 Yu Yu
Yu Yu Jing Li
Jing Li Qingqing Liu
Qingqing Liu Rongrong Zhai
Rongrong Zhai Yuzhu Dai
Yuzhu Dai Lei Sun
Lei Sun- 1Department of Clinical Laboratory, Lu'an People's Hospital, Lu'an, China
- 2Department of Clinical Laboratory, Fuyang Second People's Hospital, Fuyang, China
- 3Department of Clinical Research, The 903rd Hospital of PLA, Hangzhou, China
- 4Department of Clinical Laboratory, Qilu Hospital of Shandong University, Jinan, China
Severe fever with thrombocytopenia syndrome (SFTS) is an acute infectious disease caused by the SFTS virus (SFTSV). Since the first reported case, SFTSV has spread globally, particularly in Asian regions such as China, South Korea, and Japan, with an increasing number of cases and a high mortality rate among severe patients. SFTSV is an RNA virus capable of rapid biological evolution through genetic mutations, reassortment, and homologous recombination. The disease primarily occurs in mountainous, forested, and hilly areas. Due to limited clinical research, the clinical characteristics and pathogenesis of SFTS remain incompletely understood. This review summarizes recent advances in the regional epidemiological characteristics, clinical features, genotyping, pathogenesis, and rapid detection methods of SFTSV.
1 Introduction
Severe fever with thrombocytopenia syndrome (SFTS) is an acute infectious disease characterized by fever, thrombocytopenia, and leukopenia (Seo et al., 2021). The virus isolated from the serum of SFTS patients, initially named Huaiyangshan virus, was later reclassified as Dabie bandavirus (DBV) in 2019, belonging to the Phenuiviridae family and Bandavirus genus (Niu et al., 2024). SFTSV is primarily transmitted through tick bites but can also spread via contact with infected blood or bodily fluids (Kim et al., 2024; Kim and Park, 2023). Although the International Committee on Taxonomy of Viruses (ICTV) has reclassified the virus, the terms “SFTSV” and “SFTS” remain widely used. For consistency with previous studies, this review will use “SFTSV” and “SFTS” to refer to the virus and the disease, respectively.
SFTSV is an enveloped virus with a diameter of 80–120 nm, containing three single-stranded negative-sense RNA segments: large (L), medium (M), and small (S), with lengths of 6,368 bp, 3,378 bp, and 1746 bp (Wang et al., 2022; Liu et al., 2016). The complementary ends of the genome form a circular structure. The L segment encodes the RNA-dependent RNA polymerase (RdRp), the M segment encodes a membrane protein precursor that is cleaved into Gn and Gc proteins, and the S segment is a bicistronic RNA encoding the non-structural protein (NSs) and the nucleocapsid protein (NP) (Williams et al., 2023). The SFTSV Gn and Gc exist as a heterodimer on the surface of viral particles, and further assemble into pentameric and hexameric peplomers with the dimer as the structural unit, thus constituting a virus particle similar to an icosahedron (Du et al., 2023). This indicates that adjacent Gn/Gc dimers form a closely packed structure rather than a simple dimeric form. After SFTSV infection, viral particles bind to receptors on the surface of host cells through their membrane glycoproteins Gn and Gc. Studies have shown that dendritic cell-specific intercellular adhesion molecule-3-grabbing non-integrin (DC-SIGN) and Nonmuscle Myosin Heavy Chain IIA (NMMHCIIA) promote viral adsorption by recognizing the glycosylation sites of Gn (Spiegel et al., 2016; Yuan and Zheng, 2017). After SFTSV binds to receptors on the surface of host cells, it enters the host cells through a clathrin-dependent endocytic pathway (Liu et al., 2019). In a low pH environment, the Gc protein undergoes a conformational change, exposing the fusion loop. The fusion loop of Gc inserts into the endosomal membrane of the host cell, promoting the fusion of the viral envelope with the endosomal membrane (Wu et al., 2017). After membrane fusion is completed, the viral genomic RNA is released into the cytoplasm of the host cell, and NP and RdRp work together to initiate the replication and transcription of the viral genome. The above processes indicate that the glycoproteins Gn and Gc play a major role in viral replication. They are also important targets for specific neutralizing antibodies (Chang et al., 2024). In addition, during the process of viral entry into host cells, Gc mediates the fusion of the viral envelope with the endosomal membrane. In this process, some Gc subunits may dissociate from the dimeric structure and participate in processes such as membrane fusion in an independent form. Under low pH conditions, Gc may exist in the form of independent subunits to form trimers, promoting the fusion of the virus with the host cell membrane (Yuan and Zheng, 2017). NP and RdRp form ribonucleoprotein complexes (RNPs) that protect the virus from degradation by nucleases and the host immune system. NP and NSs play important roles in evading host immune responses and promoting viral replication. NP can inhibit the RIG-I/MDA5 pathway to block IFN production (Min et al., 2020). SFTSV NSs are potent IFN antagonists, which exert inhibitory effects on IFN by binding to several host molecules and sequestering them into inclusion bodies (IBs) (Ren et al., 2021).
Since its first identification, SFTSV has spread widely, particularly in China, South Korea, and Japan, posing a significant public health threat (Li et al., 2021). The transmission dynamics and pathogenesis of SFTSV are not fully understood (Casel et al., 2021). What is certain is that the segmented nature of the SFTSV genome allows for homologous recombination and reassortment between different genotypes during viral replication, which enhances viral genetic diversity and leads to the emergence of new viral strains, thereby facilitating rapid viral spread (Kwon et al., 2024). Furthermore, previous studies have shown that different SFTSV genotypes exhibit significant differences in pathogenicity and case fatality rates. Therefore, unified and accurate genotyping of SFTSV holds important practical significance for the selection of clinical treatment approaches and the implementation of public health interventions (Dai et al., 2022). This review will summarize the regional epidemiological characteristics, clinical features, genotyping, pathogenesis, and rapid detection methods of SFTSV.
2 Regional distribution of SFTS
SFTSV is transmitted through tick bites. Haemaphysalis longicornis is widely recognized as the primary vector, followed by Haemaphysalis flava, Rhipicephalus microplus, Amblyomma testudinarium, Dermacentor nuttalli, Hyalomma asiaticum, and Ixodes nipponensis (Casel et al., 2021; Zhu et al., 2019). Ticks have a broad host range, and SFTSV is thought to circulate in a tick-animal-tick transmission cycle. Currently, SFTSV RNA or anti-SFTSV antibodies have been detected in wild animals such as hedgehogs, rodents, and some bird species, as well as domestic animals like cattle, sheep, and pigs (Chen et al., 2019; Huang et al., 2019; Zhao et al., 2022). This indicates a high zoonotic transmission potential of SFTSV. Additionally, studies have shown that exposure to body fluids and secretions of infected patients can lead to SFTSV infection, suggesting human-to-human transmission. SFTSV infections predominantly occur from spring to autumn, with higher incidence rates in people living in mountainous, forested, and hilly areas or working outdoors—consistent with the main habitats of ticks (Peng et al., 2025). In high-altitude areas (averaging over 4,000 m), the spread of SFTSV is restricted due to the reduced geographical distribution of ticks and low population density, which is consistent with the results of SFTS case distribution shown in Figure 1. Tick growth and reproduction are closely associated with climatic factors, including light, humidity, and temperature (Pérez et al., 2024). For example, H. hystricis prefers warm and humid environments, while H. longicornis exhibits stronger environmental adaptability, widely distributed in rural landscapes and urban areas (Miao et al., 2020). Changes in environmental factors, particularly climatic-ecological and geographical landscape factors, may provide suitable ecological conditions for natural tick population growth, contributing to the seasonal variation characteristics of SFTSV infections. Furthermore, host animals carrying ticks expand their survival range through natural migration, further accelerating cross-regional transmission of SFTSV (Ji et al., 2024).
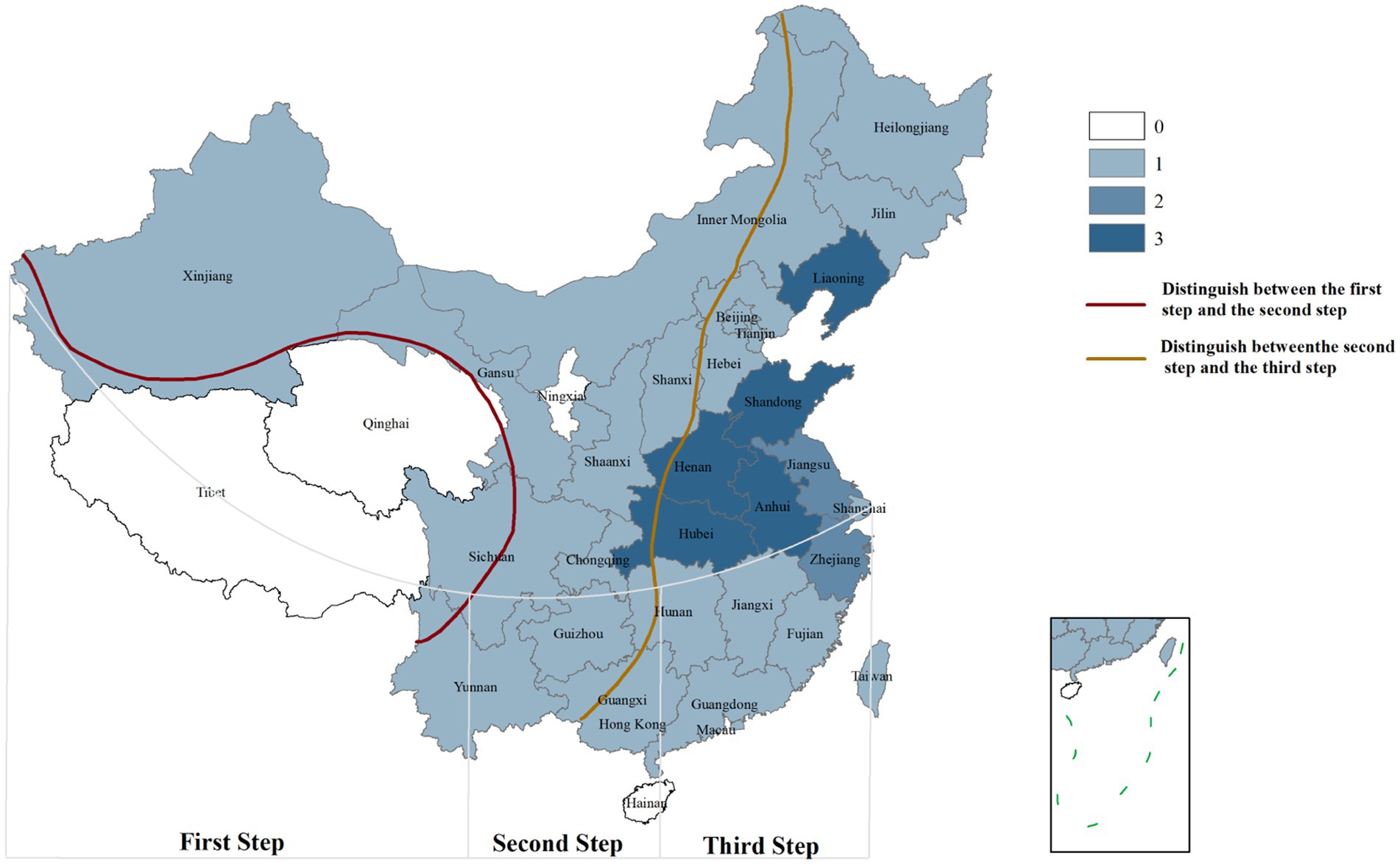
Figure 1. Geographical distribution of SFTS cases in China. 0: Area with 0 cases of infection; 1: Area with 1–100 cases of infection; 2: Area with 101–1,000 cases of infection; 3: Area with>1,000 cases of infection. First Step: Average altitude above 4,000 m; Second Step: Average altitude above 1,000-2000 m; Third Step: Average altitude below 500 m [The data comes from the study of Chen et al. (2022), created by Arcmap V10.2.2].
The reported cases of SFTS are mainly concentrated in East Asia. SFTS was first reported in central China between March and July 2009, with subsequent cases reported across various provinces. Japan reported its first case in 2012, primarily in eastern and southern regions, while South Korea reported its first fatal case in 2013, although infections likely occurred earlier (Liu et al., 2014). Most cases are concentrated in China, South Korea, and Japan, with sporadic reports from other regions. Retrospective analyses of serum samples from patients with acute febrile illnesses in Southeast Asian countries such as Vietnam and Thailand have detected SFTSV (Tran et al., 2019; Rattanakomol et al., 2022). SFTSV has also been detected in Rhipicephalus microplus and domestic poultry in Taiwan, China (Lin et al., 2020). Additionally, in the United States, cases infected with Heartland virus recorded in 2009 showed disease progression similar to SFTSV infection. Heartland virus and SFTSV are both in the Bandavirus genus within the Phenuiviridae family, Bunyavirale. Field sampling and laboratory work identified Amblyomma americanum as the vector of Heartland virus (Staples et al., 2020; Godsey et al., 2016).
In China, the national reported incidence of SFTS has shown an upward trend. As of 2023, the cumulative number of reported cases and deaths nationwide has reached 27,447 and 1,326 respectively, affecting 27 provinces. The average case-fatality rate is 4.83% (Yue et al., 2024; Chen et al., 2022; Huang et al., 2024). SFTSV cases are predominantly clustered in mountainous, forested, and hilly areas of central China, although reports from other regions are increasing, raising concerns about potential public health crises (Liang et al., 2023; Huang et al., 2021). In Japan, epidemiological surveys from 2013 to 2017 identified 310 cases, with an average mortality rate of 7.8%, and 60 to 100 new cases are added annually (Saijo, 2022; Crump and Tanimoto, 2020; Kobayashi et al., 2020). In South Korea, 1,203 cases and 231 deaths were reported by August 2020 (Jang et al., 2024). The reporting rate of SFTS in South Korea is on the rise, with a fatality rate of 18.54% (Cui et al., 2024). The geographical distribution of SFTS cases in China is illustrated in Figure 1. Additionally, seroprevalence studies have revealed significant regional variations. A meta-analysis of SFTSV antibody prevalence in China reported an overall seroprevalence of 4.3%, indicating widespread transmission and the presence of unreported mild or asymptomatic infections (Li et al., 2017). In contrast, the seroprevalence in Japan is much lower, ranging from 0.14 to 0.3% (Maslow et al., 2019). A seroscreening trial for SFTSV conducted in rural areas of South Korea by Han et al showed an SFTSV IgG antibody seroprevalence rate of 4.1%, slightly lower than that in China (Han et al., 2020). These differences may reflect variations in regional transmission patterns, such as the predominance of different genotypes or differences in antibody detection methods.
3 Clinical features of SFTS
The initial clinical symptoms of SFTSV infection are persistent high fever and respiratory or gastrointestinal symptoms, followed by a gradual decrease in platelets and white blood cells (Li et al., 2021). These symptoms are similar to those of other infectious fevers caused by different pathogens, lacking specificity, and often lead to inappropriate treatment, resulting in disease progression to severe cases and even death. SFTS cases mainly distribute in the population aged 35–80. In SFTS patients, the incidence increases with age, and most fatal cases occur in patients over 50 years old, suggesting that age is a risk factor related to both incidence and mortality (Casel et al., 2021). Asymptomatic carriers also exist in SFTSV infections. DU et al found asymptomatic SFTSV infections in serological surveys of healthy populations, which may have an important impact on the dynamics of SFTS outbreaks (Du et al., 2019). Currently, the research on the transmission capacity of asymptomatic carriers is insufficient. Asymptomatic carriers may provide the possibility for the continuous existence of SFTSV in the population and play a non-negligible role in the SFTSV transmission chain. Future work should focus on studying the potential factors for the development of asymptomatic SFTSV infections and preventing transfusion safety to avoid the spread of SFTS epidemics by asymptomatic cases (Zeng et al., 2015). The typical course of SFTSV infection is generally divided into three stages: the fever stage, the multiple-organ dysfunction (MOD) stage, and the convalescence stage (Yuan and Zheng, 2017). The fever stage can last for 5–11 days, during which patients present with influenza-like symptoms, including persistent high fever (body temperature 38–41°C), myalgia, and gastrointestinal symptoms such as anorexia, vomiting, and diarrhea, accompanied by thrombocytopenia (<100.0 × 109/L), leukopenia (<4.0 × 109/L), and lymphadenopathy. The high viral load during the fever stage is an important means for clinical confirmation. Patients typically progress from fever to MOD within 3–5 days. The progression of MOD is rapid, initially affecting the liver and heart, followed by the lungs and kidneys. Rapid elevations in clinical biochemical markers, including serum alanine aminotransferase (>40 IU/L), aspartate aminotransferase (>40 IU/L), creatine kinase (>200 IU/L), creatine kinase isoenzyme (>25 IU/L), lactate dehydrogenase (>245 IU/L), and activated partial thromboplastin time (>43.5 s), indicate liver and kidney dysfunction, myocardial damage, and coagulation disorders (Zhang et al., 2024). Patients with a gradual decline in serum viral load or self-limiting infections enter the recovery stage, with approximately 85% of patients having a good prognosis and their biochemical indicators returning to normal within 3–4 weeks (Zhang et al., 2024). In contrast, clinical laboratory indices continue to rise, including viral load (>1 × 106 copies/mL), activated partial thromboplastin time (>62.6 s), aspartate aminotransferase (>288 IU/L), etc. Elderly patients with underlying diseases who develop neurological symptoms, disseminated intravascular coagulation (DIC), and multiple organ failure (MOF) are more likely to die (Liang et al., 2024).
Additionally, previous studies have reported atypical and special cases of SFTS infection. Yun et al. (2019) compared the chest radiographs and CT scans of SFTS patients and scrub typhus patients, showing that SFTS patients mainly presented with cardiac enlargement, with or without pericardial effusion and patchy consolidation with ground-glass opacity (GGO), while scrub typhus presented with interstitial pneumonia on chest radiographs, which helps in early differentiation between SFTS and scrub typhus. Although most cases present with leukopenia, occasional leukocytosis has been observed in SFTS patients, possibly due to secondary infections. Lee et al. (2024) reported a case of thrombocytopenia with leukocytosis. PCR and antibody titer tests confirmed SFTS, while blood culture results indicated an Escherichia coli infection, suggesting that the patient had SFTS complicated by E. coli bacteremia. SFTS complicated by encephalitis may be due to the presence of SFTSV in cerebrospinal fluid, with patients presenting with headache and epilepsy and other central nervous system symptoms. Although these central nervous system symptoms have only been reported in a few cases, they are believed to be related to disease severity and death. However, the mechanism by which SFTSV causes central nervous system symptoms remains to be further investigated.
4 Pathogenesis of SFTS
The pathogenesis of SFTS is not yet fully understood. A common pathogenic feature of bunyaviruses is their ability to inhibit the host immune response, facilitating rapid viral replication. As an antiviral cytokine, IFN induces multiple antiviral responses to inhibit viral replication (Schneider et al., 2014). The IFN pathway comprises two stages: IFN induction and signal transduction. IFN induction detects viruses through pattern recognition receptors (PRRs) identifying pathogen-associated molecular patterns (PAMPs), while IFN signal transduction is activated by secreted IFN binding to relevant receptors expressed on adjacent cells, leading to antiviral protein expression (Lee and Ashkar, 2018). SFTSV interferes with IFN-I production via multiple mechanisms. During the IFN induction stage, SFTSV NSs protein specifically traps tripartite motif-containing protein 25 (TRIM25) into inclusion bodies (IBs), hinders TRIM25-mediated Lys-63 ubiquitination and RIG-I activation, and suppresses the production of interferon-stimulated genes (ISGs) (Min et al., 2020; Liu et al., 2019). Studies also show that the C-terminal of SFTSV NS protein specifically binds to TANK-binding kinase 1 (TBK1) to form IBs. These IBs not only serve as viral replication sites but also sequester key proteins in the IFN signaling pathway, such as TBK1, NF-κB kinase inhibitor (IKK), and IFN regulatory factor-3 (IRF-3), thereby blocking IFN-I production and promoting viral replication (Khalil et al., 2021; Chen et al., 2017). Additionally, SFTSV NSs protein captures mitochondrial antiviral signaling protein (MAVS) into IBs, disrupting IFN signal transduction and inhibiting NF-κB signaling pathway activation (Wuerth and Weber, 2016).
During the IFN signal transduction stage, SFTSV NS protein suppresses IFN signaling and ISG expression by sequestering STAT2 into IBs and impairing STAT2 heterodimer phosphorylation and nuclear translocation (Kitagawa et al., 2018). It also inhibits exogenous IFN-α-induced Jak/STAT signaling by suppressing STAT1 phosphorylation and activation, thereby blocking type I and III IFN signaling (Chen et al., 2017). Further research indicates that SFTSV NS protein hijacks STAT1 into viral IBs and reduces its expression, inhibiting type II IFN responses (Chen et al., 2023). These evidences indicate that SFTSV NSs is an effective IFN antagonist, which exerts inhibitory effects on IFN by binding to several host molecules and sequestering them into IBs, such as RIG-I, TBK1, IKK, IRF3, TRIM25, STAT1 and STAT2.
SFTSV evades immunity by influencing immune responses of immune cells through diverse mechanisms. The main target organs of SFTSV include the spleen, lymph nodes, liver, and bone marrow, while the lungs, kidneys, and heart can also be affected. Macrophages in the spleen and liver are likely the primary target cells for SFTSV infection (Xu et al., 2021) (Figure 2). SFTSV activates STAT1 to induce monocyte immune responses and stimulate macrophage differentiation into the M1 phenotype, leading to inflammatory cytokine production. However, post-infection, it suppresses M1 macrophage differentiation and drives macrophages toward the M2 phenotype, promoting viral shedding and transmission (Zhang et al., 2019). Natural killer (NK) cells control viral load by releasing perforin, granzyme, proinflammatory cytokines, and chemokines to induce host immune responses (Jost and Altfeld, 2013). Studies show that CD3−CD16+56+ NK cells significantly decrease in the early stage of SFTSV infection, indicating their involvement in early immune responses. Depletion of NK cells may contribute to disease progression (Li et al., 2020).
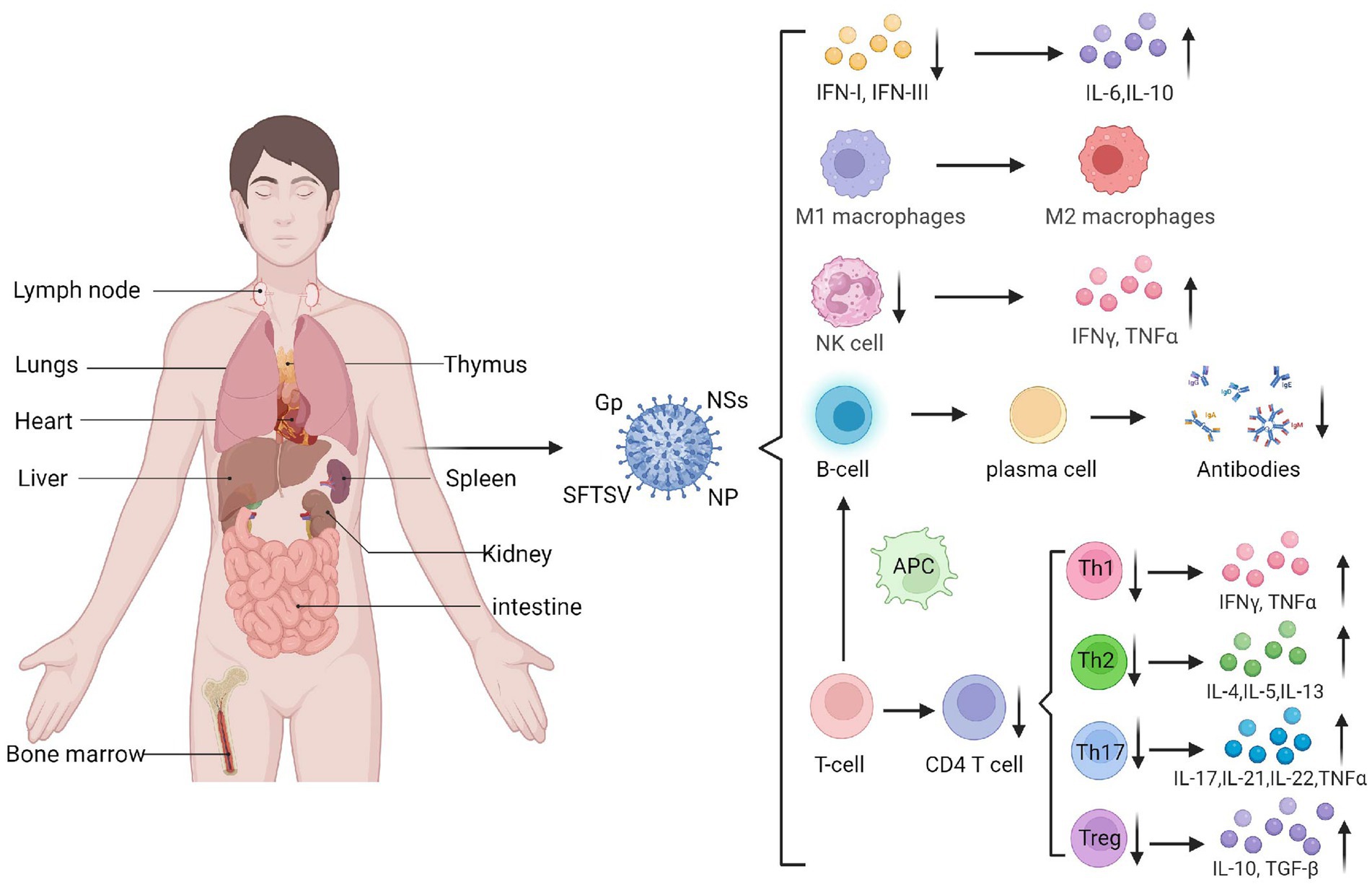
Figure 2. The target organs of SFTSV infection and its impact on immune cells. SFTSV infection impairs the production and function of various immune cells and cytokines, such as IFN, macrophages, and NK cells. It can also suppress the secretion and maturation of T cells and B cells, mediating the occurrence of a cytokine storm (Created by biorender.com).
Adaptive immune responses to the virus involve T and B cells, which specifically recognize and eliminate viral pathogens. T lymphocytes are the main cells mediating cellular immunity. Li et al. (2018) found that the number of T lymphocytes, especially CD4+ T cells, significantly decreases in SFTS patients, with the reduction magnitude positively correlated with disease severity. Further analysis of naive CD4+ T cell subsets showed that the counts of helper T (Th)1, Th2, and regulatory T (Treg) cells in SFTSV fatal cases were significantly lower than those in surviving patients. Increases in Th2 and Th17 cells within CD4+ T cell populations lead to abnormal Th1/Th2 and Th17/Treg ratios, which correlate positively with disease severity. Elevated Th17 cells can suppress effector T cell-mediated killing of target cells, hindering host antiviral responses. Additionally, research (Hou et al., 2014) shows that Th17 cells upregulate anti-apoptotic molecules to promote persistent viral replication in SFTSV-infected cells. Thus, SFTSV evades immunity by suppressing T cell responses and regulating infected cell apoptosis.
B cell responses are regulated by antigen-presenting cells and Tfh cells. However, early infection-induced apoptosis reduces dendritic cell (DC)-mediated antigen presentation, impairing Tfh differentiation and function, which significantly weakens humoral immunity. Studies indicate that peripheral blood mononuclear cells in SFTS patients contain transient plasmablasts, and in vitro induction shows these atypical lymphocytes are activated B cells, suggesting that SFTSV-infected B cells release factors driving B cell differentiation into plasmablasts (Wada et al., 2022). Most SFTSV in lethal infections resides in plasma B cells, and these activated/differentiated B cells do not express IgM or IgG, leading to insufficient humoral responses in fatal cases. Collectively, these findings show that SFTSV infection suppresses antibody secretion and B cell maturation, thereby evading effective humoral immunity against viral escape from the host immune system (Figure 2).
The cytokine storm induced by SFTSV infection is also believed to play an important role in worsening the condition of SFTS patients. When the host immune response fails to suppress viral replication, SFTSV can induce the release of large amounts of cytokines from target cells, leading to pathological lesions. In the cytokine storm, abnormal expression of several inflammatory cytokines, including IL-6, IL-8, and IL-10, is associated with the severity of SFTS (Song et al., 2024). He et al. (2021) found that monocyte chemoattractant protein-1 (MCP-1), macrophage inflammatory protein-1α (MIP-1α), and transforming growth factor-β1 (TGF-β1) were significantly elevated in severe SFTS patients compared to asymptomatic SFTS patients. These cytokines induce abnormal inflammatory disorders, exacerbating multi-organ damage in the body. The low expression of platelet-derived growth factor and secretory factors stored in platelets may be related to thrombocytopenia in peripheral blood, and these cytokines return to normal ranges when patients enter the recovery stage (Li et al., 2021). The bleeding tendency in SFTS patients is also related to elevated tumor necrosis factor (TNF-α). TNF-α acts on endothelial cells, inducing vasodilatory substances, stimulating nitric oxide synthase, increasing capillary endothelial permeability, affecting coagulation function in patients, and thereby increasing the risk of DIC (Xing et al., 2018). Cytokine secretion can also be regulated by multiple cellular signaling pathways, such as JAK/STAT3, MAPK, NF-κB, mTOR, and TLR4 signaling pathways. The crosstalk between these different signals can modulate cytokine expression. These previous findings indicate that cytokine/chemokine-mediated inflammatory responses, characterized by imbalances in cytokine and chemokine expression, play a crucial role in the progression of SFTS.
5 Genotyping of SFTSV
Takahashi et al. (2014) conducted a retrospective analysis of SFTS cases in Japan, showed that all Japanese SFTSV isolates clustered into two separate branches from those of Chinese SFTSV isolates, consistent with geographical distribution, suggesting independent natural transmission of SFTSV in Japan. Shi et al. (2017) further included 139 SFTSV isolates from Japan, South Korea, and China in GenBank for phylogenetic analysis. They clustered into five separate clades designated as C1, C2, C3, C4, and J. The study indicated that SFTSV isolates in China and Japan evolved independently within their respective regions. Lv et al. (2017) further divided SFTSV into genotypes C1-C5 and sublineages J1-J3 based on whole-genome sequence analysis for homologous recombination and gene reassortment. Additionally, two new sublineages, C6 and J4, were identified in the S gene segment of SFTSV.
Following the publication of SFTSV sequences from more regions, SFTSV has demonstrated broader genetic diversity. A six-genotype classification (A-F) is now widely used. Fu et al. (2016) classified SFTSV into six genotypes: A, B, C, D, E, and F. The prevalence of different SFTSV genotypes varies among countries. The main genotypes in China are A, D, E, and F, with genotype E being unique to China. In contrast, genotype B is only found in South Korea, Japan, and other regions. Yun et al. (2020) further divided genotype B SFTSV into three subtypes, B1, B2, and B3, based on genetic distance in South Korea. Park et al. (2024) identified the presence of genotype B4 in China, South Korea, and Japan and emphasizing its potential importance in viral evolution. It is noteworthy that there is a significant correlation between SFTSV genotype and mortality rates in different regions. The case fatality rate (CFR) of genotype B2 is the highest among genotype B and its subtypes, at 47.9%. The CFRs of genotypes D and F in China range from 17.3 to 22.7%, while genotype E has a CFR of only 12.5%, and this trend also exists in recombinant strains (Moon et al., 2023). The distribution of SFTSV genotypes in China, South Korea, and Japan is shown in Table 1.
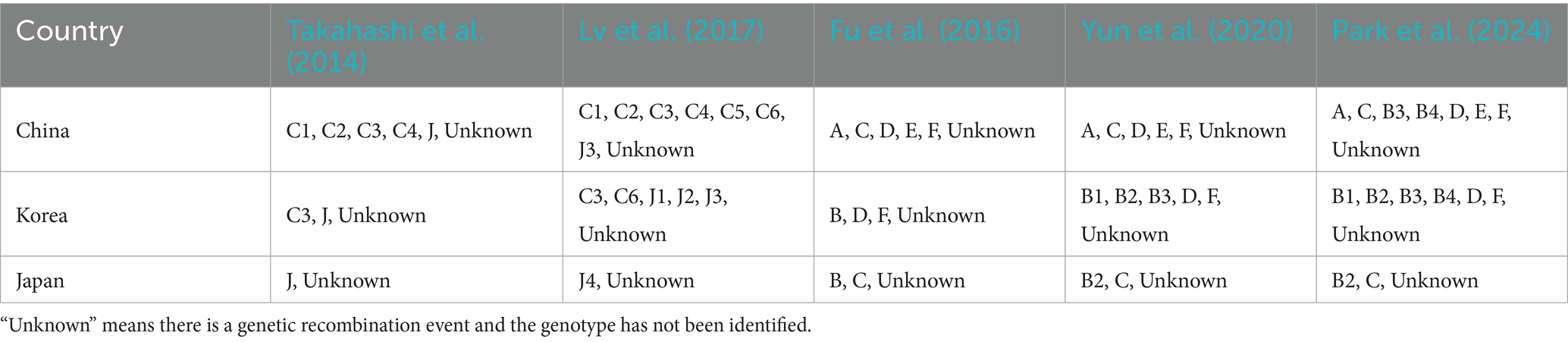
Table 1. SFTSV genotype distribution.
Pérez et al. (2024) classified SFTSV genomic evolution into three lineages. Lineage I comprised predominantly Chinese sequences (~95%), while Lineages II and III consisted mainly of South Korean variants. Clade II additionally contained Japanese isolates (~39%), with Lineage III incorporating Chinese and Thai sequences (~32%). Extensive homologous recombination was shown to accelerate SFTSV genomic evolution. The R624W and R962S mutations in the SFTSV glycoprotein precursor (GP) mediate pH-dependent cell membrane fusion (Tsuda et al., 2017; Tani et al., 2019). The N1891K mutation in the RNA-dependent RNA polymerase (RdRp) enhances enzymatic activity (Noda et al., 2020). The NS protein mutations P102A and K211R impair TPL2 signalling and IL-10 production, consequently reducing viral pathogenicity. These findings demonstrate that extensive genomic variations in SFTSV may contribute to regional differences in the pathogenesis and mortality rates of Severe Fever with Thrombocytopenia Syndrome (SFTS) (Ma et al., 2025).
6 Rapid detection of SFTSV
Quantitative real-time reverse transcription polymerase chain reaction (qRT-PCR) can be used to detect viral load in patients’ blood and serves as a predictive indicator for disease. However, since the increase in viral load during the early stage of SFTS infection occurs later than the onset of clinical symptoms, timely diagnosis is often not possible in clinical practice, leading to disease progression. Therefore, early and accurate diagnosis is crucial for effective treatment and disease management. Jang et al. (2022) developed a point-of-care molecular diagnostic method based on loop-mediated isothermal amplification (LAMP) technology to differentiate SFTSV infection from scrub typhus, with results available within 30 min and high specificity and sensitivity. Recombinase polymerase amplification (RPA) is an isothermal nucleic acid amplification technique that can rapidly and sensitively detect viral nucleic acids under constant temperature conditions. Zhou et al. (2020) evaluated the performance of RT-RPA for detecting SFTSV in serum samples from suspected SFTS patients, with sensitivity and specificity of 96.00 and 98.95%, respectively, showing good application prospects. Serological detection involves detecting SFTSV-specific antibodies in patients’ serum or plasma to diagnose the disease. Huang et al. (2019) developed an up-converting phosphor technology-based lateral flow (UPT-LF) detection method, coating SFTSV recombinant N protein on a biosensor to detect total SFTSV antibodies in serum through fluorescence signal collection, which can be used for on-site rapid detection. Zuo et al. (2018) developed a lateral flow immunochromatographic strip for ultra-sensitive detection of SFTSV nucleocapsid protein NP, with a detection limit as low as 1 ng/mL, suitable for rapid detection in remote areas.
RT-PCR is the most widely used molecular method for SFTSV diagnosis, offering high specificity and sensitivity. However, its primary limitations include the requirement for expensive thermal cycling equipment and specialised laboratory personnel, restricting its application in resource-limited settings (Zeng et al., 2024). Isothermal amplification technologies, such as LAMP and RPA, eliminate the need for costly thermal cyclers by enabling rapid amplification at a single fixed temperature. When coupled with simple readout devices, these methods facilitate rapid diagnosis in low-resource environments, such as primary healthcare facilities or field screenings in endemic areas. Nevertheless, isothermal amplification generates substantial by-products, particularly non-specific amplification, resulting in relatively lower sensitivity compared to other detection methods (Rolando et al., 2024). Serological assays for detecting SFTSV-specific antibodies are suitable for large-scale epidemiological screening and rapid field testing in resource-limited areas due to their speed, low cost, and minimal equipment requirements (Varghese et al., 2023). However, cross-reactivity with related viruses may lead to false-positive results, while false negatives can occur during the early infection window (Piantadosi and Kanjilal, 2020; Chan et al., 2022). Thus, serological testing should be supplemented with molecular methods to improve diagnostic accuracy. Although significant progress has been made in detecting SFTSV, there is still much work to be done in standardization, automation, and the development of multiplex detection methods to improve detection efficiency and accuracy (Ai et al., 2023).
7 Conclusion
SFTSV infection has become a global public health issue. Early diagnosis of SFTS based on typical clinical features and laboratory findings is crucial for improving patient survival rates in clinical practice. Further research on the pathogenesis of SFTS will help elucidate the mechanisms of DIC and MOF to reduce mortality and develop new therapeutic molecules. Comparative studies of viral isolates from different regions may clarify the genetic diversity and variation characteristics of SFTSV. Additionally, the development of rapid detection methods for SFTSV will aid in rapid diagnosis to contain and prevent viral spread.
Author contributions
YY: Conceptualization, Writing – original draft, Investigation, Writing – review & editing. JL: Formal analysis, Writing – review & editing. QL: Formal analysis, Visualization, Writing – review & editing. RZ: Supervision, Methodology, Writing – review & editing. YD: Conceptualization, Supervision, Writing – review & editing, Resources, Project administration. LS: Resources, Supervision, Project administration, Writing – review & editing, Conceptualization.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Gen AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ai, L., Wang, W., and Teng, Z. (2023). Advancements in the worldwide detection of severe fever with thrombocytopenia syndrome virus infection from 2009 to 2023. China CDC Wkly. 5, 687–693. doi: 10.46234/ccdcw2023.132
Casel, M. A., Park, S. J., and Choi, Y. K. (2021). Severe fever with thrombocytopenia syndrome virus: emerging novel phlebovirus and their control strategy. Exp. Mol. Med. 53, 713–722. doi: 10.1038/s12276-021-00610-1
Chan, K. R., Ismail, A. A., Thergarajan, G., Raju, C. S., Yam, H. C., Rishya, M., et al. (2022). Serological cross-reactivity among common flaviviruses. Front. Cell. Infect. Microbiol. 12:975398. doi: 10.3389/fcimb.2022.975398
Chang, Z., Gao, D., Liao, L., Sun, J., Zhang, G., Zhang, X., et al. (2024). Bispecific antibodies targeting two glycoproteins on SFTSV exhibit synergistic neutralization and protection in a mouse model. Proc. Natl. Acad. Sci. USA 121:e2400163121. doi: 10.1073/pnas.2400163121
Chen, L., Chen, T., Li, R., Xu, Y., and Xiong, Y. (2023). Recent advances in the study of the immune escape mechanism of SFTSV and its therapeutic agents. Viruses 15:940. doi: 10.3390/v15040940
Chen, C., Li, P., Li, K. F., Wang, H. L., Dai, Y. X., Cheng, X., et al. (2019). Animals as amplification hosts in the spread of severe fever with thrombocytopenia syndrome virus: a systematic review and meta-analysis. Int. J. Infect. Dis. 79, 77–84. doi: 10.1016/j.ijid.2018.11.017
Chen, X., Ye, H., Li, S., Jiao, B., Wu, J., Zeng, P., et al. (2017). Severe fever with thrombocytopenia syndrome virus inhibits exogenous type I IFN signaling pathway through its NSs in vitro. PLoS One 12:e0172744. doi: 10.1371/journal.pone.0172744
Chen, Q. L., Zhu, M. T., Chen, N., Yang, D., Yin, W. W., Mu, D., et al. (2022). Epidemiological characteristics of severe fever with thtrombocytopenia syndrome in China, 2011-2021. Zhonghua Liu Xing Bing Xue Za Zhi 43, 852–859. doi: 10.3760/cma.j.cn112338-20220325-00228
Crump, A., and Tanimoto, T. (2020). Severe fever with thrombocytopenia syndrome: Japan under threat from life-threatening emerging tick-borne disease. JMA J. 3, 295–302. doi: 10.31662/jmaj.2019-0073
Cui, H., Shen, S., Chen, L., Fan, Z., Wen, Q., Xing, Y., et al. (2024). Global epidemiology of severe fever with thrombocytopenia syndrome virus in human and animals: a systematic review and meta-analysis. Lancet Reg. Health. Western Pacific. 48:101133. doi: 10.1016/j.lanwpc.2024.101133
Dai, Z. N., Peng, X. F., Li, J. C., Zhao, J., Wu, Y. X., Yang, X., et al. (2022). Effect of genomic variations in severe fever with thrombocytopenia syndrome virus on the disease lethality. Emerg Microbes Infect. 11, 1672–1682. doi: 10.1080/22221751.2022.2081617
Du, Y., Cheng, N., Li, Y., Wang, H., You, A., Su, J., et al. (2019). Seroprevalance of antibodies specific for severe fever with thrombocytopenia syndrome virus and the discovery of asymptomatic infections in Henan Province, China. PLoS Neglected Tropical Diseases. 13:e0007242. doi: 10.1371/journal.pntd.0007242
Du, S., Peng, R., Xu, W., Qu, X., Wang, Y., Wang, J., et al. (2023). Cryo-EM structure of severe fever with thrombocytopenia syndrome virus. Nat. Commun. 14:6333. doi: 10.1038/s41467-023-41804-7
Fu, Y., Li, S., Zhang, Z., Man, S., Li, X., Zhang, W., et al. (2016). Phylogeographic analysis of severe fever with thrombocytopenia syndrome virus from Zhoushan Islands, China: implication for transmission across the ocean. Sci. Rep. 6:19563. doi: 10.1038/srep19563
Godsey, M. S., Savage, H. M., Burkhalter, K. L., Bosco-Lauth, A. M., and Delorey, M. J. (2016). Transmission of heartland virus (Bunyaviridae: Phlebovirus) by experimentally infected Amblyomma americanum (Acari: Ixodidae). J. Med. Entomol. 53, 1226–1233. doi: 10.1093/jme/tjw080
Han, S. W., Kang, J. G., Byeon, A. R., Cho, Y. K., Choi, K. S., and Chae, J. S. (2020). Severe fever with thrombocytopenia syndrome in canines from the Republic of Korea. Ticks Tick Borne Dis. 11:101454. doi: 10.1016/j.ttbdis.2020.101454
He, Z., Wang, B., Li, Y., Hu, K., Yi, Z., Ma, H., et al. (2021). Changes in peripheral blood cytokines in patients with severe fever with thrombocytopenia syndrome. J. Med. Virol. 93, 4704–4713. doi: 10.1002/jmv.26877
Hou, W., Jin, Y. H., Kang, H. S., and Kim, B. S. (2014). Interleukin-6 (IL-6) and IL-17 synergistically promote viral persistence by inhibiting cellular apoptosis and cytotoxic T cell function. J. Virol. 88, 8479–8489. doi: 10.1128/JVI.00724-14
Huang, X. X., Du, S. S., Li, A. Q., Li, C., Tian, T. T., Liu, T. Z., et al. (2024). Epidemiological characteristics of severe fever with thrombocytopenia syndrome in China, 2018-2021. Zhonghua Liu Xing Bing Xue Za Zhi 45, 112–116. doi: 10.3760/cma.j.cn112338-20230504-00274
Huang, X. Y., Du, Y. H., Wang, H. F., You, A. G., Li, Y., Su, J., et al. (2019). Prevalence of severe fever with thrombocytopenia syndrome virus in animals in Henan Province, China. Inf. Dis. Poverty. 8:56. doi: 10.1186/s40249-019-0569-x
Huang, X., Li, J., Li, A., Wang, S., and Li, D. (2021). Epidemiological characteristics of severe fever with thrombocytopenia syndrome from 2010 to 2019 in mainland China. Int. J. Environ. Res. Public Health 18:3092. doi: 10.3390/ijerph18063092
Huang, C., Wei, Q., Hu, Q., Wen, T., Xue, L., Li, S., et al. (2019). Rapid detection of severe fever with thrombocytopenia syndrome virus (SFTSV) total antibodies by up-converting phosphor technology-based lateral-flow assay. Luminescence 34, 162–167. doi: 10.1002/bio.3588
Jang, H., Casel, M. A. B., Jang, S.-g., Choi, J. H., Gil, J., Rollon, R., et al. (2024). Seasonal dynamics of Haemaphysalis tick species as SFTSV vectors in South Korea. Microbiol. Spectr. 12:e0048924. doi: 10.1128/spectrum.00489-24
Jang, W. S., Lim, D. H., Choe, Y. L., Nam, J., Moon, K. C., Kim, C., et al. (2022). Developing a multiplex loop-mediated isothermal amplification assay (LAMP) to determine severe fever with thrombocytopenia syndrome (SFTS) and scrub typhus. PLoS One 17:e0262302. doi: 10.1371/journal.pone.0262302
Ji, S. R., Byun, H. R., Rieu, M. S., Han, S. W., Nam, H. Y., Seo, S., et al. (2024). First detection of Bandavirus dabieense in ticks collected from migratory birds in the Republic of Korea. Acta Trop. 257:107279. doi: 10.1016/j.actatropica.2024.10727
Jost, S., and Altfeld, M. (2013). Control of human viral infections by natural killer cells. Annu. Rev. Immunol. 31, 163–194. doi: 10.1146/annurev-immunol-032712-100001
Khalil, J., Kato, H., and Fujita, T. (2021). The role of non-structural protein NSs in the pathogenesis of severe fever with thrombocytopenia syndrome. Viruses 13:876. doi: 10.3390/v13050876
Kim, D., Lai, C. J., Cha, I., and Jung, J. U. (2024). Current Progress of severe fever with thrombocytopenia syndrome virus (SFTSV) vaccine development. Viruses 16:128. doi: 10.3390/v16010128
Kim, E. H., and Park, S. J. (2023). Emerging tick-borne Dabie bandavirus: virology, epidemiology, and prevention. Microorganisms 11:2309. doi: 10.3390/microorganisms11092309
Kitagawa, Y., Sakai, M., Shimojima, M., Saijo, M., Itoh, M., and Gotoh, B. (2018). Nonstructural protein of severe fever with thrombocytopenia syndrome phlebovirus targets STAT2 and not STAT1 to inhibit type I interferon-stimulated JAK-STAT signaling. Microbes Infect. 20, 360–368. doi: 10.1016/j.micinf.2018.05.007
Kobayashi, Y., Kato, H., Yamagishi, T., Shimada, T., Matsui, T., Yoshikawa, T., et al. (2020). Severe fever with thrombocytopenia syndrome, Japan, 2013-2017. Emerg. Infect. Dis. 26, 692–699. doi: 10.3201/eid2604.191011
Kwon, J. S., Kim, J. Y., Jang, C. Y., Son, J. Y., Kim, W., Kim, T., et al. (2024). Effect of severe fever with thrombocytopenia syndrome virus genotype on disease severity, viral load, and cytokines in South Korea. Open Forum Infect. Dis. 11:ofae508. doi: 10.1093/ofid/ofae508
Lee, A. J., and Ashkar, A. A. (2018). The dual nature of type I and type II interferons. Front. Immunol. 9:2061. doi: 10.3389/fimmu.2018.02061
Lee, M., Lee, E., Kim, S. W., Kim, Y. K., Bae, I. G., Kim, J., et al. (2024). Severe fever with thrombocytopenia syndrome in South Korea, 2016-2021: clinical features of severe progression and complications. Am. J. Trop. Med. Hyg. 111, 661–670. doi: 10.4269/ajtmh.24-0062
Li, H., Li, X., Lv, S., Peng, X., Cui, N., Yang, T., et al. (2021). Single-cell landscape of peripheral immune responses to fatal SFTS. Cell Rep. 37:110039. doi: 10.1016/j.celrep.2021.110039
Li, J., Li, S., Yang, L., Cao, P., and Lu, J. (2021). Severe fever with thrombocytopenia syndrome virus: a highly lethal bunyavirus. Crit. Rev. Microbiol. 47, 112–125. doi: 10.1080/1040841X.2020.1847037
Li, P., Tong, Z. D., Li, K. F., Tang, A., Dai, Y. X., and Yan, J. B. (2017). Seroprevalence of severe fever with thrombocytopenia syndrome virus in China: a systematic review and meta-analysis. PLoS One 12:e0175592. doi: 10.1371/journal.pone.0175592
Li, M., Xiong, Y., Li, M., Zhang, W., Liu, J., Zhang, Y., et al. (2020). Depletion but activation of CD56dimCD16+ NK cells in acute infection with severe fever with thrombocytopenia syndrome virus. Virol. Sin. 35, 588–598. doi: 10.1007/s12250-020-00224-3
Li, M. M., Zhang, W. J., Weng, X. F., Li, M. Y., Liu, J., Xiong, Y., et al. (2018). CD4 T cell loss and Th2 and Th17 bias are associated with the severity of severe fever with thrombocytopenia syndrome (SFTS). Clin. Immunol. 195, 8–17. doi: 10.1016/j.clim.2018.07.009
Liang, S., Li, Z., Zhang, N., Wang, X., Qin, Y., Xie, W., et al. (2023). Epidemiological and spatiotemporal analysis of severe fever with thrombocytopenia syndrome in eastern China, 2011-2021. BMC Public Health 23:508. doi: 10.1186/s12889-023-15379-3
Liang, B., Xu, L., Li, M., Wang, H., Lu, S., Fan, L., et al. (2024). The association between elevated myocardial injury-related biomarker (TnI) and increased mortality in patients with severe fever with thrombocytopenia syndrome. Crit. Care Med. 52, 1509–1519. doi: 10.1097/CCM.0000000000006367
Lin, T. L., Ou, S. C., Maeda, K., Shimoda, H., Chan, J. P., Tu, W. C., et al. (2020). The first discovery of severe fever with thrombocytopenia syndrome virus in Taiwan. Emerg. Microbes Infect. 9, 148–151. doi: 10.1080/22221751.2019.1710436
Liu, Q., He, B., Huang, S. Y., Wei, F., and Zhu, X. Q. (2014). Severe fever with thrombocytopenia syndrome, an emerging tick-borne zoonosis. Lancet Infect. Dis. 14, 763–772. doi: 10.1016/S1473-3099(14)70718-2
Liu, S., Liu, H., Zhang, K., Li, X., Duan, Y., Wang, Z., et al. (2019). Proteasome inhibitor PS-341 effectively blocks infection by the severe fever with thrombocytopenia syndrome virus. Virol. Sin. 34, 572–582. doi: 10.1007/s12250-019-00162-9
Liu, J., Xu, M., Tang, B., Hu, L., Deng, F., Wang, H., et al. (2019). Single-particle tracking reveals the sequential entry process of the Bunyavirus severe fever with thrombocytopenia syndrome virus. Small 15:e1803788. doi: 10.1002/smll.201803788
Liu, J. W., Zhao, L., Luo, L. M., Liu, M. M., Sun, Y., Su, X., et al. (2016). Molecular evolution and spatial transmission of severe fever with thrombocytopenia syndrome virus based on complete genome sequences. PLoS One 11:e0151677. doi: 10.1371/journal.pone.0151677
Lv, Q., Zhang, H., Tian, L., Zhang, R., Zhang, Z., Li, J., et al. (2017). Novel sub-lineages, recombinants and reassortants of severe fever with thrombocytopenia syndrome virus. Ticks Tick Borne Dis. 8, 385–390. doi: 10.1016/j.ttbdis.2016.12.015
Ma, W., Hao, Y., Peng, C., Zhang, D., Yuan, Y., Xiao, P., et al. (2025). Analysis of gene differences between F and B epidemic lineages of Bandavirus Dabieense. Microorganisms. 13:292. doi: 10.3390/microorganisms13020292
Maslow, J. N., Kwon, J. J., Mikota, S. K., Spruill, S., Cho, Y., and Jeong, M. (2019). Severe fever and thrombocytopenia syndrome virus infection: considerations for vaccine evaluation of a rare disease. Hum. Vaccin. Immunother. 15, 2249–2257. doi: 10.1080/21645515.2019.1633875
Miao, D., Dai, K., Zhao, G. P., Li, X. L., Shi, W. Q., Zhang, J. S., et al. (2020). Mapping the global potential transmission hotspots for severe fever with thrombocytopenia syndrome by machine learning methods. Emerg. Microbes Inf. 9, 817–826. doi: 10.1080/22221751.2020.1748521
Min, Y. Q., Ning, Y. J., Wang, H., and Deng, F. (2020). A RIG-I-like receptor directs antiviral responses to a bunyavirus and is antagonized by virus-induced blockade of TRIM25-mediated ubiquitination. J. Biol. Chem. 295, 9691–9711. doi: 10.1074/jbc.RA120.013973
Moon, M. Y., Kim, H. K., Chung, S. J., Byun, J. H., Kim, H. N., Lee, W., et al. (2023). Genetic diversity, regional distribution, and clinical characteristics of severe fever with thrombocytopenia syndrome virus in Gangwon Province, Korea, a highly prevalent region, 2019-2021. Microorganisms. 11:2288. doi: 10.3390/microorganisms11092288
Niu, Y., Liu, Y., Huang, L., Liu, W., Cheng, Q., Liu, T., et al. (2024). Antiviral immunity of severe fever with thrombocytopenia syndrome: current understanding and implications for clinical treatment. Front. Immunol. 15:1348836. doi: 10.3389/fimmu.2024.1348836
Noda, K., Tsuda, Y., Kozawa, F., Igarashi, M., Shimizu, K., Arikawa, J., et al. (2020). The polarity of an amino acid at position 1891 of severe fever with thrombocytopenia syndrome virus L protein is critical for the polymerase activity. Viruses 13:33. doi: 10.3390/v13010033
Park, D., Kim, K. W., Kim, Y. I., Casel, M. A. B., Jang, H., Kwon, W., et al. (2024). Deciphering the evolutionary landscape of severe fever with thrombocytopenia syndrome virus across East Asia. Virus Evol. 10:veae054. doi: 10.1093/ve/veae054
Peng, C., Hao, Y., Yuan, Y., Ma, W., Zhang, D., Kong, J., et al. (2025). Bandavirus dabieense: a review of epidemiology, clinical characteristics, pathophysiology, treatment and prevention. Virulence 16:2520343. doi: 10.1080/21505594.2025.2520343
Pérez, L. J., Baele, G., Hong, S. L., Cloherty, G. A., and Berg, M. G. (2024). Ecological changes exacerbating the spread of invasive ticks has driven the dispersal of severe fever with thrombocytopenia syndrome virus throughout Southeast Asia. Mol. Biol. Evol. 41:msae173. doi: 10.1093/molbev/msae173
Piantadosi, A., and Kanjilal, S. (2020). Diagnostic approach for arboviral infections in the United States. J. Clin. Microbiol. 58:e01926-19. doi: 10.1128/JCM.01926-19
Rattanakomol, P., Khongwichit, S., Linsuwanon, P., Lee, K. H., Vongpunsawad, S., and Poovorawan, Y. (2022). Severe fever with thrombocytopenia syndrome virus infection, Thailand, 2019-2020. Emerg. Infect. Dis. 28, 2572–2574. doi: 10.3201/eid2812.221183
Ren, F., Shen, S., Ning, Y. J., Wang, Q., Dai, S., Shi, J., et al. (2021). Non-structural proteins of severe fever with thrombocytopenia syndrome virus suppress RNA synthesis in a transcriptionally active cDNA-derived viral RNA synthesis system. Front. Microbiol. 12:709517. doi: 10.3389/fmicb.2021.709517
Rolando, J. C., Melkonian, A. V., and Walt, D. R. (2024). The present and future landscapes of molecular diagnostics. Ann. Rev. Anal. Chem. 17, 459–474. doi: 10.1146/annurev-anchem-061622-015112
Saijo, M. (2022). Severe fever with thrombocytopenia syndrome, a viral hemorrhagic fever, endemic to Japan: achievements in and directions for medical research. Jpn. J. Infect. Dis. 75, 217–227. doi: 10.7883/yoken.JJID.2021.851
Schneider, W. M., Chevillotte, M. D., and Rice, C. M. (2014). Interferon-stimulated genes: a complex web of host defenses. Annu. Rev. Immunol. 32, 513–545. doi: 10.1146/annurev-immunol-032713-120231
Seo, J. W., Kim, D., Yun, N., and Kim, D. M. (2021). Clinical update of severe fever with thrombocytopenia syndrome. Viruses 13:1213. doi: 10.3390/v13071213
Shi, J., Hu, S., Liu, X., Yang, J., Liu, D., Wu, L., et al. (2017). Migration, recombination, and reassortment are involved in the evolution of severe fever with thrombocytopenia syndrome bunyavirus. Infect. Genet. Evol. 47, 109–117. doi: 10.1016/j.meegid.2016.11.015
Song, L., Zou, W., Wang, G., Qiu, L., and Sai, L. (2024). Cytokines and lymphocyte subsets are associated with disease severity of severe fever with thrombocytopenia syndrome. Virol. J. 21:126. doi: 10.1186/s12985-024-02403-0
Spiegel, M., Plegge, T., and Pöhlmann, S. (2016). The role of Phlebovirus glycoproteins in viral entry, assembly and release. Viruses 8:202. doi: 10.3390/v8070202
Staples, J. E., Pastula, D. M., Panella, A. J., Rabe, I. B., Kosoy, O. I., Walker, W. L., et al. (2020). Investigation of heartland virus disease throughout the United States, 2013-2017. Open Forum Infect. Dis. 7:ofaa125. doi: 10.1093/ofid/ofaa125
Takahashi, T., Maeda, K., Suzuki, T., Ishido, A., Shigeoka, T., Tominaga, T., et al. (2014). The first identification and retrospective study of severe fever with thrombocytopenia syndrome in Japan. J. Infect. Dis. 209, 816–827. doi: 10.1093/infdis/jit603
Tani, H., Kawachi, K., Kimura, M., Taniguchi, S., Shimojima, M., Fukushi, S., et al. (2019). Identification of the amino acid residue important for fusion of severe fever with thrombocytopenia syndrome virus glycoprotein. Virology 535, 102–110. doi: 10.1016/j.virol.2019.06.014
Tran, X. C., Yun, Y., Van An, L., Kim, S. H., Thao, N. T. P., Man, P. K. C., et al. (2019). Endemic severe fever with thrombocytopenia syndrome. Vietnam. Emerg Infect Dis. 25, 1029–1031. doi: 10.3201/eid2505.181463
Tsuda, Y., Igarashi, M., Ito, R., Nishio, S., Shimizu, K., Yoshimatsu, K., et al. (2017). The amino acid at position 624 in the glycoprotein of SFTSV (severe fever with thrombocytopenia virus) plays a critical role in low-pH-dependent cell fusion activity. Biomed. Res. 38, 89–97. doi: 10.2220/biomedres.38.89
Varghese, J., De Silva, I., and Millar, D. S. (2023). Latest advances in arbovirus diagnostics. Microorganisms 11:1159. doi: 10.3390/microorganisms11051159
Wada, Y., Miyamoto, S., Iida, S., Sano, K., Sato, Y., Ainai, A., et al. (2022). Propagation of activated B cells by in vitro severe fever with thrombocytopenia syndrome virus infection of human peripheral blood mononuclear cells. J. Infect. Dis. 225, 269–281. doi: 10.1093/infdis/jiab343
Wang, M., Tan, W., Li, J., Fang, L., and Yue, M. (2022). The endless wars: severe fever with thrombocytopenia syndrome virus, host immune and genetic factors. Front. Cell. Infect. Microbiol. 12:808098. doi: 10.3389/fcimb.2022.808098
Williams, H. M., Thorkelsson, S. R., Vogel, D., Milewski, M., Busch, C., Cusack, S., et al. (2023). Structural insights into viral genome replication by the severe fever with thrombocytopenia syndrome virus L protein. Nucleic Acids Res. 51, 1424–1442. doi: 10.1093/nar/gkac1249
Wu, Y., Zhu, Y., Gao, F., Jiao, Y., Oladejo, B. O., Chai, Y., et al. (2017). Structures of phlebovirus glycoprotein Gn and identification of a neutralizing antibody epitope. Proc. Natl. Acad. Sci. USA 114, E7564–E7573. doi: 10.1073/pnas.1705176114
Wuerth, J. D., and Weber, F. (2016). Phleboviruses and the type I interferon response. Viruses 8:174. doi: 10.3390/v8060174
Xing, B., Li, X. K., Zhang, S. F., Lu, Q. B., Du, J., Zhang, P. H., et al. (2018). Polymorphisms and haplotypes in the promoter of the TNF-α gene are associated with disease severity of severe fever with thrombocytopenia syndrome in Chinese Han population. PLoS Negl. Trop. Dis. 12:e0006547. doi: 10.1371/journal.pntd.0006547
Xu, S., Jiang, N., Nawaz, W., Liu, B., Zhang, F., Liu, Y., et al. (2021). Infection of humanized mice with a novel phlebovirus presented pathogenic features of severe fever with thrombocytopenia syndrome. PLoS Pathog. 17:e1009587. doi: 10.1371/journal.ppat.1009587
Yuan, F., and Zheng, A. (2017). Entry of severe fever with thrombocytopenia syndrome virus. Virol. Sin. 32, 44–50. doi: 10.1007/s12250-016-3858-6
Yue, Y., Ren, D., and Lun, X. (2024). Epidemic characteristics of fatal cases of severe fever with thrombocytopenia syndrome in China from 2010 to 2023. J. Trop. Dis. Parasitol. 22:257. doi: 10.20199/j.issn.1672-2302.2024.05.001
Yun, J. H., Hwang, H. J., Jung, J., Kim, M. J., Chong, Y. P., Lee, S. O., et al. (2019). Comparison of chest radiographic findings between severe fever with thrombocytopenia syndrome and scrub typhus: single center observational cross-sectional study in South Korea. Medicine 98:e17701. doi: 10.1097/MD.0000000000017701
Yun, S. M., Park, S. J., Kim, Y. I., Park, S. W., Yu, M. A., Kwon, H. I., et al. (2020). Genetic and pathogenic diversity of severe fever with thrombocytopenia syndrome virus (SFTSV) in South Korea. JCI Insight 5:e129531. doi: 10.1172/jci.insight.129531
Zeng, W., Chen, W., Liu, Y., Zhang, T., Zhai, C., Li, W., et al. (2024). Preamplification-free ultra-fast and ultra-sensitive point-of-care testing via LwaCas13a. Biosens. Bioelectron. 259:116400. doi: 10.1016/j.bios.2024.116400
Zeng, P., Ma, L., Gao, Z., Wang, J., Liu, J., Huang, X., et al. (2015). A study of seroprevalence and rates of asymptomatic viremia of severe fever with thrombocytopenia syndrome virus among Chinese blood donors. Transfusion 55, 965–971. doi: 10.1111/trf.12953
Zhang, L., Fu, Y., Wang, H., Guan, Y., Zhu, W., Guo, M., et al. (2019). Severe fever with thrombocytopenia syndrome virus-induced macrophage differentiation is regulated by mi R-146. Front. Immunol. 10:1095. doi: 10.3389/fimmu.2019.01095
Zhang, Z., Hu, X., Du, Q., Liu, J., Chen, X., Mo, P., et al. (2024). Rhabdomyolysis in severe fever with thrombocytopenia syndrome: associations with acute kidney injury and mortality. J. Med. Virol. 96:e70095. doi: 10.1002/jmv.70095
Zhang, S., Wang, J., Zhang, Q., Pan, Y., Zhang, Z., Geng, Y., et al. (2024). Association of liver function and prognosis in patients with severe fever with thrombocytopenia syndrome. PLoS Negl. Trop. Dis. 18:e0012068. doi: 10.1371/journal.pntd.0012068
Zhao, C., Zhang, X., Si, X., Ye, L., Lawrence, K., Lu, Y., et al. (2022). Hedgehogs as amplifying hosts of severe fever with thrombocytopenia syndrome virus, China. Emerg. Infect. Dis. 28, 2491–2499. doi: 10.3201/eid2812.220668
Zhou, J., Wang, Q., Zhu, L., Li, S., Li, W., Fu, Y., et al. (2020). Development and evaluation of a rapid detection assay for severe fever with thrombocytopenia syndrome virus based on reverse-transcription recombinase polymerase amplification. Mol. Cell. Probes 52:101580. doi: 10.1016/j.mcp.2020.101580
Zhu, L., Yin, F., Moming, A., Zhang, J., Wang, B., Gao, L., et al. (2019). First case of laboratory-confirmed severe fever with thrombocytopenia syndrome disease revealed the risk of SFTSV infection in Xinjiang, China. Emerg. Microbes Infect. 8, 1122–1125. doi: 10.1080/22221751.2019.1645573
Keywords: severe fever with thrombocytopenia syndrome virus, genotype, clinical features, pathogenesis, rapid detection
Citation: Yu Y, Li J, Liu Q, Zhai R, Dai Y and Sun L (2025) Advances in research on severe fever with thrombocytopenia syndrome virus. Front. Microbiol. 16:1622394. doi: 10.3389/fmicb.2025.1622394
Edited by:
Antonio Battisti, Institute of Experimental Zooprophylactic of the Lazio and Tuscany Regions (IZSLT), ItalyReviewed by:
Shouwen Du, Guangdong Academy of Agricultural Sciences, ChinaMichael G. Berg, Abbott, United States
Copyright © 2025 Yu, Li, Liu, Zhai, Dai and Sun. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yuzhu Dai, ZHl6NTg5NUBxcS5jb20=; Lei Sun, MjAyMDEyMDA1MEBtYWlsLnNkdS5lZHUuY24=