 Jiangqi Wu
Jiangqi Wu Haiyan Wang2
Haiyan Wang2 Guang Li
Guang Li- 1College of Forestry, Gansu Agricultural University, Lanzhou, China
- 2College of Urban Environment, Lanzhou City University, Lanzhou, China
Alpine wet meadows are known as N2O sinks due to nitrogen (N) limitation. However, phosphate addition and N deposition can modulate this limitation, and little is known about their combinative effects on N2O emission from the Qinghai-Tibet Plateau in wet meadows. This study used natural wet meadow as the control treatment (CK) and conducted experiments with N (CON2H4 addition, N15), P (NaH2PO4 addition, P15), and their combinations (CON2H4 and NaH2PO4 addition, N15P15) to investigate how N and P supplementation affected soil N2O emissions in wet meadow of QTP. Contrary to previous studies on grasslands, the effect of phosphate addition treatment on soil N2O flux was not detectable during the growing seasons of 2019 and 2020. Over a span of two years, the N addition treatment significantly increased the N2O flux by 3.45 μg⋅m–2⋅h–1 due to increased soil N availability. Noticeably, phosphate addition intensified the effect of N deposition treatment on soil N2O flux with high significance in the early growth season of 2020. This augmentation can be attributed to the alleviation of limiting factors imposed by plants and microorganisms on soil N and P, fostering the mineralization and decomposition of litter and soil nutrients by microorganisms. Consequently, the results showed that total nitrogen and nitrate nitrogen were the main controls on soil N2O emission under N and P addition. In addition, redundancy analysis showed that the relative abundance of NirK genes in soil microorganisms (Bradyrhizobium, Devosia, Ochrobactrum, Alcaligenes, Rhizobium) is the main factor affecting N2O flux and available nitrogen. We project that if nutrient input continues to increase, the main limiting factor of soil will change from N restriction to P restriction due to the unique microbial nitrogen conversion process in the alpine meadow, significantly increasing N2O emissions. Consequently, the heightened contribution of alpine wet meadows to global warming and ozone depletion hinges on the dynamics of nutrient input regimes, spotlighting the urgent need for informed environmental management strategies.
1 Introduction
Nitrous oxide (N2O) is a significant driver of global warming, with a warming potential 298 times that of carbon dioxide per unit mass (Wu et al., 2023). Recent studies underscore the alarming reality that atmospheric N2O accounts for approximately 6.2% of anthropogenic global warming and is on a trajectory of annual increase at a rate of 0.2–0.3% (Kim et al., 2022). This potent greenhouse gas not only contributes to global warming but also poses a risk to the ozone layer, amplifying the impact of ultraviolet radiation and imperiling the health and survival of both human beings and other organisms. In wetlands characterized by abundant organic matter and surface water, N2O production predominantly occurs through denitrification processes (Mehnaz and Dijkstra, 2016; Martínez-Espinosa et al., 2021). Recognized as a crucial natural source of N2O, wetland soil denitrification significantly influences the global nitrogen biogeochemical cycle (Xie et al., 2015). Studies have shown that nutrient addition enhances the diversity and complexity of soil bacteria, especially altering the functional bacteria related to soil nutrient cycling (Wen et al., 2022; Zhao et al., 2023). Consequently, the inhibition of the denitrification process in wetland soils emerges as a key strategy for curbing N2O emissions, thereby mitigating environmental pollution risks and enhancing nitrogen use efficiency (Bardon et al., 2017).
Nitrogen and phosphorus are the key nutrient factors that limit plant growth in wetland ecosystems, and play a very important role in wetland productivity and soil ecological process (Gong and Wu, 2021). In recent years, nitrogen and phosphorus inputs to global ecosystems have increased dramatically due to excessive use of chemical nitrogen fertilizers and human overgrazing. N can be input into the wetland ecosystem through natural settlement and plant N fixation, while P is mostly input through livestock manure and artificial fertilization (Jiang et al., 2013). The high imbalance of N and P inputs in the 20th century (Peñuelas and Sardans, 2022), which is more pronounced in areas of high N settlement (Sun et al., 2017), may lead to a shift from N constraint to a broader P constraint or NP co-constraint (Wu et al., 2023). Studies have shown that nitrogen and phosphorus input can disrupt the nutrient balance of wetland ecosystems, which may affect soil nutrient status, soil microbial diversity and vegetation productivity in the ecosystem (Poeplau et al., 2019), and these can promote (Kuypers et al., 2018) or inhibit (Mori et al., 2014) N2O emissions in wetlands. Although the P input can alleviate the P limit of the system and the imbalance between N and P, it also affects the denitrification of wetland soil N (Kim et al., 2015) and increases N2O emissions (Wang et al., 2017). In addition, P input can reduce the P limitation of plants and soil microorganisms, promote nitrogen absorption and fixation of plants and microorganisms, and reduce the nitrogen substrate available for N2O production, thus inhibiting the emission of N2O from the system (Yu et al., 2017). Furthermore, P input can alleviate the P-limiting effect of denitrifying bacteria, promote bacterial activity (DeForest and Otuya, 2020), increase the abundance of functional genes produced by N2O, and promote N2O emission (Zhang et al., 2019). Therefore, an accurate understanding of the N2O emission characteristics and driving mechanism under the background of future N and P input is a vital prerequisite for slowing down wetland N2O emissions and coping with future climate change.
The Qinghai-Tibet Plateau (QTP) is a crucial ecological security barrier in China. It is home to a distinctive alpine meadow ecosystem, covering 53% of its total area (Zhou et al., 2020). Perennial low temperatures limit N and P mineralization in this area, configuring a nutrient limitation model marked by phosphorus or combined nitrogen and phosphorus limitation (Chen et al., 2018; Wu et al., 2020). Over time, global nitrogen deposition and large-scale overfertilization have propelled a surge in nitrogen and phosphorus deposition on the Qinghai-Tibet Plateau, escalating from 1 kg N ha–1 y–1 in 1980 to 5 kg N ha–1 y–1 in 2010, and projections indicating a twofold to threefold increase in the future (Liu et al., 2013). In addition, QTP is an essential base for developing animal husbandry in China, which produces a large amount of livestock manure and increases the input of phosphorus in wetland soil (Zhang et al., 2019). N deposition and increased P input not only affect the N and P content of wetland soil but also influence its physical and chemical properties and relative abundance of microbial dominant genera (Qiu et al., 2023; Voigt et al., 2020; Gao et al., 2019), which in turn impacts wetland N2O emissions. N addition can induce changes in soil N and P availability, influencing the structure and relative abundance of soil microbial communities, resulting in positive (Wu et al., 2023) or negative (Gao et al., 2014) effects on N2O flux. Conversely, the impact of increased phosphorus input on wetland N2O emissions varies, with some studies suggesting a promotion and others noting no significant effect (Gao et al., 2015; Zhang et al., 2019). The high water content and rich organic matter characteristic of the QTP area create a low-oxygen environment in high-altitude soils, fostering N2O production primarily through denitrification (Wrage-Mönnig et al., 2018). Central to this process is the NirK gene, acknowledged as the pivotal gene in denitrification, exceeding the significance of the nirS gene in soil denitrification processes (Wu et al., 2022). The presence and activity of the NirK gene directly influences the soil nitrogen cycle, thereby shaping the dynamics of soil N2O flux. In addition, soil water content and temperature are major factors affecting the diversity of denitrifying bacteria by regulating soil nitrogen content and oxygen content (Na et al., 2019). Therefore, it was crucial to investigate how soil nutrients and NirK genes respond to changes in N and P addition for a clearer understanding of N2O dynamics.
Therefore, this study focuses on the Gahai wet meadow as the research area in which to study the effects of N and P addition on N2O flux, soil nitrogen components, and denitrification functional genes. Our research objectives are threefold: (1) To determine how soil N2O flux responds to N and P addition, and whether this relationship changes with temperature and soil water content; (2) To clarify the relationship between the changes in surface soil nitrogen components and denitrification functional genes after the N and P addition; (3) To explore the effects of N and P addition on N2O flux through key abiotic and biological factors. We hypothesize that: (1) phosphate addition increases N2O emissions due to stimulating the activity of denitrifying microorganisms (Cui et al., 2018); (2) Nitrogen deposition promotes soil N2O emissions due to the lifting of nitrogen restrictions, and phosphate addition intensifies the impact of N deposition on soil N2O release; (3) N and P addition can change the soil N2O emissions by changing the relative abundance of dominant species in the soil denitrification bacterial community; (4) Soil nitrogen components, temperature, and water content are the main environmental controls for N2O emissions.
2 Material and methods
2.1 Study area
The research area was located in the nature reserve of Gahai-Zecha (33° 58′ 12″−34° 32′ 16″ N, 102° 05′ 00″−102° 47′ 39″ E), near the northeast border of the QTP. The distribution area of alpine meadows in the area was over 80%, with an average elevation of 3,430–4,300 m and an area of 57,846 hm2. The Gahai Wetland belongs to the highly cold and humid climate zone of the Qinghai-Tibet Plateau. The average temperature in the area from 1981 to 2020 was 2.9°C, with an average annual precipitation of 785 mm.1 More than 70% of the rainfall was concentrated during the growing season (Wang et al., 2022). The soil was mainly composed of meadow soil, peat soil, and swamp soil, with a high organic matter content.
2.2 Experimental design
In May 2019, we selected an area characterized by flat terrain and evenly distributed vegetation as the experimental area. To implement nitrogen and phosphorus treatments, we utilized urea (CON2H4) and sodium dihydrogen phosphate (NaH2PO4) to convert these substances into their respective nitrogen and phosphorus content (Camenzind et al., 2014). Studies have shown that the current background value for both nitrogen deposition and phosphorus addition in this area was 5.0 kg N ha–1 y–1, and the rate of nitrogen deposition and phosphorus addition is expected to increase 2–3 times in the future (Zhai et al., 2024; Wu et al., 2023). Based on this, we added 15 kg N ha–1 y–1 and 15 kg P ha–1 y–1 as nutrient additions under different treatments. A total of four treatments were set up, including no fertilization as the control (CK), N15 treatment (CON2H4 addition), P15 treatment (NaH2PO4 addition), and N15P15 treatment (CON2H4 and NaH2PO4 addition). Each treatment had three replicates, a total of 12 plots. The plot size was 2 m × 2 m, and to avoid mutual interference, the distance between two adjacent plots was 5 m. At the end of May 2019 and 2020, the corresponding mass of fertilizer was dissolved in 2 L of water, and the sprayer was used to uniformly distribute the fertilizer solution across the experimental plots, while the control plots received an even spray of 2 L of water.
2.3 N2O flux and soil sample collection
Throughout the plant growth season, spanning from June to October 2019 and 2020, we conducted comprehensive measurements of N2O fluxes for each treatment. Gas samples were meticulously collected at 15-day intervals, employing the closed static chamber technique (Ma et al., 2018). The chambers used in our study were designed with dimensions of 50 cm in length, width, and height. After chamber closure, gas samples were taken at specific time intervals (0, 3, 10, 25, and 40 min) after chamber closure. Following collection, the N2O gas samples of the air samples were quantified using a gas chromatograph (Echrom A90, China). For N2O flux, the concentration slope versus time was accepted if R2 > 0.80 (Schneider et al., 2023). The N2O fluxes were then calculated using linear regression of N2O in the chamber concentrations versus time (Shi et al., 2021). Overall, 99.5% of the fluxes were calculated by linear regression and 0.5% by non-linear regression, and all N2O flux data were used for further analysis. In addition, a portable digital thermometer (JM624) and a soil moisture content analyzer (RS232) were used to measure soil temperature (ST) and soil water content (SWC) at 10 cm depth.
We collected 0–10 cm soil samples from June to October 2019 and 2020 (mid-month; June is the early growth period, EG; July to August is the middle growth period, MG; September to October is the late growth period, LG). The soil characteristics, including soil ammonium nitrogen (NH4+), nitrate nitrogen (NO3–), total nitrogen (TN), total phosphorus (TP), microbial biomass nitrogen (MBN), and microbial biomass carbon (MBC) were determined using previously established methods described (Saha et al., 2018; Wu et al., 2023). Additionally, on 20 June, 1 August, and 22 September 2020, liquid nitrogen was used to transport 0–10 cm soil samples back to the laboratory for testing the NirK gene sequence in each treated soil using a detection method as described in previous research (Zhang et al., 2018; Wu et al., 2022). DNA was extracted from 0.25 g of soil samples using the soil DNA extraction kit (MoBio Laboratories, Carlsbad, CA, USA), and use 1% agarose gel electrophoresis solution was used to detect DNA quality. The eligible DNA genes were amplified using the primer combination nirK1F-nirK5R (Chen et al., 2010), and the amplified sequencing was performed using the Illumina MiSeq 250 Sequencer (generating 2 × 250 bp paired-end reads) from Shanghai Tianhao Biotechnology Co., Ltd.
2.4 Statistical analysis
A normality test was conducted on the data using SPSS 22.0 software. The effects of treatments on soil N2O emissions and soil characteristic variables (ST, SWC, NH4+, NO3–, TP, MBC, and MBN) were tested with a one-way ANOVA. A linear regression model was used to explain the variability of environmental variables to N2O flux. Additionally, the differences in the NirK gene community structure were displayed by principal coordinate analysis (PCoA) based on Bray-Curtis distances using the PCoA function in the “ape” package in R software (significance level was P < 0.05). Redundancy analysis (RDA) was used to explore the relationships between soil NirK gene community structure, N2O fluxes, and soil characteristic factors.
3 Results
3.1 Soil characteristics
The soil characteristics of the 0–10 cm soil layer in the wet meadow on the QTP were significantly affected by the N and P addition treatments (Table 1 and Figure 1). Compared with CK, P15 and N15P15 treatments significantly increased the contents of soil MBN, NH4+-N, NO3–-N, and MBC (P < 0.05); P15 treatments significantly increased the soil TP content (P < 0.05), while N15 treatments significantly decreased the contents of TN, TP, NO3–-N and MBC (P < 0.05). ST showed a trend of increasing first and then decreasing after a month, while there was no clear SWC trend. In addition, the soil characteristics content under four treatments showed significant seasonal variations (P < 0.05, Table 2). Except for the TP content in the soil treated with CK and N15, the content of MBN, TN, NH4+, and NO3– in the soil showed a trend of first increasing and then decreasing with the extension of the season, with the larger values appearing in MG. The soil MBC content under the four treatments showed a trend of decreasing and increasing with the extension of seasons. Repeated analysis of variance demonstrated that N and P addition and season significantly interaction influenced the soil characteristics content in wet meadow soil (Table 2).
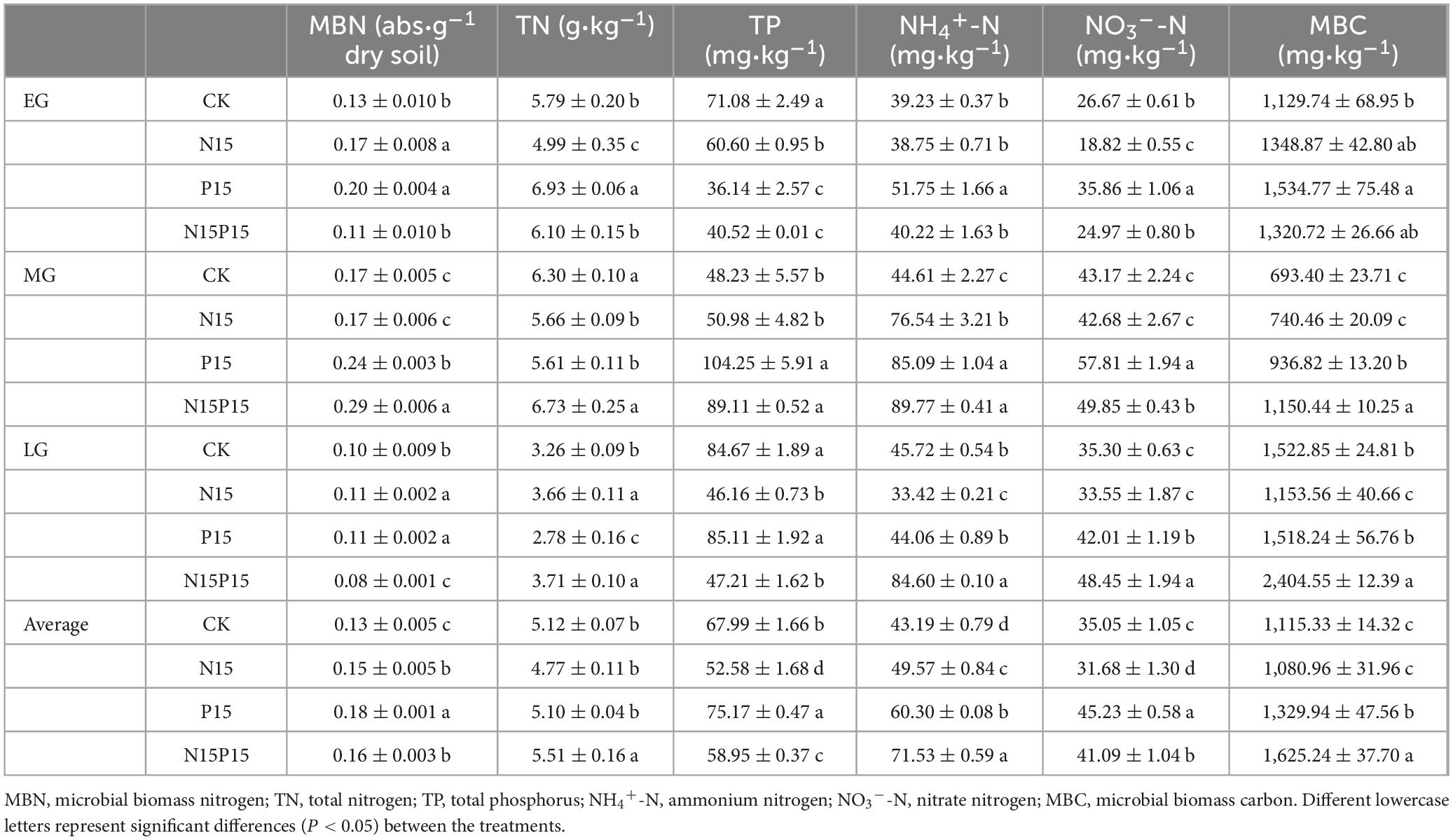
Table 1. Changes in surface soil characteristics during different vegetation growth seasons under nitrogen and phosphate addition for two consecutive years (mean ± standard errors).
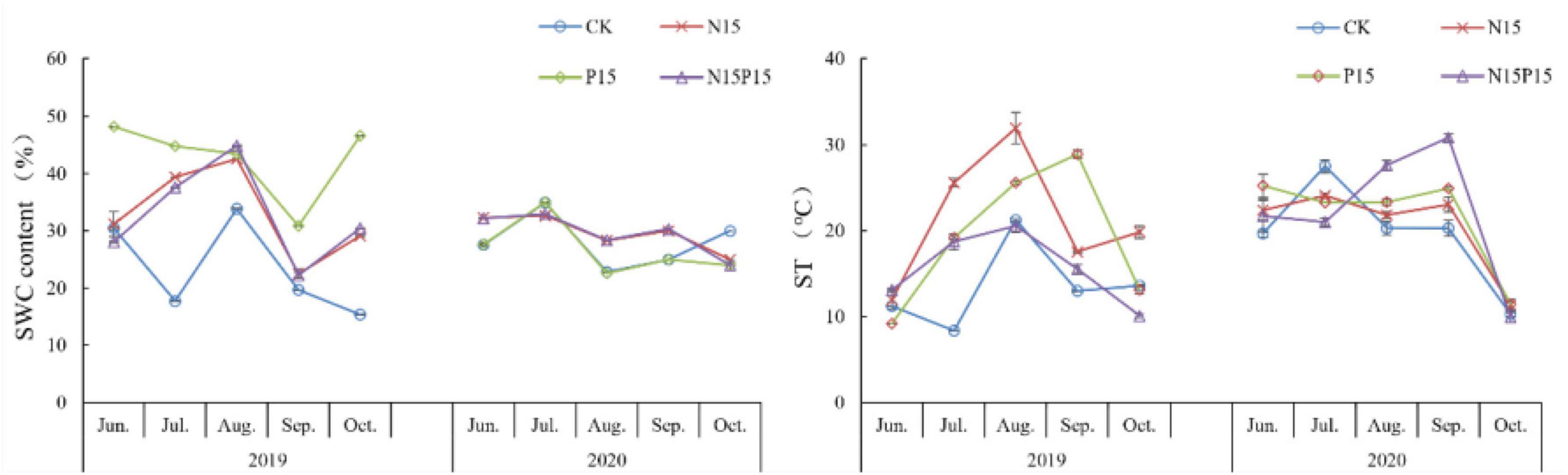
Figure 1. Changes in soil water content and soil temperature under nitrogen and phosphate addition. Error bars show the standard error. CK, Control; N15, CON2H4 addition treatment; P15, NaH2PO4 addition treatment; N15P15, CON2H4 and NaH2PO4 addition treatment.
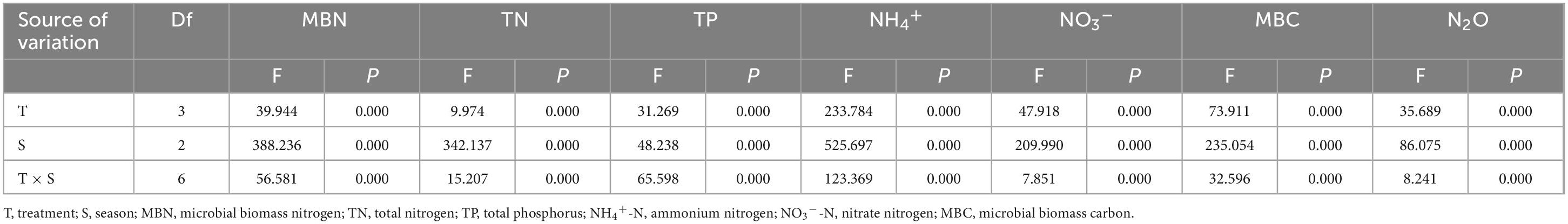
Table 2. Results of a repeated-measures ANOVA testing for differences in surface soil characteristics (TN, NO3–, NH4+, MBN, DON) among nitrogen and phosphate addition using season as the repeated variable.
3.2 N and P addition effects on N2O flux
High levels of N addition and NP addition significantly impacted the 2-year average N2O flux, but P addition did not significantly affect N2O flux (Figure 2). Throughout the entire plant growing seasons (June to October) of QTP in 2019 and 2020, the N2O emissions varied between the four treatments. Compared with CK (10.08 μg⋅m–2⋅h–1), N15, P15, and N15P15 increased N2O average emission by 3.46, 0.86, and 5.53 μg⋅m–2⋅h–1 in wet meadow. Moreover, P addition increased the increase of N2O emission resulting from N fertilization by 2.07 μg⋅m–2⋅h–1 (Figure 2).
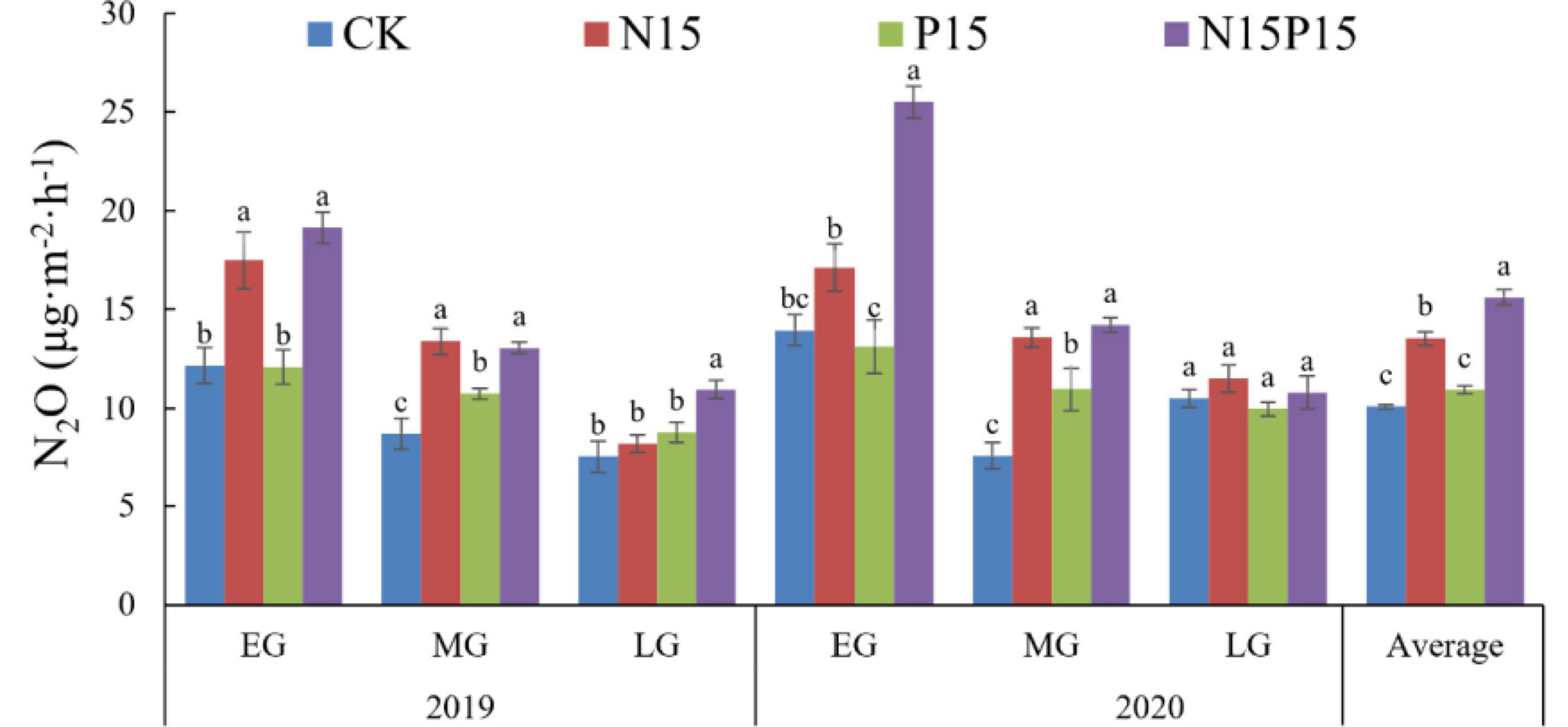
Figure 2. Average N2O flux in the 2019 and 2020 early (EG), middle (MG), and late (LG) growing season under four treatments. Average N2O flux in the 2019 and 2020 early (EG), middle (MG), and late (LG) growing season under four treatments. Different lowercase letters represent significant differences (P < 0.05) between the treatments, error bars show the standard error. CK, Control; N15, CON2H4 addition treatment; P15, NaH2PO4 addition treatment; N15P15, CON2H4 and NaH2PO4 addition treatment.
Fertilization and seasonal changes also significantly impacted soil N2O flux in QTP wet meadow (Table 2). Compared with the CK treatments in the EG (12.17 ± 0.90 μg⋅m–2⋅h–1) and MG periods (8.71 ± 0.79 μg⋅m–2⋅h–1) of 2019, the N15 treatment significantly increased average N2O emissions by 5.34 μg⋅m–2⋅h–1 during the EG period and 4.67 μg⋅m–2⋅h–1 during the MG period. Likewise, soil N2O emissions under N15 treatment increased by 3.16 μg⋅m–2⋅h–1 during the EG period and 5.99 μg⋅m–2⋅h–1 during the MG period in 2020. In addition, soil N2O emissions under the interactive treatment of N15 and P15 fertilization treatment were 6.95 μg⋅m–2⋅h–1 and 11.56 μg⋅m–2⋅h–1 higher than under the CK treatment in the EG of 2019 and 2020, respectively (Figure 2). Additionally, P fertilization together significantly increased the N2O flux by 2.02 and 3.37 μg⋅m–2⋅h–1 compared with the CK treatment only during the MG period in 2019 and 2020 (P = 0.03 and P = 0.01). In the late growth season, all other treatments had no significant effect on N2O flux except for N15P15 (P > 0.05).
3.3 Effect of N and P addition on soil NirK genes
There were significant differences in the composition of soil microbial communities under the four N and P addition treatments (Figure 3). Principal coordinates analysis (PCoA) of Bray-Curtis distance showed that there were significant differences in soil microbial community structure between N and P addition treatments and CK (P = 0.024), while the differences in the microbial community structure between N15 and N15P15 treatments did not reach significant (P = 0.152). Moreover, the dominant genera among the four treatments in wet meadows were Bradyrhizobium, Devosia, Ochrobactrum, Alcaligenes, and Rhizobium (Figure 4). N and P addition (N15, P15, N15P15) significantly reduced the relative abundance of Bradyrhizobium, and increased the relative abundance of Devosia, Ochrobactrum, Alcaligenes, and Rhizobium.
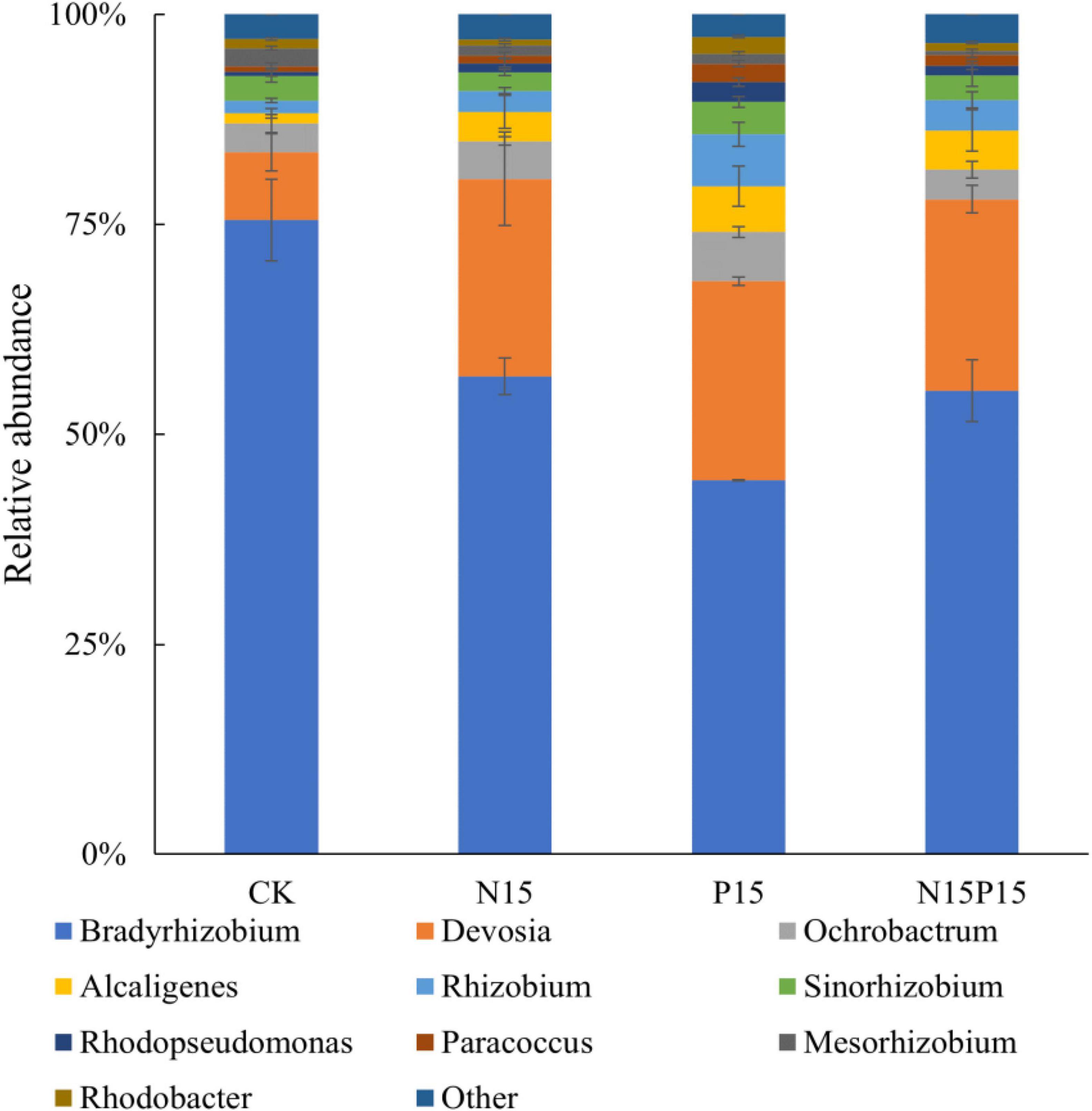
Figure 3. Histogram of soil NirK bacterial community structure (genus level) under nitrogen and phosphate addition treatment. CK, Control; N15, CON2H4 addition treatment; P15, NaH2PO4 addition treatment; N15P15, CON2H4 and NaH2PO4 addition treatment.
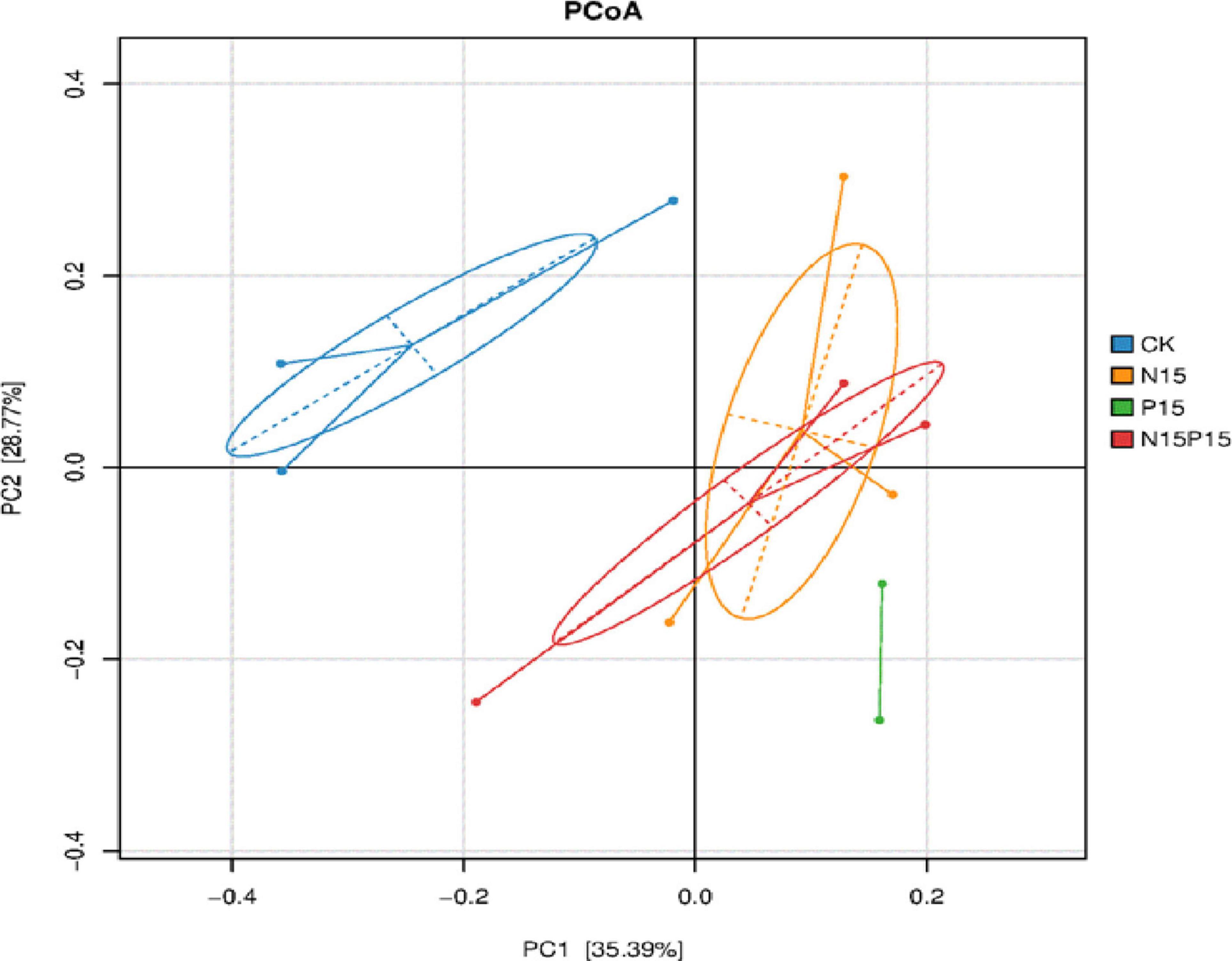
Figure 4. Principal coordinates analysis (PCoA) of bacterial community structure based on Bray-Curtis distances under different treatment. CK, Control; N15, CON2H4 addition treatment; P15, NaH2PO4 addition treatment; N15P15, CON2H4 and NaH2PO4 addition treatment. We used the first two samples from the P15 treatment for PCoA analysis due to the third soil sample from the P15 treatment was contaminated.
3.4 Relationships between N2O flux and soil characteristics and NirK genes
After two years of nitrogen and phosphorus addition treatments, significant correlations between N2O flux and TN at 10 cm depth, and between N2O flux and NO3– at 10 cm depth were observed (Table 3), explaining 16.99 and 24.78% of N2O variation, respectively (Figure 5). Furthermore, the relationships between N2O flux and other soil characteristics (ST, SWC, MBN, TP, NH4+ and MBC at 10 cm depth) were undetectable in this study. Moreover, at the level of dominant genera, the N2O flux was significantly negatively correlated with the relative abundance of Ochrobactrum, significantly positively correlated with the relative abundance of Alcaligenes (P < 0.05, Figure 6), and weakly correlated with other dominant genera (P > 0.05). Bradyrhizobium was significantly positively correlated with NH4+, NO3–, MBN, and significantly negatively correlated with MBC. Devosia and Rhizobium were significantly positively correlated with MBC, while negatively correlated with NH4+, NO3–, and MBN. Ochrobactrum was positively correlated with TP content but negatively correlated with SWC, ST, and TN.
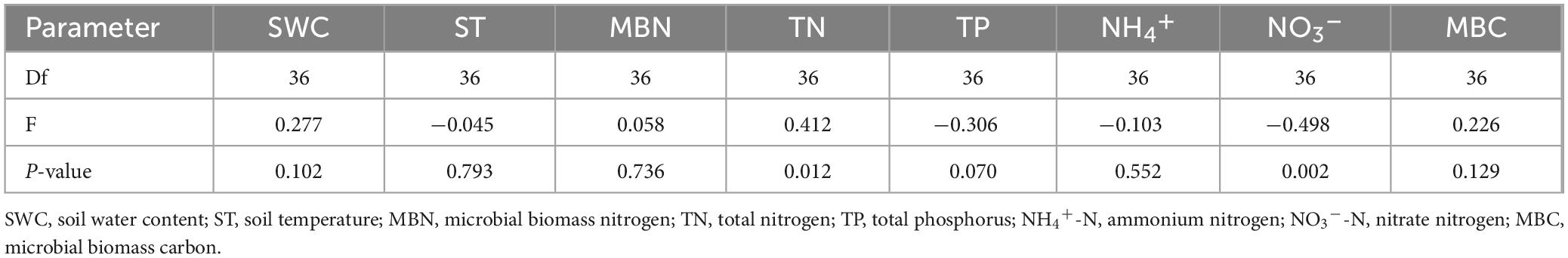
Table 3. Correlation analysis between soil characteristics and N2O flux in different vegetation growth seasons.
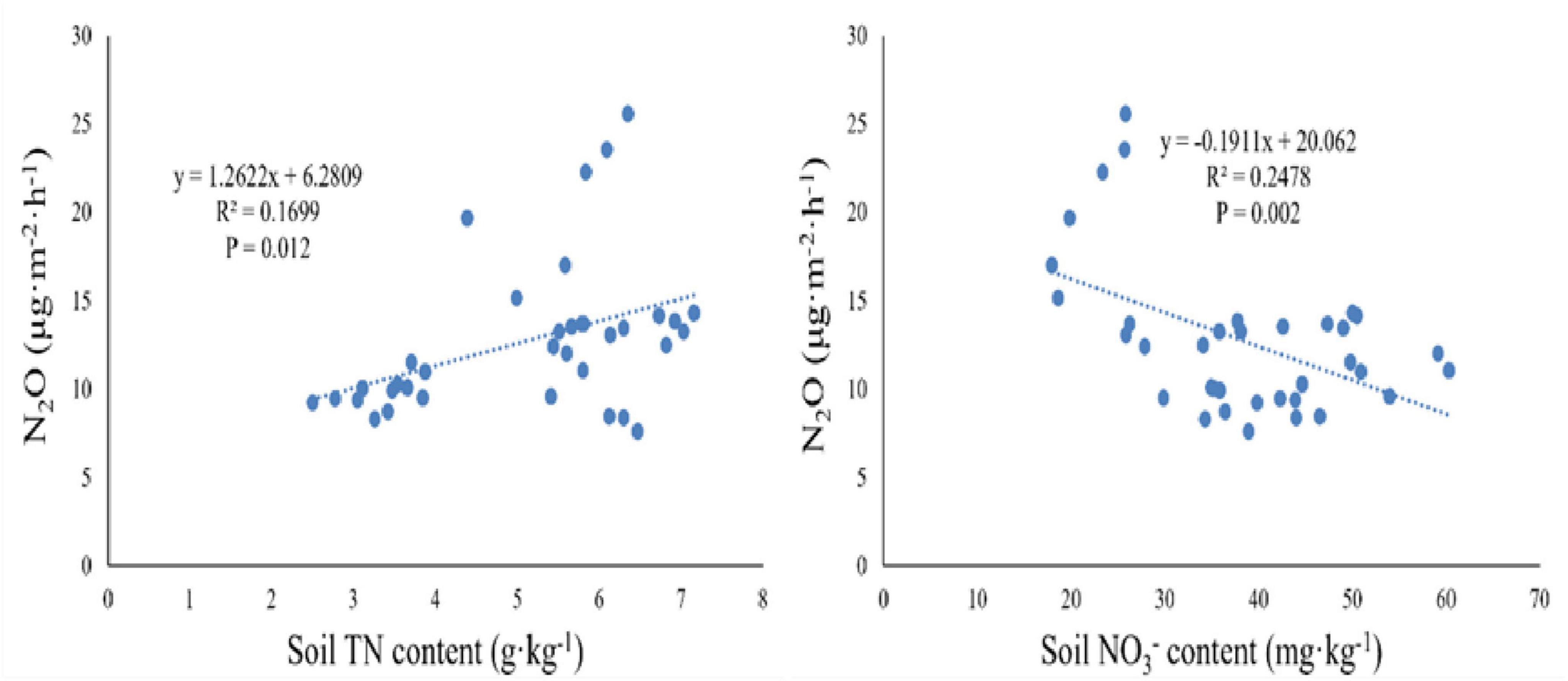
Figure 5. Relationship between soil N2O flux and two environmental variables (total nitrogen (TN) and nitrate nitrogen (NO3–) at 10 cm soil depths).
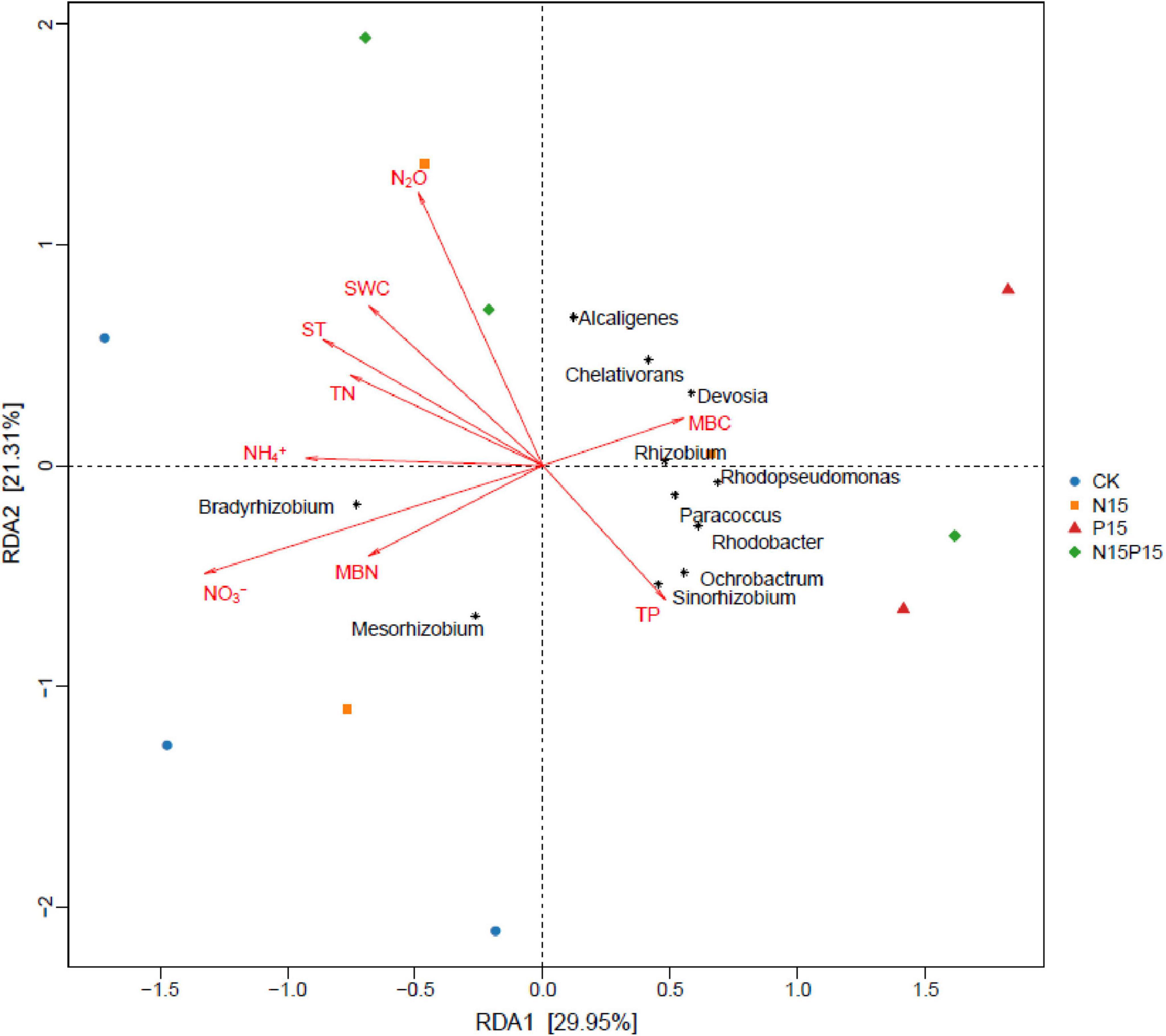
Figure 6. Redundancy analysis (RDA) between N2O emissions and NirK bacterial community structure (genus level) and soil characteristics. The length and angle of the arrows indicates the magnitude and direction of the correlation, respectively. CK, Control; N15, CON2H4 addition treatment; P15, NaH2PO4 addition treatment; N15P15, CON2H4 and NaH2PO4 addition treatment.
4 Discussion
4.1 Effects of nitrogen and phosphorus addition on N2O flux in wet meadow
N2O is mainly produced by two biological processes, nitrification and denitrification, which are affected by the soil environment and substrate nutrient content (Cui et al., 2016). After two years of phosphate addition treatment alone in the Qinghai-Tibet Plateau wet meadow, no significant effects on N2O fluxes were found. This fails to confirm our first hypothesis but is consistent with previous studies on alpine grasslands and wetlands (Wang et al., 2018; Zhang et al., 2019). The significant positive effect of phosphate addition on N2O flux has been reported in grasslands. It is attributed to the increase in soil-denitrifying microbes biomass and activity under the phosphate addition treatment (Cui et al., 2018). Although soil total phosphorus content was increased due to the phosphate addition treatment, no effect on N2O fluxes has been detected in this study. Contrary to the phosphate addition treatment, N fertilization (N15) significantly increased N2O emission, which confirms our second hypothesis that N fertilization relieves N element limitation in wet meadow systems and promotes N2O emissions. On the one hand, N and P addition treatment can increase soil available nitrogen (Table 1), promote the abundance of soil microbial functional genes, and increase the reaction matrix available for N2O production (Mori et al., 2017). On the other hand, nitrogen addition will weaken the limiting effect of nutrient elements, accelerate the decomposition rate of litter, increase the content and distribution of soil nutrients (Jiang et al., 2021), and promote the emission of N2O.
Similar to the second hypothesis, we found that phosphate addition trended to intensify the effect of N deposition treatment (N15P15) and it was significant in the early growth season of 2020 (Figure 2). The comprehensive impact of nitrogen and phosphorus addition on N2O emissions is mainly due to the competition between plants and soil microorganisms for nutrient availability (Shen and Zhu, 2022). The simultaneous addition of NP not only relieved the N limitation of the soil in the Gahai wet meadow, but also increased the available nutrients in the soil. Adequate P element increased the activity of soil microorganisms and extracellular enzymes (Mehnaz et al., 2019; Camenzind et al., 2016), promoted the mineralization of nitrogen in the ecosystem (Anderson et al., 2023), and stimulated the occurrence of denitrification (Gao et al., 2015), resulting in that the addition of P enhanced the influence of nitrogen deposition on N2O emission. To survive, soil microorganisms will promote the mineralization and decomposition of soil TN into available nitrogen (MBN, NH4+, NO3–), resulting in a lower N2O flux under phosphorus addition conditions than under nitrogen addition treatment. This result is consistent with Baral et al.’s (2014) finding that phosphorus addition reduces N2O emissions, mainly due to the Gahai wet meadow being an N limited ecosystem (Wu et al., 2021). Phosphorus addition reduces the absorption of soil mineral nitrogen by plant roots and nitrogen assimilation by soil microorganisms (Guan et al., 2024; Chen et al., 2023), ultimately reducing N2O emissions. Additionally, the N2O flux under N and P addition gradually decreased with the extension of the season (Figure 3), with the minimum value appearing at the end of plant growth (September-October), which was mainly related to the rapid depletion of nutrients after N and P addition (Gebremichael et al., 2022; Wei et al., 2020). On the one hand, the available nutrients in the soil gradually decrease with the extension of nitrogen and phosphorus addition time, and the nitrification and denitrification processes gradually weaken due to the decrease in substrate concentration, resulting in the peak of N2O flux at the beginning of plant growth after nitrogen and phosphorus addition. On the other hand, this temporal trend is linked to the rainfall pattern characteristic of the Gahai wet meadow area (Wu et al., 2023), with rainfall mainly concentrated in the plant growing season (May-October). The elevated temperatures during this period enhance soil microbial activity, fostering the microbial decomposition of nitrogen and phosphorus nutrients and litter. This, in turn, supplements the substrate concentration for soil microbial nitrification and denitrification processes (Luo et al., 2020), leading to an initial increase in N2O flux. However, as the growing season progresses, nutrient competition between plants and soil microorganisms emerges (Jones et al., 2018). Consequently, soil nutrient content and soil N2O flux gradually decline. In contrast, the N2O flux under the CK treatment displayed a trend of first decreasing and then increasing with the extension of the season. Because plant growth absorbs the available nutrients in the soil, the competition between soil microorganisms and plants for soil nutrients is intensified (Das et al., 2022). In addition, P addition alone significantly increased N2O emissions in the middle growing season, because higher temperatures increased N mineralization and decomposition by soil microorganisms and increased nutrient content in the soil (Table 1), P addition promoted soil microbial and extracellular enzyme activities that enhanced soil denitrification (Mori, 2022; Mehnaz et al., 2019; Baral et al., 2014), and contributed to N2O emissions. During the late growth period, aboveground plants will turn yellow or even die; the decomposition of aboveground litter by soil microorganisms increases the effective nutrients in the soil (Ochoa-Hueso et al., 2020), promotes soil nutrient cycling processes and increases the substrate content of N2O production (Pandeya et al., 2020), leads to an increase in N2O emissions under CK treatment at the late growth period.
4.2 Effects of environmental and microorganism factors on N2O flux
N and P addition not only induces shifts in the soil environmental factors of wet meadows (e.g., SWC and ST) but also leads to changes in soil nitrogen components (e.g., TN, NH4+, NO3–, MBN) and soil microbial community structure, ultimately changing soil N2O flux. Prior research has shown that soil total nitrogen and nitrate are substrates for nitrification and denitrification processes that produce N2O and soil microorganisms usually absorb mineral nitrogen for nitrification and denitrification, resulting in reduced nitrogen component content in the soil and promoting N2O emission (Kuang et al., 2018). Consistent with this understanding, our study reveals a significant correlation between soil N2O flux, nitrate nitrogen, and TN content (Table 2 and Figure 5). Moreover, prior research has consistently underscored soil temperature and humidity as principal drivers influencing N2O flux. This correlation stems from the direct impact of temperature on soil microbial activity, and the indirect influence of soil water content on denitrification processes by regulating anaerobic conditions (Bååth, 2018; Wu et al., 2022). Surprisingly, unlike the anticipated association outlined in our four hypotheses, no significant correlation was observed between N2O flux and temperature, NH4+, and MBN. This incongruity challenges the notion that soil temperature and nitrogen composition are the primary controllers of N2O flux. In contrast, our findings reveal a substantial positive correlation between N2O flux and SWC. This alignment emphasizes the influential role of soil water content in shaping the redox state and microbial activity within wet meadow soils. Increased SWC promotes denitrification processes, thereby influencing the production and transport of N2O (Xu et al., 2016).
Similar to the third hypothesis, we found that N addition and NP co-addition can promote soil N2O emissions by changing the relative abundance of dominant species in the soil denitrification bacterial community. Although P addition significantly altered the relative abundance of dominant species in the soil denitrification bacterial community, no significant effect was observed on the average N2O emission. Specifically, N and P addition resulted in a decrease in the relative abundance of Bradyrhizobium and an increase in the relative abundance of Devosia and Rhizobium genera. This observed microbial community restructuring aligns with prior research by Xu et al. (2019): as Bradyrhizobium is a slow-growing bacteria, N and P addition increases the available nitrogen content in the soil. Plants absorb a large amount of mineral nitrogen for root growth, which increases soil ventilation (Table 1). A higher concentration of oxygen will cause Bradyrhizobium to consume more energy to protect the nitrogenase from oxygen inactivation (Lin et al., 2018), decreasing the abundance and nitrogen component content of Bradyrhizobium. Consistent with previous studies showing that reduced microbial nitrogen fixation capacity increases N2O emissions (Mori et al., 2014), this confirms our second hypothesis that N and P addition alters N2O emissions by modifying the proportion of prevailing species within soil denitrifying bacterial communities. Moreover, as reflected in Table 1, the observed increase in the relative abundance of fast-growing nitrogen-fixation bacteria, such as Devosia and Rhizobium, due to nitrogen and phosphorus addition contributes to heightened soil microbial activity (MBN and MBC). This stimulation, in turn, facilitated the rapid propagation of Devosia and Rhizobium, directly promoting the soil nitrogen cycle (Kuypers et al., 2018; Fang et al., 2019). The consequence is an indirect increase in N2O emission. Notably, Devosia and Rhizobium are known to promote plant growth (Zhou et al., 2017; Yang et al., 2021). Consequently, the accelerated absorption of available nitrogen by wet meadow plants, facilitated by these bacteria, reduces soil nitrogen component content. This finding is consistent with the results demonstrating a negative correlation between Devosia, Rhizobium, and soil NH4+, NO3–, MBN, alongside a significant positive correlation with MBC (Figure 6). Contrastingly, Ochrobactrum exhibits a negative correlation with N2O, attributing to its capacity to degrade aromatic and hydrocarbons (Veeranagouda et al., 2006). This microbial activity converts high levels of NO2– to N2 (Doi et al., 2009) through denitrification under anaerobic conditions. In this study, the addition of nitrogen and phosphorus resulted in higher soil available nutrient content, promoting the growth of soil microorganisms and increasing the relative abundance of Ochrobactrum (Figure 3). This, in turn, stimulated the conversion of NO2– into N2, and reduced the production of N2O. Therefore, the availability of soil nitrogen and the relative abundance of microorganisms emerge as pivotal limiting factors for soil denitrification in QTP wet meadows. These intricate microbial and nutrient dynamics underscore the importance of considering alpine wet meadows in the prediction of global greenhouse gas emissions and climate models, particularly in relation to N2O emissions. The unique geography and sensitive climatic conditions of the Gahai wet meadow system have resulted in much higher N2O emissions than elsewhere (Tiemeyer et al., 2016; Jiang et al., 2010), soil microbes and effective nutrients are the main factors influencing N2O emissions. The N2O warming potential is about 296 times that of carbon dioxide, contributing about 7% to global warming (Feng and Li, 2023), becoming the main destroyer of the ozone layer. To accurately assess the contribution of global climate change and nutrient inputs to the warming effect, the model needs to clarify the response of soil N2O emissions to nutrient inputs (Prentice et al., 2012).
5 Conclusion
This study examined the effects of high levels of N and P additions on N2O flux within alpine wet meadow ecosystems. The impact of phosphate addition treatment on soil N2O flux was not detectable, while a significant effect of N deposition treatment was shown. A trend that phosphate addition intensified the effect of N deposition treatment on soil N2O flux was observed, which was significant in the early growth season. Among different abiotic factors, soil TN and NO3– were the main controls for N2O emission, while SWC has a weaker impact on N2O flux. Furthermore, our research revealed a strong correlation between N2O flux and soil available nitrogen and the relative abundance of NirK microorganisms (Bradyrhizobium, Devosia, Ochrobactrum, Alcaligenes, Rhizobium). In the alpine wet meadow ecosystem, the denitrification process is constrained by nitrogen availability and microbial biomass carbon and phosphorus content. Our results indicated a shift in the main limiting factor from nitrogen to phosphorus in response to nutrient addition, suggesting a change in the ecological dynamics of this area. Nitrogen alone and nitrogen-phosphorus interactions were all found to significantly amplify the environmental pressure associated with N2O emission in wet meadows. This insight underscores the importance of considering the intricate interplay between nutrient dynamics, microbial communities, and environmental factors for a comprehensive understanding of the consequences of nutrient additions in alpine wet meadows.
Data availability statement
The original contributions presented in this study are included in this article, further inquiries can be directed to the corresponding author.
Author contributions
JW: Data curation, Funding acquisition, Methodology, Writing – original draft. HW: Methodology, Data curation, Investigation, Writing – original draft. GL: Funding acquisition, Methodology, Supervision, Writing – review and editing.
Funding
The authors declare that financial support was received for the research, authorship, and/or publication of this article. This work was supported by the National Natural Science Foundation of China (42307348), Fuxi Young Talents Project of Gansu Agricultural University (Gaufx-05Y05), the Top-notch Leading Talent Project in Gansu Province (GSBJLJ-2023-09), and the Doctoral Research Fund Project of Lanzhou City University (LZCU-BS2024-04).
Acknowledgments
We would like to thank all the people who were involved in the field and laboratory work.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Footnotes
References
Anderson, F. C., Clough, T. J., Condron, L. M., Richards, K. G., and Rousset, C. (2023). Nitrous oxide responses to long-term phosphorus application on pasture soil. N. Zeal. J. Agr. Res. 66, 171–188. doi: 10.1080/00288233.2021.2014528
Bå,åth, E. (2018). Temperature sensitivity of soil microbial activity modeled by the square root equation as a unifying model to differentiate between direct temperature effects and microbial community adaptation. Global Change Biol. 24, 2850–2861. doi: 10.1111/gcb.14285
Baral, B. R., Kuyper, T. W., and Van Groenigen, J. W. (2014). Liebig’s law of the minimum applied to a greenhouse gas: alleviation of P-limitation reduces soil N2O emission. Plant Soil 374, 539–548. doi: 10.1007/s11104-013-1913-8
Bardon, C., Poly, F., el Zahar Haichar, F., Le Roux, X., Simon, L., Meiffren, G., et al. (2017). Biological denitrification inhibition (BDI) with procyanidins induces modification of root traits, growth and N status in Fallopia x bohemica. Soil Biol. Biochem. 107, 41–49. doi: 10.1016/j.soilbio.2016.12.009
Camenzind, T., Hempel, S., Homeier, J., Horn, S., Velescu, A., Wilcke, W., et al. (2014). Nitrogen and phosphorus additions impact arbuscular mycorrhizal abundance and molecular diversity in a tropical montane forest. Glob. Change Biol. 20, 3646–3659. doi: 10.1111/gcb.12618
Camenzind, T., Homeier, J., Dietrich, K., Hempel, S., Hertel, D., Krohn, A., et al. (2016). Opposing effects of nitrogen versus phosphorus additions on mycorrhizal fungal abundance along an elevational gradient in tropical montane forests. Soil Biol Biochem. 94, 37–47. doi: 10.1016/j.soilbio.2015.11.011
Chen, H., Li, D., Zhao, J., Xiao, K., and Wang, K. (2018). Effects of nitrogen addition on activities of soil nitrogen acquisition enzymes: a meta-analysis. Agr. Ecosyst. Environ. 252, 126–131. doi: 10.1016/j.agee.2017.09.032
Chen, J., Ma, X., Lu, X., Xu, H., Chen, D., Li, Y., et al. (2023). Long-term phosphorus addition alleviates CO2 and N2O emissions via altering soil microbial functions in secondary rather primary tropical forests. Environ. Pollut. 323:121295. doi: 10.1016/j.envpol.2023.121295
Chen, Z., Luo, X., Hu, R., Wu, M., Wu, J., and Wei, W. (2010). Impact of long-term fertilization on the composition of denitrifier communities based on nitrite reductase analyses in a paddy soil. Microb. Ecol. 60, 850–861. doi: 10.1007/s00248-010-9700-z
Cui, P., Fan, F., Yin, C., Song, A., Huang, P., Tang, Y., et al. (2016). Long-term organic and inorganic fertilization alters temperature sensitivity of potential N2O emissions and associated microbes. Soil Biol Biochem. 93, 131–141. doi: 10.1016/j.soilbio.2015.11.005
Cui, Y., Xiao, R., Xie, Y., and Zhang, M. (2018). Phosphorus fraction and phosphate sorption-release characteristics of the wetland sediments in the Yellow River Delta. Phys. Chem. Earth 103, 19–27. doi: 10.1016/j.pce.2017.06.005
Das, P. P., Singh, K. R., Nagpure, G., Mansoori, A., Singh, R. P., Ghazi, I. A., et al. (2022). Plant-soil-microbes: a tripartite interaction for nutrient acquisition and better plant growth for sustainable agricultural practices. Environ. Res. 214:113821. doi: 10.1016/j.envres.2022.113821
DeForest, J. L., and Otuya, R. K. (2020). Soil nitrification increases with elevated phosphorus or soil pH in an acidic mixed mesophytic deciduous forest. Soil Biol Biochem. 142:107716. doi: 10.1016/j.soilbio.2020.107716
Doi, Y., Takaya, N., and Takizawa, N. (2009). Novel denitrifying bacterium Ochrobactrum anthropi YD50. 2 tolerates high levels of reactive nitrogen oxides. Appl. Environ. Microb. 75, 5186–5194. doi: 10.1128/AEM.00604-09
Fang, W., Yan, D., Wang, Q., Huang, B., Ren, Z., Wang, X., et al. (2019). Changes in the abundance and community composition of different nitrogen cycling groups in response to fumigation with 1, 3-dichloropropene. Sci. Total Environ. 650, 44–55. doi: 10.1016/j.scitotenv.2018.08.432
Feng, R., and Li, Z. (2023). Current investigations on global N2O emissions and reductions: prospect and outlook. Environ. Pollut. 338:122664. doi: 10.1016/j.envpol.2023.122664
Gao, G. F., Li, P. F., Zhong, J. X., Shen, Z. J., Chen, J., Li, Y. T., et al. (2019). Spartina alterniflora invasion alters soil bacterial communities and enhances soil N2O emissions by stimulating soil denitrification in mangrove wetland. Sci. Total Environ. 653, 231–240. doi: 10.1016/j.scitotenv.2018.10.277
Gao, Y., Chen, H., and Zeng, X. (2014). Effects of nitrogen and sulfur deposition on CH4 and N2O fluxes in high-altitude peatland soil under different water tables in the Tibetan Plateau. Soil Sci. Plant Nutr. 60, 404–410. doi: 10.1080/00380768.2014.893812
Gao, Y., Chen, H., Schumann, M., Wu, Y., and Zeng, X. (2015). Short-term responses of nitrous oxide fluxes to nitrogen and phosphorus addition in a peatland on the Tibetan Plateau. Environ. Eng. Manag. J. 14, 121–127. doi: 10.30638/eemj.2015.014
Gebremichael, A. W., Wall, D. P., O’Neill, R. M., Krol, D. J., Brennan, F., Lanigan, G., et al. (2022). Effect of contrasting phosphorus levels on nitrous oxide and carbon dioxide emissions from temperate grassland soils. Sci. Rep. 12:2602. doi: 10.1038/s41598-022-06661-2
Gong, Y., and Wu, J. (2021). Vegetation composition modulates the interaction of climate warming and elevated nitrogen deposition on nitrous oxide flux in a boreal peatland. Glob. Change Biol. 27, 5588–5598. doi: 10.1111/gcb.15865
Guan, Z. H., Cao, Z., Li, X. G., Scholten, T., Kühn, P., Wang, L., et al. (2024). Soil phosphorus availability mediates the effects of nitrogen addition on community-and species-level phosphorus-acquisition strategies in alpine grasslands. Sci. Total Environ. 906:167630. doi: 10.1016/j.scitotenv.2023.167630
Jiang, C., Yu, G., Fang, H., Cao, G., and Li, Y. (2010). Short-term effect of increasing nitrogen deposition on CO2, CH4 and N2O fluxes in an alpine meadow on the Qinghai-Tibetan Plateau, China. Atmos Environ. 44, 2920–2926. doi: 10.1016/j.atmosenv.2010.03.030
Jiang, J., Wang, Y. P., Liu, F., Du, Y., Zhuang, W., Chang, Z., et al. (2021). Antagonistic and additive interactions dominate the responses of belowground carbon-cycling processes to nitrogen and phosphorus additions. Soil Biol. Biochem. 156:108216. doi: 10.1016/j.soilbio.2021.108216
Jiang, J., Zong, N., Song, M., Shi, P., Ma, W., Fu, G., et al. (2013). Responses of ecosystem respiration and its components to fertilization in an alpine meadow on the Tibetan Plateau. Eur. J. Soil Biol. 56, 101–106. doi: 10.1016/j.ejsobi.2013.03.001
Jones, D. L., Magthab, E. A., Gleeson, D. B., Hill, P. W., Sánchez-Rodríguez, A. R., Roberts, P., et al. (2018). Microbial competition for nitrogen and carbon is as intense in the subsoil as in the topsoil. Soil Biol. Biochem. 117, 72–82. doi: 10.1016/j.soilbio.2017.10.024
Kim, G. W., Lim, J. Y., Bhuiyan, M. S. I., Das, S., Khan, M. I., and Kim, P. J. (2022). Investigating the arable land that is the main contributor to global warming between paddy and upland vegetable crops under excessive nitrogen fertilization. J. Clean Prod. 346:131197. doi: 10.1016/j.jclepro.2022.131197
Kim, S. Y., Veraart, A. J., Meima-Franke, M., and Bodelier, P. L. (2015). Combined effects of carbon, nitrogen and phosphorus on CH4 production and denitrification in wetland sediments. Geoderma 259, 354–361. doi: 10.1016/j.geoderma.2015.03.015
Kuang, W., Gao, X., Gui, D., Tenuta, M., Flaten, D. N., Yin, M., et al. (2018). Effects of fertilizer and irrigation management on nitrous oxide emission from cotton fields in an extremely arid region of northwestern China. Field Crop Res. 229, 17–26. doi: 10.1016/j.fcr.2018.09.010
Kuypers, M. M., Marchant, H. K., and Kartal, B. (2018). The microbial nitrogen-cycling network. Nat. Rev. Microbiol. 16, 263–276. doi: 10.1038/nrmicro.2018.9
Lin, Y., Ye, G., Liu, D., Ledgard, S., Luo, J., Fan, J., et al. (2018). Long-term application of lime or pig manure rather than plant residues suppressed diazotroph abundance and diversity and altered community structure in an acidic Ultisol. Soil Biol. Biochem. 123, 218–228. doi: 10.1016/j.soilbio.2018.05.018
Liu, X., Zhang, Y., Han, W., Tang, A., Shen, J., Cui, Z., et al. (2013). Enhanced nitrogen deposition over China. Nature 494, 459–462. doi: 10.1038/nature11917
Luo, G., Xue, C., Jiang, Q., Xiao, Y., Zhang, F., Guo, S., et al. (2020). Soil carbon, nitrogen, and phosphorus cycling microbial populations and their resistance to global change depend on soil C: N: P stoichiometry. Msystems 5, e00162–20. doi: 10.1128/msystems.00162-20
Ma, W., Alhassan, A. R. M., Wang, Y., Li, G., Wang, H., and Zhao, J. (2018). Greenhouse gas emissions as influenced by wetland vegetation degradation along a moisture gradient on the eastern Qinghai-Tibet Plateau of North-West China. Nutr. Cycl. Agroecosys. 112, 335–354. doi: 10.1007/s10705-018-9950-6
Martínez-Espinosa, C., Sauvage, S., Al Bitar, A., Green, P. A., Vörösmarty, C. J., and Sánchez-Pérez, J. M. (2021). Denitrification in wetlands: a review towards a quantification at global scale. Sci. Total Environ. 754:142398. doi: 10.1016/j.scitotenv.2020.142398
Mehnaz, K. R., and Dijkstra, F. A. (2016). Denitrification and associated N2O emissions are limited by phosphorus availability in a grassland soil. Geoderma 284, 34–41. doi: 10.1016/j.geoderma.2016.08.011
Mehnaz, K. R., Corneo, P. E., Keitel, C., and Dijkstra, F. A. (2019). Carbon and phosphorus addition effects on microbial carbon use efficiency, soil organic matter priming, gross nitrogen mineralization and nitrous oxide emission from soil. Soil Biol. Biochem. 134, 175–186. doi: 10.1016/j.soilbio.2019.04.003
Mori, T. (2022). The ratio of β-1, 4-glucosidase activity to phosphomonoesterase activity remains low in phosphorus-fertilized tropical soils: a meta-analysis. Appl. Soil Ecol. 180:104635. doi: 10.1016/j.apsoil.2022.104635
Mori, T., Ohta, S., Ishizuka, S., Konda, R., Wicaksono, A., and Heriyanto, J. (2014). Phosphorus application reduces N2O emissions from tropical leguminous plantation soil when phosphorus uptake is occurring. Biol. Fert. Soils. 50, 45–51. doi: 10.1007/s00374-013-0824-4
Mori, T., Wachrinrat, C., Staporn, D., Meunpong, P., Suebsai, W., Matsubara, K., et al. (2017). Effects of phosphorus addition on nitrogen cycle and fluxes of N2O and CH4 in tropical tree plantation soils in Thailand. Agric. Nat. Reso. 51, 91–95. doi: 10.1016/j.anres.2016.03.002
Na, X., Yu, H., Wang, P., Zhu, W., Niu, Y., and Huang, J. (2019). Vegetation biomass and soil moisture coregulate bacterial community succession under altered precipitation regimes in a desert steppe in northwestern China. Soil Biol. Biochem. 136:107520. doi: 10.1016/j.soilbio.2019.107520
Ochoa-Hueso, R., Borer, E. T., Seabloom, E. W., Hobbie, S. E., Risch, A. C., Collins, S. L., et al. (2020). Microbial processing of plant remains is co-limited by multiple nutrients in global grasslands. Glob. Change Biol. 26, 4572–4582. doi: 10.1111/gcb.15146
Pandeya, H. R., Friedl, J., De Rosa, D., Asis, C. T., Tilbrook, J., Scheer, C., et al. (2020). Combined effect of nitrogen fertiliser and leaf litter carbon drive nitrous oxide emissions in tropical soils. Nutr. Cycl. Agroecosys. 118, 207–222. doi: 10.1007/s10705-020-10094-6
Peñuelas, J., and Sardans, J. (2022). The global nitrogen-phosphorus imbalance. Science. 375, 266–267. doi: 10.1126/science.abl4827
Poeplau, C., Helfrich, M., Dechow, R., Szoboszlay, M., Tebbe, C. C., Don, A., et al. (2019). Increased microbial anabolism contributes to soil carbon sequestration by mineral fertilization in temperate grasslands. Soil Biol. Biochem. 130, 167–176. doi: 10.1016/j.soilbio.2018.12.019
Prentice, I. C., Spahni, R., and Niu, H. S. (2012). Modelling terrestrial nitrous oxide emissions and implications for climate feedback. N. Phytol. 196, 472–488. doi: 10.1111/j.1469-8137.2012.04269.x
Qiu, L., Gou, X., Kong, Y., Tu, F., Peng, X., Xu, L., et al. (2023). Nitrogen addition stimulates N2O emissions via changes in denitrification community composition in a subtropical nitrogen-rich forest. J. Environ. Manage. 348:119274. doi: 10.1016/j.jenvman.2023.119274
Saha, U. K., Sonon, L., and Biswas, B. K. (2018). A comparison of diffusion-conductimetric and distillation-titration methods in analyzing ammonium-and nitrate-nitrogen in the KCl-extracts of Georgia soils. Commun. Soil Sci. Plan. 49, 63–75. doi: 10.1080/00103624.2017.1421647
Schneider, N., Islam, M., Wehrle, R., Pätzold, S., Brüggemann, N., Töpfer, R., et al. (2023). Deep incorporation of organic amendments into soils of a ‘Calardis Musqué’vineyard: effects on greenhouse gas emissions, vine vigor, and grape quality. Front. Plant Sci. 14:1253458. doi: 10.3389/fpls.2023.1253458
Shen, Y., and Zhu, B. (2022). Effects of nitrogen and phosphorus enrichment on soil N2O emission from natural ecosystems: a global meta-analysis. Environ. Pollut. 301:118993. doi: 10.1016/j.envpol.2022.118993
Shi, Y., Wang, J., Zhang, J., Ao, Y., Guo, Z., Mu, C., et al. (2021). Rainfall-associated chronic N deposition induces higher soil N2O emissions than acute N inputs in a semi-arid grassland. Agr. Forest Meteorol. 304:108434. doi: 10.1016/j.agrformet.2021.108434
Sun, Y., Peng, S., Goll, D. S., Ciais, P., Guenet, B., Guimberteau, M., et al. (2017). Diagnosing phosphorus limitations in natural terrestrial ecosystems in carbon cycle models. Earths Fut. 5, 730–749. doi: 10.1002/2016EF000472
Tiemeyer, B., Albiac Borraz, E., Augustin, J., Bechtold, M., Beetz, S., Beyer, C., et al. (2016). High emissions of greenhouse gases from grasslands on peat and other organic soils. Glob. Change Biol. 22, 4134–4149. doi: 10.1111/gcb.13303
Veeranagouda, Y., Emmanuel Paul, P. V., Gorla, P., Siddavattam, D., and Karegoudar, T. B. (2006). Complete mineralisation of dimethylformamide by Ochrobactrum sp. DGVK1 isolated from the soil samples collected from the coalmine leftovers. Appl. Microbiol. Biot. 71, 369–375. doi: 10.1007/s00253-005-0157-9
Voigt, C., Marushchak, M. E., Abbott, B. W., Biasi, C., Elberling, B., Siciliano, S. D., et al. (2020). Nitrous oxide emissions from permafrost-affected soils. Nat. Rev. Earth Env. 1, 420–434. doi: 10.1038/s43017-020-0063-9
Wang, G., Liang, Y., Ren, F., Yang, X., Mi, Z., Gao, Y., et al. (2018). Greenhouse gas emissions from the Tibetan alpine grassland: effects of nitrogen and phosphorus addition. Sustainability 10:4454. doi: 10.3390/su10124454
Wang, H., Wu, J., Li, G., Yan, L., and Wei, X. (2022). Effects of rainfall frequency on soil labile carbon fractions in a wet meadow on the Qinghai-Tibet Plateau. J. Soil Sediment. 22, 1489–1499. doi: 10.1007/s11368-022-03170-7
Wang, H., Yu, L., Zhang, Z., Liu, W., Chen, L., Cao, G., et al. (2017). Molecular mechanisms of water table lowering and nitrogen deposition in affecting greenhouse gas emissions from a Tibetan alpine wetland. Glob. Change Biol. 23, 815–829. doi: 10.1111/gcb.13467
Wei, S., Tie, L., Liao, J., Liu, X., Du, M., Lan, S., et al. (2020). Nitrogen and phosphorus co-addition stimulates soil respiration in a subtropical evergreen broad-leaved forest. Plant Soil 450, 171–182. doi: 10.1007/s11104-020-04498-0
Wen, X., Sun, R., Cao, Z., Huang, Y., Li, J., Zhou, Y., et al. (2022). Synergistic metabolism of carbon and nitrogen: cyanate drives nitrogen cycle to conserve nitrogen in composting system. Bioresour. Technol. 361, 127708. doi: 10.1016/j.biortech.2022.127708
Wrage-Mönnig, N., Velthof, G. L., Van Beusichem, M. L., and Oenema, O. (2018). The role of nitrifier denitrification in the production of nitrous oxide revisited. Soil Biol. Biochem. 123, A3–A16. doi: 10.1016/j.soilbio.2018.03.020
Wu, J., Wang, H., and Li, G. (2023). Effects of nitrogen deposition on N2O emission in a wet meadow on the Qinghai-Tibet Plateau. Appl. Soil Ecol. 191:105049. doi: 10.1016/j.apsoil.2023.105049
Wu, J., Wang, H., Li, G., Ma, W., Wu, J., Gong, Y., et al. (2020). Vegetation degradation impacts soil nutrients and enzyme activities in wet meadow on the Qinghai-Tibet Plateau. Sci. Rep. 10:21271. doi: 10.1038/s41598-020-78182-9
Wu, J., Wang, H., Li, G., Wu, J., and Ma, W. (2021). Vertical and seasonal changes in soil carbon pools to vegetation degradation in a wet meadow on the Qinghai-Tibet Plateau. Sci. Rep. 11:12268. doi: 10.1038/s41598-021-90543-6
Wu, J., Wang, H., Li, G., Wu, J., Gong, Y., and Wei, X. (2022). Unimodal response of N2O flux to changing rainfall amount and frequency in a wet meadow in the Tibetan Plateau. Ecol. Eng. 174:106461. doi: 10.1016/j.ecoleng.2021.106461
Xie, W. X., Cui, Y. Q., Du, H. N., Chen, J. L., and Zhu, K. J. (2015). The research progress of N2O emission regularity in wetlands and the nitrogen interaction on its emission. Adv. Mater. Res. 1073, 154–158.
Xu, Y., Wang, T., Li, H., Ren, C., Chen, J., Yang, G., et al. (2019). Variations of soil nitrogen-fixing microorganism communities and nitrogen fractions in a Robinia pseudoacacia chronosequence on the Loess Plateau of China. Catena 174, 316–323. doi: 10.1016/j.catena.2018.11.009
Xu, Y., Zhan, M., Cao, C., Tian, S., Ge, J., Li, S., et al. (2016). Improved water management to reduce greenhouse gas emissions in no-till rapeseed-rice rotations in Central China. Agr. Ecosyst. Environ. 221, 87–98. doi: 10.1016/j.agee.2016.01.021
Yang, W., Li, C., Wang, S., Zhou, B., Mao, Y., Rensing, C., et al. (2021). Influence of biochar and biochar-based fertilizer on yield, quality of tea and microbial community in an acid tea orchard soil. Appl. Soil Ecol. 166:104005. doi: 10.1016/j.apsoil.2021.104005
Yu, L., Wang, Y., Zhang, X., Dörsch, P., and Mulder, J. (2017). Phosphorus addition mitigates N2O and CH4 emissions in N-saturated subtropical forest, SW China. Biogeosciences 14, 3097–3109. doi: 10.5194/bg-14-3097-2017
Zhai, J., Zhou, H., Wu, Y., Wang, G., and Xue, S. (2024). Effect of nitrogen and phosphorus on soil enzyme activities and organic carbon stability in Qinghai–Tibet Plateau. Agronomy 14, 1376. doi: 10.3390/agronomy14071376
Zhang, Q., Kong, D., Shi, P., Singh, V. P., and Sun, P. (2018). Vegetation phenology on the Qinghai-Tibetan Plateau and its response to climate change (1982-2013). Agr. Forest Meteorol. 248, 408–417. doi: 10.1016/j.agrformet.2017.10.026
Zhang, Y., Wang, C., and Li, Y. (2019). Contrasting effects of nitrogen and phosphorus additions on soil nitrous oxide fluxes and enzyme activities in an alpine wetland of the Tibetan Plateau. PLoS One 14:e0216244. doi: 10.1371/journal.pone.0216244
Zhao, M., Zhao, Y., Gao, W., Xie, L., Zhang, G., Song, C., et al. (2023). Exploring the nitrogen fixing strategy of bacterial communities in nitrogen cycling by adding calcium superphosphate at various periods during composting. Sci. Total Environ. 901:166492. doi: 10.1016/j.scitotenv.2023.166492
Zhou, J., Jiang, X., Wei, D., Zhao, B., Ma, M., Chen, S., et al. (2017). Consistent effects of nitrogen fertilization on soil bacterial communities in black soils for two crop seasons in China. Sci. Rep. 7:3267. doi: 10.1038/s41598-017-03539-6
Keywords: nitrogen addition, phosphate addition, N2O flux, NirK gene, alpine wet meadow, Qinghai-Tibet Plateau
Citation: Wu J, Wang H and Li G (2024) Phosphate addition intensifies the increase in N2O emission under nitrogen deposition in wet meadows of the Qinghai-Tibet Plateau. Front. Microbiol. 15:1472792. doi: 10.3389/fmicb.2024.1472792
Received: 30 July 2024; Accepted: 02 December 2024;
Published: 18 December 2024.
Edited by:
Ming Jiang, Chinese Academy of Sciences (CAS), ChinaReviewed by:
Jianghua Wu, Memorial University of Newfoundland, CanadaYongcui Deng, Nanjing Normal University, China
Copyright © 2024 Wu, Wang and Li. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Guang Li, bGlnZ3NhdUBmb3htYWlsLmNvbQ==