Mini-review plant-beneficial effects of Trichoderma and of its genes. Microbiology 158, 17–25. doi: 10.1099/mic.0.052274-0
Herrera-Estrella, A. (2014). “Genome-wide approaches toward understanding mycotrophic Trichoderma species,” in Biotechnology and Biology of Trichoderma, eds V. Gupta, M. Schmoll, A. Herrera-Estrella, R. S. Upadhyay, and M. G. Tuohy (Oxford: Elsevier), 455–464. doi: 10.1111/j.1365-2958.1993.tb01142.x
Herrera-Estrella, A., Goldman, G. H., van Montagu, M., and Geremia, R. A. (1993). Electrophoretic karyotype and gene assignment to resolved chromosomes of Trichoderma spp. Mol. Microbiol. 7, 515–521.
Ho, C.-L., Tan, Y.-C., Yeoh, K.-A., Ghazali, A.-K., Yee, W.-Y., and Hoh, C.-C. (2016). De novo transcriptome analyses of host-fungal interactions in oil palm (Elaeis guineensis Jacq.). BMC Genomics 17:66. doi: 10.1186/s12864-016-2368-0
Hoell, I. A., Klemsdal, S. S., Vaaje-Kolstad, G., Horn, S. J., and Eijsink, V. G. H. (2005). Overexpression and characterization of a novel chitinase from Trichoderma atroviride strain P1. Biochim. Biophys. Acta 1748, 180–190. doi: 10.1016/j.bbapap.2005.01.002
Howell, C. R. (2006). Understanding the mechanisms employed by Trichoderma virens to effect biological control of cotton diseases. Phytopathology 96, 178–180. doi: 10.1094/PHYTO-96-0178
Ihrmark, K., Asmail, N., Ubhayasekera, W., Melin, P., Stenlid, J., and Karlsson, M. (2010). Comparative molecular evolution of Trichoderma chitinases in response to mycoparasitic interactions. Evol. Bioinform. 6, 1–26. doi: 10.1017/S1355838202026018
Ike, M., Nagamatsu, K., Shioya, A., Nogawa, M., Ogasawara, W., Okada, H., et al. (2006). Purification, characterization, and gene cloning of 46 kDa chitinase (Chi46) from Trichoderma reesei PC-3-7 and its expression in Escherichia coli. Appl. Microbiol. Biotechnol. 71, 294–303. doi: 10.1007/s00253-005-0171-y
Inada, T., Winstall, E., Tarun, S. Z. Jr., Yates, J. R. III, Schieltz, D., and Sachs, A. B. (2002). One-step affinity purification of the yeast ribosome and its associated proteins and mRNAs. RNA 8, 948–958.
Ingolia, N. T., Brar, G. A., Rouskin, S., McGeachy, A. M., and Weissman, J. S. (2012). The ribosome profiling strategy for monitoring translation in vivo by deep sequencing of ribosome-protected mRNA fragments. Nat. Protoc. 7, 1534–1550. doi: 10.1038/nprot.2012.086
Ingolia, N. T., Ghaemmaghami, S., Newman, J. R., and Weissman, J. S. (2009). Genome-wide analysis in vivo of translation with nucleotide resolution using ribosome profiling. Science 324, 218–223. doi: 10.1126/science.1168978
Jackson, R. J., Hellen, C. U., and Pestova, T. V. (2010). The mechanism of eukaryotic translation initiation and principles of its regulation. Nat. Rev. Mol. Cell Biol. 11, 113–127. doi: 10.1038/nrm2838
Jayapal, K. P., Philp, R. J., Kok, Y. J., Yap, M. G., Sherman, D. H., Griffin, T. J., et al. (2008). Uncovering genes with divergent mRNA–protein dynamics in Streptomyces coelicolor. PLoS ONE 3:e2097. doi: 10.1371/journal.pone.0002097
Jiao, Y., and Meyerowitz, E. M. (2010). Cell-type specific analysis of translating RNAs in developing flowers reveals new levels of control. Mol. Syst. Biol. 6, 419. doi: 10.1038/msb.2010.76
Karginov, F. V., and Hannon, G. J. (2013). Remodeling of Ago2-mRNA interactions upon cellular stress reflects miRNA complementarity and correlates with altered translation rates. Genes Dev. 27, 1624–1632. doi: 10.1101/gad.215939.113
Kim, D. J., Baek, J. M., Uribe, P., Kenerley, C. M., and Cook, D. R. (2002). Cloning and characterization of multiple glycosyl hydrolase genes from Trichoderma virens. Curr. Genet. 40, 374–384. doi: 10.1007/s00294-001-0267-6
King, H. A., and Gerber, A. P. (2016). Translatome profiling: methods for genome-scale analysis of mRNA translation. Brief. Funct. Genomics 15, 22–31. doi: 10.1093/bfgp/elu045
Klemsdal, S. S., Clarke, J. L., Hoell, I., Eijsink, V. G. H., and Brurberg, M. B. (2006). Molecular cloning, characterization, and expression studies of a novel chitinase gene (ech30) from the mycoparasite Trichoderma atroviride strain P1. FEMS Microbiol. Lett. 256, 282–289. doi: 10.1111/j.1574-6968.2006.00132.x
Koritzinsky, M., and Wouters, B. G. (2007). Hypoxia and regulation of messenger RNA translation. Methods Enzymol. 435, 247–273. doi: 10.1016/S0076-6879(07)35013-1
Kottb, M., Gigolashvili, T., Großkinsky, D. K., and Piechulla, B. (2015). Trichoderma volatiles effecting Arabidopsis: from inhibition to protection against phytopathogenic fungi. Front. Microbiol. 6:995. doi: 10.3389/fmicb.2015.00995
Kubicek, C. P., Herrera-Estrella, A., Seidl-seiboth, V., Martinez, D. A., Druzhinina, I. S., Thon, M., et al. (2011). Comparative genome sequence analysis underscores mycoparasitism as the ancestral life style of Trichoderma. Genome Biol. 12:R40. doi: 10.1186/gb-2011-12-4-r40
Kulminskaya, A. A., Thomsen, K. K., Shabalin, K. A., Sidorenko, I. A., Eneyskaya, E. V., Savel, A. N., et al. (2001). Isolation, enzymatic properties, and mode of action of an exo-1,3-β-glucanase from T. viride. FEBS J. 268, 6123–6131.
Lamdan, N.-L., Shalaby, S., Ziv, T., Kenerley, C. M., and Horwitz, B. A. (2015). Secretome of Trichoderma interacting with maize roots: role in induced systemic resistance. Mol. Cell Proteomics 14, 1054–1063. doi: 10.1074/mcp.M114.046607
Lee, S., Yap, M., Behringer, G., Hung, R., and Bennett, J. W. (2016). Volatile organic compounds emitted by Trichoderma species mediate plant growth. Fungal Biol. Biotechnol. 3:7. doi: 10.1186/s40694-016-0025-7
Lin, Y. R., Lo, C. T., Liu, S. Y., and Peng, K. C. (2012). Involvement of pachybasin and emodin in self-regulation of Trichoderma harzianum mycoparasitic coiling. J. Agric. Food Chem. 60, 2123–2128. doi: 10.1021/jf202773y
Liu, P. G., and Yang, Q. (2005). Identification of genes with a biocontrol function in Trichoderma harzianum mycelium using the expressed sequence tag approach. Res. Microbiol. 156, 416–423. doi: 10.1016/j.resmic.2004.10.007
Liu, Z., Yang, X., Sun, D., Song, J., Chen, G., Juba, O., et al. (2010). Expressed sequence tags-based identification of genes in a biocontrol strain Trichoderma asperellum. Mol. Biol. Rep. 37, 3673–3681. doi: 10.1007/s11033-010-0019-0
Loc, N. H., Quang, H. T., Hung, N. B., Huy, N. D., Phuong, T. T. B., and Ha, T. T. T. (2011). Trichoderma asperellum Chi42 genes encode chitinase. Mycobiology 39, 182–186. doi: 10.5941/MYCO.2011.39.3.182
Longa, C. M., Savazzini, F., Tosi, S., Elad, Y., and Pertot, I. (2009). Evaluating the survival and environmental fate of the biocontrol agent Trichoderma atroviride SC1 in vineyards in northern Italy. J. Appl. Microbiol. 106, 1549–1557. doi: 10.1111/j.1365-2672.2008.04117.x
Lopez-Mondejar, R., Catalano, V., Kubicek, C. P., and Seidl, V. (2009). The β-N-acetylglucosaminidases NAG1 and NAG2 are essential for growth of Trichoderma atroviride on chitin. FEBS J. 276, 5137–5148. doi: 10.1111/j.1742-4658.2009.07211.x
Lorito, M., Woo, S. L., Harman, G. E., and Monte, E. (2010). Translational research on Trichoderma: from ’omics to the field. Annu. Rev. Phytopathol. 48, 395–417. doi: 10.1146/annurev-phyto-073009-114314
Maischak, H., Zimmermann, M. R., Felle, H. H., and Boland, W. (2010). Alamethicin-induced electrical long distance signaling in plants. Plant Signal. Behav. 5, 988–990. doi: 10.1104/pp.108.133884
Mantyla, A. L., Rossi, K. H., Vanhanen, S. A., Penttila, M. E., Suominen, P. L., and Nevalainen, K. M. H. (1992). Electrophoretic karyotyping of wild-type and mutant Trichoderma longibrachiatum (reesei) strains. Curr. Genet. 21, 471–477. doi: 10.1007/BF00351657
Marra, R., Ambrosino, P., Carbone, V., Vinale, F., Woo, S. L., Ruocco, M., et al. (2006). Study of the three- way interaction between Trichoderma atroviride, plant and fungal pathogens by using a proteomic approach. Curr. Genet. 50, 307–321. doi: 10.1007/s00294-006-0091-0
Martinez, D., Berka, R. M., Henrissat, B., Saloheimo, M., Arvas, M., Baker, S. E., et al. (2008). Genome sequencing and analysis of the biomass-degrading fungus Trichoderma reesei (syn. Hypocrea Jecorina). Nat. Biotechnol. 26, 553–560. doi: 10.1038/nbt1403
Martínez-Medina, A., Appels, F. V. W., and Van Wees, S. C. M. (2017a). Impact of salicylic acid- and jasmonic acid-regulated defences on root colonization by Trichoderma harzianum T-78. Plant Signal. Behav. doi: 10.1080/15592324.2017.1345404 [Epub ahead of print].
Martínez-Medina, A., Van Wees, S. C. M., and Pieterse, C. M. J. (2017b). Airborne signals by Trichoderma fungi stimulate iron uptake responses in roots resulting in priming of jasmonic acid-dependent defences in shoots of Arabidopsis thaliana and Solanum lycopersicum. Plant Cell Environ. doi: 10.1111/pce.13016 [Epub ahead of print].
Mathys, J., De Cremer, K., Timmermans, P., Van Kerckhove, S., Lievens, B., Vanhaecke, M., et al. (2012). Genome-Wide characterization of ISR induced in Arabidopsis thaliana by Trichoderma hamatum T382 against Botrytis cinerea infection. Front. Plant Sci. 3:108. doi: 10.3389/fpls.2012.00108
Mehrabi-Koushki, M., Rouhani, H., and Mahdikhani-Moghaddam, E. (2012). Differential display of abundantly expressed genes of Trichoderma harzianum during colonization of tomato-germinating seeds and roots. Curr. Microbiol. 65, 524–533. doi: 10.1007/s00284-012-0189-1
Merino, S., and Cherry, J. (2007). “Progress and challenges in enzyme development for biomass utilization,” in Biofuels, ed. L. Olsson (Berlin: Springer), 95–120.
Meteignier, L. V., El Oirdi, M., Cohen, M., Barff, T., Matteau, D., Lucier, J. F., et al. (2017). Translatome analysis of an NB-LRR immune response identifies important contributors to plant immunity in Arabidopsis. J. Exp. Bot. 68, 2333–2344. doi: 10.1093/jxb/erx078
Montero, M., Sanz, L., Rey, M., Llobell, A., and Monte, E. (2007). Cloning and characterization of bgn16.3, coding for a β-1,6-glucanase expressed during Trichoderma harzianum mycoparasitism. J. Appl. Microbiol. 103, 1291–1300. doi: 10.1111/j.1365-2672.2007.03371.x
Montero, M., Sanz, L., Rey, M., Monte, E., and Llobell, A. (2005). BGN16.3, a novel acidic β-1,6-glucanase from mycoparasitic fungus Trichoderma harzianum CECT 2413. FEBS J. 272, 3441–3448. doi: 10.1111/j.1742-4658.2005.04762.x
Morán-Diez, E., Rubio, B., Domínguez, S., Hermosa, R., Monte, E., and Nicolás, C. (2012). Transcriptomic response of Arabidopsis thaliana after 24h incubation with the biocontrol fungus Trichoderma harzianum. J. Plant Physiol. 169, 614–620. doi: 10.1016/j.jplph.2011.12.016
Morán-Diez, M. E., Cardoza, R. E., Gutiérrez, S., Monte, E., and Hermosa, R. (2010). TvDim1 of Trichoderma virens is involved in redox-processes and confers resistance to oxidative stresses. Curr. Genet. 56, 63–73. doi: 10.1007/s00294-009-0280-8
Morán-Diez, M. E., Trushina, N., Lamdan, N. L., Rosenfelder, L., Mukherjee, P. K., Kenerley, C. M., et al. (2015). Host-specific transcriptomic pattern of Trichoderma virens during interaction with maize or tomato roots. BMC Genomics 16:8. doi: 10.1186/s12864-014-1208-3
Mukherjee, M. (2012). “Trichoderma genes involved in interactions with fungi and plants,” in Biotechnology of Fungal Genes, eds V. K. Gupta and M. Ayyachamy (New York, NY: CRC Press), 153–171.
Mukherjee, P. K., Horwitz, B. A., Herrera-Estrella, A., Schmoll, M., and Kenerley, C. M. (2013). Trichoderma research in the genome era. Annu. Rev. Phytopathol. 51, 105–129. doi: 10.1146/annurev-phyto-082712-102353
Mukherjee, P. K., Horwitz, B. A., and Kenerley, C. M. (2012). Secondary metabolism in Trichoderma - a genomic perspective. Microbiology 158, 35–45. doi: 10.1099/mic.0.053629-0
Nobe, R., Sakakibara, Y., Ogawa, K., and Suiko, M. (2004). Cloning and expression of a novel Trichoderma viride laminarinase AI gene (lamAI). Biosci. Biotechnol. Biochem. 68, 2111–2119. doi: 10.1271/bbb.68.2111
Palmieri, M. C., Perazzolli, M., Matafora, V., Moretto, M., Bachi, A., and Pertot, I. (2012). Proteomic analysis of grapevine resistance induced by Trichoderma harzianum T39 reveals specific defence pathways activated against downy mildew. J. Exp. Bot. 63, 6237–6251. doi: 10.1093/jxb/ers279
Panizel, I., Yarden, O., Ilan, M., and Carmeli, S. (2013). Eight new peptaibols from sponge-associated Trichoderma atroviride. Mar. Drugs 11, 4937–4960. doi: 10.3390/md11124937
Papavizas, G. C. (1985). Trichoderma and Gliocladium: biology, ecology, and potential for biocontrol. Annu. Rev. Phytopathol. 23, 23–54. doi: 10.1146/annurev.py.23.090185.000323
Payne, S. H. (2015). The utility of protein and mRNA correlation. Trends Biochem. Sci. 40, 1–3. doi: 10.1016/j.tibs.2014.10.010
Pelagio-Flores, R., Esparza-Reynoso, S., Garnica-Vergara, A., López-Bucio, J., and Herrera-Estrella, A. (2017). Trichoderma-induced acidification is an early trigger for changes in Arabidopsis root growth and determines fungal phytostimulation. Front. Plant Sci. 17:822. doi: 10.3389/fpls.2017.00822
Perazzolli, M., Moretto, M., Fontana, P., Ferrarini, A., Velasco, R., Moser, C., et al. (2012). Downy mildew resistance induced by Trichoderma harzianum T39 in susceptible grapevines partially mimics transcriptional changes of resistant genotypes. BMC Genomics 13:660. doi: 10.1186/1471-2164-13-660
Picard, F., Loubière, P., Girbal, L., and Cocaign-Bousquet, M. (2013). The significance of translation regulation in the stress response. BMC Genomics 14:588. doi: 10.1186/1471-2164-14-588
Piccirillo Ciriaco, A., Bjur, E., Topisirovic, I., Sonenberg, N., and Larsson, O. (2014). Translational control of immune responses: from transcripts to translatomes. Nat. Immunol. 15, 503–511. doi: 10.1038/ni.2891
Przylucka, A., Bayram, G., Chenthamara, K., Cai, F., Grujic, M., Karpenko, J., et al. (2017). HFB7 – a novel orphan hydrophobin of the Harzianum and Virens clades of Trichoderma, is involved in response to biotic and abiotic stresses. Fungal Genet. Biol. 102, 63–76. doi: 10.1016/j.fgb.2017.01.002
Puglisi, I., Faedda, R., Sanzaro, V., Lo Piero, A. R., Petrone, G., and Cacciola, S. O. (2012). Identification of differentially expressed genes in response to mercury I and II stress in Trichoderma harzianum. Gene 15, 325–330. doi: 10.1016/j.gene.2012.06.091
Qi, W., and Zhao, L. (2013). Study of the siderophore-producing Trichoderma asperellum Q1 on cucumber growth promotion under salt stress. J. Basic Microbiol. 53, 355–364. doi: 10.1002/jobm.201200031
Reino, J. L., Guerrero, R. F., Hernández-Galan, R., and Collado, I. G. (2008). Secondary metabolites from species of the biocontrol agent Trichoderma. Phytochemistry 7, 89–123. doi: 10.1007/s11101-006-9032-2
Reithner, B., Ibarra-Laclette, E., Mach, R. L., and Herrera-Estrella, A. (2011). Identification of mycoparasitism-related genes in Trichoderma atroviride. Appl. Environ. Microbiol. 77, 4361–4370. doi: 10.1128/AEM.00129-11
Röhrich, C. R., Voglmayr, H., Iversen, A., and Vilcinskas, A. (2015). Front line defenders of the ecological niche! Screening the structural diversity of peptaibiotics from saprotrophic and fungicolous Trichoderma/Hypocrea species. Fungal Divers. 69, 117–146. doi: 10.1007/s13225-013-0276-z
Rosales-Saavedra, T., Esquivel-Naranjo, E. U., Casas-Flores, S., Martinez-Hernandez, P., Ibarra-Laclette, E., Cortes-Penagos, C., et al. (2006). Novel light-regulated genes in Trichoderma atroviride: a dissection by cDNA microarrays. Microbiology 152, 3305–3317. doi: 10.1099/mic.0.29000-0
Rubio, M. B., Dominguez, S., Monte, E., and Hermosa, R. (2012). Comparative study of Trichoderma gene expression in interactions with tomato plants using high-density oligonucleotide microarrays. Microbiology 158(Pt 1), 119–128. doi: 10.1099/mic.0.052118-0
Rubio, M. B., Hermosa, R., Reino, J. L., Collado, I. G., and Monte, E. (2009). Thctf1 transcription factor of Trichoderma harzianum is involved in 6-pentyl-2H-pyran-2-one production and antifungal activity. Fungal Genet. Biol. 46, 17–27. doi: 10.1016/j.fgb.2008.10.008
Rubio, M. B., Quijada, N. M., Pérez, E., Domínguez, S., Monte, E., and Hermosa, R. (2014). Identifying beneficial qualities of Trichoderma parareesei for plants. Appl. Environ. Microbiol. 80, 1864–1873. doi: 10.1128/AEM.03375-13
Ruocco, M., Lanzuise, S., Lombardi, N., Woo, S. L., Vinale, F., and Marra, R. (2015). Multiple roles and effects of a novel Trichoderma hydrophobin. Mol. Plant Microbe Interact. 28, 167–179. doi: 10.1094/MPMI-07-14-0194-R
Ruocco, M., Lanzuise, S., Vinale, F., Marra, R., Turrà, D., Woo, S. L., et al. (2009). Identification of a new biocontrol gene in Trichoderma atroviride: the role of an ABC transporter membrane pump in the interaction with different plant-pathogenic fungi. Mol. Plant Microbe Interact. 22, 291–301. doi: 10.1094/MPMI-22-3-0291
Samolski, I., de Luis, A., Vizcaino, J. A., Monte, E., and Suarez, M. B. (2009). Gene expression analysis of the biocontrol fungus Trichoderma harzianum in the presence of tomato plants, chitin, or glucose using a high-density oligonucleotide microarray. BMC Microbiol. 9:217. doi: 10.1186/1471-2180-9-217
Sanna, A. (2006). Characterization of the Trichoderma reesei Hydrophobins HFBI and HFBII. Doctoral dissertation, Helsinki University of Technology, Espoo. doi: 10.1007/s00294-008-0226-6
Scherm, B., Schmoll, M., Balmas, V., Kubicek, C. P., and Migheli, Q. (2009). Identification of potential marker genes for Trichoderma harzianum strains with high antagonistic potential against Rhizoctonia solani by a rapid subtraction hybridization approach. Curr. Genet. 55, 81–91. doi: 10.1007/s00294-008-0226-6
Schmoll, M., Dattenböck, C., Carreras-Villaseñor, N., Mendoza-Mendoza, A., Tisch, D., Alemán, M. I., et al. (2016). The genomes of three uneven siblings: footprints of the lifestyles of three Trichoderma species. Microbiol. Mol. Biol. Rev.? 80, 205–327. doi: 10.1128/MMBR.00040-15
Schuster, A., and Schmoll, M. (2010). Biology and biotechnology of Trichoderma. Appl. Microbiol. Biotechnol. 87, 787–799. doi: 10.1007/s00253-010-2632-1
Segarra, G., Casanova, E., Bellido, D., Odena, M. A., Oliveira, E., and Trillas, I. (2007). Proteome, salicylic acid, and jasmonic acid changes in cucumber plants inoculated with Trichoderma asperellum strain T34. Proteomics 7, 3943–3952. doi: 10.1002/pmic.200700173
Seidl, V., Huemer, B., Seiboth, B., and Kubicek, C. P. (2005). A complete survey of Trichoderma chitinases reveals three distinct subgroups of family 18 chitinases. FEBS J. 272, 5923–5939. doi: 10.1111/j.1742-4658.2005.04994.x
Seidl, V., Song, L., Lindquist, E., Gruber, S., Koptchinskiy, A., Zeilinger, S., et al. (2009). Transcriptomic response of the mycoparasitic fungus Trichoderma atroviride to the presence of a fungal prey. BMC Genomics 10:567. doi: 10.1186/1471-2164-10-567
Sharma, V., Bhandari, P., Singh, B., Bhatacharya, A., and Shanmugam, V. (2013). Chitinase expression due to reduction in fusaric acid level in an antagonistic Trichoderma harzianum S17TH. Indian J. Microbiol. 53, 214–220. doi: 10.1007/s12088-012-0335-2
Sharma, V., and Salwan, R. (2017). “Molecular markers and their use in taxonomic characterization of Trichoderma spp,” in Molecular Markers in Mycology. Fungal Biology, eds B. Singh and V. Gupta (Cham: Springer).
Sharma, V., Salwan, R., and Sharma, P. N. (2017a). The comparative mechanistic aspects of Trichoderma and probiotics: scope for future research. Physiol. Mol. Plant Pathol. 100, 84–96. doi: 10.1016/j.pmpp.2017.07.005
Sharma, V., Salwan, R., and Sharma, P. N. (2016a). Differential response of extracellular proteases of Trichoderma harzianum against fungal phytopathogens. Curr. Microbiol. 73, 419–425. doi: 10.1007/s00284-016-1072-2
Sharma, V., Salwan, R., Sharma, P. N., and Kanwar, S. S. (2017b). Elucidation of biocontrol mechanisms of Trichoderma harzianum against different plant fungal pathogens: universal yet host specific response. Int. J. Biol. Macromol. 95, 72–79. doi: 10.1016/j.ijbiomac.2016.11.042
Sharma, V., Salwan, R., Sharma, P. N., and Kanwar, S. S. (2016b). Molecular cloning and characterization of ech46 endochitinase from Trichoderma harzianum. Int. J. Biol. Macromol. 92, 615–624. doi: 10.1016/j.ijbiomac.2016.07.067
Sharma, V., and Shanmugam, V. (2012). Purification and characterization of an extracellular 24 kDa chitobiosidase from the mycoparasitic fungus Trichoderma saturnisporum. J. Basic Microbiol. 52, 324–331. doi: 10.1002/jobm.201100145
Shaw, S., Cocq, K. L., Paszkiewicz, K., Moore, K., Winsbury, R., de Torres Zabala, M., et al. (2016). Transcriptional reprogramming underpins enhanced plant growth promotion by the biocontrol fungus Trichoderma hamatum GD12 during antagonistic interactions with Sclerotinia sclerotiorum in soil. Mol. Plant Pathol. 17, 1425–1441. doi: 10.1111/mpp.12429
Shentu, X.-P., Liu, W.-P., Zhan, X.-H., Xu, Y. P., Xu, J. F., Yu, X. P., et al. (2014). Transcriptome sequencing and gene expression analysis of Trichoderma brevicompactum under different culture conditions. PLoS ONE 9:e94203. doi: 10.1371/journal.pone.0094203
Shi, M., Chen, L., Wang, X., Zhang, T., Zhao, P., Song, X., et al. (2012). Antimicrobial peptaibols from Trichoderma pseudokoningii induce programmed cell death in plant fungal pathogens. Microbiology 158, 166–175. doi: 10.1099/mic.0.052670-0
Shi, W.-L., Chen, X.-L., Wang, L.-X., Gong, Z.-T., Li, S., Li, C.-L., et al. (2016). Cellular and molecular insight into the inhibition of primary root growth of Arabidopsis induced by peptaibols, a class of linear peptide antibiotics mainly produced by Trichoderma spp. J. Exp. Bot. 67, 2191–2205. doi: 10.1093/jxb/erw023
Shoresh, M., and Harman, G. E. (2008). The molecular basis of shoot responses of maize seedlings to Trichoderma harzianum T22 inoculation of the root: a proteomic approach. Plant Physiol. 147, 2147–2163. doi: 10.1104/pp.108.123810
Shoresh, M., Harman, G. E., and Mastouri, F. (2010). Induced systemic resistance and plant responses to fungal biocontrol agents. Annu. Rev. Phytopathol. 48, 21–43. doi: 10.1146/annurev-phyto-073009-114450
Simkovi, M., Kurucová, A., Hunová, M., and Vare, L’. (2008). Induction of secretion of extracellular proteases from Trichoderma viride. Acta Chim. Slovaca 1, 250–264.
Singh, A., Taylor, L. E. II, Vander Wall, T. A., Linger, J., Himmel, M. E., Podkaminer, K., et al. (2015). Heterologous protein expression in Hypocrea jecorina: a historical perspective and new developments. Biotechnol. Adv. 33, 142–154. doi: 10.1016/j.biotechadv.2014.11.009
Spangenberg, L., Shigunov, P., Abud, A. P., Cofré, A. R., Stimamiglio, M. A., Kuligovski, C., et al. (2013). Polysome profiling shows extensive posttranscriptional regulation during human adipocyte stem cell differentiation into adipocytes. Stem Cell Res. 11, 902–912. doi: 10.1016/j.scr.2013.06.002
Spriggs, K. A., Bushell, M., and Willis, A. E. (2010). Translational regulation of gene expression during conditions of cell stress. Mol. Cell. 40, 228–237. doi: 10.1016/j.molcel.2010.09.028
Steindorff, A. S., Ramada, M. H., Coelho, A. S., Miller, R. N., Pappas, G. J. Jr., Ulhoa, C. J., et al. (2014). Identification of mycoparasitism-related genes against the phytopathogen Sclerotinia sclerotiorum through transcriptome and expression profile analysis in Trichoderma harzianum. BMC Genomics 15:204. doi: 10.1186/1471-2164-15-204
Steindorff, A. S., Silva, R. N., Coelho, A. S. G., Nagata, T., Noronha, E. F., and Ulhoa, C. J. (2012). Trichoderma harzianum expressed sequence tags for identification of genes with putative roles in mycoparasitism against Fusarium solani. Biol. Control 61, 134–140. doi: 10.1016/j.biocontrol.2012.01.014
Suárez, M. B., Vizcaíno, J. A., Llobell, A., and Monte, E. (2007). Characterization of genes encoding novel peptidases in the biocontrol fungus Trichoderma harzianum CECT 2413 using the TrichoEST functional genomics approach. Curr. Genet. 51, 331–342. doi: 10.1007/s00294-007-0130-5
Szekeres, A., Kredics, L., Antal, Z., Kevei, F., and Manczinger, L. (2004). Isolation and characterization of protease overproducing mutants of Trichoderma harzianum. FEMS Microbiol. Lett. 233, 215–222. doi: 10.1111/j.1574-6968.2004.tb09485.x
Teresa, M., Bara, F., Lima, A. L., and Ulhoa, C. J. (2003). Purification and characterization of an exo- β-1,3-glucanase produced by Trichoderma asperellum. FEMS Microbiol. Lett. 219, 81–85. doi: 10.1016/S0378-1097(02)01191-6
Terman, S. A. (1970). Relative effect of transcription-level and translation-level control of protein synthesis during early development of the sea urchin. Proc. Natl. Acad. Sci. U.S.A. 65, 985–992. doi: 10.1073/pnas.65.4.985
Thomas, J. D., and Johannes, G. J. (2007). Identification of mRNAs that continue to associate with polysomes during hypoxia. RNA 13, 1116–1131. doi: 10.1261/rna.534807
Tian, Q., Stepaniants, S. B., Mao, M., Mao, M., Weng, L., Feetham, M. C., et al. (2004). Integrated genomic and proteomic analyses of gene expression in Mammalian cells. Mol. Cell. Proteomics 3, 960–969. doi: 10.1074/mcp.M400055-MCP200
Tucci, M., Ruocco, M., De Masi, L., De Palma, M., and Lorito, M. (2011). The beneficial effect of Trichoderma spp. on tomato is modulated by the plant genotype. Mol. Plant Pathol. 12, 341–354. doi: 10.1111/j.1364-3703.2010.00674.x
Vazqez-Garciduen, S., Leal-morales, C. A., and Herrera-Estrella, A. (1998). Analysis of the β-1,3-glucanolytic system of the biocontrol agent Trichoderma harzianum. Appl. Environ. Microbiol. 64, 1442–1446.
Vieira, P. M., Coelho, A. S. G., Steindorff, A. S., de Siqueira, S. J. L., Silva, R., do, N., et al. (2013). Identification of differentially expressed genes from Trichoderma harzianum during growth on cell wall of Fusarium solani as a tool for biotechnological application. BMC Genomics 14:177. doi: 10.1186/1471-2164-14-177
Vinale, F., Sivasithamparam, K., Ghisalberti, E. L., Woo, S. L., Nigro, M., Marra, R., et al. (2014). Trichoderma secondary metabolites active on plants and fungal pathogens. Open Mycol. J. 8, 127–139. doi: 10.1111/j.1472-765X.2009.02599.x
Viterbo, A., Harel, M., and Chet, I. (2004). Isolation of two aspartyl proteases from Trichoderma asperellum expressed during colonization of cucumber roots. FEMS Microbiol. Lett. 238, 151–158.
Viterbo, A., Wiest, A., Brotman, Y., Chet, I., and Kenerley, C. (2007). The 18mer peptaibols from Trichoderma virens elicit plant defence responses. Mol. Plant. Pathol. 8, 737–746. doi: 10.1111/j.1364-3703.2007.00430.x
Viterbo, A., Ramot, O., Chemin, L., and Chet, I. (2002). Significance of lytic enzymes from Trichoderma spp. in the biocontrol of fungal plant pathogens. Antonie Van Leeuwenhoek 81, 549–556. doi: 10.1023/A:1020553421740
Vizcaíno, J. A., González, F. J., Suárez, M. B., Redondo, J., Heinrich, J., Delgado-Jarana, J., et al. (2006). Generation, annotation and analysis of ESTs from Trichoderma harzianum CECT 2413. BMC Genomics 7:193. doi: 10.1186/1471-2164-7-193
Vizcaíno, J. A., Redondo, J., Suárez, M. B., Cardoza, R. E., Hermosa, R., González, F. J., et al. (2007). Generation, annotation, and analysis of ESTs from four different Trichoderma strains grown under conditions related to biocontrol. Appl. Microbiol. Biotechnol. 75, 853–862. doi: 10.1007/s00253-007-0885-0
Vogel, C., and Marcotte, E. M. (2012). Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat. Rev. Genet. 13, 227–232. doi: 10.1038/nrg3185
Wu, Q., Sun, R., Ni, M., Yu, J., Li, Y., Yu, C., et al. (2017). Identification of a novel fungus, Trichoderma asperellum GDFS1009, and comprehensive evaluation of its biocontrol efficacy. PLoS ONE 12:e0179957. doi: 10.1371/journal.pone.0179957
Xie, B.-B., Li, D., Shi, W.-L., Qin, Q. L., Wang, X., Rong, J. C., et al. (2015). Deep RNA sequencing reveals a high frequency of alternative splicing events in the fungus Trichoderma longibrachiatum. BMC Genomics 16:54. doi: 10.1186/s12864-015-1251-8
Xie, B.-B., Qin, Q.-L., Shi, M., Chen, L.-L., Shu, Y.-L., Luo, Y., et al. (2014). Comparative genomics provide insights into evolution of Trichoderma nutrition style. Genome Biol. Evol. 6, 379–390. doi: 10.1093/gbe/evu018
Yanguez, E., Castro-Sanz, A. B., Ferna´ndez-Bautista, N., Oliveros, J. C., and Castellano, M. M. (2013). Analysis of genome-wide changes in the translatome of Arabidopsis seedlings subjected to heat stress. PLoS ONE 8:e71425. doi: 10.1371/journal.pone.0071425
Yao, L., Yang, Q., Song, J., Tan, C., Guo, C., Wang, L., et al. (2013). Cloning, annotation and expression analysis of mycoparasitism-related genes in Trichoderma harzianum 88. J. Microbiol. 51, 174–182. doi: 10.1007/s12275-013-2545-7
Yedidia, I. I., Benhamou, N., and Chet, I. I. (1999). Induction of defense responses in cucumber plants (Cucumis sativus L.) by the biocontrol agent Trichoderma harzianum. Appl. Environ. Microbiol. 65, 1061–1070.
Zeilinger, S., Gruber, S., Bansal, R., and Mukherjee, P. K. (2016). Secondary metabolism in Trichoderma – chemistry meets genomics. Fungal Biol. Rev. 30, 74–90. doi: 10.1016/j.fbr.2016.05.001
Zhang, F., Yang, X., Ran, W., and Shen, Q. (2014). Fusarium oxysporum induces the production of proteins and volatile organic compounds by Trichoderma harzianum T-E5. FEMS Microbiol. Lett. 359, 116–123. doi: 10.1111/1574-6968.12582
Zhang, T., Chaturvedi, V., and Chaturvedi, S. (2015). Novel Trichoderma polysporum strain for the biocontrol of Pseudogymnoascus destructans, the fungal etiologic agent of bat white nose syndrome. PLoS ONE 10:e0141316. doi: 10.1371/journal.pone.0141316
Keywords: transcripts, active mRNA, regulation, integrated omic, translatome
Citation: Sharma V, Salwan R, Sharma PN and Gulati A (2017) Integrated Translatome and Proteome: Approach for Accurate Portraying of Widespread Multifunctional Aspects of Trichoderma. Front. Microbiol. 8:1602. doi: 10.3389/fmicb.2017.01602
Received: 21 April 2017; Accepted: 07 August 2017;
Published: 29 August 2017.
Edited by:
Katarzyna Turnau, Jagiellonian University, PolandReviewed by:
Somayeh Dolatabadi, Westerdijk Fungal Biodiversity Institute, NetherlandsRavindra Nath Kharwar, Banaras Hindu University, India
Copyright © 2017 Sharma, Salwan, Sharma and Gulati. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) or licensor are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Vivek Sharma, YW5rdml2ZWtAZ21haWwuY29t