 Mikołaj Kokociński1*
Mikołaj Kokociński1* Caio Graco-Roza2,3
Caio Graco-Roza2,3 Iwona Jasser4Jūratė Karosienė5Jūratė Kasperovičienė5
Iwona Jasser4Jūratė Karosienė5Jūratė Kasperovičienė5 Justyna Kobos6Judita Koreivienė5
Justyna Kobos6Judita Koreivienė5 Joanna Mankiewicz-Boczek7
Joanna Mankiewicz-Boczek7 Janne Soininen2Agnieszka Szczurowska8
Janne Soininen2Agnieszka Szczurowska8- 1Department of Hydrobiology, Institute of Environmental Biology, Adam Mickiewicz University, Poznań, Poland
- 2Department of Geosciences and Geography, University of Helsinki, Helsinki, Finland
- 3Lammi Biological Station, University of Helsinki, Lammi, Finland
- 4Faculty of Biology, Institute of Environmental Biology, University of Warsaw, Warsaw, Poland
- 5Laboratory of Algology and Microbial Ecology, State Scientific Research Institute Nature Research Centre, Vilnius, Lithuania
- 6Division of Marine Biotechnology, Faculty of Oceanography and Geography, University of ń, ń, Gdynia
- 7European Regional Centre for Ecohydrology of the Polish Academy of Sciences, Łódź, Poland
- 8Department of Botany and Plant Physiology, University of Life Sciences in Lublin, Lublin, Poland
Objective: In recent decades, the invasive cyanobacteria Raphidiopsis raciborskii and Raphidiopsis mediterranea have expanded their distribution globally, particularly in temperate regions. Understanding the ecological drivers of Raphidiopsis distribution is imperative to addressing the challenges associated with these species. Here, we aimed to characterize the distribution and biomass of R. raciborskii and R. mediterranea across 112 lakes in Poland and Lithuania in relation to local and regional factors.
Research design and methods: Integrated water samples were collected from 102 Polish and 10 Lithuanian lakes from different regions for phytoplankton and chemical analyses. The lakes varied in surface area, and exhibited diverse mixing regimes, trophic states, and morphometries. Phytoplankton was identified and quantified using a Fuchs-Rosenthal or Nageotte chamber. Additionally, we characterized the degree of human pressures the climatic constraints experienced by each lake.
Results: R. raciborskii occurrence has increased in eastern regions of Poland but biomass is relatively low compared to western Poland, likely due to lower air temperatures and nutrient concentrations, especially phosphorus. In contrast, R. mediterranea only occurred in a small number of lakes in Poland, and in a single lake in Lithuania, with no relation to measured local and regional variables.
Conclusions: Our study shows contrasting patterns in the distribution of two invasive cyanobacteria species in Europe, highlighting the importance of climate and nutrients to the distribution of R. raciborskii, the most widespread species, and providing relevant information for decision making and conservation strategies.
1 Introduction
Rising lake water temperatures related to global warming and eutrophication are two major factors enhancing the expansion of invasive cyanobacteria and the development of harmful algae blooms (HABs) (Paerl and Huisman, 2009; Kosten et al., 2011; Lürling et al., 2017). Increasing freshwater productivity due to higher level of nutrients relates strongly to changes in catchment land use (Hupfer and Hilt, 2008). Intensive land use (especially agriculture and urban activities) accelerates nutrient delivery to water bodies, particularly in the areas with poor wastewater treatment (de Jonge and Elliott, 2001). Consequently, biodiversity typically decreases, with harmful changes in ecosystem function. Furthermore, new threats may emerge when toxic cyanobacteria strains dominate and release toxins into the water.
Among the species that often dominate HABs, Raphidiopsis raciborskii (Wołoszyńska) Aguilera, Berrendero Gómez, Kástovsky, Echenique and Salerno originates from tropical and subtropical regions, but it has expanded into temperate zones over the last several decades (Padisák, 1997; Sinha et al., 2012; Burford et al., 2016; Aguilera et al., 2018). R. raciborskii was recorded in Europe for the first time at Lake Kastoria in Greece (Skuja, 1937), from where colonization of other European countries likely started (Padisák, 1997). In many countries, it is considered an invasive species and a serious threat to native phytoplankton communities, disrupting natural ecosystem processes (Sukenik et al., 2012; Wilk-Woźniak et al., 2016). Recent studies showed increasing occurrence of R. raciborskii in both hemispheres with the northernmost (57°N) recorded case in Nero Lake, Russia suggesting it is now a cosmopolitan species (Panou et al., 2018; Sidelev et al., 2020). Previous studies have also indicated that R. raciborskii, can become dominant within a relatively short period (Padisák, 1997).
Evidence suggests that R. raciborskii exhibits phenotypic plasticity, allowing it to colonize diverse freshwater ecosystems. This adaptability relates to its highly flexible ecotypes, enabling it to cope with environmental changes, including shifts in climate, explaining the successful expansion toward more northern regions (Burford et al., 2016). Studies show that R. raciborskii thrives across a wide range of water temperatures and light conditions. Nevertheless, its environmental optima is warmer waters, typically ranging between 29°C and 32°C (Thomas and Litchman, 2016; Xiao et al., 2017; Zheng et al., 2023). Blooms of R. raciborskii usually occur when water temperature exceeds 25°C (Recknagel et al., 2014; Jia et al., 2021); however, in Langer See, Germany, R. raciborskii reached its highest abundances at 24°C, even when higher temperatures were recorded during the summer seasons (Wiedner et al., 2007; Rücker et al., 2009). In tropical regions, where temperatures typically exceed 20°C, perennial blooms commonly occur (Recknagel et al., 2014; Jia et al., 2021). Despite optima in warmer waters, there are records of viable R. raciborskii populations at temperatures as low as 8°C–11°C (Bonilla et al., 2012; Dokulil, 2015).
Light availability also plays an important role in R. raciborskii distribution, with evidence of growth at light intensities ranging from 8.5 to hundreds of μmol photons m−2 s−2 (Briand et al., 2004; Carneiro et al., 2009; Bonilla et al., 2012; Mehnert et al., 2010). However, high water temperature and irradiance levels (above 2,000 μmol photons m−2 s−2) may inhibit growth and bloom formation (Kehoe et al., 2015). Available data on growth rate response to light intensity originate from different temperatures, suggesting an interactive effect between light and temperature on R. raciborskii. The ongoing rise in air temperature and light attenuation in lakes due to anthropogenic climate change may play significant role in further successful spatial expansion of R. raciborskii.
In addition to broad preferences for light and temperature, R. raciborskii also exhibits high uptake affinity and storage capacity for phosphorus (Antunes et al., 2015), which proves beneficial under conditions of pulsed discharge or fluctuating concentrations of dissolved phosphorus (Marinho et al., 2013; Amaral et al., 2014). As for nitrogen, R. raciborskii shows a preference for dissolved inorganic (ammonia and nitrate) and organic (urea) forms, but also can fix N2 nitrogen by terminal heterocytes under nitrogen depletion (Plominsky et al., 2013; Ammar et al., 2014; Burford et al., 2018). The capacity to regulate buoyancy and akinete formation support growth and dominance even under suboptimal environmental conditions (Sukenik et al., 2012). Altogether, these traits reflect multiple strains with differing physiological responses across range of environmental conditions (Pierangelini et al., 2015; Willis et al., 2015; Xiao et al., 2017), resulting in successful range expansion.
In addition to R. raciborskii, Raphidiopsis mediterranea Skuja has increased occurrence in temperate zones. Like R. raciborskii, it was initially described in Lake Kastoria, Greece, suggesting a Mediterranean origin (Kaštovský et al., 2010). R. mediterranea occurs worldwide, from tropical and subtropical regions (e.g., Australia, Brazil, China) to temperate regions (e.g., Bulgaria, Czech Republic, Serbia) (Wilk-Woźniak et al., 2016). Notably, continuous populations have been documented in six Lithuanian lakes (Kasperovičienė et al., 2010; Kaštovský et al., 2010). Despite these global reports, our knowledge of its ecological preferences and current distribution remains incomplete. R. mediterranea thrives in a variety of aquatic environments, including eutrophic to hypertrophic, shallow, and deep lakes or ponds (Kasperovičienė et al., 2010; Wilk-Woźniak et al., 2016; Aguilera et al., 2018). It exhibits adaptability to a broad temperature range, with growth observed between 0.5°C and 17.9°C (Cronberg, 1973). While some studies indicate year-round growth, R. mediterranea has only been documented in the temperate zone, for example in the Lithuanian lakes, during warmer summer months (July–August) when water temperature ranges between 20°C and 25°C (Kasperovičienė et al., 2010). Similarly to R. raciborskii, R. mediterranea has the potential to coexist with other cyanobacteria during toxic blooms (Watanabe et al., 2003; Namikoshi et al., 2003; Mohamed, 2007).
R. raciborskii and R. mediterranea are both potential cylindrospermopsin producers (McGregor et al., 2011; de La Cruz et al., 2013), but such production has not yet been reported in Europe. There are also known strains from outside Europe capable of producing neurotoxins, including saxitoxin by R. raciborskii (Lorenzi et al., 2016) and anatoxin-a by R. mediterranea (Namikoshi et al., 2003). Moreover, some European strains of R. raciborskii can produce chemical compounds of yet unknown structure that may cause oxidative stress and cytotoxic and neurotoxic effects in human or animal cells (Poniedziałek et al., 2015; Rzymski et al., 2017a; Rzymski et al., 2017b; Falfushynska et al., 2019).
A 2014 study showed widespread occurrence of R. raciborskii in lakes impacted by agriculture and urbanization in Western Poland (Kokociński et al., 2017), with R. raciborskii recorded in 25 out of the 117 lakes sampled. Notably, this study highlighted an increasing contribution of R. raciborskii to the overall phytoplankton biomass in Western Polish lakes and no occurrence of this species in Eastern Poland, including the north-eastern lake districts. R. raciborskii was less prevalent in Lithuania, observed in only one lake (Kokociński et al., 2017). In contrast, R. mediterranea exhibited limited distribution in Polish lakes compared to Lithuanian water bodies in recent years (Kasperovičienė et al., 2010). Since 2004, R. mediterranea has been identified in six out of 18 lakes investigated, contributing up to 15% of the total phytoplankton biomass (Kasperovičienė et al., 2005).
To summarize, it is imperative to deepen our understanding of the distribution and ecological factors influencing the proliferation of R. raciborskii and R. mediterranea. We therefore aim to (i) characterize the current distribution of R. raciborskii and R. mediterranea in lakes across Poland and Lithuania, (ii) explore the roles played by local environmental variables, land use and climate in shaping the distribution of R. raciborskii and R. mediterranea, and (iii) investigate whether the occurrence and biomass of R. raciborskii have increased in recent years.
2 Materials and methods
2.1 Study sites
To examine the environmental factors driving the distribution of R. raciborskii and R. mediterranea in the temperate zone, we sampled 102 Polish and 10 Lithuanian lakes from different regions (Supplementary Table 1). The lakes varied in surface area, ranging from 5 to 11.4 ha, and exhibited diverse mixing regimes, trophic states, and morphometries. Among the 112 lakes surveyed, 25 were classified as very shallow (max. depth < 5 m), 34 as shallow (5–10 m), and 53 as deep (> 10 m). Additionally, land use showed considerable variation, with Western Poland featuring agricultural and urban catchments, while Eastern Poland and Lithuanian catchments were predominantly characterized by forest land use.
2.2 Sample collection
Samples were collected once for each lake during the summer (July–September) 2020. Integrated phytoplankton samples were collected from the water column in polymictic lakes or epilimnion in stratified lakes from one sampling station using bathometer. The phytoplankton samples were preserved with acidified Lugol’s solution with a final concentration of 1% (APHA et al., 2012; Hötzel and Croome, 1999) and stored under cool and dark conditions until analysis.
2.3 Phytoplankton analysis
Phytoplankton was identified and quantified using a Fuchs-Rosenthal or Nageotte chamber. Two subsamples were analyzed for each lake. To ensure accuracy, a minimum of 400 cells or filaments were counted, reducing the error to less than 10% (Olenina et al., 2006). Phytoplankton biomass, measured in fresh weight, was determined through cell volumetric analysis using geometric approximation and expressed as a wet weight following Wetzel and Likens (2000).
2.4 Physico-chemical analysis
Simultaneously with the collection of phytoplankton samples, water samples were collected for chemical analyses. Total nitrogen (TN), total phosphorus (TP), dissolved nitrogen (DN) and total reactive phosphorus (TRP) concentrations were determined by a spectrophotometric method. For a chlorophyll a (chl a) analysis, 200–500 mL of water was filtered through a GF/C Whatman filter. The concentration was determined spectrophotometrically after extraction with 90% acetone, and calculations were based on Lorenzen’s formula (Wetzel and Likens, 2000). Field measurements included water temperature, pH, and conductivity using a multiparameter probe, while water transparency was measured using a Secchi disk (SD).
2.5 Land use and climatic variables
We characterized the degree of human pressures experienced by each lake by estimating the proportion of urban and agricultural environments within a radial buffer of 2 km around the lake centroid. The human-associated land cover classes (two out of 17 landcover classes in total) were obtained from the “Land_Cover_Type_1” raster layer of the “MCD12Q1” data product of NASA’s Moderate Resolution Imaging Spectroradiometer (MODIS) (Friedl and Sulla-Menashe, 2022). In addition, we characterized the climatic constraints of each lake by using measures of the mean annual temperature (Air temperature; °C) and total annual precipitation (Precipitation; mm) from a 1 km radial buffer around each lake centroid. The climatic variables used (two out of 19 variables) were obtained from WorldClim v.2.0 (Fick and Hijmans, 2017).
3 Data analysis
We analyzed data using R version 4.2.0 (R Core Team, 2022) and the packages stats v.4.2.0 (R Core Team, 2022), “nlme” v.3.1–157 (Pinheiro and Bates, 2000; Pinheiro and Bates, 2022), and the “tidyverse” suite of packages (Wickham et al., 2019).
To determine the role of local environmental variables, land use and climate in the distribution of R. raciborskii and R. mediterranea we modeled the relationship between R. raciborskii biomass and distribution as a function of area (Log km2), mean depth (Log m), conductivity (log μS cm–1), Secchi depth (log m), total phosphorus (log mg l−1), total reactive phosphorus (log mg l−1), total nitrogen (log mg l−1), air and water temperature (°C), precipitation (mm), proportion of urban area (% in 2 km radial buffer), and proportion of agricultural area (% in 2 km radial buffer) using a two-step modeling approach. First, we modeled the local and broad scale factors explaining R. raciborskii presence. We included the distribution of R. raciborskii across lakes as a response variable to be explained by the set of environmental factors using a general linear model (GLM) following a binomial distribution of the errors fitted with the package “stats.” Second, we modeled the effects of local and broad scale factors on R. raciborskii biomass using generalized least squares regression (GLS) fitted with the package “nlme.” To account for spatial autocorrelation, we introduced an exponential correlation structure on the geographic coordinates (longitude and latitude) of each lake. We validated our models using a suite of diagnostic tests via the check-model function from the performance package (version 0.12.3) in R. Specifically, we evaluated residual normality (using Q–Q plots), homoscedasticity (by plotting residuals versus fitted values), and the influence of individual observations (using leverage and Cook’s distance). In addition, we computed Variance Inflation Factors to assess multicollinearity. Collectively, these diagnostics confirmed that our models satisfied the necessary assumptions for reliable inference (Zuur and Ieno, 2016). Model performance was moderate, explaining 0.26 and 0.30 of the variation in biomass and distribution of R. raciborskii, respectively. In addition, we used Generalized Additive Model to test for potential non-linear relationships between predictor variables and the biomass and distribution of R. raciborskii. Analysis did not show any non-linear relationships and did not explain more variability in species occurrence and thus results are not shown here.
We note that R. mediterranea was identified in only seven lakes, and therefore provided insufficient sample size for modeling the occurrence and biomass of this species.
4 Results and discussion
4.1 Distribution pattern of R. raciborskii and R. mediterranea
Our study contributes to the understanding of R. raciborskii and R. mediterranea distribution in Central East Europe. We identified R. raciborskii in 31 out of 112 surveyed lakes (Figure 1A) and occurrence was related mainly to decreasing TRP and increasing TN (Table 1). Kokociński et al. (2017) showed that since 2006, R. raciborskii biomass has consistently increased, although it is not a dominant species, in contrast to southern European regions, where R. raciborskii frequently dominates the phytoplankton community (Bolius et al., 2017). Our research identifies new occurrences of R. raciborskii in lakes within the Warmia and Masuria district and other regions of eastern Poland since 2007, when this species was only reported in two lakes (Jakubowska et al., 2013).
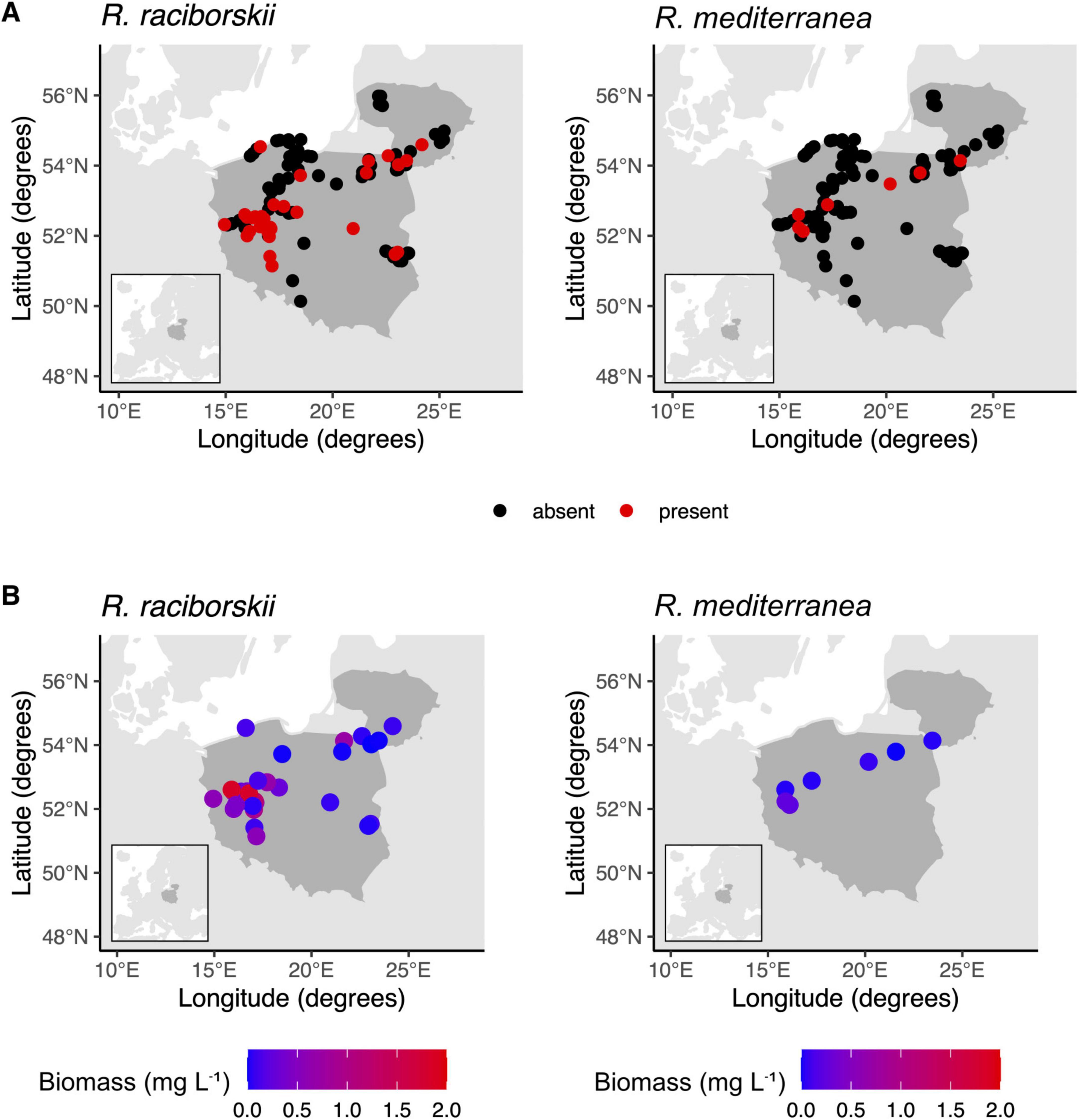
Figure 1. The occurrence (A) and biomass (B) of Raphidiopsis raciborskii and Raphidiopsis mediterranea in Polish and Lithuanian lakes.
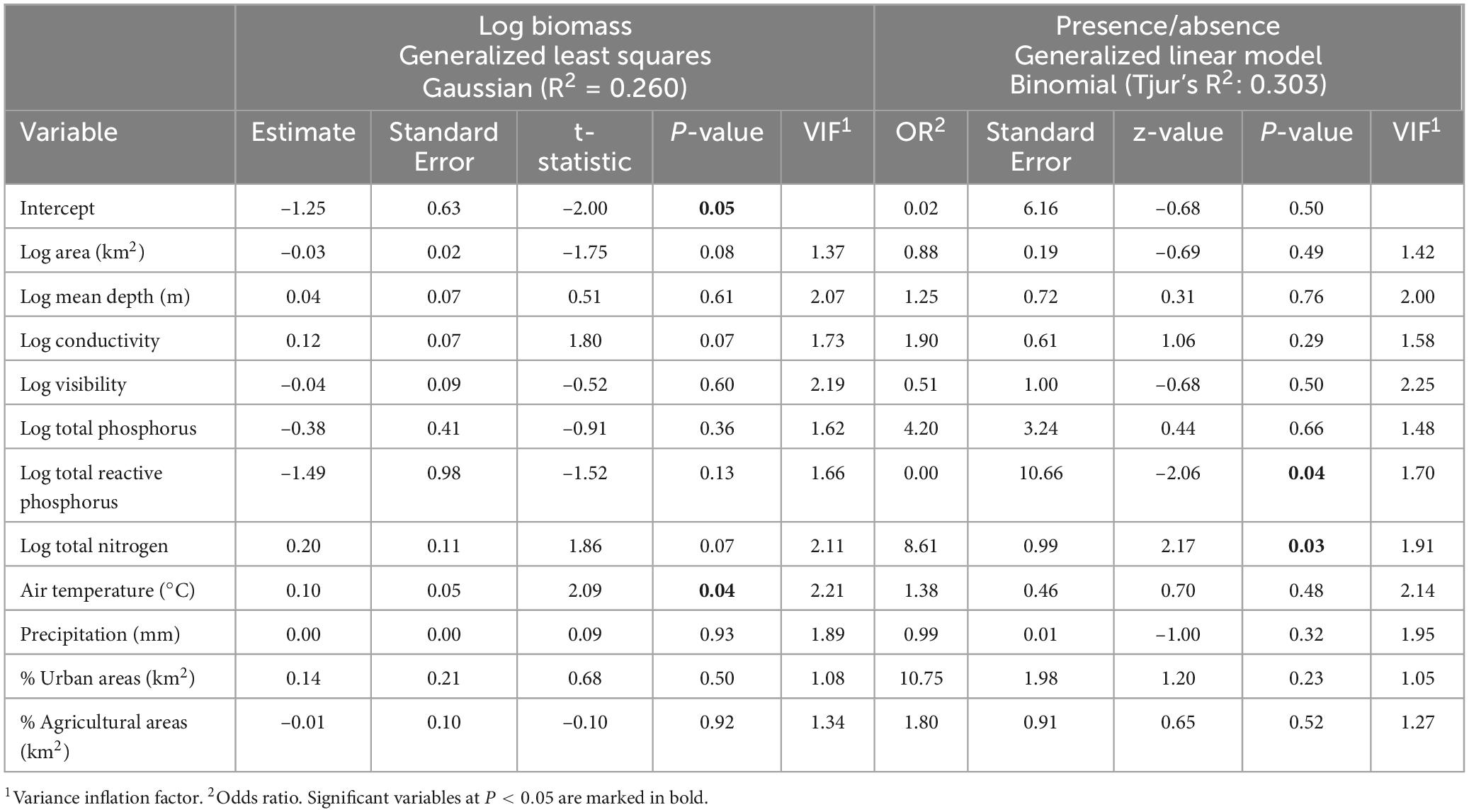
Table 1. Modeling the distribution of the species Raphidiopsis raciborskii.
We did not find any new occurrences of either species in Lithuanian lakes. Despite expanding occurrence of R. raciborskii to the east, our data show that biomass differed between regions. R. raciborskii biomass ranged from 0.02 to 5.90 mg L−1 and was much higher in lakes in Western Poland (on average 1.08 mg L−1) than Eastern Poland (on average 0.14 mg L−1) (Figure 1B and Supplementary Table 1), mainly driven by increased air temperature (Table 1). Contribution to total phytoplankton biomass ranged from 0.1% to 31% and was also higher in Western Poland (on average 7.4%) than Eastern Poland (on average 0.9%). Moreover, lower occurrence and contribution to phytoplankton biomass in Eastern Poland, along with the sole occurrence in Lithuania, suggest environmental barriers inhibiting colonization toward eastern continental zone of Europe.
Compared with R. raciborskii, R. mediterranea was recorded much less frequently (Figure 1A). We identified individuals in seven of 112 lakes, too small of a sample size to associate these occurrences with possible drivers. Still, we report three new occurrences of R. mediterranea in lakes located in the eastern part of the country (Figure 1A). In Poland, it occurred in different regions without any clear distribution pattern. Biomass and relative contribution of R. mediterranea to total phytoplankton biomass were also consistently low, ranging from 0.004 to 0.02 mg L−1 (Supplementary Table 1) and 0.10%–2.6%, respectively, aligning with prior reports that underscore limited range expansion (Wilk-Woźniak and Najberek, 2013). In Lithuania, the species was found in only one lake, diverging from the findings of previous years when relatively high biomass was observed in several lakes (Kasperovičienė et al., 2010). This observation shows that, akin to other non-native species such as Chrysosporum bergii, Sphaerospermopsis aphanizomenoides, and Cuspidothrix issatchenkoi, R. mediterranea is expanding toward the temperate zone. However, occurrence and contribution to total phytoplankton biomass vary notably across years.
We acknowledge that our sampling protocol may miss some of the species and thus some false absences may appear, but importantly, our sampling effort was the same for all lakes. Moreover, the sampling of large number (117) of lakes should alleviate the problems related to potential false absence. Furthermore, as temporal trends maybe important in plankton, we collected all samples in the middle of vegetative season to further reduce the potential false absences. To sum up, we think that our sampling design comprising large number of lakes is sufficient to provide reliable overview of the species regional distribution and biomass.
4.2 Environmental factors driving the abundance and occurrence of R. raciborskii and R. mediterranea
Among the possible factors influencing the abundance and occurrence of R. raciborskii, air temperature, TRP and TN were significant (Table 1). Specifically, air temperature exhibited a positive association with R. raciborskii biomass, while nutrient concentrations were related to occurrence. Notably, our findings contrast with many earlier studies emphasizing the important role played by water temperature, which impacts R. raciborskii growth and bloom formation (Soares et al., 2012; Recknagel et al., 2014).
The observed positive relationship between R. raciborskii biomass and air temperature aligns with a previous study in the same region (Kokociński et al., 2017), highlighting the pivotal role of climate on distribution. Our findings reveal a consistent longitudinal pattern of R. raciborskii distribution and biomass in Poland and Lithuania, extending from the west to the east. This pattern can be attributed to the influence of warmer air masses from the Atlantic Ocean, leading to higher temperatures in Western Poland, in contrast to Eastern Poland, where colder air masses from Eurasian landmass prevail (Błażejczyk, 2006). Furthermore, western lakes experience earlier warming in the spring and prolonged warmth in the autumn, resulting in an extended vegetative season compared to lakes in eastern Poland and Lithuania.
Regarding nutrients, we found a negative relationship between orthophosphates and the probability of R. raciborskii occurrence. This finding aligns with expectations, considering that R. raciborskii employs multiple strategies to thrive under diverse phosphorus conditions (Bonilla et al., 2012; Wu et al., 2022). Notably, it exhibits a particularly high affinity for dissolved inorganic phosphorus (Isvánovics et al., 2000; Wu et al., 2009, 2012) and possesses a substantial phosphorus storage capacity (Posselt et al., 2009; Willis et al., 2017, 2019). Due to this storage capacity, the pulsed addition of dissolved phosphorus is deemed more favorable for growth than constant phosphorus input (Posselt et al., 2009; Amaral et al., 2014). Collectively, these adaptive strategies allow R. raciborskii to successfully compete with native cyanobacteria such as Microcystis aeruginosa and Aphanizomenon flos-aquae (Wu et al., 2009). Our results provide further evidence that R. raciborskii has high capability to inhabit and sustain abundant populations in new lakes even under conditions of depleted dissolved phosphorus provided that other environmental conditions are suitable.
Our study also showed that R. raciborskii is more likely to be found in lakes with higher TN concentrations. This agrees with previous findings for the same region, where TN played a pivotal role in R. raciborskii distribution (Kokociński et al., 2017). While R. raciborskii can fix atmospheric nitrogen to thrive under low dissolved nitrogen conditions, it still exhibits preference for certain nitrogen forms (Burford et al., 2006; Ammar et al., 2014). For instance, ammonium nitrogen increased R. raciborskii growth rate, allowing it to outcompete native Planktothrix agardhii populations (Ammar et al., 2014). Nitrogen concentrations as low as 0.50 mg L−1, however, may limit R. raciborskii growth (Dai et al., 2015).
Note that we lack data for zooplankton in the study lakes. We acknowledge that the inclusion of zooplankton data (e.g., number of individuals) might have increased somewhat the explained variability by the species distribution model. We think, however, that this study based on abiotic variables only still contribute significantly to our understanding of R. raciborskii expansion.
R. mediterranea is considered a taxonomically close species or even a non-heterocytous form of R. raciborskii (Moustaka-Gouni et al., 2009). However, Aguilera et al. (2018) observed stable native populations of R. mediterranea without heterocytes and found no differentiation of heterocytes in isolated strains exposed to nitrogen starvation, suggesting that R. mediterranea is likely a distinct species. In this study, R. mediterranea was detected in seven water bodies in Poland, and both Raphidiopsis species co-occurred in five lakes. However, in most lakes, R. mediterranea biomass was 2–200 times lower than R. raciborskii biomass. The results also show that water transparency was lower and total and reactive phosphorus concentrations were higher in lakes where R. mediterranea or both species were present (mean Secchi depth 0.74 ± 0.55 m; mean TP 0.180 ± 0.138 mg L–1; mean TRP 0.063 ± 0.040 mg L–1) compared to lakes containing only R. raciborskii (mean Secchi depth 1.40 ± 1.54 m; mean TP 0.121 ± 0.105 mg L–1; mean TRP 0.040 ± 0.039 mg L–1). Similarly, Aguilera et al. (2019) demonstrated that water temperature, light availability and phosphorus are key factors influencing the proliferation of R. mediterranea isolates and competition with other cyanobacteria species. Previously, R. mediterranea was detected in six Lithuanian lakes (Kasperovičienė et al., 2005, 2010; Kasperovičienė, 2007) and correlated with water temperature. In 2020, however, R. mediterranea was not observed in the same lakes, although temperature ranges were similar to previous studies. The fragmentary occurrence of the species in both countries does not provide clear evidence of the species’ environmental optima.
In addition to nutrient concentrations and temperature, intraspecific diversity and naturally occurring ecotypes should be included among factors determining the distribution of R. raciborskii in eastern Europe. A recent study on the global occurrence of R. raciborskii ecotypes indicated a few major ecotypes with unclear distribution (Baxter et al., 2022). This may indirectly explain the distribution pattern we observed: the expansion of ecotypes that inhabit western regions with milder climate was hampered in eastern regions with more severe climate. Only some strains could establish and sustain small populations under these conditions.
5 Conclusion
Although our study showed increasing prevalence of R. raciborskii in the eastern regions of Poland, expansion into this region appears to be constrained by the more severe climate conditions. Although R. raciborskii occurred in several new lakes, biomass was similar across all lakes, the maximum contribution to total phytoplankton biomass in western regions of Poland was higher than observed in the 2016 study. Air temperature was again a significant factor determining biomass. As such, the contribution of R. raciborskii to total phytoplankton biomass was much lower in cooler eastern regions than warmer western regions with longer vegetation season. In addition to air temperature, nutrient concentrations played a major role as R. raciborskii occurred more frequently in lakes with low TRP and high TN concentrations than more eutrophic lakes. Catchment area land use, both agricultural and urban, had no direct effect on the occurrence or biomass of R. raciborskii. However, these catchment types should not be neglected when considering non-point sources of nutrients. In addition to environmental variables, the role of ecotypes at the edge of species geographical range should be considered when explaining range expansion. R. mediterranea was only detected in small number of lakes in Poland without a clear distributional pattern. In Lithuania, it was present in only one lake, compared to previous years when it was more widely distributed. In both countries, R. mediterranea biomass was low and therefore relationships with environmental factors could not be quantified. In sum, these results highlight the importance of climate variables for the distribution of invasive cyanobacteria.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary material, further inquiries can be directed to the corresponding author.
Author contributions
MK: Conceptualization, Investigation, Methodology, Supervision, Writing – original draft, Writing – review and editing. CG-R: Data curation, Visualization, Writing – review and editing. IJ: Investigation, Writing – review and editing. JKaro: Investigation, Visualization, Writing – review and editing. JKasp: Investigation, Writing – review and editing. JKobo: Investigation, Writing – review and editing. JKore: Investigation, Writing – review and editing. JM-B: Writing – review and editing. JS: Software, Writing – review and editing. AS: Investigation, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was partly funded by the National Science Centre, project no. UMO-2012/07/B/NZ8/03991 - “Application of reporter cell biosensors in ecotoxicology of cyanobacteria: new tar-gets for bioactivity.”
Acknowledgments
We would like to thank Łukasz Wejnerowski (Department of Hydrobiology, Adam Mickiewicz University) for his help during sampling campaign and Jennifer L. Graham from New York Water Science Center, United States (United States Geological Survey) for careful proofreading and editorial suggestions that improve English language. We also thank the associate editor and the reviewers for their useful feedback that improve this manuscript.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fmicb.2025.1533716/full#supplementary-material
References
Aguilera, A., Aubriot, L., Echenique, R. O., Donadelli, J. L., and Salerno, G. L. (2019). Raphidiopsis mediterranea (Nostocales) exhibits a flexible growth strategy under light and nutrient fluctuations in contrast to Planktothrix agardhii (Oscillatoriales). Hydrobiologia 839, 145–157. doi: 10.1007/s10750-019-04002-5
Aguilera, A., Gómez, E. B., Kăstovský, J., Echenique, R. O., and Salerno, G. L. (2018). The polyphasic analysis of two native Raphidiopsis isolates supports the unification of the genera Raphidiopsis and Cylindrospermopsis (Nostocales, Cyanobacteria). Phycologia 57, 130–146. doi: 10.2216/17-2.1
Amaral, V., Bonilla, S., and Aubriot, L. (2014). Growth optimization of the invasive cyanobacterium Cylindrospermopsis raciborskii in response to phosphate fluctuations. Eur. J. Phycol. 49, 134–141. doi: 10.1080/09670262.2014.897760
Ammar, M., Comte, K., Tran, T. D. C., and Bour, M. E. (2014). Initial growth phases of two bloom-forming cyanobacteria (Cylindrospermopsis raciborskii and Planktothrix agardhii) in monocultures and mixed cultures depending on light and nutrient conditions. Ann. Limnol. Int. J. Limol. 50, 231–240. doi: 10.1051/limn/2014096
Antunes, J. T., Leäo, P. N., and Vasconcelos, V. M. (2015). Cylindrospermopsis raciborskii: Review of the distribution, phylogeography, and ecophysiology of a global invasive species. Front. Microbiol. 6:473. doi: 10.3389/fmicb.2015.00473
APHA, AWWA, and WEF. (2012). Standard Methods for Examination of Water and Wastewater, 22nd Edn. Washington, DC: American Public Health Association, 1360.
Baxter, K., Jameson, I. D., and Willis, A. (2022). Towards defining global ecotypes of the toxic cyanobacterium Raphidiopsis raciborskii. Appl. Phycol. 3, 149–158. doi: 10.1080/26388081.2020.1740891
Błażejczyk, K. (2006). “Climate and bioclimate of Poland,” in Natural and Human Environment of Poland. A Geographical Overview, ed. M. Degórski (Warsaw: Polish Academy of Sciences), 31–48.
Bolius, S., Wiedner, C., and Weithoff, G. (2017). High local trait variability in a globally invasive cyanobacterium. Freshw. Biol. 62, 1879–1890. doi: 10.1111/fwb.13028
Bonilla, S., Aubriot, L., Soares, M. C. S., Gonzalez-Piana, M., Fabre, A., Huszar, V. L., et al. (2012). What drives the distribution of the bloom-forming cyanobacteria Planktothrix agardhii and Cylindrospermopsis raciborskii? FEMS Microbiol. Ecol. 79, 594–607. doi: 10.1111/j.1574-6941.2011.01242.x
Briand, J. F., Leboulanger, C., Humbert, J. F., Bernard, C., and Dufour, P. (2004). Cylindrospermopsis raciborskii (Cyanobacteria) invasion at mid-latitudes: Selection, wide physiological tolerance, or global warming? J. Phycol. 40, 231–238. doi: 10.1111/j.1529-8817.2004.03118.x
Burford, M. A., Beardall, J., Willis, A., Orr, P. T., Magalhaes, V. F., Rangel, L. M., et al. (2016). Understanding the winning strategies used by the bloom-forming cyanobacterium Cylindrospermopsis raciborskii. Harmful Algae 54, 44–53. doi: 10.1016/j.hal.2015.10.012
Burford, M. A., McNeale, K. L., and McKenzie-Smith, F. J. (2006). The role of nitrogen in promoting the toxic cyanophyte Cylindrospermopsis raciborskii in a subtropical water reservoir. Fresh. Biol. 51, 2143–2153. doi: 10.1111/j.1365-2427.2006.01630
Burford, M. A., Willis, A., Chuang, A., Man, X., and Orr, P. T. (2018). Recent insights into physiological responses to nutrients by the cylindrospermopsin producing cyanobacterium. Chin. J. Oceanol. Limnol. 36, 1032–1039. doi: 10.1007/s00343-018-7179-5
Carneiro, R. L., Santos, M. E. V., Pacheco, A. B. F., and Azevedo, S. M. F. O. (2009). Effects of light intensity and light quality on growth and circadian rhythm of saxitoxins production in Cylindrospermopsis raciborskii (Cyanobacteria). J. Plank. Res. 31, 481–488. doi: 10.1093/plankt/fbp006
Cronberg, G. (1973). Development and ecology of Raphidiopsis mediterranea Skuja in the Swedish lake Trummen. Svensk Botnici Tidskrift 67, 59–64.
Dai, J. J., Peng, L., Yu, T., and Lei, L. M. (2015). The effects of phosphorus and nitrogen on the growth of Cylindrospermopsis raciborskii N8 isolated from the Zhenhai reservoir. Acta Hydrobiol. Sin. 39, 533–539. doi: 10.7541/2015.70
de Jonge, V. N., and Elliott, M. (2001). “Eutrophication,” in Encyclopedia of Marine Sciences, eds J. Steele, S. Thorpe, and K. Turekian (London: Academic Press), 852–870.
de La Cruz, A. A., Hiskia, A., Kaloudis, H. A., Chernoff, N., Hill, D., and Antoniou, M. G. (2013). A review on cylindrospermopsin: The global occurrence, detection, toxicity and degradation of a potent cyanotoxin. Environ. Sci. Processes Impacts 15, 1979–2003. doi: 10.1039/c3em00353a
Dokulil, M. T. (2015). Vegetative survival of Cylindrospermopsis raciborskii (Cyanobacteria) at low temperature and low light. Hydrobiologia 764, 241–247. doi: 10.1007/s10750-015-2228-y
Falfushynska, H., Horyn, O., Brzozowska, A., Fedoruk, O., Buyak, B., and Poznansky, D. (2019). Is the presence of Central European strains of Raphidiopsis (Cylindrospermopsis) raciborskii a threat to a freshwater fish? An in vitro toxicological study in common carp cells. Aquat. Toxicol. 206, 105–113. doi: 10.1016/j.aquatox.2018.11.012
Fick, S. E., and Hijmans, R. J. (2017). Worldclim 2: New 1-km spatial resolution climate surfaces for global land areas. Int. J. Climatol. 37, 4302–4315. doi: 10.1002/joc.5086
Friedl, M., and Sulla-Menashe, D. (2022). MCD12Q1 MODIS/Terra+Aqua Land Cover Type Yearly L3 Global 500m SIN Grid V061. NASA EOSDIS Land Processes DAAC. doi: 10.5067/MODIS/MCD12Q1.061
Hötzel, G., and Croome, R. (1999). A Phytoplankton Methods Manual for Australian Waters, Land and Water Resources Research and Development Corporation Occasional Paper Series, 22/99. Wodonga: Land and Water Resources Research and Development Corporation.
Hupfer, M., and Hilt, S. (2008). “Lake restoration,” in Encyclopedia of Ecology, Vol. 3, eds S. E. Jørgensen and B. D. Fath (Oxford: Elsevier), 2080–2093.
Isvánovics, V., Shafik, H. M., Présing, M., and Juhos, S. (2000). Growth and phosphate uptake kinetics of the cyanobacterium, Cylindrospermopsis raciborskii (Cyanophyceae) in throughflow cultures. Freshw. Biol. 43, 257–275. doi: 10.1046/j.1365-2427.2000.00549
Jakubowska, N., Zagajewski, P., and Gołdyn, R. (2013). Water blooms and cyanobacterial toxins in lakes. Pol. J. Environ. Stud. 22, 1077–1082.
Jia, N., Wang, Y., Guan, Y., Chen, Y., Li, R., and Yu, G. (2021). Occurrence of Raphidiopsis raciborskii blooms in cool waters: Synergistic effects of nitrogen availability and ecotypes with adaptation to low temperature. Environ. Pollut. 270:116070. doi: 10.1016/j.envpol.2020.116070
Kasperovičienė, J. (2007). Assessment of Ecological status of the Dovinė river catchment area lakes according to phytoplankton data. Ekologia 53, 13–21.
Kasperovičienė, J., Koreivienė, J., and Karosienė, J. (2010). “Recent appearance of Raphidiopsis mediterranea (Cyanobacteria) in eutrophic Lithuanian lakes. – Biological invasions in a changing world from science to management,” in Proceedings of the 6th NEOBIOTA Conference, (Copenhagen), 14–17.
Kasperovičienė, J., Koreivienė, J., and Paškauskas, R. (2005). Cyanoprocariotes and microcystins dynamics in shallow hypertrophic lake (south-Eastern Lithuania). Oceanol. Hydrobiol. Stud. 34, 93–104.
Kaštovský, J., Hauer, T., Mareš, J., Krautová, M., Bašta, T., Komárek, J., et al. (2010). A review of the alien and expansive species of freshwater cyanobacteria and algae in the Czech Republic. Biol. Invasions 12, 3599–3625. doi: 10.1007/s10530-010-9754-3
Kehoe, M., O’Brien, K. R., Grinham, A., and Burford, M. A. (2015). Primary production of lake phytoplankton, dominated by the cyanobacterium Cylindrospermopsis raciborskii, in response to irradiance and temperature. Inland Waters 5, 93–100. doi: 10.5268/iw-5.2.778
Kokociński, M., Ga̧gała, I., Jasser, I., Karosienė, J., Kasperovičienė, J., Kobos, J., et al. (2017). Distribution of invasive Cylindrospermopsis raciborskii in the East-Central Europe is driven by climatic and local environmental variables. FEMS Microbiol. Ecol. 93:fix035. doi: 10.1093/femsec/fix035
Kosten, S., Huszar, V., Bècares, E., Costa, L., van Donk, E., Hansson, L. A., et al. (2011). Warmer climates boost cyanobacterial dominance in shallow lakes. Glob. Change. Biol. 18, 118–126. doi: 10.1111/j.1365-2486.2011.02488.x
Lorenzi, A. S., Silva, G. G. Z., Lopes, F. A. C., Chia, M. A., Edwards, R. A., and Bittencourt-Oliveira, M. C. (2016). Draft genome sequence of Cylindrospermopsis raciborskii (Cyanobacteria) strain ITEP-A1 isolated from a Brazilian semiarid freshwater body: Evidence of saxitoxin and cylindrospermopsin synthetase genes. Genome Announc. 4:e00228-16. doi: 10.1128/genomeA.00228-16
Lürling, M., van Oosterhout, F., and Faassen, E. (2017). Eutrophication and warming boost cyanobacterial biomass and microcystins. Toxins 9:64. doi: 10.3390/toxins9020064
Marinho, M. M., Souza, M. B. G., and Lürling, M. (2013). Light and phosphate competition between Cylindrospermopsis raciborskii and Microcystis aeruginosa is strain dependent. Microb. Ecol. 66, 479–488. doi: 10.1007/s00248-013-0232-1
McGregor, G. B., Sendall, B. C., Hunt, L. T., and Eaglesham, G. K. (2011). Report of the cyanotoxins cylindrospermopsin and deoxy-cylindrospermopsin from Raphidiopsis mediterranea Skuja (Cyanobacteria/Nostocales). Harmful Algae 10, 402–410. doi: 10.1016/j.hal.2011.02.002
Mehnert, G., Leunert, F., Cirés, S., Jöhnk, K., Rücker, J., Nixdorf, B., et al. (2010). Competitiveness of invasive and native cyanobacteria from temperate freshwaters under various light and temperature conditions. J. Plankton Res. 32, 1009–1021. doi: 10.1093/plankt/fbq033
Mohamed, Z. A. (2007). First report of toxic Cylindrospermopsis raciborskii and Raphidiopsis mediterranea (Cyanoprokaryota) in Egyptian fresh waters. FEMS Microbiol. Ecol. 59, 749–761. doi: 10.1111/j.1574-6941.2006.00226.x
Moustaka-Gouni, M., Kormas, K., Vardaka, E., Katsiapi, M., and Gkelis, S. (2009). Raphidiopsis mediterranea Skuja represents non-heterocytous life-cycle stages of Cylindrospermopsis raciborskii (Woloszynska) Seenayya et Subba Raju in Lake Kastoria (Greece), its type locality: Evidence by morphological and phylogenetic analysis. Harmful Algae 8, 864–872. doi: 10.1016/j.hal.2009.04.003
Namikoshi, M., Murakami, T., Watanabe, M. F., Oda, T., Yamada, J., Tsujimura, S., et al. (2003). Simultaneous production of homoanatoxin-a, anatoxin-a, and a new non-toxic 4-hydroxyhomoanatoxin-a by the cyanobacterium Raphidiopsis mediterranea Skuja. Toxicon 42, 533–538. doi: 10.1016/S0041-0101(03)00233-2
Olenina, I., Hajdu, S., Edler, L., Andersson, A., Wasmund, N., and Busch, S. (2006). Biovolumes and size-classes of phytoplankton in the Baltic Sea. HELCOM Balt. Sea Environ. Proc. 106:144.
Padisák, J. (1997). Cylindrospermopsis raciborskii (Woloszynska) Seenayya et Subba Raju, an expanding, highly adaptive cyanobacterium: Worldwide distribution and review of its ecology. Arch. Hydrobiol. 107, 563–593.
Paerl, H. W., and Huisman, J. (2009). Climate change: A catalyst for global expansion of harmful cyanobacterial blooms. Environ. Microbiol. Rep. 1, 27–37. doi: 10.1016/j.scitotenv.2017.08.149
Panou, M., Zervou, S. K., Kaloudis, T., Hiskia, A., and Gkelis, S. (2018). A Greek Cylindrospermopsis raciborskii strain: Missing link in tropic invader’s phylogeography tale. Harmful Algae 80, 96–106. doi: 10.1016/j.hal.2018.10.002
Pierangelini, M., Sinha, R., Willis, A., Burford, M. A., Orr, P. T., and Beardall, J. (2015). Constitutive cylindrospermopsin cellular pool size in Cylindrospermopsis raciborskii under different light and pCO2 conditions. Appl. Environ. Microbiol. 81, 3069–3076. doi: 10.1128/AEM.03556-14
Pinheiro, J. C., and Bates, D. M. (2000). Mixed-Effects Models in S and S-PLUS. Springer: New York, doi: 10.1007/b98882
Pinheiro, J., and Bates, D. (2022). R Core Team. Nlme: Linear and Nonlinear Mixed Effects Models. R package version 3.1-157. Available online at: https://CRAN.R-project.org/package=nlme>
Plominsky, ÁM., Larsson, J., Bergman, B., Delherbe, N., Osses, I., and Vásquez, M. (2013). Dinitrogen fixation is restricted to the terminal heterocysts in the invasive cyanobacterium Cylindrospermopsis raciborskii CS-505. PLoS One 8:e51682. doi: 10.1371/journal.pone.0051682
Poniedziałek, B., Rzymski, P., Kokociński, M., and Karczewski, J. (2015). Toxic potencies of metabolite(s) of non-cylindrospermopsin producing Cylindrospermopsis raciborskii isolated from temperate zone in human white cells. Chemosphere 120, 608–614. doi: 10.1016/j.chemosphere.2014.09.067
Posselt, A. J., Burford, M. A., and Shaw, G. (2009). Pulses of phosphate promote dominance of the toxic cyanophyte Cylindrospermopsis raciborskii in a subtropical water reservoir. J. Phycol. 45, 540–546. doi: 10.1111/j.1529-8817.2009.00675.x
R Core Team. (2022). R: A Language and Environment for Statistical Computing. Vienna: R Foundation for Statistical Computing.
Recknagel, F., Orr, F. T., and Cao, H. (2014). Inductive reasoning and forecasting of population dynamics of Cylindrospermopsis raciborskii in three sub-tropical reservoirs by evolutionary computation. Harmful Algae 31, 26–34. doi: 10.1016/j.hal.2013.09.004
Rücker, J., Tingwey, E. I., Wiedner, C., Anu, C. M., and Nixdorf, B. (2009). Impact of the inoculum size on the population of Nostocales cyanobacteria in a temperate lake. J. Plankton Res. 31, 1151–1159. doi: 10.1093/plankt/fbp067
Rzymski, P., Brygider, A., and Kokociński, M. (2017a). On the occurrence and toxicity of Cylindrospermopsis raciborskii in Poland. Limnol. Rev. 17, 23–29. doi: 10.1515/limre-2017-0003
Rzymski, P., Poniedziałek, B., Mankiewicz-Boczek, J., Faassen, E. J., Jurczak, T., and Ga̧gała-Borowska, I. (2017b). Polyphasic toxicological screening of Cylindrospermopsis raciborskii and Aphanizomenon gracile isolated in Poland. Algal Res. 24, 72–80. doi: 10.1016/j.algal.2017.02.011
Sidelev, S., Koksharova, O., Babanazarova, O., Fastner, J., Chernova, E., and Gusev, E. (2020). Phylogeographic, toxicological and ecological evidence for the global distribution of Raphidiopsis raciborskii and its northernmost presence in Lake Nero, Central Western Russia. Harmful Algae 98:101889. doi: 10.1016/j.hal.2020.101889
Sinha, R., Pearson, L. A., Davis, T. W., Burford, M. A., Orr, P. T., and Neilan, B. (2012). Increased incidence of Cylindrospermopsis raciborskii in temperate zones — is climate change responsible? Water. Res. 46, 1408–1419. doi: 10.1016/j.watres.2011.12.019
Soares, M. C. S., Lürling, M., and Huszar, V. L. M. (2012). Growth and temperature related phenotypic plasticity in the cyanobacterium Cylindrospermopsis raciborskii. Phycol. Res. 61, 61–67. doi: 10.1111/pre.12001
Sukenik, A., Hadas, O., Kaplan, A., and Quesada, A. (2012). Invasion of Nostocales (cyanobacteria) to subtropical and temperate freshwater lakes - physiological, regional, and global driving forces. Front. Microbiol. 3:86. doi: 10.3389/fmicb.2012.00086
Thomas, M. K., and Litchman, E. (2016). Effects of temperature and nitrogen availability on the growth of invasive and native cyanobacteria. Hydrobiologia 763, 357–369. doi: 10.1007/s10750-015-2390-2
Watanabe, M. F., Tsujimura, S., Oishi, S., Niki, T., and Namikoshi, M. (2003). Isolation and identification of homoanatoxin-A from a toxic strain of the cyanobacterium Raphidiopsis mediterranea Skuja isolated from Lake Biwa, Japan. Phycologia 42, 364–369. doi: 10.2216/i0031-8884-42-4-364.1
Wickham, H., Averick, M., Bryan, J., Chang, W., McGowan, L. D. A., François, R., et al. (2019). Welcome to the Tidyverse. J. Open Source Softw. 4:1686. doi: 10.21105/joss.01686
Wiedner, C., Rücker, J., Brüggemann, R., and Nixdorf, B. (2007). Climate change affects timing and size of populations of an invasive cyanobacterium in temperate regions. Oecologia 152, 473–484. doi: 10.1007/s00442-007-0683-5
Wilk-Woźniak, E., and Najberek, K. (2013). Towards clarifying the presence of alien algae in inland waters – Can we predict places of their occurrence? Biologia 68, 838–844. doi: 10.2478/s11756-013-0221-3
Wilk-Woźniak, Solarz, W., Najberek, K., and Pociecha, A. (2016). Alien cyanobacteria: An unsolved part of the “expansion and evolution” jigsaw puzzle? Hydrobiologia 764, 65–79. doi: 10.1007/s10750-015-2395-x
Willis, A., Adams, M. P., Chuang, A. W., and Orr, P. T. (2015). Constitutive toxin production under various nitrogen and phosphorus regimes of three ecotypes of Cylindrospermopsis raciborskii (Wołoszynska) Seenayya et Subba Raju. Harmful Algae 47, 27–34. doi: 10.1016/j.hal.2015.05.011
Willis, A., Chuang, A., Dyhrman, S., and Burford, M. A. (2019). Differential expression of phosphorus acquisition genes in response to phosphorus stress in two Raphidiopsis raciborskii strains. Harmful Algae 82, 19–25. doi: 10.1016/j.hal.2018.12.003
Willis, A., Ind, A., and Burford, M. A. (2017). Variations in carbon-to-phosphorus ratios of two Australian strains of Cylindrospermopsis raciborskii. Eur. J. Phycol. 52, 1–8. doi: 10.1080/09670262.2017.1286524
Wu, Z. X., Shi, J. Q., and Li, R. H. (2009). Comparative studies on photosynthesis and phosphate metabolism of Cylindrospermopsis raciborskii with Microcystis aeruginosa and Aphanizomenon flos-aquae. Harmful Algae 8, 910–915. doi: 10.1016/j.hal.2009.05.002
Wu, Z. X., Zeng, B., Li, R. H., and Song, L. (2012). Physiological regulation of Cylindrospermopsis raciborskii (Nostocales, Cyanobacteria) in response to inorganic phosphorus limitation. Harmful Algae 15, 53–58. doi: 10.1016/j.hal.2011.11.005
Wu, Z., Yang, S., and Shi, J. (2022). Overview of the distribution and adaptation of a bloomforming cyanobacterium Raphidiopsis raciborskii: Integrating genomics, toxicity, and ecophysiology. J. Oceanol. Limnol. 40, 1774–1791. doi: 10.1007/s00343-022-2003-7
Xiao, M., Willis, A., and Burford, M. A. (2017). Differences in cyanobacterial strain responses to light and temperature reflect species plasticity. Harmful Algae 62, 84–93. doi: 10.1016/j.hal.2016.12.008
Zheng, B., He, S., Zhao, L., Li, J., Du, Y., Li, Y., et al. (2023). Does temperature favour the spread of Raphidiopsis raciborskii, an invasive bloom-forming cyanobacterium, by altering cellular trade-offs? Harmful Algae 124:02406. doi: 10.1016/j.hal.2023.102406
Keywords: cyanobacteria expansion, freshwater lakes, nutrients, non-native cyanobacteria, temperature
Citation: Kokociński M, Graco-Roza C, Jasser I, Karosienė J, Kasperovičienė J, Kobos J, Koreivienė J, Mankiewicz-Boczek J, Soininen J and Szczurowska A (2025) Environmental factors determining the distribution patterns of invasive Raphidiopsis raciborskii and R. mediterranea in central east Europe. Front. Microbiol. 16:1533716. doi: 10.3389/fmicb.2025.1533716
Received: 24 November 2024; Accepted: 14 April 2025;
Published: 07 May 2025.
Edited by:
Jin Zhou, Tsinghua University, ChinaReviewed by:
Abdiel Jover Capote, Universidad de Oriente, CubaMatina Katsiapi, Aristotle University of Thessaloniki, Greece
Copyright © 2025 Kokociński, Graco-Roza, Jasser, Karosienė, Kasperovičienė, Kobos, Koreivienė, Mankiewicz-Boczek, Soininen and Szczurowska. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mikołaj Kokociński, a29rQGFtdS5lZHUucGw=