 Baiyila Wu
Baiyila Wu Zongfu Hu
Zongfu Hu Manlin Wei
Manlin Wei Huaxin Niu
Huaxin Niu- College of Animal Science and Technology, Inner Mongolia Minzu University, Tongliao, China
Leymus chinensis is an important crop that can be fed to ruminants. The purpose of this study was to investigate the roles of Lactiplantibacillus plantarum and Lentilactobacillus buchneri in fermentation quality, aerobic stability, and dynamics of wilted L. chinensis silage microorganisms. Wilted L. chinensis silages were ensiled with/without L. plantarum and L. buchneri. After 14 and 56 days of ensiling, the silos were opened and subjected to a 7-day aerobic deterioration test. This study looked at the composition of fermentation products as well as the microbial communities in silage. Silage inoculated with L. plantarum and L. buchneri had an increased lactic acid content as well as lactic acid bacterial (LAB) quantity, but a decrease in pH and levels of butyric acid, 2,3-butanediol, and ethanol was observed during ensiling. Non-treated and L. plantarum-treated silages deteriorated in the 7-day spoilage test after opening day-14 silos, whereas L. buchneri-inoculated silage showed no signs of deterioration. Lactobacillus abundance increased in the 7-day spoilage test after opening day-56 silos, while undesirable microorganisms such as Acetobacter, Bacillus, and molds, namely, Aspergillus and Penicillium were inhibited within L. plantarum- and L. buchneri-inoculated silages. The composition of fermentation products was related to the bacterial community, particularly Lactobacillus, Enterococcus, and Acetobacter. To summarize, L. plantarum- and L. buchneri-inoculated silage enhanced fermentation quality during ensiling and inhibited aerobic spoilage in a 7-day spoilage test of 56 days ensiling within wilted L. chinensis silage.
Introduction
Leymus chinensis (Trin.) Tzvel. is a Gramineae perennial grass with a wide range of adaptations that is a major source of animal feed in China (Li et al., 2019). Due to its excellent palatability, tender stems and leaves, and capacity to meet ruminant nutrition demands, L. chinensis is one of the most important forage grasses in China’s Inner Mongolian Plateau, Korea, Russia, the Northern Plain, and Mongolia (Peng et al., 2013; Zhu et al., 2020). Despite its high yield in these areas, L. chinensis has a low lactic acid bacteria (LAB) count and water-soluble carbohydrates (WSCs) level, which can hinder ensilage as a method of grass conservation (Zhang et al., 2014). Forages must develop an appropriate ensilage process because feeding high-quality silage is supposed to increase the productivity of livestock such as beef and dairy cattle.
Lactic acid bacteria inoculants have been widely used to stimulate the fermentation process in order to produce high-quality silage. Previous studies have reported the classification of inoculants as homofermentative and heterofermentative bacteria based on fermentation profiles (Muck et al., 2018). Many homofermentative LAB strains, such as Lactiplantibacillus plantarum, Pediococcus species, and Enterococcus faecium, have been widely used for dominating ensiling fermentation by rapidly producing lactic acid while lowering pH, preventing yeast, mold, and other undesirable microorganisms, and preserving forage nutrients during storage (Ranjit and Kung, 2000). However, aerobic spoilage of sorghum, wheat, corn, and Italian ryegrass silages is increased when homofermentative LAB is used because the elevated lactic acid is rapidly oxidized by yeasts and other aerobic microorganisms after silage air exposure (Weinberg et al., 2002; Tabacco et al., 2009; Li and Nishino, 2011a). One of the heterofermentative LAB species, Lentilactobacillus buchneri, was developed as a silage inoculant to prevent aerobic degradation. Under anaerobic conditions, L. buchneri inhibited aerobic deterioration by fermenting lactic acid into 1,2-propanediol and acetic acid (Oude Elferink et al., 2001). A large amount of acetic acid was produced to protect the silage from aerobic molds and yeasts that cause aerobic spoilage (Nishino et al., 2003).
Over the last few decades, culture-dependent/independent approaches, such as 16S-rRNA gene sequences, real-time PCR, ribosomal intergenic spacer analysis, and characterizing denaturing gradient gel electrophoresis bands, have been used to investigate the epiphytic microbiota in pre-ensiled crops and the microbiota in ensiled forages (Ennahar et al., 2003; Stevenson et al., 2006; Brusetti et al., 2011; Wu et al., 2014). Although the preceding studies provide information on microorganism composition in silage processes, the associated analyses only discovered a few major operational taxonomic units (OTUs) and failed to provide detailed information on microbial community composition (Ni et al., 2017). Next-generation sequencing (NGS; e.g., the ITS unit for fungi and 16S rRNA for bacteria) has proven to be a quick and accurate molecular tool for defining silage ecology, such as a description of microorganisms within silage (Wang et al., 2020b).
Furthermore, Zhang et al. (2016) discovered dynamic changes in bacterial communities in L. chinensis at different stages of silage and after aerobic exposure. To the best of our knowledge, however, only a few studies have looked at dynamic changes in fungal populations during L. chinensis silage and after aerobic exposure. This study looked at how microbial quantities, fermentation products, and Illumina MiSeq sequencing changed during the silo opening and the aerobic deterioration test in wilted L. chinensis inoculated with L. plantarum and L. buchneri. This study sought to determine whether changes in microbial communities could account for changes in aerobic stability and ensiling fermentation.
Materials and methods
Ensiling and bacterial inoculants
At the boot stage on 29 July 2021, L. chinensis was collected from an experimental field at China’s Inner Mongolia Minzu University. After a 5-h wilting period, the grass was chopped to an estimated particle length of 20 mm with a crop cutter to achieve the desired dry matter (DM) of 436 g/kg. Both L. plantarum (No. 6026) and L. buchneri (No. 20294) were successfully inoculated in L. chinensis silage at 1 × 106 colony-forming unit (CFU)/g of fresh matter (FM). For the control group, the same amount of distilled water was sprayed. Each 300 g of non-inoculated and inoculated materials were packed tightly into plastic pouches (BN-10, 250 × 350 mm; Wangnuo, Beijing, China) using a commercial vacuum sealer (ZK-320; Ouxin, Beijing, China). Triplicate silages were prepared for each treatment, and silos were stored at room temperature for 14 and 56 days, respectively.
Aerobic spoilage
After opening, 150 g of silage (half the amount) was placed in a 500-ml polyethylene (PE) bottle without being compacted. These uncapped bottles were exposed to air for 7 days at 25°C.
Chemical analysis and microbial enumeration
For pH and organic acid determination, each fresh sample (20 g) was homogenized with sterilized water (180 ml) within the blender for 1 min, followed by filtration using a membrane (0.22 μm). The pH of this extract was determined immediately using the glass electrode pH meter (SX-620, Sanxin, Shanghai, China). An ion-exclusion polymeric high-performance liquid chromatography (HPLC) equipped with an refractive index (RI) detector was used to evaluate the composition of fermented products (Wu and Nishino, 2016).
The fresh material was oven-dried under 65°C for 48 h to obtain DM content. After that, each sample was grounded in a Wiley mill with a 1-mm sieve (ZM200, Retsch GmbH). The standard Association of Official Analytical Chemists (AOAC, 1990) procedures were utilized for measuring crude protein (CP) amount. Acid detergent fiber (ADF) and neutral detergent fiber (NDF) were assessed using Van Soest et al.’s (1991) method, whereas WSCs were quantified using the phenol-sulfuric acid assay (Wu and Nishino, 2016).
A serial dilution (from 10–1 to 10–6) was performed on a clean bench. To count LAB, de Man, Rogosa, and Sharpe agar (CM0361B, Oxoid Ltd., United Kingdom) was used, while violet red bile agar (CM0107B, Oxoid Ltd., United Kingdom) was used for enterobacterial counting. Molds and yeasts were quantified using spread plates in potato dextrose agar (CM0139B, Oxoid Ltd., United Kingdom) at pH 3.5 (maintained with sterile lactic acid). Colony counts in CFU/g of FM were used to calculate the viable microbial number.
Microbial community analysis
The refrigerated silage (10 g) was blended for 2 h at 120 rpm with sterile PBS (40 ml; pH 7.4) using an electronic oscillator. The sample was then filtered using double-gauze masks. After 10 min of centrifugation of the filtrate at 4°C and 13,000 × g, the supernatant was removed and the pellet was kept on dry ice. Majorbio Bio-Pharm Technology in Shanghai, China, was used for metagenomic sequencing, which included PCR amplification and DNA extraction, followed by Illumina MiSeq sequencing and final sequencing data processing. The UPARSE version 7.1 (Edgar, 2013) was used to cluster the OTUs at the 97% similarity threshold. Following the identification and elimination of Chimeric sequences, the RDP Classifier (version 2.2) was used for the taxonomic analysis of typical OTU sequences against a 16S rRNA database such as Silva version 138, with a confidence level of 0.7 (Yang et al., 2019). The free and open Majorbio Cloud Platform was used to analyze sequencing data from fungi and bacteria.
Statistical analyses
The data were analyzed statistically using JMP version 13 software (SAS Institute, Japan) and bidirectional ANOVA, with inoculation and ensilage duration as primary factors. To assess the inoculation effect, the unidirectional ANOVA and subsequent Tukey’s test-based multiple comparisons were used. To determine significance, P < 0.05 was used.
Results and discussion
Microbial and chemical compositions of pre-ensiled Leymus chinensis
Table 1 shows microbial and chemical compositions of pre-ensiled L. chinensis. DM level in L. chinensis was 436 g/kg of FM, which was comparatively higher than that of tropical legume and grass silages (Heinritz et al., 2012). CP level was found to be 93.71 g/kg of DM, which is greater than the 84.40 g/kg of DM found by Zhang et al. (2016). WSC served as the critical substrate in silage fermentation, with a level of greater than 60 g/kg of DM ensuring the desired grade of fermentation. This study determined WSC as 32.58 g/kg of DM in pre-ensiled L. chinensis, indicating that it was difficult to achieve good preservation without adding anything during the ensilage process. The epiphytic LAB count was 3.67 log CFU/g of FM, yeast count was 5.47 log CFU/g of FM, and enterobacterial count was 5.12 log CFU/g of FM. The epiphytic LAB count on fresh crops was usually considered an important factor in deciding whether bacterial inoculants should be applied in silage materials and predicting silage fermentation quality. The LAB count on forage material must be above 5.00 log CFU/g of FM to ensure a successful ensilage (Cai et al., 1998). The L. chinensis silage fermentation quality was improved by introducing LAB inoculants because of the low LAB count and high harmful bacteria count.
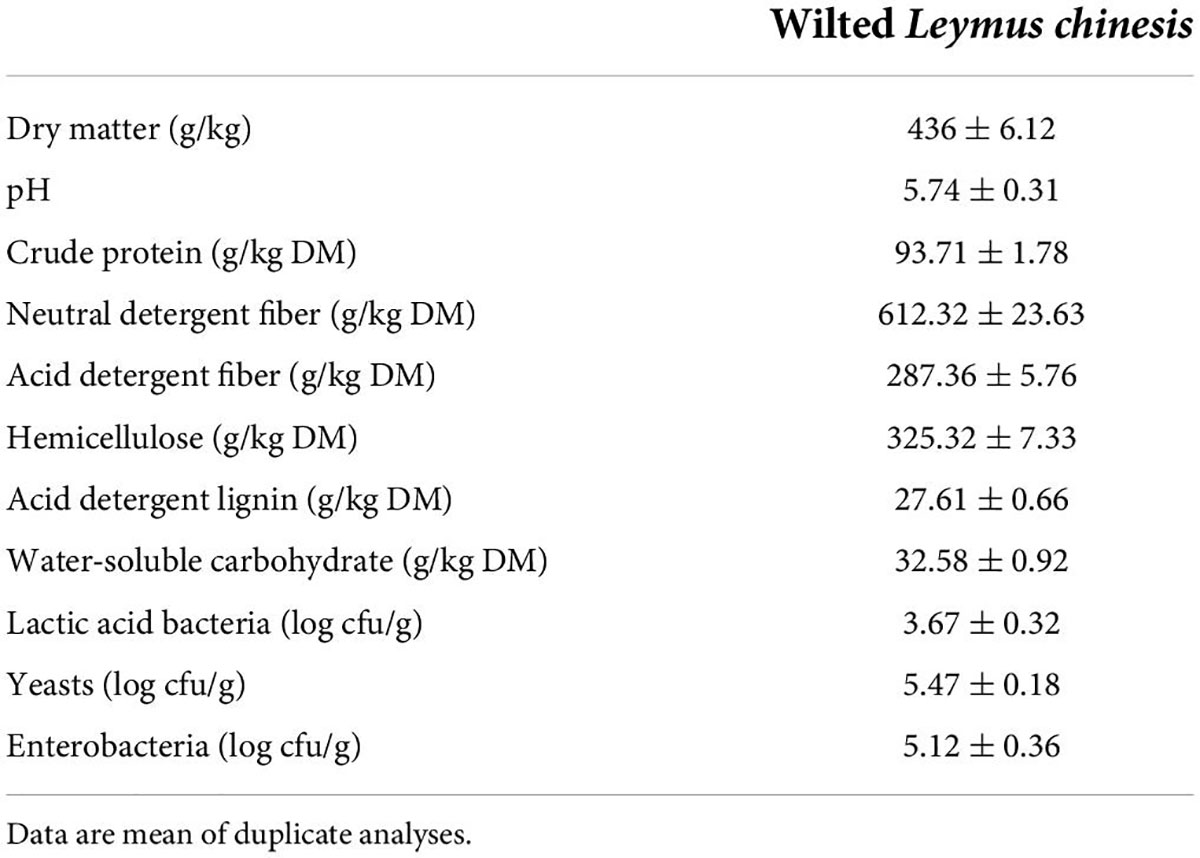
Table 1. Chemical and microbial compositions of pre-ensiling Leymus chinesis.
Roles of Lactiplantibacillus plantarum and Lentilactobacillus buchneri in silage quality during ensiling and after aerobic exposure
According to Table 2, the lactic acid level was 8.33 g/kg of DM on the 14th day for the untreated silage, while the acetic acid level was 3.38 g/kg of DM. Both contents increased when ensilage in untreated silage was prolonged, but propionic acid was not detected at 14 or 56 days. The content of ethanol was 3.42 g/kg of DM, while the content of 2,3-butanediol was 2.66 g/kg of DM. It was reported that 2,3-butanediol can be produced from sugar by yeasts, Bacillus spp., and enterobacteria, and citric and malic acids by the two types of LAB (Mcdonald et al., 1991). L. plantarum, a homofermentative LAB species, was previously used as an additive to improve alfalfa fermentation quality (Filya et al., 2007; Contreras-Govea et al., 2013; Zhang et al., 2015). In comparison with the untreated control, inoculation with L. plantarum increased LAB and lactic acid and decreased pH and levels of butyric acid, 2,3-butanediol, and ethanol. Heterofermentative LAB-treated silages had higher levels of lactic acid, 1,2-propanediol, and acetic acid (Li and Nishino, 2011a). In this study, L. buchneri inoculation increased lactic acid, acetic acid, 1,2-propanediol, and LAB number while decreasing pH, butyric acid, 2,3-butanediol, and ethanol content. Regardless of the ensiling process, enterobacteria could become dominant in untreated silage. However, because of the rapid acidification and antagonistic effects of L. plantarum and L. buchneri, the enterobacterial count rapidly decreased, and the ethanol production efficiency was limited.
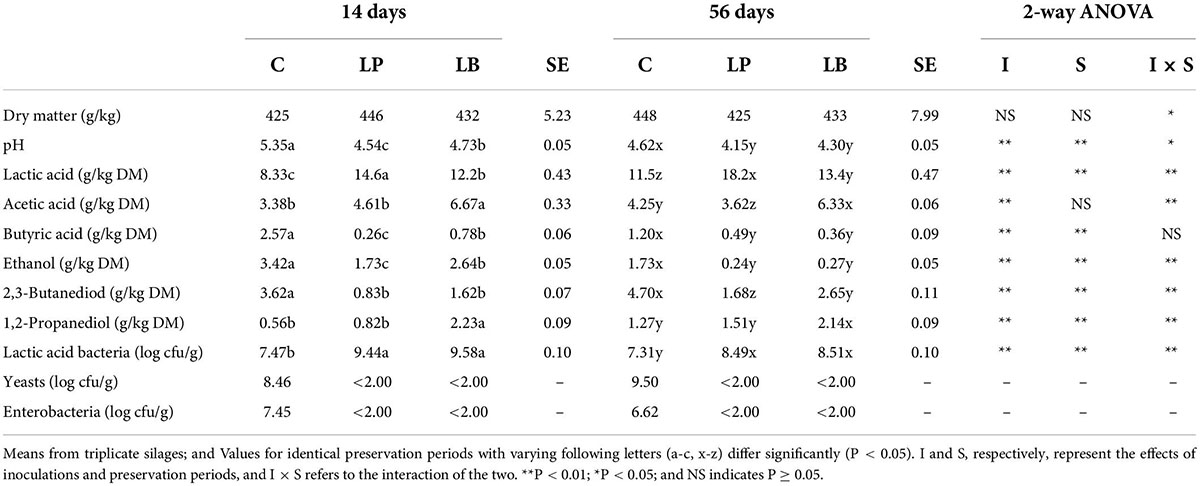
Table 2. Fermentation products and microbial counts of wilted Leymus chinesis silage without and with Lactiplantibacillus plantarum (LP) or Lentilactobacillus buchneri (LB) and subjected to 14 and 56 days of preservation.
Roles of Lactiplantibacillus plantarum and Lentilactobacillus buchneri in aerobic stability
Heating was observed on the third day after silo opening for L. plantarum-treated and non-inoculated silages, which were opened on the 14th day. The pH and yeast counts of the L. plantarum-treated silages were raised to > 6.0 and > 108 CFU/g, respectively (Table 3). However, no heating was observed, followed by aerobic exposure in L. buchneri-treated silages opened on the 14th day, and significantly higher acetic acid and 1,2-propanediol contents were observed in the 7-day spoilage test after silo opening. In the 56-day non-inoculated silage, heating was observed on the 5th day after silo opening because yeast quantities exceeded 108 levels in the 7-day spoilage test. Yeast counts exceeded 105, indicating that the aerobic stability of the silage had been compromised (Mcdonald et al., 1991). Whole crop corn silage showed decreasing aerobic stability after 14 days of ensiling, regardless of LAB inoculation, as the yeast count increased (Li and Nishino, 2011b). There was no heating due to aerobic spoilage in the 56-day silage inoculated with L. plantarum and L. buchneri. Despite the improved preservation quality, Cai et al. (1999) discovered that the L. plantarum inoculation was ineffective at preventing yeast growth, resulting in aerobically deteriorated alfalfa, Italian ryegrass, and sorghum silages. However, Li et al. (2020) discovered that L. plantarum-producing class IIa bacteriocins improved alfalfa silage fermenting quality, reduced mold and yeast levels, and improved aerobic stability. Acetic acid promotes aerobic stability by inhibiting the growth of unfavorable bacteria such as molds and yeasts (Wilkinson and Davies, 2013). Therefore, the aerobic stability of the L. buchneri-treated silage of L. chinensis is likely.
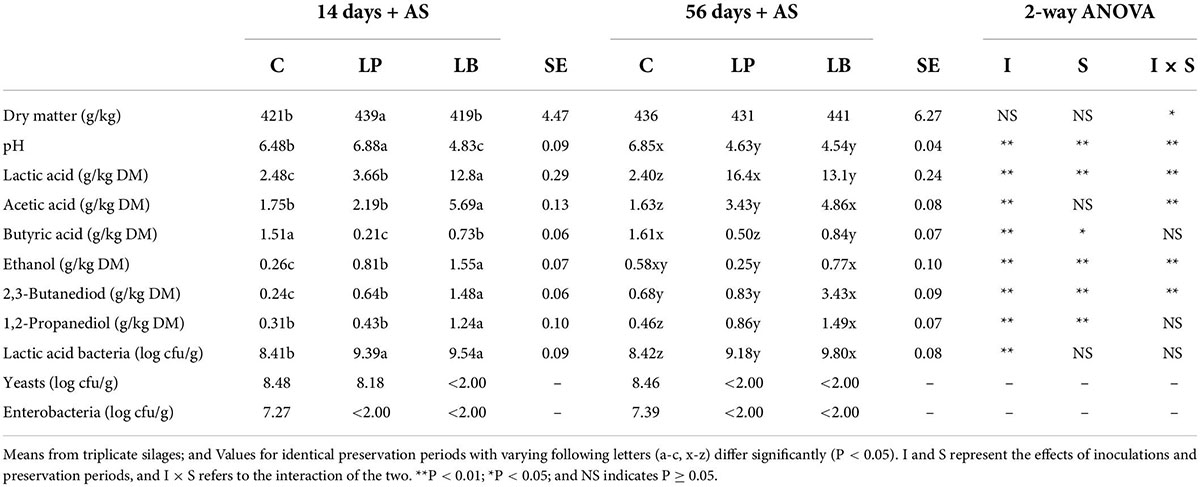
Table 3. Fermentation products and microbial counts of wilted Leymus chinesis silage without and with Lactiplantibacillus plantarum (LP) or Lentilactobacillus buchneri (LB) and subjected to 14 and 56 days of preservation followed by 7-day AS (aerobic stability) test.
Role of Lactiplantibacillus plantarum and Lentilactobacillus buchneri in microbial community diversities in ensiling and after aerobic exposure
Next-generation sequencing can provide detailed information on the microflora composition of whole crop corn, alfalfa, and sugarcane top silages (Guo et al., 2018; Hu et al., 2018; Wang et al., 2020b). However, there is currently no research on how homofermenter L. plantarum or heterofermenter L. buchneri affects dynamic changes in microbial communities in L. chinensis silage and after aerobic exposure. In Table 4, all samples had coverage values around 0.999, indicating that the majority of bacterial and fungal microflora were collected. For all samples, the 16S rRNA gene and the ITS gene were sequenced using high-throughput amplicon sequencing, yielding a total of 3,157,850 quality sequence reads. These readings were clustered into 8,935 OTUs with a dissimilarity of 3%. At 14 days, OTUs and Chao1 levels were higher after L. buchneri and L. plantarum inoculation than in the non-treated control, implying an increase in bacterial abundance with LAB inoculation compared to untreated silage. At 56 days, there was a reduction in OTUs and Chao1 compared to untreated silage after L. buchneri and L. plantarum inoculation. Guo et al. (2018) found that at 60 days, the levels of OTUs and Chao1 were higher in the LAB-inoculated alfalfa silage than in the non-inoculated samples, regardless of the fermentation method. On the 14th day, the Shannon index for bacterial diversity in L. buchneri-treated silage was the lowest, most likely due to microbial growth inhibition and subsequent bacterial diversity decline caused by lower pH in L. chinensis silage. The Chao, ACE, and number of OUTs decreased in spoilage silage after a 7-day aerobic deterioration test, indicating that aerobic microorganisms were not resistant to acids multiplied throughout the aerobic degradation test (Wilkinson and Davies, 2013). In addition, a denaturing gradient gel electrophoresis study revealed that when dominant bacteria become common in Italian ryegrass and alfalfa silages, the microbial community becomes less diverse (Li and Nishino, 2011a; Wu and Nishino, 2016).
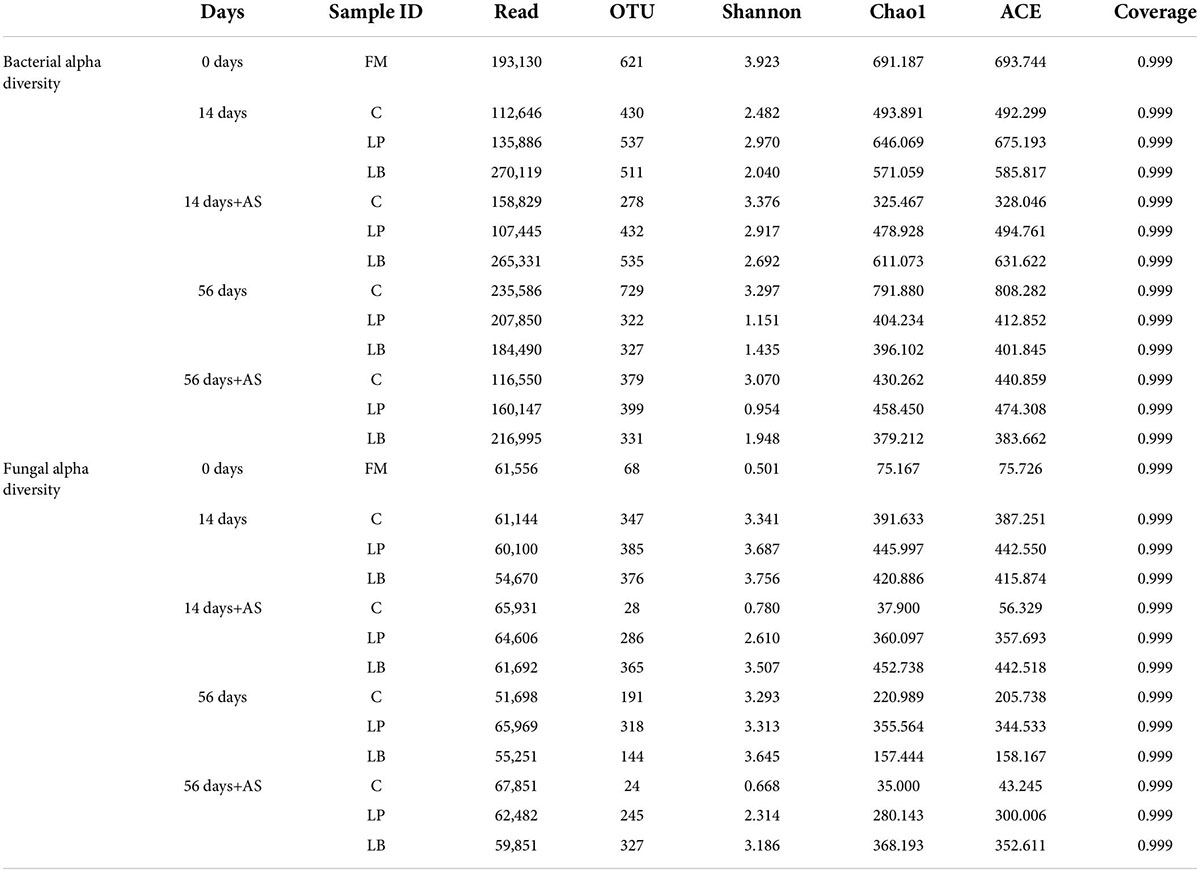
Table 4. The bacterial and fungal alpha diversity of wilted Leymus chinesis silage during ensiling and aerobic stability (AS) test.
During the aerobic deterioration test in L. chinensis silage, the richness and diversity of the fungi differed from that of the bacteria. The number of OTUs, Chao, and ACE indexes of fungi in untreated silage decreased after aerobic deterioration test than silo opening regardless of ensiling time, which was in accordance with Wang et al. (2020b). However, fewer changes in the fungi alpha diversity were observed in silage-inoculated L. buchneri on the 7-day aerobic test compared with silo opening at 14 days. One possible reason is that the high levels of acetic acid and 1,2-propanediol in silage-inoculated L. buchneri prevented certain fungi from dominating.
The principal coordinates the fungal and bacterial microflora analysis in wilted silages of L. chinensis inoculated with and without L. plantarum or L. buchneri were shown in Figure 1. As shown in Figure 1A, the inoculation of L. plantarum and L. buchneri affected the bacterial microflora of silage during ensiling. Similar results were observed in Broussonetia papyrifera leaves silage inoculated with L. plantarum (He et al., 2021). As the CAS, CAE, and LPAS samples were detected heating after aerobic exposure, they were separated from the other samples. There was a significant separation of fungal populations between fresh material and silages (Figure 1B). As fungal community changes in the CAS and CAE samples induced aerobic degradations, they were separated from the other samples.
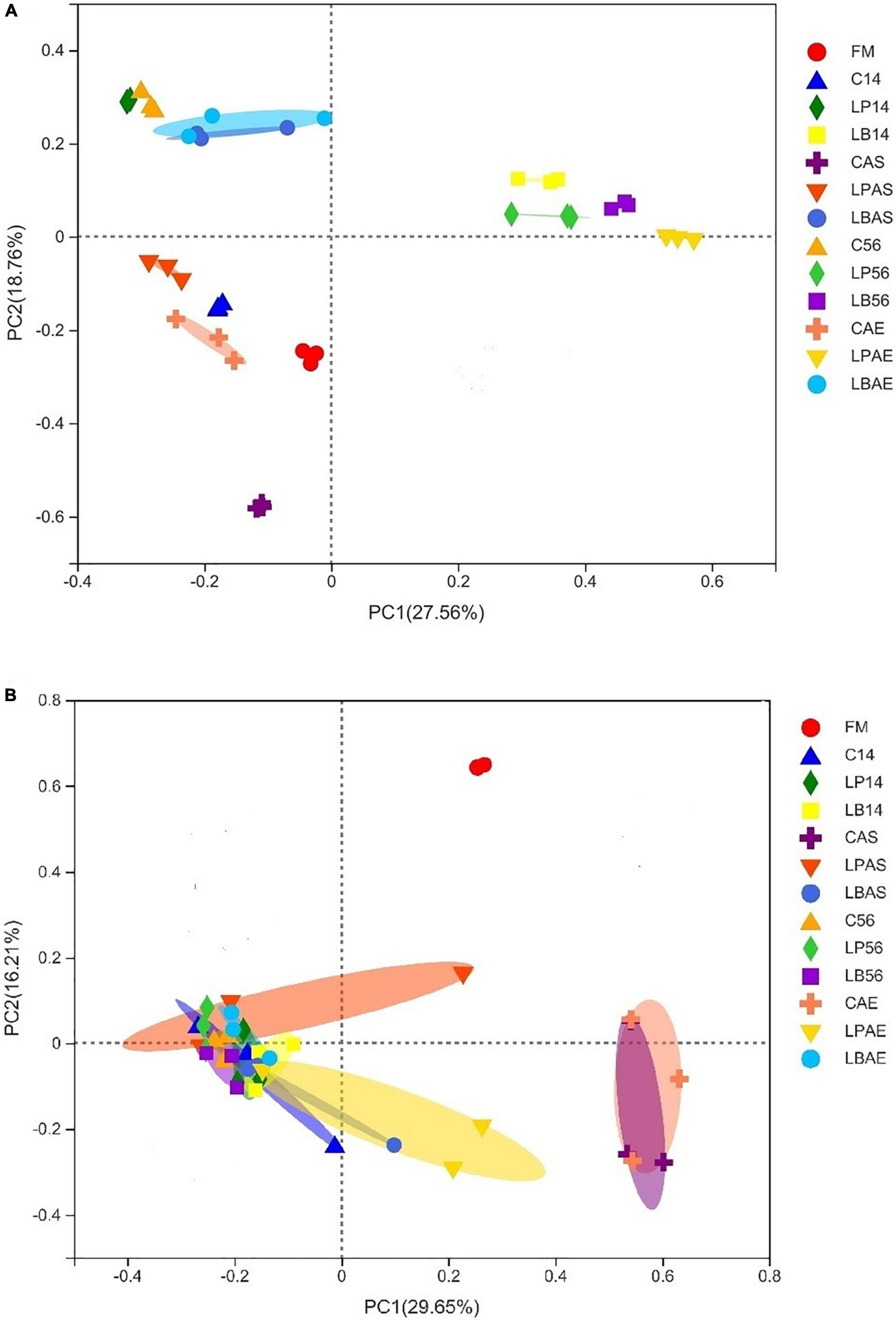
Figure 1. Principal coordinate analysis (PCoA) findings for the bacterial (A) and fungal (B) community in wilted Leymus chinesis silage without and with L. plantarum or L. buchneri. FM stands for fresh material; C stands for control; LP refers to the L. plantarum inoculation, and LB refers to the L. buchneri inoculation. The numbers following C, LP, and LB stand for the ensiling days. AS and AE of the following C, LP, and LB represent the aerobic stability test.
Roles of Lactiplantibacillus plantarum and Lentilactobacillus buchneri in microbial community dynamics in ensiling as well as after aerobic exposure
Figures 2A, 3A show the bacterial communities of pre-ensiled crop, silages, and after aerobic exposure at the phylum and family levels. Proteobacteria, Firmicutes, Bacteroidota, and Actinobacteriota were the most common phyla in pre-ensiled crops (Figure 2A). However, Proteobacteria and Firmicutes were the most common phyla in all samples after ensiling and aerobic exposure, accounting for more than 84.65% of all detected sequences, according to Figure 3A. The dominant families in pre-ensiled crops were Pseudomonadaceae and Erwiniaceae, which were found in fresh materials such as alfalfa, corn, and Italian ryegrass (Ni et al., 2017; Wang et al., 2020a). Despite the fact that their sections died during the ensiling process, Lactobacillacae and Leuconostocaceae emerged as the most dominant families after 14 and 56 days of ensiling, with or without LAB inoculants, respectively. Similarly, Ni et al. (2017) discovered a 70% overall Lactobacillacae and Leuconostocaceae ratio in Italian ryegrass silage after 60 days of ensiling. Lactobacillacae levels were higher in silage inoculated with L. plantarum and L. buchneri than in untreated silage at opening day-14 and -56 silos. Similar results were obtained in whole plant hybrid Pennisetum silage that was ensiled with L. plantarum (Shah et al., 2020). However, after aerobic exposure, the bacterial microflora composition of LAB-inoculated silages differed significantly from that of non-inoculated silages. The non-inoculated and L. plantarum-treated silages had higher Acetobacteraceae and Bacillaceae richness but lower Lactobacillaceae richness than the L. buchneri-treated silage in the 7-day spoilage test after opening day-14 silos. In the 7-day spoilage test after opening day-56 silos, untreated silage had more Acetobacteraceae and Bacillaceae and less Lactobacillaceae than LAB-inoculated silage. Acetobacteraceae, an organic acid-consuming bacterial species that occasionally causes aerobic deterioration, was discovered in silage by da Silva et al. (2020). Furthermore, after the start of spoiling, the Bacillaceae, which were generally present in the air-exposed silage, increased (Okatsu et al., 2019). After aerobic exposure, the L. buchneri-treated silage had lower Acetobacteraceae and Bacillaceae richness, as well as higher Lactobacillaceae richness, than the non-inoculated silage, regardless of ensiling process, implying that L. buchneri inoculation after aerobic exposure improves silage aerobic stability.
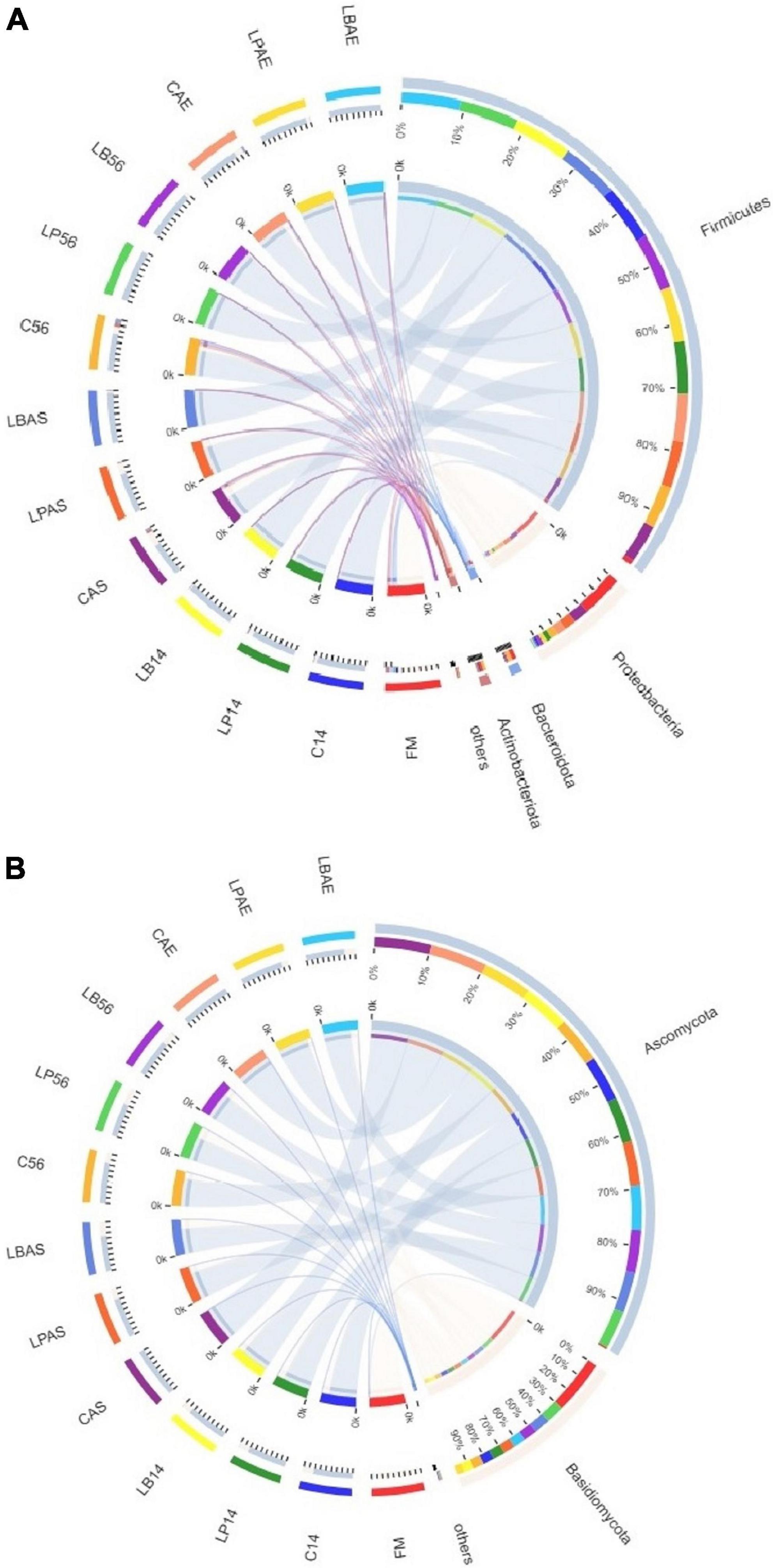
Figure 2. The phylum-level bacterial (A) and fungal (B) community in wilted Leymus chinensis silage inoculated with and without L. plantarum or L. buchneri. FM stands for fresh material; C stands for control; LP refers to the L. plantarum inoculation, and LB refers to the L. buchneri inoculation. The numbers following C, LP, and LB are the ensiling days. AS and AE of C, LP, and LB represent the aerobic stability test.
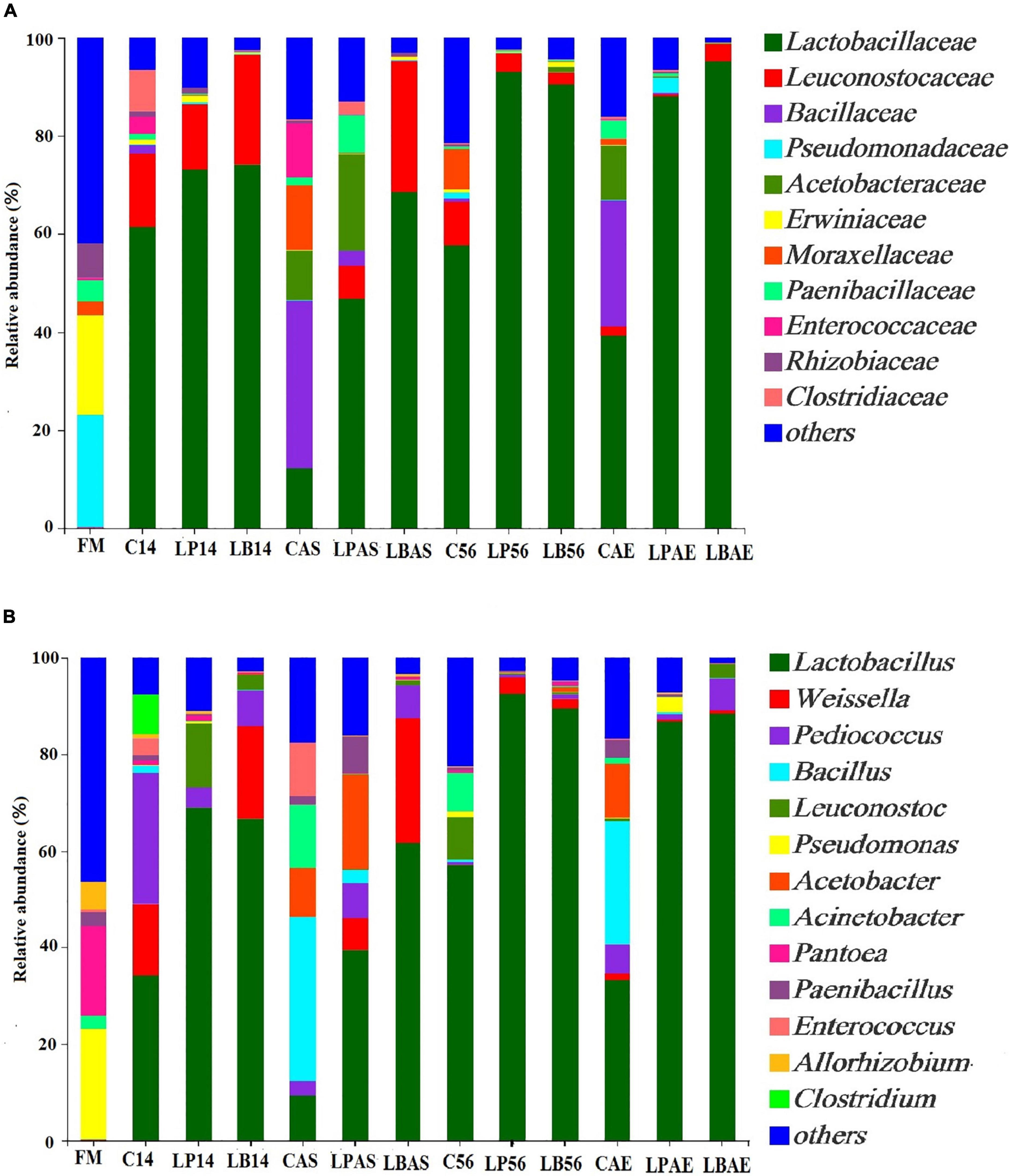
Figure 3. (A) Family- and (B) genus-level bacterial community in wilted Leymus chinensis silage inoculated with and without L. plantarum or L. buchneri. FM stands for fresh material; C stands for control; LP refers to the L. plantarum inoculation, and LB refers to the L. buchneri inoculation. The numbers following C, LP, and LB indicate the ensiling days. AS and AE following C, LP, and LB represent the aerobic stability test.
Figure 3B depicts the genus-level bacterial community of pre-ensiled crops, silages, and after aerobic exposure. Those predominant genera of pre-ensiled crops included Pseudomonas, Pantoea, Acinetobacter, Paenibacillus, and Pedobacter. Shah et al. (2020) reported similar results that Pseudomonas and Pantoea were dominated in fresh material of whole-plant hybrid Pennisetum. Lactobacillus was the most prevalent genus in every sample taken between the 14th and 56th days of ensiling, which is consistent with previous findings (Liu et al., 2019; Wang et al., 2020b). However, after aerobic exposure, the non-inoculated silage had higher Acetobacteraceae and Bacillaceae richness and lower Lactobacillaceae richness than the L. buchneri-treated silage. In the 7-day spoilage test after opening day-14 silos, the Acetobacter proportion increased from 0.32 to 19.66%, while the Lactobacillus proportion decreased from 68.99 to 39.41% in L. plantarum-inoculated silage. Therefore, these findings indicated that aerobic exposure decreased the Lactobacillus proportion in silage and that L. plantarum inoculation did not maintain relative Lactobacillus richness after silos were opened at 14 days. Following a 7-day aerobic exposure, ensiling with L. buchneri helped maintain high relative Lactobacillus richness (66.71% at 14 days, 89.52% at 56 days), implying that L. buchneri plays a Lactobacillus-enriching role in the silage. This was similar to what was discovered in alfalfa silage (Guo et al., 2018). In this study, once aerobic deterioration occurred, the amount of Acetobacter and Bacillus increased in untreated silage. Acetobacter, a non-fermenting aerobic bacterium, can be detected using denaturing gradient gel electrophoresis in whole crop corn silage (Li and Nishino, 2011b). The Acinetobacter bacteria were visible in all non-inoculated silage after 7 days of aerobic exposure, as well as in L. plantarum-treated silage after 7 days of aerobic exposure in 14 days of ensilage, though they did not grow in L. buchneri-inoculated silage after 7 days of aerobic exposure. Therefore, after a 7-day aerobic exposure, a high abundance of Acinetobacter promoted spoiling microbial development, reducing the aerobic stability of L. chinensis silage. Similar to Acinetobacter, Bacillus, and Lysinibacillus, increased aerobic species richness was observed in non-inoculated silage following aerobic spoilage, but not in L. plantarum-and L. buchneri-treated silages. Serratia nematodiphila, Myroides odoratimimus, Lysinibacillus fusiformis, Stenotrophomonas maltophilia, Bacillus pumilus, and Acinetobacter soli were discovered in the silage of corn stalk at the silo opening, and the aerobic bacteria were attributed as the likely cause of the aerobic spoilage (Liu et al., 2013).
Figures 2B, 4A show the fungal population of the pre-ensiled crop, silages, and after aerobic exposure at the phylum and family levels. Ascomycota and Basidiomycota were the most common phyla of pre-ensiled crops and silages, respectively (Figure 2B). Following aerobic exposure, the dominant phylum in CAS and CAE samples was Ascomycota alone, accounting for 99.89% of all sequences found. The Filobasidiaceae family was the most abundant in the pre-ensiled crop, accounting for more than 94.45% of total abundance (Figure 4A). However, the Aspergillaceae, Pleosporaceae, Cadosporiacaeae, and Bulleribasidiaceae were the dominant families in every sample after ensilage, accounting for more than half of all detected sequences. After aerobic exposure, changes in the fungal microflora were observed in the non-inoculated silage, with Aspergillaceae and Sordariaceae emerging as the dominant families, accounting for more than 97% of the total abundance. Fungi were also found in the corn silage, but there was no evidence of Sordariaceae involvement in the aerobic deterioration (Sandona et al., 2019). Mycotoxins, according to Gallo et al. (2021), are a group of secondary metabolites produced by Aspergillaceae during aerobic exposure to corn silage. Therefore, secondary fermentation was most likely triggered in the non-inoculated silage. When compared to 7 days after aerobic exposure, the changes in fungal microflora in L. buchneri-treated silage were insignificant at silo opening. The total amount of yeast after aerobic exposure of L. buchneri-inoculated silage was less than 102 CFU/g, which could explain why the silage did not heat after 7 days of aerobic exposure. Furthermore, Wang et al. (2020b) discovered that the total amount of yeasts in Lactobacillus hilgardii-inoculated silage was lower than in untreated silage, which could explain why the pH and temperature of the silage treated with L. hilgardii did not reach their maximums after aerobic exposure. Figure 4B depicts the fungal population of pre-ensiled crop, silage, and after aerobic exposure at the genus level. Filobasidium was the most common genus in the fungal microflora of the pre-ensiled crop, appearing in nearly 94% of the sequences. The findings of Zhang et al. (2021) were also consistent, with Filobasidium being found to be the richest fungus in the fresh wheat sample. Despite the fact that Aspergillus, Filobasidium, and Penicillium were the most common fungi discovered in this study, Aspergillus was the most common fungus at the genus level after 56 days of silage preservation. After aerobic exposure, the proportion of Aspergillus was higher than at silo opening in untreated silage. However, after aerobic exposure, the proportion of Aspergillus in the L. buchneri-treated silage was lower than in the non-inoculated silage because LAB (heterofermentative) inoculation into the silage of L. chinensis likely inhibited the aerobic deterioration caused by fungi post-spoilage growth. This finding contradicts the findings of Wang et al. (2020b). Fungal spoilage of animal feed silage is common during feed-out. Aspergillus fumigatus and Penicillium roqueforti were the most common spoilage molds in sugar beet press pulp and maize silage (Nout et al., 1993). The proportion of Penicillium was higher after aerobic exposure than after 14 days of ensiling with L. plantarum-inoculated silage in this study. Therefore, this could explain why heating was detected on the third day after silo opening for L. plantarum-inoculated silage.
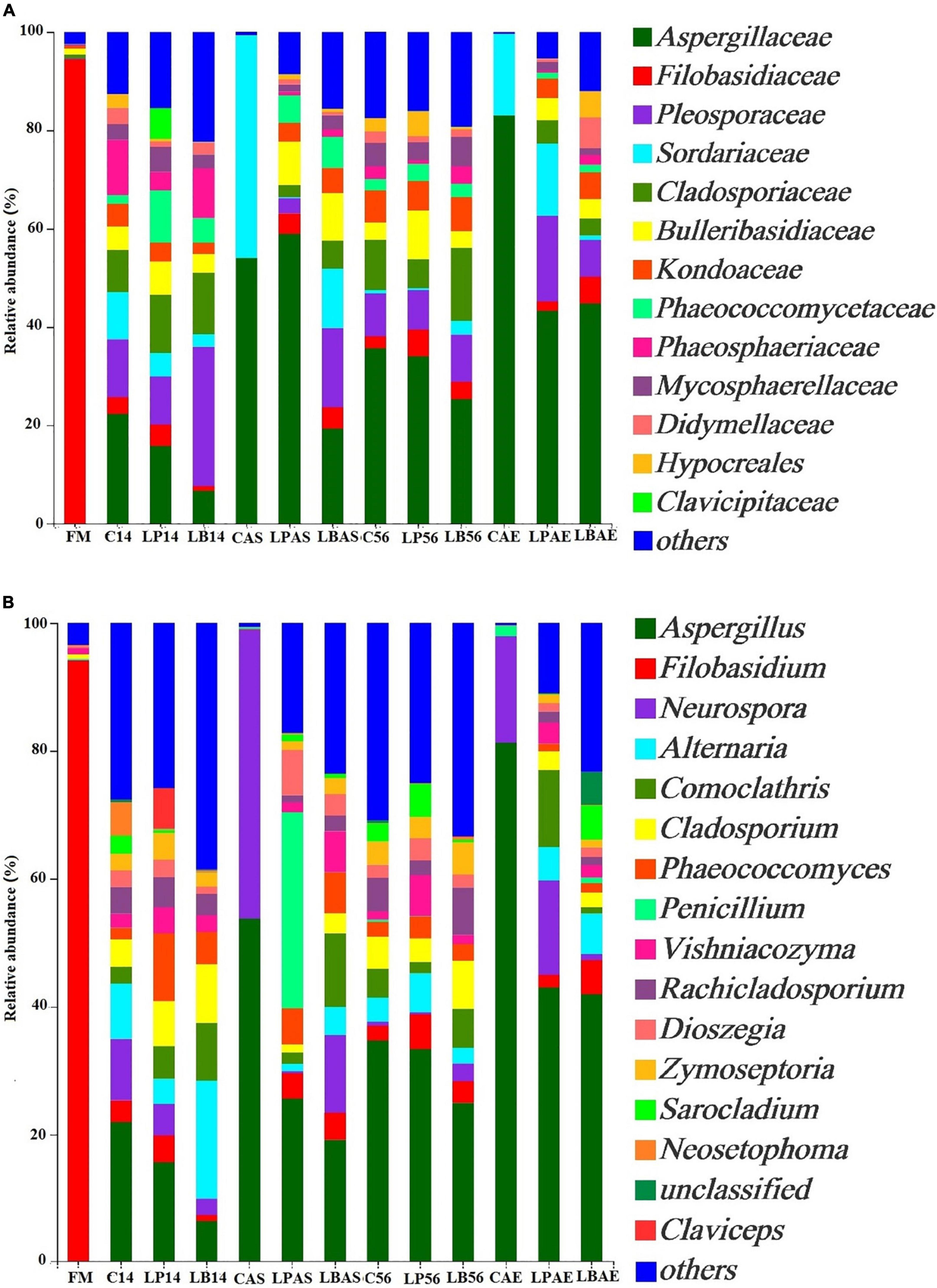
Figure 4. Fungal community at the family (A) and genus (B) levels in wilted Leymus chinensis silage without and with L. plantarum or L. buchneri. FM stands for fresh material; C stands for control; LP refers to the L. plantarum inoculation, and LB refers to the L. buchneri inoculation. The numbers following C, LP, and LB indicate the ensiling days. AS and AE following C, LP, and LB represent the aerobic stability test.
Correlation analysis of the microbial community with fermentation products
Ensilage and aerobic exposure were both complex processes involving interactions between various fermentation products and microflora. Beneficial bacteria reduce aerobic deterioration by producing a variety of metabolites, in addition to improving fermentation quality. For example, LAB-producing class IIa bacteriocins could improve silage fermentation quality, lower mold and yeast counts, and improve aerobic stability (Li et al., 2020). Mold has been linked to mycotoxin production, whereas enterobacteria can ferment WSCs into products such as lactic and acetic acids (Mcdonald et al., 1991). To better understand the correlations between microbial dynamics and fermented products (e.g., acetic acid, lactic acid, butyric acid, ethanol, 2,3-butanediol, and 1,2-propanediol) during L. chinensis ensilage, and following aerobic exposure, a genus-level heatmap of the Spearman correlation was plotted for the major bacteria and fungi (Figures 5, 6). The Lactobacillus and Leuconostoc genera had positive relationships with acetate acid, 2,3-butanediol, and 1,2-propanediol concentrations, according to the Spearman correlation analysis (Figure 5). In contrast, the Paenibacillus, Bacillus, Lysinibacillus, and Acetobacter genera had no relationships with acetate acid, 2,3-butanediol, or 1,2-propanediol levels. Lactic acid content was found to be positively correlated with the Lactobacillus genus (R = 0.88), whereas Enterococcus, Pediococcus, and Acetobacter richness were found to be negatively correlated with lactic acid, with correlation values of –0.79, –0.44, and –0.50, respectively. The relationship between lactic acid and bacterial richness can be described in a variety of ways based on the above findings. Despite the fact that lactic acid-producing cocci (e.g., Leuconostocs and Pediococcus) began lactic fermentation early in the ensilage stage, these bacteria were unable to survive at low pH. The Lactobacillus bacterium produces lactic acid in silage and has a significant impact on pH reduction during the late ensilage stage (Cai et al., 1998). Wang et al. (2020b) discovered a positive relationship between silage lactic acid content and Lactobacillus richness. Silage is deemed undesirable when Acetobacter spp. is present due to its ability to cause aerobic deterioration as well as acetic and lactic acid breakdown, resulting in CO2 and water production (Li and Nishino, 2011c). The Kondoa, Alternaria, Cladosporium, Rachicladosporium, and Zymoseptoria genera were positively correlated with lactic acid, 2,3-butanediol, and 1,2-propanediol concentrations, according to the Spearman correlation analysis (Figure 6). The acetate acid and the richness of Comoclathris, Phaeococcomyces, Alternaria, Cladosporium, Rachicladosporium, and Zymoseptoria were found to have positive correlation factors of 0.60, 0.56, 0.55, 0.65, 0.61, and 0.56. In contrast, negative relationships of acetate acid contents with the Aspergillus (R = –0.48) and Penicillium (R = –0.37) genera were discovered. Although Aspergillus has been shown to inhibit acetic acid fermentation, no studies have demonstrated that it has a significant impact on acetic acid reduction. In this study, Aspergillus, a common mold that produces mycotoxin in corn silages (Gallo et al., 2021), was found to have high richness after aerobic exposure to untreated silage. In the early stages of silage production, Alternaria and Cladosporium were discovered (Liu et al., 2019). There is less information on the fungal microbiota during the ensilage process and after aerobic exposure. Therefore, the roles of these fungi during forage ensilage, followed by aerobic exposure, are unknown.
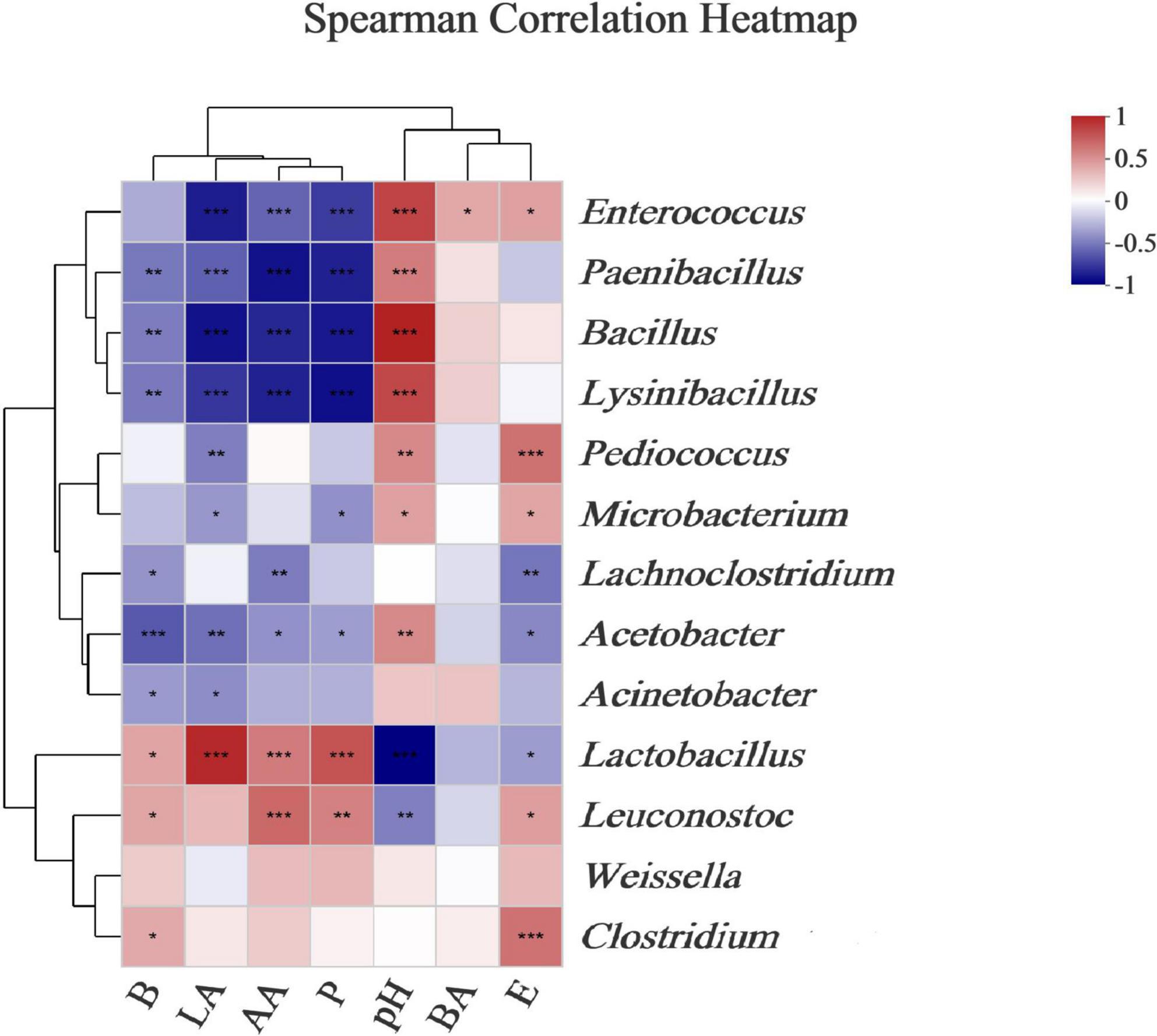
Figure 5. Correlations of bacterial richness with the fermented products. Heatmaps analyzing Spearman’s rho, as well as p-values for the pairwise comparisons of bacterial genera and fermented products. *, 0.01 < P < 0.05; **, 0.001 < P < 0.01; and ***, P < 0.001. LA, AA, and BA, respectively, stand for the lactic, acetic, and butyric acids; E, Ethanol; B, 2,3-Butanediod; P, 1,2-Propanediol.
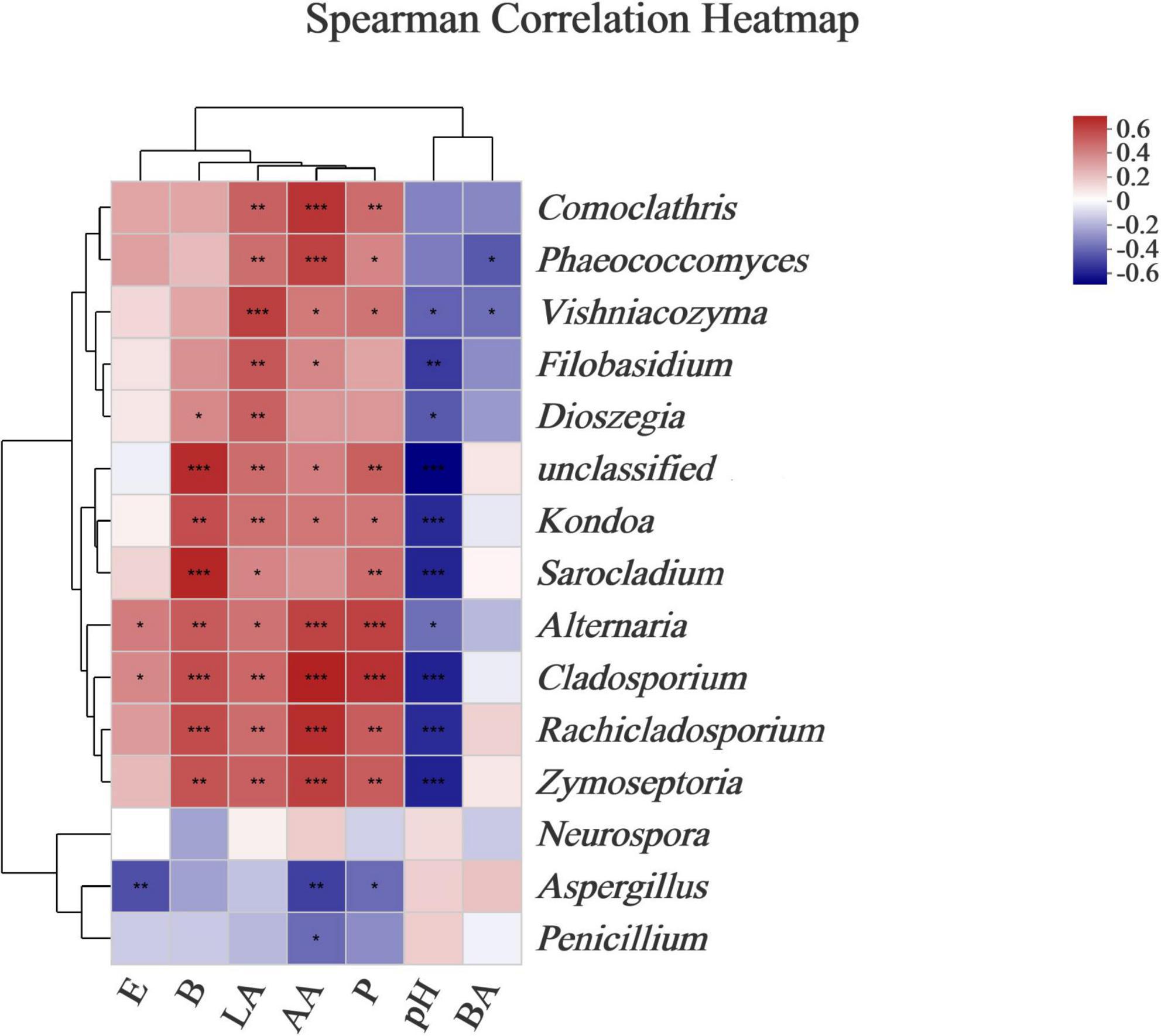
Figure 6. Correlations of fungal richness with the fermented products. Heatmaps analyzing Spearman’s rho, as well as p-values for the pairwise comparisons of fungal genera and fermented products. *, 0.01 < P < 0.05; **, 0.001 < P < 0.01; and ***, P < 0.001. LA, AA, and BA, respectively, stand for the lactic, acetic, and butyric acids; E, Ethanol; B, 2,3-Butanediod; P, 1,2-Propanediol.
Conclusion
During ensiling, silages inoculated with L. plantarum and L. buchneri had higher lactic acid and LAB counts and lower pH, butyric acid, ethanol, and 2,3-butanediol contents. Heating was observed on the third day after silo opening for L. plantarum-treated and non-inoculated silages, which were opened on the 14th day. However, no heating was observed after silo opening in L. buchneri-inoculated silages. In the 7-day spoilage test after opening 56-day silos, silages inoculated with L. plantarum and L. buchneri had higher Lactobacillus richness and lower hazardous microorganisms, namely, Acetobacter, Bacillus, Aspergillus, and Penicillium, which were linked to silage deterioration. The composition of fermentation products was related to the bacterial community, particularly Lactobacillus, Enterococcus, and Acetobacter.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Author contributions
BW and HN designed the experiment. ZH, MW, and MY conducted the experiments. ZH, MW, MY, and HN analyzed the data. BW wrote the manuscript. All authors read and approved the manuscript.
Funding
This study was funded by the Doctoral Funding of the Inner Mongolia Minzu University (Grant no. BS607), the Technical Services for the Agricultural Planting Industry (Grant no. ZG2020C0019), and the Young Talents of Science and Technology in Universities of Inner Mongolia Autonomous Region (Grant no. NJYT22054).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Brusetti, L., Crotti, E., Tamburini, A., Cittaro, D., Garavaglia, V., Rolli, E., et al. (2011). Influence of transgenic Bt176 and non-transgenic corn silage on the structure of rumen bacterial communities. Ann. Microbiol. 61, 925–930.
Cai, Y., Benno, Y., Ogawa, M., and Kumai, S. (1999). Effect of applying lactic acidbacteria isolated from forage crops on fermentation characteristics and aerobicdeterioration of silage. J. Dairy Sci. 82, 520–526. doi: 10.3168/jds.S00220302(99)75263-X
Cai, Y., Benno, Y., Ogawa, M., Ohmomo, S., and Nakase, T. (1998). Influence of Lactobacillus spp. from an inoculant and of Weissella and Leuconostoc spp. from forage crops on silage fermentation. Appl. Environ. Microbiol. 64, 2982–2987. doi: 10.1128/AEM.64.8.2982-2987.1998
Contreras-Govea, F. E., Muck, R. E., Broderick, G. A., and Weimer, P. J. (2013). Lactobacillus plantarum effects on silage fermentation and in vitro microbial yield. Anim. Feed Sci. Technol. 179, 61–68. doi: 10.1016/j.anifeedsci.2012.11.008
da Silva, E. B., Savage, R. M., Biddle, A. S., Polukis, S. A., Smith, M. L., and Kung, L. Jr. (2020). Effects of a chemical additive on the fermentation, microbial communities, and aerobic stability of corn silage with or without air stress during storage. J. Anim. Sci. 98:skaa246. doi: 10.1093/jas/skaa246
Edgar, R. C. (2013). UPARSE: highly accurate OTU sequences from microbial amplicon reads. Nat. Methods 10, 996–998. doi: 10.1038/nmeth.2604
Ennahar, S., Cai, Y., and Fujita, Y. (2003). Phylogenetic diversity of lactic acid bacteria associated with paddy rice silage as determined by 16S ribosomal DNA analysis. Appl. Environ. Microbiol. 69, 444–451.
Filya, I., Muck, R. E., and Contreras-Govea, F. E. (2007). Inoculant effects on alfalfa silage: fermentation products and nutritive value. J. Dairy Sci. 90, 5108–5114.
Gallo, A., Ghilardelli, F., Atzori, A. S., Zara, S., Novak, B., Faas, J., et al. (2021). Co-Occurrence of regulated and emerging mycotoxins in corn silage: relationships with fermentation quality and bacterial communities. Toxins 13:232. doi: 10.3390/toxins13030232
Guo, X. S., Ke, W. C., Ding, W. R., Ding, L. M., Xu, D. M., Wang, W. W., et al. (2018). Profiling of metabolome and bacterial community dynamics in ensiled Medicago sativa inoculated without or with Lactobacillus plantarum or Lactobacillus buchneri. Sci. Rep. 8:357. doi: 10.1038/s41598-017-18348-0
He, Q., Zhou, W., Chen, X., and Zhang, Q. (2021). Chemical and bacterial composition of Broussonetia papyrifera leaves ensiled at two ensiling densities with or without Lactobacillus plantarum. J. Cleaner Prod. 329:12972. doi: 10.1016/j.jclepro.2021.12972
Heinritz, S. N., Martens, S. D., Avila, P., and Hoedtke, S. (2012). The effect of inoculant and sucrose addition on the silage quality of tropical forage legumes with varying ensilability. Anim. Feed Sci. Technol. 174, 201–210. doi: 10.1016/j.anifeedsci.2012.03.017
Hu, Z., Chang, J., Yu, J., Li, S., and Niu, H. (2018). Diversity of bacterial community during ensiling and subsequent exposure to air in whole-plant maize silage, Asian-Australas. J. Anim. Sci. 31, 1464–1473. doi: 10.5713/ajas.17.0860
Li, F., Ding, Z., Adesogan, A. T., Ke, W., Jiang, Y., Bai, J., et al. (2020). Effects of class IIa bacteriocin-producing Lactobacillus species on fermentation quality and aerobic stability of Alfalfa Silage. Animals 10:1575. doi: 10.3390/ani10091575
Li, L., Yang, H., Peng, L., Ren, W., Gong, J., Liu, P., et al. (2019). Comparative study reveals insights of sheepgrass (Leymus chinensis) coping with phosphate deprived stress condition. Front. Plant Sci. 10:170. doi: 10.3389/fpls.2019.00170
Li, Y., and Nishino, N. (2011a). Bacterial and fungal communities of wilted Italian ryegrass silage inoculated with and without Lactobacillus rhamnosus or Lactobacillus buchneri. Lett. Appl. Microbiol. 52, 314–321. doi: 10.1111/j.1472-765X.2010.03000.x
Li, Y., and Nishino, N. (2011b). Effects of inoculation of Lactobacillus rhamnosus and Lactobacillus buchneri on fermentation, aerobic stability and microbial communities in whole crop corn silage. Grassl. Sci. 57, 184–191.
Li, Y., and Nishino, N. (2011c). Monitoring the bacterial community of maize silage stored in a bunker silo inoculated with Enterococcus faecium, Lactobacillus plantarum and Lactobacillus buchneri. J. Appl. Microbiol. 110, 1561–1570. doi: 10.1111/j.1365-2672.2011.05010.x
Liu, B., Huan, H., Gu, H., Xu, N., Shen, Q., and Ding, C. (2019). Dynamics of a microbial community during ensiling and upon aerobic exposure in lactic acid bacteria inoculation-treated and untreated barley silages. Bioresour. Technol. 273, 212–219. doi: 10.1016/j.biortech.2018.10.041
Liu, Q. H., Shao, T., and Zhang, J. G. (2013). Determination of aerobic deterioration of corn stalk silage caused by aerobic bacteria. Anim. Feed Sci. Technol. 183, 124–131. doi: 10.1016/j.anifeedsci.2013.05.012
Mcdonald, P., Henderson, A. R., and Heron, S. J. E. (1991). The biochemistry of silage, 2nd Edn. Lincon: Chalcombe Publications.
Muck, R. E., Nadeau, E. M. G., McAllister, T. A., Contreras-Govea, F. E., Santos, M. C., and Kung, L. Jr. (2018). Silage review: recent advances and future uses of silage additives. J. Dairy Sci. 101, 3980–4000. doi: 10.3168/jds.2017-13839
Ni, K., Minh, T. T., Tu, T. T., Tsuruta, T., Pang, H., and Nishino, N. (2017). Comparative microbiota assessment of wilted Italian ryegrass, whole crop corn, and wilted alfalfa silage using denaturing gradient gel electrophoresis and next-generation sequencing. Appl. Microbiol. Biotechnol. 101, 1385–1394. doi: 10.1007/s00253-016-7900-2
Nishino, N., Yoshida, M., Shiota, H., and Sakaguchi, E. (2003). Accumulation of 1,2 propanediol and enhancement of aerobic stability in whole crop maize silage inoculated with Lactobacillus buchneri. J. Appl. Microbiol. 94, 800–807. doi: 10.1046/j.1365-2672.2003.01810.x
Nout, M. J. R., Bouwmeester, H. M., Haaksma, J., and Dijk, V. H. (1993). Fungal growth in silages of sugarbeet press pulp and maize. J. Agric. Sci. 121, 323–326.
Okatsu, Y., Swanepoel, N., Maga, E. A., and Robinson, P. H. (2019). Impacts of some factors that effect spoilage of silage at the periphery of the exposed face of corn silage piles. Anim. Feed Sci. Technol. 247, 234–247.
Oude Elferink, S. J., Krooneman, J., Gottschal, J. C., Spoelstra, S. F., Faber, F., and Driehuis, F. (2001). Anaerobic conversion of lactic acid to acetic acid and 1, 2 propanediol by Lactobacillus buchneri. Appl. Environ. Microbiol. 67, 125–132. doi: 10.1128/AEM.67.1.125-132.2001
Peng, X., Zhang, L., Zhang, L., Liu, Z., Cheng, L., Yang, Y., et al. (2013). The transcriptional factor LcDREB2 cooperates with LcSAMDC2 to contribute to salt tolerance in Leymus chinensis. Plant Cell Tissue Organ Cult. 113, 245–256. doi: 10.1007/s11240-012-0264-0
Ranjit, N. K., and Kung, L. (2000). The effect of Lactobacillus buchneri, Lactobacillus plantarum, or a chemical preservative on the fermentation and aerobic stability of corn silage. J. Dairy Sci. 83, 526–535.
Sandona, K., Billingsley Tobias, T. L., Hutchinson, M. I., Natvig, D. O., and Porras- Alfaro, A. (2019). Diversity of thermophilic and thermotolerant fungi in corn grain. Mycologia 111, 719–729. doi: 10.1080/00275514.2019.1631137
Shah, A. A., Wu, J., Qian, C., Liu, Z., Mobashar, M., and Tao, Z. (2020). Ensiling of whole-plant hybrid pennisetum with natamycin and Lactobacillus plantarum impacts on fermentation characteristics and meta genomic microbial community at low temperature. J. Sci. Food Agric. 100, 3378–3385. doi: 10.1002/jsfa.10371
Stevenson, D. M., Muck, R. E., Shinners, K. J., and Weimer, P. J. (2006). Use of real time PCR to determine population profiles of individual species of lactic acid bacteria in alfalfa silage and stored corn stover. Appl. Microbiol. Biotechnol. 71, 329–338. doi: 10.1007/s00253-005-0170-z
Tabacco, E., Piano, S., Cavallarin, L., Bernardes, T. F., and Borreani, G. (2009). Clostridia spore formation during aerobic deterioration of maize and sorghum silages as influenced by Lactobacillus buchneri and Lactobacillus plantarum inoculants. J. Appl. Microbiol. 107, 1632–1641. doi: 10.1111/j.1365-2672.2009.04344.x
Van Soest, P. J., Robertson, J. B., and Lewis, B. A. (1991). Methods for dietary fiber, neutral detergent fiber, and nonstarch polysaccharides in relation to animal nutrition. J. Dairy Sci. 74, 3583–3597. doi: 10.3168/jds.S0022-0302(91)78551-2
Wang, T., Teng, K., Cao, Y., Shi, W., Xuan, Z., Zhou, J., et al. (2020b). Effects of Lactobacillus hilgardii 60TS-2, with or without homofermentative Lactobacillus plantarum B90, on the aerobic stability, fermentation quality and microbial community dynamics in sugarcane top silage. Bioresour. Technol. 312:123600. doi: 10.1016/j.biortech.2020.123600
Wang, M., Franco, M., Cai, Y., and Yu, Z. (2020a). Dynamics of fermentation profile and bacterial community of silage prepared with alfalfa, whole-plant corn and their mixture. Anim. Feed Sci. Technol. 270:114702. doi: 10.1016/j.anifeedsci.2020.114702
Weinberg, Z. G., Ashbell, G., Hen, Y., Azrieli, A., Szakacs, G., and Filya, I. (2002). Ensiling whole-crop wheat and corn in large containers with Lactobacillus plantarum and Lactobacillus buchneri. J. Ind. Microbiol. Biotechnol. 28, 7–11. doi: 10.1038/sj/jim/7000207
Wilkinson, J. M., and Davies, D. R. (2013). The aerobic stability of silage: key findings and recent developments. Grass Forage Sci. 68, 1–19. 2494.2012.00891.x doi: 10.1111/j.1365-2494.2012.00891.x
Wu, B., and Nishino, N. (2016). Identification and isolation of Lactobacillus fructivorans from wilted alfalfa silage with and without molasses. J. Appl. Microbiol. 120, 543–551. doi: 10.1111/jam.13031
Wu, B., Zhang, Q., Liu, Z., Yu, Z., and Nishino, N. (2014). Bacterial communities in alfalfa and corn silages produced in large-scale stack and bunker silos in China. Grassl. Sci. 60, 247–251. doi: 10.1111/grs.12063
Yang, L., Yuan, X., Li, J., Dong, Z., and Shao, T. (2019). Dynamics of microbial community and fermentation quality during ensiling of sterile and nonsterile alfalfa with or without Lactobacillus plantarum inoculant. Bioresour. Technol. 275, 280–287. doi: 10.1016/j.biortech.2018.12.067
Zhang, M., Wu, G., Wang, L., Zhang, B., Chen, J., Liu, Y., et al. (2021). Characteristics of Lactobacillus plantarum QZW5 and its effects on wheat silage under multigelation. Chem. Biol Technol. Agric. 8, 52. doi: 10.1186/s40538-021-00251-6
Zhang, Q., Li, X. J., Zhao, M. M., and Yu, Z. (2014). Isolating and evaluating lactic acid bacteria strains for effectiveness of Leymus chinensis silage fermentation. Lett. Appl. Microbiol. 59, 391–397. doi: 10.1111/grs.12097
Zhang, Q., Wu, B., Nishino, N., Wang, X., and Yu, Z. (2016). Fermentation and microbial population dynamics during the ensiling of native grass and subsequent exposure to air. Anim. Sci. J. 87, 389–397. doi: 10.1111/asj.12427
Zhang, Q., Yu, Z., and Wang, X. (2015). Isolating and evaluating lactic acid bacteria strains with or without sucrose for effectiveness of silage fermentation. Grassl. Sci. 61, 167–176.
Keywords: Leymus chinensis silage, fermentation quality, microbial community, aerobic stability, Lactiplantibacillus plantarum, Lentilactobacillus buchneri
Citation: Wu B, Hu Z, Wei M, Yong M and Niu H (2022) Effects of inoculation of Lactiplantibacillus plantarum and Lentilactobacillus buchneri on fermentation quality, aerobic stability, and microbial community dynamics of wilted Leymus chinensis silage. Front. Microbiol. 13:928731. doi: 10.3389/fmicb.2022.928731
Received: 26 April 2022; Accepted: 11 July 2022;
Published: 29 July 2022.
Edited by:
Jan Kopečný, Academy of Sciences of the Czech Republic (ASCR), CzechiaReviewed by:
Atte Von Wright, University of Eastern Finland, FinlandYanbing Li, Heilongjiang Bayi Agricultural University, China
Copyright © 2022 Wu, Hu, Wei, Yong and Niu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Huaxin Niu, bml1aHhAaW11bi5lZHUuY24=