 Peilin Yao
Peilin Yao Effarizah Mohd Esah
Effarizah Mohd Esah Chuanping Zhao
Chuanping Zhao- 1Food Technology Division, School of Industrial Technology, Universiti Sains Malaysia, Penang, Malaysia
- 2School of Biotechnology and Food Engineering, Suzhou University, Suzhou, China
Lactobacillus is widely recognized for its probiotic benefits and has been widely used in food production. While biofilms are typically associated with pathogenic bacteria, they also served as a self-protective mechanism formed by microorganisms in an adverse environments. In recent years, relevant studies have revealed the excellent characteristics of Lactobacillus biofilms, offering new insights into their potential applications in the food industry. The Lactobacillus biofilms is important in improving fermentation processes and enhancing the resilience of Lactobacillus in various conditions. This paper reviews how quorum sensing regulates the formation of Lactobacillus biofilms and explores their roles in stress resistance, bacteriostasis and food production. Additionally, it highlights the emerging concept of fourth-generation probiotics, developed through biofilm technology, as a novel approach to probiotic applications.
1 Introduction
In 2002, the joint FAO/WTO Expert Committee introduced a scientifically grounded definition of probiotics, describing them as living microorganisms that confer health benefits upon the host when ingested in adequate quantities (FAO/WHO, 2002). The most widely used probiotic strains belong to Lactobacillus spp., Bifidobacterium spp., Lactococcus spp., Leuconostoc spp., Streptococcus spp., and various species, with Lactobacillus spp being the most extensively studied. As a representative of probiotics, some Lactobacillus have good probiotic properties, including lowering intestinal pH, regulating intestinal flora, preventing lactose intolerance, and inhibiting the growth of harmful microbes in the dairy products (Garnier et al., 2020), meat products (Danielski et al., 2020), aquatic products (Aymerich et al., 2019), and fruit and vegetable products (Ma et al., 2019), among others. Furthermore, these bacteria play an important role in the fermentation processes of common foods, such as yogurt and kimchi (Mathur et al., 2020; Wang et al., 2021). To demonstrate efficient biological activities, a daily dose of probiotics of about 108–109 colony-forming units (CFU) is required during their passage through the gastrointestinal tract (Liu et al., 2019). Over time, methods to improve the stress resistance have evolved. These approaches include the use of improved strain protective agent (Sánchez et al., 2017), the addition of oligosaccharides (Dong et al., 2023), gene recombination technology (Wang et al., 2013a) and the encapsulation of probiotics in biofilms, referred to as fourth-generation probiotics (Salas-Jara et al., 2016). Both domestically and internationally, especially within the fermentation industry (Rosche et al., 2009), bioreactors-based biofilms are widely used in sewage treatment (Bao and Dai, 2013), biological fermentation (Cheng et al., 2011), and microbial fuel cells (Raganati et al., 2016).
Probiotics can form complex communities known as biofilms, which offer several advantages for microbial populations when facing various abiotic or biotic stress factors (Salas-Jara et al., 2016). The formation of biofilms allows bacteria to preferentially adhere to specific epithelium surfaces, such as the intestinal mucosa, prolonging and stabilizing their presence in the epithelium and helping to exclude pathogenic bacteria by competitive inhibition or steric hindrance (Saxelin et al., 2005). A key characteristic of biofilms is the formation of extracellular polymers, which provide mechanical stability and promote the formation of a microenvironment, which triggers quorum sensing and further regulates the maturation of the biofilm (Allison, 2003). A mature biofilm has stronger antibacterial activity and stress resistance (Lewis, 2001).
The formation of Lactobacillus biofilms is beneficial to the environmental adaptability and probiotic properties of Lactobacillus. Therefore, understanding the regulatory mechanisms of Lactobacillus biofilm formation and the biofilm’s characteristics contributes to the potential role of Lactobacillus on food fermentation industry and promoting human health. However, the relevant knowledge on this topic remains fragmented, highlighting the need for a review on the Lactobacillus biofilm and their practical applications.
2 Regulatory mechanisms of Lactobacillus biofilm formation
The formation of bacterial biofilms is dependent on the regulation of signaling systems, including quorum sensing (QS), second messenger signaling system and cyclic adenosine phosphate-receptor protein (cAMP-CRP) signaling system (Liu et al., 2020; Li and Tian, 2012) (Figure 1). In the existing reports, the QS system is mainly involved in the formation and regulation of Lactobacillus biofilm. QS, also known as “autoinduction,” refers to the induction phenomenon in which bacteria employ self-inducing molecules to communicate and coordinate their group behavior (Papenfort and Bassler, 2016). As bacteria grow, they secrete a signal molecule sensing the surrounding environment. By detecting variations in the concentration of these signal molecules, bacteria can modulate the expression of related genes, thereby regulating associated behaviors. Consequently, interfering in bacterial QS signaling to either promote or inhibit the development of lactic acid bacteria (LAB) biofilms holds substantial significance in terms of enhancing intestinal immunity, promoting health, and elevating the quality of fermented food.
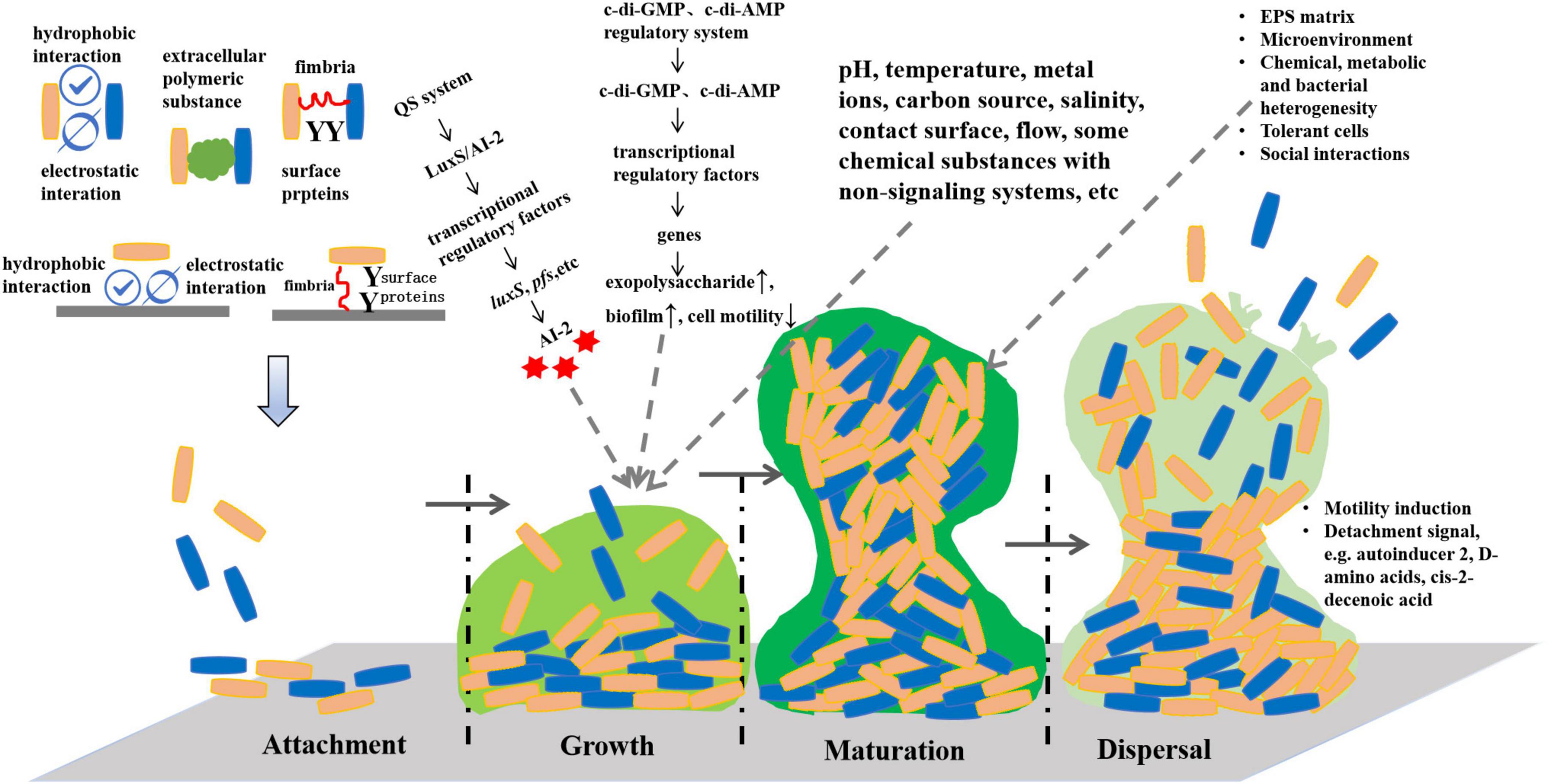
Figure 1. Formation and regulation of lactic acid bacteria biofilms.
2.1 AI-2 mediated QS system
The information communication mode of LAB is carried out in a density-dependent manner, which can be divided into intraspecific and interspecific information communication. Intraspecific communication used autoinducing peptides (AIPs) as signal molecules. The precursor AIPs are transcribed and modified to form specific AIPs, which cannot pass through the cell wall freely and must reach the extracellular via the ABC transport system or other membrane channel proteins. At optimal levels, AIPs activate the two-component system (TCS), which in turn controls the expression of target genes (Sturme et al., 2007). Interspecific signal communication involves the exchange of signals between different bacterial cells. The QS system, encompassing AI-2 and the enzyme LuxS responsible for AI-2 synthesis, plays a crucial role in the intraspecific and interspecific information exchange among various bacteria. AI-2 serves as the signal molecule for sensing environmental changes and facilitating inter-bacterial communication between Gram-positive and Gram-negative bacteria (Pereira et al., 2013). It has also been proved that luxS gene in QS regulates various physiological activities of LAB (Gu et al., 2018).
In recent years, research on the regulation of the LuxS/AI-2 mediated QS system in LAB mainly focuses on biofilm formation. Lebeer et al. (2009) reported that the ability of mutant strains luxS gene knockout of Lactobacillus rhamnosus GG to form biofilms was decreased. Adding exogenous AI-2 precursor molecules or wild-type strains could partially restored the biofilm formation of mutant strains, but could not fully revert them to their original state. Deng et al. (2022) published similar findings after knocking out luxS gene of L. rhamnosus. On the other hand, the overexpression of the luxS gene in Lactobacillus paraplantarum L-ZS9 resulted in an increased concentration of the AI-2 signaling molecule, promoting biofilm formation. These findings demonstrated that the luxS gene can regulate several genes responsible for encoding transporters, membrane proteins and transcriptional regulators (Liu et al., 2018). Zhang et al. (2022) reported that the synthetic AI-2 increased the cell density of Lactobacillus sanfranciscensis, enhanced the bacterial cohesion, and promoted the formation of biofilms. However, the luxS gene deletion strain of Lactobacillus reuteri 100-23C formed a thicker biofilm than the wild-type strain, yet the addition of exogenous AI-2 could not fully restore the biofilm thickness to that of the wild-type strain (Tannock et al., 2005). It can be seen that LuxS/AI-2 mediated QS system plays varying roles in biofilm formation among different bacteria, including those in the LAB group. It can either promote or inhibit biofilm formation, which needs further research. In the context of the LuxS/AI-2 mediated QS system, the pfs gene, in addition to luxS, serves as a key component in the synthesis of the signaling molecule AI-2 (Han and Lu, 2009). The pfs gene encodes the S-adenosine homocysteine nucleotide enzyme (Pfs), responsible for the hydrolysis of S-adenosine homocysteine (SAH) into S-nucleoside homocysteine (SRH) and adenine. SRH continues to participate in the synthesis of AI-2 in the presence of LuxS protein (Zhao et al., 2018). For example, expressing the pfs gene in Streptococcus suis serotype 2 could recover AI-2 synthesis, not the luxS gene (Wang et al., 2013b). Consequently, it becomes evident that the key genes governing AI-2 synthesis vary among different strains.
2.2 Second messenger molecules involved in regulation
Cyclic di-AMP (c-di-AMP) and cyclic di-GMP (c-di-GMP) are second messengers present in the cytoplasm, with c-di-GMP being particularly involved in the regulation of bacterial biofilm formation (Hickman and Harwood, 2008). An increase in cytoplasmic c-di-GMP concentration inhibits bacterial motility, reducing it to low or no activity. This reduced motility not only facilitates cell adhesion to surfaces, but also plays a crucial role in biofilm maturation (Samrot et al., 2021). Additionally, the increase in c-di-GMP levels enhance the transcription of genes responsible for exopolysaccharide synthesis, thereby promoting the formation of biofilms (Dial et al., 2021). However, the role of c-di-GMP in the formation of Lactobacillus biofilm remains underexplored. He et al. (2018) confirmed the genes related to c-di-GMP metabolism in Lactobacillus acidophilus, including dgcA, pdeA, pdeB, nrnA, gtsA and gtsB, and also proved that these genes jointly participated in regulating the formation and co-aggregation of extracellular polymers of Lactobacillus acidophilus through bioinformatics and biochemical analysis tests.
2.3 Multiple genes involvement in macro-regulation
The regulatory mechanisms involved in each stage of LAB, from planktonic state to biofilm state, are very complex. Whether in the adhesion, growth or maturity stage, a multitude of gene expressions and the transfer of signal molecules participate in regulating biofilm formation. The key genes regulating biofilm formation are different in different strains, so it is necessary to study them from an overall perspective. Recent advancements in modern biotechnology, such as the genome, transcriptome, proteomics and metabolomics in microorganisms research has deepened understanding of the regulatory mechanisms in LAB biofilm. This is expected to lay a theoretical foundation for the targeted improvement of the production performance of LAB in the food industry. Yang et al. (2021) analyzed the key genes of AI-2 synthesis in Leuconostoc citrem 37 using whole genome sequencing. Five genes (metK, DNMT/dcm, pfs, luxS and mmuM/BHMT2) were involved in AI-2 synthesis, in which the pfs and luxS played an important role. In addition, the signaling pathways involved in biofilm and quorum sensing may also involve genes related to carbon metabolisms, energy metabolisms, amino acid metabolisms, signal transduction and cell membrane transport, such as ciaH, ciaR and 26 other genes. Sun et al. (2020) described the characteristics of the biofilm formed by L. plantarum J26 and clarified its metabolic pathway based on transcriptome sequencing. The findings revealed that 1,051 genes significantly differed in the planktonic and biofilm state, among which 513 genes were up-regulated, and 538 were down-regulated. These genes were closely related to pyrimidine and glycerol metabolism, amino acid synthesis, stress response, enzyme synthesis and quorum sensing. Liu et al. (2021) analyzed the metabolic characteristics of planktonic and biofilm cells of Lactobacillus paraplantarum L-ZS9 by untargeted metabolomics, and the results suggested a significant distinctions, with biofilm cells displaying higher activity in amino acids and carbohydrate metabolism compared to planktonic cells. This suggest that Lactobacillus biofilms undergo notable metabolic adaptations to support biofilm formation and stability.
2.4 Influence of other factors on biofilm formation
The formation of Lactobacillus biofilm is not only related to QS system and c-di-GMP signaling system, but also to bacterial hydrophobicity, acid-base charge, self-polymerization and environmental factors, such as pH value, temperature, carbon source, metal ion, contact surface, and water erosion. The hydrophobicity and electrostatic interaction between the cell surface or between the cell surface and the object surface played a leading role in the non-specific adhesion process of Lactobacillus. These interactions influence how Lactobacillus cells adhere to various surfaces, which is essential for biofilm formation and colonization in different environments. In particular, hydrophobicity has been used as an important index to evaluate the adhesion of Lactobacillus. Li et al. (2023) found that L. plantarum Y42 exhibits different phenotypic properties in its planktonic and biofilm forms. In particular, the biofilm form demonstrated enhanced surface properties, including increased autoaggregation ability, hydrophobicity, acid-base charge, and adhesiveness, compared to its planktonic form. Therefore, surface physicochemical modification of materials can improve the adhesion of Lactobacillus. For example, the hydrophilicity of a dopamine-modified polypropylene fiber membrane was improved, which in turn promoted the biofilm formation of Lactobacillus paracasei (Zhao et al., 2016).
The effect of pH value on the adhesion ability of Lactobacillus was also different among strains, and an acidic environment is more favorable to adhesion. In acidic environments, L. acidophilus BG2FO4 secretes a protein molecule that acts as a bridging protein, mediating the connection between bacteria and cell receptors (Coconnier et al., 1992). Additionally, the effect of temperature on both the growth and biofilm formation of Lactobacillus strain varies. Ramírez et al. (2015) studied the effects of temperature on seven L. plantarum strains and found distinct differences among them. In 5 strains, as the temperature increased, their growth decreased, but the amount of biofilm formation increased. This suggests that while higher temperatures may inhibit cell proliferation in certain Lactobacillus strain, they can also stimulate biofilm formation, possibly as a stress response mechanism.
The carbon source had great influence on the biofilm formation, and it was related to the utilization rate of carbon source and the preference of strain. Glucose is the carbon source commonly used for culturing Lactobacillus, and most Lactobacillus show an increase in biofilm formation as the amount of glucose increases (Ramírez et al., 2015; Akoglu, 2020). The metal ions exert a substantial influence on LAB biofilm formation, and the effects varied among strains. Lactobacillus gasseri, Lactobacillus delbrueckii, L. reuteri and L. rhamnosus isolated from the vagina did not produce biofilms in MRS without MnSO4 (Terraf et al., 2012). Similarly, the same result was found in the study of L. plantarum WCFS1 and six L. plantarum strains isolated from food spoilage, but L. rhamnosus Gr18 increased the biofilm amount by 27.9% in manganese deficient environment compared to normal environment (Liu et al., 2024).
The contact surface materials commonly used in the food industry are stainless steel, glass and polystyrene. Studies had shown that L. rhamnosus GG had the strongest adhesion on stainless steel compared to glass, and the weakest was on polystyrene. The luxS gene, which is associated with the regulation of biofilm formation, was upregulated at 24 and 48 h on polystyrene and stainless steel supports, respectively (Nahle et al., 2023). In addition, the contact surface roughness has a significant influence on the adhesion of cells. The surface roughness of nano- and micro-roughness can enhance the initial adhesion of cells by increasing the contact area between the cells and the interface (Ionescu et al., 2012). Hu et al. (2019) cultivated the biofilm of L. plantarum on electrospun nanofiber membranes, which had excellent resistance to gastrointestinal fluids. Yogurt produced using nanofiber membranes containing L. plantarum biofilm as a starter culture has shown excellent fermentation properties, which shortens the fermentation time and makes the survival rate of probiotics higher during the shelf life of yogurt. According to Huijboom et al. (2024), L. plantarum biofilm structure and matrix, physiological state and stress resistance of cells is strain dependent and strongly affected under flow conditions.
Some non-signaling system-related chemicals can also alter Lactobacillus biofilms formation. The resveratrol can change the physicochemical properties of the surface of Lactobacillus paracasei, thereby enhancing the aggregation of cells and promoting adhesion and biofilm formation (Al Azzaz et al., 2020). The serotonin can promote the transport of quorum sensing signaling peptides in Enterococcus faecium, increase the abundance of some proteins related adhesion, and enhance the ability of cell self-aggregation and biofilm formation (Scardaci et al., 2021). The organic selenium regulated QS system of L. paracasei by binding two crucial receptor proteins (histidine protein kinase HPK and periplasmic binding protein LuxP) from specific sites, and promoted the biofilm formation (Lou et al., 2024). Therefore, Lactobacillus biofilm formation can be regulated by exogenous addition of these chemicals.
3 Relationship between Lactobacillus biofilm and adhesion
The adhesion of Lactobacillus to the surface is the first step for Lactobacillus to play a probiotic role. At the early stage of adhesion, whether or not the bacteria can attach to the cell surface is determined by the physicochemical properties of the bacterial surface, mainly by the properties of surface proteins. Bacterial initial adhesion and biofilm formation are controlled by different genes, and bacterial initial adhesion is an important prerequisite for biofilm formation (van Merode et al., 2006). For instance, the addition of mucin to MRS medium without glucose increased the biofilm formation ability of L. rhamnosus by 20% (Lebeer et al., 2007). At the later stage of adhesion, more extracellular matrix is secreted to the outside of the cell with the continuous proliferation of bacteria, so that the bacteria accumulate in large numbers, and finally form biofilm. The formation of Lactobacillus biofilm can greatly improve the adhesion ability of the bacteria. The extracellular matrix plays a key role in the later stage, especially exopolysaccharides. It had been reported in comparison to the wild-type L. rhamnosus, the mutant strain containing genes related to extracellular polysaccharide synthesis exhibited stronger biofilm formation ability (Lebeer et al., 2009).
The adhesion ability of Lactobacillus is related to its surface polysaccharides and membrane proteins. It had been found that L. plantarum WCFS1 had a similar QS system lamBDCA to Staphylococcus aureus. The adhesion of mutant lamA to glass surface is lower than that of wild-type strains, which is the first time that non-pathogenic agr systems have been found to encode self-inducible peptides and participate in the regulation of adhesion (Sturme et al., 2005). Because biofilm formation is closely related to the QS system mediated by the signaling molecule AI-2, it has been found that the signaling molecules of the QS system can influence the adhesion of LAB. The AI-2 signaling molecules produced by L. acidophilus NCFM can enhance the formation of biofilms and ultimately increase the adhesion to intestinal epithelial cells (Buck et al., 2009). Zhang et al. (2022) compared the effects of endogenous and synthetic AI-2 on the growth of L. sanfranciscensis. The synthetic AI-2 increased the cell density and cohesive force of L. sanfranciscensis. The adherence of luxS mutant L. acidophilus to Caco-2 decreased by 58% compared with the wild type (Buck et al., 2009). The ability of L. plantarum KLDS1.0391 to adhere to Caco-2 cells was drastically decreased by the luxS gene knockout, speculating that luxS/AI-2 mediated QS may affect the adhesion of LAB and regulate biofilm (Jia et al., 2018). Therefore, it is of great significance to utilize QS system to regulate the formation of Lactobacillus biofilms, thereby improving the adhesion ability of Lactobacillus and enhancing the probiotic effect of Lactobacillus.
4 Stress resistance of Lactobacillus biofilm
Biofilm is a self-protective mechanism formed by microorganisms in an adverse environment. They exhibit a natural capacity for self-production of extracellular polymeric substances (EPS), which include exopolysaccharides, proteins, and extracellular DNA. These substances enhance their adhesion to surfaces, forming a thick protective barrier. In addition to providing a structural barrier, Lactobacillus also tend to survive in the form of biofilm when faced with adverse environmental stress, thus enhancing the stress resistance of the bacteria. The stress resistance of Lactobacillus biofilm to different environments is shown in Table 1. The protective effect of biofilm is thought to be mainly due to the fact that the extracellular polymers of biofilm shield the damage of toxic compounds, antibiotics and enzymes to the embedded bacteria, and the diffusion of harmful substances can be hindered by the extracellular polymers (Nguyen et al., 2022). Therefore, strains located deeper in the biofilm can be better protected. In addition, L. plantarum biofilms have been shown to withstand harsh conditions in fermented foods due to their robust EPS matrix, which protects against desiccation and nutrient depletion (Wang et al., 2021).
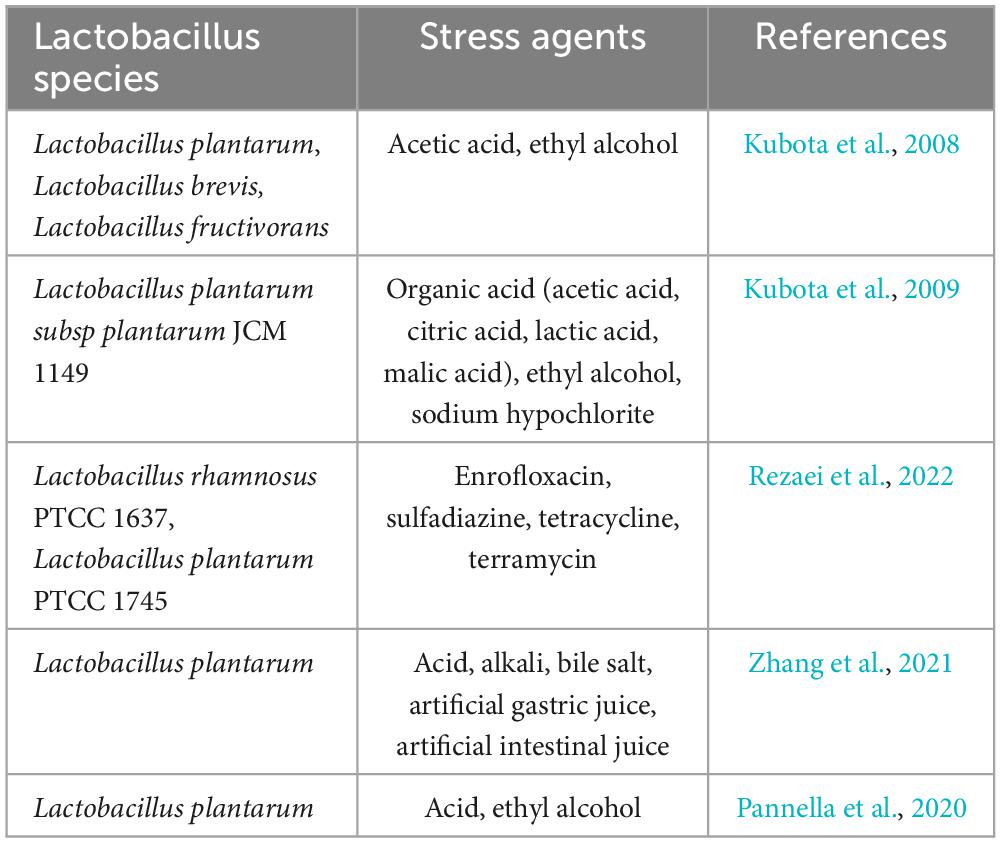
Table 1. Types of stress that Lactobacillus biofilms are able to withstand.
Recent research shows that when exposed to acidic environment, Lactobacillus activate a complex stress response that leads to significant physiological adaptations. This stress response often involves upregulation of acid tolerance mechanisms, including the F0F1-ATPase system and the production of chaperones that stabilize cellular proteins on low pH conditions (Zhang et al., 2022; Wang et al., 2021). This adaptation not only enhances survival but also improves intercellular communication and resource sharing through quorum sensing, particularly via the AI-2 signaling pathway (Papadimitriou et al., 2016). Furthermore, the physiological changes triggered by acidic stress responses in Lactobacillus, such as modifications in the cell surface properties and membrane liquid composition have been shown to provide cross-protection against additional stresses, like oxidative and osmotic pressures (Papadimitriou et al., 2016).
As an important regulatory mechanism involved in bacterial metabolism, it is of utmost importance to study the stress mechanism of LAB under different environmental stresses from the LuxS/AI-2 mediated QS system. At present, some progress has been made in this aspect. A total of 72 genes showed differential expression in L. reuteri after being treated in the acidic environment, among which the expression level was luxS gene up-regulated by approximately 3–4 times (Wall et al., 2007). The luxS gene expression was significantly up-regulated in all three strains of L. plantarum F, Lactobacillus sakei L4 and L. plantarum R, in which transcription level was positively associated with salt concentration under high nitrate stress, but differentially among the three strains (Lin et al., 2015). Transcription level of the luxS gene also appear to be upregulated under nutrient deficient conditions (Gu et al., 2018). Given that the luxS gene regulates the synthesis of the signaling molecule AI-2, the influence of AI-2 on the stress resistance of LAB exhibits a certain strain specificity and concentration dependence. Adding the exogenous signaling molecule AI-2 can improve the bile salt tolerance of L. sanfranciscensis (Zhang et al., 2022). The exogenous addition of AI-2 by Gu et al. (2021) significantly improved the tolerance of L. plantarum to bile salts but did not improve the tolerance to acid. Overexpression of luxS gene promoted AI-2 synthesis and enhanced the heat tolerance of L. paraplantarum L-ZS9 (Liu et al., 2018). Meanwhile, the deletion of luxS gene will have some effect on the stress resistance and normal physiological function of LAB. Bove et al. (2012) knocked out the luxS gene of L. plantarum KLDS1.0391 and showed no significant difference in the number of viable cells compared to the wild-type strain under normal culture conditions, but the acid and bile salt tolerance of the strain with the deletion of luxS gene was significantly reduced under high acid and high bile salt environments.
In addition to the luxS gene, other genes related to the QS system were also associated with the stress resistance of strains, such as the gene ftsH encoding FtsH protein in the AAA (ATPase associated with different cellular activities) family, a ubiquitous membrane-bound, ATP-dependent metalloproteinase. Some studies have shown that the mutation of ftsH caused the loss of FtsH protease in L. plantarum WCFS1, which resulted in poor heat and salt tolerance of the mutant and reduced the ability to form biofilm. On the contrary, the overexpression of FtsH enhanced the heat tolerance, salt tolerance and biofilm formation ability of L. plantarum (Del Pozo, 2018). In the L. sanfranciscensis, the addition of artificial AI-2, can up-regulate the expression of ftsH gene, promoting biofilm formation and improving the bile salt tolerance (Zhang et al., 2022). In addition, in the mixed-species biofilms formed by Streptococcus thermophilus with Lactobacillus bulgaricus, Lactobacillus helveticus, Lactobacillus equinosus and L. paracasei, have been shown to offer better microbial protection due to the increased biofilm mass (Yao et al., 2022).
A large number of studies have proved that Lactobacillus biofilms exhibit strong tolerance to the environment. Studies on Lactobacillus stress responses have been relatively comprehensive (Baig et al., 2022), especially regarding acid resistance mechanism (Wang X. et al., 2022). However, studies specifically examining the link between stress responses and biofilm formation in Lactobacillus remains limited. Therefore, future research on stress resistance of Lactobacillus biofilms should focus on exploring the specific stress responses induced by various environmental factors and elucidating the mechanisms behind biofilms’ enhanced resistance. This mechanistic insight will support strategies to improve the resilience of Lactobacillus through biofilm-based approaches.
5 Application of Lactobacillus biofilms in food industry
Historically, Lactobacillus has been mostly used in food industry in the form of planktonic or cell-free metabolites, such as food starter culture and oral bacterial solution. Although most Lactobacillus can play a probiotic role in food, the strain will form a biofilm due to changes of environment, which leads to food spoilage. It has been reported that many Lactobacillus can form biofilms that can affect the quality of meat, cheese, sake, beer and salads. For example, Lactobacillus fructivorans can cause mayonnaise and miso to deteriorate. Lactobacillus acetotolerans and Lactobacillus brevis can cause the deterioration of vinegar, and L. plantarum subsp. plantarum may cause the contamination of common food production plants as well as spoilage of pickles (Somers et al., 2001; Kubota et al., 2009). However, Lactobacillus biofilms also exhibit unique features in terms of stress resistance and antibacterial, which have been used for food production and human health. The application of Lactobacillus biofilm in food mainly involves three aspects, prevent adhesion of pathogens, control spoilage organisms and improve food safety (Jara et al., 2020). Studies have shown that Lactobacillus biofilm can control the growth of Listeria monocytogenes, Salmonella Typhimurium and Escherichia coli O157:H7 (Silva et al., 2020). In addition, pathogen inhibition analysis of Escherichia coli, Staphylococcus aureus, and L. monocytogenes suggested a significant distinction between the planktonic and biofilm of L. reuteri DSM 17938 (Hu et al., 2022a). In addition to inhibiting food spoilage bacteria, it has also been found to remove common toxins in foods. L. rhamnosus biofilm was found to be able to remove aflatoxin M1 from milk by binding it to its cell surface components, reducing the toxin’s bioavailability and thereby potentially enhancing the safety of dairy products (Assaf et al., 2019). The tremendous benefits of Lactobacillus biofilms have not only been noticed against bacterial pathogens but also against fungi, especially in clinical environments. Therefore, the antifungal activities of Lactobacillus biofilms are also important in order to expand the therapeutical approaches to fungi diseases.
Biofilms formed by probiotics are considered an effective strategy against biofilms of pathogenic bacteria in certain disease intervention. They can compete with pathogenic bacteria for nutrients and space through different mechanisms of action, such as producing bacteriostatic substances, playing an immunomodulatory role, and competing with pathogenic bacteria for adhesion sites. Many researchers are trying to use probiotics as an alternative to control infection and introduce promising treatment alternative to infections (Guerrieri et al., 2009). Table 2 shows some examples of lactic acid bacteria biofilm inhibiting foodborne or pathogenic microorganisms.
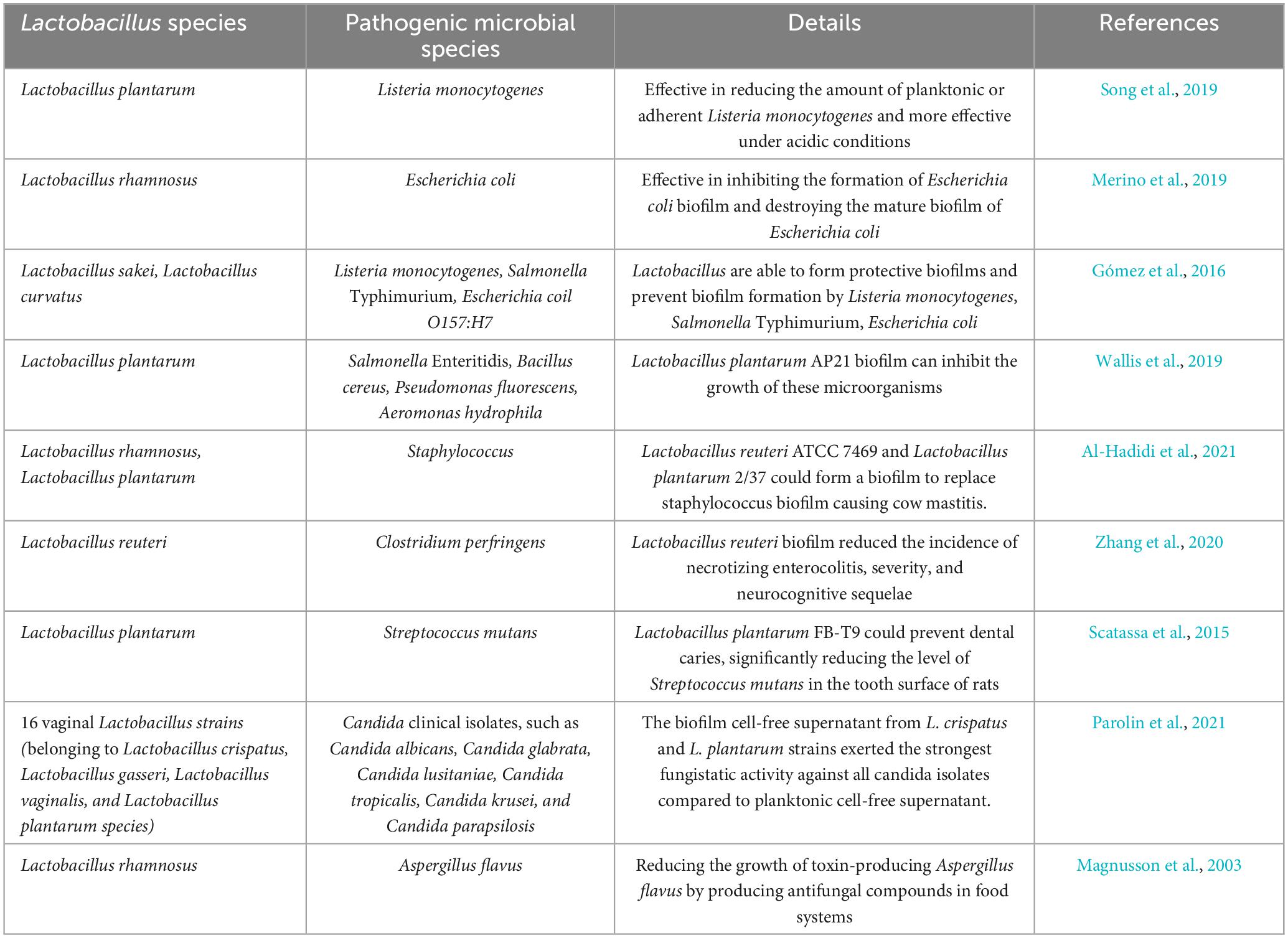
Table 2. Examples of pathogens inhibited by Lactobacillus biofilms.
The use of Lactobacillus biofilms can extend the shelf life of microbial starters, reduce fermentation times, and enhance the characteristic flavors of fermented food. Hu et al. (2019) cultivated the biofilm of L. plantarum on electrospun nanofiber membranes, which had excellent resistance to gastrointestinal fluids. Yogurt produced using nanofiber membranes containing L. plantarum biofilm as a starter culture has shown excellent fermentation properties, which shortens the fermentation time and makes the survival rate of probiotics higher during the shelf life of yogurt. In the production of traditional cheeses, Italian Ragusano cheese and French Salers cheese are made from raw milk, while oak barrels used for fermentation and maturation are rich in microbial biofilms, which are mainly composed of LAB and have an important influence on the unique flavor of cheese (Carpino et al., 2017; Furukawa, 2015).
Lactobacillus biofilm is used to produce metabolites, which is conducive to improving the production efficiency. Bastarrachea et al. (2022) use the biofilm formed by L. paracasei on the chitosan-modified polypropylene material to ferment lactic acid. The findings revealed that the production rate of lactic acid has been significantly improved. The fermentation system has the characteristics of high yield, good continuity and stability, which can maintain a high yield for a long time. Similarly, Zhao et al. (2016) found that a dopamine-modified polypropylene fiber membrane can promote the biofilm formation of L. paracasei, and the biofilm had excellent stress resistance. The purity of L-lactic acid produced by fermentation in the biofilm reactor was maintained at more than 99%. In contrast, the reactor had no fermentation delay period, effectively improving biological fermentation efficiency.
The fermentation process often involves multiple microbial species including Lactobacillus, yeast, and acetic acid bacteria. These microbial communities always exist and play a role in the form of mixed-species biofilm. The application of biofilms formed by Lactobacillus and other species of microorganisms in fermented food production are shown in Table 3. A better understanding of the formation, spatial distribution, systematic succession and function of multi-species biofilms can help researchers better control and utilize biofilms during fermentation and help improve the yield, quality, and safety of fermented products.
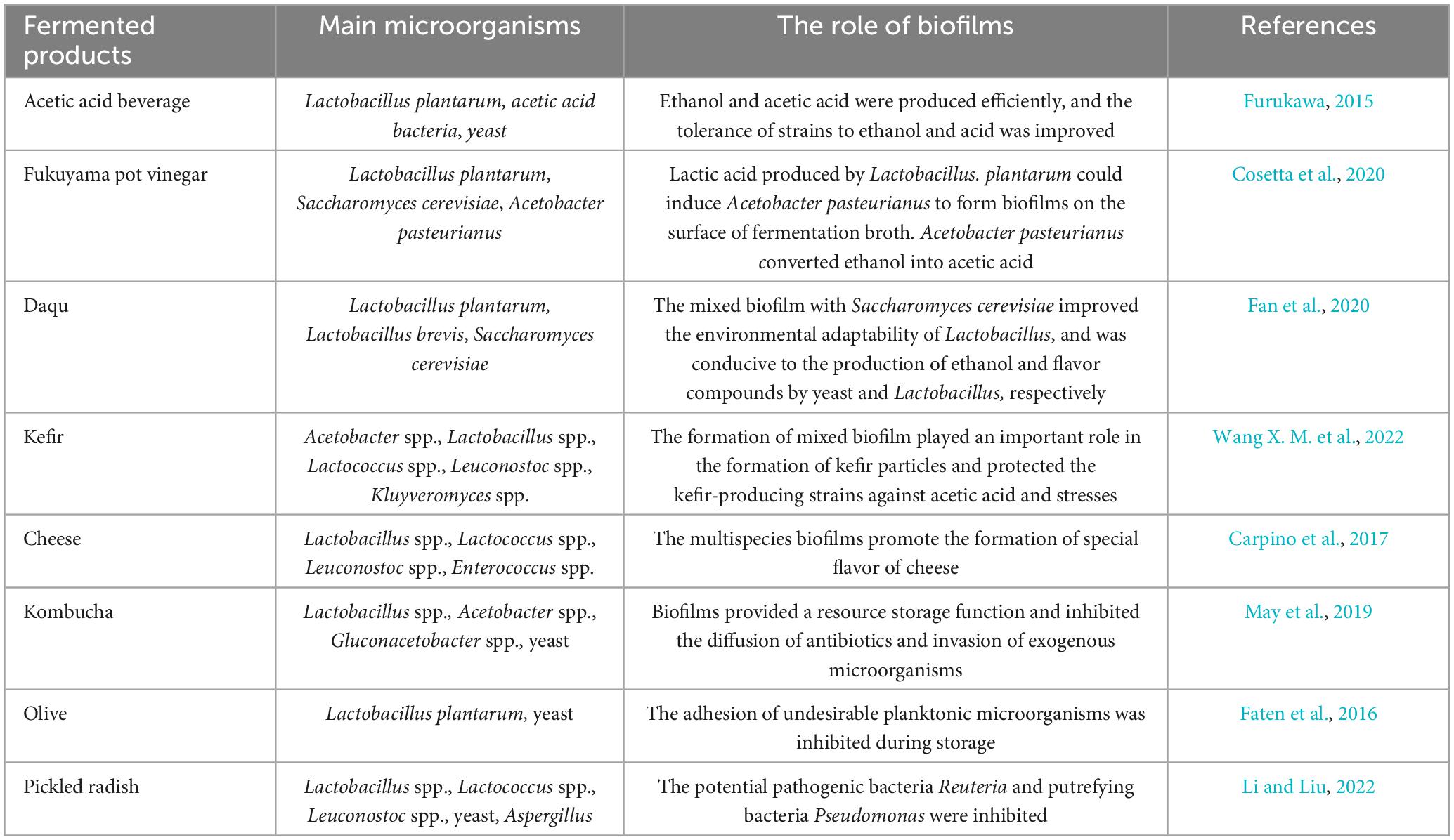
Table 3. Application of multispecies biofilms in food productions.
6 Lactobacillus biofilm encapsulation
To have any biological effect, a daily dose of probiotics of about 108–109 colony-forming units (CFU) is required prior and during passage through the gastrointestinal tract (Liu et al., 2019). Strategies for preserving the viability of probiotic strains have changed over time, leading to the development of the fourth-generation probiotics. Fourth-generation probiotics are represented by encapsulated probiotics, whereby bacterial cells exist in the form of biofilms. Encapsulation of LAB in the form of a biofilm involves embedding the bacteria within a protective matrix of edible colloidal substances like alginate, chitosan, or gelatin. Lactobacillus biofilm encapsulation protects LAB from the external environment and achieves release at the target site under conditions of active metabolism (Salas-Jara et al., 2016). Previous studies have verified that Lactobacillus can be encapsulated and form biofilms within capsules. These encapsulated biofilms show improved resistance to heat, acid, and storage environments. Cheow and Hadinoto (2013) encapsulated L. rhamnosus biofilm with a double-layer coating, which found stronger resistance than the planktonic strains encapsulated with a double-layer coating. Heumann et al. (2020) successfully prepared calcium pectin beads in which encapsulated Lactobacillus paracasei ATCC334 cells showed higher resistance to acid stress, freezing-drying stress, and combined stress. He et al. (2021) encapsulated LAB with low-methoxyl pectin, stimulated the biofilm formation of encapsulated lactobacilli upon in situ cultivation on microcapsules. The findings revealed that the microcapsules formed by biofilm-forming bacteria are more resistant to thermal shock, freeze-drying, gastrointestinal digestion, and drugs than those formed by planktonic bacteria. Vega-Sagardía et al. (2018) encapsulated Lactobacillus fermentum UCO-979C with alginate, xanthan gum, and vegetable oil, and allowed the strain to form biofilm within the microcapsules, and subjected to pH 3.0 maintained the anti-H. pylori inhibitory activity.
Probiotic encapsulation mainly depends on the choice of wall materials, such as emulsion, gel, nanocoating, and liposomes. Each encapsulating material has its advantages and disadvantages with respect to chemical, biological, mechanical, and physical properties (Razavi et al., 2021). Generally, the wall material of microcapsules should not affect the growth and metabolism of probiotics or human health. In addition, it should also have the advantages of being widely available, affordable, edible, and able to ensure targeted release in the colon while simultaneously resisting various adverse environments. Electrospun nanofibrous scaffolds have demonstrated excellent properties in facilitating the biofilm formation of probiotics (Hu et al., 2022b; Hu et al., 2019). However, the output of electrospun nanofibrous scaffolds is limited by the existing production method. Recent studies have explored using natural materials such as polysaccharides and proteins as encapsulation materials (Table 4). Seeking green, inexpensive, and sustainable encapsulation materials that support biofilm formation is important for the successful cultivation and large-scale production of biofilm-based probiotics.
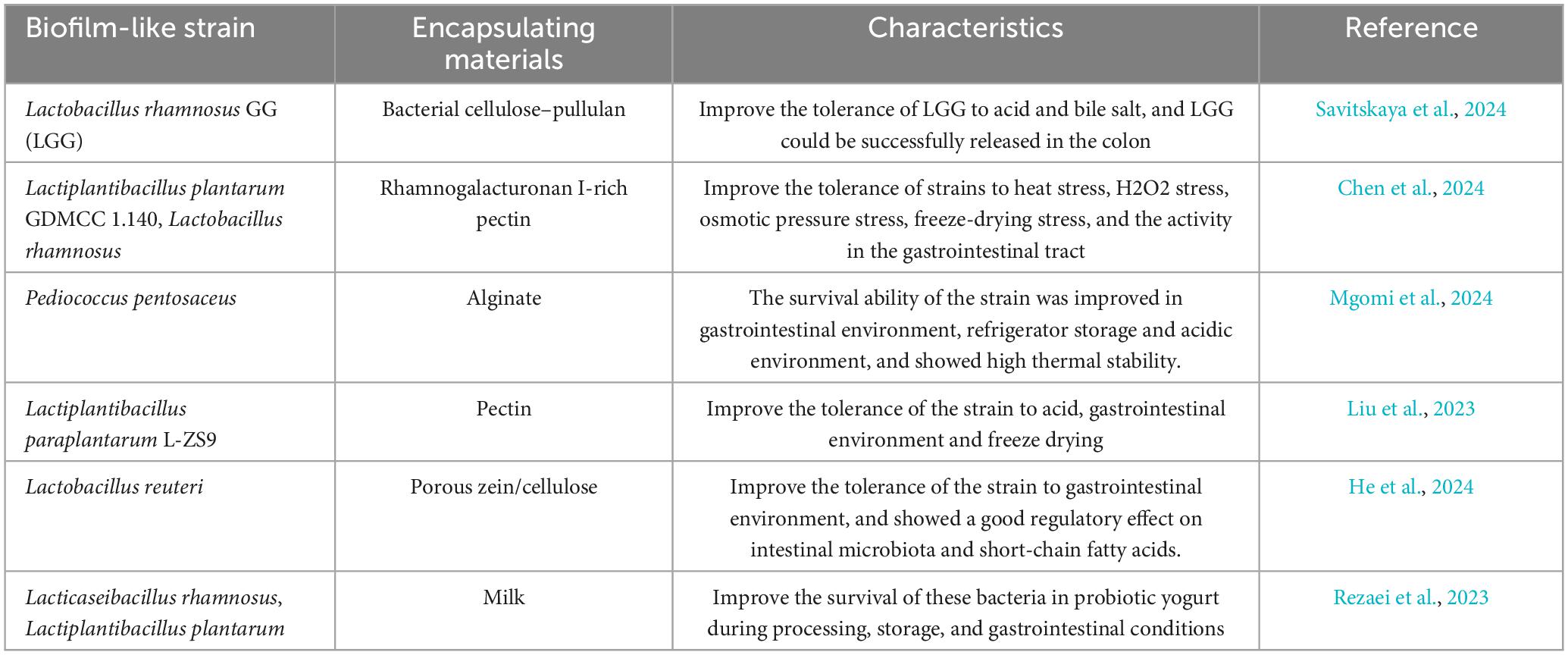
Table 4. Capsule materials and characteristics of Lactobacillus biofilm encapsulation.
7 Conclusion
In conclusion, research on the Lactobacillus biofilms has grown significantly in recent years, highlighting their beneficial properties, such as promoting bacterial adhesion, enhancing stress resistance and preventing colonization by pathogens. Despite this progress, substantial doubts remain to be resolved. Existing studies of LAB biofilms have focused mainly on strains of the genus Lactobacillus, whereas reports on biofilm in strains of the genera Bifidobacterium, Streptococcus, Leuconostoc, and Pediococcus are rare. Species-specific biofilm formation mechanisms and biofilm function should be investigated. Meanwhile, the applications of Lactobacillus biofilm is limited by the low cell viability of planktonic cells, the detachment of surface cells, the cell release after biofilm maturation, the incomplete regulatory mechanism and the limited material diffusion within biofilms. Another issue is determining the optimal encapsulation materials and parameters for developing fourth-generation probiotics. Survival of fourth-generation probiotics has been assessed only in vitro and thus requires further studies. These limitations also present opportunities for advancement. The rapid development of emerging technologies, such as multi-omics, gene-editing systems, electrostatic spinning and electrostatic spraying, has provided more options for studying the Lactobacillus biofilms. Continuous increases in the understanding of the Lactobacillus biofilm will undoubtedly help to enhance resistance to stress, inhibit harmful microorganisms, and enable more remarkable breakthroughs in improving product quality and flavor, and producing high-value products.
Author contributions
PY: Conceptualization, Formal analysis, Funding acquisition, Investigation, Writing – original draft, Writing – review and editing. EM: Supervision, Writing – review and editing. CZ: Investigation, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by the Major Natural Science Foundation of the Anhui Educational Committee (2023AH052217), the Key Research Project of Suzhou University (2022yzd04), and the National College Student Innovation and Entrepreneurship Plan Project (202310379010).
Acknowledgments
We would like to thank Min Kang (Universiti Sains Malaysia) and Bo Su (Suzhou University) for playing an integral role in production of the presented scientific data.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Akoglu, A. (2020). The effect of some environmental conditions on planktonic growth and biofilm formation by some lactic acid bacteria isolated from a local cheese in Turkey. Biotechnol. Lett. 42, 481–492. doi: 10.1007/s10529-020-02794-4
Al Azzaz, J., Al Tarraf, A., Heumann, A., Barreira, D. D., Laurent, J., Assifaoui, A., et al. (2020). Resveratrol favors adhesion and biofilm formation of Lacticaseibacillus paracasei subsp. paracasei strain ATCC334. Int. J. Mol. Sci. 21:5423. doi: 10.3390/ijms21155423
Al-Hadidi, A., Navarro, J., Goodman, S. D., Bailey, M. T., and Besner, G. E. (2021). Lactobacillus reuteri in its biofilm state improves protection from experimental necrotizing enterocolitis. Nutrients 13:918. doi: 10.3390/nu13030918
Assaf, J. C., El Khoury, A., Chokr, A., Louka, N., and Atoui, A. (2019). A novel method for elimination of aflatoxin M1 in milk using Lactobacillus rhamnosus GG biofilm. Int. J. Dairy Technol. 72, 248–256. doi: 10.1111/1471-0307.12578
Aymerich, T., Rodríguez, M., Garriga, M., and Bover-Cid, S. (2019). Assessment of the bioprotective potential of lactic acid bacteria against Listeria monocytogenes on vacuum-packed cold-smoked salmon stored at 8°C. Food Microbiol. 83, 64–70. doi: 10.1016/j.fm.2019.04.011
Baig, M., Turner, M. S., Liu, S. Q., Shah, N. N., and Ayyash, M. M. (2022). Heat, cold, acid, and bile salt induced differential proteomic responses of a novel potential probiotic Lactococcus garvieae C47 isolated from camel milk. Food Chem. 397:133774. doi: 10.1016/j.foodchem.2022.133774
Bao, Y. L., and Dai, G. Z. (2013). Time-gradient nitric acid modification of CF biofilm-carrier and surface nature effects on microorganism immobilization behavior in wastewater. Biotechnol. Biotechnol. Equipm. 27, 3918–3922. doi: 10.5504/BBEQ.2013.0054
Bastarrachea, L. J., Britt, D. W., Ward, R. E., and Demirci, A. (2022). Development of bioactive solid support for immobilized Lactobacillus casei biofilms and the production of lactic acid. Bioprocess Biosyst. Eng. 45, 217–226. doi: 10.1007/s00449-021-02654-z
Bove, P., Capozzi, V., Garofalo, C., Rieu, A., Spano, G., and Fiocco, D. (2012). Inactivation of the ftsH gene of Lactobacillus plantarum WCFS1: effects on growth, stress tolerance, cell surface properties and biofilm formation. Microbiol. Res. 167, 187–193. doi: 10.1016/j.micres.2011.07.001
Buck, B. L., Azcarate-Peril, M. A., and Klaenhammer, T. R. (2009). Role of autoinducer-2 on the adhesion ability of Lactobacillus acidophilus. J. Appl. Microbiol. 107, 269–279. doi: 10.1111/j.1365-2672.2009.04204.x
Carpino, S., Randazzo, C. L., Pino, A., Russo, N., Rapisarda, T., Belvedere, G., et al. (2017). Influence of PDO Ragusano cheese biofilm microbiota on flavour compounds formation. Food Microbiol. 61, 126–135. doi: 10.1016/j.fm.2016.09.006
Chen, P., Tian, J. H., Ren, Y. M., Cheng, H., Pan, H. B., Chen, S. G., et al. (2024). Enhance the resistance of probiotics by microencapsulation and biofilm construction based on rhamnogalacturonan I rich pectin. Int. J. Biol. Macromol. 258:128777. doi: 10.1016/j.ijbiomac.2023.128777
Cheng, K. C., Demirci, A., and Catchmark, J. M. (2011). Continuous pullulan fermentation in a biofilm reactor. Appl. Microbiol. Biotechnol. 90, 921–927. doi: 10.1007/s00253-011-3151-4
Cheow, W. S., and Hadinoto, K. (2013). Biofilm-like Lactobacillus rhamnosus probiotics encapsulated in alginate and carrageenan microcapsules exhibiting enhanced thermotolerance and freeze-drying resistance. Biomacromolecules 14, 3214–3222. doi: 10.1021/bm400853d
Coconnier, M. H., Klaenhammer, T. R., Kerneis, S., Bernet, M. F., and Servin, A. L. (1992). Protein-mediated adhesion of lactobacillus-acidophilus bg2fo4 on human enterocyte and mucus-secreting cell-lines in culture. Appl. Environ. Microbiol. 58, 2034–2039. doi: 10.1128/AEM.58.6.2034-2039.1992
Cosetta, C. M., Kfoury, N., Robbat, A., and Wolfe, B. E. (2020). Fungal volatiles mediate cheese rind microbiome assembly. Environ. Microbiol. 22, 4745–4760. doi: 10.1111/1462-2920.15223
Danielski, G. M., Imazaki, P. H., Cavalari, C. M. D., Daube, G., Clinquart, A., and de Macedo, R. E. F. (2020). Carnobacterium maltaromaticum as bioprotective culture in vitro and in cooked ham. Meat Sci. 162:108035. doi: 10.1016/j.meatsci.2019.108035
Del Pozo, J. L. (2018). Biofilm-related disease. Expert Rev. Antiinfect. Ther. 16, 51–65. doi: 10.1080/14787210.2018.1417036
Deng, Z. X., Hou, K. W., Valencak, T. G., Luo, X. M., Liu, J. X., and Wang, H. F. (2022). AI-2/LuxS quorum sensing system promotes biofilm formation of Lactobacillus rhamnosus GG and enhances the resistance to enterotoxigenic Escherichia coli in germ-free zebrafish. Microbiol. Spectr. 10:e0061022. doi: 10.1128/spectrum.00610-22
Dial, C. N., Speare, L., Sharpe, G. C., Gifford, S. M., Septer, A. N., and Visick, K. L. (2021). Para-aminobenzoic acid, calcium, and c-di-GMP induce formation of cohesive, syp-polysaccharide-dependent biofilms in Vibrio fischeri. Mbio 12:e0203421. doi: 10.1128/mBio.02034-21
Dong, Y., Han, M., Fei, T., Liu, H., and Gai, Z. H. (2023). Utilization of diverse oligosaccharides for growth by Bifidobacterium and Lactobacillus species and their in vitro co-cultivation characteristics. Int. Microbiol. 27, 941–952. doi: 10.1007/s10123-023-00446-x
Fan, Y., Huang, X. N., Chen, J. Y., and Han, B. Z. (2020). Formation of a mixed-species biofilm is a survival strategy for unculturable lactic acid bacteria and Saccharomyces cerevisiae in Daqu, a Chinese traditional fermentation starter. Front. Microbiol. 11:138. doi: 10.3389/fmicb.2020.00138
FAO/WHO (2002). Joint FAO/WHO Working Group Report on Drafting Guidelines for the Evaluation of Probiotics in Food. London: FAO/WHO.
Faten, K., Hamida, K., Soumya, E., Saad, I. K., Hasna, M., Hassan, L., et al. (2016). Lactobacillus plantarum: effect of a protective biofilm on the surface of olives during storage. Brazil. J. Microbiol. 47, 202–209. doi: 10.1016/j.bjm.2015.11.028
Furukawa, S. (2015). Studies on formation, control and application of biofilm formed by food related microorganisms. Biosci. Biotechnol. Biochem. 79, 1050–1056. doi: 10.1080/09168451.2015.1018126
Garnier, L., Penland, M., Thierry, A., Maillard, M. B., Jardin, J., Coton, M., et al. (2020). Antifungal activity of fermented dairy ingredients: identification of antifungal compounds. Int. J. Food Microbiol. 322:108574. doi: 10.1016/j.ijfoodmicro.2020.108574
Gómez, N. C., Ramiro, J. M. P., Quecan, B. X. V., and de Melo Franco, B. D. G. (2016). Use of potential probiotic Lactic Acid Bacteria (LAB) biofilms for the control of Listeria monocytogenes, Salmonella Typhimurium, and Escherichia coil O157:H7 biofilms formation. Front. Microbiol. 7:863. doi: 10.3389/fmicb.2016.00863
Gu, Y., Li, B., Tian, J. J., Wu, R., and He, Y. F. (2018). The response of LuxS/AI-2 quorum sensing in Lactobacillus fermentum 2-1 to changes in environmental growth conditions. Ann. Microbiol. 68, 287–294. doi: 10.1007/s13213-018-1337-z
Gu, Y., Tian, J. J., Zhang, Y., Wu, R., Li, L. J., Zhang, B. J., et al. (2021). Dissecting signal molecule AI-2 mediated biofilm formation and environmental tolerance in Lactobacillus plantarum. J. Biosci. Bioeng. 131, 153–160. doi: 10.1016/j.jbiosc.2020.09.015
Guerrieri, E., de Niederhäusern, S., Messi, P., Sabia, C., Iseppi, R., Anacarso, I., et al. (2009). Use of lactic acid bacteria (LAB) biofilms for the control of Listeria monocytogenes in a small-scale model. Food Control 20, 861–865. doi: 10.1016/j.foodcont.2008.11.001
Han, X. G., and Lu, C. P. (2009). In vitro biosynthesis of autoinducer 2 of Steptococcus suis Serotype 2 using recombinant LuxS and Pfs. Enzyme Microb. Technol. 44, 40–45. doi: 10.1016/j.enzmictec.2008.09.009
He, C. A., Sampers, I., Van de Walle, D., Dewettinck, K., and Raes, K. (2021). Encapsulation of lactobacillus in low-methoxyl pectin-based microcapsules stimulates biofilm formation: enhanced resistances to heat shock and simulated gastrointestinal digestion. J. Agric. Food Chem. 69, 6281–6290. doi: 10.1021/acs.jafc.1c00719
He, F., Ma, X. K., Tu, C. K., Teng, H., Shao, X., Chen, J., et al. (2024). Lactobacillus reuteri biofilms formed on porous zein/cellulose scaffolds: synbiotics to regulate intestinal microbiota. Int. J. Biol. Macromol. 262:130152. doi: 10.1016/j.ijbiomac.2024.130152
He, J. H., Ruan, W. H., Sun, J. L., Wang, F., and Yan, W. J. (2018). Functional characterization of c-di-GMP signaling-related genes in the probiotic Lactobacillus acidophilus. Front. Microbiol. 9:1935. doi: 10.3389/fmicb.2018.01935
Heumann, A., Assifaoui, A., Barreira, D. D., Thomas, C., Briandet, R., Laurent, J., et al. (2020). Intestinal release of biofilm-like microcolonies encased in calcium-pectinate beads increases probiotic properties of Lacticaseibacillus paracasei. NPJ Biofilms Microbiomes 6:44. doi: 10.1038/s41522-020-00159-3
Hickman, J. W., and Harwood, C. S. (2008). Identification of FleQ from Pseudomonas aeruginosa as a c-di-GMP-responsive transcription factor. Mol. Microbiol. 69, 376–389. doi: 10.1111/j.1365-2958.2008.06281.x
Hu, M. X., He, F., Guo, Y. X., Mo, L. Z., and Zhu, X. (2022a). Lactobacillus reuteri biofilms inhibit pathogens and regulate microbiota in In Vitro fecal fermentation. J. Agric. Food Chem. 70, 11935–11943. doi: 10.1021/acs.jafc.2c02372
Hu, M. X., He, F., Zhao, Z. S., Guo, Y. X., Ma, X. K., Tu, C. K., et al. (2022b). Electrospun nanofibrous membranes accelerate biofilm formation and probiotic enrichment: enhanced tolerances to pH and antibiotics. ACS Appl. Mater. Interfaces 14, 31601–31612. doi: 10.1021/acsami.2c04540
Hu, M. X., Li, J. N., Guo, Q., Zhu, Y. Q., and Niu, H. M. (2019). Probiotics biofilm-integrated electrospun nanofiber membranes: a new starter culture for fermented milk production. J. Agric. Food Chem. 67, 3198–3208. doi: 10.1021/acs.jafc.8b05024
Huijboom, L., Rashtchi, P., Tempelaars, M., Boeren, S., van der Linden, E., Habibi, M., et al. (2024). Phenotypic and proteomic differences in biofilm formation of two Lactiplantibacillus plantarum strains in static and dynamic flow environments. Biofilm 7:100197. doi: 10.1016/j.bioflm.2024.100197
Ionescu, A., Wutscher, E., Brambilla, E., Schneider-Feyrer, S., Giessibl, F. J., and Hahnel, S. (2012). Influence of surface properties of resin-based composites on in vitro Streptococcus mutans biofilm development. Eur. J. Oral Sci. 120, 458–465. doi: 10.1111/j.1600-0722.2012.00983.x
Jara, J., Pérez-Ramos, A., del Solar, G., Rodríguez, J. M., Fernández, L., and Orgaz, B. (2020). Role of Lactobacillus biofilms in Listeria monocytogenes adhesion to glass surfaces. Int. J. Food Microbiol. 334:108804.
Jia, F. F., Zheng, H. Q., Sun, S. R., Pang, X. H., Liang, Y., Shang, J. C., et al. (2018). Role of luxS in stress tolerance and adhesion ability in Lactobacillus plantarum KLDS1.0391. Biomed. Res. Int. 2018:4506829. doi: 10.1155/2018/4506829
Kubota, H., Senda, S., Nomura, N., Tokuda, H., and Uchiyama, H. (2008). Biofilm formation by lactic acid bacteria and resistance to environmental stress. J. Biosci. Bioeng. 106, 381–386. doi: 10.1263/jbb.106.381
Kubota, H., Senda, S., Tokuda, H., Uchiyama, H., and Nomura, N. (2009). Stress resistance of biofilm and planktonic Lactobacillus plantarum subsp plantarum JCM 1149. Food Microbiol. 26, 592–597. doi: 10.1016/j.fm.2009.04.001
Lebeer, S., Verhoeven, T. L. A., Francius, G., Schoofs, G., Lambrichts, I., Dufrêne, Y., et al. (2009). Identification of a gene cluster for the biosynthesis of a long, galactose-rich exopolysaccharide in Lactobacillus rhamnosus GG and functional analysis of the priming glycosyltransferase. Appl. Environ. Microbiol. 75, 3554–3563. doi: 10.1128/AEM.02919-08
Lebeer, S., Verhoeven, T. L. A., Vélez, M. P., Vanderleyden, J., and De Keersmaecker, S. C. J. (2007). Impact of environmental and genetic factors on Biofilm formation by the Probiotic strain Lactobacillus rhamnosus GG. Appl. Environ. Microbiol. 73, 6768–6775. doi: 10.1128/AEM.01393-07
Lewis, K. (2001). Riddle of biofilm resistance. Antimicrob. Agents Chemother. 45, 999–1007. doi: 10.1128/AAC.45.4.999-1007.2001
Li, J. Y., Mu, G. Q., and Tuo, Y. (2023). Phenotypic traits and probiotic functions of Lactiplantibacillus plantarum Y42 in planktonic and biofilm forms. Foods 12:1516. doi: 10.3390/foods12071516
Li, X. Q., and Liu, D. Q. (2022). Effects of wheat bran co-fermentation on the quality and bacterial community succession during radish fermentation. Food Res. Int. 157:111229. doi: 10.1016/j.foodres.2022.111229
Li, Y. H., and Tian, X. L. (2012). Quorum sensing and bacterial social interactions in biofilms. Sensors 12, 2519–2538. doi: 10.3390/s120302519
Lin, M., Zhou, G. H., Wang, Z. G., and Yun, B. (2015). Functional analysis of AI-2/LuxS from bacteria in Chinese fermented meat after high nitrate concentration shock. Eur. Food Res. Technol. 240, 119–127. doi: 10.1007/s00217-014-2313-x
Liu, C., Sun, D., Zhu, J. R., Liu, J. W., and Liu, W. J. (2020). The regulation of bacterial biofilm formation by cAMP-CRP: a mini-review. Front. Microbiol. 11:802. doi: 10.3389/fmicb.2020.00802
Liu, H., Cui, S. W., Chen, M. S., Li, Y., Liang, R., Xu, F. F., et al. (2019). Protective approaches and mechanisms of microencapsulation to the survival of probiotic bacteria during processing, storage and gastrointestinal digestion: a review. Crit. Rev. Food Sci. Nutr. 59, 2863–2878. doi: 10.1080/10408398.2017.1377684
Liu, L., Deng, J., Guo, S. Y., Li, Y. F., Wang, L. Y., Liu, Y. Y., et al. (2023). Stress tolerance and characterization of biofilm-like probiotic bacteria encapsulated in calcium pectin beads. LWT Food Sci. Technol. 184:115072. doi: 10.1016/j.lwt.2023.115072
Liu, L., Guo, S. Y., Chen, X., Yang, S. H., Deng, X., Tu, M. X., et al. (2021). Metabolic profiles of Lactobacillus paraplantarum in biofilm and planktonic states and investigation of its intestinal modulation and immunoregulation in dogs. Food Funct. 12, 5317–5332. doi: 10.1039/d1fo00905b
Liu, L., Wu, R. Y., Zhang, J. L., and Li, P. L. (2018). Overexpression of luxS promotes stress resistance and biofilm formation of Lactobacillus paraplantarum L-ZS9 by regulating the expression of multiple genes. Front. Microbiol. 9:2628. doi: 10.3389/fmicb.2018.02628
Liu, Y. S., Shen, K. S., Liu, Q., Diao, X. J., Ma, W. Y., and Liu, G. R. (2024). Transcriptomics analysis of the mechanism behind Lactobacillus rhamnosus Gr18 biofilm formation in an Mn 2+-deficient environment. Lwt Food Sci. Technol. 200:116163. doi: 10.1016/j.lwt.2024.116163
Lou, Z. X., Dong, J. L., Tao, H. W., Tan, Y. X., and Wang, H. X. (2024). Regulation and mechanism of organic selenium on quorum sensing, biofilm, and antioxidant effects of Lactobacillus paracasei. Cell Biochem. Funct. 42:e3975. doi: 10.1002/cbf.3975
Ma, J. H., Hong, Y., Deng, L. L., Yi, L. H., and Zeng, K. F. (2019). Screening and characterization of lactic acid bacteria with antifungal activity against Penicillium digitatum on citrus. Biol. Control 138:104044. doi: 10.1016/j.biocontrol.2019.104044
Magnusson, J., Ström, K., Roos, S., Sjögren, J., and Schnürer, J. (2003). Broad and complex antifungal activity among environmental isolates of lactic acid bacteria. FEMS Microbiol. Lett. 219, 129–135. doi: 10.1016/S0378-1097(02)01207-7
Mathur, H., Beresford, T. P., and Cotter, P. D. (2020). Health benefits of lactic acid bacteria (LAB) fermentates. Nutrients 12:1679. doi: 10.3390/nu12061679
May, A., Narayanan, S., Alcock, J., Varsani, A., Maley, C., and Aktipis, A. (2019). Kombucha: a novel model system for cooperation and conflict in a complex multi-species microbial ecosystem. PEERJ 7:e7565. doi: 10.7717/peeej.7565
Merino, L., Trejo, F. M., De Antoni, G., and Golowczyc, M. A. (2019). Lactobacillus strains inhibit biofilm formation of Salmonella sp. isolates from poultry. Food Res. Int. 123, 258–265. doi: 10.1016/j.foodres.2019.04.067
Mgomi, F. C., Yuan, L., Farooq, R., Lu, C. L., and Yang, Z. Q. (2024). Survivability and characterization of the biofilm-like probiotic Pediococcus pentosaceus encapsulated in calcium alginate gel beads. Food Hydrocolloids 156:110253. doi: 10.1016/j.foodhyd.2024.110253
Nahle, S., Atoui, A., Assaf, J. C., El Khoury, A., Louka, N., and Chokr, A. (2023). Time-dependent effect of surface material on Lactobacillus rhamnosus GG biofilm formation and gene expression. Microbiology 92, 55–65. doi: 10.1134/S0026261721102142
Nguyen, P. T., Nguyen, T. T., Nguyen, T. T. U., Hoang, Q. K., and Nguyen, H. T. (2022). Improve the viability and extracellular polymeric substances bioactivity of Lactiplantibacillus plantarum VAL6 using the environmental adaptation. Food Bioprod. Process. 131, 149–155. doi: 10.1016/j.fbp.2021.11.006
Pannella, G., Lombardi, S. J., Coppola, F., Vergalito, F., Iorizzo, M., Succi, M., et al. (2020). Effect of biofilm formation by Lactobacillus plantarum on the malolactic fermentation in model wine. Foods 9:797. doi: 10.3390/foods9060797
Papadimitriou, K., Alegría, Á., Bron, P. A., de Angelis, M., Gobbetti, M., Kleerebezem, M., et al. (2016). Stress physiology of lactic acid bacteria. Microbiol. Mol. Biol. Rev. MMBR 80, 837–890. doi: 10.1128/MMBR.00076-15
Papenfort, K., and Bassler, B. L. (2016). Quorum sensing signal-response systems in Gram-negative bacteria. Nat. Rev. Microbiol. 14, 576–588. doi: 10.1038/nrmicro.2016.89
Parolin, C., Croatti, V., Laghi, L., Giordani, B., Tondi, M. R., De Gregorio, P. R., et al. (2021). Lactobacillus biofilms influence anti-Candida activity. Front. Microbiol. 12:750368. doi: 10.3389/fmicb.2021.750368
Pereira, C. S., Thompson, J. A., and Xavier, K. B. (2013). AI-2-mediated signalling in bacteria. FEMS Microbiol. Rev. 37, 156–181. doi: 10.1111/j.1574-6976.2012.00345.x
Raganati, F., Procentese, A., Olivieri, G., Russo, M. E., Gotz, P., Salatino, P., et al. (2016). Butanol production by Clostridium acetobutylicum in a series of packed bed biofilm reactors. Chem. Eng. Sci. 152, 678–688. doi: 10.1016/j.ces.2016.06.059
Ramírez, M. D. F., Smid, E. J., Abee, T., and Groot, M. N. N. (2015). Characterisation of biofilms formed by Lactobacillus plantarum WCFS1 and food spoilage isolates. Int. J. Food Microbiol. 207, 23–29. doi: 10.1016/j.ijfoodmicro.2015.04.030
Razavi, S., Janfaza, S., Tasnim, N., Gibson, D. L., and Hoorfar, M. (2021). Microencapsulating polymers for probiotics delivery systems: preparation, characterization, and applications. Food Hydrocolloids 120:106882. doi: 10.1016/j.foodhyd.2021.106882
Rezaei, Z., Khanzadi, S., and Salari, A. (2022). A survey on biofilm formation of Lactobacillus rhamnosus (PTCC 1637) and Lactobacillus plantarum (PTCC 1745) as a survival strategy of probiotics against antibiotic in vitro and yogurt. J. Food Process. Preserv. 46:e15991. doi: 10.1111/jfpp.15991
Rezaei, Z., Salari, A., Khanzadi, S., Jong-Whan, R., and Shamloo, E. (2023). Preparation of milk-based probiotic lactic acid bacteria biofilms: a new generation of probiotics. Food Sci. Nutr. 11, 2915–2924. doi: 10.1002/fsn3.3273
Rosche, B., Li, X. Z., Hauer, B., Schmid, A., and Buehler, K. (2009). Microbial biofilms: a concept for industrial catalysis? Trends Biotechnol. 27, 636–643. doi: 10.1016/j.tibtech.2009.08.001
Salas-Jara, M. J., Ilabaca, A., Vega, M., and García, A. (2016). Biofilm forming Lactobacillus: new challenges for the development of probiotics. Microorganisms 4:35. doi: 10.3390/microorganisms4030035
Samrot, A. V., Mohamed, A. A., Faradjeva, E., Jie, L. S., Sze, C. H., Arif, A., et al. (2021). Mechanisms and impact of biofilms and targeting of biofilms using bioactive compounds-A review. Medicina Lithuania 57:839. doi: 10.3390/medicina57080839
Sánchez, M. T., Ruiz, M. A., Lasserrot, A., Hormigo, M., and Morales, M. E. (2017). An improved ionic gelation method to encapsulate Lactobacillus spp. bacteria: protection, survival and stability study. Food Hydrocolloids 69, 67–75. doi: 10.1016/j.foodhyd.2017.01.019
Savitskaya, I., Zhantlessova, S., Kistaubayeva, A., Ignatova, L., Sinyavskiy, Y., Shokatayeva, D., et al. (2024). Prebiotic cellulose-pullulan matrix as a “Vehicle” for probiotic biofilm delivery to the host large intestine. Polymers 16:30. doi: 10.3390/polym16010030
Saxelin, M., Tynkkynen, S., Mattila-Sandholm, T., and de Vos, W. M. (2005). Probiotic and other functional microbes: from markets to mechanisms. Curr. Opin. Biotechnol. 16, 204–211. doi: 10.1016/j.copbio.2005.02.003
Scardaci, R., Manfredi, M., Barberis, E., Scutera, S., Marengo, E., and Pessione, E. (2021). Serotonin exposure improves stress resistance, aggregation, and biofilm formation in the probiotic Enterococcus faecium NCIMB10415. Microbiol. Res. 12, 606–625. doi: 10.3390/microbiolres12030043
Scatassa, M. L., Gaglio, R., Macaluso, G., Francesca, N., Randazzo, W., Cardamone, C., et al. (2015). Transfer, composition and technological characterization of the lactic acid bacterial populations of the wooden vats used to produce traditional stretched cheeses. Food Microbiol. 52, 31–41. doi: 10.1016/j.fm.2015.06.008
Silva, D. R., Sardi, J. D. O., Pitangui, N. D., Roque, S. M., da Silva, A. C. B., and Rosalen, P. L. (2020). Probiotics as an alternative antimicrobial therapy: current reality and future directions. J. Funct. Foods 73:104080. doi: 10.1016/j.jff.2020.104080
Somers, E. B., Johnson, M. E., and Wong, A. C. L. (2001). Biofilm formation and contamination of cheese by nonstarter lactic acid bacteria in the dairy environment. J. Dairy Sci. 84, 1926–1936. doi: 10.3168/jds.S0022-0302(01)74634-6
Song, H. Y., Zhang, J. B., Qu, J. L., Liu, J. J., Yin, P. Y., Zhang, G. X., et al. (2019). Lactobacillus rhamnosus GG microcapsules inhibit Escherichia coli biofilm formation in coculture. Biotechnol. Lett. 41, 1007–1014. doi: 10.1007/s10529-019-02694-2
Sturme, M. H. J., Francke, C., Siezen, R. J., de Vos, W. M., and Kleerebezem, M. (2007). Making sense of quorum sensing in lactobacilli:: a special focus on Lactobacillus plantarum WCFS1. Microbiol. 153, 3939–3947. doi: 10.1099/mic.0.2007/012831-0
Sturme, M. H. J., Nakayama, J., Molenaar, D., Murakami, Y., Kunugi, R., Fujii, T., et al. (2005). An agr-like two-component regulatory system in Lactobacillus plantarum is involved in production of a novel cyclic peptide and regulation of adherence. J. Bacteriol. 187, 5224–5235. doi: 10.1128/JB.187.15.5224-5235.2005
Sun, L. L., Zhang, Y., Guo, X. J., Zhang, L. D., Zhang, W., Man, C. X., et al. (2020). Characterization and transcriptomic basis of biofilm formation by Lactobacillus plantarum J26 isolated from traditional fermented dairy products. Lwt Food Sci. Technol. 125:109333. doi: 10.1016/j.lwt.2020.109333
Tannock, G. W., Ghazally, S., Walter, J., Loach, D., Brooks, H., Cook, G., et al. (2005). Ecological behavior of Lactobacillus reuteri 100-23 is affected by mutation of the luxS gene. Appl. Environ. Microbiol. 71, 8419–8425. doi: 10.1128/AEM.71.12.8419-8425.2005
Terraf, M. C. L., Tomás, M. S. J., Nader-Macías, M. E. F., and Silva, C. (2012). Screening of biofilm formation by beneficial vaginal lactobacilli and influence of culture media components. J. Appl. Microbiol. 113, 1517–1529. doi: 10.1111/j.1365-2672.2012.05429.x
van Merode, A. E. J., van der Mei, H. C., Busscher, H. J., and Krom, B. P. (2006). Influence of culture heterogeneity in cell surface charge on adhesion and biofilm formation by Enterococcus faecalis. J. Bacteriol. 188, 2421–2426. doi: 10.1128/JB.188.7.2421-2426.2006
Vega-Sagardía, M., Rocha, J., Sáez, K., Smith, C. T., Gutierrez-Zamorano, C., and García-Cancino, A. (2018). Encapsulation, with and without oil, of biofilm forming Lactobacillus fermentum UCO-979C strain in alginate-xanthan gum and its anti-Helicobacter pylori effect. J. Funct. Foods 46, 504–513. doi: 10.1016/j.jff.2018.04.067
Wall, T., Båth, M., Britton, R. A., Jonsson, H., Versalovic, J., and Roos, S. (2007). The early response to acid shock in Lactobacillus reuteri involves the ClpL chaperone and a putative cell wall-altering esterase. Appl. Environ. Microbiol. 73, 3924–3935. doi: 10.1128/AEM.01502-06
Wallis, J. K., Krömker, V., and Paduch, J. H. (2019). Biofilm challenge: lactic acid bacteria isolated from bovine udders versus Staphylococci. Foods 8:79. doi: 10.3390/foods8020079
Wang, H. K., Sun, Y., Chen, C., Sun, Z., Zhou, Y. C., Shen, F. D., et al. (2013a). Genome shuffling of Lactobacillus plantarum for improving antifungal activity. Food Control 32, 341–347. doi: 10.1016/j.foodcont.2012.12.020
Wang, X. M., Li, W. P., Xu, M. J., Tian, J. J., and Li, W. (2022). The microbial diversity and biofilm-forming characteristic of two traditional tibetan kefir grains. Foods 11:12. doi: 10.3390/foods11010012
Wang, X., Cui, Y. H., and Qu, X. J. (2022). Mechanisms and improvement of acid resistance in lactic acid bacteria. Arch. Microbiol. 200, 195–201. doi: 10.1007/s00203-017-1446-2
Wang, Y. Q., Wu, J. T., Lv, M. X., Shao, Z., Hungwe, M., Wang, J. J., et al. (2021). Metabolism characteristics of lactic acid bacteria and the expanding applications in food industry. Front. Bioeng. Biotechnol. 9:612285. doi: 10.3389/fbioe.2021.612285
Wang, Y., Yi, L., Zhang, Z. C., Fan, H. J., Cheng, X. C., and Lu, C. P. (2013b). Overexpression of luxS cannot increase autoinducer-2 production, only affect the growth and biofilm formation in Streptococcus suis. Sci. World J. 2013:924276. doi: 10.1155/2013/924276
Yang, Q. L., Wang, Y., An, Q., Sa, R., Zhang, D. J., and Xu, R. H. (2021). Research on the role of LuxS/AI-2 quorum sensing in biofilm of Leuconostoc citreum 37 based on complete genome sequencing. 3 Biotech 11:189. doi: 10.1007/s13205-021-02747-2
Yao, C. Q., Li, J., E, J., Wang, R. X., Zhang, Q. L., and Wang, J. G. (2022). The symbiosis among, and the storage stabilities of, starter lactic acid bacterial strains in biofilms. Lwt Food Sci. Technol. 155:112896. doi: 10.1016/j.lwt.2021.112896
Zhang, Q. X., Qin, S. J., Huang, Y., Xu, X. Y., Zhao, J. X., Zhang, H., et al. (2020). Inhibitory and preventive effects of Lactobacillus plantarum FB-T9 on dental caries in rats. J. Oral Microbiol. 12:1703883. doi: 10.1080/20002297.2019.1703883
Zhang, Y., Gu, Y., Wu, R., Zheng, Y. X., Wang, Y., Nie, L. L., et al. (2022). Exploring the relationship between the signal molecule AI-2 and the biofilm formation of Lactobacillus sanfranciscensis. Lwt Food Sci. Technol. 154:112704. doi: 10.1016/j.lwt.2021.112704
Zhang, Y., He, Y. F., Gu, Y., Wang, Y., and Zheng, Y. X. (2021). Stress resistance and antioxidant properties of lactic acid bacteria with high biofilm production. Trans. Chin. Soc. Agric. Eng. 37, 282–288. doi: 10.11975/j.issn.1002-6819.2021.06.034 (in Chinese with English abstract).
Zhao, J., Quan, C. S., Jin, L. M., and Chen, M. (2018). Production, detection and application perspectives of quorum sensing autoinducer-2 in bacteria. J. Biotechnol. 268, 53–60. doi: 10.1016/j.jbiotec.2018.01.009
Zhao, Z. J., Xie, X. N., Wang, Z., Tao, Y. C., Niu, X. D., Huang, X. R., et al. (2016). Immobilization of Lactobacillus rhamnosus in mesoporous silica-based material: an efficiency continuous cell-recycle fermentation system for lactic acid production. J. Biosci. Bioeng. 121, 645–651. doi: 10.1016/j.jbiosc.2015.11.010
Keywords: Lactobacillus, biofilm, quorum sensing, stress resistance, encapsulation
Citation: Yao P, Mohd Esah E and Zhao C (2025) Regulatory mechanisms and applications of Lactobacillus biofilms in the food industry. Front. Microbiol. 15:1465373. doi: 10.3389/fmicb.2024.1465373
Received: 16 July 2024; Accepted: 26 November 2024;
Published: 07 January 2025.
Edited by:
Moshe Shemesh, Agricultural Research Organization (ARO), IsraelReviewed by:
Angela Racioppo, University of Foggia, ItalyYigal Achmon, Guangdong Technion-Israel Institute of Technology (GTIIT), China
Copyright © 2025 Yao, Mohd Esah and Zhao. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Effarizah Mohd Esah, ZWZmYXJpemFoQHVzbS5teQ==