 Gizem Kezer1,2
Gizem Kezer1,2 Spiros Paramithiotis3
Spiros Paramithiotis3 Khaoula Khwaldia4
Khaoula Khwaldia4 Iskandar Azmy Harahap1,5
Iskandar Azmy Harahap1,5 Martina Čagalj6
Martina Čagalj6 Vida Šimat6
Vida Šimat6 Slim Smaoui7
Slim Smaoui7 Walid Elfalleh8
Walid Elfalleh8 Fatih Ozogul9,10
Fatih Ozogul9,10 Tuba Esatbeyoglu1*
Tuba Esatbeyoglu1*- 1Department of Molecular Food Chemistry and Food Development, Institute of Food and One Health, Gottfried Wilhelm Leibniz University, Hannover, Germany
- 2Department of Agricultural Biotechnology, Faculty of Agriculture, Kırşehir Ahi Evran University, Kırşehir, Türkiye
- 3Laboratory of Microbiology, Department of Biological Applications and Technology, University of Ioannina, Ioannina, Greece
- 4Laboratoire des Substances Naturelles (LSN), Institut National de Recherche et d’Analyse Physico-chimique (INRAP), BiotechPole Sidi Thabet 2020, Sidi Thabet, Tunisia
- 5Research Organization for Health, National Research and Innovation Agency, Bogor, Indonesia
- 6University Department of Marine Studies, University of Split, Split, Croatia
- 7Laboratory of Microbial and Enzymatic Biotechnologies and Biomolecules, Center of Biotechnology of Sfax (CBS), University of Sfax, Sfax, Tunisia
- 8Department of Biology, College of Science, Imam Mohammad Ibn Saud Islamic University (IMSIU), Riyadh, Saudi Arabia
- 9Department of Seafood Processing Technology, Faculty of Fisheries, Cukurova University, Adana, Türkiye
- 10Biotechnology Research and Application Center, Cukurova University, Adana, Türkiye
The gut-brain axis (GBA) represents a complex bidirectional communication system connecting the gastrointestinal tract and the central nervous system through neural, endocrine, immune, and metabolic pathways. Emerging evidence suggests that dietary interventions, particularly probiotics, prebiotics, and synbiotics, can influence the composition and activity of the gut microbiota, thereby modulating GBA function. Such modulation has been linked to potential benefits for cognitive performance, emotional regulation, and resilience against neurodegenerative and neuropsychiatric disorders. In addition, these interventions may contribute to immune homeostasis and the management of chronic conditions such as inflammatory bowel disease, irritable bowel syndrome, and multiple sclerosis. Nevertheless, the mechanisms underlying these effects and their long-term clinical relevance remain incompletely understood. In this narrative review, we systematically synthesize current clinical and preclinical evidence on the role of probiotics, prebiotics, and synbiotics in regulating the GBA. Particular attention is given to their impact on neurocognitive outcomes and systemic health, highlighting both the therapeutic potential and the existing gaps that warrant further investigation.
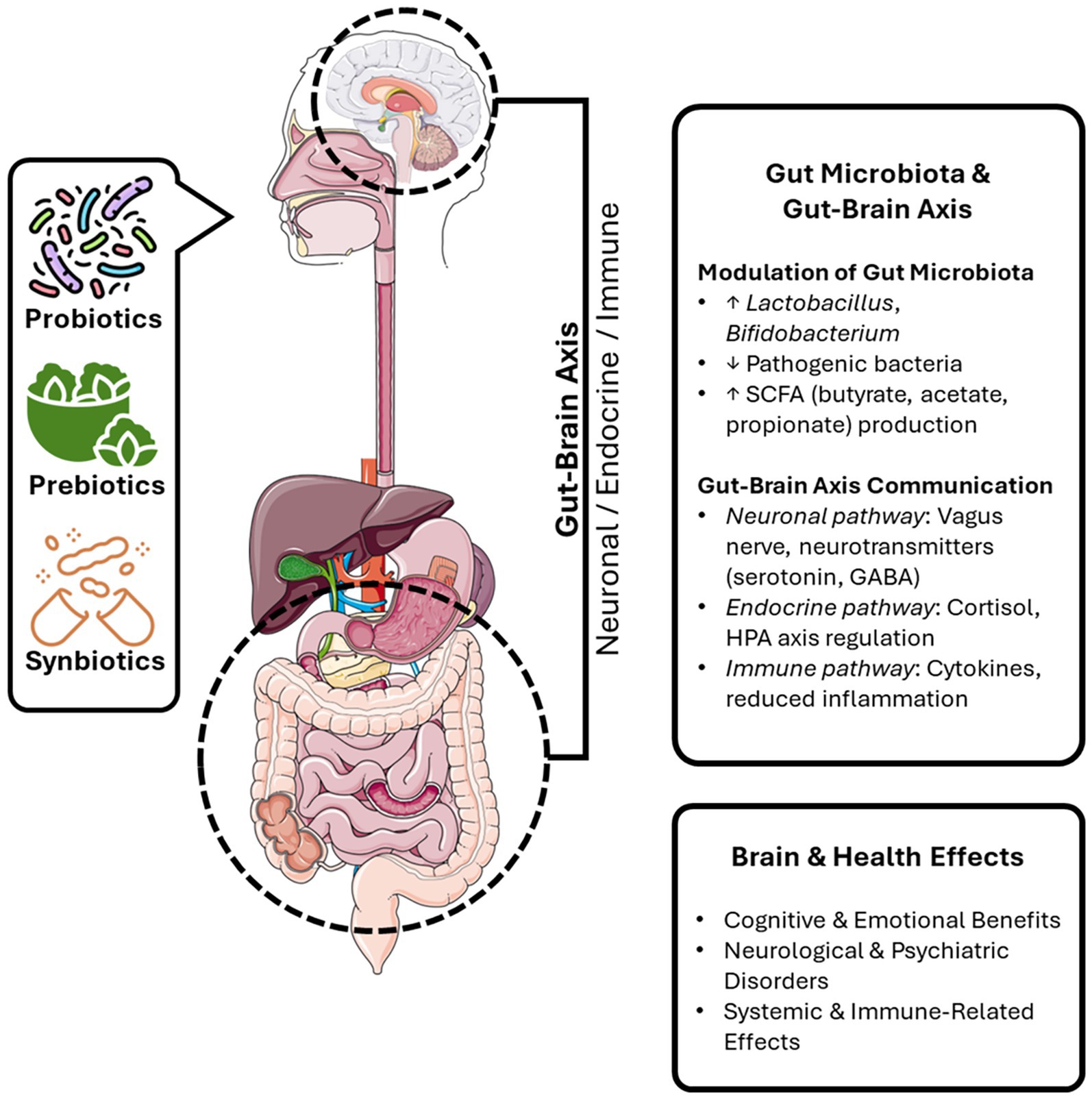
Graphical Abstract.
1 Introduction
The microbiota is the community of microorganisms colonizing the human body and interacting with the host. The mammalian gut alone contains around 2000 bacterial species, forming a complex ecosystem known as the gut microbiota (Góralczyk-Bińkowska et al., 2022; Maiuolo et al., 2021; Loh et al., 2024). This ecosystem is not randomly assembled; rather, it is shaped by selective pressures that reflect host genetics, diet, environment, and evolutionary history. In healthy adults, the gut microbiota is dominated by five phyla: Firmicutes (79.4%), Bacteroidetes (16.9%), Actinobacteria (2.5%), Proteobacteria (1%), and Verrucomicrobia (0.1%; Szymczak-Pajor et al., 2025; Maiuolo et al., 2021).
Members of the gut microbiota contribute to a wide range of metabolic functions by expressing enzymes and genes that facilitate nutrient conversion, energy harvest, and biosynthesis of essential compounds. These include amino acids, vitamins, short-chain fatty acids (SCFAs), and lipids, which play key roles in host physiology. Furthermore, the microbiota produces antimicrobial substances that protect against pathogenic colonization and supports intestinal barrier maturation and immune system regulation. A balanced and diverse microbiota is therefore critical for maintaining host health. Conversely, microbial diversity and community structure vary markedly among individuals due to factors such as mode of birth, early-life nutrition, lifestyle, pharmacological exposure, and genetic background (Morais et al., 2021; Maiuolo et al., 2021).
Recent insights suggest that the bioactive landscape of microbiota-derived metabolites is much more complex than previously assumed, with specialized peptides exhibiting antibacterial, immunomodulatory, and signaling roles (Shah and Shim, 2025; Shah et al., 2025). These discoveries highlight the microbiome not only as a determinant of host well-being but also as a source of novel therapeutic strategies.
The gastrointestinal tract remains the primary habitat for this microbial community, containing trillions of microorganisms, which outnumber host cells by nearly tenfold. Dysbiosis, defined as an imbalance of the gut microbiota, has been implicated in a range of metabolic, immunological, and neurological conditions (Verma et al., 2025; Bhagwat et al., 2025). The microbiota is now recognized as an active participant in host physiology, influencing systemic metabolism, immune development, and organ function.
One of the most studied aspects of host–microbe interaction is the bidirectional communication between the gut microbiota and the central nervous system (CNS), termed the microbiota-gut-brain axis. Although the gut and brain are anatomically distinct, multiple biological pathways facilitate this crosstalk, including neural (vagus nerve, enteric nervous system), immune, and endocrine signaling. Through the production of neurotransmitters, metabolites, and hormones, gut microbes are capable of modulating CNS activity (Liang et al., 2018; Loh et al., 2024; Ashique et al., 2024).
The microbiota-gut-brain axis thus represents a complex communication network that integrates microbial, immune, endocrine, and neural signaling to maintain homeostasis. However, perturbations in this system may contribute to the pathogenesis of neuropsychiatric and neurodegenerative diseases (Morais et al., 2021). Against this background, probiotics, prebiotics, and synbiotics have gained attention for their potential to beneficially modulate gut microbiota composition and function (Ansari et al., 2023). Probiotics are defined as live microorganisms that confer health benefits when consumed in adequate amounts, with Lactobacillus and Bifidobacterium among the most studied genera (Fekete et al., 2024; Ansari et al., 2023). Prebiotics, including galacto-oligosaccharides (GOS), fructo-oligosaccharides (FOS), and xylo-oligosaccharides, are nondigestible substrates that selectively stimulate the growth or activity of beneficial microbes (Ansari et al., 2023; Fekete et al., 2024). Synbiotics combine probiotics and prebiotics to act synergistically, with benefits ranging from improved digestion to potential roles in neuropsychiatric health (Markowiak and Śliżewska, 2017; Fekete et al., 2024; Ansari et al., 2023).
Although preclinical and clinical evidence suggests that microbiota-targeted interventions can influence CNS outcomes such as mood, cognition, and stress resilience (Chudzik et al., 2021; Radford-Smith and Anthony, 2023; Fekete et al., 2024), findings remain inconsistent. Randomized controlled trials have reported both positive effects and null results, reflecting strain-specificity, dosage variability, treatment duration, and methodological differences (Hofmeister et al., 2021; Alli et al., 2022; Nikolova et al., 2019). Moreover, the strong effects observed in animal studies have not always translated to humans (Forssten et al., 2022; Slykerman et al., 2025). Limitations including small sample sizes, heterogeneous populations, and lack of standardized protocols reduce the generalizability of current findings. There is also debate about whether beneficial effects arise from direct microbial activity, modulation of host–microbe interactions, or downstream immunological and metabolic changes.
Taken together, these controversies underscore that while probiotics, prebiotics, and synbiotics are promising as adjunctive approaches to CNS disorders, their clinical efficacy remains inconclusive. Alongside established strategies such as diet modification and fecal microbiota transplantation, these interventions represent an emerging frontier in brain-gut research. Future investigations should focus on large-scale, multicenter clinical trials, and mechanistic studies to clarify pathways of action and define clinical relevance. This review therefore aims to provide a critical synthesis of the current literature, highlighting mechanisms, therapeutic potential, limitations, and research gaps concerning the effects of probiotics, prebiotics, and synbiotics on the gut-brain axis. In preparing this narrative review, we performed a comprehensive search of relevant peer-reviewed literature using databases such as PubMed, Scopus, and Google Scholar. Articles were identified through combinations of keywords including probiotics, prebiotics, synbiotics, gut-brain axis, and neurocognitive health. Priority was given to recent publications (within the past 10–15 years), landmark studies, and mechanistic reports that provide insight into underlying pathways. Reference lists of pertinent papers were also examined to ensure inclusion of additional relevant works. Figures were generated and adapted using Microsoft PowerPoint and BioRender, based on data synthesized from the reviewed studies. This approach ensures both breadth and depth of coverage, while maintaining the narrative character of the review.
2 Probiotics
Among the many microbes that are an essential part of human life, probiotics have been recognized and extensively studied for their health benefits, particularly for the prevention of various gastrointestinal, metabolic and chronic diseases (Swanson et al., 2020). To be classified as a probiotic, the strain must be non-pathogenic, non-toxic, free from transferable antibiotic resistance genes, adequately characterized, tested for safety and technical characteristics for the intended use, maintain a viable population throughout its shelf life, and be proven to confer health benefits (Hill et al., 2014). In addition, a suitable and efficient probiotic must fulfill several functional criteria, such as maintenance of genetic integrity, resistance to exposure to low pH and bile salts, effective adherence to intestinal epithelial cells, production of beneficial metabolites, stability under industrial processing conditions, and the ability to multiply efficiently in the intestinal environment (de Melo Pereira et al., 2018).
Antimicrobial resistance must be considered in the safety assessment of probiotics, as strains carrying transmissible antibiotic resistance genes (ARG) could trigger horizontal gene transfer (HGT) in the gut, meaning that ARG can be transferred to pathogenic bacteria in the gut microbiome, resulting in drug-resistant strains. Therefore international FAO/WHO guidelines and the EFSA Qualified Presumption of Safety (QPS), emphasize that probiotics intended for human use must be free of ARG, while intrinsic, non-transferable resistance is generally acceptable. For example, many lactobacilli are intrinsically resistant to vancomycin due to their cell wall characteristics that result in reduced vancomycin binding. The current best practice recognizes the importance of rigorous screening through a multi-step approach: Whole genome sequencing for species identification and in silico ARG/mobilome analysis; phenotypic susceptibility testing to confirm resistance patterns; and, if indicated, conjugation or transmissibility testing to assess HGT potential [Tóth et al., 2021; Merenstein et al., 2023; Byakika et al., 2019; EFSA Panel on Contaminants in the Food Chain (CONTAM), 2018; EFSA BIOHAZ Panel et al., 2025; FAO/WHO, 2006].
Throughout history, fermented foods such as yoghurt and fermented vegetables have been the main source of probiotics in the human diet. Today, probiotics are available as dietary supplements and are used to fortify foods in a variety of strains and dosages. The psychobiotic effects of probiotics are strain- and dose-specific, and have been studied to identify potential therapeutic applications and to develop more efficient delivery systems. The most commonly used probiotics belong primarily to the genera Lactobacillus and Bifidobacterium. Common species include Lacticaseibacillus casei (L. casei), Lactiplantibacillus plantarum (L. plantarum), Lactobacillus acidophilus (L. acidophilus), Lactobacillus helveticus (L. helveticus), Lacticaseibacillus rhamnosus (L. rhamnosus), Bifidobacterium longum (B. longum), Bifidobacterium bifidum (B. bifidum), and Bifidobacterium breve (B. breve). Other probiotic strains that are gaining interest are Saccharomyces cerevisiae var. boulardii (a beneficial yeast), Streptococcus thermophilus, and strains of Bacillus, Lactococcus, Enterococcus and some Escherichia coli (E. coli) (Sarita et al., 2025).
2.1 Mechanisms
Probiotics (sometimes referred to as psychobiotics in this context) can modulate communication between the microbiota, gut and brain through multiple pathways. They influence neuronal signaling (e.g., via the vagus nerve and the enteric nervous system), hormonal responses (such as modulation of cortisol and the HPA axis) and immune activity (including cytokine regulation and inflammation; Mörkl et al., 2020). Certain probiotic bacteria, particularly Lactobacillus and Bifidobacterium species, produce neuroactive compounds - for instance, neurotransmitters such as GABA and serotonin, as well as short-chain fatty acids - that can affect brain function (Rahmannia et al., 2024). In addition, probiotics strengthen the gut barrier by reducing intestinal permeability and systemic inflammation, thereby protecting the brain from inflammatory stress (Rahmannia et al., 2024). Through these mechanisms—such as reducing neuroinflammation, modulating neurotransmitter levels, and influencing neuronal circuitry via the vagus nerve—probiotics contribute to a more favorable biochemical environment for brain health and emotional regulation.
2.2 Clinical evidence and limitations
A review by Dronkers et al. (2020) reported that probiotics (the most studied strains were Lacticaseibacillus rhamnosus GG (LGG) and Bifidobacterium animalis subsp. lactis BB12) were administered in over 1,000 clinical trials with an average sample size of 74 participants. These clinical studies addressed 700 different diseases and conditions and were registered at ClinicalTrials.gov run by the United States National Library of Medicine and/or the World Health Organization’s International Clinical Trials Registry Platform (Dronkers et al., 2020). In addition to strain and dosage activity, the potential benefits of probiotics are often limited by interactions with the host microbiome (Suez et al., 2019). In the last 5 years, there have been more than 500 case studies/year, systematic reviews and meta-analyses covering a wide range of conditions, confirming that probiotic interventions have beneficial effects in various gastrointestinal, metabolic, immunological, neuropsychiatric and various other conditions (Bagdadi et al., 2025). In particular, probiotics have been shown to exert condition-specific benefits through multiple mechanisms. In psychiatric disorders (excluding schizophrenia), clinical studies reported improvements in mood regulation, anxiety reduction, and cognitive performance, possibly mediated by modulation of the gut-brain axis and reduction of systemic inflammation (Hong et al., 2022; de Lima et al., 2025). In allergic diseases, probiotics were associated with decreased symptom severity and improved immunological tolerance, potentially via restoration of gut microbial balance and enhancement of regulatory T-cell responses (Xi et al., 2025). For patients with type 2 diabetes mellitus, probiotic supplementation improved glycemic control, insulin sensitivity, and inflammatory markers, highlighting their role in metabolic regulation (Wang et al., 2024). In gastrointestinal disorders such as irritable bowel syndrome, probiotics alleviated abdominal pain, bloating, and irregular bowel habits, likely through normalization of gut motility and modulation of the gut microbiota (Ceccherini et al., 2022; Li et al., 2020). Similarly, in inflammatory bowel diseases, osteoarthritis, and chronic kidney disease, clinical evidence supports their role in reducing disease activity, inflammatory biomarkers, and oxidative stress, thereby contributing to improved quality of life (Karim, 2025; Liu C, et al., 2024). However, for diseases such as COVID-19, systemic sclerosis, Crohn’s disease and scleroderma, the results of the studies are inadequate. Clinical efficacy remains limited, and interpretation of results is compromised by the lack of standardization when using different strains (even within the same species), the inability to determine individual contributions in multi-strain formulations and the lack of consistency in dosage and duration of probiotic use. Furthermore, reproducibility of clinical trials is limited as they differ in design (small numbers of participants and heterogeneous groups, short duration), do not capture participants’ health status, age, diet and baseline microbiota, focus on short-term symptom improvement and report symptom relief without investigating mechanisms, e.g., metabolomics and immunomodulation. Common probiotic strains have been extensively studied and are well known for their health benefits (Table 1).
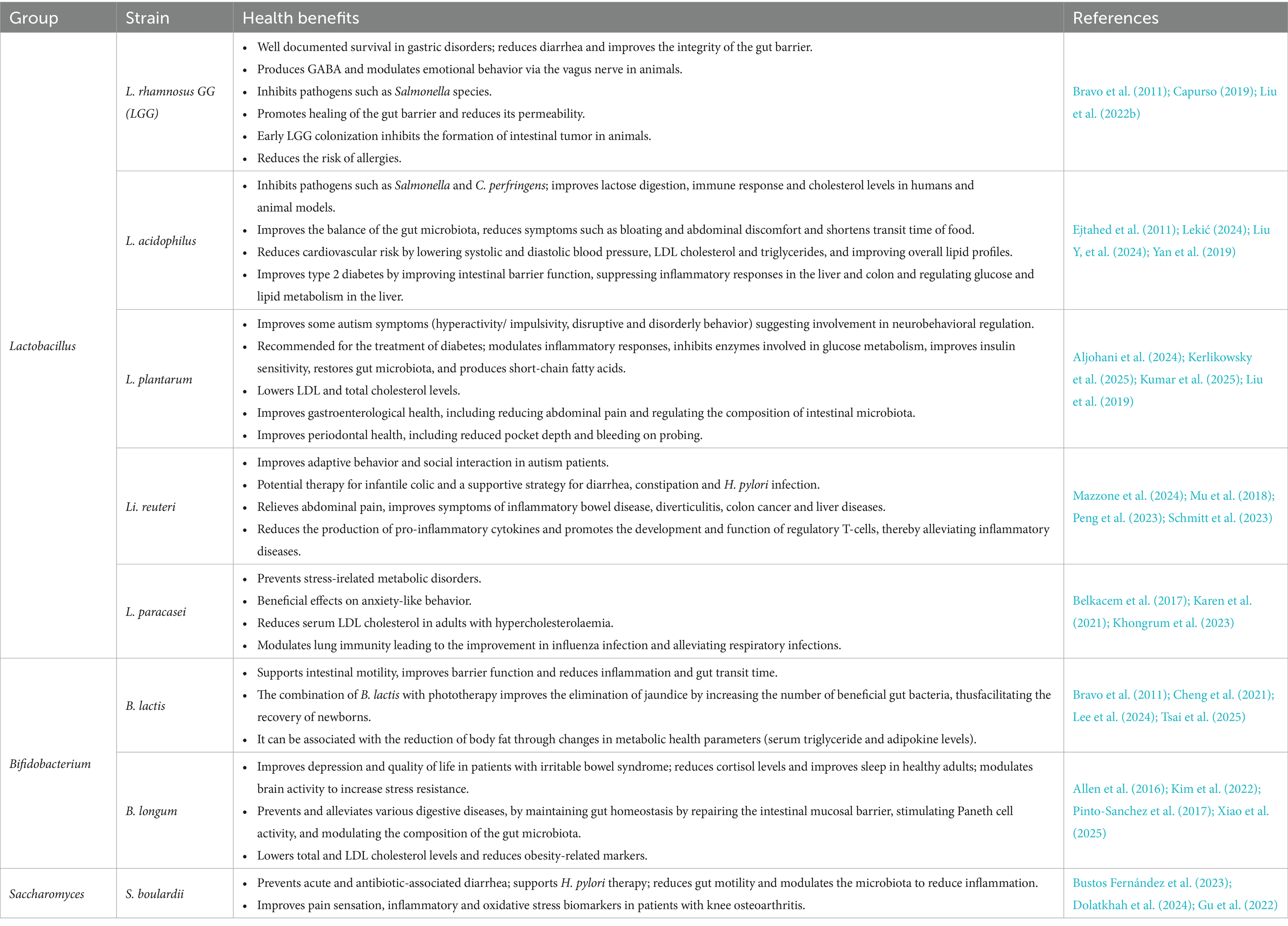
Table 1. Documented health benefits of specific probiotic strains from clinical and preclinical studies (covered period 2020–2025).
2.3 Innovations
Recent innovations in the use of probiotics include the development of multi-strain and customized psychobiotic formulations aimed at influencing anxiety and depression, cognition and neuroinflammation (Kamal et al., 2025; Messaoudi et al., 2011); the combination of probiotics with prebiotics (fiber-based substrates that promote the growth of probiotics), resulting in synbiotic therapies for enhanced effects on the gut and brain; next-generation encapsulation and delivery technologies (e.g., microbiotics); and the development of new products (e.g., microencapsulation, enteric coatings, phage-probiotic combinations) to improve strain viability and targeted release in the gut (D’Amico et al., 2025; Ranadheera et al., 2017); precision approaches using specific strains such as Limosilactobacillus reuteri (L. reuteri) for behavioral modulation in autism or B. longum for stress resistance; and ongoing research on genetically engineered probiotics that produce neuroactive compounds (e.g., GABA, serotonin) or immunomodulatory molecules directly in the gut (Charbonneau et al., 2020). Taken together, these innovations are moving probiotics from general gut health agents to targeted therapeutics for gut- and brain-related conditions such as anxiety, depression, Alzheimer’s, Parkinson’s, and autism spectrum disorders. They are increasingly recognized as modulators of gut and systemic health, including the gut-brain axis (GBA; Ali et al., 2025). Future studies should conduct longer-term studies considering the standardized strain-specific design and protocols, and personalized approaches based on patients’ microbiome profiles.
3 Prebiotics
Prebiotics have long been used to modulate gut microbiota metabolic function, and enhance host health (Sanders et al., 2019). Microbial enzymes and substrates could provide important potential for prebiotics production, with extensive use in the food and pharmaceutical industries. Among the most extensively employed prebiotics are galacto-oligosaccharides (GOS), xylo-oligosaccharides (XOS), and fructo-oligosaccharides (FOS).
3.1 Galacto-oligosaccharides
Several innovative microbial approaches have been extensively explored for the production of galacto-oligosaccharides (GOS), offering sustainable and efficient alternatives to conventional synthesis methods. One remarkable process includes the β-galactosidase immobilized on different substrates (Hackenhaar et al., 2021; Carević et al., 2018). Geiger et al. (2016) stated that a simple recombinant β-galactosidase from Streptococcus thermophilus DSM 20259 could convert 80% of whey lactose to GOS in 5 h. They also reported that ≈ 1 kg of GOS was produced from 3 kg of whey permeate powder. From Greek yogurt, the β-galactosidase synthesized by Cryptococcus laurentii whole-cells produced the GOS whey at 36% (w/w), and 50% of the initial lactose was converted at a specific productivity equal to 2.3 mg/U·h (Fischer and Kleinschmidt, 2021). Based on an expression system developed on the T7 RNA polymerase promoter in E. coli, Kittibunchakul et al. (2019) noted a high recombinant β-galactosidase activity (26,000 U/L). This value was 28-fold and 1,000-fold higher than the production of native β-galactosidase from L. helveticus DSM 20075 when grown on lactose and glucose, respectively. To improve their catalytic activity and GOS production, seven β-glucosidase mutants were obtained from Thermotoga naphthophila RKU-10. Interestingly, the F414S mutant showed efficient properties since the GOS production was improved from 140 mM to 207 Mm using 0.2 mM lactose (Yang et al., 2018). On free or immobilized forms, commercial enzymes have been employed in GOS production. The primary commercial enzymes used in GOS synthesis process are bacterial β-galactosidases (e.g., Kluyveromyces lactis) and fungal (e.g., Aspergillus oryzae) sources (Maráz et al., 2022). From different microbial sources, Mano et al. (2019) confirmed that Kluyveromyces lactis commercial enzyme named Lactozyme™ 2,600 L could display an optimal performance for lactose conversion, yield and specific productivity (50 g GOS/g enzyme×h).
3.2 Xylo-oligosaccharides
Due to their superior properties compared to other prebiotics, xylo-oligosaccharides (XOS) have gained more attention. XOS are stable across a broad pH range (2.5–8.0) and at temperatures up to 100 °C. Moreover, xylobiose has 0.3–0.4 times the sweetness of sucrose, and the approved daily dietary intake of XOS (2.1 g) is lower than that of most other oligosaccharides (Amorim et al., 2019). Notwithstanding these merits, XOS is more costly than the other prebiotics (Yegin, 2023). Because of the diverse enzyme systems they possess, each microorganism has a distinct XOS utilization pattern. For instance, XOS containing uronic acid are utilized by only a few bifidobacteria of human origin. Bifidobacterium adolescentis can metabolize both arabino-XOS and linear XOS (Falck et al., 2013), whereas Levilactobacillus brevis grows preferentially on linear XOS (Precup et al., 2022).
To date, most commercial xylanases are produced by bacteria (e.g., Bacillus and Streptomyces) and fungi (e.g., Thermomyces, Trichoderma, and Aspergillus). As a result, xylanases from these sources became dominant in the enzyme market. To categorize the extracellular enzymes including xylanases synthesized by Bacillus sp. AR03, Hero et al. (2021) exploited a proteomic approach. By LC–MS/MS identification, these authors reported a glucuronoxylanase GH30-8 and an endoglucanase GH5-2. From Bacillus sp. strain BP-7. Gallardo et al. (2010) reported that GH5 xylanase (Xyn5B) was able to act on linear XOS, and xylooligomers with methylglucuronic groups were generated. A novel xylanase produced by Streptomyces spp. (B6) was able to generate two xylanases attributed to GH10 and GH11 families (Liu et al., 2022a). In another study conducted by Boonchuay et al. (2014), Streptomyces thermovulgaris TISTR1948 used xylanase for XOS production. The enzyme production was conducted at 50 °C and 250 rpm for 96 h utilizing the rice straw as a carbon source, and the principal oligomer was xylobiose with 85.15 mg/g. Adsul et al. (2009) described an efficient process for the hydrolysis of xylan by using Streptomyces matensis xylanase, yielding mainly xylotriose and xylobiose as the predominant XOS products. Enzymes from Aspergillus have also been employed for XOS production using lignocellulosic biomass. For example, Akpinar et al. (2007) applied a two-stage approach to produce XOS from cotton stalk, first extracting xylan with KOH and then hydrolyzing it with commercial Aspergillus niger xylanase. Maximum XOS production was observed at 40 °C and pH 5.4 with 2% xylan (10 mL) and 4.4 U/mL enzyme (1 mL). To produce XOS by a packed bed reactor in continuous mode, Aragon et al. (2013) employed xylanase immobilization of Aspergillus versicolor by using several support materials. The glyoxyl agarose constitutes the most effective support for xylanase immobilization and maintained till 85% of its catalytic activity. After incubation at 60 °C, the immobilized enzyme was nearly 700 times more stable than the free fraction, and retained full activity after 10 cycles of 1-h process. In addition, the immobilized xylanase delivered 2.5-fold higher xylobiose production as compared to free fraction.
The xylan bioconversion from agricultural residues into XOS without prior pre-treatment has been revealed to be possibly economical alternate for industrial application. Crude xylanase from Aspergillus fumigatus R1 was qualified to yield 1.08% (w/w) of XOS from raw wheat husk xylan. XOS with a DP up to 5 were detected in the final hydrolysate, being xylobiose the greatest principal oligosaccharide during the entire reaction time (Chavan et al., 2023). This enzymatic process avoids the formation of unwanted by-products typical of chemical extraction.
3.3 Fructo-oligosaccharides
Fructo-oligosaccharides (FOS) are enzymatically produced from sucrose through a transfructosylating reaction catalyzed by β-fructofuranosidase (FFase) or fructosyltransferase (FTase) enzymes. Aspergillus flavus NFCCI 2364 FTase was explored to produce FOS from 16 agro-wastes (Ganaie et al., 2017). Smaali et al. (2012) used β- Ffase from Aspergillus awamori NBRC4033, demonstrating high yields and indicating the efficiency of agro-residue biomasses as substrates. Silva et al. (2013) stated that inulinase from Aspergillus niger (A. niger) and Kluyveromyces marxianus (K. marxianus) NRRL Y 7571 was able to generate FOS from inulin, with specific yields of kestose, nystose, and fructosyl nystose. Diez-Municio et al. (2013) showed that the inulosucrase from Lactobacillus gasseri DSM 20604 could generate FOS and maltosylfructosides (MFOS) from sucrose and sucrose/maltose mixtures. For short-chain FOS (scFOS) and oligolevans production, an inventive two-phase system of levansucrase (from Bacillus amyloliquefaciens)/endo-inulinase (from A. niger) used the sucrose. This system permitted levansucrase to create levans, while endoinulinase monitored molecule size, with 6-kestose being the primary scFOS (Ni et al., 2021). In addition, the immobilization of levansucrase improved the levan production over scFOS (Ni et al., 2021). To produce FOS from sucrose, Soliman et al. (2017) studied the immobilization of inulinase isolated from A. niger. The used material was on polyurethane foam, attaining a 30% total FOS yield, including GF2, GF3, and GF4. Bersaneti et al. (2018) revealed that levansucrase from B. subtilis could generate FOS and levan concurrently at 41 g/L of FOS and 87 g/L, respectively. Huang et al. (2016) stated that Aspergillus aculeatus M105 produced extracellular FTase, achieving FOS yields of 68 and 66% (w/w), respectively. By using the inulosucrase (IslA4), issued from Leuconostoc citreum, and sucrose, Peña-Cardeña et al. (2015) produced FOS containing compounds f-nystose, nystose, neokestose, 1-kestose and 6-kestose.
3.4 Other prebiotics
Many additional produced prebiotics can be isolated or produced from several microbial sources. Manno-oligosaccharides (MOS) production emphasizes a substantial progress in the utilization of renewable resources for producing appreciated prebiotic. A genetically engineered endo-β-(1,4)-mannanase, isolated from B. subtilis and expressed in Escherichia coli, could generate mannans to MOS with a degree of polymerization arraying from 4 to 7 (Sathitkowitchai et al., 2022). The β-mannanase from Penicillium aculeatum APS1 can degrade glucomannan (from konjac) and galactomannan (from locust guar and bean gums). The enzyme produces low molecular weight at DP ≤ 4 (Bangoria et al., 2021). From Streptomyces cyaenus, a mannanase hydrolyzed palm cake kernel with oligo-mannans (DP ≤ 7). In addition, the mannotriose and mannobiose were detected throughout the reaction period (up to 8 h; Purnawan et al., 2017).
To improve the production of isomalto-oligosaccharides (IMOs), recombinant enzymes engineering showed an important progression. In this line, Kaulpiboon et al. (2015) used pullulanase, a modified amylomaltase Y101S, and transglucosidase from A. niger. This enzyme blend enabled the production of long-chain IMOs at pH 7.0 and 40 °C.
Table 2 summarizes some examples of major microbial enzymatic producing prebiotic (GOS, XOS and FOS). Besides the biochemical and production aspects described above, the physiological and clinical effects of prebiotics have also been investigated extensively. Several studies have shown that the intake of prebiotics such as GOS, FOS, and XOS can positively alter the composition of the gut microbiota, increase short-chain fatty acid production, and improve intestinal barrier function. In clinical trials, prebiotics have been associated with positive results in gastrointestinal health, including relief from constipation and irritable bowel syndrome, with beneficial effects on metabolic disorders such as obesity and type 2 diabetes (Sanders et al., 2019; Markowiak and Śliżewska, 2017; Davani-Davari et al., 2019; Gibson et al., 2017). These evidences highlight the importance of prebiotics not only as industrially significant compounds but also as key regulators of human health.
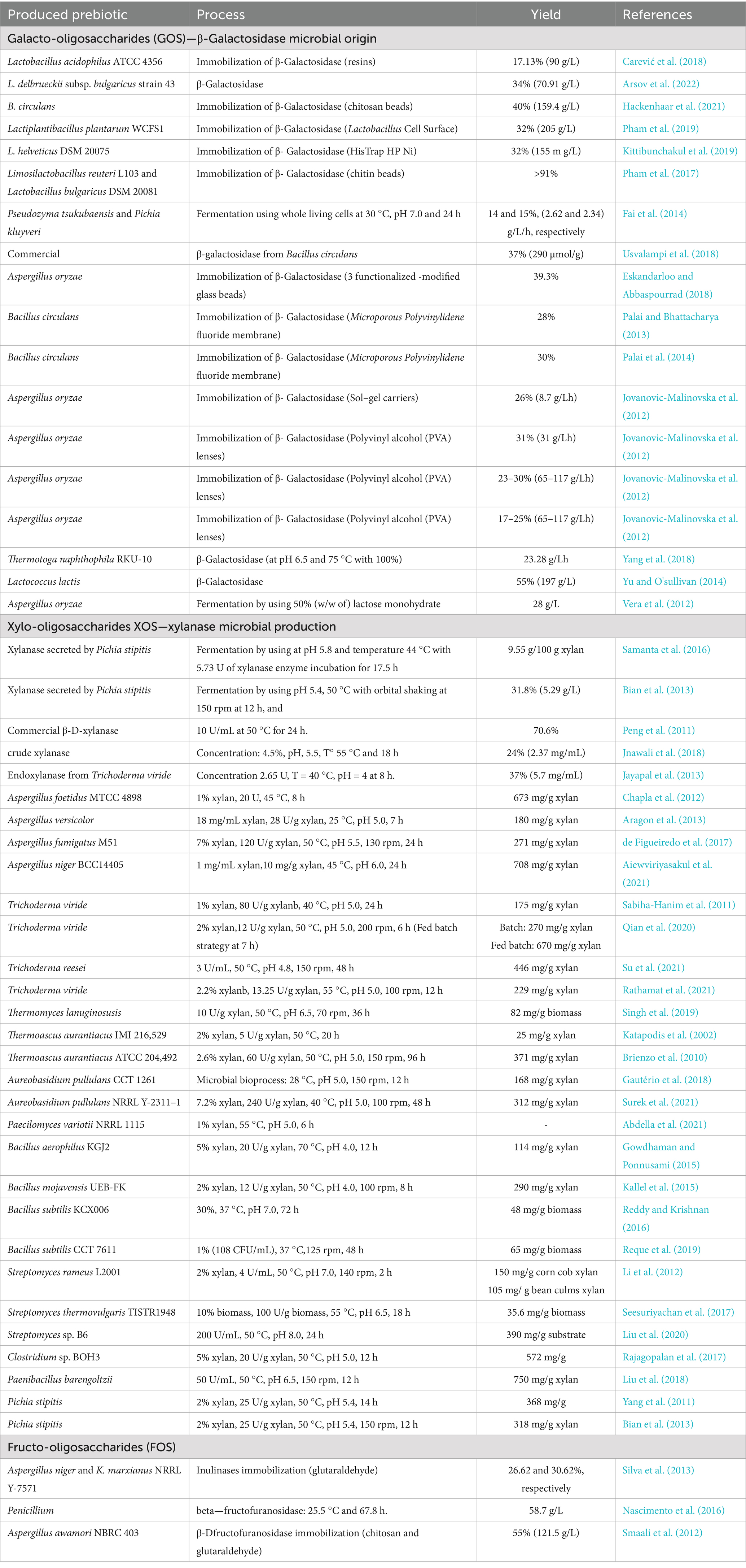
Table 2. Examples of major enzymatic produced prebiotic (GOS, XOS and FOS), including microorganisms, enzyme sources, processes and yields.
4 Synbiotics
The term Synbiotic, first defined in 1995 by Gibson and Roberfroid to refer to a combination of probiotics and prebiotics, describes a combination in which prebiotics enhance the activity of probiotics (Dahiya and Nigam, 2022; Ashique et al., 2024). At a meeting of the International Scientific Association for Probiotics and Prebiotics (ISAPP) in 2019, this phrase was revised to a mixture containing live microorganisms and substrate(s) that are selectively utilized by the host microorganisms and provide a health benefit to the host (Gibson et al., 2017; Swanson et al., 2020; Dahiya and Nigam, 2022). Synbiotics are classified into two forms. Complementary synbiotics consist of a combination of probiotic and prebiotic components that each independently meet minimum efficacy criteria; these components act independently to support host health. In comparison, synergistic synbiotics are defined as a system in which the coordinated interaction of selected probiotic microorganism strains and prebiotic substrates that specifically promote the growth and activity of these strains work together to provide targeted physiological benefits (Kleerebezem and Führen, 2024). Mechanically, synbiotics represent the combination of the biochemical properties of prebiotics and the functional effects of probiotics. While prebiotics selectively stimulate the growth of beneficial microbes, the presence of specific probiotic strains ensures direct interaction with the host. This synergistic interaction provides both metabolic and immunological benefits and creates a more comprehensive effect than either component alone (Swanson et al., 2020; Kolida and Gibson, 2011).
Studies show that synbiotic intake contributes to the alleviation of irritable bowel syndrome, metabolic syndrome, inflammatory bowel diseases, diarrhea and skin problems such as atopic dermatitis (Markowiak and Śliżewska, 2017). Wang et al. (2024), investigated whether the symptoms of Autism Spectrum Disorder (ASD) could be improved through modulation of gut microbiota. They treated mouse models with the disorder with a daily synbiotic treatment consisting of a combination of Limosilactobacillus reuteri (L. reuteri) and inulin for 4 weeks. They reported that this combination alleviated the social impairments associated with ASD, in part through its regulatory effects on the gut-brain axis (Wang et al., 2024). Palepu et al. (2024) investigated the effect of Faecalibacterium prausnitzii (ATCC-27766) in combination with fructo-oligosaccharides (FOS) and galacto-oligosaccharides (GOS) in 2024 treatment-resistant depression (TRD) rat models and suggested that this synbiotic may reverse TRD-like symptoms in rats by positively affecting gut health, neuroinflammation, neurotransmitters and gut microbial composition. Bonfili et al. (2017), supplemented the diet of 3xTg-AD Alzheimer’s disease mice with a red lentil (prebiotic) based cookie enriched with neuroprotective probiotics and performed behavioral, biochemical and molecular tests. They reported that short-term memory improved after the treatments and that the combined use was successful compared to the individual ingredients. Johnson et al. (2025), investigated the effect of dietary synbiotic supplementation on behavioral, neurochemical and microbial parameters in W36 Hy-Line laying hens, focusing on modulation of the microbiota-gut-brain (MGB) axis. Over a 10-week intervention, they found that in chickens receiving a synbiotic diet, agonistic behaviors, including pecking and fighting, were significantly reduced, while dopamine and serotonin levels were increased. They stated that MGB axis regulation contributed to improved welfare and reduced stress-induced behaviors. Ghorbani et al. (2018) aimed to evaluate the efficacy of a 6-week synbiotic supplement in the treatment of moderate depression and gave fluoxetine (20 mg/day) to all patients for 4 weeks. They then added a synbiotic capsule (plus fluoxetine) or placebo (plus fluoxetine) to the treatment for 6 weeks.
And they found the efficacy of the synbiotic as an adjuvant treatment for moderate depression. Shinde et al. (2020), wanted to develop dietary strategies to help address the increasing cases of inflammatory bowel diseases (IBD) and investigated the efficacy of green banana resistant starch (GBRS) and probiotic Bacillus coagulans (B. coagulans) MTCC5856 spores for improving dextran-sulfate sodium (DSS)-induced colitis in mice. They found that synbiotic supplementation with B. coagulans and GBRS improved the overall inflammatory status of the experimental IBD model through synergistic functioning. They recommended investigating the effect of this practice in reducing inflammation in human IBD. Polakowski et al. (2019), investigated the effect of preoperative synbiotic administration in colorectal cancer patients with colorectal resection. They randomized patients to receive synbiotics (Simbioflora, Farmoquimica, São Paulo, Brazil) or placebo (maltodextrin) 8 days before surgery. They found that 7 days of preoperative synbiotic use in colorectal cancer patients relieved the inflammatory state and reduced morbidity, length of hospital stay and antibiotic use.
When the above findings are evaluated, it is understood that synbiotics exhibit beneficial effects in various conditions, including gastrointestinal, metabolic, dermatological, and neuropsychiatric disorders. Despite the diversity of tested models and combinations, several mechanisms consistently emerge, such as modulation of the gut-brain axis via neurotransmitter regulation, alleviation of systemic and local inflammation, and improvement of gut microbial balance. However, several limitations must be acknowledged. Most studies are based on animal models, limiting translation to humans. Furthermore, existing clinical studies are generally short-term and have limited sample sizes. Moreover, heterogeneity in strain-substrate combinations and dosages makes direct comparisons between studies difficult and hinders the establishment of standardized protocols. Future research should prioritize conducting large-scale, long-term clinical studies to establish the clinical efficacy and safety of synbiotics.
5 Gut microbiota and brain communications
Probiotic research has expanded far beyond gut microbiota balance, revealing their roles in strengthening gut barrier integrity, regulating immune responses, producing bioactive compounds, and interacting directly with host cells, thus broadening their therapeutic potential (Piccioni et al., 2023). While well-established strains include Lactobacillus, Bifidobacterium, and Saccharomyces, emerging candidates such as Roseburia spp., Akkermansia spp., and Faecalibacterium spp. show potential for future applications (Sanders et al., 2019). Many of these functional roles are supported primarily by in vitro and animal model data, and while preclinical results are encouraging, translation into clinically validated outcomes remains inconsistent and strain-specific, as not all effects observed in experimental models are confirmed in humans, and even among clinical trials, heterogeneity in study design, dosage, strain selection, and host response complicates definitive conclusions. Additional limitations include the lack of long-term safety data, and potential interactions between strains or with the host that may diminish efficacy, and the need for personalized approaches to optimize benefits. Advances in genome sequencing, microbiota analysis, and real-time in vivo sampling are expected to help address these gaps, leading to a clearer understanding of their mechanisms and health benefits (Sanders et al., 2019).
Prebiotics are substances selectively utilized by host microorganisms to promote health benefits, including the modulation of gut microbiota and the production of beneficial metabolites such as short-chain fatty acids (SCFAs) and tryptophan (Galica et al., 2022). Initially known for stimulating the growth of bifidobacteria and lactobacilli, prebiotics are now recognized for their broader effects on metabolic and physiological systems, such as immune modulation, defense against pathogens, improved intestinal function, and enhanced mineral absorption (Piccioni et al., 2023). Common prebiotics, such as fructo-oligosaccharides (FOS), inulin, and resistant starches, are commercially available and contribute to optimizing the human microbial environment. However, while the mechanisms of action have been outlined through in vitro and animal models, confirming these effects in humans remains challenging (Sanders et al., 2019).
Synbiotics, a combination of probiotics and prebiotics, support the growth and activity of beneficial gut bacteria, promoting digestive health and strengthening immune function. By enhancing gut microbiota balance, they offer a comprehensive approach to improving overall well-being through dietary supplementation (Al-Habsi et al., 2024).
To clarify the specific effects of probiotics, prebiotics, and synbiotics on gut-brain axis and their mechanisms of action, Table 3 summarizes their health benefits and potential roles in various physiological processes. Figure 1 illustrates the bidirectional communication pathways of the gut-brain axis, showing how neuronal, endocrine, and immune signaling mediate interactions between the gut microbiota and the central nervous system. The gut microbiota modulates brain function through neuronal (vagal and enteric nervous system), endocrine (gut-derived hormones and HPA axis), and immune (cytokines, inflammation) pathways. In return, the brain influences gut physiology through stress, emotion, and autonomic regulation. Microbial metabolites such as SCFAs, neurotransmitters, and tryptophan derivatives act as mediators in this bidirectional dialog.
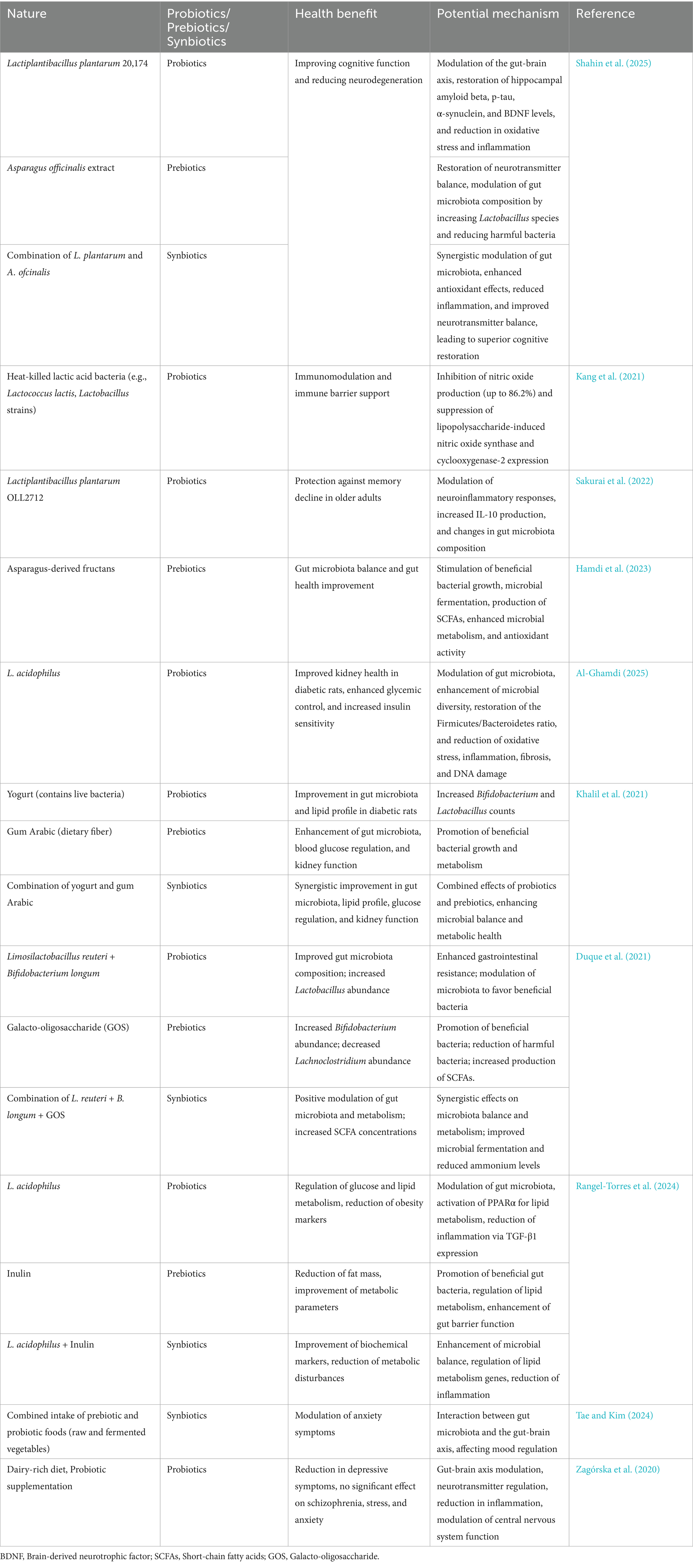
Table 3. Impact of probiotics, prebiotics, and synbiotics on the gut-brain axis.
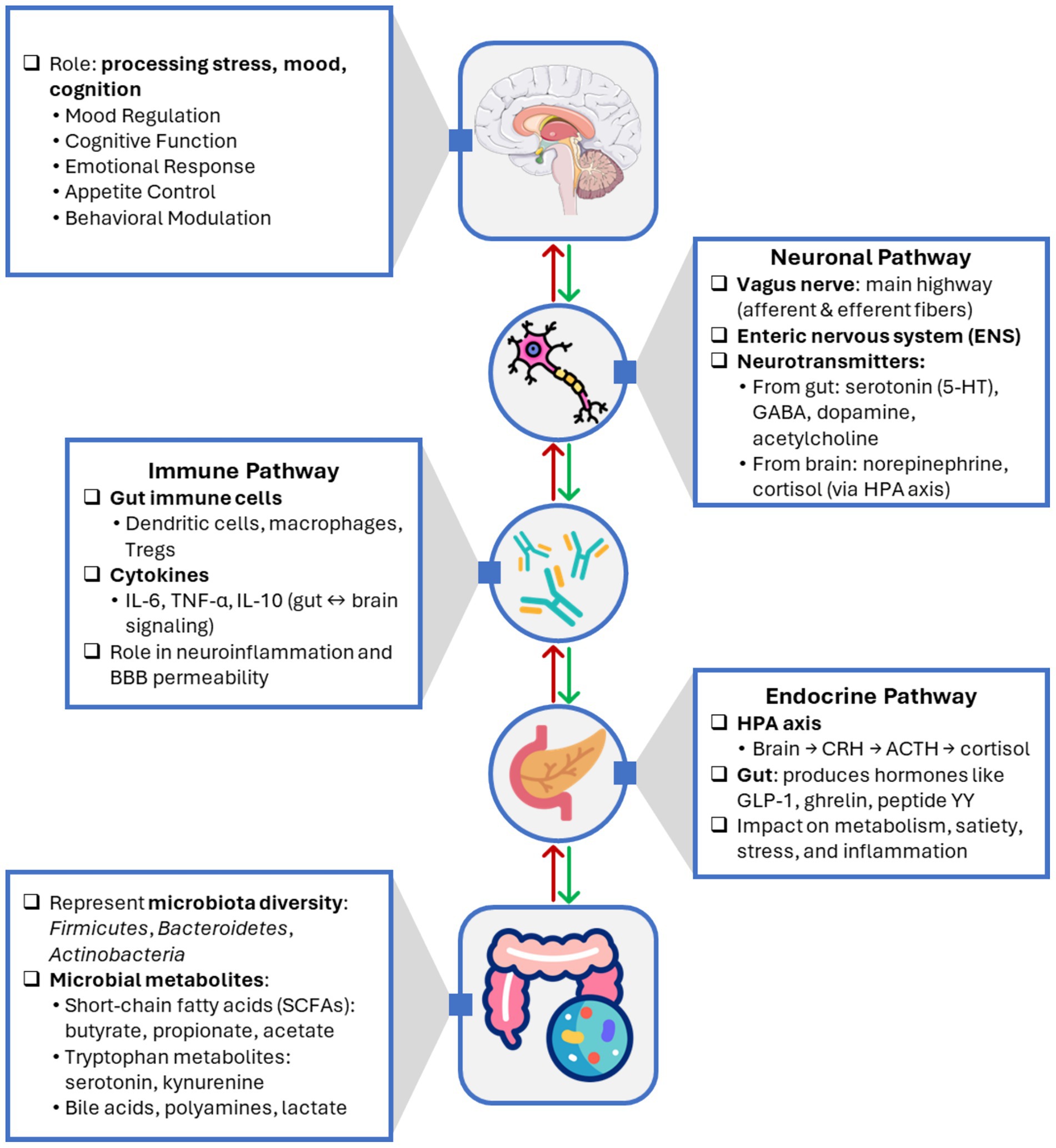
Figure 1. Bidirectional communication pathways of the gut-brain axis.
5.1 Effects on gut microbiota
5.1.1 Mechanisms of action
Probiotic strains influence gut microbiota through nutrient competition, antagonism, cross-feeding, and stability support. Lactobacillus and Bifidobacterium species produce lactic and acetic acids as key metabolites of carbohydrate fermentation, which lower luminal pH and inhibit pathogen growth. SCFAs, such as acetate, butyrate, and propionate, are generated through the colonic fermentation of dietary fiber and resistant starch. These SCFAs play a crucial role in anti-inflammatory pathways and signaling across various organs, contributing to overall host health (Markowiak-Kopeć and Śliżewska, 2020). Figure 2 exemplifies the modulatory effects of probiotics, prebiotics, and synbiotics on gut microbiota composition and function, highlighting their downstream impacts on gut health, immune response, metabolic regulation, and neurocognitive outcomes. These dietary interventions promote the growth of beneficial microbes while suppressing harmful species, enhancing short-chain fatty acid production, gut barrier integrity, immune tolerance, and neurotransmitter synthesis. The resulting microbial balance contributes to improvements in gut health, immune regulation, metabolic homeostasis, and cognitive and emotional function.
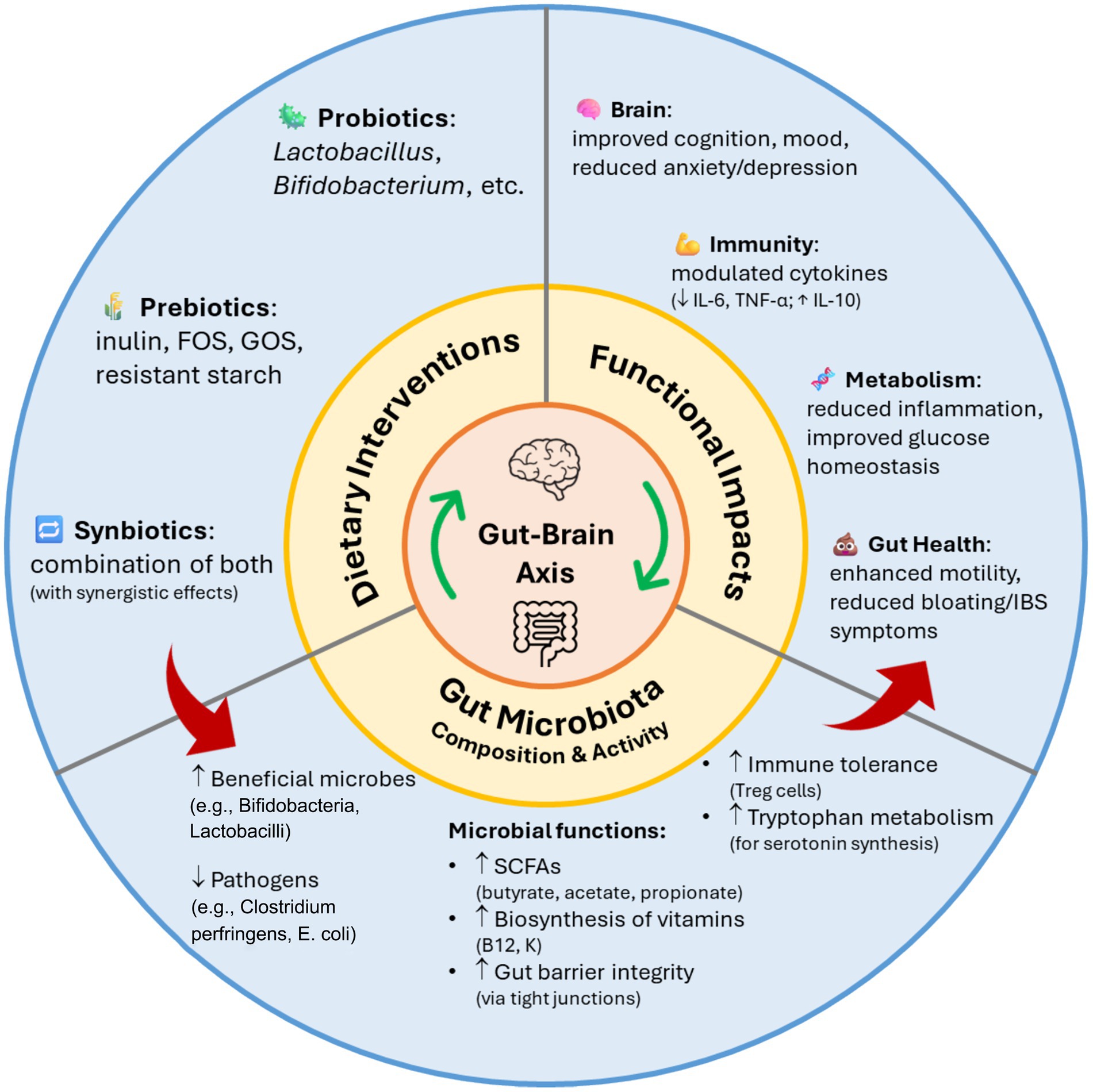
Figure 2. Impact of probiotics, prebiotics, and synbiotics on gut microbiota composition and function.
5.1.2 Key microbial metabolites and strain-spesific effects
Probiotic supplementation with Lactobacillus acidophilus, Lactiplantibacillus plantarum, Lacticaseibacillus rhamnosus, and Enterococcus faecium for 3 weeks resulted in colonization in both the luminal and mucosal compartments of the colon, increasing lactate levels in the proximal and distal regions. This promoted the growth of lactate-consuming bacteria, enhancing SCFA production, particularly butyrate (Moens et al., 2019). Lactiplantibacillus plantarum strains, commonly found in meat, dairy, fruits, and vegetables, are known to support gut microbiota, modulate immune function, lower blood cholesterol, and reduce cancer risk (Zare et al., 2024). Additionally, L. acidophilus supplementation improved kidney health in diabetic rats by enhancing gut microbiota diversity and restoring the Firmicutes/Bacteroidetes ratio. This led to better glycemic control, increased insulin sensitivity, reduced oxidative stress, and improved kidney structure with less inflammation and fibrosis, suggesting its potential in mitigating diabetes-related renal complications (Al-Ghamdi, 2025).
Many probiotics exert antagonistic effects by producing organic acids and bacteriocins through saccharolytic metabolism. These bacteriocins help inhibit pathogenic bacteria in the intestines and urinary tract while also contributing to gut homeostasis (Ballan et al., 2020).
5.1.3 Prebiotics and their impact
Prebiotics, such as inulin and polyphenols, are selectively fermented by gut microbiota, generating SCFAs and other beneficial metabolites. These compounds help lower intestinal pH, suppress pathogen growth, and enhance mineral and vitamin absorption (Ballan et al., 2020). Asparagus-derived fructans, similar to commercial fructans, promote beneficial bacterial growth (Hamdi et al., 2023). Despite having a lower degree of polymerization (DP up to 25), they undergo microbial fermentation, supporting gut balance and SCFA production. Additionally, their rich protein and phenolic content may enhance microbial metabolism and antioxidant activity, further benefiting gut health. Phenolic compounds also function as prebiotics by selectively modulating gut microbiota composition through dual mechanisms. They stimulate the growth of beneficial bacteria such as Lactobacillus, Bifidobacterium, and Akkermansia, enhancing the production of SCFAs, which support gut health. Simultaneously, they exert antimicrobial effects by disrupting the structural integrity of pathogenic bacteria (e.g., Clostridium, Staphylococcus, and Escherichia coli; Chiu et al., 2021). This occurs through inhibition of key bacterial proteins (PBP2 and PBP4), impairing peptidoglycan cross-linking, and altering membrane function via proton donation and H+-ATPase depletion, thereby creating an unfavorable environment for harmful microbes (Cano et al., 2024). The relationship between polyphenols and gut microbiota is bidirectional, as gut microbes also influence phenolic compound metabolism and bioavailability. Although preclinical studies suggest their potential in preventing and treating disorders and diseases, more clinical research is needed to confirm these benefits (Chiu et al., 2021).
5.1.4 Animal and human studies
Several studies have explored the effects of probiotics and prebiotics across different models. Martinez-Porchas et al. (2023) conducted a meta-analysis revealing minimal variations in the gut microbiota structure of tilapia exposed to feed additives (probiotics, prebiotics, and biofloc) across 221 samples from multiple studies. Despite the diversity of the datasets and potential methodological biases, this comprehensive analysis identified consistent core microbiota taxa, including Proteobacteria, Fusobacteria, Actinobacteria, Firmicutes, and Bacteroidetes, suggesting a resilient microbial community capable of adapting to dietary interventions without compromising host physiological function. These findings provide foundational insight for sustainable aquaculture practices that leverage microbial modulation. Likewise, Khalil et al. (2021) found that yogurt and gum Arabic supplementation improved gut microbiota composition in rats, enhancing beneficial bacteria such as Bifidobacterium and Lactobacillus. This change in microbiota was associated with better blood glucose and lipid control, highlighting the importance of gut health in managing diabetes. Using an in vitro gut microbiome model, Duque et al. (2021) investigated the combined effects of the probiotic strains Limosilactobacillus reuteri and Bifidobacterium longum with the prebiotic galacto-oligosaccharides (GOS) in children with autism spectrum disorder. The treatment enhanced gastrointestinal resistance, increased Lactobacillus abundance, and promoted Bifidobacterium growth while reducing potentially harmful genera such as Lachnoclostridium. Importantly, prebiotic and synbiotic interventions raised short-chain fatty acid concentrations and lowered ammonium levels, indicating a favorable shift in microbial metabolism with potential implications for gut-brain axis modulation in neurodevelopmental disorders. Finally, Qureshi et al. (2024) highlighted those prebiotics and probiotics regulate gut microbiota composition in individuals with obesity by increasing beneficial bacteria while reducing harmful microorganisms. This modulation helps alleviate gut dysbiosis, which is associated with inflammation and excessive fat accumulation. By restoring microbial balance, these therapies contribute to improved metabolic health and weight management.
5.1.5 Future tools like AI/ML
Machine learning enables large-scale analysis of gut microbes and prebiotic sources, allowing rational selection of substrates or synbiotic formulations based on genomic and metabolic predictions, without the need for preliminary in vitro tests. Additionally, they allow for a more precise assessment of microbiota composition and functional activity, such as bioactive metabolite production, paving the way for personalized nutrition and targeted therapeutic strategies (Sabater et al., 2021).
5.2 Effects on the nervous system
5.2.1 Neurodevelopmental disorders
The gut microbiota communicate with the nervous system through the vagus nerve, immune signaling, and endocrine pathways. Microbial metabolites such as SCFAs, serotonin, and lipopolysaccharides influence neurotransmission, neuroinflammation, and brain function. When gut permeability increases, they can cross the blood–brain barrier and affect neural activity (Nie et al., 2024).
Microbial modulation shows promise in neurological health. Probiotic and prebiotic supplementation, individually or in combination, has shown positive effects on autism-related behaviors and molecular markers. These treatments improved social interaction, anxiety, and repetitive behaviors, while also increasing anti-inflammatory IL-10 levels. Probiotics restored the Bacteroidetes/Firmicutes ratio and reduced IL-6 levels, while the combined treatment additionally increased 5-HT levels in the prefrontal cortex (Adıgüzel et al., 2022). Similarly, L. plantarum OLL2712 improved memory function in older adults by reducing inflammation-associated gut bacteria and enhancing cognitive scores (Sakurai et al., 2022). After 12 weeks, participants who consumed heat-treated OLL2712 exhibited significant improvements in composite memory and visual memory scores compared to the placebo group. Additionally, the gut microbiota of the active group showed a reduced abundance of inflammation-associated genera, including Lachnoclostridium, Monoglobus, and Oscillibacter. These results suggest that OLL2712 may mitigate memory decline by modulating gut microbiota and reducing neuroinflammation.
5.2.2 Cognitive function and memory
The impact of L. plantarum (probiotic), Asparagus officinalis extract (prebiotic), and their synbiotic combination on high-fat diet (HFD)-induced cognitive dysfunction and neurodegeneration in rats was investigated (Shahin et al., 2025). The findings indicate that these interventions effectively restored cognitive function and alleviated neurodegeneration by modulating key markers such as amyloid beta, p-tau, α-synuclein, and brain-derived neurotrophic factor (BDNF) in the hippocampus. Additionally, the treatments improved the disrupted lipid profile and mitigated oxidative stress, inflammation, and neurotransmitter imbalances. Notably, synbiotic treatment demonstrated superior effects by restoring gut microbiota balance, increasing beneficial Lactobacillus species, and reducing harmful bacteria (e.g., coliform and staphylococci), suggesting that the combined impact on gut health and brain function was more potent than the individual therapies. Flavonoids and phenolic compounds further support brain health by enhancing the production of key metabolites such as SCFAs, γ-aminobutyric acid (GABA), and BDNF. Certain Gram-positive bacteria, including Lactobacillus and Bifidobacterium spp., convert glutamate into GABA, a major inhibitory neurotransmitter. Additionally, phenolic compounds stimulate beneficial bacteria such as Streptococcus, Escherichia, and Enterococcus spp., promoting SCFA synthesis and neurotransmitter production, including serotonin via tryptophan metabolism, thereby influencing brain function through the gut-brain axis (Cano et al., 2024).
5.2.3 Mood and anxiety disorders
Tae and Kim (2024) analyzed the impact of prebiotic and probiotic food consumption on anxiety in 4317 adults and found that higher intake of these foods was linked to lower anxiety levels. However, prebiotic consumption was associated with higher anxiety in both men and women, while probiotic food intake significantly reduced anxiety in men. These results suggest that prebiotics and probiotics may influence the nervous system, particularly through the gut-brain axis, with gender differences in their effects.
According to Zhao et al. (2024), the gut microbiota influence mood disorders such as major depressive disorder and bipolar disorder through the microbe-gut-brain axis, a bidirectional communication system. Gut microbes can regulate brain function, impacting mental health. Therapeutic strategies such as probiotics, prebiotics, synbiotics, and fecal microbiota transplantation may help restore microbial balance and alleviate symptoms of these disorders.
Despite promising findings, current research on probiotics, prebiotics, and synbiotics in neurodevelopmental and neuropsychiatric disorders faces several limitations. These include the lack of strain-specific evidence, variability in host response due to genetic and environmental factors, and the scarcity of long-term, large-scale human clinical trials. Furthermore, most studies rely on small sample sizes and short intervention periods, making it difficult to draw definitive conclusions regarding efficacy and safety. Addressing these gaps will require standardized protocols, extended follow-up durations, and multi-center collaborations to ensure reproducibility and generalizability of results (Marco et al., 2021).
5.3 Effects on the immune barrier
The gut microbiota play a crucial role in systemic immunity by modulating cytokine production, regulating immune cell activity, and strengthening the intestinal barrier (Sivan et al., 2015). Certain probiotics further enhance immune defenses by stimulating phagocytosis, activating natural killer cells, and interacting with dendritic cells (Piccioni et al., 2023). They also boost antibody production, improve vaccine responses, and promote anti-inflammatory cytokine release, potentially reducing the risk of colon cancer and colitis. Additionally, gut microbes contribute to pathogen defense through competitive exclusion, antimicrobial compound production, and nutrient metabolism, impacting overall immunity and health (Cano et al., 2024).
While the exact mechanisms by which probiotics exert their immunomodulatory effects are not yet fully understood, several potential pathways have been proposed. Probiotics are believed to influence immune function through the inhibition of Toll-Like Receptors (TLRs), which play a central role in the recognition of microbial components and the activation of inflammatory responses. By downregulating TLR expression, probiotics can reduce the activation of inflammatory pathways such as NF-κB, which is involved in the transcription of pro-inflammatory cytokines (Plaza-Diaz et al., 2019). Additionally, probiotics may modulate the activity of innate immune cells, such as Natural Killer cells, enhancing their cytotoxic potential and improving immune surveillance (Kwok et al., 2022). Probiotic supplementation has also been shown to impact oxidative stress markers, reducing oxidative damage and improving the balance between antioxidants and oxidants. By modulating factors such as nitric oxide and C-reactive protein, probiotics help mitigate the risk of inflammatory diseases, cardiovascular dysfunction, and metabolic disorders. These mechanisms highlight the potential of probiotics to regulate immune responses, although further research is needed to clarify strain-specific effects and optimal intervention strategies (Kwok et al., 2022).
While many studies report the general immunomodulatory benefits of probiotics, it is important to emphasize the significant strain specificity in their effects on immunity. Variability in host response due to genetic, environmental, and lifestyle factors further complicates the translation of findings. Moreover, there is a notable lack of long-term human clinical trials assessing safety, efficacy, and optimal dosing regimens of specific probiotic strains in diverse populations. Addressing these research gaps is crucial for advancing the clinical application of probiotics in immune-related conditions and for developing personalized probiotic therapies tailored to individual immune profiles (Kim et al., 2014; Sempach et al., 2024). Application-based studies have demonstrated the immune-enhancing potential of specific probiotic strains. For instance, L. plantarum supplementation in mice enhanced immune organ activity, modulated immune cell populations, and increased antimicrobial substances and immunoglobulin levels (Sivan et al., 2015). Additionally, this strain strengthens mucosal immunity while maintaining immune homeostasis, making it a promising antigen delivery carrier. It enhances antigen immunogenicity, boosts defense against harmful antigens, and has been recognized for its potential as a mucosal vaccine carrier due to its ability to modulate immune tolerance (Zare et al., 2024). Similarly, Kang et al. (2021) observed that heat-killed Lactococcus lactis MG5125 and various Lactobacillus strains suppressed nitric oxide production by up to 86.2% and reduced the expression of nitric oxide synthase and cyclooxygenase-2 induced by lipopolysaccharides. This suggests that heat-killed probiotics may offer a stable alternative to live probiotics in functional foods, while still modulating immune responses effectively.
Early-life gut microbiota composition influences allergy development. Intestinal dendritic cells regulate Treg cells, which are linked to immune tolerance. Additionally, oligosaccharide supplementation has alleviated atopic dermatitis symptoms in children, with improvements associated with changes in peripheral eosinophil levels, highlighting the immunomodulatory role of prebiotics (Kim et al., 2024). Similarly, probiotics play a crucial role in immune regulation beyond early childhood, including in physically active individuals. In athletes, probiotic supplementation has been shown to influence immune regulation in several ways. Tavakoly et al. (2021) demonstrated that probiotics modulate key immune cell populations, including reductions in T cytotoxic lymphocytes and monocytes, while multi-strain formulations increase leukocyte counts. Complementing these findings, Guo et al. (2022) reported that probiotics enhance immune defense by increasing IFN-γ and salivary IgA levels while reducing TNF-α and IL-10, particularly in short interventions. The absence of significant effects on other inflammatory markers suggests that probiotics selectively regulate immune responses, highlighting their potential role in optimizing immune function in athletes. These findings underscore the importance of selecting appropriate strains and tailoring interventions based on the target population and desired immune outcomes.
5.4 Metabolites and neurotransmitters
A series of microbial metabolites have been implicated in the regulation of brain function, including branched-chain amino acids, trimethylamine-N-oxide, short-chain fatty acids, tryptophan metabolites, gamma-aminobutyric acid, bile acid metabolites and choline (Meher et al., 2024). Their mode of action is principally indirect, for example by improving intestinal health, exerting anti-inflammatory effects and modulating the production of metabolites such as serotonin, leptin and insulin that affect brain function. However, they may also have a direct effect, for example through the activation of aryl hydrocarbon receptor that takes place through the production of indoles (Aoki et al., 2018; Pappolla et al., 2021). Research has mainly focused on short-chain fatty acids, tryptophan metabolites, and ghrelin as well as the impact of probiotic, prebiotic and symbiotic supplementation on their production. These compounds are important due to their protective effects against obesity, depression, anxiety, colitis, atopic dermatitis and cancer. Table 4 presents recent studies demonstrating these protective effects. These compounds also mediate gut-brain axis communication via neuronal, endocrine, and immune pathways, engaging diverse microbial, neural, hormonal, and immune mediators (Table 5). These mechanisms enable bidirectional signaling between the gut microbiota and the brain, influencing mood, cognition, behavior, and neurological health.
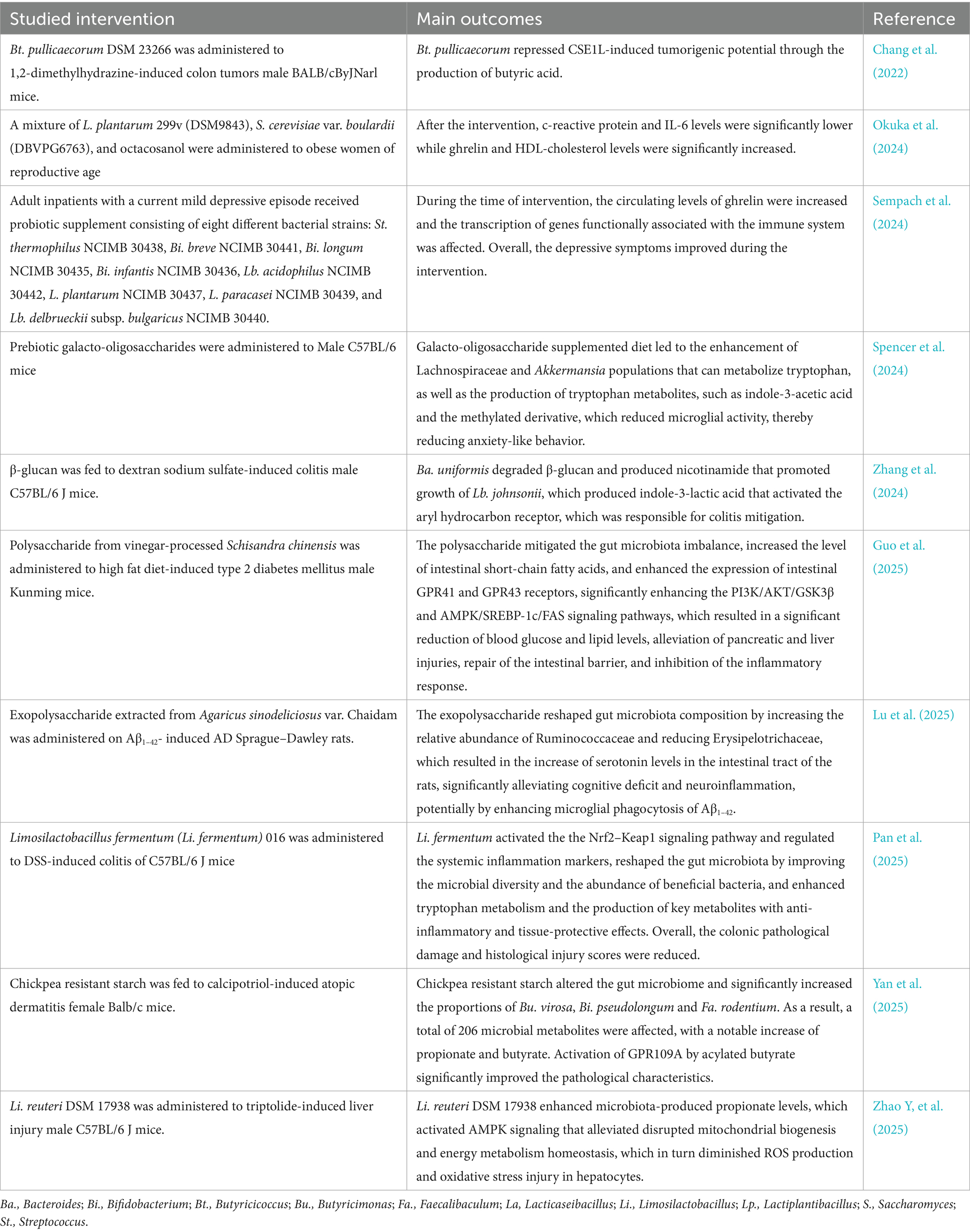
Table 4. Recent studies highlighting the protective effects of short-chain fatty acids, tryptophan metabolites and ghrelin against adverse health situations.
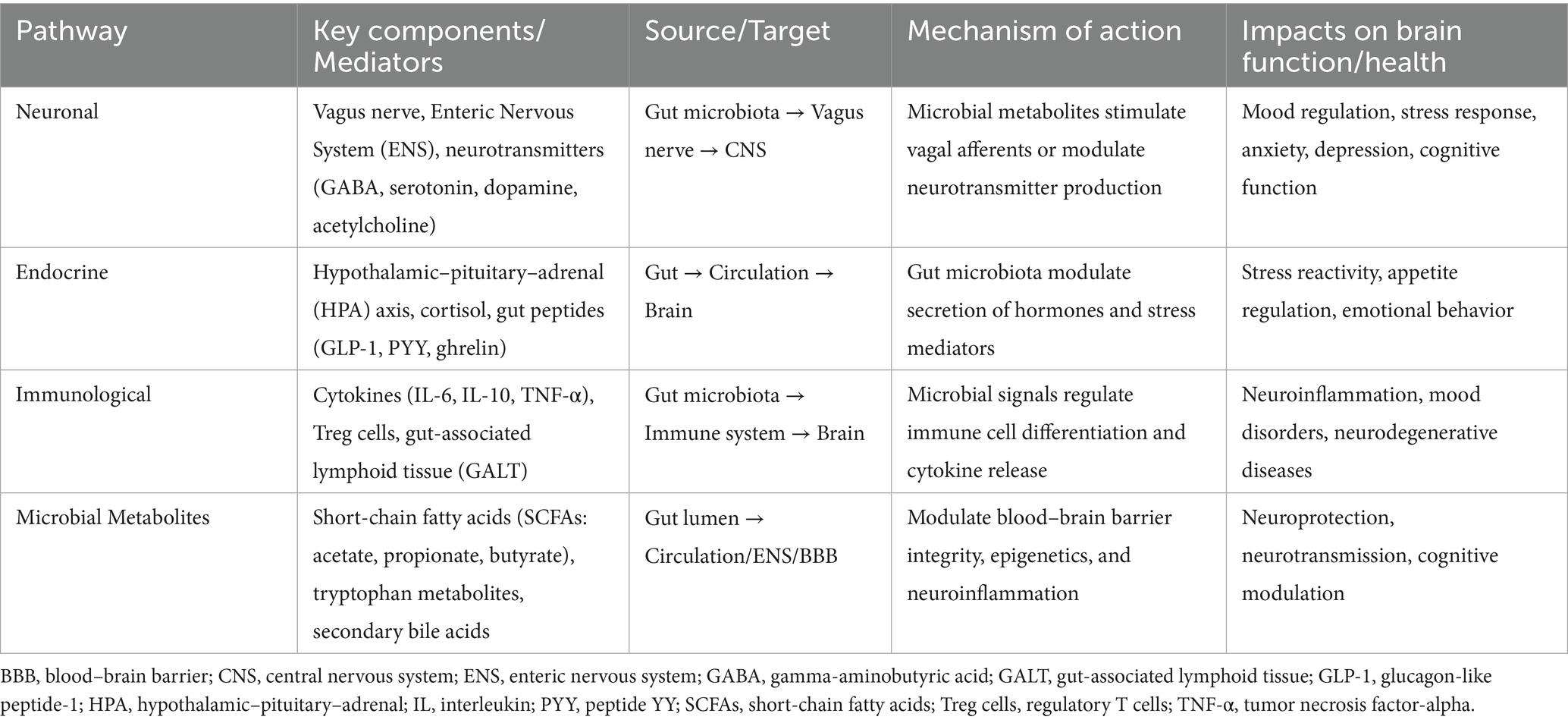
Table 5. Key pathways of gut-brain axis communication.
5.5 Short-chain fatty acids
Short-chain fatty acids (SCFA) are microbial metabolites with fewer than six carbon atoms, produced in the colon. The most represented ones are acetate, propionate and butyrate; formate and lactate are also produced, but at lower quantities. In the cecum and the proximal colon there is an increased availability of fermentable substrates compared to the distal colon. These substrates include resistant starch and components of plant cell walls, which have escaped digestion in the small intestine. Thus, the concentration of SCFA tends to be higher in the proximal colon and is depleted toward the distal colon (van der Beek et al., 2017). SCFA concentrations are estimated at 70 and 140 mM in the proximal colon and 20–70 mM in the distal colon (Wong et al., 2006). Their ratio along the colon is similar, namely 3:1:1 (acetate:propionate:butyrate), which reflects their efficient and concentration-dependent absorption (Topping and Clifton, 2001). The factors that affect qualitatively and quantitatively the production of SCFA are the ones that affect the composition of the microecosystem and the metabolic activity of the producer microorganisms, such as diet (Flint et al., 2015; Garcia-Mantrana et al., 2018; Selma-Royo et al., 2019; Fusco et al., 2023; Yi et al., 2025), gut transit time (Wong et al., 2006) pH value (Wong et al., 2006; Yi et al., 2025) and bile salt concentration (Flint et al., 2015).
The pathways for acetate production, namely acetogenesis and the Wood-Ljungdahl pathway, seem to be widely distributed among the phyla that comprise the human gut microbiome. On the contrary, production of butyrate and propionate is substrate specific, and the respective pathways seem to be restricted to a few species. Butyrate is mainly produced through the CoA-transferase pathway and only a few species use the butyrate kinase pathway (Flint et al., 2015). Butyrate production via the CoA-transferase pathway is mainly driven by Eubacterium hallii, Eubacterium rectale, Faecalibacterium prausnitzii, Roseburia faecis, and other Lachnospiraceae species (Louis et al., 2010; Reichardt et al., 2014). On the other hand, the occurrence of the butyrate kinase pathway has only been reported in Coprococcus eutactus and Coprococcus comes (Reichardt et al., 2014). Interestingly, the presence and metabolic activity of Ruminococcus bromii is particularly important when resistant starch is available (Ze et al., 2012). On the other hand, propionate can be produced by three pathways, the succinate pathway used by Bacteroidetes and some Firmicutes, the propanediol pathway that operates only when fucose and rhamnose serve as carbon sources and is used by some members of the Lachnospiraceae family, and the acrylate pathway that is restricted to only a few members of the Firmicutes and is used to convert lactate to propionate (Reichardt et al., 2014; Flint et al., 2015).
As far as formate and lactate are concerned, the first is mainly produced by bifidobacteria and Eubacterium hallii (Schwab et al., 2017) while the second mainly by bifidobacteria, lactobacilli, streptococci and staphylococci (Jost et al., 2012; Pham et al., 2016). The lactate is then catabolized by bacteria such as Eubacterium hallii and Anaerostipes caccae toward the production of propionate and butyrate (Reichardt et al., 2014).
SCFA are considered as possible mediators of the communication between gut microbiota and the brain (Dalile et al., 2019). SCFA produced in the colon are rapidly absorbed by the colonocytes and used for energy production (Schonfeld and Wojtczak, 2016). The ones that are not catabolized in the colonocytes are transported to the liver, where they are used for energy production by the hepatocytes, with the exception of propionate that can also be used for gluconeogenesis and acetate that can also be used to produce fatty acids and cholesterol (Boets et al., 2017). As a result, only a small percentage of the SCFA produced in the colon can reach peripheral organs through systemic circulation, which has been calculated at 2, 9 and 36% for butyrate, propionate and acetate, respectively (Boets et al., 2015). SCFA modulate brain function through immune, endocrine and vagal pathways (Dalile et al., 2019). The interactions of SCFA with a variety of immune cells may modulate brain activity. More specifically, SCFA directly affect neutrophils by regulating the production of inflammatory cytokines and by acting as neutrophil chemoattractants (Rodrigues et al., 2016), inhibit the maturation of monocytes, macrophages and dendritic cells (Correa-Oliveira et al., 2016; Chang et al., 2014) and affect the differentiation and proliferation of T cells (Kim et al., 2014). Although a variety of mechanisms have been proposed, the inhibition of histone deacetylases appears to play a key role. The endocrine pathway is activated by the secretion of gastrointestinal as well as other metabolic hormones, which is affected by colonic SCFA. The production of SCFA in the colon activates orphan G protein-coupled receptors which results in the production of peptide tyrosine tyrosine (PYY) and glucagon-like peptide 1 (GLP1) by enteroendocrine L cells (Tolhurst et al., 2012; Larraufie et al., 2018). On the other hand, there are indications that the production of hormones such as leptin, ghrelin and insulin is modulated by colonic SCFA (Xiong et al., 2004; Robertson et al., 2005; Rahat-Rozenbloom et al., 2017); however, the underlying mechanism is yet to be fully elucidated. The mechanisms by which these hormones affect brain function have been extensively assessed, and especially in the case of PYY and GLP1, have already been proposed (Koda et al., 2005; Katsurada and Yada, 2016). In the case οf ghrelin, SCFA have been reported to interfere with ghrelinergic signaling, most likely by antagonistic binding to its receptor GHSR-1a (Torres-Fuentes et al., 2019). Indications that SCFA stimulate the vagal afferents have been repeatedly reported (Bercik et al., 2011; Bravo et al., 2011; Goswami et al., 2018). This activation may be mediated by the free fatty acid receptor 3 (FFAR3; Bonaz et al., 2018) that is expressed in nodose ganglion neurons (Nohr et al., 2015).
The aforementioned interactions of SCFA are considered as the main mechanisms through which they contribute to the reduction of symptom severity or treatment of gastrointestinal, metabolic, cardiovascular, neurological and other disorders that have been associated with the gut-brain axis (Xiong et al., 2022; Zhang et al., 2023; Facchin et al., 2024).
5.6 Tryptophan and metabolites
Tryptophan is an essential amino acid; therefore, humans rely on dietary intake. Dietary tryptophan is mainly used for protein synthesis. Free tryptophan, i.e., the tryptophan that is not used for protein synthesis is mainly catabolized through the kynurenine pathway to produce a wide range of biologically active metabolites, collectively termed kynurenines. Over 95% of free tryptophan catabolism occurs via this pathway (Polyzos and Ketelhuth, 2015). The kynurenines have wide physiological and often opposing roles that are essential in immune responses, inflammation, oxidative stress and neurodegeneration, affecting, thus, brain function (Tanaka et al., 2024). Tryptophan may also be used for the biosynthesis of serotonin and melatonin, the modulation of the brain function by both has been well documented (Carhart-Harris and Nutt, 2017; Lee et al., 2019). Finally, the gut microbiota may use tryptophan for the production of indoles and their derivatives. Colonic microbiota may shift from saccharolytic to proteolytic metabolism-depending on protein intake, carbohydrate availability, transit time, and pH-leading to protein degradation and tryptophan catabolism. This catabolic shift has been reported as more intense toward the distal colon (Smith and Macfarlane, 1996; Geypens et al., 1997; Zelante et al., 2013; Roager et al., 2016; Vieira-Silva et al., 2016). The capacity of several Gram-positive and -negative species to catabolize tryptophan and produce indoles and their derivatives has been reported; most of them belong to the genera Anaerostipes, Bacteroides, Bifidobacterium, Butyrivibrio, Clostridium, Desulfovibrio, Enteroroccus, Escherichia, Eubacterium, Faecalibacterium, Fusobacterium, Haemophilus, Lactobacillus sensu lato, Megamonas, Parabacteroides, Peptostreptococcus and Ruminococcus (Roager and Licht, 2018).
The gut microbiota may affect directly or indirectly tryptophan metabolism by the host. The direct effect may result from the reduction of tryptophan availability for the host, which may lead to decreased serotonin and 5-hydroxyindoleacetic acid production, which, in turn, may lead to depressive-like behavior (Lukic et al., 2019). Interestingly, a causal link has been suggested, as depressive phenotypes can be transferred via gut microbiota transplantation (Kelly et al., 2016). Serotonin production may also be modulated, either toward stimulation that has been reported to occur by spore-forming bacteria including Clostridium ramosum (Yano et al., 2015; Mandic et al., 2019), or toward disruption (Golubeva et al., 2017). Similarly, modulation of the kynurenine pathway may also take place, as in the case of Lactobacillus johnsonii N6.2, which reduced the production of indoleamine-2,3-deoxygenase that catalyzes the oxidation of L-tryptophan to N-formylkynurenine, the first step of the kynurenine pathway (Valladares et al., 2013). The indirect effect has been reported to occur either through butyrate production, which has been reported to suppress kynurenine production (Martin-Gallausiaux et al., 2018), or through the maintenance of gut integrity that may prevent gastrointestinal disorders, such as inflammatory bowel disease and irritable bowel syndrome, which have been associated with disruption of the serotonergic signaling pathways (Gracie et al., 2019). Maintenance of gut integrity can be achieved through a number of mechanisms including the promotion of cytokine release, such as IL-6, IL-17 and IL-22, by indole derivatives through the activation of the aryl hydrocarbon receptor (Zelante et al., 2013; Schiering et al., 2017; Busbee et al., 2020; Zhang et al., 2024).
These direct and indirect effects of gut microbiota on tryptophan metabolism by the host regulate intestinal and systemic homeostasis in both health and disease (Zhao P, et al., 2025). More specifically, the development of many diseases including digestion, respiratory, blood, neoplastic and non-neoplastic ones has been associated with disruption of tryptophan metabolism. Therefore, the therapeutic potential of restoration of tryptophan metabolism has been indicated (Platten et al., 2019; Chen et al., 2024).
5.7 Ghrelin
Ghrelin is a 28 amino acid hormone primarily produced in the stomach. The acylated form of ghrelin binds with high affinity to the growth hormone secretagogue receptor (GHSR), and more specifically GHSR-1a. This receptor is ubiquitously expressed in central and peripheral nervous system and has been implicated in the regulation of an extended array of functions related to feeding behavior, metabolism and energy storage. Therefore, it is considered as a key molecule that communicates nutrition-related information along the gut-brain axis (Leeuwendaal et al., 2021).
Studies report both positive and negative correlations between gut microbiota and ghrelin levels (Parnell and Reimer, 2012; Hooda et al., 2013; Gomez-Arango et al., 2016; Kang et al., 2016; Liu et al., 2017; Massot-Cladera et al., 2017; Yanagi et al., 2017; Yang et al., 2019; Bo et al., 2019) suggesting a regulatory relationship (Mahana et al., 2016; Ikenoya et al., 2018). More specifically, ghrelin levels seem to be affected by the lipopolysaccharides of Gram-negative bacteria, as well as by metabolites such as formylated peptides, amino acids, hydrogen sulfide and SCFA. The first has been adequately exhibited in the case of Helicobacter pylori, whose lipopolysaccharide seems to activate an inflammatory response through TRL-4 stimulation and ghrelin-mediated GHSR-1a activation (Slomiany and Slomiany, 2017). Similarly, formylated peptides may also have an indirect effect on ghrelin levels as they activate the epithelial GPCR formyl peptide receptor 1 (FPR1) stimulating ROS generation by epithelial cells, which in turns increases plasma ghrelin concentration (Suzuki et al., 2011; Alam et al., 2014). Microbial proteolysis of dietary proteins produces amino acids that affect plasma ghrelin levels in a residue-specific manner. More specifically, L-glutamine, L-glutamic acid, L-lysine, L-threonine and L-valine increase ghrelin plasma levels while L-cysteine, L-leucine and L-tryptophan reduces them (McGavigan et al., 2015; Steinert et al., 2017; Elsabagh et al., 2018; Yin et al., 2018a, 2018b). L-cysteine holds an additional role as its degradation is the major pathway for hydrogen sulfide production. The latter has been reported to negatively affect the ghrelin secretion (Slade et al., 2018). A negative correlation has also been reported between SCFAs and ghrelin levels (Rahat-Rozenbloom et al., 2017). Two mechanisms have been proposed, a direct that includes antagonism for the GHSR-1a receptor and an indirect that includes FFAR2-mediated regulation (Torres-Fuentes et al., 2019).
6 Current challenges and future directions
Although evidence on probiotics, prebiotics, and synbiotics in modulating the gut-brain axis (GBA) is growing, key challenges still hinder the translation of preclinical findings into clinical practice. First, heterogeneity in dosage, duration, and strain selection complicates interpretation and undermines reproducibility. Second, many studies are conducted in small, homogeneous populations, reducing the generalizability of findings across diverse age groups, lifestyles, and clinical conditions. Third, although animal studies provide valuable mechanistic insights, their predictive value for human physiology and neurocognitive outcomes remains limited. Moreover, findings are inconsistent: some studies show clear benefits while others report null or adverse outcomes, fueling ongoing debate.
Another critical limitation is the lack of standardized biomarkers and validated clinical endpoints for assessing GBA-related benefits. Current measures mainly depend on subjective self-reports or indirect proxies, which fail to fully capture the multidimensional nature of gut-brain interactions. In addition, the long-term safety and efficacy of chronic probiotic or synbiotic supplementation are still unclear, as most trials are of relatively short duration.
Future research should prioritize large-scale, well-controlled, and multi-center trials with clearly defined outcomes to establish clinical relevance. Integrating omics technologies, such as metabolomics, metagenomics, and transcriptomics, may provide a systems-level understanding of host-microbiome interactions. Furthermore, there is a need to explore personalized approaches, as individual differences in microbiome composition, genetics, and lifestyle factors likely influence responsiveness to interventions. Addressing these challenges is essential to translate promising findings into evidence-based strategies for brain and systemic health.
7 Conclusion
Studies confirm that the gut microbiota play an essential function in regulating the two-way communication between the gastrointestinal tract and the central nervous system, commonly referred to as the gut-brain axis. Probiotics, prebiotics, and synbiotics modulate the gut-brain axis by influencing microbial composition, metabolic activity, immune responses, and neurochemical pathways. Studies suggest that their inclusion in the diet may alleviate clinical manifestations of various neurological and psychiatric disorders, enhance cognitive function and improve systemic immune function. However, key issues must be resolved before clinical implementation. Upcoming research must give priority to clarifying how hosts and microbiota interact, variability between microbial strains and substrates, the influence of individual genetic and environmental factors, and standardized clinical trials should be performed. Large-scale randomized trials integrating multi-omics approaches are needed to clarify the functional effects of these interventions. In addition, personalized approaches that take into account individual variability in microbiota composition, genetics, diet and lifestyle are important to maximize therapeutic efficacy and minimize adverse outcomes. It is also important to focus on optimizing microbial formulations and defining precise therapeutic windows for different disease states. Additionally, emerging technologies such as machine learning and systems biology will open up different opportunities to predict host responses and design tailored microbial therapies. Moreover, the development of next-generation probiotics and designer synbiotics targeting specific pathways involved in neuroinflammation, neurotransmitter synthesis and immune modulation represents a promising direction for future research. In conclusion, regulating the gut-brain axis is a promising strategy for managing neurological, psychiatric, and systemic diseases. Interdisciplinary research and technological advances are crucial for translating findings into clinical practice.
Author contributions
GK: Conceptualization, Data curation, Visualization, Writing – original draft, Writing – review & editing. SP: Data curation, Writing – review & editing. KK: Data curation, Writing – review & editing. IH: Data curation, Writing – review & editing. MČ: Data curation, Writing – review & editing. VŠ: Data curation, Writing – review & editing. SS: Data curation, Writing – review & editing. WE: Data curation, Writing – review & editing. FO: Conceptualization, Project administration, Supervision, Writing – review & editing. TE: Conceptualization, Funding acquisition, Project administration, Supervision, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was supported by the Scientific and Technological Research Council of Türkiye (TUBITAK); grant no: 123 N064 (PRIMA Programme Section 2) and by the PRIMA program under project InnoSol4Med (Project ID 1836). The PRIMA programme is supported by the European Union. The publication of this article was funded by the open access fund of Leibniz Universität Hannover.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abdella, A., Ramadan, S., Hamouda, R. A., Saddiq, A. A., Alhazmi, N. M., and Al-Saman, M. A. (2021). Paecilomyces variotii xylanase production, purification and characterization with antioxidant xylo-oligosaccharides production. Sci. Rep. 11:16468. doi: 10.1038/s41598-021-95965-w
Adıgüzel, E., Çiçek, B., Ünal, G., Aydın, M. F., and Barlak-Keti, D. (2022). Probiotics and prebiotics alleviate behavioral deficits, inflammatory response, and gut dysbiosis in prenatal VPA-induced rodent model of autism. Physiol. Behav. 256:113961. doi: 10.1016/j.physbeh.2022.113961
Adsul, M. G., Bastawde, K. B., and Gokhale, D. V. (2009). Biochemical characterization of two xylanases from yeast Pseudozyma hubeiensis producing only xylooligosaccharides. Bioresour. Technol. 100, 6488–6495. doi: 10.1016/j.biortech.2009.07.064
Aiewviriyasakul, K., Bunterngsook, B., Lekakarn, H., Sritusnee, W., Kanokratana, P., and Champreda, V. (2021). Biochemical characterization of xylanase GH11 isolated from Aspergillus niger BCC14405 (XylB) and its application in xylooligosaccharide production. Biotechnol. Lett. 43, 2299–2310. doi: 10.1007/s10529-021-03202-1
Akpinar, O., Ak, O., Kavas, A., Bakir, U., and Yilmaz, L. (2007). Enzymatic production of xylooligosaccharides from cotton stalks. J. Agric. Food Chem. 55, 5544–5551. doi: 10.1021/jf063580d
Alam, A., Leoni, G., Wentworth, C. C., Kwal, J. M., Wu, H., Ardita, C. S., et al. (2014). Redox signaling regulates commensal-mediated mucosal homeostasis and restitution and requires formyl peptide receptor. Mucosal Immunol. 7, 645–655. doi: 10.1038/mi.2013.84
Al-Ghamdi, S. B. (2025). Probiotics as renal guardians: modulating gut microbiota to combat diabetes-induced kidney damage. Biology 14:122. doi: 10.3390/BIOLOGY14020122
Al-Habsi, N., Al-Khalili, M., Haque, S. A., Elias, M., Al Olqi, N., and Al Uraimi, T. (2024). Health benefits of prebiotics, probiotics, synbiotics, and postbiotics. Nutrients 16:3955. doi: 10.3390/NU16223955
Ali, S., Hamayun, M., Siraj, M., Khan, S. A., Kim, H. Y., and Lee, B. (2025). Recent advances in prebiotics: classification, mechanisms, and health applications. Future Foods 12:100680. doi: 10.1016/j.fufo.2025.100680
Aljohani, A., Rashwan, N., Vasani, S., Alkhawashki, A., Wu, T. T., Lu, X., et al. (2024). The health benefits of probiotic Lactiplantibacillus plantarum: a systematic review and meta-analysis. Probiotics Antimicrob. Proteins. doi: 10.1007/s12602-024-10287-3
Allen, A. P., Hutch, W., Borre, Y. E., Kennedy, P. J., Temko, A., Boylan, G., et al. (2016). Bifidobacterium longum 1714 as a translational psychobiotic: modulation of stress, electrophysiology and neurocognition in healthy volunteers. Transl. Psychiatry 6:e939. doi: 10.1038/tp.2016.191
EFSA BIOHAZ Panel Allende, A., Alvarez-Ordóñez, A., Bover-Cid, S., Chemaly, M., and De Cesare, A. (2025). Protocol for extensive literature search (ELS) for the maintenance and update of list of QPS-recommended biological agents. Zenodo. doi: 10.5281/zenodo.3607188
Alli, S. R., Gorbovskaya, I., Jonathan Kolla, N. J., Brown, L., and Müller, D. J. (2022). The gut microbiome in depression and potential benefit of prebiotics, probiotics and synbiotics: a systematic review of clinical trials and observational studies. Int. J. Mol. Sci. 23:4494. doi: 10.3390/ijms23094494
Amorim, C., Silvério, S. C., Prather, K. L., and Rodrigues, L. R. (2019). From lignocellulosic residues to market: production and commercial potential of xylooligosaccharides. Biotechnol. Adv. 37:107397. doi: 10.1016/j.biotechadv.2019.05.003
Ansari, F., Neshat, M., Pourjafar, H., Jafari, S. M., Samakkhah, S. A., and Mirzakhani, E. (2023). The role of probiotics and prebiotics in modulating of the gut-brain axis. Front. Nutr. 10:1173660. doi: 10.3389/fnut.2023.1173660
Aoki, R., Aoki-Yoshida, A., Suzuki, C., and Takayama, Y. (2018). Indole-3-pyruvic acid, an aryl hydrocarbon receptor activator, suppresses experimental colitis in mice. J. Immunol. 201, 3683–3693. doi: 10.4049/jimmunol.1701734
Aragon, C. C., Santos, A. F., Ruiz-Matute, A. I., Corzo, N., Guisan, J. M., Monti, R., et al. (2013). Continuous production of xylooligosaccharides in a packed bed reactor with immobilized–stabilized biocatalysts of xylanase from aspergillus versicolor. J. Mol. Catal. B Enzym. 98, 8–14. doi: 10.1016/j.molcatb.2013.09.017
Arsov, A., Ivanov, I., Tsigoriyna, L., Petrov, K., and Petrova, P. (2022). In vitro production of galactooligosaccharides by a novel β-galactosidase of Lactobacillus bulgaricus. Int. J. Mol. Sci. 23:14308. doi: 10.3390/ijms232214308
Ashique, S., Mohanto, S., Ahmed, M. G., Mishra, N., Garg, A., Chellappan, D. K., et al. (2024). Gut-brain axis: a cutting-edge approach to target neurological disorders and potential synbiotic application. Heliyon 10:e34092. doi: 10.1016/j.heliyon.2024.e34092
Bagdadi, B., Alqazlane, A., Alotaibi, M., Alamoudi, A., Baghdadi, L., Mohammad Mahmood, A., et al. (2025). The effectiveness of probiotics or synbiotics in the prevention and treatment of diarrhea among critically ill adults: a systematic review and meta-analysis. Clin. Nutr. ESPEN 65, 218–226. doi: 10.1016/j.clnesp.2024.11.025
Ballan, R., Battistini, C., Xavier-Santos, D., and Saad, S. M. I. (2020). Interactions of probiotics and prebiotics with the gut microbiota. Prog. Mol. Biol. Transl. Sci. 171, 265–300. doi: 10.1016/bs.pmbts.2020.03.008
Bangoria, P., Divecha, J., and Shah, A. R. (2021). Production of mannooligosaccharides producing β-mannanase by newly isolated Penicillium aculeatum APS1 using oil seed residues under solid state fermentation. Biocatal. Agric. Biotechnol. 34:102023. doi: 10.1016/j.bcab.2021.102023
Belkacem, N., Serafini, N., Wheeler, R., Derrien, M., Boucinha, L., Couesnon, A., et al. (2017). Lactobacillus paracasei feeding improves immune control of influenza infection in mice. PLoS One 12:e0184976. doi: 10.1371/journal.pone.0184976
Bercik, P., Park, A. J., Sinclair, D., Khoshdel, A., Lu, J., Huang, X., et al. (2011). The anxiolytic effect of Bifidobacterium longum NCC3001 involves vagal pathways for gut brain communication. Neurogastroenterol. Motil. 23, 1132–1139. doi: 10.1111/j.1365-2982.2011.01796.x
Bersaneti, G. T., Pan, N. C., Baldo, C., and Celligoi, M. A. P. C. (2018). Co-production of fructooligosaccharides and Levan by levansucrase from Bacillus subtilis natto with potential application in the food industry. Appl. Biochem. Biotechnol. 184, 838–851. doi: 10.1007/s12010-017-2587-0
Bhagwat, A., Haldar, T., Kanojiya, P., and Saroj, S. D. (2025). Bacterial metabolism in the host and its association with virulence. Virulence 16:2459336. doi: 10.1080/21505594.2025.2459336
Bian, J., Peng, F., Peng, X. P., Peng, P., Xu, F., and Sun, R. C. (2013). Structural features and antioxidant activity of xylooligosaccharides enzymatically produced from sugarcane bagasse. Bioresour. Technol. 127, 236–241. doi: 10.1016/j.biortech.2012.09.112
Boets, E., Deroover, L., Houben, E., Vermeulen, K., Gomand, S. V., Delcour, J. A., et al. (2015). Quantification of in vivo colonic short chain fatty acid production from inulin. Nutrients 7, 8916–8929. doi: 10.3390/nu7115440
Boets, E., Gomand, S. V., Deroover, L., Preston, T., Vermeulen, K., De Preter, V., et al. (2017). Systemic availability and metabolism of colonic- derived short-chain fatty acids in healthy subjects: a stable isotope study. J. Physiol. 595, 541–555. doi: 10.1113/JP272613
Bonaz, B., Bazin, T., and Pellissier, S. (2018). The vagus nerve at the interface of the microbiota- gut-brain axis. Front. Neurosci. 12:49. doi: 10.3389/fnins.2018.00049
Bonfili, L., Cecarini, V., Berardi, S., Scarpona, S., Suchodolski, J. S., Nasuti, C., et al. (2017). Microbiota modulation counteracts Alzheimer’s disease progression influencing neuronal proteolysis and gut hormones plasma levels. Sci Rep. 7:2426. doi: 10.1038/s41598-017-02587-2
Boonchuay, P., Techapun, C., Seesuriyachan, P., and Chaiyaso, T. (2014). Production of xylooligosaccharides from corncob using a crude thermostable endo-xylanase from Streptomyces thermovulgaris TISTR1948 and prebiotic properties. Food Sci. Biotechnol. 23, 1515–1523. doi: 10.1007/s10068-014-0207-0
Bo, T. B., Zhang, X. Y., Wen, J., Deng, K., Qin, X. W., and Wang, D. H. (2019). The microbiota-gut-brain interaction in regulating host metabolic adaptation to cold in male Brandt’s voles (Lasiopodomys brandtii). ISME J. 13, 3037–3053. doi: 10.1038/s41396-019-0492-y
Bravo, J. A., Forsythe, P., Chew, M. V., Escaravage, E., Savignac, H. M., Dinan, T. G., et al. (2011). Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. USA 108, 16050–16055. doi: 10.1073/pnas.1102999108
Brienzo, M., Carvalho, W., and Milagres, A. M. (2010). Xylooligosaccharides production from alkali-pretreated sugarcane bagasse using xylanases from Thermoascus aurantiacus. Appl. Biochem. Biotechnol. 162, 1195–1205. doi: 10.1007/s12010-009-8892-5
Busbee, P. B., Menzel, L., Alrafas, H. R., Dopkins, N., Becker, W., Miranda, K., et al. (2020). Indole-3-carbinol prevents colitis and associated microbial dysbiosis in an IL-22-dependent manner. JCI Insight 5:e127551. doi: 10.1172/jci.insight.127551
Bustos Fernández, L. M., Man, F., and Lasa, J. S. (2023). Impact of Saccharomyces boulardii CNCM I-745 on bacterial overgrowth and composition of intestinal microbiota in diarrhea-predominant irritable bowel syndrome patients: results of a randomized pilot study. Dig. Dis. 41, 798–809. doi: 10.1159/000528954
Byakika, S., Mukisa, I. M., Byaruhanga, Y. B., and Muyanja, C. (2019). A review of criteria and methods for evaluating the probiotic potential of microorganisms. Food Rev. Int. 35, 427–466. doi: 10.1080/87559129.2019.1584815
Cano, R., Bermúdez, V., Galban, N., Garrido, B., Santeliz, R., Gotera, M. P., et al. (2024). Dietary polyphenols and gut microbiota cross-talk: molecular and therapeutic perspectives for cardiometabolic disease: a narrative review. Int. J. Mol. Sci. 25:9118. doi: 10.3390/ijms25169118
Capurso, L. (2019). Thirty years of Lactobacillus rhamnosus GG. J. Clin. Gastroenterol. 53, S1–S41. doi: 10.1097/MCG.0000000000001170
Carević, M., Vukašinović-Sekulić, M., Ćorović, M., Rogniaux, H., Ropartz, D., Veličković, D., et al. (2018). Evaluation of β-galactosidase from Lactobacillus acidophilus as biocatalyst for galacto-oligosaccharides synthesis: product structural characterization and enzyme immobilization. J. Biosci. Bioeng. 126, 697–704. doi: 10.1016/j.jbiosc.2018.06.003
Carhart-Harris, R. L., and Nutt, D. J. (2017). Serotonin and brain function: a tale of two receptors. J. Psychopharmacol. 31, 1091–1120. doi: 10.1177/0269881117725915
Ceccherini, C., Daniotti, S., Bearzi, C., and Re, I. (2022). Evaluating the efficacy of probiotics in IBS treatment using a systematic review of clinical trials and multi-criteria decision analysis. Nutrients 14:2689. doi: 10.3390/nu14132689
Chang, C. C., Kao, W. Y., Liu, C. Y., Su, H. H., Kan, Y. A., Lin, P. Y., et al. (2022). Butyrate supplementation regulates expression of chromosome segregation 1-like protein to reverse the genetic distortion caused by p53 mutations in colorectal cancer. Int. J. Oncol. 60:64. doi: 10.3892/ijo.2022.5354
Chang, P. V., Hao, L., Offermanns, S., and Medzhitov, R. (2014). The microbial metabolite butyrate regulates intestinal macrophage function via histone deacetylase inhibition. Proc. Natl. Acad. Sci. USA 111, 2247–2252. doi: 10.1073/pnas.1322269111
Chapla, D., Pandit, P., and Shah, A. (2012). Production of xylooligosaccharides from corncob xylan by fungal xylanase and their utilization by probiotics. Bioresour. Technol. 115, 215–221. doi: 10.1016/j.biortech.2011.10.083
Charbonneau, M. R., Isabella, V. M., Li, N., and Kurtz, C. B. (2020). Developing a new class of engineered live bacterial therapeutics to treat human diseases. Nat. Commun. 11:1738. doi: 10.1038/s41467-020-15508-1
Chavan, A. R., Singh, A. K., Gupta, R. K., Nakhate, S. P., Poddar, B. J., Gujar, V. V., et al. (2023). Recent trends in the biotechnology of functional non-digestible oligosaccharides with prebiotic potential. Biotechnol. Genet. Eng. Rev. 39, 465–510. doi: 10.1080/02648725.2022.2152627
Cheng, J., Laitila, A., and Ouwehand, A. C. (2021). Bifidobacterium animalis subsp. lactis HN019 effects on gut health: a review. Front. Nutr. 8:790561. doi: 10.3389/fnut.2021.790561
Chen, X., Xu, D., Yu, J., Song, X. J., Li, X., and Cui, Y. L. (2024). Tryptophan metabolism disorder-triggered diseases, mechanisms, and therapeutic strategies: a scientometric review. Nutrients 16:3380. doi: 10.3390/nu16193380
Chiu, H. F., Venkatakrishnan, K., Golovinskaia, O., and Wang, C. K. (2021). Gastroprotective effects of polyphenols against various gastro-intestinal disorders: a mini-review with special focus on clinical evidence. Molecules 26:2090. doi: 10.3390/molecules26072090
Chudzik, A., Orzyłowska, A., Rola, R., and Stanisz, G. J. (2021). Probiotics, prebiotics and postbiotics on mitigation of depression symptoms: modulation of the brain–gut–microbiome Axis. Biomolecules 11:1000. doi: 10.3390/biom11071000
Correa-Oliveira, R., Fachi, J. L., Vieira, A., Sato, F. T., and Vinolo, M. A. (2016). Regulation of immune cell function by short- chain fatty acids. Clin. Translational Immunol. 5:e73. doi: 10.1038/cti.2016.17
D’Amico, V., Cavaliere, M., Ivone, M., Lacassia, C., Celano, G., Vacca, M., et al. (2025). Microencapsulation of probiotics for enhanced stability and health benefits in dairy functional foods: a focus on pasta filata cheese. Pharmaceutics 17:185. doi: 10.3390/pharmaceutics17020185
Dahiya, D., and Nigam, P. S. (2022). The gut microbiota ınfluenced by the ıntake of probiotics and functional foods with prebiotics can sustain wellness and alleviate certain ailments like gut-inflammation and colon-cancer. Microorganisms 10:665. doi: 10.3390/microorganisms10030665
Dalile, B., Van Oudenhove, L., Vervliet, B., and Verbeke, K. (2019). The role of short- chain fatty acids in microbiota–gut–brain communication. Nat. Rev. Gastroenterol. Hepatol. 16, 461–478. doi: 10.1038/s41575-019-0157-3
Davani-Davari, D., Negahdaripour, M., Karimzadeh, I., Seifan, M., Mohkam, M., Masoumi, S. J., et al. (2019). Prebiotics: definition, types, sources, mechanisms, and clinical applications. Foods 8:92. doi: 10.3390/foods8030092
de Figueiredo, F. C., Carvalho, A. F. A., Brienzo, M., Campioni, T. S., and de Oliva-Neto, P. (2017). Chemical input reduction in the arabinoxylan and lignocellulose alkaline extraction and xylooligosaccharides production. Bioresour. Technol. 228, 164–170. doi: 10.1016/j.biortech.2016.12.097
de Melo Pereira, G. V., de Oliveira Coelho, B., Magalhães Júnior, A. I., Thomaz-Soccol, V., and Soccol, C. R. (2018). How to select a probiotic? A review and update of methods and criteria. Biotechnol. Adv. 36, 2060–2076. doi: 10.1016/j.biotechadv.2018.09.003
Diez-Municio, M., De Las Rivas, B., Jimeno, M. L., Muñoz, R., Moreno, F. J., and Herrero, M. (2013). Enzymatic synthesis and characterization of fructooligosaccharides and novel maltosylfructosides by inulosucrase from Lactobacillus gasseri DSM 20604. Appl. Environ. Microbiol. 79, 4129–4140. doi: 10.1128/AEM.00854-13
Dolatkhah, N., Jafari, A., Eslamian, F., Toopchizadeh, V., Saleh, P., and Hashemian, M. (2024). Saccharomyces boulardii improves clinical and paraclinical indices in overweight/obese knee osteoarthritis patients: a randomized triple-blind placebo-controlled trial. Eur. J. Nutr. 63, 2291–2305. doi: 10.1007/s00394-024-03428-5
Dronkers, T. M. G., Ouwehand, A. C., and Rijkers, G. T. (2020). Global analysis of clinical trials with probiotics. Heliyon 6:e04467. doi: 10.1016/j.heliyon.2020.e04467
Duque, A. L. R. F., Demarqui, F. M., Santoni, M. M., Zanelli, C. F., Adorno, M. A. T., Milenkovic, D., et al. (2021). Effect of probiotic, prebiotic, and synbiotic on the gut microbiota of autistic children using an in vitro gut microbiome model. Food Res. Int. 149:110657. doi: 10.1016/j.foodres.2021.110657
EFSA Panel on Contaminants in the Food Chain (CONTAM) (2018). Scientific opinion on the risks for animal health related to the presence of deoxynivalenol and its acetylated and modified forms in feed. EFSA J. 16:5206. doi: 10.2903/j.efsa.2018.5206
Ejtahed, H. S., Mohtadi-Nia, J., Homayouni-Rad, A., Niafar, M., Asghari-Jafarabadi, M., Mofid, V., et al. (2011). Effect of probiotic yogurt containing Lactobacillus acidophilus and Bifidobacterium lactis on lipid profile in individuals with type 2 diabetes mellitus. J. Dairy Sci. 94, 3288–3294. doi: 10.3168/jds.2010-4128
Elsabagh, M., Ishikake, M., Sakamoto, Y., Haruno, A., Miura, M., Fujieda, T., et al. (2018). Postruminal supply of amino acids enhances ghrelin secretion and lipid metabolism in feed-deprived sheep. Anim. Sci. J. 89, 1663–1672. doi: 10.1111/asj.13114
Eskandarloo, H., and Abbaspourrad, A. (2018). Production of galacto-oligosaccharides from whey permeate using β-galactosidase immobilized on functionalized glass beads. Food Chem. 251, 115–124. doi: 10.1016/j.foodchem.2018.01.068
Facchin, S., Bertin, L., Bonazzi, E., Lorenzon, G., De Barba, C., Barberio, B., et al. (2024). Short-chain fatty acids and human health: from metabolic pathways to current therapeutic implications. Life 14:559. doi: 10.3390/life14050559
Fai, A. E. C., da Silva, J. B., de Andrade, C. J., Bution, M. L., and Pastore, G. M. (2014). Production of prebiotic galactooligosaccharides from lactose by Pseudozyma tsukubaensis and Pichia kluyveri. Biocatal. Agric. Biotechnol. 3, 343–350. doi: 10.1016/j.bcab.2014.04.005
Falck, P., Precha-Atsawanan, S., Grey, C., Immerzeel, P., Stålbrand, H., Adlercreutz, P., et al. (2013). Xylooligosaccharides from hardwood and cereal xylans produced by a thermostable xylanase as carbon sources for Lactobacillus brevis and Bifidobacterium adolescentis. J. Agric. Food Chem. 61, 7333–7340. doi: 10.1021/jf401249g
FAO/WHO (2006). Probiotics in food. Health and nutritional properties and guidelines for evaluation. FAO Food Nutr 85, 1–50.
Fekete, M., Lehoczki, A., Major, D., Fazekas-Pongor, V., Tamás Csípő,, Tarantini, S., et al. (2024). Exploring the influence of gut–brain axis modulation on cognitive health: a comprehensive review of prebiotics, probiotics, and symbiotics. Nutrients 16:789. doi: 10.3390/nu16060789
Fischer, C., and Kleinschmidt, T. (2021). Synthesis of galactooligosaccharides by Cryptococcus laurentii and Aspergillus oryzae using different kinds of acid whey. Int. Dairy J. 112:104867. doi: 10.1016/j.idairyj.2020.104867
Flint, H. J., Duncan, S. H., Scott, K. P., and Louis, P. (2015). Links between diet, gut microbiota composition and gut metabolism. Proc. Nutr. Soc. 74, 13–22. doi: 10.1017/S0029665114001463
Forssten, S. D., Ouwehand, A. C., Griffin, S. M., and Patterson, E. (2022). One giant leap from mouse to man: the microbiota–gut–brain axis in mood disorders and translational challenges moving towards human clinical trials. Nutrients 14:568. doi: 10.3390/nu14030568
Fusco, W., Lorenzo, M. B., Cintoni, M., Porcari, S., Rinninella, E., Kaitsas, F., et al. (2023). Short-chain fatty-acid-producing bacteria: key components of the human gut microbiota. Nutrients 15:2211. doi: 10.3390/nu15092211
Galica, A. N., Galica, R., and Dumitrașcu, D. L. (2022). Diet, fibers, and probiotics for irritable bowel syndrome. J. Med. Life 15, 174–179. doi: 10.25122/jml-2022-0028
Gallardo, O., Fernández-Fernández, M., Valls, C., Valenzuela, S. V., Roncero, M. B., Vidal, T., et al. (2010). Characterization of a family GH5 xylanase with activity on neutral oligosaccharides and evaluation as a pulp bleaching aid. Appl. Environ. Microbiol. 76, 6290–6294. doi: 10.1128/AEM.00871-10
Ganaie, M. A., Soni, H., Naikoo, G. A., Oliveira, L. T. S., Rawat, H. K., Mehta, P. K., et al. (2017). Screening of low cost agricultural wastes to maximize the fructosyltransferase production and its applicability in generation of fructooligosaccharides by solid state fermentation. Int. Biodeterior. Biodegradation 118, 19–26. doi: 10.1016/j.ibiod.2017.01.006
Garcia-Mantrana, I., Selma-Royo, M., Alcantara, C., and Collado, M. C. (2018). Shifts on gut microbiota associated to mediterranean diet adherence and specific dietary intakes on general adult population. Front. Microbiol. 9:890. doi: 10.3389/fmicb.2018.00890
Gautério, G. V., Vieira, M. C., da Silva, L. G. G., Hübner, T., Sanzo, A. V. L., and Kalil, S. J. (2018). Production of xylanolitic enzymes and xylooligosaccharides by Aureobasidium pullulans CCT 1261 in submerged cultivation. Ind. Crop. Prod. 125, 335–345. doi: 10.1016/j.indcrop.2018.09.011
Geiger, B., Nguyen, H. M., Wenig, S., Nguyen, H. A., Lorenz, C., Kittl, R., et al. (2016). From by-product to valuable components: efficient enzymatic conversion of lactose in whey using β-galactosidase from Streptococcus thermophilus. Biochem. Eng. J. 116, 45–53. doi: 10.1016/j.bej.2016.04.003
Geypens, B., Claus, D., Evenepoel, P., Hiele, M., Maes, B., Peeters, M., et al. (1997). Influence of dietary protein supplements on the formation of bacterial metabolites in the colon. Gut 41, 70–76.
Ghorbani, Z., Nazari, S., Etesam, F., Nourimajd, S., Ahmadpanah, M., Jahromi, S. R., et al. (2018). The effect of synbiotic as an adjuvant therapy to fluoxetine in moderate depression: A randomized multicenter trial. Arch Neurosci. 5:e60507. doi: 10.5812/archneurosci.60507
Gibson, G. R., Hutkins, R., Sanders, M. E., Prescott, S. L., Reimer, R. A., Salminen, S. J., et al. (2017). The international scientific association for probiotics and prebiotics (ISAPP) consensus statement on the definition and scope of prebiotics. Nat. Rev. Gastroenterol. Hepatol. 14, 491–502. doi: 10.1038/nrgastro.2017.75
Golubeva, A. V., Joyce, S. A., Moloney, G., Burokas, A., Sherwin, E., Arboleya, S., et al. (2017). Microbiota-related changes in bile acid & tryptophan metabolism are associated with gastrointestinal dysfunction in a mouse model of autism. EBioMedicine 24, 166–178. doi: 10.1016/j.ebiom.2017.09.020
Gomez-Arango, L. F., Barrett, H. L., McIntyre, H. D., Callaway, L. K., Morrison, M., and Nitert, M. D. (2016). Connections between the gut microbiome and metabolic hormones in early pregnancy in overweight and obese women. Diabetes 65, 2214–2223. doi: 10.2337/db16-0278
Góralczyk-Bińkowska, A., Szmajda-Krygier, D., and Kozłowska, E. (2022). The microbiota-gut-brain axis in psychiatric disorders. Int. J. Mol. Sci. 23:11245. doi: 10.3390/ijms231911245
Goswami, C., Iwasaki, Y., and Yada, T. (2018). Short- chain fatty acids suppress food intake by activating vagal afferent neurons. J. Nutr. Biochem. 57, 130–135. doi: 10.1016/j.jnutbio.2018.03.009
Gowdhaman, D., and Ponnusami, V. (2015). Production and optimization of xylooligosaccharides from corncob by Bacillus aerophilus KGJ2 xylanase and its antioxidant potential. Int. J. Biol. Macromol. 79, 595–600. doi: 10.1016/j.ijbiomac.2015.05.046
Gracie, D. J., Hamlin, P. J., and Ford, A. C. (2019). The influence of the brain–gut axis in inflammatory bowel disease and possible implications for treatment. Lancet Gastroenterol. Hepatol. 4, 632–642. doi: 10.1016/S2468-1253(19)30089-5
Guo, X., Su, Y., Du, Y., Zhang, F., Yu, W., Ren, W., et al. (2025). Vinegar-processed Schisandra chinensis polysaccharide ameliorates type 2 diabetes via modulation serum metabolic profiles, gut microbiota, and fecal SCFAs. Int. J. Biol. Macromol. 294:139514. doi: 10.1016/j.ijbiomac.2025.139514
Guo, Y. T., Peng, Y. C., Yen, H. Y., Wu, J. C., and Hou, W. H. (2022). Effects of probiotic supplementation on immune and inflammatory markers in athletes: a meta-analysis of randomized clinical trials. Medicina (Kaunas) 58:1188. doi: 10.3390/medicina58091188
Gu, Y., Wang, C., Qin, X., Zhou, B., Liu, X., Liu, T., et al. (2022). Saccharomyces boulardii, a yeast probiotic, inhibits gut motility through upregulating intestinal serotonin transporter and modulating gut microbiota. Pharmacol. Res. 181:106291. doi: 10.1016/j.phrs.2022.106291
Hackenhaar, C. R., Spolidoro, L. S., Flores, E. E. E., Klein, M. P., and Hertz, P. F. (2021). Batch synthesis of galactooligosaccharides from co-products of milk processing using immobilized β-galactosidase from Bacillus circulans. Biocatal. Agric. Biotechnol. 36:102136. doi: 10.1016/j.bcab.2021.102136
Hamdi, A., Viera-Alcaide, I., Guillén-Bejarano, R., Rodríguez-Arcos, R., Muñoz, M. J., Monje Moreno, J. M., et al. (2023). Asparagus fructans as emerging prebiotics. Foods 12:81. doi: 10.3390/foods12010081
Hero, J. S., Pisa, J. H., Raimondo, E. E., and Martínez, M. A. (2021). Proteomic analysis of secretomes from Bacillus sp. AR03: characterization of enzymatic cocktails active on complex carbohydrates for xylooligosaccharides production. Prep. Biochem. Biotechnol. 51, 871–880. doi: 10.1080/10826068.2020.1870136
Hill, C., Guarner, F., Reid, G., Gibson, G. R., Merenstein, D. J., Pot, B., et al. (2014). The international scientific association for probiotics and prebiotics consensus statement on the scope and appropriate use of the term probiotic. Nat. Rev. Gastroenterol. Hepatol. 11, 506–514. doi: 10.1038/nrgastro.2014.66
Hofmeister, M., Clement, F., Patten, S., Li, J., Dowsett, L. E., Farkas, B., et al. (2021). The effect of interventions targeting gut microbiota on depressive symptoms: a systematic review and meta-analysis. CMAJ Open 9, E1195–E1204. doi: 10.9778/cmajo.20200283
Hong, C. T., Chen, J. H., and Huang, T. W. (2022). Probiotics treatment for Parkinson disease: a systematic review and meta-analysis of clinical trials. Aging 14, 7014–7025. doi: 10.18632/aging.204266
Hooda, S., Vester Boler, B. M., Kerr, K. R., Dowd, S. E., and Swanson, K. S. (2013). The gut microbiome of kittens is affected by dietary protein:carbohydrate ratio and associated with blood metabolite and hormone concentrations. Br. J. Nutr. 109, 1637–1646. doi: 10.1017/S0007114512003479
Huang, M. P., Wu, M., Xu, Q. S., Mo, D. J., and Feng, J. X. (2016). Highly efficient synthesis of fructooligosaccharides by extracellular fructooligosaccharide-producing enzymes and immobilized cells of Aspergillus aculeatus M105 and purification and biochemical characterization of a fructosyltransferase from the fungus. J. Agric. Food Chem. 64, 6425–6432. doi: 10.1021/acs.jafc.6b02115
Ikenoya, C., Takemi, S., Kaminoda, A., Aizawa, S., Ojima, S., Gong, Z., et al. (2018). Β-Oxidation in ghrelin-producing cells is important for ghrelin acyl-modification. Sci. Rep. 8:9176. doi: 10.1038/s41598-018-27458-2
Jayapal, N., Samanta, A. K., Kolte, A. P., Senani, S., Sridhar, M., Suresh, K. P., et al. (2013). Value addition to sugarcane bagasse: Xylan extraction and its process optimization for xylooligosaccharides production. Ind. Crop. Prod. 42, 14–24. doi: 10.1016/j.indcrop.2012.05.019
Jnawali, P., Kumar, V., Tanwar, B., Hirdyani, H., and Gupta, P. (2018). Enzymatic production of xylooligosaccharides from brown coconut husk treated with sodium hydroxide. Waste Biomass Valoriz. 9, 1757–1766. doi: 10.1007/s12649-017-9963-4
Johnson, K. V., and Steenbergen, L. (2025). Probiotics reduce negative mood over time: The value of daily self-reports in detecting effects. Npj Ment Health Res. 4:10. doi: 10.1038/s44184-025-00123-z
Jost, T., Lacroix, C., Braegger, C. P., and Chassard, C. (2012). New insights in gut microbiota establishment in healthy breast fed neonates. PLoS One 7:e44595. doi: 10.1371/journal.pone.0044595
Jovanovic-Malinovska, R., Fernandes, P., Winkelhausen, E., and Fonseca, L. (2012). Galacto-oligosaccharides synthesis from lactose and whey by β-galactosidase immobilized in PVA. Appl. Biochem. Biotechnol. 168, 1197–1211. doi: 10.1007/s12010-012-9850-1
Kallel, F., Driss, D., Bouaziz, F., Neifer, M., Ghorbel, R., and Chaabouni, S. E. (2015). Production of xylooligosaccharides from garlic straw xylan by purified xylanase from Bacillus mojavensis UEB-FK and their in vitro evaluation as prebiotics. Food Bioprod. Process. 94, 536–546. doi: 10.1016/j.fbp.2014.07.012
Kamal, N., Saharan, B. S., Duhan, J. S., Kumar, A., Chaudhary, P., Goyal, C., et al. (2025). Exploring the promise of psychobiotics: bridging gut microbiota and mental health for a flourishing society. Med. Microecol. 23:100118. doi: 10.1016/j.medmic.2024.100118
Kang, C. H., Kim, J. S., Kim, H., Park, H. M., and Paek, N. S. (2021). Heat-killed lactic acid bacteria inhibit nitric oxide production via inducible nitric oxide synthase and cyclooxygenase-2 in RAW 264.7 cells. Probiotics Antimicrob. Proteins 13, 1530–1538. doi: 10.1007/s12602-021-09781-9
Kang, C., Zhang, Y., Zhu, X., Liu, K., Wang, X., Chen, M., et al. (2016). Healthy subjects differentially respond to dietary capsaicin correlating with specific gut enterotypes. J. Clin. Endocrinol. Metab. 101, 4681–4689. doi: 10.1210/jc.2016-2786
Karen, C., Shyu, D. J. H., and Rajan, K. E. (2021). Lactobacillus paracasei supplementation prevents early life stress-induced anxiety and depressive-like behavior in maternal separation model-possible involvement of microbiota-gut-brain axis in differential regulation of MicroRNA124a/132 and glutamate receptors. Front. Neurosci. 15:719933. doi: 10.3389/fnins.2021.719933
Karim, A. (2025). Unveiling the potential of probiotics in osteoarthritis management. Curr. Rheumatol. Rep. 27:2. doi: 10.1007/s11926-024-01166-5
Katapodis, P., Kavarnou, A., Kintzios, S., Pistola, E., Kekos, D., Macris, B. J., et al. (2002). Production of acidic xylo-oligosaccharides by a family 10 endoxylanase from Thermoascus aurantiacus and use as plant growth regulators. Biotechnol. Lett. 24, 1413–1416. doi: 10.1023/A:1019898414801
Katsurada, K., and Yada, T. (2016). Neural effects of gut- and brain- derived glucagon- like peptide-1 and its receptor agonist. J. Diabetes Investig. 7, 64–69. doi: 10.1111/jdi.12464
Kaulpiboon, J., Rudeekulthamrong, P., Watanasatitarpa, S., Ito, K., and Pongsawasdi, P. (2015). Synthesis of long-chain isomaltooligosaccharides from tapioca starch and an in vitro investigation of their prebiotic properties. J. Mol. Catal. B Enzym. 120, 127–135. doi: 10.1016/j.molcatb.2015.07.004
Kelly, J. R., Borre, Y., O’ Brien, C., Patterson, E., El Aidy, S., Deane, J., et al. (2016). Transferring the blues: depression-associated gut microbiota induces neurobehavioural changes in the rat. J. Psychiatr. Res. 82, 109–118. doi: 10.1016/j.jpsychires.2016.07.019
Kerlikowsky, F., Müller, M., Greupner, T., Amend, L., Strowig, T., and Hahn, A. (2025). Distinct microbial taxa are associated with LDL-cholesterol reduction after 12 weeks of Lactobacillus plantarum intake in mild hypercholesterolemia: results of a randomized controlled study. Probiotics Antimicrob. Proteins 17, 1086–1095. doi: 10.1007/s12602-023-10191-2
Khalil, N. A., Eltahan, N. R., Elaktash, H. M., Aly, S., and Sarbini, S. R. (2021). Prospective evaluation of probiotic and prebiotic supplementation on diabetic health associated with gut microbiota. Food Biosci. 42:101149. doi: 10.1016/j.fbio.2021.101149
Khongrum, J., Yingthongchai, P., Boonyapranai, K., Wongtanasarasin, W., Aobchey, P., Tateing, S., et al. (2023). Safety and effects of Lactobacillus paracasei TISTR 2593 supplementation on ımproving cholesterol metabolism and atherosclerosis-related parameters in subjects with hypercholesterolemia: a randomized, double-blind, placebo-controlled clinical trial. Nutrients 15:661. doi: 10.3390/nu15030661
Kim, C. H., Park, J., and Kim, M. (2014). Gut microbiota- derived short- chain fatty acids, T cells, and inflammation. Immune Netw. 14, 277–288. doi: 10.4110/in.2014.14.6.277
Kim, S., Kang, B. G., Sa, S., Park, S. Y., Ryu, K., Lee, J., et al. (2024). Advanced fructo-oligosaccharides improve itching and aberrant epidermal lipid composition in children with atopic dermatitis. Front. Microbiol. 15:1383779. doi: 10.3389/fmicb.2024.1383779
Kim, Y. T., Kim, C. H., Kwon, J. G., Cho, J. H., Shin, Y. S., Kim, H. B., et al. (2022). In vivo trial of Bifidobacterium longum revealed the complex network correlations between gut microbiota and health promotional effects. Front. Microbiol. 13:886934. doi: 10.3389/fmicb.2022.886934
Kittibunchakul, S., Pham, M. L., Tran, A. M., and Nguyen, T. H. (2019). β-Galactosidase from Lactobacillus helveticus DSM 20075: biochemical characterization and recombinant expression for applications in dairy industry. Int. J. Mol. Sci. 20:947. doi: 10.3390/ijms20040947
Kleerebezem, M., and Führen, J. (2024). Synergistic vs. complementary synbiotics: the complexity of discriminating synbiotic concepts using a Lactiplantibacillus plantarum exemplary study. Microbiome Res Rep. 3:46. doi: 10.20517/mrr.2024.48
Koda, S., Date, Y., Murakami, N., Shimbara, T., Hanada, T., Toshinai, K., et al. (2005). The role of the vagal nerve in peripheral PYY3-36-induced feeding reduction in rats. Endocrinology 146, 2369–2375. doi: 10.1210/en.2004-1266
Kolida, S., and Gibson, G. R. (2011). Synbiotics in health and disease. Annu. Rev. Food Sci. Technol. 2, 373–393. doi: 10.1146/annurev-food-022510-133739
Kumar, A., Kumar, V., Pramanik, J., Rustagi, S., Prajapati, B., Jebreen, A., et al. (2025). Lactiplantibacillus plantarum as a complementary approach for diabetes treatment and management. Curr. Nutr. Rep. 14:72. doi: 10.1007/s13668-025-00664-x
Kwok, K. O., Fries, L. R., Thakkar, S. K., Iroz, A., and Blanchard, C. (2022). Effects of probiotic intervention on markers of inflammation and health outcomes in women of reproductive age and their children. Front. Nutr. 9:889040. doi: 10.3389/fnut.2022.889040
Larraufie, P., Martin-Gallausiaux, C., Lapaque, N., Dore, J., Gribble, F. M., Reimann, F., et al. (2018). SCFAs strongly stimulate PYY production in human enteroendocrine cells. Sci. Rep. 8:74. doi: 10.1038/s41598-017-18259-0
Lee, J. G., Woo, Y. S., Park, S. W., Seog, D. H., Seo, M. K., and Bahk, W. M. (2019). The neuroprotective effects of melatonin: possible role in the pathophysiology of neuropsychiatric disease. Brain Sci. 9:285. doi: 10.3390/brainsci9100285
Lee, M., Bok, M. K., Son, K., Lee, M., Park, H., Yang, J., et al. (2024). Bifidobacterium lactis IDCC 4301 (B. lactis FitTM) supplementation effects on body fat, serum triglyceride, and adipokine ratio in obese women: a randomized clinical trial. Food Funct. 15, 8448–8458. doi: 10.1039/D4FO00535J
Leeuwendaal, N. K., Cryan, J. F., and Schellekens, H. (2021). Gut peptides and the microbiome: focus on ghrelin. Curr. Opin. Endocrinol. Diabetes Obes. 28, 243–252. doi: 10.1097/MED.0000000000000616
Lekić, L. (2024). The multifaceted effects of Lactobacillus acidophilus on endocrine, gastrointestinal and cardiovascular health: a clinical study. IJISRT 9, 160–163. doi: 10.5281/zenodo.14500380
Liang, S., Wu, X., and Jin, F. (2018). Gut-brain psychology: rethinking psychology from the microbiota-gut-brain axis. Front. Integr. Neurosci. 12:33. doi: 10.3389/fnint.2018.00033
Li, B., Liang, L., Deng, H., Guo, J., Shu, H., and Zhang, L. (2020). Efficacy and safety of probiotics in ırritable bowel syndrome: a systematic review and meta-analysis. Front. Pharmacol. 11, 362–372. doi: 10.3389/fphar.2020.00332
Lima, A. A.de, Santos, S. S., Araújo, M. A.de, Vinderola, G., Villarreal, C. F., and Viana, M. D. M. (2025). Probiotics as technological innovations in psychiatric disorders: patents and research reviews Front. Nutr. 12:1567097. doi: 10.3389/fnut.2025.1567097
Liu, C., Yang, L., Wei, W., and Fu, P. (2024). Efficacy of probiotics/synbiotics supplementation in patients with chronic kidney disease: a systematic review and meta-analysis of randomized controlled trials. Front. Nutr. 11:1434613. doi: 10.3389/fnut.2024.1434613
Liu, L., Xu, M., Cao, Y., Wang, H., Shao, J., Xu, M., et al. (2020). Biochemical characterization of xylanases from Streptomyces sp. B6 and their application in the xylooligosaccharide production from viscose fiber production waste. J. Agric. Food Chem. 68, 3184–3194. doi: 10.1021/acs.jafc.9b06704
Liu, R., Zhang, C., Shi, Y., Zhang, F., Li, L., Wang, X., et al. (2017). Dysbiosis of gut microbiota associated with clinical parameters in polycystic ovary syndrome. Front. Microbiol. 8:324. doi: 10.3389/fmicb.2017.00324
Liu, X., Jin, G., Tang, Q., Huang, S., Zhang, Y., Sun, Y., et al. (2022a). Early life Lactobacillus rhamnosus GG colonisation inhibits intestinal tumour formation. Br. J. Cancer 126, 1421–1431. doi: 10.1038/s41416-021-01562-z
Liu, X., Liu, Y., Jiang, Z., Liu, H., Yang, S., and Yan, Q. (2018). Biochemical characterization of a novel xylanase from Paenibacillus barengoltzii and its application in xylooligosaccharides production from corncobs. Food Chem. 264, 310–318. doi: 10.1016/j.foodchem.2018.05.023
Liu, X., Yan, Q., Xue, Y., Wang, S., Yang, H., and Jiang, Z. (2022b). Biochemical characterization of a novel glycoside hydrolase family 11 xylanase from Chaetomium sp. suitable for bread making. Process Biochem. 117, 1–9. doi: 10.1016/j.procbio.2022.03.017
Liu, Y., Nawazish, H., Farid, M. S., Abdul Qadoos, K., Habiba, U. E., Muzamil, M., et al. (2024). Health-promoting effects of Lactobacillus acidophilus and its technological applications in fermented food products and beverages. Fermentation 10:380. doi: 10.3390/fermentation10080380
Liu, Y. W., Liong, M. T., Chung, Y. C. E., Huang, H. Y., Peng, W. S., Cheng, Y. F., et al. (2019). Effects of Lactobacillus plantarum PS128 on children with autism spectrum disorder in Taiwan: a randomized, double-blind, placebo-controlled trial. Nutrients 11:820. doi: 10.3390/nu11040820
Li, X., Li, E., Zhu, Y., Teng, C., Sun, B., Song, H., et al. (2012). A typical endo-xylanase from Streptomyces rameus L2001 and its unique characteristics in xylooligosaccharide production. Carbohydr. Res. 359, 30–36. doi: 10.1016/j.carres.2012.05.005
Loh, J. S., Mak, W. Q., Tan, L. K. S., Ng, C. X., Chan, H. H., Yeow, S. H., et al. (2024). Microbiota-gut-brain axis and its therapeutic applications in neurodegenerative diseases. Signal Transduct Target Ther. 9:37. doi: 10.1038/s41392-024-01743-1
Louis, P., Young, P., Holtrop, G., and Flint, H. J. (2010). Diversity of human colonic butyrate-producing bacteria revealed by analysis of the butyryl-CoA:acetate CoA-transferase gene. Environ. Microbiol. 12, 304–314. doi: 10.1111/j.1462-2920.2009.02066.x
Lu, H., Liu, S., Zhang, S., Chen, J., and Chen, Q. (2025). Suppression of Alzheimer’s disease by Agaricus sinodeliciosus var. chaidam exopolysaccharide with amyloid-β clearance activity via gut microbiota-metabolite regulation. Int. J. Biol. Macromol. 13:141048. doi: 10.1016/j.ijbiomac.2025.14104
Lukic, I., Getselter, D., Koren, O., and Elliott, E. (2019). Role of tryptophan in microbiota-induced depressive-like behavior: evidence from tryptophan depletion study. Front. Behav. Neurosci. 13:123. doi: 10.3389/fnbeh.2019.00123
Mahana, D., Trent, C. M., Kurtz, Z. D., Bokulich, N. A., Battaglia, T., Chung, J., et al. (2016). Antibiotic perturbation of the murine gut microbiome enhances the adiposity, insulin resistance, and liver disease associated with high-fat diet. Genome Med. 8:48. doi: 10.1186/s13073-016-0297-9
Maiuolo, J., Gliozzi, M., Musolino, V., Carresi, C., Scarano, F., Nucera, S., et al. (2021). The contribution of gut microbiota-brain axis in the development of brain disorders. Front. Neurosci. 15:616883. doi: 10.3389/fnins.2021.616883
Mandic, A. D., Woting, A., Jaenicke, T., Sander, A., Sabrowski, W., Rolle-Kampcyk, U., et al. (2019). Clostridium ramosum regulates enterochromaffin cell development and serotonin release. Sci. Rep. 9:1177. doi: 10.1038/s41598-018-38018-z
Mano, M. C. R., Paulino, B. N., and Pastore, G. M. (2019). Whey permeate as the raw material in galacto-oligosaccharide synthesis using commercial enzymes. Food Res. Int. 124, 78–85. doi: 10.1016/j.foodres.2018.09.019
Maráz, A., Kovács, Z., Benjamins, E., and Pázmándi, M. (2022). Recent developments in microbial production of high-purity galacto-oligosaccharides. World J. Microbiol. Biotechnol. 38:95. doi: 10.1007/s11274-022-03279-4
Marco, M. L., Sanders, M. E., Gänzle, M., Arrieta, M. C., Cotter, P. D., De Vuyst, L., et al. (2021). The international scientific Association for Probiotics and Prebiotics (ISAPP) consensus statement on fermented foods. Nat. Rev. Gastroenterol. Hepatol. 18, 196–208. doi: 10.1038/s41575-020-00390-5
Markowiak-Kopeć, P., and Śliżewska, K. (2020). The effect of probiotics on the production of short-chain fatty acids by human intestinal microbiome. Nutrients 12:1107. doi: 10.3390/nu12041107
Markowiak, P., and Śliżewska, K. (2017). Effects of probiotics, prebiotics, and synbiotics on human health. Nutrients 9:1021. doi: 10.3390/nu9091021
Martinez-Porchas, M., Preciado-Álvarez, A., Vargas-Albores, F., Gracia-Valenzuela, M. H., Cicala, F., Martinez-Cordova, L. R., et al. (2023). Microbiota plasticity in tilapia gut revealed by meta-analysis evaluating the effect of probiotics, prebiotics, and biofloc. PeerJ 11:e16213. doi: 10.7717/peerj.16213
Martin-Gallausiaux, C., Larraufie, P., Jarry, A., Beguet-Crespel, F., Marinelli, L., Ledue, F., et al. (2018). Butyrate produced by commensal bacteria down-regulates indolamine 2,3-dioxygenase 1 (IDO-1) expression via a dual mechanism in human intestinal epithelial cells. Front. Immunol. 9:2838. doi: 10.3389/fimmu.2018.02838
Massot-Cladera, M., Mayneris-Perxachs, J., Costabile, A., Swann, J. R., Franch, À., Pérez-Cano, F. J., et al. (2017). Association between urinary metabolic profile and the intestinal effects of cocoa in rats. Br. J. Nutr. 117, 623–634. doi: 10.1017/S0007114517000496
Mazzone, L., Dooling, S. W., Volpe, E., Uljarević, M., Waters, J. L., Sabatini, A., et al. (2024). Precision microbial intervention improves social behavior but not autism severity: a pilot double-blind randomized placebo-controlled trial. Cell Host Microbe 32, 106–116.e6. doi: 10.1016/j.chom.2023.11.021
McGavigan, A. K., O’hara, H. C., Amin, A., Kinsey-Jones, J., Spreckley, E., Alamshah, A., et al. (2015). L-cysteine suppresses ghrelin and reduces appetite in rodents and humans. Int. J. Obes. 39, 447–455. doi: 10.1038/ijo.2014.172
Meher, A. K., Acharya, B., and Sahu, P. K. (2024). Probiotics: bridging the interplay of a healthy gut and psychoneurological well-being. Food Bioeng. 3, 126–147. doi: 10.1002/fbe2.12081
Merenstein, D., Pot, B., Leyer, G., Ouwehand, A. C., Preidis, G. A., Elkins, C. A., et al. (2023). Emerging issues in probiotic safety: 2023 perspectives. Gut Microbes 15:2185034. doi: 10.1080/19490976.2023.2185034
Messaoudi, M., Lalonde, R., Violle, N., Javelot, H., Desor, D., Nejdi, A., et al. (2011). Assessment of psychotropic-like properties of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in rats and human subjects. Br. J. Nutr. 105, 755–764. doi: 10.1017/S0007114510004319
Moens, F., Van den Abbeele, P., Basit, A. W., Dodoo, C., Chatterjee, R., Smith, B., et al. (2019). A four-strain probiotic exerts positive immunomodulatory effects by enhancing colonic butyrate production in vitro. Int. J. Pharm. 555, 1–10. doi: 10.1016/j.ijpharm.2018.11.020
Morais, L. H., Schreiber, H. L., and Mazmanian, S. K. (2021). The gut microbiota-brain axis in behaviour and brain disorders. Nat. Rev. Microbiol. 19, 241–255. doi: 10.1038/s41579-020-00460-0
Mörkl, S., Butler, M. I., Holl, A., Cryan, J. F., and Dinan, T. G. (2020). Probiotics and the microbiota-gut-brain axis: focus on psychiatry. Curr. Nutr. Rep. 9, 171–182. doi: 10.1007/s13668-020-00313-5
Mu, Q., Tavella, V. J., and Luo, X. M. (2018). Role of Lactobacillus reuteri in human health and diseases. Front. Microbiol. 9:757. doi: 10.3389/fmicb.2018.00757
Nascimento, A. K. C., Nobre, C., Cavalcanti, M. T. H., Teixeira, J. A., and Porto, A. L. (2016). Screening of fungi from the genus Penicillium for production of β-fructofuranosidase and enzymatic synthesis of fructooligosaccharides. J. Mol. Catal. B Enzym. 134, 70–78. doi: 10.1016/j.molcatb.2016.09.005
Ni, D., Xu, W., Zhu, Y., Pang, X., Lv, J., and Mu, W. (2021). Insight into the effects and biotechnological production of kestoses, the smallest fructooligosaccharides. Crit. Rev. Biotechnol. 41, 34–46. doi: 10.1080/07388551.2020.1844622
Nie, T., You, L., Tang, F., Duan, Y., Nepovimova, E., Kuca, K., et al. (2024). Microbiota-gut-brain axis in age-related neurodegenerative diseases. Curr. Neuropharmacol. doi: 10.2174/1570159X23666241101093436
Nikolova, V., Zaidi, S. Y., Young, A. H., Cleare, A. J., and Stone, J. M. (2019). Gut feeling: randomized controlled trials of probiotics for the treatment of clinical depression: systematic review and meta-analysis. Therapeutic Advan. Psychopharmacol. 9:2045125319859963. doi: 10.1177/2045125319859963
Nohr, M. K., Egerod, K. L., Christiansen, S. H., Gille, A., Offermanns, S., Schwartz, T. W., et al. (2015). Expression of the short chain fatty acid receptor GPR41/FFAR3 in autonomic and somatic sensory ganglia. Neuroscience 290, 126–137. doi: 10.1016/j.neuroscience.2015.01.040
Okuka, N., Milinkovic, N., Velickovic, K., Polovina, S., Sumarac-Dumanovic, M., Minic, R., et al. (2024). Beneficial effects of a new probiotic formulation on adipocytokines, appetite-regulating hormones, and metabolic parameters in obese women. Food Funct. 15, 7658–7668. doi: 10.1039/d4fo01269k
Palai, T., and Bhattacharya, P. K. (2013). Kinetics of lactose conversion to galacto-oligosaccharides by β-galactosidase immobilized on PVDF membrane. J. Biosci. Bioeng. 115, 668–673. doi: 10.1016/j.jbiosc.2012.12.014
Palai, T., Singh, A. K., and Bhattacharya, P. K. (2014). Enzyme, β-galactosidase immobilized on membrane surface for galacto-oligosaccharides formation from lactose: kinetic study with feed flow under recirculation loop. Biochem. Eng. J. 88, 68–76. doi: 10.1016/j.bej.2014.03.017
Palepu, M. S. K., Bhalerao, H. A., Sonti, R., and Dandekar, M. P. (2024). Faecalibacterium prausnitzii, FOS and GOS loaded synbiotic reverses treatment-resistant depression in rats: Restoration of gut-brain crosstalk. Eur. J. Pharmacol. 983:176960. doi: 10.1016/j.ejphar.2024.176960
Pan, H., Yang, S., Kulyar, M. F., Ma, H., Li, K., Zhang, L., et al. (2025). Lactobacillus fermentum 016 alleviates mice colitis by modulating oxidative stress, gut microbiota, and microbial metabolism. Nutrients 17:452. doi: 10.3390/nu17030452
Pappolla, M. A., Perry, G., Fang, X., Zagorski, M., Sambamurti, K., and Poeggeler, B. (2021). Indoles as essential mediators in the gut-brain axis. Their role in Alzheimer’s disease. Neurobiol. Dis. 156:105403. doi: 10.1016/j.nbd.2021.105403
Parnell, J. A., and Reimer, R. A. (2012). Prebiotic fibres dose-dependently increase satiety hormones and alter Bacteroidetes and Firmicutes in lean and obese JCR:LAcp rats. Br. J. Nutr. 107, 601–613. doi: 10.1017/S0007114511003163
Peña-Cardeña, A., Rodríguez-Alegría, M. E., Olvera, C., and Munguía, A. L. (2015). Synthesis of Fructooligosaccharides by IslA4, a truncated inulosucrase from Leuconostoc citreum. BMC Biotechnol. 15, 1–9. doi: 10.1186/s12896-015-0116-1
Peng, P., Peng, F., Bian, J., Xu, F., and Sun, R. (2011). Studies on the starch and hemicelluloses fractionated by graded ethanol precipitation from bamboo Phyllostachys bambusoides f. shouzhu Yi. J. Agric. Food Chem. 59, 2680–2688. doi: 10.1021/jf1045766
Peng, Y., Ma, Y., Luo, Z., Jiang, Y., Xu, Z., and Yu, R. (2023). Lactobacillus reuteri in digestive system diseases: focus on clinical trials and mechanisms. Front. Cell. Infect. Microbiol. 13:1254198. doi: 10.3389/fcimb.2023.1254198
Pham, M. L., Leister, T., Nguyen, H. A., Do, B. C., Pham, A. T., Haltrich, D., et al. (2017). Immobilization of β-galactosidases from Lactobacillus on chitin using a chitin-binding domain. J. Agric. Food Chem. 65, 2965–2976. doi: 10.1021/acs.jafc.6b04982
Pham, M. L., Tran, A. M., Kittibunchakul, S., Nguyen, T. T., Mathiesen, G., and Nguyen, T. H. (2019). Immobilization of β-galactosidases on the Lactobacillus cell surface using the peptidoglycan-binding motif LysM. Catalysts 9:443. doi: 10.3390/catal9050443
Pham, V. T., Lacroix, C., Braegger, C. P., and Chassard, C. (2016). Early colonization of functional groups of microbes in the infant gut. Environ. Microbiol. 18, 2246–2258. doi: 10.1111/1462-2920.13316
Piccioni, A., Covino, M., Candelli, M., Ojetti, V., Capacci, A., Gasbarrini, A., et al. (2023). How do diet patterns, single foods, prebiotics and probiotics impact gut microbiota? Microbiol. Res. 14, 390–408. doi: 10.3390/microbiolres14010030
Pinto-Sanchez, M. I., Hall, G. B., Ghajar, K., Nardelli, A., Bolino, C., Lau, J. T., et al. (2017). Probiotic Bifidobacterium longum NCC3001 reduces depression scores and alters brain activity: a pilot study in patients with irritable bowel syndrome. Gastroenterology 153, 448–459.e8. doi: 10.1053/j.gastro.2017.05.003
Platten, M., Nollen, E. A. A., and Röhrig, U. F. (2019). Tryptophan metabolism as a common therapeutic target in cancer, neurodegeneration and beyond. Nat. Rev. Drug Discov. 18, 379–401. doi: 10.1038/s41573-019-0016-5
Plaza-Diaz, J., Ruiz-Ojeda, F. J., Gil-Campos, M., and Gil, A. (2019). Mechanisms of action of probiotics. Adv. Nutr. 10, S49–S66. doi: 10.1093/advances/nmy063
Polakowski, C. B., Kato, M., Preti, V. B., Schieferdecker, M. E. M., and Ligocki Campos, A. C. (2019). Impact of the preoperative use of synbiotics in colorectal cancer patients: A prospective, randomized, double-blind, placebo-controlled study. Nutrition. 58:40–46. doi: 10.1016/j.nut.2018.06.004
Polyzos, K. A., and Ketelhuth, D. F. J. (2015). The role of the kynurenine pathway of tryptophan metabolism in cardiovascular disease. An emerging field. Hamostaseologie 35, 128–136. doi: 10.5482/HAMO-14-10-0052
Precup, G., Teleky, B. E., Ranga, F., and Vodnar, D. C. (2022). Assessment of physicochemical and rheological properties of xylo-oligosaccharides and glucose-enriched doughs fermented with BB-12. Biology 11:553. doi: 10.3390/biology11040553
Purnawan, A., Yopi, Y., and Irawadi, T. T. (2017). Production of manooligomannan from palm kernel cake by mannanase produced from Streptomyces cyaenus. Biosaintifika J. Biol. Biol. Educ. 9, 73–80. doi: 10.15294/biosaintifika.v9i1.9201
Qian, S., Zhou, J., Chen, X., Ji, W., Zhang, L., Hu, W., et al. (2020). Evaluation of an efficient fed-batch enzymatic hydrolysis strategy to improve production of functional xylooligosaccharides from maize straws. Ind. Crop. Prod. 157:112920. doi: 10.1016/j.indcrop.2020.112920
Qureshi, A., Umer, K., Tahir, S., Jebreen, A., Tahir, H., Ullah, R., et al. (2024). “Reduction of body fat and alteration of intestinal microbiota with probiotics and prebiotics in patients with obesity and overweight” in Gut heath, microbiota and animal diseases. ed. P. Liu (Faisalabad, Pakistan: Unique Scientific Publishers), 51–57.
Radford-Smith, D. E., and Anthony, D. C. (2023). Prebiotic and probiotic modulation of the microbiota–gut–brain axis in depression. Nutrients 15:1880. doi: 10.3390/nu15081880
Rahat-Rozenbloom, S., Fernandes, J., Cheng, J., and Wolever, T. M. S. (2017). Acute increases in serum colonic short-chain fatty acids elicited by inulin do not increase GLP-1 or PYY responses but may reduce ghrelin in lean and overweight humans. Eur. J. Clin. Nutr. 71, 953–958. doi: 10.1038/ejcn.2016.249
Rahmannia, M., Poudineh, M., Mirzaei, R., Aalipour, M. A., Shahidi Bonjar, A. H., Goudarzi, M., et al. (2024). Strain-specific effects of probiotics on depression and anxiety: a meta-analysis. Gut Pathog. 16:46. doi: 10.1186/s13099-024-00634-8
Rajagopalan, G., Shanmugavelu, K., and Yang, K. L. (2017). Production of prebiotic-xylooligosaccharides from alkali pretreated mahogany and mango wood sawdust by using purified xylanase of Clostridium strain BOH3. Carbohydr. Polym. 167, 158–166. doi: 10.1016/j.carbpol.2017.03.021
Ranadheera, C., Vidanarachchi, J., Rocha, R., Cruz, A., and Ajlouni, S. (2017). Probiotic delivery through fermentation: dairy vs. non-dairy beverages. Fermentation 3:67. doi: 10.3390/fermentation3040067
Rangel-Torres, B. E., García-Montoya, I. A., Rodríguez-Tadeo, A., and Jiménez-Vega, F. (2024). The symbiosis between Lactobacillus acidophilus and inulin: metabolic benefits in an obese murine model. Probiotics Antimicrob. Proteins 16, 26–34. doi: 10.1007/s12602-022-10012-y
Rathamat, Z., Choorit, W., Chisti, Y., and Prasertsan, P. (2021). Two-step isolation of hemicellulose from oil palm empty fruit bunch fibers and its use in production of xylooligosaccharide prebiotic. Ind. Crop. Prod. 160:113124. doi: 10.1016/j.indcrop.2020.113124
Reddy, S. S., and Krishnan, C. (2016). Production of xylooligosaccharides in SSF by Bacillus subtilis KCX006 producing β-xylosidase-free endo-xylanase and multiple xylan debranching enzymes. Prep. Biochem. Biotechnol. 46, 49–55. doi: 10.1080/10826068.2014.970694
Reichardt, N., Duncan, S. H., Young, P., Belenguer, A., McWilliam Leitch, C., Scott, K. P., et al. (2014). Phylogenetic distribution of three pathways for propionate production within the human gut microbiota. ISME J. 8, 1323–1335. doi: 10.1038/ismej.2014.14
Reque, P. M., Pinilla, C. M. B., Gautério, G. V., Kalil, S. J., and Brandelli, A. (2019). Xylooligosaccharides production from wheat middlings bioprocessed with Bacillus subtilis. Food Res. Int. 126:108673. doi: 10.1016/j.foodres.2019.108673
Roager, H. M., Hansen, L. B., Bahl, M. I., Frandsen, H. L., Carvalho, V., Gøbel, R. J., et al. (2016). Colonic transit time is related to bacterial metabolism and mucosal turnover in the gut. Nat. Microbiol. 1:16093. doi: 10.1038/nmicrobiol.2016.93
Roager, H. M., and Licht, T. R. (2018). Microbial tryptophan catabolites in health and disease. Nat. Commun. 9:3294. doi: 10.1038/s41467-018-05470-4
Robertson, M. D., Bickerton, A., Dennis, A. L., Vidal, H., and Frayn, K. N. (2005). Insulin-sensitizing effects of dietary resistant starch and effects on skeletal muscle and adipose tissue metabolism. Am. J. Clin. Nutr. 82, 559–567. doi: 10.1093/ajcn.82.3.559
Rodrigues, H. G., Takeo Sato, F., Curi, R., and Vinolo, M. A. R. (2016). Fatty acids as modulators of neutrophil recruitment, function and survival. Eur. J. Pharmacol. 785, 50–58. doi: 10.1016/j.ejphar.2015.03.098
Sabater, C., Calvete-Torre, I., Villamiel, M., Moreno, F. J., Margolles, A., and Ruiz, L. (2021). Vegetable waste and by-products to feed a healthy gut microbiota: current evidence, machine learning and computational tools to design novel microbiome-targeted foods. Trends Food Sci. Technol. 118, 399–417. doi: 10.1016/j.tifs.2021.10.002
Sabiha-Hanim, S., Noor, M. A. M., and Rosma, A. (2011). Effect of autohydrolysis and enzymatic treatment on oil palm (Elaeis guineensis Jacq.) frond fibres for xylose and xylooligosaccharides production. Bioresour. Technol. 102, 1234–1239. doi: 10.1016/j.biortech.2010.08.017
Sakurai, K., Toshimitsu, T., Okada, E., Anzai, S., Shiraishi, I., Inamura, N., et al. (2022). Effects of Lactiplantibacillus plantarum OLL2712 on memory function in older adults with declining memory: a randomized placebo-controlled trial. Nutrients 14:4300. doi: 10.3390/nu14204300
Samanta, A. K., Kolte, A. P., Elangovan, A. V., Dhali, A., Senani, S., Sridhar, M., et al. (2016). Value addition of corn husks through enzymatic production of xylooligosaccharides. Braz. Arch. Biol. Technol. 59:e16160078. doi: 10.1590/1678-4324-2016160078
Sanders, M. E., Merenstein, D. J., Reid, G., Gibson, G. R., and Rastall, R. A. (2019). Probiotics and prebiotics in intestinal health and disease: from biology to the clinic. Nat. Rev. Gastroenterol. Hepatol. 16, 605–616. doi: 10.1038/s41575-019-0173-3
Sarita, B., Samadhan, D., Hassan, M. Z., and Kovaleva, E. G. (2025). A comprehensive review of probiotics and human health-current prospective and applications. Front. Microbiol. 15:1487641. doi: 10.3389/fmicb.2024.1487641
Sathitkowitchai, W., Sathapondecha, P., Angthong, P., Srimarut, Y., Malila, Y., Nakkongkam, W., et al. (2022). Isolation and characterization of mannanase-producing bacteria for potential synbiotic application in shrimp farming. Animals 12:2583. doi: 10.3390/ani12192583
Schiering, C., Wincent, E., Metidji, A., Iseppon, A., Li, Y., Potocnik, A. J., et al. (2017). Feedback control of AHR signalling regulates intestinal immunity. Nature 542, 242–245. doi: 10.1038/nature21080
Schmitt, L. M., Smith, E. G., Pedapati, E. V., Horn, P. S., Will, M., Lamy, M., et al. (2023). Results of a phase Ib study of SB-121, an investigational probiotic formulation, a randomized controlled trial in participants with autism spectrum disorder. Sci. Rep. 13:5192. doi: 10.1038/s41598-023-30909-0
Schonfeld, P., and Wojtczak, L. (2016). Short- and medium- chain fatty acids in energy metabolism: the cellular perspective. J. Lipid Res. 57, 943–954. doi: 10.1194/jlr.R067629
Schwab, C., Ruscheweyh, H. J., Bunesova, V., Pham, V. T., Beerenwinkel, N., and Lacroix, C. (2017). Trophic interactions of infant bifidobacteria and Eubacterium hallii during L-fucose and fucosyllactose degradation. Front. Microbiol. 8:95. doi: 10.3389/fmicb.2017.00095
Seesuriyachan, P., Kawee-Ai, A., and Chaiyaso, T. (2017). Green and chemical-free process of enzymatic xylooligosaccharide production from corncob: enhancement of the yields using a strategy of lignocellulosic destructuration by ultra-high pressure pretreatment. Bioresour. Technol. 241, 537–544. doi: 10.1016/j.biortech.2017.05.193
Selma-Royo, M., Tarrazó, M., García-Mantrana, I., Gómez-Gallego, C., Salminen, S., and Collado, M. C. (2019). Shaping microbiota during the first 1000 days of life. Adv. Exp. Med. Biol. 1125, 3–24. doi: 10.1007/5584_2018_312
Sempach, L., Doll, J. P. K., Limbach, V., Marzetta, F., Schaub, A. C., Schneider, E., et al. (2024). Examining immune-inflammatory mechanisms of probiotic supplementation in depression: secondary findings from a randomized clinical trial. Transl. Psychiatry 14:305. doi: 10.1038/s41398-024-03030-7
Shah, A. B., Cho, H., and Shim, S. H. (2025). Exploring the bioactive landscape: peptides and non-peptides from the human microbiota. NPJ Biofilms Microbiomes 11:76. doi: 10.1038/s41522-025-00713-x
Shah, A. B., and Shim, S. H. (2025). Human microbiota peptides: important roles in human health. Nat. Prod. Rep. 42, 151–194. doi: 10.1039/d4np00042k
Shahin, N. N., Ahmed-Farid, O. A., Sakr, E. A. E., Kamel, E. A., and Mohamed, M. M. (2025). Oral supplements of combined Lactobacillus plantarum and Asparagus officinalis modulate gut microbiota and alleviate high-fat diet–induced cognitive deficits and neurodegeneration in rats. Probiotics Antimicrob. Proteins. doi: 10.1007/s12602-024-10429-7
Shinde, T., Hansbro, P. M., Sohal, S. S., Dingle, P., Eri, R., Stanley, R., et al. (2020). Microbiota modulating nutritional approaches to countering the effects of viral respiratory infections including SARS-CoV-2 through promoting metabolic and immune fitness with probiotics and plant bioactives. Microorganisms 8:921. doi: 10.3390/microorganisms8060921
Silva, M. F., Rigo, D., Mossi, V., Golunski, S., de Oliveira Kuhn, G., Di Luccio, M., et al. (2013). Enzymatic synthesis of fructooligosaccharides by inulinases from Aspergillus niger and Kluyveromyces marxianus NRRL Y-7571 in aqueous–organic medium. Food Chem. 138, 148–153. doi: 10.1016/j.foodchem.2012.09.118
Singh, R. D., Nadar, C. G., Muir, J., and Arora, A. (2019). Green and clean process to obtain low degree of polymerisation xylooligosaccharides from almond shell. J. Clean. Prod. 241:118237. doi: 10.1016/j.jclepro.2019.118237
Sivan, A., Corrales, L., Hubert, N., Williams, J. B., Aquino-Michaels, K., Earley, Z. M., et al. (2015). Commensal Bifidobacterium promotes antitumor immunity and facilitates anti–PD-L1 efficacy. Science 350, 1084–1089. doi: 10.1126/science.aac4255
Slade, E., Williams, L., and Gagnon, J. (2018). Hydrogen sulfide suppresses ghrelin secretion in vitro and delays postprandial ghrelin secretion while reducing appetite in mice. Phys. Rep. 6:e13870. doi: 10.14814/phy2.13870
Slomiany, B. L., and Slomiany, A. (2017). Role of LPS-elicited signaling in triggering gastric mucosal inflammatory responses to H. pylori: modulatory effect of ghrelin. Inflammopharmacology 25, 415–429. doi: 10.1007/s10787-017-0360-1
Slykerman, R. F., Davies, N., Vlckova, K., O’Riordan, K. J., Bassett, S. A., Dekker, J., et al. (2025). Precision psychobiotics for gut–brain axis health: advancing the discovery pipelines to deliver mechanistic pathways and proven health efficacy. Microb. Biotechnol. 18:e70079. doi: 10.1111/1751-7915.70079
Smaali, I., Jazzar, S., Soussi, A., Muzard, M., Aubry, N., and Marzouki, M. N. (2012). Enzymatic synthesis of fructooligosaccharides from date by-products using an immobilized crude enzyme preparation of β-D-fructofuranosidase from Aspergillus awamori NBRC 4033. Biotechnol. Bioprocess Eng. 17, 385–392. doi: 10.1007/s12257-011-0388-9
Smith, E. A., and Macfarlane, G. T. (1996). Enumeration of human colonic bacteria producing phenolic and indolic compounds: effects of pH, carbohydrate availability and retention time on dissimilatory aromatic amino acid metabolism. J. Appl. Bacteriol. 81, 288–302. doi: 10.1111/j.1365-2672.1996.tb04331.x
Soliman, A. B., Hassan, M. H., Huan, T. N., Abugable, A. A., Elmehalmey, W. A., Karakalos, S. G., et al. (2017). Pt immobilization within a tailored porous-organic polymer–graphene composite: opportunities in the hydrogen evolving reaction. ACS Catal. 7, 7847–7854. doi: 10.1021/acscatal.7b02246
Spencer, K. D., Bline, H., Chen, H. J., Verosky, B. G., Hilt, M. E., Jaggers, R. M., et al. (2024). Modulation of anxiety-like behavior in galactooligosaccharide-fed mice: a potential role for bacterial tryptophan metabolites and reduced microglial reactivity. Brain Behav. Immun. 121, 229–243. doi: 10.1016/j.bbi.2024.07.024
Steinert, R. E., Ullrich, S. S., Geary, N., Asarian, L., Bueter, M., Horowitz, M., et al. (2017). Comparative effects of intraduodenal amino acid infusions on food intake and gut hormone release in healthy males. Phys. Rep. 5:e13492. doi: 10.14814/phy2.13492
Suez, J., Zmora, N., Segal, E., and Elinav, E. (2019). The pros, cons, and many unknowns of probiotics. Nat. Med. 25, 716–729. doi: 10.1038/s41591-019-0439-x
Surek, E., Buyukkileci, A. O., and Yegin, S. (2021). Processing of hazelnut (Corylus avellana L.) shell autohydrolysis liquor for production of low molecular weight xylooligosaccharides by Aureobasidium pullulans NRRL Y–2311–1 xylanase. Ind. Crop. Prod. 161:113212. doi: 10.1016/j.indcrop.2020.113212
Su, Y., Fang, L., Wang, P., Lai, C., Huang, C., Ling, Z., et al. (2021). Efficient production of xylooligosaccharides rich in xylobiose and xylotriose from poplar by hydrothermal pretreatment coupled with post-enzymatic hydrolysis. Bioresour. Technol. 342:125955. doi: 10.1016/j.biortech.2021.125955
Suzuki, H., Matsuzaki, J., and Hibi, T. (2011). Ghrelin and oxidative stress in gastrointestinal tract. J. Clin. Biochem. Nutr. 48, 122–125. doi: 10.3164/jcbn.10-16GFR
Swanson, K. S., Gibson, G. R., Hutkins, R., Reimer, R. A., Reid, G., Verbeke, K., et al. (2020). The international scientific association for probiotics and prebiotics (ISAPP) consensus statement on the definition and scope of synbiotics. Nat. Rev. Gastroenterol. Hepatol. 17, 687–701. doi: 10.1038/s41575-020-0344-2
Szymczak-Pajor, I., Drzewoski, J., Kozłowska, M., Krekora, J., and Śliwińska, A. (2025). The gut microbiota-related antihyperglycemic effect of metformin. Pharmaceuticals 18:55. doi: 10.3390/ph18010055
Tae, H., and Kim, T. (2024). The effect of prebiotic and probiotic food consumption on anxiety severity: a nationwide study in Korea. Front. Nutr. 11:1385518. doi: 10.3389/fnut.2024.1385518
Tanaka, M., Szabó, Á., and Vécsei, L. (2024). Redefining roles: A paradigm shift in tryptophan–kynurenine metabolism for innovative clinical applications. Int. J. Mol. Sci. 25:12767. doi: 10.3390/ijms252312767
Tavakoly, R., Hadi, A., Rafie, N., Talaei, B., Marx, W., and Arab, A. (2021). Effect of probiotic consumption on immune response in athletes: A meta-analysis. Int. J. Sports Med. 42, 769–781. doi: 10.1055/a-1463-3303
Tolhurst, G., Heffron, H., Lam, Y. S., Parker, H. E., Habib, A. M., Diakogiannaki, E., et al. (2012). Short- chain fatty acids stimulate glucagon- like peptide-1 secretion via the G- proteincoupled receptor FFAR2. Diabetes 61, 364–371. doi: 10.2337/db11-1019
Topping, D. L., and Clifton, P. M. (2001). Short-chain fatty acids and human colonic function: roles of resistant starch and nonstarch polysaccharides. Physiol. Rev. 81, 1031–1064. doi: 10.1152/physrev.2001.81.3.1031
Torres-Fuentes, C., Golubeva, A. V., Zhdanov, A. V., Wallace, S., Arboleya, S., Papkovsky, D. B., et al. (2019). Short-chain fatty acids and microbiota metabolites attenuate ghrelin receptor signaling. FASEB J. 33, 13546–13559. doi: 10.1096/fj.201901433R
Tóth, A. G., Csabai, I., Judge, M. F., Maróti, G., Becsei, Á., Spisák, S., et al. (2021). Mobile antimicrobial resistance genes in probiotics. Antibiotics 10:1287. doi: 10.3390/antibiotics10111287
Tsai, M. L., Shen, S. P., Chen, Y. T., Chiu, H. Y., Lin, H. Y., Cheng, H. W., et al. (2025). Effects of phototherapy combined with Lactobacillus salivarius AP-32 or Bifidobacterium animalis subsp. lactis CP-9 on improving neonatal jaundice and gut microbiome health: a randomized double-blind clinical study. Nutr. J. 24:73. doi: 10.1186/s12937-025-01126-4
Usvalampi, A., Maaheimo, H., Tossavainen, O., and Frey, A. D. (2018). Enzymatic synthesis of fucose-containing galacto-oligosaccharides using β-galactosidase and identification of novel disaccharide structures. Glycoconj. J. 35, 31–40. doi: 10.1007/s10719-017-9794-3
Valladares, R., Bojilova, L., Potts, A. H., Cameron, E., Gardner, C., Lorca, G., et al. (2013). Lactobacillus johnsonii inhibits indoleamine 2,3-dioxygenase and alters tryptophan metabolite levels in biobreeding rats. FASEB J. 27, 1711–1720. doi: 10.1096/fj.12-223339
van der Beek, C. M., Dejong, C. H. C., Troost, F. J., Masclee, A. A. M., and Lenaerts, K. (2017). Role of short-chain fatty acids in colonic inflammation, carcinogenesis, and mucosal protection and healing. Nutr. Rev. 75, 286–305. doi: 10.1093/nutrit/nuw067
Vera, C., Guerrero, C., Conejeros, R., and Illanes, A. (2012). Synthesis of galacto-oligosaccharides by β-galactosidase from Aspergillus oryzae using partially dissolved and supersaturated solution of lactose. Enzym. Microb. Technol. 50, 188–194. doi: 10.1016/j.enzmictec.2011.12.003
Verma, J., Anwar, M. T., Linz, B., Backert, S., and Pachathundikandi, S. K. (2025). The influence of gastric microbiota and probiotics in Helicobacter pylori infection and associated diseases. Biomedicine 13:61. doi: 10.3390/biomedicines13010061
Vieira-Silva, S., Falony, G., Darzi, Y., Lima-Mendez, G., Garcia Yunta, R., Okuda, S., et al. (2016). Species-function relationships shape ecological properties of the human gut microbiome. Nat. Microbiol. 1:16088. doi: 10.1038/nmicrobiol.2016.88
Wang, X., Chen, L., Zhang, C., Shi, Q., Zhu, L., Zhao, S., et al. (2024). Effect of probiotics at different intervention time on glycemic control in patients with type 2 diabetes mellitus: a systematic review and meta-analysis. Front. Endocrinol. 15:1392306. doi: 10.3389/fendo.2024.1392306
Wong, J. M. W., de Souza, R., Kendall, C. W. C., Emam, A., and Jenkins, D. J. A. (2006). Colonic health: fermentation and short chain fatty acids. J. Clin. Gastroenterol. 40, 235–243. doi: 10.1097/00004836-200603000-00015
Xiao, Y., Huang, L., Zhao, J., Chen, W., and Lu, W. (2025). The gut core microbial species Bifidobacterium longum: colonization, mechanisms, and health benefits. Microbiol. Res. 290:127966. doi: 10.1016/j.micres.2024.127966
Xiong, R. G., Zhou, D. D., Wu, S. X., Huang, S. Y., Saimaiti, A., Yang, Z. J., et al. (2022). Health benefits and side effects of short-chain fatty acids. Foods 11:2863. doi: 10.3390/foods11182863
Xiong, Y., Miyamoto, N., Shibata, K., Valasek, M. A., Motoike, T., Kedzierski, R. M., et al. (2004). Short-chain fatty acids stimulate leptin production in adipocytes through the G protein coupled receptor GPR41. Proc. Natl. Acad. Sci. USA 101, 1045–1050. doi: 10.1073/pnas.2637002100
Xi, Z., Fenglin, X., Yun, Z., and Chunrong, L. (2025). Efficacy of probiotics in the treatment of allergic diseases: a meta-analysis. Front. Nutr. 12:1502390. doi: 10.3389/fnut.2025.1502390
Yanagi, H., Tsuda, A., Matsushima, M., Takahashi, S., Ozawa, G., Koga, Y., et al. (2017). Changes in the gut microbiota composition and the plasma ghrelin level in patients with Helicobacter pylori-infected patients with eradication therapy. BMJ Open Gastroenterol. 4:182. doi: 10.1136/bmjgast-2017-000182
Yan, F., Li, N., Shi, J., Li, H., Yue, Y., Jiao, W., et al. (2019). Lactobacillus acidophilus alleviates type 2 diabetes by regulating hepatic glucose, lipid metabolism and gut microbiota in mice. Food Funct. 10, 5804–5815. doi: 10.1039/C9FO01062A
Yang, F., Li, J., Pang, G., Ren, F., and Fang, B. (2019). Effects of diethyl phosphate, a nonspecific metabolite of organophosphorus pesticides, on serum lipid, hormones, inflammation, and gut microbiota. Molecules 24:2003. doi: 10.3390/molecules24102003
Yang, H., Wang, K., Song, X., and Xu, F. (2011). Production of xylooligosaccharides by xylanase from Pichia stipitis based on xylan preparation from triploid Populas tomentosa. Bioresour. Technol. 102, 7171–7176. doi: 10.1016/j.biortech.2011.03.110
Yang, J., Gao, R., Zhou, Y., Anankanbil, S., Li, J., Xie, G., et al. (2018). β-Glucosidase from Thermotoga naphthophila RKU-10 for exclusive synthesis of galactotrisaccharides: kinetics and thermodynamics insight into reaction mechanism. Food Chem. 240, 422–429. doi: 10.1016/j.foodchem.2017.07.155
Yano, J. M., Yu, K., Donaldson, G. P., Shastri, G. G., Ann, P., Ma, L., et al. (2015). Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell 161, 264–276. doi: 10.1016/j.cell.2015.02.047
Yan, Q., Wang, W., Fan, Z., Wei, Y., Yu, R., Pan, T., et al. (2025). Chickpea-resistant starch exhibits bioactive function for alleviating atopic dermatitis via regulating butyrate production. Int. J. Biol. Macromol. 303:140661. doi: 10.1016/j.ijbiomac.2025.140661
Yegin, S. (2023). Microbial xylanases in xylooligosaccharide production from lignocellulosic feedstocks. Biomass Convers. Biorefinery 13, 3619–3658. doi: 10.1007/s13399-022-03190-w
Yi, C., Huang, S., Zhang, W., Guo, L., Xia, T., Huang, F., et al. (2025). Synergistic interactions between gut microbiota and short chain fatty acids: pioneering therapeutic frontiers in chronic disease management. Microb. Pathog. 199:107231. doi: 10.1016/j.micpath.2024.107231
Yin, J., Han, H., Li, Y., Liu, Z., Zhao, Y., Fang, R., et al. (2018a). Lysine restriction affects feed intake and amino acid metabolism via gut microbiome in piglets. Cell. Physiol. Biochem. 44, 1749–1761. doi: 10.1159/000485782
Yin, J., Li, Y., Han, H., Liu, Z., Zeng, X., Li, T., et al. (2018b). Long-term effects of lysine concentration on growth performance, intestinal microbiome, and metabolic profiles in a pig model. Food Funct. 9, 4153–4163. doi: 10.1039/c8fo00973b
Yu, L., and O’sullivan, D. J. (2014). Production of galactooligosaccharides using a hyperthermophilic β-galactosidase in permeabilized whole cells of Lactococcus lactis. J. Dairy Sci. 97, 694–703. doi: 10.3168/jds.2013-7492
Zagórska, A., Marcinkowska, M., Jamrozik, M., Wiśniowska, B., and Paśko, P. (2020). From probiotics to psychobiotics - the gut-brain axis in psychiatric disorders. Benefic. Microbes 11, 717–732. doi: 10.3920/BM2020.0063
Zare, D., Aryaee, H., Mirdamadi, S., and Shirkhan, F. (2024). The benefits and applications of Lactobacillus plantarum in food and health: a narrative review. Iran. J. Public Health 53, 2201–2213. doi: 10.18502/ijph.v53i10.16698
Zelante, T., Iannitti, R. G., Cunha, C., De Luca, A., Giovannini, G., Pieraccini, G., et al. (2013). Tryptophan catabolites from microbiota engage aryl hydrocarbon receptor and balance mucosal reactivity via interleukin-22. Immunity 39, 372–385. doi: 10.1016/j.immuni.2013.08.003
Ze, X., Duncan, S. H., Louis, P., and Flint, H. J. (2012). Ruminococcus bromii is a keystone species for the degradation of resistant starch in the human colon. ISME J. 6, 1535–1543. doi: 10.1038/ismej.2012.4
Zhang, D., Jian, Y. P., Zhang, Y. N., Li, Y., Gu, L. T., Sun, H. H., et al. (2023). Short-chain fatty acids in diseases. Cell Commun. Signal 21:212. doi: 10.1186/s12964-023-01219-9
Zhang, S., Nie, Q., Sun, Y., Zuo, S., Chen, C., Li, S., et al. (2024). Bacteroides uniformis degrades β-glucan to promote Lactobacillus johnsonii improving indole-3-lactic acid levels in alleviating colitis. Microbiome 12:177. doi: 10.1186/s40168-024-01896-9
Zhao, J., Liu, J., Feng, J., Liu, X., and Hu, Q. (2024). The gut microbiota-brain connection: insights into major depressive disorder and bipolar disorder. Front. Psychol. 15:1421490. doi: 10.3389/fpsyt.2024.1421490
Zhao, P., Chen, Y., Zhou, S., and Li, F. (2025). Microbial modulation of tryptophan metabolism links gut microbiota to disease and its treatment. Pharmacol. Res. 219:107896. doi: 10.1016/j.phrs.2025.107896
Keywords: probiotic, prebiotic, synbiotic, gut-brain axis, health
Citation: Kezer G, Paramithiotis S, Khwaldia K, Harahap IA, Čagalj M, Šimat V, Smaoui S, Elfalleh W, Ozogul F and Esatbeyoglu T (2025) A comprehensive overview of the effects of probiotics, prebiotics and synbiotics on the gut-brain axis. Front. Microbiol. 16:1651965. doi: 10.3389/fmicb.2025.1651965
Edited by:
João Miguel Rocha, Laboratory of Process Engineering, Environment, Biotechnology and Energy, Faculty of Engineering, University of Porto, PortugalReviewed by:
Kimon Andreas Karatzas, University of Reading, United KingdomAbdul Bari Shah, Al-Farabi Kazakh National University, Kazakhstan
Copyright © 2025 Kezer, Paramithiotis, Khwaldia, Harahap, Čagalj, Šimat, Smaoui, Elfalleh, Ozogul and Esatbeyoglu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Tuba Esatbeyoglu, ZXNhdGJleW9nbHVAZm9oLnVuaS1oYW5ub3Zlci5kZQ==