 Yu Zhang1
Yu Zhang1 Hailong Lun2
Hailong Lun2 Naiqiang Zhu3,4*†Ning Yang3Kaikai Ding1Bin Chen4Chengbing Chang4Haipeng Gu4Yanqi Liu4
Naiqiang Zhu3,4*†Ning Yang3Kaikai Ding1Bin Chen4Chengbing Chang4Haipeng Gu4Yanqi Liu4- 1Graduate School of Chengde Medical University, Chengde, Hebei, China
- 2Tangshan Nanhu Hospital, Department of Orthopedic, Tangshan, Hebei, China
- 3Hebei Key Laboratory of Panvascular Diseases, Affiliated Hospital of Chengde Medical University, Chengde, Hebei, China
- 4Department of Minimally Invasive Spine Surgery, Affiliated Hospital of Chengde Medical University, Chengde, Hebei, China
Introduction: Osteosarcoma (OS), a prevalent metastatic cancer among young individuals, is associated with a grim prognosis. Long non-coding RNAs (lncRNAs), including C1QTNF1-AS1, are pivotal regulators of cancer cell proliferation and motility. As an oncogene, C1QTNF1-AS1 is implicated in various tumor types, such as colorectal, pancreatic, hepatocellular carcinomas, and OS. The aim of this study was to investigate the functions and underlying mechanisms of C1QTNF1-AS1 in the progression of osteosarcoma.
Methods: This investigation focused on elucidating the functional roles and mechanisms of C1QTNF1-AS1 in OS cells. Bioinformatics tools were utilized to identify the interaction between microRNA miR-34a-5p and C1QTNF1-AS1, as well as the targeting of LDHA and PDK3 by miR-34a-5p. Dual-luciferase reporter assays and RNA immunoprecipitation were employed to validate these interactions. Expression profiles of C1QTNF1-AS1, miR-34a-5p, LDHA, and PDK3 in osteosarcoma cells were analyzed using RT-PCR and western blot analyses, revealing their intricate relationships. The impact of these molecules on OS cell proliferation, invasion, and migration was assessed through CCK-8, Transwell, and Cell scratch assay. Moreover, the effects on aerobic glycolysis in OS cells were examined by quantifying ATP levels, lactate production, glucose uptake capacity, and the extracellular acidification rate.
Results: The findings indicated a significant decrease in C1QTNF1-AS1 expression levels in OS cells compared to normal osteoblasts. A parallel downregulation trend of miR-34a-5p was also observed in OS cells. Silencing C1QTNF1-AS1 led to a marked upregulation of LDHA and PDK3 in OS cells, which was partially attenuated by miR-34a-5p mimics. Functional evaluations demonstrated that suppression of C1QTNF1-AS1 accelerated OS cell growth, motility, invasiveness, and the Warburg effect. Conversely, the overexpression of miR-34a-5p mitigated these stimulatory effects, suggesting a regulatory role in modulating OS progression.
Discussion: Our research emphasizes the critical role of C1QTNF1-AS1 in the pathogenesis of osteosarcoma (OS). We discovered that the downregulation of C1QTNF1-AS1 indirectly upregulates the expression of LDHA and PDK3 by suppressing miR-34a-5p, which functions as a regulator of the Warburg effect. This cascade of events promotes OS progression by enhancing glycolytic metabolism and supplying energy for cancer cell growth, migration, and invasion. These findings suggest a potential therapeutic target and highlight the importance of understanding the regulatory network involving lncRNAs in cancer metabolism and progression.
1 Introduction
Osteosarcoma, commonly abbreviated as OS, represents a highly aggressive form of connective tissue malignancy characterized by the capability of cancerous cells to produce bone and osteoid tissues (1). It is the leading primary malignancy affecting bone tissues. Posing significant challenges for treatment due to its aggressive nature and propensity for dissemination (2). OS continues to yield poor patient outcomes despite rigorous therapeutic interventions (3). Consequently, there is an urgent need to unravel the intricate molecular mechanisms underpinning OS development, identify novel biomarkers to facilitate early diagnosis, and explore innovative therapeutic strategies to address this formidable disease.
To elucidate the Warburg effect, it implies that cancer cells, under hypoxic conditions, favor lactate fermentation over oxidative phosphorylation for energy generation (4). This metabolic shift is marked by intensified glycolysis, augmented glucose consumption, escalated lactate excretion, and decreased oxygen utilization within tumor cells (5, 6). Extensive research has confirmed the manifestation of the Warburg effect in cancer cells, emphasizing its pivotal contribution to tumor progression and expansion. This phenomenon underscores the metabolic reprogramming of tumor cells, pivotal for their survival and aggressive growth.
PDK3, or pyruvate dehydrogenase kinase isoform 3, serves as a key regulator of intracellular energy metabolism, particularly within tumor cells (7). By inhibiting pyruvate dehydrogenase activity, PDK3 exerts a pronounced influence on the metabolic profile of cancer cells (8). This enzyme plays a central role in perpetuating the Warburg effect, a phenomenon intimately tied to energy metabolism and tumor cell vitality (9). Meanwhile, lactate dehydrogenase A (LDHA), a pivotal player in intracellular lactate dynamics, is intricately linked to both glycolysis and the Warburg effect (10). As a gene and enzyme implicated in glycolysis, LDHA facilitates the generation and accumulation of lactate, thereby bolstering cell survival and proliferation (11). This metabolic pathway is fundamental to cellular energy procurement and, as previous investigations have attested, is a hallmark of OS, underscoring its critical role in the disease.
Long non-coding RNAs (lncRNAs), comprising RNA molecules longer than 200 nucleotides without protein-coding capability, pervade the human genome (12). These transcripts exert substantial influence on the initiation, progression, and development of tumors, either bolstering or retarding these processes (13, 14). For example, FEZF1-AS1, through its interaction with miR-4443, modulates the NUPR1-axis, thereby fostering the development of OS (15). In breast cancer, augmented levels of C1QTNF1-AS1 perturb growth, invasion, and dissemination of cancer cells by regulating various signaling cascades, such as Wnt/β-catenin, PI3K/Akt, and NF-κB (16–19). Likewise, in lung cancer, high expression of C1QTNF1-AS1 orchestrates critical cellular functions, including proliferation, invasion, and angiogenesis, by targeting genes like EGFR, HIF-1α, and VEGF (20–22). However, the precise function of C1QTNF1-AS1 in OS remains an enigma, necessitating further investigation.
MicroRNAs (miRNAs), consisting of approximately 22 nucleotides, are single-stranded, non-coding RNAs that have garnered substantial research attention (23). miR-323a-3p, for instance, amplifies LDHA expression, thereby augmenting lactate generation and fostering metastatic and invasive capabilities in osteosarcoma (24). Conversely, miR-199b-3p retards the expansion of OS by targeting PDK1 (25). Notably, miR-34a-5p exhibits abnormal expression patterns in OS cells, and its levels are intimately tied to cellular responsiveness, tumor stage, lung metastasis potential, and patient prognosis (26, 27).
Competitive endogenous RNA (ceRNA) are RNA molecules that modulate gene expression by interacting with microRNA (miRNA) (28). This class includes various types such as mRNA, lncRNA, pseudogenes, and circRNA (29). The ceRNA mechanism centers on the “sponge effect,” where these molecules bind to miRNA, preventing it from binding to and inhibiting target mRNAs (30, 31). This interaction is based on complementary sequences between ceRNA and miRNA targeting sites (32). The biological significance of ceRNA lies in its role within a gene regulatory network, where ceRNAs compete for miRNAs, creating a complex regulatory system (33). This discovery enhances our comprehension of RNA functions and gene regulation, offering insights into disease mechanisms.
The study unveiled intriguing findings regarding the roles of C1QTNF1-AS1 and miR-34a-5p in OS cells. Specifically, it was observed that both C1QTNF1-AS1 and miR-34a-5p exhibited reduced expression levels in OS cells compared to normal osteoblasts. Furthermore, silencing C1QTNF1-AS1 led to a marked enhancement in OS cell proliferation and the Warburg effect, a phenomenon wherein cancer cells preferentially utilize glycolysis for energy generation even in the presence of oxygen. Bioinformatics analysis illuminated a potential interaction between C1QTNF1-AS1 and miR-34a-5p, suggesting that they partially base pair. Moreover, it was hypothesized that LDHA and PDK3, enzymes implicated in glycolysis and energy metabolism, could be target genes of miR-34a-5p. This interaction was subsequently validated using dual-luciferase reporter assays. The study delved deeper into the underlying mechanisms by exploring whether suppressing C1QTNF1-AS1 indirectly elevated LDHA and PDK3 expression levels through downregulation of miR-34a-5p. This, in turn, was postulated to influence the Warburg effect and accelerate tumor progression.
The research provides valuable insights into the complex interplay between lncRNAs, miRNAs, and their target genes in modulating cancer cell behavior. By shedding light on the regulatory mechanisms governing glycolysis and energy metabolism in OS cells, this study opens new avenues for the development of targeted therapies aimed at disrupting these pathways and ultimately retarding tumor growth and progression.
2 Methods
2.1 Cell culture and reagents
Human osteosarcoma cell lines (including MG63, Saos-2, U2OS and HOS) and normal osteoblasts (HFOB1.19) were obtained from the ATCC Cell Bank located in Manassas, Virginia, USA. These cells were cultivated in Eagle’s medium supplemented with 10% fetal bovine serum sourced from Thermo Fisher Scientific (Manassas, VA, USA), along with 0.1% penicillin and 0.1% streptomycin procured from Invitrogen (Carlsbad, CA, USA). All osteosarcoma cell lines were cultured at 37°C, whereas the HFOB1.19 cells were maintained at 34°C. All cells were incubated in an environment containing 5% carbon dioxide.
2.2 Bioinformatics analysis
The dataset GSE42352 originates from the Gene Expression Omnibus (GEO) database, which is accessible at http://www.ncbi.nlm.nih.gov/geo. The downloaded data comprised 84 disease samples and three healthy controls. The SangerBox platform (http://sangerbox.com) and the limma package (version 3.46.0, Linear Models for Microarray Data, from Bioconductor) were utilized to analyze the two sample groups.
2.3 Database analysis
Data pertaining to potential target genes of miR-34a-5p were acquired utilizing the online software tools TarBase (accessible at https://dianalab.e-ce.uth.gr), miRDB (located at https://mirdb.org), and TargetScan (found at https://www.targetscan.org). Analysis conducted with RNAhybrid and miRanda indicated that C1QTNF1-AS1 targets miR-34a-5p. Furthermore, predictions from TargetScan, miRanda, and miRWalk databases suggested that LDHA and PDK3 are potential targets of miR-34a-5p.
2.4 Real-time quantitative PCR
In accordance with the manufacturer’s protocol, RNA extraction from osteosarcoma cell lines including MG63, Saos-2, U2OS and HOS was accomplished using the TRIzol reagent sourced from Thermo Fisher Scientific. Subsequently, the RNA samples were reverse transcribed into cDNA using the PrimeScript RT Kit provided by Takara. Gene expression levels were then quantified via qRT-PCR on an ABI7500 Quantitative PCR instrument from ABI Corporation, employing the SYBR Prex Ex Taq II Kit also from Takara. GAPDH served as the stable reference gene for normalization during the analysis.
2.5 Western blotting
Total protein extraction was carried out using RIPA buffer (Sigma) supplemented with a protease inhibitor from Roche. The extracted proteins were then resolved on a 12% SDS-PAGE gel. After electrophoresis, the proteins were transferred to membranes which were subsequently blocked with 5% skim milk for an hour. Overnight incubation with the primary antibody at 4°C was followed by rinsing the membranes with TBST. Subsequently, the membranes were incubated with secondary antibodies and washed again with TBST. Luminescence was detected using an ECL detection kit from Share-bio. For quantification, densitometric analysis of the immunoblotted proteins was conducted utilizing ImageJ software.
2.6 Cell count kit-8 test
Following transfection, U2OS and MG63 cells were plated in 96-well plates at a concentration of 3,000 cells per well. Each well received 10μL of CCK-8 solution (Dojindo Molecular Technologies) and was incubated at 37°C with 5% CO2 for 0, 24, and 48 hours. The optical density at 450 nm was then determined using a BioTek microplate reader. The findings reflect three separate trials.
2.7 Cell transfection
Cells with C1QTNF1-AS1 knockdown (si-lnc) and those with a negative control (si-NC) were produced. GenePharma (Shanghai, China) supplied the miR-34a-5p mimic (miR-mim) along with its control (miR-NC). Lipofectamine 3000 reagent (Invitrogen) was utilized for transfection, following the guidelines provided by the manufacturer.
2.8 Measurement of glucose metabolism, lactate production, and intracellular ATP levels
The rate of glucose metabolism was assessed using the colorimetric glucose uptake test kit provided by Sigma-Aldrich. The determination of intracellular ATP concentration was carried out according to the protocol provided by Promega Corporation (Madison, Wisconsin, USA), using their ATP Assay Kit. Fluorescence measurements of bioluminescence were taken using a fluorometer produced by Perkin Elmer located in Waltham, Massachusetts, USA. ATP levels were calculated based on the standard curve method. Furthermore, the extracellular lactate levels were measured utilizing a lactate assay kit sourced from BioVision (Zurich, Switzerland), in accordance with the manufacturer’s protocol. All measurements were normalized to the cellular protein levels. The experiment was independently replicated three times.
2.9 Transwell assay
A matrix-coated 24-well span chamber (8μm aperture) was prepared for cell invasion assays. U2OS and MG63 cells were incubated in a serum-free medium in the upper chamber. The lower chamber received medium with 10% FBS. Following a 48-hour incubation period, the cells that had migrated to the lower chamber were treated with methanol and stained using a 0.1% crystal violet solution. The invading cells were examined using an Olympus inverted microscope.
2.10 Cell scratch assay
Cell scratch tests were utilized to evaluate the movement of OS cell lines. Once the U2OS and MG63 cells achieved 90% confluence in 24-well plates, a sterile plastic tip was used to scrape the monolayer, followed by two washes with phosphate-buffered saline (PBS) to eliminate cell debris. The cells were then incubated in complete growth medium. Ultimately, the cells that moved to the damaged region were gathered at 0 and 24 hours following the initial scratch and examined using an inverted microscope (Olympus) for each injury. The relative distances of the cell scratches were analyzed using ImageJ software.
2.11 Dual-luciferase activity measurement
A dual-luciferase reporter assay was conducted by inserting either wild-type or mutant lncRNA-C1QTNF1-AS1/LDHA/PDK3 into the pmirGLO vector (Universal Biotech, China). MG63 cells were planted in 48-well plates at a concentration of 5×104 cells per well. For 48 hours, the luciferase reporter plasmid along with miR-34a-5p mimic or mimic-NC were introduced using Liposome 3000 (Invitrogen). The activities of firefly and Renilla luciferase were evaluated using a dual-luciferase reporter assay kit from Promega, USA. The activity of Firefly luciferase was adjusted relative to that of Renilla luciferase. Every test was conducted three times.
2.12 Measurement of glycolysis
The extracellular acidification rate (ECAR) of cultured cells was measured using an XF96 metabolic flux analyzer from Seahorse Biosciences, located in Billerica, MA, USA, in accordance with the manufacturer’s instructions. Briefly, 80 µL of a suspension containing 3 × 10^4 target cells were dispensed into each well of an XF96 96-well plate provided by Seahorse Biosciences and incubated overnight at 37°C. The XF sensor box, manufactured by Seahorse Bioscience, was prepared with XF calibration solution and left to incubate overnight at 37°C, without the addition of CO2. On the subsequent day, the standard medium was replaced with XF assay-adapted DMEM (containing 1 g/mL glucose, pH 7.4; sourced from Seahorse Biosciences), and the cells were further incubated for an hour at 37°C in a CO2-free environment. Sequential additions of 10 mM glucose, 1 mM oligomycin (provided by Sigma-Aldrich), and 80 mM 2-deoxyglucose (D8375; Sigma-Aldrich) were made to assess ECAR. The obtained data were analyzed using XFe Wave software, also provided by Seahorse Biosciences.
2.13 RNA immunoprecipitation assay
This study employed the RIP Kit manufactured by Bersinbio Company (Guangzhou, China), adhering strictly to the manufacturer’s operating instructions, to assess the interactions among LDHA, PDK3, miR-34a-5p, and the AGO2 protein. During the experimental procedure, cells subjected to oxidative stress were initially washed with phosphate-buffered saline (PBS). Following this, cell lysis was conducted in RIP buffer supplemented with proteasome inhibitors. The resultant lysate was then co-incubated overnight with antibodies specifically targeting the AGO2 protein or with control IgG antibodies, to facilitate the formation of RNA-protein complexes. Subsequently, these complexes were bound to protein A/G magnetic beads and subjected to proteinase K treatment to eliminate the protein components. Ultimately, the purified RNA was analyzed using qRT-PCR.
2.14 Statistical analysis
Data were analyzed and graphed using GraphPad Prism 9 (version 9.4.0). Data are presented as mean ± standard deviation, and group differences were analyzed using the t-test Or the One-way test. A p-value of less than 0.05 was considered statistically significant.
3 Results
3.1 C1QTNF1-AS1 expression was significantly downregulated in OS cells
The SangerBox platform was utilized to analyze gene expression profiles from the GSE42352 dataset on GEO, aiming to pinpoint genes with differential expression in OS cells. Differentially expressed genes between OS cells and normal osteoblasts were identified using the limma package (log FC > 1 and p < 0.05) and volcano and heatmap plots were prepared in The R Project for Statistical Computing (R version 4.4.0) (Figures 1A, B). Among these differentially expressed genes, C1QTNF1-AS1 was significantly downregulated in OS cells compared to that in normal osteoblasts (Figure 1C). Initially, we assessed the expression levels of C1QTNF1-AS1 in both osteoblasts (HFOB 1.19) and various osteosarcoma cell lines (Saos2, MG63, HOS, and U2OS) through qRT-PCR analysis. The findings indicated that C1QTNF1-AS1 expression was notably reduced in osteosarcoma cell lines relative to normal human osteogenic cell lines, with a marked decrease observed in MG63 and U2OS cell lines (Figure 1D).
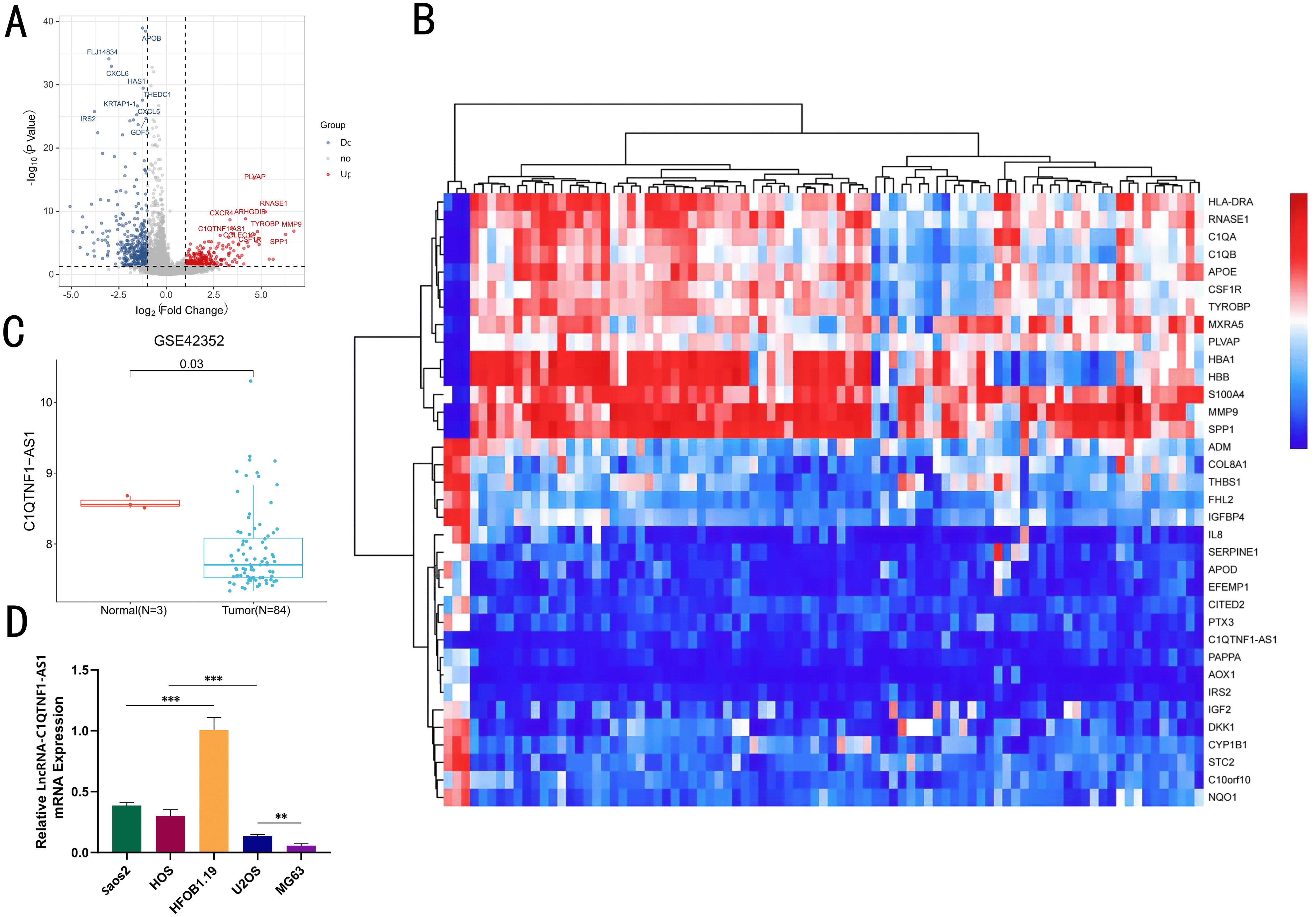
Figure 1. The levels of LncRNA‐C1QTNF1-AS1 are reduced in OS. A volcano plot showcasing the LncRNA with differential expression in GEO datasets (A). A heatmap illustrating the variably expressed LncRNA in GSE42352, sourced from the GEO database (B). Expression of LncRNA C1QTNF1-AS1 in normal cells and OS cells of dataset (C). Quantitative real-time PCR (D) was employed to measure the levels of LncRNA-C1QTNF1-AS1 in U2OS, Saos2, MG63, and HOS cells compared to hFOB1.19 cells. Results are shown as average ± standard deviation. **P<0.01, ***P<0.001.
3.2 Silencing of C1QTNF1-AS1 significantly promoted OS cell development and the Warburg effect in vitro
To investigate the precise role of C1QTNF1-AS1 in osteosarcoma cells, we created a cell line with C1QTNF1-AS1 knocked down (si-lnc) and a corresponding negative control (si-NC). The results of the CCK-8 proliferation assay (Figure 2A), cell scratch tests (Figures 2B, C), along with the Transwell experiment (Figures 2D, E) demonstrated that reducing C1QTNF1-AS1 levels enhanced the growth, movement, and invasive capabilities of MG63 and U2OS cells. The Warburg effect is essential for tumor progression and aids in the proliferation of cancer cells. We investigated the potential correlation between C1QTNF1-AS1 and the Warburg effect in the development of osteosarcoma (OS) by examining glucose levels, ATP production, and lactate production in the supernatants of OS cell cultures. To ascertain whether the silencing of C1QTNF1-AS1 contributes to the regulation of aerobic glycolysis, we utilized a metabolic flux analyzer to measure extracellular acidification rates (ECAR). Our research revealed that the silencing of C1QTNF1-AS1 led to a substantial increase in ATP production (Figure 2F) and lactate production (Figure 2G) in MG63 and U2OS cells. Simultaneously, it notably decreased the glucose levels in the supernatants of these cells (Figure 2H), indicating that the cells consumed more glucose. Furthermore, we found that the knockdown of C1QTNF1-AS1 resulted in enhanced glycolysis levels in U2OS and MG63 cells (Figure 2I). Taken together, these data suggest that the silencing of C1QTNF1-AS1 promotes aerobic glycolysis in OS cells.
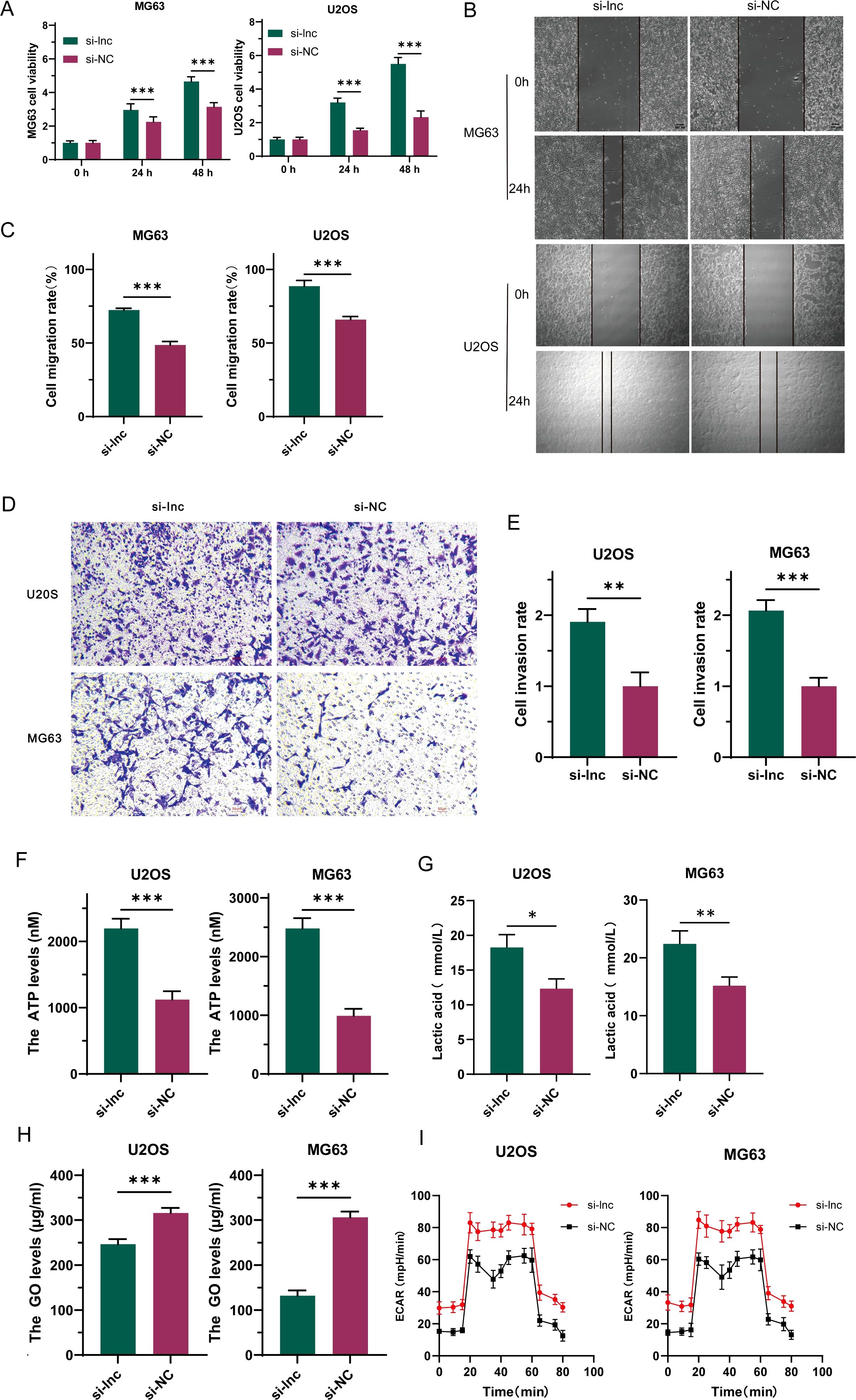
Figure 2. LncRNA-C1QTNF1-AS1 exhibits inhibitory effects on the progression of osteosarcoma (OS) in laboratory conditions. The CCK-8 assay was utilized to evaluate the growth rate of OS cells following the inhibition of C1QTNF1-AS1 (A). The migration capability of cells after C1QTNF1-AS1 knockdown was assessed through a cell scratch assay (B, C). The invasive potential post-inhibition of C1QTNF1-AS1 was further examined using a Transwell assay (D, E). To gain insights into the metabolic alterations, ATP (F) and lactate production levels (G) in C1QTNF1-AS1 knockdown cells were measured using respective assays, while glucose concentrations in cell supernatants were determined via a glucose assay (H). To definitively determine the impact of C1QTNF1-AS1 silencing on glycolysis rates, the extracellular acidification rate (ECAR) was assessed using a metabolic flux analyzer (I). Results are shown as average ± standard deviation. *P<0.05, **P<0.01, ***P<0.001.
3.3 miR-34a-5p was identified as a direct target of C1QTNF1-AS1 and showed consistent expression trends in OS cells
In order to investigate how C1QTNF1-AS1 influences aerobic glycolysis in OS, we employed three gene prediction tools—TarBase, miRDB, and TargetScan—to identify mRNA candidates targeted by C1QTNF1-AS1.Intersection of the three databases revealed that only miR-34a-5p was associated with aerobic glycolysis (Figure 3A). The binding site of C1QTNF1-AS1 to miR-34a-5p was predicted using the RNAhybrid and miRanda algorithms, and a mutated sequence, mut, was designed. The dual-luciferase assay results showed that the miR-34a-5p mimic could bind to C1QTNF1-AS1 wild-type to increase luciferase activity compared to the expression of the NC mimic. Nonetheless, altering the binding site resulted in the miR-34a-5p mimic showing no notable impact on luciferase activity (Figure 3B). This suggested that miR-34a-5p attaches to C1QTNF1-AS1 at this location (Figure 3C). In order to investigate the connection between C1QTNF1-AS1 and miR-34a-5p in osteosarcoma cells, we created a stable cell line (si-lnc) and a negative control (si-NC). The qRT-PCR analysis indicated that miR-34a-5p levels in osteosarcoma cells decreased following the silencing of C1QTNF1-AS1 (Figure 3D).
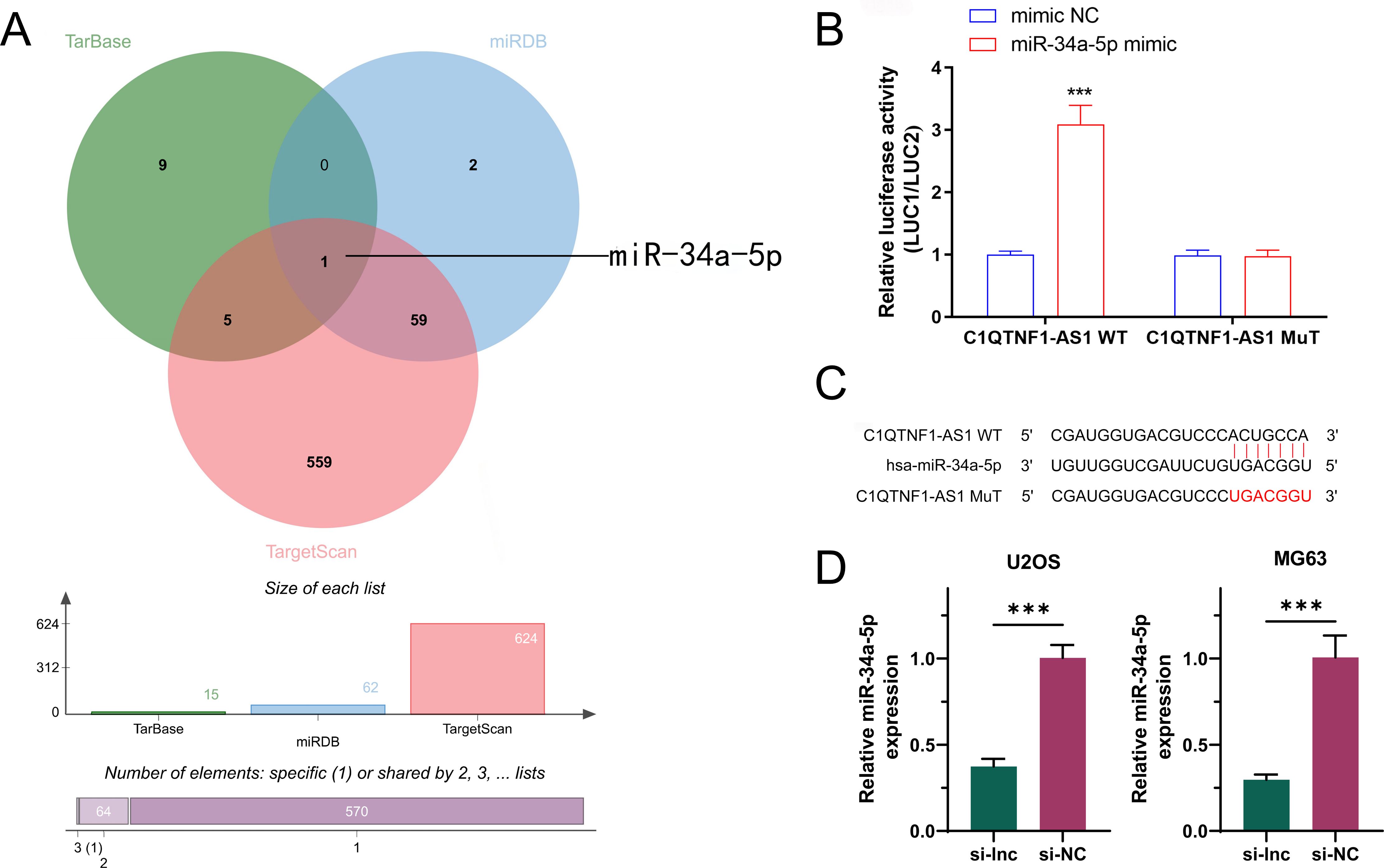
Figure 3. miR-34a-5p is an authentic target of LncRNA‐C1QTNF1-AS1.A Venn diagram illustrating the anticipated glycolysis-associated target genes of lncRNA C1QTNF1-AS1, derived from the TarBase miRDB, and TargetScan databases (A). Dual-luciferase reporter experiments were conducted on OS cells transfected with either miR-34a-5p UTR wild-type (WT) or mutant (MUT) forms, alongside lncRNA C1QTNF1-AS1 plasmid or a control plasmid (B). miR-34a-5p 3′‐UTR contains one predicted LncRNA C1QTNF1-AS1 binding site (C). Expression levels of miR-34a-5p in OS cells after transfection with either a negative control or a C1QTNF1-AS1 knockdown plasmid (D). Results are shown as the average ± standard deviation. Results are shown as average ± standard deviation. ***P<0.001.
3.4 miR-34a-5p suppressed the growth and progression of OS cells by modulating the Warburg effect
To explore the function of miR-34a-5p in osteosarcoma, this study constructed stable cell lines overexpressing miR (miR-mim) and negative controls (mim-NC) and performed CCK-8 proliferation assays. The results showed that increased expression levels of miR-34a-5p inhibited the growth of OS cells (Figure 4A). Transwell assay results indicated that elevated miR-34a-5p expression levels suppressed the invasion of OS cells (Figures 4B, C). Scratch assays demonstrated that overexpression of miR-34a-5p inhibited the migration of OS cells (Figures 4D, E). This study found that the overexpression of miR-34a-5p significantly reduced ATP production (Figure 4F) and lactate production (Figure 4G) in MG63 and U2OS cell lines, while significantly increasing the glucose level in the cell supernatant (Figure 4H). This indicated a reduction in glucose consumption by osteosarcoma cells. Metabolic flux analysis revealed that the overexpression of miR-34a-5p significantly decreased the extracellular acidification rate (ECAR) (Figure 4I) in both MG63 and U2OS cell lines, thereby inhibiting the Warburg effect. In summary, miR-34a-5p inhibited the proliferation, migration, and invasion of osteosarcoma cells, as well as aerobic glycolysis.
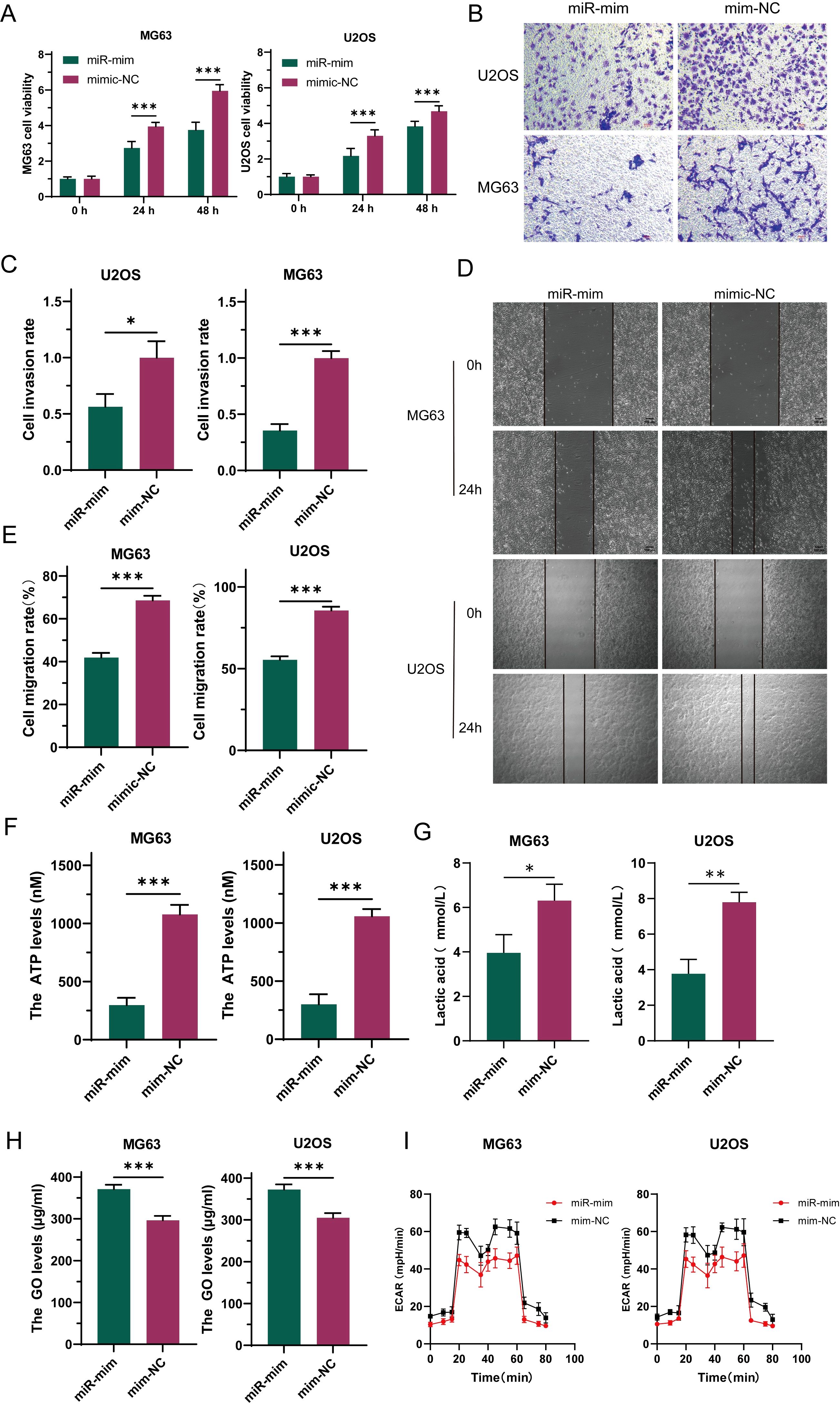
Figure 4. Under laboratory conditions, miR-34a-5p has been demonstrated to inhibit the progression of osteosarcoma (OS). To investigate this, the proliferation rate of OS cells was assessed following miR-34a-5p overexpression using CCK-8 assays (A). Furthermore, the invasive potential of these cells with miR-34a-5p overexpression was evaluated through Transwell experiments (B, C). To determine the effect of miR-34a-5p overexpression on cell migration, scratch assays were conducted on the cells (D, E). Additionally, to assess the production of ATP and lactate in cells overexpressing miR-34a-5p, ATP levels (F), lactate levels (G), and glucose content in the cell supernatant (H) were measured using appropriate assays. Moreover, ECAR detection was performed to evaluate the glycolytic rate of the cells after miR-34a-5p overexpression (I). Results are shown as average ± standard deviation. *P<0.05, **P<0.01, ***P<0.001.
3.5 Silencing of C1QTNF1-AS1 promoted OS progression through miR-34-a-5p–mediated glycolysis
To deeply explore whether C1QTNF1-AS1 inhibits the development and progression of osteosarcoma cells by targeting miR-34a-5p, this study selected U2OS and MG63 cell lines for rescue experiments. By comparing the data from CCK-8 assays (Figure 5A), Transwell assays (Figures 5B, C), and cell scratch assays (Figures 5D, E), it was found that knockdown of C1QTNF1-AS1 significantly promoted the proliferation, migration, and invasion abilities of osteosarcoma cells, while the overexpression of miR-34a-5p partially reversed this effect. Similarly, the elevation of miR-34a-5p levels partially alleviated the impact of C1QTNF1-AS1 inhibition on the Warburg effect of osteosarcoma cells. Further confirmation of this effect was provided by measuring ATP production (Figure 5F), lactate production (Figure 5G), glucose consumption (Figure 5H), and ECAR (Figure 5I). Based on these results, this study revealed that inhibiting C1QTNF1-AS1 enhances the proliferation, migration, and invasion abilities of osteosarcoma cells by blocking the glycolysis driven by miR-34a-5p.
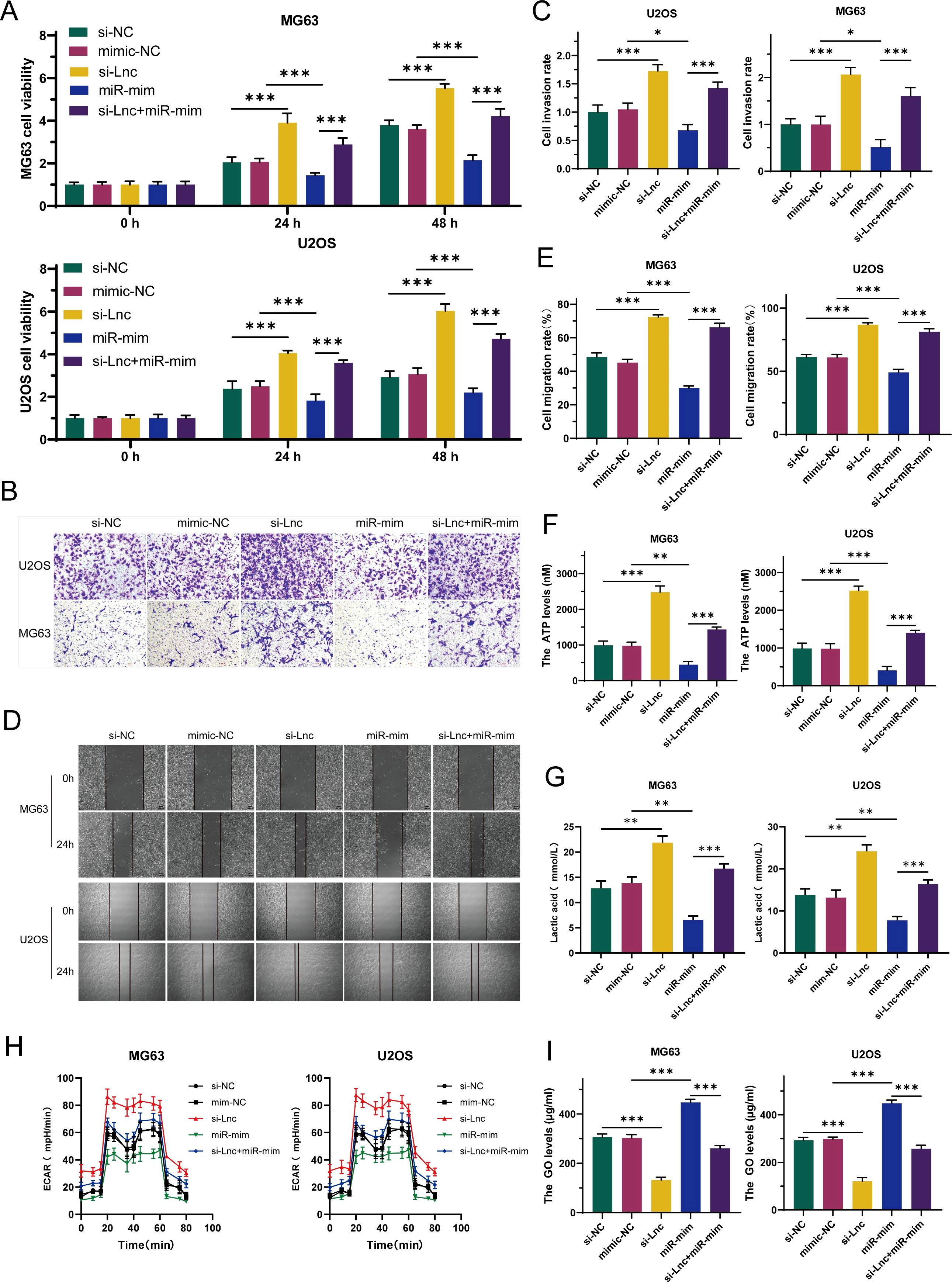
Figure 5. Elevated levels of miR-34a-5p partially restore the oncogenic role suppressed by the downregulation of C1QTNF1-AS1 in osteosarcoma. CCK-8 tests (A), Transwell assays (B, C), and cell scratch tests (D, E) demonstrate that the overexpression of miR-34a-5p somewhat counteracts the enhancement of OS cell proliferation, invasion, and migration induced by the knockdown of C1QTNF1-AS1. Furthermore, assessments of ATP levels (F), lactate production (G), glucose consumption (H), and ECAR (I) indicate that the overexpression of miR-34a-5p also somewhat mitigates the impact of C1QTNF1-AS1 silencing on the Warburg effect in osteosarcoma cells. Results are shown as average ± standard deviation. *P<0.05, **P<0.01, ***P<0.001.
3.6 LDHA and PDK3 were identified as direct targets of miR-34a-5p and showed opposite expression trends in OS cells
The binding sites for LDHA, PDK3, and miR-34a-5p were predicted utilizing the TargetScan, miRanda, and miRWalk algorithms, and a mutated sequence (mut) was subsequently designed. The dual-luciferase assay revealed that the miR-34a-5p mimic attached to both wild-type LDHA and PDK3, resulting in a reduction in luciferase activity compared to the NC mimic expression. However, mutating the binding site rendered the miR-34a-5p mimic ineffective in altering luciferase activity (Figures 6A, B). These findings demonstrated that miR-34a-5p binds to LDHA and PDK3 at the predicted sites (Figures 6C, D). Furthermore, stable cell lines overexpressing miR-34a-5p (miR-mim) and a negative control (mim-NC) were established. The qRT-PCR analysis results indicated that the expression levels of both PDK3 and LDHA were significantly downregulated in osteosarcoma cells following the overexpression of miR-34a-5p (Figure 6E). Western blot analysis further confirmed this observation, demonstrating a significant decrease in the protein expression of PDK3 and LDHA in osteosarcoma cells due to the overexpression of miR-34a-5p (Figures 6F, G). RNA immunoprecipitation (RIP) experiments revealed that the enrichment levels of LDHA and PDK3 were significantly increased in the group overexpressing miR-34a-5p and targeting Argonaute2 (Ago2) (Figures 6H, I). These experimental data collectively demonstrated that miR-34a-5p inhibited the expression of LDHA and PDK3 by directly interacting with their 3′ untranslated regions (3′UTR).
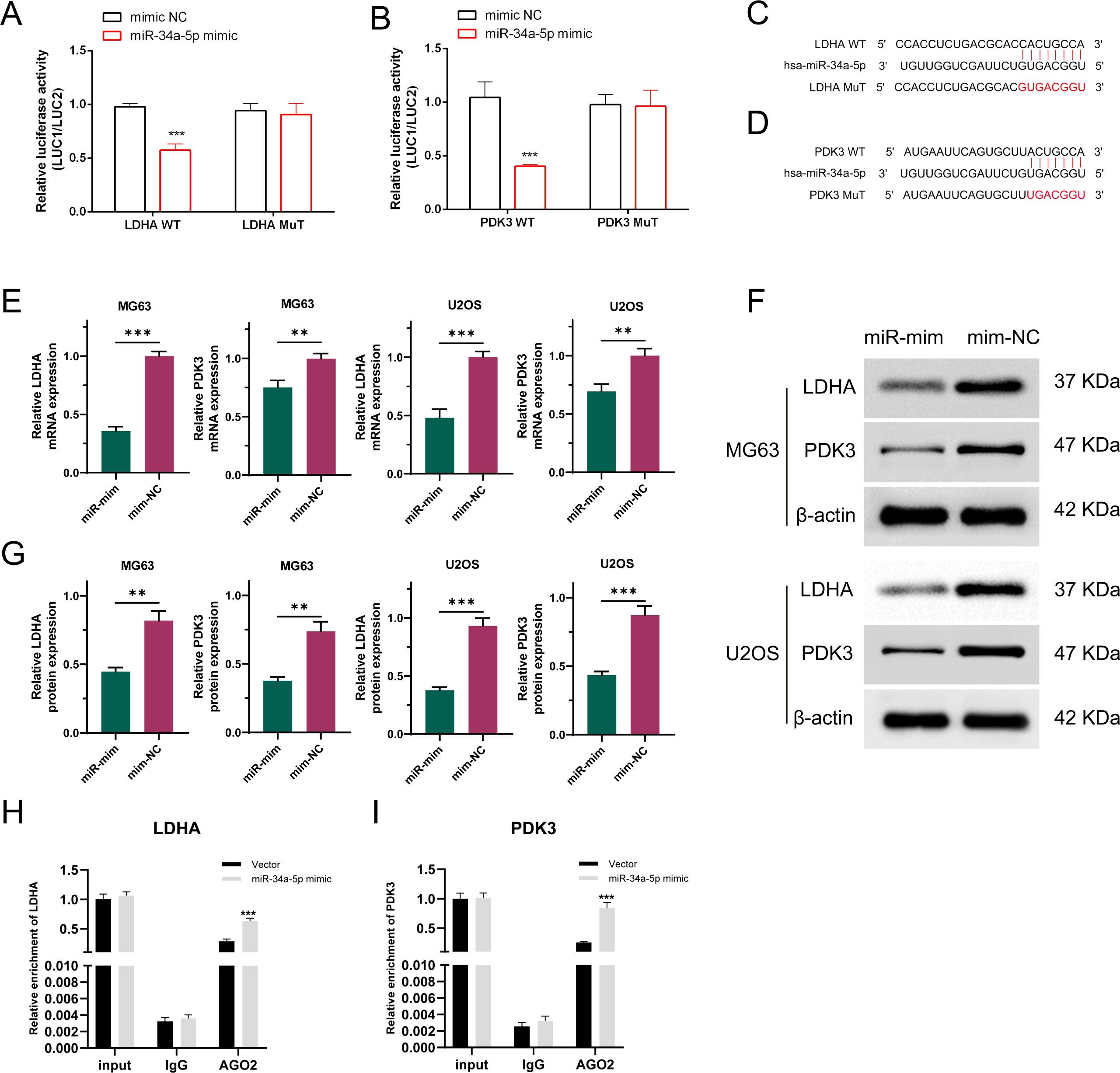
Figure 6. LDHA and PDK3 are authentic targets of miR-34a-5p. Dual-luciferase reporter experiments were conducted on OS cells transfected with either wild-type or mutant LDHA and PDK3 UTR, alongside miR-34a-5p plasmid or a control plasmid (A, B). The 3′-UTRs of LDHA and PDK3 both have a predicted binding site for miR-34a-5p (C, D). As demonstrated by qRT-PCR (E) and Western blot analyses (F, G), the expression levels of LDHA and PDK3 were assessed in osteosarcoma cells transfected with either a negative control plasmid or a plasmid overexpressing miR-34a-5p. RIP experiments were conducted using MG63 cell lysates with AGO2 or IgG antibodies. qRT-PCR results showed the relative enrichment of LDHA and PDK3 mRNA in cells transfected with miR-34a-5p mimics compared to those transfected with NC mimics (H, I). Results are shown as average ± standard deviation. **P<0.01, ***P<0.001.
3.7 Silencing of C1QTNF1-AS1 upregulates LDHA and PDK3 expression in OS cells through inhibition of miR-34a-5p
To explore the interactions between C1QTNF1-AS1, miR-34a-5p, LDHA, and PDK3 in OS cells, we constructed C1QTNF1-AS1 knockdown stable cell lines (si-lnc) and miR-34a-5p overexpression stable cell lines (miR-mim). As demonstrated by qRT-PCR (Figure 7A) and Western blot analyses (Figures 7B, C), the suppression of C1QTNF1-AS1 in OS cells led to a notable rise in LDHA and PDK3 levels. As demonstrated by qRT-PCR (Figure 7D) and Western blot analyses (Figures 7E, F), our rescue experiments revealed that elevating miR-34a-5p levels partially mitigated the enhancement induced by LDHA and PDK3 following the knockdown of C1QTNF1-AS1.
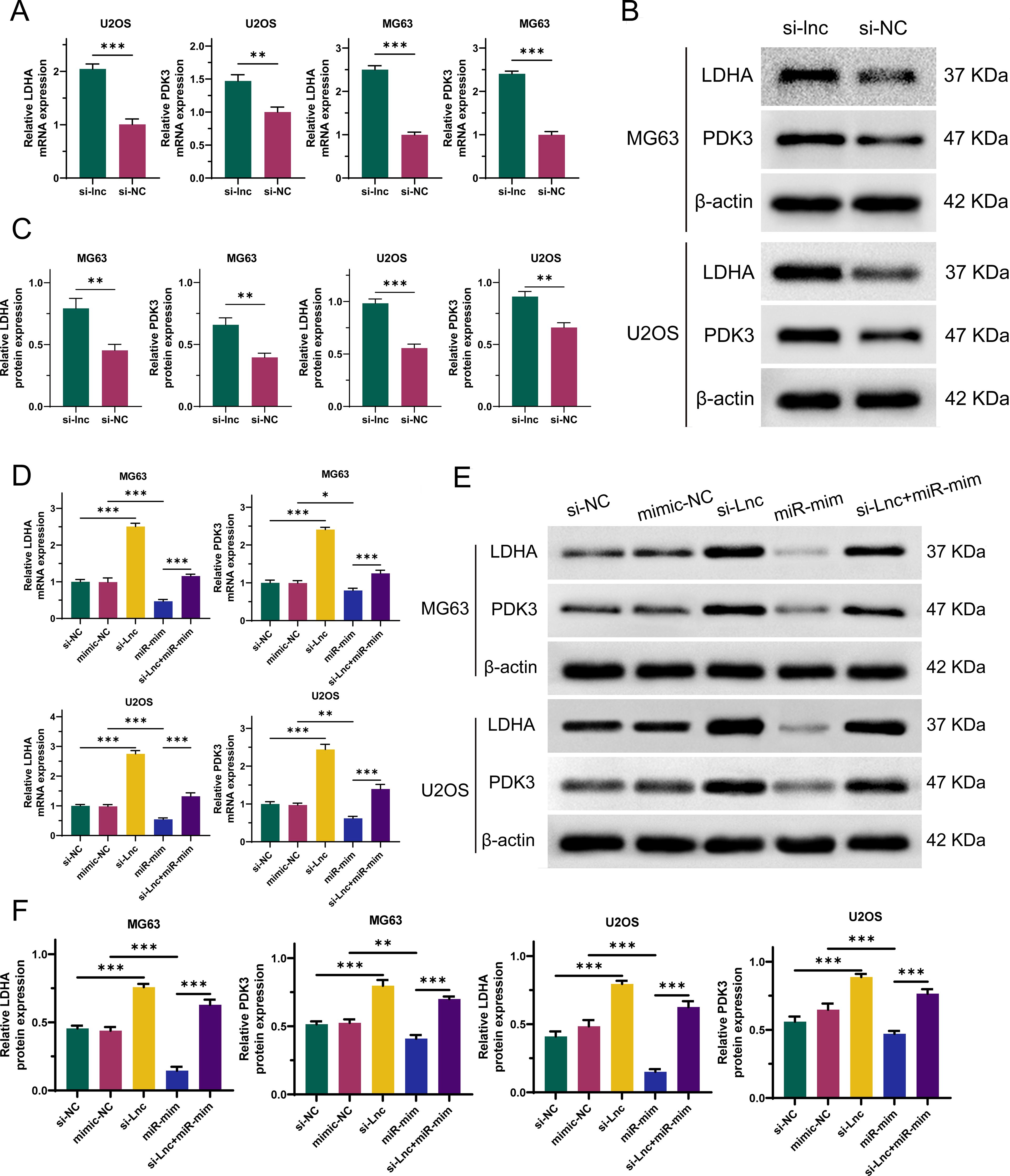
Figure 7. Silencing of C1QTNF1-AS1 upregulates the expression of LDHA and PDK3 in osteosarcoma (OS) cells by inhibiting miR-34a-5p. The altered expression levels of LDHA and PDK3 in OS cells subsequent to C1QTNF1-AS1 silencing are demonstrated through qRT-PCR (A) and Western blot analysis (B, C). As illustrated by qRT-PCR (D) and Western blot analysis (E, F), overexpression of miR-34a-5p partially reverses the enhanced expression of LDHA and PDK3 that is observed following the knockdown of C1QTNF1-AS1. Results are shown as average ± standard deviation. *P<0.05, **P<0.01, ***P<0.001.
4 Discussion
OS, a common form of primary bone malignancy encountered in clinical practice, predominantly strikes adolescents and children and boasts the highest incidence rate among its kind. Characterized by rapid progression and a high tendency for metastasis (34). OS is considered a highly aggressive cancer. In the realm of cancer development, long non-coding RNAs (lncRNAs) occupy a pivotal position (35). This particular study delves into the functional roles and underlying mechanisms of the lncRNA, C1QTNF1-AS1, in the context of osteosarcoma. The findings of this investigation reveal that the inhibition or silencing of C1QTNF1-AS1 leads to an upregulation of LDHA and PDK3 expressions, a phenomenon mediated through the sequestration of miR-34a-5p. This observation underscores the intricate interplay between various regulatory elements within the cancer cell landscape.
The provided text discusses the role of C1QTNF1-AS1 in various types of cancer, including OS. It highlights that C1QTNF1-AS1 expression is reduced in certain cancer cells and that it inhibits multiple types of cancer by affecting proliferation, invasion, epithelial-mesenchymal transition, and apoptosis induction (36, 37). Additionally, C1QTNF1-AS1 counteracts the Warburg effect in hepatocellular carcinoma (38). The study confirms the function of C1QTNF1-AS1 in OS cells, showing that silencing C1QTNF1-AS1 promotes proliferation, migration, invasion, and the Warburg effect in OS cells. Overall, the findings indicate that C1QTNF1-AS1 is essential in the advancement of OS.
miR-34a-5p inhibits Thyroid cancer progression by restricting cell growth and spread and also hampers Head and neck squamous cell carcinoma development by targeting Flotillin-2 (39). In this research, miR-34a-5p was identified as a target of C1QTNF1-AS1, confirmed through dual-luciferase reporter tests. Our analysis revealed a significant association between the levels of miR-34a-5p and C1QTNF1-AS1 in OS cells. Functional assays confirmed that miR-34a-5p overexpression suppresses cell growth, movement, invasion, and the Warburg effect in OS cells. Rescue experiments showed that silencing C1QTNF1-AS1 led to increased OS cell growth, invasion, migration, and the Warburg effect by sequestering miR-34a-5p. These data indicate that C1QTNF1-AS1 regulates OS progression by adsorbing miR-34a-5p.
The Warburg effect plays a pivotal role in fostering the growth and progression of malignant tumors (40). This effect reveals a biological phenomenon wherein, despite ample oxygen availability, osteosarcoma cells preferentially obtain energy through glycolysis rather than through the normal oxidative phosphorylation pathway (41). This metabolic pattern not only provides the necessary energy and biosynthetic precursors for rapid tumor cell proliferation but may also promote tumor invasion and metastasis by altering the tumor microenvironment (42). The presence of the Warburg effect offers new avenues for osteosarcoma treatment. Firstly, researchers can target key enzymes in the glycolytic pathway, such as hexokinase and lactate dehydrogenase, to develop specific inhibitors. These inhibitors can effectively suppress the energy metabolism of tumor cells, thereby inhibiting their growth (43). Secondly, by exacerbating mitochondrial dysfunction and promoting mitochondria-mediated apoptosis, the survival capacity of tumor cells can be further compromised (44). Additionally, intervention in the Warburg effect may synergize with existing chemotherapeutic agents, enhancing the sensitivity of tumor cells to drugs and thus intensifying the efficacy of chemotherapy (45). The development of these therapeutic strategies not only promises to improve the survival rates of osteosarcoma patients but may also enhance their quality of life. Through in-depth investigation of the relationship between the Warburg effect and osteosarcoma, scientists are progressively unveiling the mechanisms of aberrant tumor metabolism, laying a solid foundation for future personalized and precision medicine approaches. With ongoing research, there is good reason to believe that further breakthroughs in the field of osteosarcoma treatment will be achieved, offering more hope and possibilities for patients.
Notably, lactate dehydrogenase A (LDHA), a pivotal enzyme in the final phase of this metabolic shift, is prevalent in multiple cancer cells and is intimately linked to tumor dimensions and clinical outcomes (46, 47). Pyruvate dehydrogenase kinase (PDK), with its four isoforms, contributes significantly to the emergence of the Warburg effect and balances glycolysis with oxidative phosphorylation (48, 49). Our study pinpointed LDHA and PDK3 as direct targets of microRNA-34a-5p (miR-34a-5p), validated through dual-luciferase reporter assays and RNA immunoprecipitation. Additionally, analyzing the expression patterns of C1QTNF1-AS1, miR-34a-5p, LDHA, and PDK3 in OS cells, along with rescue experiments, revealed that the silencing of C1QTNF1-AS1 upregulates LDHA and PDK3 levels in OS cells by sequestering miR-34a-5p.
In summary, this study elucidates the potential biomarker value of C1QTNF1-AS1 and miR-34a-5p in the early diagnosis of osteosarcoma. By quantitatively analyzing the expression levels of these two molecules in patients, early identification and diagnosis of osteosarcoma can be achieved, thereby facilitating timely and effective therapeutic interventions aimed at improving patient survival rates. Furthermore, the study reveals the unique metabolic characteristics of osteosarcoma cells, namely the Warburg effect. This research provides a novel perspective for the formulation of treatment strategies: 1. The restoration or enhancement of miR-34a-5p function can effectively inhibit the proliferation and invasiveness of osteosarcoma cells, and by modulating the expression of enzymes such as LDHA and PDK3, the metabolic profile of osteosarcoma cells can be altered. This lays the theoretical groundwork for the development of miR-34a-5p-targeted therapeutic approaches, potentially representing a novel method for treating osteosarcoma. 2. Suppressing the expression of LDHA and PDK3 in vivo can disrupt the metabolic pathways of osteosarcoma cells, thereby inhibiting their proliferation and invasiveness. This provides a basis for the development of targeted drugs against LDHA and PDK3. 3. Combining miR-34a-5p-targeted therapies with current conventional treatments for osteosarcoma may yield superior therapeutic outcomes. By restoring miR-34a-5p function or inhibiting LDHA and PDK3 activity, patient sensitivity to chemotherapy or radiotherapy can be enhanced, thus improving treatment efficacy and reducing side effects. In summary, this study offers new biomarkers for clinical diagnosis through an in-depth exploration of the mechanisms of action of C1QTNF1-AS1 and miR-34a-5p within osteosarcoma cells, reveals the metabolic characteristics of osteosarcoma, and provides new targets for treatment strategies as well as theoretical support for combination therapy. These findings offer novel insights and approaches for the clinical practice and treatment of osteosarcoma, promising better therapeutic outcomes and quality of life for patients with this condition.
The mechanism diagram of this study can be found in Figure 8.
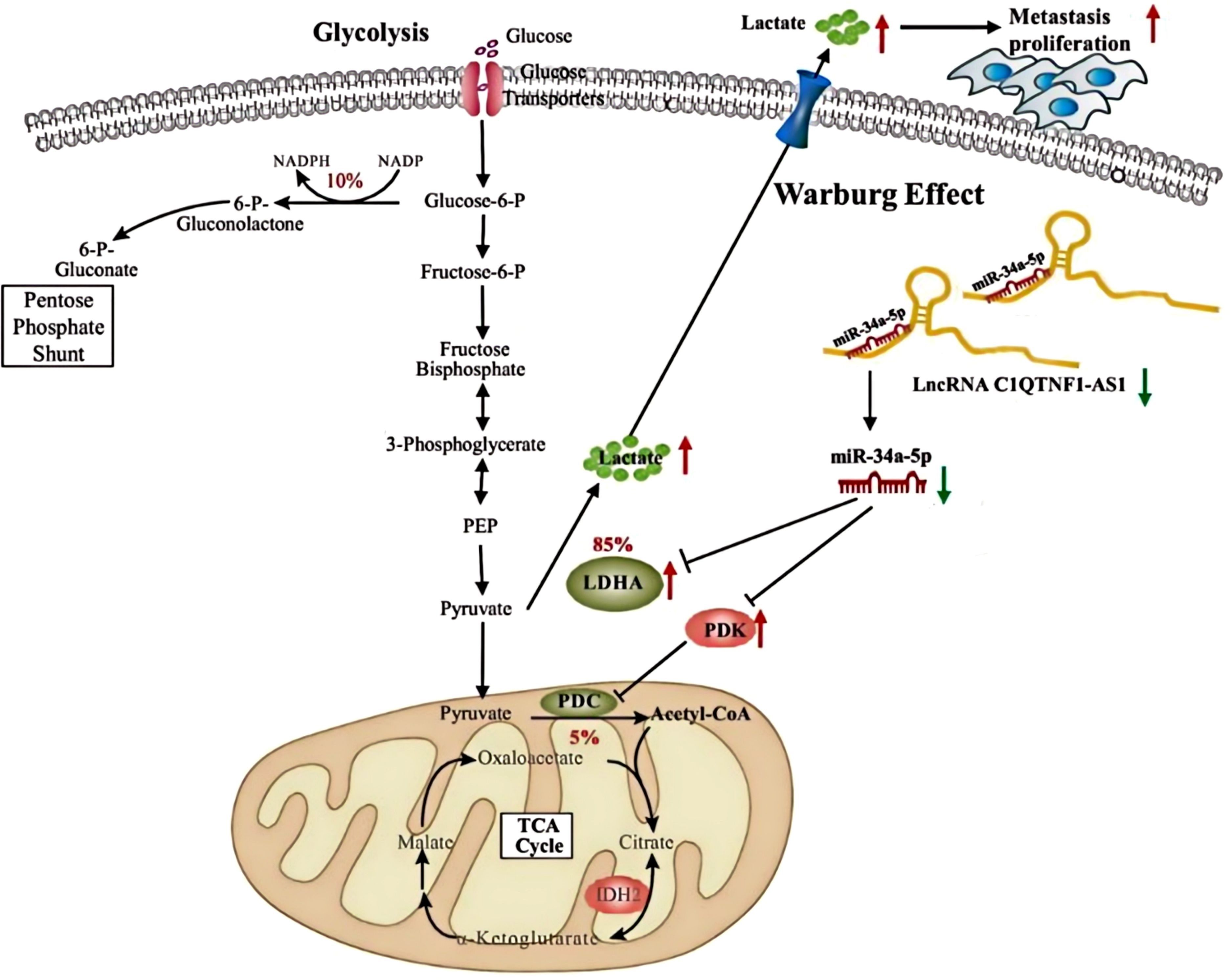
Figure 8. Silent lncRNA-C1QTNF1-AS1 promotes the expression of LDHA and PDK3 by adsorbing miR-34a-5p, indirectly regulating the Warburg effect and facilitating tumor development.
Data availability statement
The datasets presented in this study can be found in online repositories. The names of the repository/repositories and accession number(s) can be found in the article/Supplementary Material.
Author contributions
YZ: Writing – original draft, Writing – review & editing. HL: Writing – review & editing. NZ: Validation, Writing – review & editing. NY: Data curation, Supervision, Writing – review & editing. KD: Writing – review & editing. BC: Supervision, Validation, Writing – review & editing. CC: Data curation, Writing – review & editing. HG: Formal Analysis, Writing – review & editing. YL: Methodology, Writing – review & editing.
Funding
The author(s) declare financial support was received for the research, authorship, and/or publication of this article. This study was supported by Hebei Natural Science Foundation (H2022406038).
Acknowledgments
We thank all the teachers at the Central Laboratory of the Affiliated Hospital of Chengde Medical College for their contributions to this study.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fonc.2024.1485605/full#supplementary-material
Abbreviations
miR, microRNA; lncRNA, long noncoding RNA; OS, osteosarcoma; si, knockdown; mim, overexpression; LDHA, lactate dehydrogenase A; PDK3, also known as pyruvate dehydrogenase kinase isoform 3; NC, negative control; ceRNA, competing endogenous RNA.
References
1. Yu X, Duan W, Wu F, Yang D, Wang X, Wu J, et al. LncRNA-HOTAIRM1 promotes aerobic glycolysis and proliferation in osteosarcoma via the miR-664b-3p/Rheb/mTOR pathway. Cancer Sci. (2023) 114:3537–52. doi: 10.1111/cas.v114.9
2. Zheng C, Li R, Zheng S, Fang H, Xu M, Zhong L, et al. The knockdown of lncRNA DLGAP1-AS2 suppresses osteosarcoma progression by inhibiting aerobic glycolysis via the miR-451a/HK2 axis. Cancer Sci. (2023) 114:4747–62. doi: 10.1111/cas.v114.12
3. Zhu Y, Yang L, Yu Y, Xiong Y, Xiao P, Fu X, et al. Hydroxysafflor yellow A induced ferroptosis of Osteosarcoma cancer cells by HIF-1α/HK2 and SLC7A11 pathway. Oncol Res. (2024) 32:899–910. doi: 10.32604/or.2023.042604
4. Halma MTJ, Tuszynski JA, Marik PE. Cancer metabolism as a therapeutic target and review of interventions. Nutrients. (2023) 15:4245. doi: 10.3390/nu15194245
5. Mathew M, Nguyen NT, Bhutia YD, Sivaprakasam S, Ganapathy V. Metabolic signature of warburg effect in cancer: an effective and obligatory interplay between nutrient transporters and catabolic/anabolic pathways to promote tumor growth. Cancers (Basel). (2024) 16:504. doi: 10.3390/cancers16030504
6. Niepmann M. Importance of michaelis constants for cancer cell redox balance and lactate secretion—Revisiting the warburg effect. Cancers (Basel). (2024) 16:2290. doi: 10.3390/cancers16132290
7. Rai S, Roy G, Hajam YA. Melatonin: a modulator in metabolic rewiring in T-cell Malignancies. Front Oncol. (2024) 13:1248339. doi: 10.3389/fonc.2023.1248339
8. Pucci G, Minafra L, Bravata V, Calvaruso M, Turturici G, Cammarata F, et al. Glut-3 gene knockdown as a potential strategy to overcome glioblastoma radioresistance. Int J Mol Sci. (2024) 25:2079. doi: 10.3390/ijms25042079
9. Anwar S, Shamsi A, Mohammad T, Islam A, Hassan Md. I. Targeting pyruvate dehydrogenase kinase signaling in the development of effective cancer therapy. Biochim Biophys Acta (BBA) - Rev Cancer. (2021) 1876:188568. doi: 10.1016/j.bbcan.2021.188568
10. Yang W, Wei Y, Wang T, Xu Y, Jin X, Qian H, et al. Cytoplasmic localization of SETDB1-induced Warburg effect via c-MYC-LDHA axis enhances migration and invasion in breast carcinoma. Int J Mol Med. (2024) 53:40. doi: 10.3892/ijmm.2024.5364
11. Lin J, Zhong W, Lyu Z, Peng J, Rong Y, Zeng K, et al. Circular RNA circTATDN3 promotes the Warburg effect and proliferation in colorectal cancer. Cancer Lett. (2024) 589:216825. doi: 10.1016/j.canlet.2024.216825
12. Liu C, Shen A, Song J, Cheng L, Zhang M, Wang Y, et al. LncRNA-CCAT5-mediated crosstalk between Wnt/β-Catenin and STAT3 signaling suggests novel therapeutic approaches for metastatic gastric cancer with high Wnt activity. Cancer Commun (Lond). (2023) 44:76–100. doi: 10.1002/cac2.12507
13. Li L, Gao Y, Yu B, Zhang J, Ma G, Jin X. Role of LncRNA H19 in tumor progression and treatment. Mol Cell Probes. (2024) 75:101961. doi: 10.1016/j.mcp.2024.101961
14. Wan Q, Deng Y, Wei R, Ma K, Tang J, Deng Y. Tumor-infiltrating macrophage associated lncRNA signature in cutaneous melanoma: implications for diagnosis, prognosis, and immunotherapy. Aging (Albany NY). (2024) 16:4518–40. doi: 10.18632/aging.205606
15. Zhou C, Xu J, Lin J, Lin R, Chen K, Kong J, et al. Long noncoding RNA FEZF1-AS1 promotes osteosarcoma progression by regulating the miR-4443/NUPR1 axis. Oncol Res. (2018) 26:1335–43. doi: 10.3727/096504018X15188367859402
16. Yang Q, Fu Y, Wang J, Yang H, Zhang X. Roles of lncRNA in the diagnosis and prognosis of triple-negative breast cancer. J Zhejiang Univ Sci B. (2023) 24:1123–40. doi: 10.1631/jzus.B2300067
17. Cai Z, Shi Q, Li Y, Jin L, Li S, LamWong L, et al. LncRNA EILA promotes CDK4/6 inhibitor resistance in breast cancer by stabilizing cyclin E1 protein. Sci Adv. (2023) 9:eadi3821. doi: 10.1126/sciadv.adi3821
18. Zhang K-J, Tan X-L, Guo L. LncRNA TYMSOS facilitates breast cancer metastasis and immune escape through downregulating ULBP3. iScience. (2023) 26:107556. doi: 10.1016/j.isci.2023.107556
19. Dong Y, Zhang T, Li X, Yu F, Guo Y. Comprehensive analysis of coexpressed long noncoding RNAs and genes in breast cancer. J Obstetrics Gynaecology Res. (2019) 45:428–37. doi: 10.1111/jog.2019.45.issue-2
20. Braga EA, Fridman M, Burdennyy A, Loginov V, Dmitriev A, Dmitriev A, et al. Various lncRNA mechanisms in gene regulation involving miRNAs or RNA-binding proteins in non-small-cell lung cancer: main signaling pathways and networks. Int J Mol Sci. (2023) 24. doi: 10.3390/ijms241713617
21. Yang Q, Wang M, Xu J, Yu D, Li Y, Chen Y, et al. LINC02159 promotes non-small cell lung cancer progression via ALYREF/YAP1 signaling. Mol Cancer. (2023) 22:122. doi: 10.1186/s12943-023-01814-x
22. Chen Y-L, Liu Y, Lin Y, Tsai M, Wu S, Chang T, et al. LncRNA SLCO4A1-AS1 suppresses lung cancer progression by sequestering the TOX4-NTSR1 signaling axis. J Biomed Sci. (2023) 30:80. doi: 10.1186/s12929-023-00973-9
23. Yang J, Wang X, Hao W, Wang Y, Li Z, Han Q, et al. MicroRNA-488: A miRNA with diverse roles and clinical applications in cancer and other human diseases. Biomedicine Pharmacotherapy. (2023) 165:115115. doi: 10.1016/j.biopha.2023.115115
24. Chen H, Gao S, Cheng C. MiR-323a-3p suppressed the glycolysis of osteosarcoma via targeting LDHA. Hum Cell. (2018) 31:300–9. doi: 10.1007/s13577-018-0215-0
25. Zhu D, Qi H, Zhu H. hsa-miR-199b-3p suppresses osteosarcoma progression by targeting CCDC88A, inhibiting epithelial-to-mesenchymal transition, and Wnt/beta-catenin signaling pathway. Sci Rep. (2023) 13:12544. doi: 10.1038/s41598-023-39537-0
26. Zhang Y, Mi Y, He C. 2-methoxyestradiol restrains non-small cell lung cancer tumorigenesis through regulating circ_0010235/miR-34a-5p/NFAT5 axis. Thorac Cancer. (2023) 14:2105–15. doi: 10.1111/1759-7714.14993
27. Dai L, Zhang W, Wang Y, Yu K, Le Q, Wu X. circAGTPBP1 promotes the progression of papillary thyroid cancer through the notch pathway via the miR-34a-5p/notch1 axis. iScience. (2023) 26:107564. doi: 10.1016/j.isci.2023.107564
28. Li R, Xu H, Gao X. The ceRNA network regulates epithelial-mesenchymal transition in colorectal cancer. Heliyon. (2023) 9:e14143. doi: 10.1016/j.heliyon.2023.e14143
29. Mao M, Zhang J, Xiang Y, Gong M, Deng Y, Ye D. Role of exosomal competitive endogenous RNA (ceRNA) in diagnosis and treatment of Malignant tumors. Bioengineered. (2022) 13:12156. doi: 10.1080/21655979.2022.2073130
30. Yang S, Wang X, Zhou X, Hou L, Wu J, Zhang W, et al. ncRNA-mediated ceRNA regulatory network: Transcriptomic insights into breast cancer progression and treatment strategies. Biomed Pharmacother. (2023) 162:114698. doi: 10.1016/j.biopha.2023.114698
31. Zhang C, Yu Z, Yang S, Liu Y, Song J, Mao J, et al. ZNF460-mediated circRPPH1 promotes TNBC progression through ITGA5-induced FAK/PI3K/AKT activation in a ceRNA manner. Mol Cancer. (2024) 23:33. doi: 10.1186/s12943-024-01944-w
32. Wu X, Sui Z, Zhang H, Wang Y, Yu Z. Integrated Analysis of lncRNA–Mediated ceRNA Network in Lung Adenocarcinoma. Front Oncol. (2020) 10:554759. doi: 10.3389/fonc.2020.554759
33. Jiang J, Gareev I, Ilyasova T, Shumadalova A, Du W, Yang B, et al. The role of lncRNA-mediated ceRNA regulatory networks in liver fibrosis. Non-coding RNA Res. (2024) 9:463. doi: 10.1016/j.ncrna.2024.01.001
34. Yang Z, Liu Z, Lu W, Guo H, Chen J, Zhang Y. LncRNA WAC-AS1 promotes osteosarcoma Metastasis and stemness by sponging miR-5047 to upregulate SOX2. Biol Direct. (2023) 18:74. doi: 10.1186/s13062-023-00433-2
35. Wang P, Zhu J, Long Q, Wang Y, Xu H, Tao H, et al. LncRNA SATB2-AS1 promotes tumor growth and metastasis and affects the tumor immune microenvironment in osteosarcoma by regulating SATB2. J Bone Oncol. (2023) 41:100491. doi: 10.1016/j.jbo.2023.100491
36. Yu H, Yan X, Chen G, Li R, Yang Z, Liang Z, et al. Dynamic network biomarker C1QTNF1 regulates tumor formation at the tipping point of hepatocellular carcinoma. Biomolecules Biomedicine. (2024) 24:939–51. doi: 10.17305/bb.2024.10103
37. Qiu J, Wang Z, Zhao L, Zhang P, Xu Y, Xia Q. High C1QTNF1 expression mediated by potential ncRNAs is associated with poor prognosis and tumor immunity in kidney renal clear cell carcinoma. Front Mol Biosci. (2023) 10:1201155. doi: 10.3389/fmolb.2023.1201155
38. Lin J. C1QTNF1-AS1 regulates the occurrence and development of hepatocellular carcinoma by regulating miR-221-3p/SOCS3. Hepatol Int. (2021) 15:526–6. doi: 10.1007/s12072-021-10152-3
39. Lu S, Zeng L, Mo G, Lei D, Li Y, Ou G, et al. Long non-coding RNA SNHG17 may function as a competitive endogenous RNA in diffuse large B-cell lymphoma progression by sponging miR-34a-5p. PloS One. (2023) 18:e0294729. doi: 10.1371/journal.pone.0294729
40. Upadhyay S, Khan S, Hassan Md. I. Exploring the diverse role of pyruvate kinase M2 in cancer: Navigating beyond glycolysis and the Warburg effect. Biochim Biophys Acta (BBA) - Rev Cancer. (2024) 1879:189089. doi: 10.1016/j.bbcan.2024.189089
41. Pascale RM, Calvisi DF, Simile MM, Feo CF, Feo F. The warburg effect 97 years after its discovery. Cancers. (2020) 12:2819. doi: 10.3390/cancers12102819
42. Zhong X, He X, Wang Y, Hu Z, Huang H, Zhao S, et al. Warburg effect in colorectal cancer: the emerging roles in tumor microenvironment and therapeutic implications. J Hematol Oncol. (2022) 15:160. doi: 10.1186/s13045-022-01358-5
43. Liao M, Yao D, Wu L, Luo C, Wang Z, Zhang J, et al. Targeting the Warburg effect: A revisited perspective from molecular mechanisms to traditional and innovative therapeutic strategies in cancer. Acta Pharm Sinica B. (2023) 14:953. doi: 10.1016/j.apsb.2023.12.003
44. Barba I, Carrillo-Bosch L, Seoane J. Targeting the warburg effect in cancer: where do we stand? Int J Mol Sci. (2024) 25:3142. doi: 10.3390/ijms25063142
45. Kocianova E, Piatrikova V, Golias T. Revisiting the warburg effect with focus on lactate. Cancers. (2022) 14:6028. doi: 10.3390/cancers14246028
46. Wu C, Zheng C, Chen S, He Z, Hua H, Sun C, et al. FOXQ1 promotes pancreatic cancer cell proliferation, tumor stemness, invasion and metastasis through regulation of LDHA-mediated aerobic glycolysis. Cell Death Dis. (2023) 14:699. doi: 10.1038/s41419-023-06207-y
47. Chen M, Xiao Z, Huang Z, Xue K, Xia H, Zhou J, et al. Glycine decarboxylase (GLDC) plays a crucial role in regulating energy metabolism, invasion, metastasis and immune escape for prostate cancer. Int J Biol Sci. (2023) 19:4726–43. doi: 10.7150/ijbs.85893
48. Gan L, Yang Y, Liang Z, Zhang M, He Y, Zhang S. Targeting the pyruvate dehydrogenase complex/pyruvate dehydrogenase kinase (PDC/PDK) axis to discover potent PDK inhibitors through structure-based virtual screening and pharmacological evaluation. Eur J Medicinal Chem. (2024) 264:116008. doi: 10.1016/j.ejmech.2023.116008
Keywords: OS, C1QTNF1-AS1, miR-34a-5p, LDHA, PDK3, Warburg effect
Citation: Zhang Y, Lun H, Zhu N, Yang N, Ding K, Chen B, Chang C, Gu H and Liu Y (2025) Deciphering the oncogenic network: how C1QTNF1-AS1 modulates osteosarcoma through miR-34a-5p and glycolytic pathways. Front. Oncol. 14:1485605. doi: 10.3389/fonc.2024.1485605
Received: 24 August 2024; Accepted: 18 December 2024;
Published: 09 January 2025.
Edited by:
Brian D. Adams, Brain Institute of America, United StatesReviewed by:
Juliano Andreoli Miyake, Federal University of Santa Catarina, BrazilYang Wang, Sichuan Academy of Medical Sciences and Sichuan Provincial People’s Hospital, China
Copyright © 2025 Zhang, Lun, Zhu, Yang, Ding, Chen, Chang, Gu and Liu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Naiqiang Zhu, emh1bnEyMDEwQDE2My5jb20=
†ORCID: Naiqiang Zhu, orcid.org/0000-0002-9540-4408