 Marta Estévez-Rodríguez
Marta Estévez-Rodríguez Guillermo Sánchez-Garrido Campos
Guillermo Sánchez-Garrido Campos Ángela M. Zafra
Ángela M. Zafra Isabel Cordones
Isabel Cordones Javier Márquez-Ruiz*
Javier Márquez-Ruiz*- Department of Physiology, Anatomy and Cell Biology, Pablo de Olavide University, Seville, Spain
The balance between excitatory and inhibitory (E/I) activity is a fundamental property of neural circuits, ensuring precise information processing and preventing pathological states such as hyperexcitability or network silencing. Disruptions in this balance have been linked to several neurological and psychiatric disorders, including epilepsy, autism, and schizophrenia. Transcranial electrical stimulation (tES) can modulate the E/I balance through mechanisms that affect synaptic plasticity, neurotransmitter systems, and network synchronization. The main tES modalities—transcranial direct current stimulation (tDCS), transcranial alternating current stimulation (tACS), and transcranial random noise stimulation (tRNS)—operate through distinct physiological principles, enabling the modulation of neuronal excitability and oscillatory dynamics. Animal models offer controlled experimental conditions to study the effects of tES on E/I regulation at the cellular, synaptic, and network levels. Preclinical research has revealed polarity-dependent plasticity with tDCS, frequency-specific entrainment with tACS, and GABAergic modulation with tRNS. These findings are essential for validating computational models and refining stimulation protocols. Future studies should integrate multimodal technologies to enhance the translational relevance of tES and develop personalized neuromodulation strategies targeting E/I imbalance in brain disorders.
Introduction
The balance between excitatory and inhibitory (E/I) activity in the brain is essential for normal brain function and neural activity (Tatti et al., 2017). Cortical neurons integrate glutamatergic and GABAergic inputs to maintain optimal membrane potentials and enable adaptive responses (Landau et al., 2016). Disruptions in this balance contribute to disorders such as epilepsy, autism, and schizophrenia (Eichler and Meier, 2008; Yizhar et al., 2011; Nguyen et al., 2016). The E/I balance is shaped by synaptic plasticity mechanisms proposed by Hebb (1949) and later extended to include inhibitory circuits (Saraga et al., 2008; D’amour and Froemke, 2015). Non-invasive stimulation techniques like tDCS, tACS, and tRNS may help restore E/I balance depending on brain region and context (Krause et al., 2013; Van Bueren et al., 2023).
Transcranial electrical stimulation (tES) is a non-invasive technique that applies weak electrical currents to the scalp, modulating cortical excitability and inducing immediate and lasting effects on brain function (Nitsche and Paulus, 2000; Woods et al., 2016). The main tES modalities include transcranial direct current stimulation (tDCS), which uses low-intensity direct currents to modulate excitability depending on polarity, and transcranial alternating current stimulation (tACS), which applies oscillatory currents to influence brain rhythms (Paulus, 2011). tDCS has been widely studied in motor learning, cognition, depression, and stroke (Nitsche and Paulus, 2000; Nitsche et al., 2008; Brunoni et al., 2012), while tACS shows promise in conditions such as schizophrenia, epilepsy, and Alzheimer’s disease (Herrmann et al., 2013; Antal and Paulus, 2013; Kasten et al., 2016). A third variant, transcranial random noise stimulation (tRNS), delivers broadband random currents that enhance neuroplasticity and show promise in perception, memory, and clinical applications (Fertonani et al., 2011; Snowball et al., 2013; Van Der Groen and Wenderoth, 2016). The therapeutic potential of tES may lie in its ability to modulate fundamental neural processes, such as the balance between excitatory and inhibitory (E/I) activity in the brain (Krause et al., 2013).
Given the limitations of studying E/I balance directly in humans, animal models are essential for uncovering the physiological and molecular effects of tES. They offer controlled conditions to examine both immediate and long-term effects (Sánchez-León et al., 2018) and allow systematic variation of parameters such as intensity, frequency, and duration. Furthermore, they permit invasive assessments of neuronal activity, neurotransmission, and plasticity-related pathways (Jackson et al., 2016).
This mini-review highlights the contribution of animal models to uncovering how tES modulates the E/I balance, offering controlled settings to dissect the underlying physiological mechanisms.
Transcranial electrical stimulation in animal models
Animal models enable researchers to adapt tES protocols to non-human species, optimizing experimental conditions by adjusting electrode placement, current intensity, and stimulation duration to account for anatomical and physiological differences between animals and humans. To achieve targeted stimulation, researchers use smaller electrode sizes and precise skull placement, while higher current densities help compensate for differences in brain volume and conductivity (Márquez-Ruiz et al., 2014; Jackson et al., 2016).
Animal models offer versatility for studying tES across multiple levels with both in vitro and in vivo approaches. In vitro models, especially brain slices, yield insights into cellular and synaptic mechanisms, including membrane polarization, synaptic plasticity, and neurotransmitter dynamics (Bikson et al., 2004, 2012; Radman et al., 2009; Kabakov et al., 2012). In vivo models enable exploration of network and behavioral effects. Experiments in anesthetized animals permit detailed circuit-level analysis and connectivity changes (Ozen et al., 2010; Opitz et al., 2016; Vöröslakos et al., 2018), while awake animals provide key insights into neuromodulatory effects on behavior and cognition (Márquez-Ruiz et al., 2012; Monai et al., 2016; Krause et al., 2017; Sánchez-León et al., 2018, 2021a).
By integrating cutting-edge techniques such as high-density electrophysiology (Krause et al., 2022; Farahani et al., 2024; Sánchez-León et al., 2025), two-photon imaging (Monai et al., 2016; Gellner et al., 2021), optogenetics (Fröhlich and McCormick, 2010; Mabil et al., 2020; Huang et al., 2021), and chemogenetics (Su et al., 2024), animal models provide unparalleled opportunities to investigate the effects of tES. These modern approaches, when combined with classical electrophysiological techniques—including intracellular and extracellular recordings—offer unprecedented resolution for dissecting stimulation-induced changes. This multimodal strategy enables the investigation of tES-induced alterations at both the cellular and network levels, advancing our understanding of its mechanisms and potential therapeutic applications (Table 1).
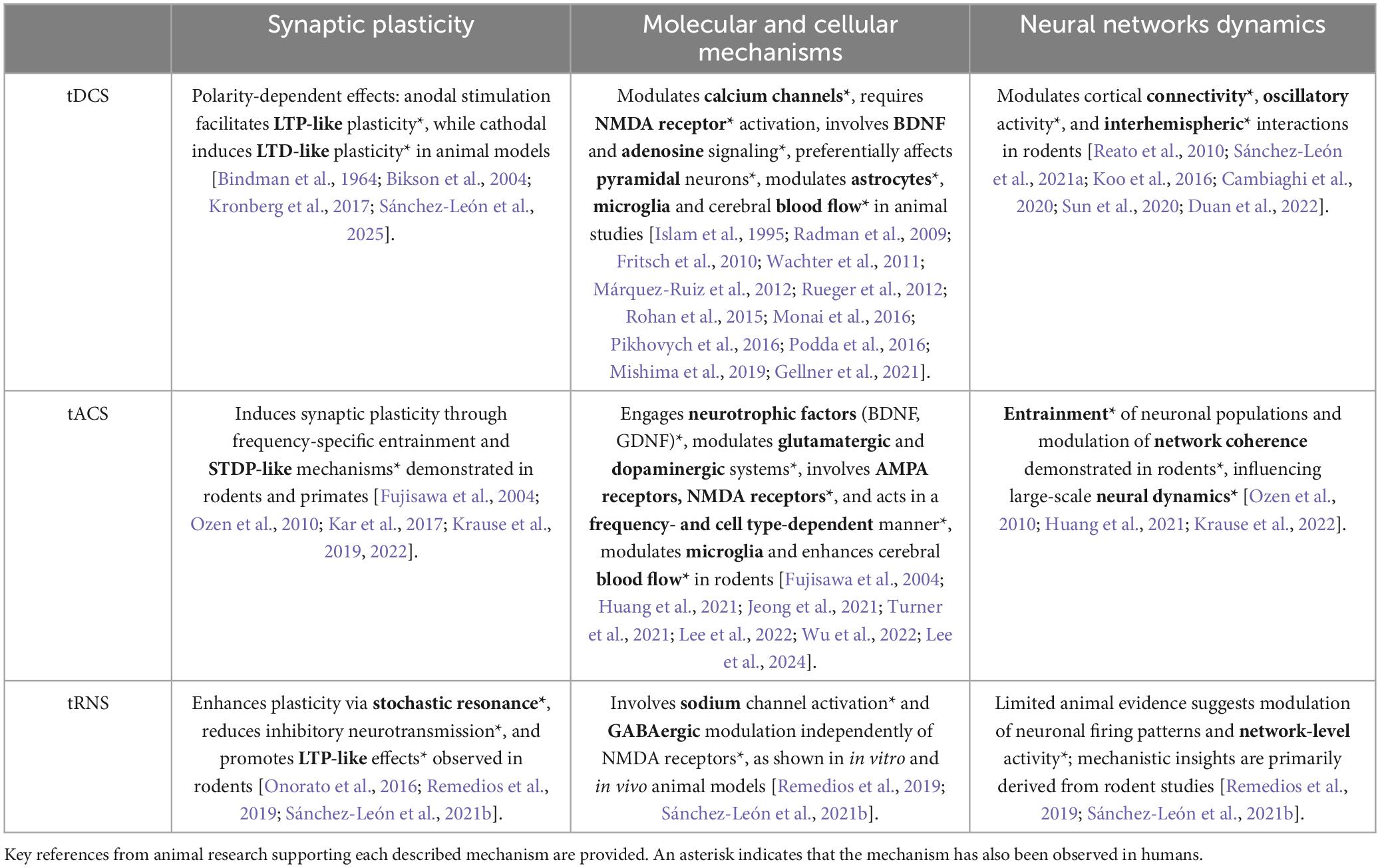
Table 1. Summary of the impact of transcranial electrical stimulation techniques (tDCS, tACS, tRNS) on synaptic plasticity, molecular and cellular mechanisms, and neural network dynamics based on animal model studies.
Animal models bridge basic and clinical research, enabling disease-specific insights (Sánchez-Garrido Campos et al., 2025). In Alzheimer’s models, gamma-frequency optogenetic stimulation of parvalbumin interneurons improved memory and synaptic plasticity, likely by reducing amyloid-beta (Iaccarino et al., 2016; Etter et al., 2019). Similarly, gamma-tACS applied for 20 min restored impaired LTP in mice (Jeong et al., 2021). Although the current density used in this animal study was not reported, human studies applying gamma-tACS for longer durations (60 min) have shown significant improvements in memory performance in patients with mild cognitive impairment due to Alzheimer’s disease (Benussi et al., 2021), supporting the direction of the findings observed in animal models. In epilepsy, cathodal tDCS (3.54 mA/cm2, 25 min) reduced seizures and restored E/I balance in anesthetized rodents (Sun et al., 2020). In clinical settings, cathodal tDCS applied for a similar duration (30 min) but with markedly lower current density (0.057 mA/cm2) has been shown to reduce seizure duration in patients with mesial temporal lobe epilepsy and hippocampal sclerosis (Tekturk et al., 2016). In Parkinson’s models, anodal tDCS (0.88 mA/cm2, 20 min) enhanced motor function through dopaminergic activation (Tamura et al., 2024). Comparable stimulation in patients—anodal tDCS over the motor cortex for 20 min—has led to significant improvements in gait speed, step length, and cadence, although at a substantially lower current density (0.057 mA/cm2) (Schabrun et al., 2016). While tRNS has shown promise in humans (Van Der Groen et al., 2022), its mechanisms remain unclear due to scarce preclinical data (Antal and Herrmann, 2016).
Building on the methodological framework established in animal models, we next investigate how tES modulates the E/I balance through synaptic-level mechanisms.
Impact of tES on E/I balance at the synaptic level
tES can modulate synaptic plasticity mechanisms essential for maintaining the E/I balance. This balance is regulated by glutamate and GABA, acting through their respective receptors (Isaacson and Scanziani, 2011), and is fine-tuned by neuromodulators like acetylcholine, dopamine, and serotonin (Froemke, 2015). Disruptions in E/I balance contribute to neurological and psychiatric disorders (Landau et al., 2016). By influencing glutamatergic and GABAergic synapses, tES can enhance or suppress synaptic strength depending on stimulation parameters and polarity (Nitsche et al., 2003; Fritsch et al., 2010). These effects are mediated by long-term potentiation (LTP) and depression (LTD), as conceptualized by Hebb (1949) and refined in models of spike-timing-dependent plasticity (STDP) (Citri and Malenka, 2008; Dan and Poo, 2006). Through these mechanisms, tES may help restore E/I balance in pathological conditions (Reato et al., 2013; Kronberg et al., 2017).
tDCS induces polarity-dependent neuromodulatory effects, first demonstrated by Bindman et al. (1964), who applied direct current to the cortex of anesthetized rats. These effects were described in humans decades later by Nitsche and Paulus (2000) using non-invasive stimulation. Their findings showed anodal tDCS enhances cortical excitability, promoting LTP-like plasticity, while cathodal tDCS reduces excitability, inducing LTD-like effects. Current evidence indicates tDCS effects depend not only on polarity but also on neuronal orientation relative to the induced electric field. Maximal effects occur when the somatodendritic axis aligns with the electric field (Bikson et al., 2004; Kronberg et al., 2017; Sánchez-León et al., 2025). tACS modulates synaptic plasticity by entraining neuronal activity at specific frequencies, reinforcing network oscillations. This effect has been observed in rodent and primate models (Fujisawa et al., 2004; Ozen et al., 2010; Krause et al., 2019, 2022). tACS-driven entrainment is presumed to promote STDP-like plasticity, contributing to E/I balance regulation, as suggested in both animal and human studies (Bland and Sale, 2019). tRNS applies randomized high-frequency currents and has been proposed to enhance synaptic plasticity via stochastic resonance, as demonstrated in vitro (Onorato et al., 2016). In addition, tRNS has been shown to reduce inhibitory responses and promotes LTP-like effects in humans (Van Der Groen and Wenderoth, 2016; Brancucci et al., 2023). Furthermore, chronic tRNS decreases GABA levels in mice, suggesting plasticity-related adaptations supporting long-term network reorganization (Sánchez-León et al., 2021b). Factors influencing tRNS-induced plasticity remain incompletely characterized, but human studies suggest intensity (Moliadze et al., 2012), frequency range (Fertonani et al., 2011; Campana et al., 2016; Moret et al., 2019), and brain state during stimulation (Jooß et al., 2016) significantly impact its effects.
By modulating synaptic plasticity, different tES protocols dynamically shift E/I balance, promoting excitation or enhancing inhibition. Since synaptic plasticity is governed by molecular and cellular events, we next explore how tES impacts these foundational mechanisms.
Molecular and cellular implications of tES on the E/I balance
The effects of tES on the E/I balance extend beyond neuronal excitability, influencing molecular pathways and cellular mechanisms that regulate synaptic plasticity. By modulating ion channels, neurotransmitter systems, and neurotrophic factors, tES induces adaptive molecular changes, which vary with stimulation modality and target cell type. Figure 1 summarizes the immediate and long-term effects of each tES modality on neuronal and glial components.
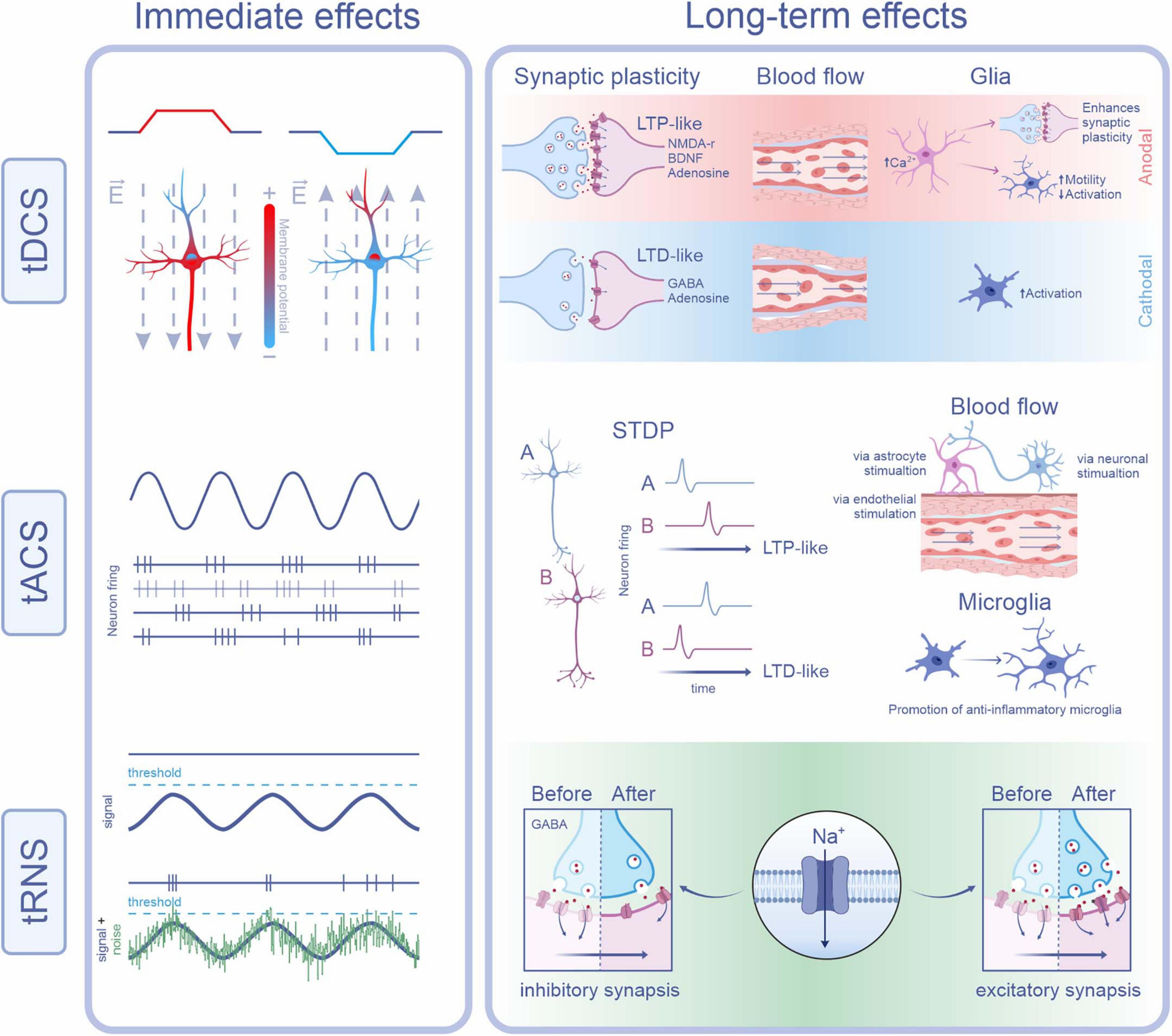
Figure 1. Physiological mechanisms underlying the immediate and long-term effects of tDCS, tACS, and tRNS. tDCS: Immediate effects result from changes in membrane potential due to redistribution of intracellular charges by the applied current. Anodal stimulation depolarizes the somatic membrane near the axon initial segment, increasing excitability by making this region more positive. In contrast, cathodal stimulation hyperpolarizes the same region, decreasing excitability. Long-term effects depend on current polarity: anodal tDCS (red background) promotes LTP-like plasticity via NMDA receptors, BDNF, and adenosine. It also enhances cerebral blood flow and astrocytic calcium levels, facilitating synaptic plasticity and microglial motility. Cathodal tDCS (blue background) favors LTD-like plasticity through GABA and adenosine receptors, reduces blood flow, and promotes a more reactive microglial state. tACS: Immediate effects are characterized by neuronal entrainment to the applied oscillatory current, depending on frequency and phase. This is illustrated by synchronous neuronal firing (purple) and a non-entrained neuron (gray-purple). Long-term effects involve synaptic plasticity through spike-timing dependent plasticity (STDP), where the relative timing of pre- and post-synaptic spikes determines whether LTP- or LTD-like changes occur. tACS also increases cerebral blood flow, potentially through activation of astrocytic, neuronal, or endothelial mechanisms, and shifts microglial phenotype toward an anti-inflammatory profile. tRNS: Immediate effects arise via stochastic resonance, whereby noise enhances the detection or transmission of weak signals. In the subpanel, the top trace shows an oscillatory membrane potential (purple) remaining below the threshold, with no action potentials generated. In contrast, the bottom trace demonstrates how the addition of random noise (green) can cause the combined signal to occasionally reach threshold, triggering action potentials. Long-term effects of tRNS may involve repeated sodium channel activation, leading to sustained depolarization and LTP-like plasticity at excitatory synapses. Additionally, LTD-like effects at inhibitory synapses may occur through modulation of GABAA receptors and decreased GABA release (Some images were created and obtained from the BioRender website: https://www.biorender.com/).
The electrical current used in tES alters membrane polarization by redistributing charges, which modulates ion channel activity and intracellular signaling (Ye and Steiger, 2015). In tDCS, anodal stimulation increases intracellular calcium—a key step in plasticity—through calcium channels, while sodium channels mediate immediate effects (Islam et al., 1995; Nitsche et al., 2003). NMDA receptor activation is also essential, as shown in vitro, in animal models, and in humans (Islam et al., 1995; Liebetanz, 2002; Nitsche et al., 2003; Rohan et al., 2015). Additionally, BDNF and adenosine contribute to long-term synaptic changes in animal studies (Fritsch et al., 2010; Márquez-Ruiz et al., 2012; Podda et al., 2016). Although the roles of BDNF and adenosine in tDCS have not been directly demonstrated in humans, the Val66Met BDNF polymorphism—which reduces BDNF release—has been associated with reduced plasticity after tDCS (Cheeran et al., 2008). The molecular mechanisms of tACS remain less defined, but studies indicate it modulates plasticity via neurotrophic and neurotransmitter systems. Beta-tACS (20 Hz) improves motor deficits in Parkinson’s models by increasing GDNF and activating neuroprotective pathways (Lee et al., 2022). Gamma-tACS (40 Hz) promotes synaptic potentiation by engaging AMPA receptors, BDNF, and CREB (Jeong et al., 2021). In humans, BDNF-dependent plasticity has been suggested to underlie some of the effects of tACS, although further investigation is needed (Riddle et al., 2020). Given that gamma oscillations rely on NMDA receptor activity, tACS may enhance synchronization through glutamatergic signaling (Fujisawa et al., 2004). Supporting this, the involvement of NMDA receptors in tACS-induced plasticity has also been demonstrated in humans (Wischnewski et al., 2019). Although less understood, tRNS appears to involve repeated sodium channel activation (Remedios et al., 2019) and reduced GABA release (Sánchez-León et al., 2021b). Unlike tDCS and tACS, it seems to act independently of NMDA receptors, relying on sodium channels and GABAergic modulation, as seen in human studies (Chaieb et al., 2015).
tES effects vary by neuronal cell type. tDCS primarily targets excitatory pyramidal neurons, likely due to their elongated morphology, as shown in vitro (Radman et al., 2009) and supported by computational models (Molaee-Ardekani et al., 2013). tACS entrains pyramidal neurons at both low (8 Hz) and high (> 100 Hz) frequencies, while interneurons show subtype-specific frequency preferences: somatostatin-positive cells respond to > 30 Hz, and parvalbumin-positive cells to ∼140 Hz, based on in vivo (Huang et al., 2021) and in vitro (Lee et al., 2024) studies. For tRNS, in vitro data suggest effects on pyramidal neurons (Remedios et al., 2019), while in vivo studies in awake animals point to possible involvement of GABAergic interneurons (Sánchez-León et al., 2021b). Beyond neurons, tES also affects glial cells, crucial for synaptic homeostasis and neuroinflammation. tDCS increases astrocytic calcium via adrenergic signaling and may influence microglia through astrocyte–microglia interactions (Monai et al., 2016; Mishima et al., 2019). It also modulates microglial activation and morphology, enhancing motility and neuron–microglia signaling via the fractalkine pathway (Rueger et al., 2012; Pikhovych et al., 2016; Gellner et al., 2021). tDCS alters cerebral blood flow, increasing it after anodal and decreasing it after cathodal stimulation in animals (Wachter et al., 2011) and in humans (Shinde et al., 2021). Similarly, tACS enhances cerebral perfusion in a frequency- and dose-dependent manner through various mechanisms, including neurovascular coupling, endothelial activation, astrocytic stimulation, and direct neuronal effects (Turner et al., 2021). These effects have also been observed in humans (Alekseichuk et al., 2016). Gamma-tACS reduces beta-amyloid plaques and promotes an anti-inflammatory microglial phenotype, suggesting therapeutic potential in neurodegeneration (Wu et al., 2022). While direct microglial changes have not been confirmed in humans, a reduction of p-tau seen after 40 Hz tACS in Alzheimer’s patients suggests microglial enhancement (Dhaynaut et al., 2022).
Modulation of the E/I balance by tES involves coordinated changes at the molecular and cellular levels. The next section explores how these local effects extend to large-scale network interactions that ultimately drive brain function.
Modulation of the E/I balance at the neural network level
Beyond its local effects, tES influences large-scale network connectivity and oscillatory activity, contributing to the modulation of the E/I balance at the neural network level. These effects extend beyond the immediate stimulation site, altering both local and distant brain regions and impacting functional and effective connectivity (Ozen et al., 2010; Reato et al., 2010; Koo et al., 2016; Cambiaghi et al., 2020; Krause et al., 2022).
Among tES modalities, tDCS has been widely studied for its ability to modulate network connectivity and oscillatory activity, thereby influencing global E/I balance. These effects were first observed in humans (Antal et al., 2004) and later replicated in animals (Reato et al., 2010). For instance, tDCS applied to the primary somatosensory cortex in mice modulates gamma activity during and after stimulation, indicating plasticity changes that could influence broader network dynamics (Sánchez-León et al., 2021a). Similarly, in Alzheimer’s disease models, prefrontal tDCS alters both alpha and gamma oscillations, suggesting its potential to restore pathological E/I imbalances and impact interconnected brain regions (Duan et al., 2022).
The network-wide effects of tDCS are linked to its layer-specific influence on cortical circuits and its capacity to modulate interhemispheric and cortico-subcortical pathways. tDCS effects are layer-dependent, with cathodal stimulation inducing LTD-like effects in superficial layers and LTP-like effects in deeper ones, as shown in animal models (Sun et al., 2020). Furthermore, tDCS influences distant regions through interhemispheric and cortico-subcortical connectivity. For example, anodal tDCS applied to the left motor cortex enhances contralateral excitability in rats, indicating plasticity-driven changes in network interactions (Koo et al., 2016). Anodal prefrontal tDCS also modulates serotonergic activity in the dorsal raphe nucleus, affecting neuromodulatory systems in remote regions (Cambiaghi et al., 2020). This ability to influence distal regions has also been confirmed in humans, where anodal prefrontal tDCS modulated activity in subcortical and contralateral cortical areas (Weber et al., 2014). Like tDCS, tACS exerts widespread effects beyond the stimulation site. In anesthetized rats, tACS entrains neuronal firing across extensive cortical networks, demonstrating its capacity to synchronize oscillatory activity at large scale (Ozen et al., 2010). Intrinsic network dynamics can amplify tACS effects, enhancing its impact on E/I balance and long-range connectivity, as supported by animal (Huang et al., 2021; Krause et al., 2022) and humans studies (Chen et al., 2021; Diedrich et al., 2025). Although tRNS is relatively new, human studies suggest it modulates neural oscillations. For example, tRNS over the auditory cortex increased theta power in frontal and parietal regions, suggesting potential network-wide modulation (Van Doren et al., 2014). However, the mechanisms remain unclear, and further animal studies are needed to clarify its impact on network-level E/I balance.
By modulating large-scale connectivity and oscillatory dynamics, tES reshapes the brain’s E/I balance, offering therapeutic potential for disorders characterized by E/I dysregulation (Krause et al., 2013). Despite significant progress in understanding the network-level effects of tES, important challenges remain that limit the extrapolation of preclinical findings to human applications.
Limitations and future perspectives
Studying the E/I balance using animal models presents inherent limitations due to anatomical and physiological differences between animal and human brains. A major challenge is the simpler geometry of animal cortices compared to the convoluted human cortex, which affects both electrical field distribution and large-scale network dynamics. For example, rodents’ smaller brains and lack of cortical gyri limit direct extrapolation to humans. Moreover, scaling issues in tES protocols often require higher current densities in animals, influenced by differences in neuronal density, axonal architecture, and cortical organization (Ozen et al., 2010; Vöröslakos et al., 2018; Asan et al., 2020). Stimulation durations also vary substantially, as preclinical studies typically employ shorter sessions than clinical protocols, complicating the assessment of long-term effects (Johnson et al., 2020; Huang et al., 2021).
Efforts are underway to bridge these gaps. Studies in non-human primates provide more translatable data on behavior, electric field distribution, and neural dynamics (Opitz et al., 2016; Krause et al., 2017). In rodents, lowering current densities (Bolzoni et al., 2013; Farahani et al., 2024) and incorporating human-relevant behavioral tasks (Márquez-Ruiz et al., 2012, 2016; Kar et al., 2017) have improved translational validity. Furthermore, animal models remain indispensable for mechanistic research at the synaptic and network levels—insights not easily attainable in humans (Jackson et al., 2016; Sánchez-León et al., 2018). Importantly, these experimental findings provide the biological foundation for developing computational models that simulate how tES modulates neuronal activity. By incorporating data on cellular and network-level mechanisms from animal studies, such models can help extrapolate stimulation effects to the human brain and guide protocol optimization with improved anatomical and physiological accuracy.
Future research should prioritize methodological refinement. Using computational modeling to estimate electric fields more accurately, adjusting stimulation parameters accordingly, and aligning experimental designs with disease-specific biomarkers will be key to increasing the translational value of animal studies. Integrating advanced neurotechnologies into these models can further accelerate progress toward clinically meaningful applications of tES.
Conclusion
In summary, tES is a promising neuromodulatory approach with significant potential for both basic neuroscience and clinical translation. By modulating the E/I balance, it influences synaptic plasticity, circuit function, and network synchronization, offering a powerful tool to probe and treat neurological and psychiatric conditions. Insights from animal models have elucidated key mechanisms underlying tES effects, including polarity-dependent modulation by tDCS, frequency-specific entrainment by tACS, and the emerging utility of tRNS.
These findings underscore the value of preclinical research for identifying how tES interacts with glutamatergic and GABAergic systems, supports oscillatory coherence, and shapes brain dynamics. Moving forward, combining tES with complementary approaches—such as pharmacological, genetic, or behavioral interventions—may enhance specificity and therapeutic efficacy. Continued integration of mechanistic insights will be essential to realize personalized neuromodulation strategies and improve clinical outcomes in disorders characterized by disrupted E/I balance.
Author contributions
ME-R: Conceptualization, Writing – original draft, Writing – review and editing. GS-GC: Writing – review and editing. ÁZ: Writing – review and editing. IC: Writing – review and editing. JM-R: Conceptualization, Writing – original draft, Writing – review and editing.
Funding
The authors declare that financial support was received for the research and/or publication of this article. This work was supported by grants from the Spanish MINECO-FEDER (BFU2017-89615-P and PID2022-141997NB-I00) and FET European Union’s Horizon 2020 research and innovation program (grant agreement No 101017716) to JM-R. GS-GC was in receipt of an FPU grant from the Spanish Government (FPU21/01025).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The authors declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The authors declare that Generative AI was used in the creation of this manuscript. ChatGPT 4o was used to enhance language clarity and polish the writing. All content generated or improved using Generative AI has been extensively reviewed, edited, and verified by the authors to ensure factual accuracy and alignment with the scientific standards of the journal.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Alekseichuk, I., Diers, K., Paulus, W., and Antal, A. (2016). Transcranial electrical stimulation of the occipital cortex during visual perception modifies the magnitude of BOLD activity: A combined tES–fMRI approach. NeuroImage 140, 110–117. doi: 10.1016/j.neuroimage.2015.11.034
Antal, A., and Herrmann, C. S. (2016). Transcranial alternating current and random noise stimulation: Possible mechanisms. Neural Plastic. 2016, 1–12. doi: 10.1155/2016/3616807
Antal, A., and Paulus, W. (2013). Transcranial alternating current stimulation (tACS). Front. Hum. Neurosci. 7:317. doi: 10.3389/fnhum.2013.00317
Antal, A., Varga, E. T., Kincses, T. Z., Nitsche, M. A., and Paulus, W. (2004). Oscillatory brain activity and transcranial direct current stimulation in humans. NeuroReport 15, 1307–1310. doi: 10.1097/01.wnr.0000127460.08361.84
Asan, A. S., Lang, E. J., and Sahin, M. (2020). Entrainment of cerebellar purkinje cells with directional AC electric fields in anesthetized rats. Brain Stimul. 13, 1548–1558. doi: 10.1016/j.brs.2020.08.017
Benussi, A., Cantoni, V., Cotelli, M. S., Cotelli, M., Brattini, C., Datta, A., et al. (2021). Exposure to gamma tACS in Alzheimer’s disease: A randomized, double-blind, sham-controlled, crossover, pilot study. Brain Stimul. 14, 531–540. doi: 10.1016/j.brs.2021.03.007
Bikson, M., Inoue, M., Akiyama, H., Deans, J. K., Fox, J. E., Miyakawa, H., et al. (2004). Effects of uniform extracellular DC electric fields on excitability in rat hippocampal slices in vitro. J. Physiol. 557, 175–190. doi: 10.1113/jphysiol.2003.055772
Bikson, M., Reato, D., and Rahman, A. (2012). “Cellular and network effects of transcranial direct current stimulation: Insights from animal models and brain slice,” in Transcranial Brain Stimulation, Vol. 20125361, ed. P. Rossini (Boca Raton, FL: CRC Press), 55–91. doi: 10.1201/b14174-5
Bindman, L. J., Lippold, O. C. J., and Redfearn, J. W. T. (1964). The action of brief polarizing currents on the cerebral cortex of the rat (1) during current flow and (2) in the production of long-lasting after-effects. J. Physiol. 172, 369–382. doi: 10.1113/jphysiol.1964.sp007425
Bland, N. S., and Sale, M. V. (2019). Current challenges: The ups and downs of tACS. Exp. Brain Res. 237, 3071–3088. doi: 10.1007/s00221-019-05666-0
Bolzoni, F., Pettersson, L., and Jankowska, E. (2013). Evidence for long-lasting subcortical facilitation by transcranial direct current stimulation in the cat. J. Physiol. 591, 3381–3399. doi: 10.1113/jphysiol.2012.244764
Brancucci, A., Rivolta, D., Nitsche, M. A., and Manippa, V. (2023). The effects of transcranial random noise stimulation on motor function: A comprehensive review of the literature. Physiol. Behav. 261:114073. doi: 10.1016/j.physbeh.2023.114073
Brunoni, A. R., Ferrucci, R., Fregni, F., Boggio, P. S., and Priori, A. (2012). Transcranial direct current stimulation for the treatment of major depressive disorder: A summary of preclinical, clinical and translational findings. Prog. Neuro-Psychopharmacol. Biol. Psychiatry 39, 9–16. doi: 10.1016/j.pnpbp.2012.05.016
Cambiaghi, M., Buffelli, M., Masin, L., Valtorta, F., and Comai, S. (2020). Transcranial direct current stimulation of the mouse prefrontal cortex modulates serotonergic neural activity of the dorsal raphe nucleus. Brain Stimul. 13, 548–550. doi: 10.1016/j.brs.2020.01.012
Campana, G., Camilleri, R., Moret, B., Ghin, F., and Pavan, A. (2016). Opposite effects of high- and low-frequency transcranial random noise stimulation probed with visual motion adaptation. Sci. Rep. 6:38919. doi: 10.1038/srep38919
Chaieb, L., Antal, A., and Paulus, W. (2015). Transcranial random noise stimulation-induced plasticity is NMDA-receptor independent but sodium-channel blocker and benzodiazepines sensitive. Front. Neurosci. 9:125. doi: 10.3389/fnins.2015.00125
Cheeran, B., Talelli, P., Mori, F., Koch, G., Suppa, A., Edwards, M., et al. (2008). A common polymorphism in the brain-derived neurotrophic factor gene (BDNF) modulates human cortical plasticity and the response to rTMS. J. Physiol. 586, 5717–5725. doi: 10.1113/jphysiol.2008.159905
Chen, C., Yuan, K., Chu, W. C., and Tong, R. K. (2021). The effects of 10 Hz and 20 Hz tACS in network integration and segregation in chronic stroke: A graph theoretical fMRI study. Brain Sci. 11:377. doi: 10.3390/brainsci11030377
Citri, A., and Malenka, R. C. (2008). Synaptic plasticity: Multiple forms, functions, and mechanisms. Neuropsychopharmacology 33, 18–41. doi: 10.1038/sj.npp.1301559
D’amour, J. A., and Froemke, R. C. (2015). Inhibitory and excitatory spike-timing-dependent plasticity in the auditory cortex. Neuron 86, 514–528. doi: 10.1016/j.neuron.2015.03.014
Dan, Y., and Poo, M.-M. (2006). Spike timing-dependent plasticity: From synapse to perception. Physiol. Rev. 86, 1033–1048. doi: 10.1152/physrev.00030.2005
Dhaynaut, M., Sprugnoli, G., Cappon, D., Macone, J., Sanchez, J. S., Normandin, M. D., et al. (2022). Impact of 40 Hz transcranial alternating current stimulation on cerebral tau burden in patients with Alzheimer’s Disease: A case series. J Alzheimer’s Dis. 85, 1667–1676. doi: 10.3233/JAD-215072
Diedrich, L., Kolhoff, H. I., Bergmann, C., Chakraborty, S., and Antal, A. (2025). Theta–gamma tACS modulates attention network synchronization, not isolated network performance. Brain Res. 1855:149550. doi: 10.1016/j.brainres.2025.149550
Duan, M., Meng, Z., Yuan, D., Zhang, Y., Tang, T., Chen, Z., et al. (2022). Anodal and cathodal transcranial direct current stimulations of prefrontal cortex in a rodent model of Alzheimer’s disease. Front. Aging Neurosci. 14:968451. doi: 10.3389/fnagi.2022.968451
Eichler, S. A., and Meier, J. C. (2008). E-I balance and human diseases – from molecules to networking. Front. Mol. Neurosci. 1:8. doi: 10.3389/neuro.02.002.2008
Etter, G., Van Der Veldt, S., Manseau, F., Zarrinkoub, I., Trillaud-Doppia, E., and Williams, S. (2019). Optogenetic gamma stimulation rescues memory impairments in an Alzheimer’s disease mouse model. Nat. Commun. 10:5322. doi: 10.1038/s41467-019-13260-9
Farahani, F., Khadka, N., Parra, L. C., Bikson, M., and Vöröslakos, M. (2024). Transcranial electric stimulation modulates firing rate at clinically relevant intensities. Brain Stimul. 17, 561–571. doi: 10.1016/j.brs.2024.04.007
Fertonani, A., Pirulli, C., and Miniussi, C. (2011). Random noise stimulation improves neuroplasticity in perceptual learning. J. Neurosci. 31, 15416–15423. doi: 10.1523/JNEUROSCI.2002-11.2011
Fritsch, B., Reis, J., Martinowich, K., Schambra, H. M., Ji, Y., Cohen, L. G., et al. (2010). Direct current stimulation promotes BDNF-dependent synaptic plasticity: Potential implications for motor learning. Neuron 66, 198–204. doi: 10.1016/j.neuron.2010.03.035
Froemke, R. C. (2015). Plasticity of cortical excitatory-inhibitory balance. Annu. Rev. Neurosci. 38, 195–219. doi: 10.1146/annurev-neuro-071714-034002
Fröhlich, F., and McCormick, D. A. (2010). Endogenous electric fields may guide neocortical network activity. Neuron 67, 129–143. doi: 10.1016/j.neuron.2010.06.005
Fujisawa, S., Matsuki, N., and Ikegaya, Y. (2004). Chronometric readout from a memory trace: Gamma-frequency field stimulation recruits timed recurrent activity in the rat CA3 network. J. Physiol. 561, 123–131. doi: 10.1113/jphysiol.2004.066639
Gellner, A.-K., Reis, J., Fiebich, B. L., and Fritsch, B. (2021). Electrified microglia: Impact of direct current stimulation on diverse properties of the most versatile brain cell. Brain Stimul. 14, 1248–1258. doi: 10.1016/j.brs.2021.08.007
Hebb, D. O. (1949). The Organization of Behavior. A Neuropsycological Theory. Hoboken, NJ: John Wiley & Sons, Inc.
Herrmann, C. S., Rach, S., Neuling, T., and Strüber, D. (2013). Transcranial alternating current stimulation: A review of the underlying mechanisms and modulation of cognitive processes. Front. Hum. Neurosci. 7:279. doi: 10.3389/fnhum.2013.00279
Huang, W. A., Stitt, I. M., Negahbani, E., Passey, D. J., Ahn, S., Davey, M., et al. (2021). Transcranial alternating current stimulation entrains alpha oscillations by preferential phase synchronization of fast-spiking cortical neurons to stimulation waveform. Nat. Commun. 12:3151. doi: 10.1038/s41467-021-23021-2
Iaccarino, H. F., Singer, A. C., Martorell, A. J., Rudenko, A., Gao, F., Gillingham, T. Z., et al. (2016). Gamma frequency entrainment attenuates amyloid load and modifies microglia. Nature 540, 230–235. doi: 10.1038/nature20587
Isaacson, J. S., and Scanziani, M. (2011). How inhibition shapes cortical activity. Neuron 72, 231–243. doi: 10.1016/j.neuron.2011.09.027
Islam, N., Aftabuddin, M., Moriwaki, A., Hattori, Y., and Hori, Y. (1995). Increase in the calcium level following anodal polarization in the rat brain. Brain Res. 684, 206–208. doi: 10.1016/0006-8993(95)00434-R
Jackson, M. P., Rahman, A., Lafon, B., Kronberg, G., Ling, D., Parra, L. C., et al. (2016). Animal models of transcranial direct current stimulation: Methods and mechanisms. Clin. Neurophysiol. 127, 3425–3454. doi: 10.1016/j.clinph.2016.08.016
Jeong, W.-H., Kim, W.-I., Lee, J.-W., Park, H.-K., Song, M.-K., Choi, I.-S., et al. (2021). Modulation of long-term potentiation by gamma frequency transcranial alternating current stimulation in transgenic mouse models of Alzheimer’s disease. Brain Sci. 11:1532. doi: 10.3390/brainsci11111532
Johnson, L., Alekseichuk, I., Krieg, J., Doyle, A., Yu, Y., Vitek, J., et al. (2020). Dose-dependent effects of transcranial alternating current stimulation on spike timing in awake nonhuman primates. Sci. Adv. 6:eaaz2747. doi: 10.1126/sciadv.aaz2747
Jooß, A., Haberbosch, L., Rönnefarth, M., Fleischmann, R., Scholz, M., Brandt, S., et al. (2016). EP 69. Brain state dependent inhibitory and facilitatory effects following transcranial random noise stimulation in two motor tasks. Clin. Neurophysiol. 127, e267–e268. doi: 10.1016/j.clinph.2016.05.120
Kabakov, A. Y., Muller, P. A., Pascual-Leone, A., Jensen, F. E., and Rotenberg, A. (2012). Contribution of axonal orientation to pathway-dependent modulation of excitatory transmission by direct current stimulation in isolated rat hippocampus. J. Neurophysiol. 107, 1881–1889. doi: 10.1152/jn.00715.2011
Kar, K., Duijnhouwer, J., and Krekelberg, B. (2017). Transcranial alternating current stimulation attenuates neuronal adaptation. J. Neurosci. 37, 2325–2335. doi: 10.1523/JNEUROSCI.2266-16.2016
Kasten, F. H., Dowsett, J., and Herrmann, C. S. (2016). Sustained aftereffect of α-tACS lasts up to 70 min after stimulation. Front. Hum. Neurosci. 10:245. doi: 10.3389/fnhum.2016.00245
Koo, H., Kim, M. S., Han, S. W., Paulus, W., Nitche, M. A., Kim, Y.-H., et al. (2016). After-effects of anodal transcranial direct current stimulation on the excitability of the motor cortex in rats. Restorative Neurol. Neurosci. 34, 859–868. doi: 10.3233/rnn-160664
Krause, B., Márquez-Ruiz, J., and Kadosh, R. C. (2013). The effect of transcranial direct current stimulation: A role for cortical excitation/inhibition balance? Front. Hum. Neurosci. 7:602. doi: 10.3389/fnhum.2013.00602
Krause, M. R., Vieira, P. G., Csorba, B. A., Pilly, P. K., and Pack, C. C. (2019). Transcranial alternating current stimulation entrains single-neuron activity in the primate brain. Proc. Natl. Acad. Sci. U. S. A. 116, 5747–5755. doi: 10.1073/pnas.1815958116
Krause, M. R., Vieira, P. G., Thivierge, J.-P., and Pack, C. C. (2022). Brain stimulation competes with ongoing oscillations for control of spike timing in the primate brain. PLoS Biol. 20:e3001650. doi: 10.1371/journal.pbio.3001650
Krause, M. R., Zanos, T. P., Csorba, B. A., Pilly, P. K., Choe, J., Phillips, M. E., et al. (2017). Transcranial direct current stimulation facilitates associative learning and alters functional connectivity in the primate brain. Curr. Biol. 27, 3086–3096.e3. doi: 10.1016/j.cub.2017.09.020
Kronberg, G., Bridi, M., Abel, T., Bikson, M., and Parra, L. C. (2017). Direct current stimulation modulates LTP and LTD: Activity dependence and dendritic effects. Brain Stimul. 10, 51–58. doi: 10.1016/j.brs.2016.10.001
Landau, I. D., Egger, R., Dercksen, V. J., Oberlaender, M., and Sompolinsky, H. (2016). The impact of structural heterogeneity on excitation-inhibition balance in cortical networks. Neuron 92, 1106–1121. doi: 10.1016/j.neuron.2016.10.027
Lee, H. J., Jung, D. H., Jung, Y. J., Shin, H. K., and Choi, B. T. (2022). Transcranial alternating current stimulation rescues motor deficits in a mouse model of Parkinson’s disease via the production of glial cell line-derived neurotrophic factor. Brain Stimul. 15, 645–653. doi: 10.1016/j.brs.2022.04.002
Lee, S. Y., Kozalakis, K., Baftizadeh, F., Campagnola, L., Jarsky, T., Koch, C., et al. (2024). Cell-class-specific electric field entrainment of neural activity. Neuron 112, 2614–2630.e5. doi: 10.1016/j.neuron.2024.05.009
Liebetanz, D. (2002). Pharmacological approach to the mechanisms of transcranial DC-stimulation-induced after-effects of human motor cortex excitability. Brain 125, 2238–2247. doi: 10.1093/brain/awf238
Mabil, P., Huidobro, N., Torres-Ramirez, O., Flores-Hernandez, J., Flores, A., Gutierrez, R., et al. (2020). Noisy light augments the Na+ current in somatosensory pyramidal neurons of optogenetic transgenic mice. Front. Neurosci. 14:490. doi: 10.3389/fnins.2020.00490
Márquez-Ruiz, J., Ammann, C., Leal-Campanario, R., Ruffini, G., Gruart, A., and Delgado-García, J. M. (2016). Synthetic tactile perception induced by transcranial alternating-current stimulation can substitute for natural sensory stimulus in behaving rabbits. Sci. Rep. 6:19753. doi: 10.1038/srep19753
Márquez-Ruiz, J., Leal-Campanario, R., Sánchez-Campusano, R., Molaee-Ardekani, B., Wendling, F., Miranda, P. C., et al. (2012). Transcranial direct-current stimulation modulates synaptic mechanisms involved in associative learning in behaving rabbits. Proc. Natl. Acad. Sci. U. S. A. 109, 6710–6715. doi: 10.1073/pnas.1121147109
Márquez-Ruiz, J., Leal-Campanario, R., Wendling, F., Ruffini, G., Gruart, A., and Delgado-García, J. M. (2014). “Transcranial electrical stimulation in animals,” in En The Stimulated Brain, ed. R. C. Kadosh (Amsterdam: Elsevier), 117–144. doi: 10.1016/B978-0-12-404704-4.00005-3
Mishima, T., Nagai, T., Yahagi, K., Akther, S., Oe, Y., Monai, H., et al. (2019). Transcranial direct current stimulation (tDCS) induces adrenergic receptor-dependent microglial morphological changes in mice. eNeuro 6:ENEURO.0204-19.2019. doi: 10.1523/ENEURO.0204-19.2019
Molaee-Ardekani, B., Márquez-Ruiz, J., Merlet, I., Leal-Campanario, R., Gruart, A., Sánchez-Campusano, R., et al. (2013). Effects of transcranial Direct Current Stimulation (tDCS) on cortical activity: A computational modeling study. Brain Stimul. 6, 25–39. doi: 10.1016/j.brs.2011.12.006
Moliadze, V., Atalay, D., Antal, A., and Paulus, W. (2012). Close to threshold transcranial electrical stimulation preferentially activates inhibitory networks before switching to excitation with higher intensities. Brain Stimul. 5, 505–511. doi: 10.1016/j.brs.2011.11.004
Monai, H., Ohkura, M., Tanaka, M., Oe, Y., Konno, A., Hirai, H., et al. (2016). Calcium imaging reveals glial involvement in transcranial direct current stimulation-induced plasticity in mouse brain. Nat. Commun. 7:11100. doi: 10.1038/ncomms11100
Moret, B., Donato, R., Nucci, M., Cona, G., and Campana, G. (2019). Transcranial random noise stimulation (tRNS): A wide range of frequencies is needed for increasing cortical excitability. Sci. Rep. 9:15150. doi: 10.1038/s41598-019-51553-7
Nguyen, B. N., McKendrick, A. M., and Vingrys, A. J. (2016). Abnormal inhibition-excitation imbalance in migraine. Cephalalgia 36, 5–14. doi: 10.1177/0333102415576725
Nitsche, M. A., and Paulus, W. (2000). Excitability changes induced in the human motor cortex by weak transcranial direct current stimulation. J. Physiol. 527, 633–639. doi: 10.1111/j.1469-7793.2000.t01-1-00633.x
Nitsche, M. A., Cohen, L. G., Wassermann, E. M., Priori, A., Lang, N., Antal, A., et al. (2008). Transcranial direct current stimulation: State of the art 2008. Brain Stimul. 1, 206–223. doi: 10.1016/j.brs.2008.06.004
Nitsche, M. A., Fricke, K., Henschke, U., Schlitterlau, A., Liebetanz, D., Lang, N., et al. (2003). Pharmacological modulation of cortical excitability shifts induced by transcranial direct current stimulation in humans. J. Physiol. 553, 293–301. doi: 10.1113/jphysiol.2003.049916
Onorato, I., D’Alessandro, G., Di Castro, M. A., Renzi, M., Dobrowolny, G., Musarò, A., et al. (2016). Noise enhances action potential generation in mouse sensory neurons via stochastic resonance. PLoS One 11:e0160950. doi: 10.1371/journal.pone.0160950
Opitz, A., Falchier, A., Yan, C.-G., Yeagle, E. M., Linn, G. S., Megevand, P., et al. (2016). Spatiotemporal structure of intracranial electric fields induced by transcranial electric stimulation in humans and nonhuman primates. Sci. Rep. 6:31236. doi: 10.1038/srep31236
Ozen, S., Sirota, A., Belluscio, M. A., Anastassiou, C. A., Stark, E., Koch, C., et al. (2010). Transcranial electric stimulation entrains cortical neuronal populations in rats. J. Neurosci. 30, 11476–11485. doi: 10.1523/jneurosci.5252-09.2010
Paulus, W. (2011). Transcranial electrical stimulation (tES - tDCS; tRNS, tACS) methods. Neuropsychol. Rehabil. 21, 602–617. doi: 10.1080/09602011.2011.557292
Pikhovych, A., Stolberg, N. P., Jessica Flitsch, L., Walter, H. L., Graf, R., Fink, G. R., et al. (2016). Transcranial direct current stimulation modulates neurogenesis and microglia activation in the mouse brain. Stem Cells Int. 2016, 2715196. doi: 10.1155/2016/2715196
Podda, M. V., Cocco, S., Mastrodonato, A., Fusco, S., Leone, L., Barbati, S. A., et al. (2016). Anodal transcranial direct current stimulation boosts synaptic plasticity and memory in mice via epigenetic regulation of Bdnf expression. Sci. Rep. 6:22180. doi: 10.1038/srep22180
Radman, T., Ramos, R. L., Brumberg, J. C., and Bikson, M. (2009). Role of cortical cell type and morphology in subthreshold and suprathreshold uniform electric field stimulation in vitro. Brain Stimul. 2:215–228.e3. doi: 10.1016/j.brs.2009.03.007
Reato, D., Rahman, A., Bikson, M., and Parra, L. C. (2010). Low-intensity electrical stimulation affects network dynamics by modulating population rate and spike timing. J. Neurosci. 30, 15067–15079. doi: 10.1523/JNEUROSCI.2059-10.2010
Reato, D., Rahman, A., Bikson, M., and Parra, L. C. (2013). Effects of weak transcranial alternating current stimulation on brain activity—A review of known mechanisms from animal studies. Front. Hum. Neurosci. 7:687. doi: 10.3389/fnhum.2013.00687
Remedios, L., Mabil, P., Flores-Hernández, J., Torres-Ramírez, O., Huidobro, N., Castro, G., et al. (2019). Effects of short-term random noise electrical stimulation on dissociated pyramidal neurons from the cerebral cortex. Neuroscience 404, 371–386. doi: 10.1016/j.neuroscience.2019.01.035
Riddle, J., McPherson, T., Atkins, A. K., Walker, C. P., Ahn, S., and Frohlich, F. (2020). Brain-derived neurotrophic factor (BDNF) polymorphism may influence the efficacy of tACS to modulate neural oscillations. Brain Stimul. 13, 998–999. doi: 10.1016/j.brs.2020.04.012
Rohan, J. G., Carhuatanta, K. A., McInturf, S. M., Miklasevich, M. K., and Jankord, R. (2015). Modulating hippocampal plasticity with in vivo brain stimulation. J. Neurosci. 35, 12824–12832. doi: 10.1523/JNEUROSCI.2376-15.2015
Rueger, M. A., Keuters, M. H., Walberer, M., Braun, R., Klein, R., Sparing, R., et al. (2012). Multi-session transcranial direct current stimulation (tDCS) elicits inflammatory and regenerative processes in the rat brain. PLoS One 7:e43776. doi: 10.1371/journal.pone.0043776
Sánchez-Garrido Campos, G., Zafra, Á. M., Estévez-Rodríguez, M., Cordones, I., Ruffini, G., and Márquez-Ruiz, J. (2025). Preclinical insights into gamma-tACS: Foundations for clinical translation in neurodegenerative diseases. Front. Neurosci. 19:1549230. doi: 10.3389/fnins.2025.1549230
Sánchez-León, C. A., Ammann, C., Medina, J. F., and Márquez-Ruiz, J. (2018). Using animal models to improve the design and application of transcranial electrical stimulation in humans. Curr. Behav. Neurosci. Rep. 5, 125–135. doi: 10.1007/s40473-018-0149-6
Sánchez-León, C. A., Campos, G. S.-G., Fernández, M., Sánchez-López, A., Medina, J. F., and Márquez-Ruiz, J. (2025). Somatodendritic orientation determines tDCS-induced neuromodulation of Purkinje cell activity in awake mice. bioRxiv [Preprint]. doi: 10.1101/2023.02.18.529047
Sánchez-León, C. A., Cordones, I., Ammann, C., Ausín, J. M., Gómez-Climent, M. A., Carretero-Guillén, A., et al. (2021a). Immediate and after effects of transcranial direct-current stimulation in the mouse primary somatosensory cortex. Sci. Rep. 11:3123. doi: 10.1038/s41598-021-82364-4
Sánchez-León, C. A., Sánchez-López, Á, Gómez-Climent, M. A., Cordones, I., Cohen Kadosh, R., and Márquez-Ruiz, J. (2021b). Impact of chronic transcranial random noise stimulation (tRNS) on GABAergic and glutamatergic activity markers in the prefrontal cortex of juvenile mice. Prog. Brain Res. 264, 323–341. doi: 10.1016/bs.pbr.2021.01.017
Saraga, F., Balena, T., Wolansky, T., Dickson, C. T., and Woodin, M. A. (2008). Inhibitory synaptic plasticity regulates pyramidal neuron spiking in the rodent hippocampus. Neuroscience 155, 64–75. doi: 10.1016/j.neuroscience.2008.05.009
Schabrun, S. M., Lamont, R. M., and Brauer, S. G. (2016). Transcranial direct current stimulation to enhance dual-task gait training in Parkinson’s Disease: A pilot RCT. PLoS One 11:e0158497. doi: 10.1371/journal.pone.0158497
Shinde, A. B., Lerud, K. D., Munsch, F., Alsop, D. C., and Schlaug, G. (2021). Effects of tDCS dose and electrode montage on regional cerebral blood flow and motor behavior. NeuroImage 237:118144. doi: 10.1016/j.neuroimage.2021.118144
Snowball, A., Tachtsidis, I., Popescu, T., Thompson, J., Delazer, M., Zamarian, L., et al. (2013). Long-term enhancement of brain function and cognition using cognitive training and brain stimulation. Curr. Biol. 23, 987–992. doi: 10.1016/j.cub.2013.04.045
Su, Y.-J., Yi, P.-L., and Chang, F.-C. (2024). Transcranial direct current stimulation (tDCS) ameliorates stress-induced sleep disruption via activating infralimbic-ventrolateral preoptic projections. Brain Sci. 14:105. doi: 10.3390/brainsci14010105
Sun, Y., Dhamne, S. C., Carretero-Guillén, A., Salvador, R., Goldenberg, M. C., Godlewski, B. R., et al. (2020). Drug-responsive inhomogeneous cortical modulation by direct current stimulation. Ann. Neurol. 88, 489–502. doi: 10.1002/ana.25822
Tamura, R., Dezawa, S., Kato, J., Nakata, M., Kunori, N., and Takashima, I. (2024). Transcranial direct current stimulation improves motor function in rats with 6-hydroxydopamine-induced Parkinsonism. Behav. Brain Res. 460:114815. doi: 10.1016/j.bbr.2023.114815
Tatti, R., Haley, M. S., Swanson, O. K., Tselha, T., and Maffei, A. (2017). Neurophysiology and regulation of the balance between excitation and inhibition in neocortical circuits. Biol. Psychiatry 81, 821–831. doi: 10.1016/j.biopsych.2016.09.017
Tekturk, P., Erdogan, E. T., Kurt, A., Vanli-yavuz, E. N., Ekizoglu, E., Kocagoncu, E., et al. (2016). The effect of transcranial direct current stimulation on seizure frequency of patients with mesial temporal lobe epilepsy with hippocampal sclerosis. Clin. Neurol. Neurosurg. 149, 27–32. doi: 10.1016/j.clineuro.2016.07.014
Turner, D., Degan, S., Galeffi, F., Schmidt, S., and Peterchev, A. V. (2021). Rapid, dose-dependent enhancement of cerebral blood flow by transcranial AC stimulation in mouse. Brain Stimul. 14, 80–87. doi: 10.1016/j.brs.2020.11.012
Van Bueren, N. E. R., Van Der Ven, S. H. G., Hochman, S., Sella, F., and Cohen Kadosh, R. (2023). Human neuronal excitation/inhibition balance explains and predicts neurostimulation induced learning benefits. PLoS Biol. 21:e3002193. doi: 10.1371/journal.pbio.3002193
Van Der Groen, O., and Wenderoth, N. (2016). Transcranial random noise stimulation of visual cortex: Stochastic resonance enhances central mechanisms of perception. J. Neurosci. 36, 5289–5298. doi: 10.1523/JNEUROSCI.4519-15.2016
Van Der Groen, O., Potok, W., Wenderoth, N., Edwards, G., Mattingley, J. B., and Edwards, D. (2022). Using noise for the better: The effects of transcranial random noise stimulation on the brain and behavior. Neurosci. Biobehav. Rev. 138:104702. doi: 10.1016/j.neubiorev.2022.104702
Van Doren, J., Langguth, B., and Schecklmann, M. (2014). Electroencephalographic effects of transcranial random noise stimulation in the auditory cortex. Brain Stimul. 7, 807–812. doi: 10.1016/j.brs.2014.08.007
Vöröslakos, M., Takeuchi, Y., Brinyiczki, K., Zombori, T., Oliva, A., Fernández-Ruiz, A., et al. (2018). Direct effects of transcranial electric stimulation on brain circuits in rats and humans. Nat. Commun. 9:483. doi: 10.1038/s41467-018-02928-3
Wachter, D., Wrede, A., Schulz-Schaeffer, W., Taghizadeh-Waghefi, A., Nitsche, M. A., Kutschenko, A., et al. (2011). Transcranial direct current stimulation induces polarity-specific changes of cortical blood perfusion in the rat. Exp. Neurol. 227, 322–327. doi: 10.1016/j.expneurol.2010.12.005
Weber, M. J., Messing, S. B., Rao, H., Detre, J. A., and Thompson-Schill, S. L. (2014). Prefrontal transcranial direct current stimulation alters activation and connectivity in cortical and subcortical reward systems: A tDCS-fMRI study. Hum. Brain Mapp. 35, 3673–3686. doi: 10.1002/hbm.22429
Wischnewski, M., Engelhardt, M., Salehinejad, M. A., Schutter, D. J. L. G., Kuo, M.-F., and Nitsche, M. A. (2019). NMDA receptor-mediated motor cortex plasticity after 20 Hz transcranial alternating current stimulation. Cereb. Cortex 29, 2924–2931. doi: 10.1093/cercor/bhy160
Woods, A. J., Antal, A., Bikson, M., Boggio, P. S., Brunoni, A. R., Celnik, P., et al. (2016). A technical guide to tDCS, and related non-invasive brain stimulation tools. Clin. Neurophysiol. 127, 1031–1048. doi: 10.1016/j.clinph.2015.11.012
Wu, L., Cao, T., Li, S., Yuan, Y., Zhang, W., Huang, L., et al. (2022). Long-term gamma transcranial alternating current stimulation improves the memory function of mice with Alzheimer’s disease. Front. Aging Neurosci. 14:980636. doi: 10.3389/fnagi.2022.980636
Ye, H., and Steiger, A. (2015). Neuron matters: Electric activation of neuronal tissue is dependent on the interaction between the neuron and the electric field. J. NeuroEng. Rehabil. 12:65. doi: 10.1186/s12984-015-0061-1
Keywords: transcranial electrical stimulation (tES), excitation-inhibition balance, tACS (transcranial alternating current stimulation), tRNS (transcranial random noise stimulation), tDCS (transcranial direct current stimulation), synaptic plasticity, animal models, translational neuroscience
Citation: Estévez-Rodríguez M, Sánchez-Garrido Campos G, Zafra ÁM, Cordones I and Márquez-Ruiz J (2025) Modulating excitation/inhibition balance through transcranial electrical stimulation: physiological mechanisms in animal models. Front. Neurosci. 19:1609679. doi: 10.3389/fnins.2025.1609679
Received: 10 April 2025; Accepted: 24 June 2025;
Published: 15 July 2025.
Edited by:
Nemanja Jovicic, University of Kragujevac, SerbiaReviewed by:
David William Carmichael, King’s College London, United KingdomMarta Carè, San Martino Hospital (IRCCS), Italy
Copyright © 2025 Estévez-Rodríguez, Sánchez-Garrido Campos, Zafra, Cordones and Márquez-Ruiz. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Javier Márquez-Ruiz, am1hcnF1ZXpAdXBvLmVz