 Adriana Reyes-León1
Adriana Reyes-León1 Eduardo Castro-Vargas1,2
Eduardo Castro-Vargas1,2 Ma. del Rocío Juárez-Velázquez1
Ma. del Rocío Juárez-Velázquez1 Daniel Martínez Anaya1Consuelo Salas-Labadía1
Daniel Martínez Anaya1Consuelo Salas-Labadía1 Dafné Moreno-Lorenzana3
Dafné Moreno-Lorenzana3 César Alejandro Galván-Díaz4Norma López-Santiago5Eduardo García-Padilla1,2
César Alejandro Galván-Díaz4Norma López-Santiago5Eduardo García-Padilla1,2 Alda Daniela García-Guzmán4
Alda Daniela García-Guzmán4 Isabel Medina-Vera6
Isabel Medina-Vera6 Patricia Pérez-Vera1*
Patricia Pérez-Vera1*- 1Laboratorio de Genética y Cáncer, Instituto Nacional de Pediatría, Ciudad de México, Mexico
- 2Becario de la Dirección General de Calidad y Educación en Salud, Secretaría de Salud, México, Mexico
- 3CONAHCYT-Instituto Nacional de Pediatría, Ciudad de México, Mexico
- 4Servicio de Oncología, Instituto Nacional de Pediatría, Ciudad de México, Mexico
- 5Servicio de Hematología, Instituto Nacional de Pediatría, Ciudad de México, Mexico
- 6Departamento de Metodología de la Investigación, Instituto Nacional de Pediatría, Ciudad de México, Mexico
Introduction: CRLF2 abnormalities are prevalent in Hispanics from the U.S. and Mexican children with pre-B acute lymphoblastic leukemia (ALL). This trait is associated with unfavorable prognosis. Furthermore, SNPs rs3824662 and rs3781093 in GATA3 have been associated with an increased risk of pre-B ALL. In particular, rs3824662 is associated with CRLF2-ALL, with a higher prevalence in Hispanic patients. Additionally, rs3824662 is associated with adipogenesis, since Hispanic patients have a high prevalence of obesity and overweight, it has been suggested that obesity predisposes to CRLF2-ALL.
Methods: In this study, we evaluated rs3824662 and rs3781093 as predisposition markers for pre-B ALL in Mexican children using Taqman probes.
Results: Both risk alleles were found to be associated with susceptibility to pre-B ALL, predisposition to CRLF2-ALL, overweight status, and overall survival. The risk alleles of both SNPs in Mexican patients were among the most frequent compared with other non-Amerindian populations. SNP rs3824662 and rs3781093 were informative for our patients. Analysis of nutritional status indicated that GATA3 alleles may impact overweight status.
Discussion: Further studies on the relationship between nutritional status and GATA3, as well as analysis of other Amerindian ALL populations, are recommended.
1 Introduction
Environmental and genetic factors are considered promoters of childhood acute lymphoblastic leukemia (ALL) (1, 2). Currently, several single nucleotide polymorphisms (SNPs) in numerous genes, such as rs3824662 and rs3781093 in GATA3 (GATA-binding protein 3), have been associated with the risk of developing this disease; for these SNPs the risk alleles described are A and C respectively (1). In particular, the germline variant rs3824662 has been widely studied in ALL patients from different populations worldwide and has been associated with the high-risk Ph-like subtype, which is part of the precursor B (pre-B) ALL. In this context, the risk homozygote AA at rs3824662 has been associated with poor prognosis in children and adolescents, and few studies have replicated the findings described for rs3781093 (3, 4).
GATA3 encodes a transcription factor that regulates T-cell development and contributes to determining the identity of hematopoietic cells (1). SNP rs3824662 is located in the transcription enhancer region of GATA3 and is considered a cis-acting regulatory element that increases its expression. At the same time, GATA3 overexpression induces the expression of the leukemia oncogene CRLF2 (cytokine receptor-like factor 2), which is altered in 50% of Ph-like patients. In contrast, rs3781093 C risk allele did not affect GATA3 transcription (3, 5).
The SNPs in GATA3 appear to have more influence on ALL susceptibility depending on the ethnic origin of patients; in this regard, Hispanic patients living in the U.S. present a higher frequency than Caucasian, Asian, and African patients (6). Inherited GATA3 variants are associated with Ph-like childhood ALL and the risk of relapse (2, 6).
Interestingly, Hispanic and Mexican patients with pre-B ALL also present a higher frequency of CRLF2 lesions, such as P2RY8::CRLF2 and IGH::CRLF2 rearrangements or CRLF2 overexpression, than Caucasian, Asian, and African-American patients (7–9). Based on this, Mexican patients could present a higher frequency of risk alleles in GATA3; nevertheless, this has not been determined in pre-B ALL cases or in a healthy population.
An independent study showed that Hispanic patients with CRLF2 rearrangements had higher obesity rates than those without CRLF2 lesions. This finding suggests that obesity and GATA3 risk alleles may contribute to CRLF2 altered pre-B ALL (CRLF2-ALL) leukemogenesis and maintenance, through obesity-induced phosphatidylinositol 3-kinase (PI3K)/AKT and mTOR signaling (10). However, GATA3 risk alleles have not yet been determined in overweight patients with CRLF2-ALL.
As germline risk variants in GATA3 co-segregate with specific somatic abnormalities in pre-B ALL (11), the aim of this study was to determine rs3824662 and rs3781093 as disease predisposition markers in Mexican patients. Here, we associated the GATA3 risk alleles with: a) susceptibility to pre-B ALL; b) the risk of developing pre-B ALL concomitantly with the overexpression of CRLF2; c) the nutritional status of pre-B ALL patients; and d) the event-free survival (EFS) and overall survival (OS) of patients.
2 Materials and methods
2.1 Patients and controls
A total of 130 patients aged <18 years who were diagnosed with pre-B ALL were included in the study (Supplementary Table 1). Patients were recruited at the time of diagnosis in the Oncology and Hematology Departments of the National Pediatrics Institute in Mexico City. The diagnosis was established using cytomorphology, immunophenotyping, and molecular biology for the most common gene fusions. Clinical and laboratory data were obtained from clinical records. The control group consisted of 130 unselected, healthy, unrelated adults with no family history of hematological malignancies. The patients and controls were Mexican mestizo residents in Mexico, with parents and grandparents born in Mexico. Patients, parents or legal tutors signed an informed consent form following the guidelines of the Declaration of Helsinki. The Institutional Research and Ethics Committee approved this study (project 076/2019; National Commission of Bioethics registration number: CONBIOETICA-09-CEI-025–20161215).
2.2 Biological samples
Saliva was obtained from pre-B ALL patients (Oragene DNA kit, DNA Genotek Inc. Ottawa, ON, Canada), and saliva or peripheral venous blood (EDTA-supplemented tubes) from the controls. Genomic DNA was extracted from saliva and blood samples (prepIT-L2P kit, DNA Genotek Inc. Ottawa, ON, Canada, and the QIAamp DNA Blood kit, QIAGEN, Hilden, Germany).
Bone marrow samples were collected from the patients with pre-B ALL at the time of diagnosis. CRLF2 expression (mRNA) was evaluated using TaqMan probes and GATA3 expression was detected in the same manner. Additionally, we determined P2RY8-CRLF2 rearrangement by RT-PCR, and when possible, samples with high CRLF2 expression and negative for P2RY8::CRLF2 were analyzed for IGH::CRLF2 rearrangement by fluorescence in situ hybridization (FISH).
2.3 Genotypification of rs3824662 and rs3781093 in GATA3
Both SNPs were genotyped by real-time PCR (StepOne Real-Time PCR, Applied Biosystems, Foster City, CA, U.S.) under standard conditions using predesigned TaqMan probes (VIC/FAM dye-labeled fluorescent probes; Applied Biosystems, Foster City, CA, U.S. ID C:27522049_10 and C:25809980_10, respectively). Each experiment included negative and positive controls for each genotype. Amplification was repeated randomly in 10% of the samples, and concordance was observed.
2.4 Analysis of CRLF2 and GATA3 expression
RNA was extracted from mononuclear cells in bone marrow samples using (RNeasy kit Qiagen, Düsseldorf, Germany) and cDNA was obtained using standard methods (Invitrogen, Waltham, MA, U.S.). The relative gene expressions of CRLF2, GATA3, and GUSβ (endogenous control) were determined in duplicate by real-time RT-PCR (LightCycler 2.0 Instrument; Roche Applied Science, Penzberg, Upper Bavaria, Germany) using TaqMan gene expression probes (Supplementary Table 2) from the Universal Probe Library System (Roche Applied Science, Penzberg, Upper Bavaria, Germany). CRLF2 overexpression was established according to the previously described criteria (12). It should be noted that the cutoff value was aligned with the IGH-CRLF2 positive patient with lower CRLF2 expression.
2.5 Detection of CRLF2 rearrangements
P2RY8::CRLF2 was assessed as previously described (13). IGH::CRLF2 was evaluated by FISH in interphase nuclei and metaphases using the dual-color break-apart probes LSI IGH (Abbott Molecular, Chicago, ILL, U.S.) and CRLF2 (CytoCell–OGT, Oxford, UK), following the manufacturer’s recommendations.
2.6 Nutritional status
Anthropometric assessment included height (cm) and weight (kg), which were measured using standard methods. Nutritional status was assessed using the following indicators from the WHO Anthro platform: weight/height (W/H), height/age (H/A), and body mass index (BMI). The classification was made according to the values established in the Official Mexican Standard NOM-008-SSA2-1993 (Control of Nutrition, Growth, and Development of Children and Adolescents). The cut-off points according to the Z score were: 1) Height for age (high +1.99 to +3; normal ≥ -1 to <+1; low -1 and lower). 2) Weight for height (obesity/overweight +1 to +3; normal ≥ -1 a <+1; malnutrition -1 and lower). 3) BMI for age (obesity/overweight ≥ +1; normal ≥ -1 a <+1; malnutrition ≥-1 and lower).
2.7 Statistical analysis
Both SNPs were analyzed for deviation from Hardy-Weinberg equilibrium (DeFinetti software (https://ihg.gsf.de/cgi-bin/hw/hwa2.pl), and the genotype and allelic frequencies were calculated for controls and patients. A two-tailed Fisher’s exact test was used to compare differences between groups (GraphPad Software, Inc. La Jolla, CA, U.S.). Odds ratios (OR) with 95% confidence intervals were calculated to estimate the risk of developing childhood pre-B ALL or pre-B ALL with higher CRLF2 expression in the presence of risk alleles and genotypes (DeFinetti Software). The association between each SNP and susceptibility to pre-B ALL was determined using p-values. CRLF2 and GATA3 expression levels were associated with different genotypes using the Kruskal–Wallis nonparametric test (IBM SPSS 29.0, Inc., Chicago, IL, USA). Nutritional status and GATA3 genotype associations were calculated using the chi-squared test. EFS and OS were calculated for patients with different genotypes using the Kaplan–Meier method and Cox regression analysis for hazard ratios (IBM SPSS 29.0). For all comparisons, statistical significance was set at p ≤ 0.05.
3 Results
Clinical and laboratory data such as gender, age, white blood cell count, leukemic infiltration site, and presence of gene fusion of the 130 patients included in this study are presented in Supplementary Table 1.
3.1 Germline variants in GATA3 and pre-B ALL susceptibility
A total of 130 patients and 130 controls were studied for pre-B ALL susceptibility. For rs3824662 and rs3781093, the genotypic frequencies of the risk homozygotes AA and CC were higher in the patients than in the controls (38.5% vs. 16.2% and 38% vs. 16.9%, p ≤ 0.0001 and p=0.0002, respectively). Similar results were observed for the frequencies of the two risk alleles A and C (0.46 vs. 0.62, p=0.0003 and 0.46 vs. 0.63, p=0.0002, respectively). In contrast, the genotypic frequencies of the non-risk homozygotes CC and TT were higher in controls than in patients (24.6% vs. 14.6%, p=0.0602 and 24.6% vs. 12.4%, p=0.0159). Similar results were obtained for the frequencies of non-risk alleles C and T (0.54 vs 0.38 and 0.54 vs 0.37, respectively) (Table 1). OR analysis revealed that risk alleles A and C confer susceptibility to the development of childhood pre-B ALL in our population (OR=1.92, p=0.0002 and OR=1.96, p=0.0002, respectively), and the risk was increased in risk homozygotes (AA OR=4.01, p=0.0004 and CC OR=4.45, p=0.0002, respectively) (Table 1).

Table 1. Genotypic and allelic frequencies of rs3824662 and rs3781093 in GATA3.
3.2 Germline variants in GATA3 and pre-B ALL with CRLF2 overexpression
Sixty-three patients without CRLF2 overexpression (No CRLF2-OE) and 51/52 patients with CRLF2 overexpression (CRLF2-OE) were analyzed (Table 1). Both risk homozygotes AA and CC were associated with pre-B ALL with CRLF2-OE (53.8% vs. 28.6%, p=0.0075 and 51% vs. 30.1%, p=0.0337, respectively). Risk alleles A and C were also more frequent in CRLF2-OE patients (Table 1). In comparison, in patients without overexpression (No CRLF2-OE), there was a trend towards a higher frequency of the non-risk alleles (C and T) and the non-risk homozygote genotypes (CC and TT) (Table 1). OR analysis revealed an association between the risk alleles of rs3824662 and rs3781093, and a predisposition to develop pre-B ALL with CRLF2-OE (Table 1). The genotypic and allelic frequencies of risk homozygotes (AA and CC) and risk alleles (A and C) for both SNPs were higher in patients with CRLF2-OE (53.8% and 51%/0.72 and 0.72, respectively), followed by the total pre-B ALL patients (38.5% and 38.0%/0.62 and 0.63) and the group of patients without CRLF2-OE (28.6% and 30.1%/0.54 and 0.56).
3.3 rs3824662 and rs3781093 genotypes and CRLF2 and GATA3 expression
Based on the availability of patient samples, it was possible to analyze 115 of 130 patients for CRLF2 expression and genotype of rs3824662. The highest levels of CRLF2 expression were observed in patients with the AA genotype for rs3824662 (p=0.004) compared to patients with AC and CC genotypes (Figure 1A). For rs3781093 and CRLF2 expression 114 patients were successfully studied. Similarly, patients with higher CRLF2 expression presented with the CC genotype (p=0.021) compared to patients with the TC and TT genotypes (Figure 1B). GATA3 expression was analyzed in 110 patients for rs3824662 and 108 patients for rs3781093; the expression levels detected among genotypes for both SNPs were heterogeneous. No associations were found between the genotypes of either SNP or GATA3 expression (Figures 1C, D), since expression levels of the three genotypes were heterogeneous. In one patient heterozygous for both SNPs, overexpression of GATA3 was ten orders of magnitude higher than in the other patients. The biological cause of this overexpression has not been investigated at this time, this sample did not show an increase in CRLF2 expression.
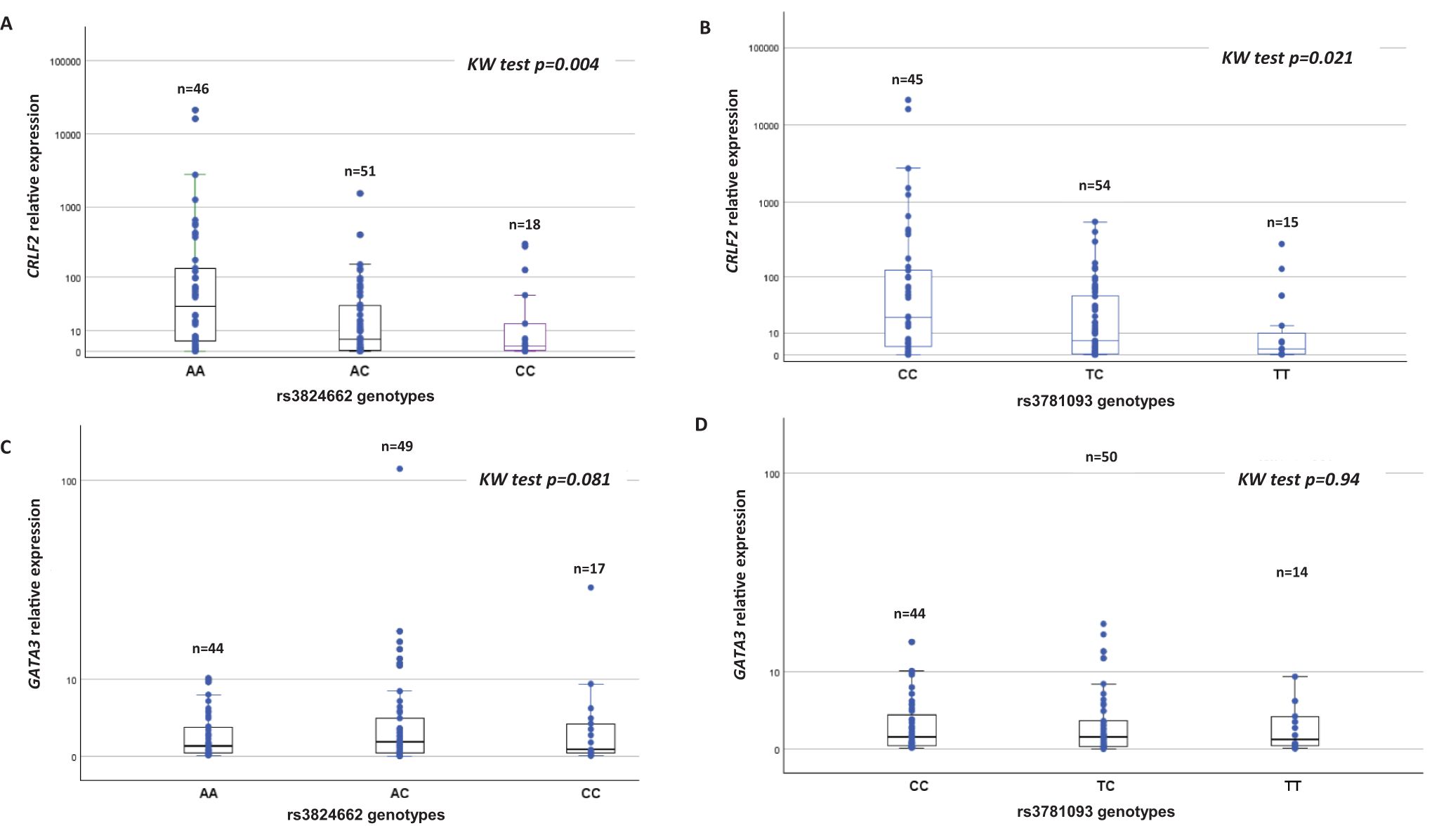
Figure 1. Association of rs3824662 and rs3781093 genotypes with CRLF2 and GATA3 expression.
3.4 rs3824662 and rs3781093 genotypes and nutritional status
We determined the nutritional status at diagnosis of 66 patients genotyped for rs3824662 and 64 for rs3781093 (Table 2). The most frequent nutritional status was normal (41 for both SNPs), followed by malnutrition, 16 and 15, and obese/overweight, 9 and 8 for rs3824662 and rs3781093 respectively. The proportion of risk homozygous genotypes AA and CC (Table 2) was higher in obese/overweight patients than in those with malnutrition and adequate nutritional status (p=0.004 and p=0.011, respectively).
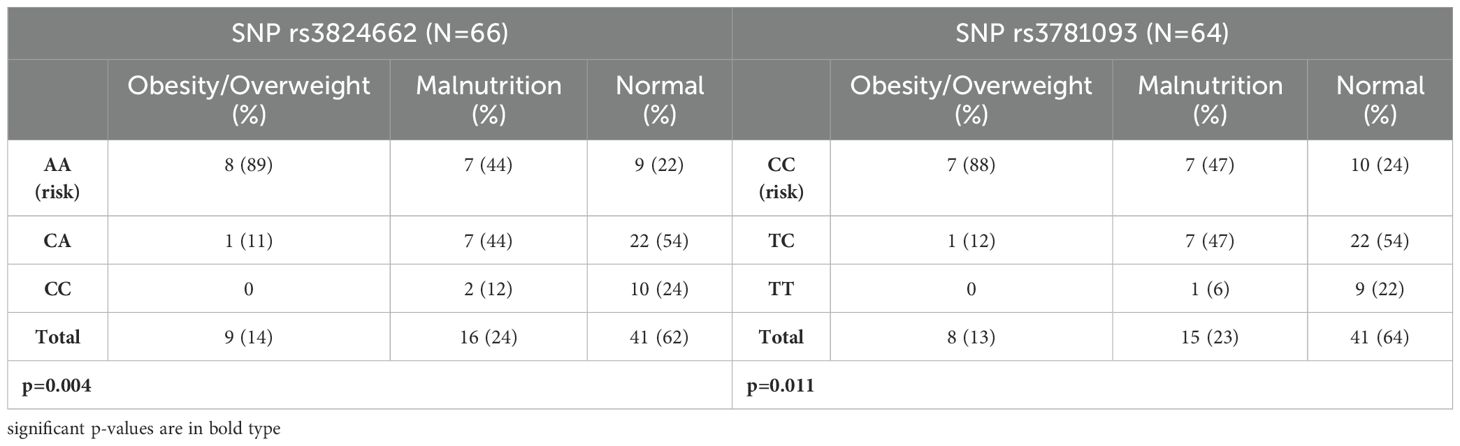
Table 2. Association of rs3824662 and rs3781093 in GATA3 with nutritional status in Pre-B ALL patients.
3.5 Survival analysis
For survival analysis, patients who discontinued chemotherapy before relapse and patients with a follow-up of <60 months were excluded. The treatment protocols for the evaluated patients were based on the Pediatric Medical Insurance Program (14). Based on this, 70 patients were included in the survival analysis. The risk homozygote genotypes AA and CC were not associated with increased EFS (Supplementary Figure 1); however, a positive association (Figure 2) with OS was observed for rs3824662 and rs3781093 (p=0.023 and p=0.022, respectively). We looked for associations between patients with risk genotypes and rearrangements in CRLF2 and survival, but did not observe any difference.
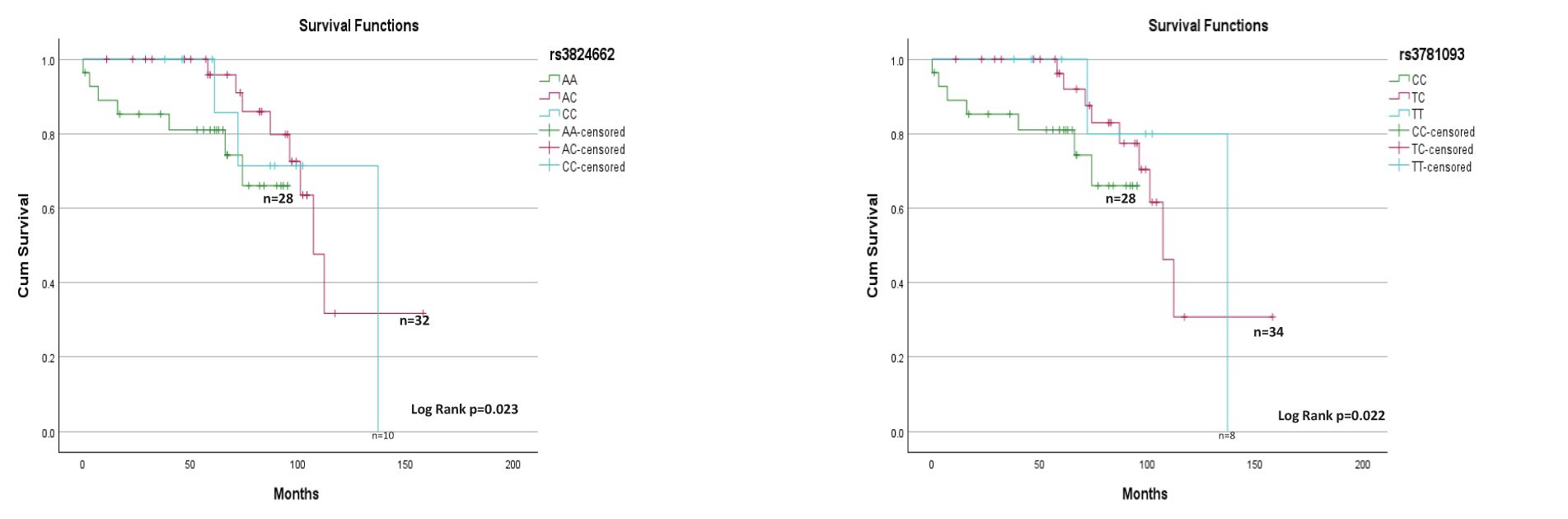
Figure 2. rs3824662 and rs3781093 genotypes and overall survival (n=70).
4 Discussion
4.1 Genotypes and alleles
Regarding the risk alleles A and C of both SNPs in GATA3, the frequencies in control populations varied widely throughout the Americas. For rs3824662, the higher frequency was observed in Guatemalans, followed by Peruvians, Colombians, Mexican Americans and Hispanic Americans (0.52, 0.45, 0.38, 0.38 and 0.33, respectively) (6, 15). In populations such as Puerto Ricans, Brazilians, and European Americans, the frequencies were lower (0.26, 0.21, and 0.17, respectively) (Figure 3A) (6, 15, 16). The frequency of the A allele in Mexican mestizos from this study (0.46) was close to those observed in Guatemalans and Peruvians, therefore it can be considered among the higher frequencies documented (Figure 3A). For rs3781093, the C allele frequencies in controls were higher in Peruvians, Mexican Americans, Colombians, and Hispanics (0.45, 0.38, 0.33, and 0.33, respectively), compared to Puerto Ricans (0.25), Brazilians (0.22), and European Americans (0.14) (6, 15, 16), but none of these were higher than that found in Mexican mestizos (0.46) (Figure 3B). It is possible that Brazilian controls have a lower frequency of risk alleles because the SNPs studied are poorly associated with African ancestry, which is predominant in this population (6, 17, 18).
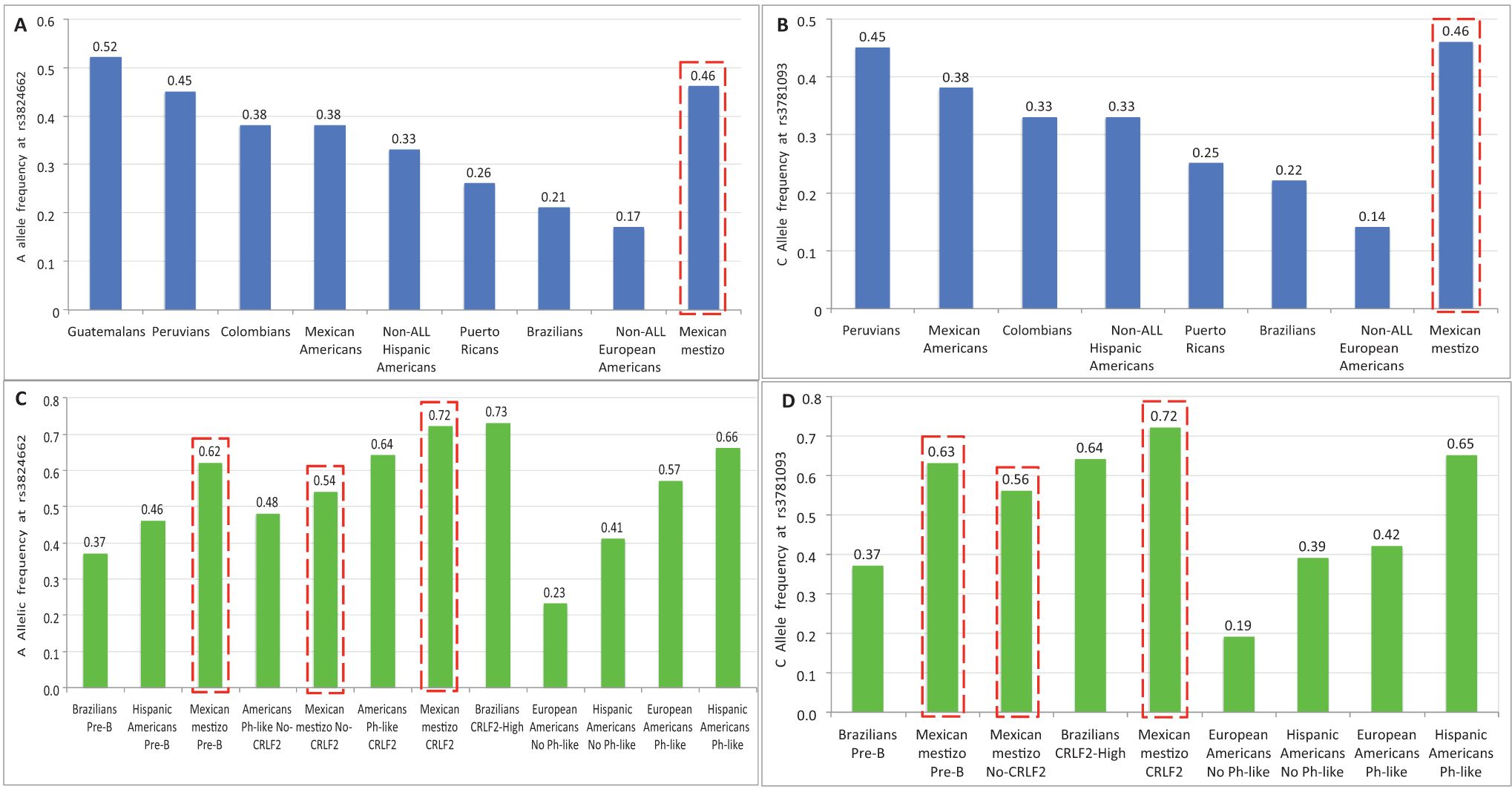
Figure 3. Allele frequencies of the risk alleles at rs3824662 and rs3781093 in different American populations. (A, B) correspond to controls and (C, D) to pre-B ALL children. Data from Colombians, Peruvians, Mexican Americans and Puerto Ricans were obtained from the 1000 Genomes Project (15). Data from Brazilians, Guatemalans, European Americans, Hispanic Americans, American controls and ALL children were obtained from Perez-Andreu, et. al., and (6) Migita, et. al., (16). ( ) Data from this study.
) Data from this study.
In childhood patients with pre-B ALL, few studies have reported the frequencies of both alleles in Latin Americans, therefore comparisons between populations are difficult. The ascending order of frequency for the A allele at rs3824662 was as follows: Brazilians (0.37), Hispanics (0.46), and Mexican mestizos (0.62) (6, 16) (Figure 3C). As expected, the frequency of the C allele in rs3781093 was similar to that found for rs3824662 (16) (Figure 3D). Considering the frequencies reported in different childhood pre-B ALL subtypes, the A allele was more frequent in Brazilians with CRLF2-high (0.73) than in CRLF2-Mexican mestizos (0.72), Ph-like CRLF2-Americans (0.64), No-CRLF2-Mexican mestizos (0.54), and Ph-like No-CRLF2-Americans (0.48) (6, 16) (Figure 3C). For rs3781093, the C allele frequency in patients was lower in No-CRLF2 Mexican mestizos (0.56) than in Brazilians with CRLF2-High and CRLF2-Mexican mestizos (0.64 and 0.72) (16) (Figure 3D). These results show that the risk alleles in rs3824662 and rs3781093 are overrepresented in our population and are associated with the risk of developing pre-B ALL and CRLF2-ALL. This suggests that the Amerindian component of Mexicans may be important for the high frequency of this subtype of leukemia in our population.
4.2 Survival and genotypes
A positive association was observed only with OS for both the SNPs, this result may be influenced by the sample size, as patients who temporarily discontinued treatment and those with less than 60 months of follow-up were excluded from the analysis. In our setting, it is of utmost importance to implement follow-up measures for patients at risk of discontinuation and nonadherence (19).
4.3 Expression assays
To our knowledge, the CRLF2 OE has only been reported in AA variant carriers (20); in our cohort, the association with OE was found not only for AA but also for CC. It has been observed that rs3824662 upregulates GATA3 transcription, which alters chromatin accessibility, indicating that GATA3 potentiates CRLF2 expression (11). Regarding GATA3, no increased expression was found in patients who were homozygous for the risk alleles of the two SNPs analyzed. In contrast to previous studies performed in the HapMap cell lines from different populations and in patient lymphoblasts from the Children’s Oncology Group cohorts (6), in this study, we observed a wide variability in GATA3 expression among the three genotypes for each SNP. These results are attributed to the different cellular physiological conditions, to undetected genetic abnormalities in GATA3, or to the transcriptional or epigenetic regulation present in the leukemic blasts of each patient. In addition, it is important to note that the increased enhancer activity was only reported for the rs3824662 risk allele, while the rs3781093 allele did not appear to have the same effect (21).
4.4 Nutritional status
The high rate of obesity in pre-B ALL Hispanic patients living in the U.S. has been considered a predisposing factor for the occurrence of CRLF2-ALL (10). It has been suggested that these characteristics may be related to the presence of the risk allele at rs3824662, which may disrupt adipogenesis, metabolism, and/or signaling pathways that contribute to the development of CRLF2 pre-B ALL (10). However, the authors did not report SNP genotyping data.
In this study, the risk homozygotes of both SNPs (AA and CC) were associated with the overweight status of the patients. To our knowledge, this is the first time that germline variations in GATA3 have been associated with the nutritional status of the Mexican pre-B ALL patients. Our results are consistent with those observed in Hispanic patients, but they must be considered with caution because: a) the method of determining nutritional status in the previous study was more complete than that used in our patients (fat mass and body fat percentage measured by whole-body dual-energy X-ray absorptiometry vs. anthropometric assessment including height, weight, and BMI); b) the number of patients analyzed for nutritional status in both studies was low; and c) the nutritional characteristics of the two populations studied may be different. The U.S. Hispanic patients have a high prevalence of obesity, whereas Mexican patients who attend our institution are overweight (20%) and malnourished (22%) (22). As expected, this study recruited a higher proportion of malnourished patients, mainly homozygotes or heterozygotes, for the risk alleles. In this context, we suggest that although the association of overweight/obesity with the analyzed risk homozygous genotypes is clear, it should not be excluded that the genotype of GATA3 variants may partly influence metabolic alterations leading to abnormal nutritional status. It has been noted that there is a link between nutritional changes and GATA3 protein, as it can alter adipogenesis and lead to insulin resistance, and inhibition of GATA3 has been shown to modify impaired adipogenesis and contribute to restoring healthy fat distribution (23). The contribution of nutritional status to ALL development through GATA3 requires further investigation.
5 Conclusion
This is the first study to investigate the association between GATA3 SNPs and predisposition to childhood pre-B ALL and CRLF2-ALL in Mexican patients. This confirms the high frequency predicted for the risk alleles in our population and shows that not only the SNP rs3824662, but also rs3781093 have a high penetrance and are effective markers of predisposition for the development of CRLF2-ALL, which is common in our patients. It also shows, for the first time, that being overweight, estimated by BMI at the time of patient diagnosis, is associated with the presence of the risk alleles of both polymorphisms. The limitations of this study include the need to refine measures of body mass and fat, to have cohorts of patients with longer follow-up to obtain more reliable survival calculations, and to perform transcriptome sequencing that will allow us to know the alterations associated with ALL Ph-like and the subgroup of patients with an aberrant Jak-Stat pathway. This will enable us to establish more specific associations between groups, allowing us to better understand our population and obtain data that can be extrapolated to populations of Amerindian ancestry.
Data availability statement
The original contributions presented in the study are publicly available. This data can be found here: https://www.ncbi.nlm.nih.gov/clinvar/; SCV006060650 - SCV006060651.
Ethics statement
The studies involving humans were approved by Institutional Research and Ethics Committee. National Commission of Bioethics registration number: CONBIOETICA-09-CEI-025–20161215). The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent for participation in this study was provided by the participants’ legal guardians/next of kin.
Author contributions
AR: Conceptualization, Formal analysis, Investigation, Methodology, Writing – original draft, Writing – review & editing, Data curation. EC: Formal analysis, Methodology, Writing – review & editing. MJ: Conceptualization, Formal analysis, Investigation, Methodology, Software, Validation, Writing – review & editing. DM: Formal analysis, Investigation, Methodology, Validation, Writing – review & editing. CS: Formal analysis, Software, Writing – review & editing. DM: Conceptualization, Investigation, Writing – review & editing. CG: Methodology, Supervision, Writing – review & editing. NL: Writing – review & editing, Methodology, Supervision. EG: Methodology, Writing – review & editing. AG: Methodology, Writing – review & editing. IM: Methodology, Writing – review & editing. PP: Conceptualization, Formal analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Supervision, Visualization, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by Convocatoria FPIS2024-INP-5345 (PP-V) and Fondos del Presupuesto Federal del Instituto Nacional de Pediatría (076/2019).
Acknowledgments
The authors thank the patients and their families.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fonc.2025.1533756/full#supplementary-material
References
1. Migliorini G, Fiege B, Hosking FJ, Ma Y, Kumar R, Sherborne AL, et al. Variation at 10p12.2 and 10p14 influences risk of childhood B-cell acute lymphoblastic leukemia and phenotype. Blood. (2013) 122:3298–307. doi: 10.1182/blood-2013-03-491316
2. Walsh KM, De Smith AJ, Welch TC, Smirnov I, Cunningham MJ, Ma X, et al. Genomic ancestry and somatic alterations correlate with age at diagnosis in Hispanic children with B-cell acute lymphoblastic leukemia. Am J Hematol. (2014) 89:721–5. doi: 10.1002/ajh.23727
3. Zhang H, Liu APY, Devidas M, Lee SH, Cao X, Pei D, et al. Association of GATA3 polymorphisms with minimal residual disease and relapse risk in childhood acute lymphoblastic leukemia. J Natl Cancer Inst. (2021) 113:408–17. doi: 10.1093/jnci/djaa138
4. Koller PB, Zhang H, Kantarjian H, Jabbour E, Pierce S, Gocho Y, et al. GATA3 rs3824662A allele in B-cell acute lymphoblastic leukemia in adults, adolescents and young adults: association with CRLF2 rearrangement and poor prognosis. Am J Hematol. (2021) 96:E71–4. doi: 10.1002/ajh.26065
5. Hou Q, Liao F, Zhang S, Zhang D, Zhang Y, Zhou X, et al. Regulatory network of GATA3 in pediatric acute lymphoblastic leukemia. Oncotarget. (2017) 8:36040–53. doi: 10.18632/oncotarget.16424
6. Perez-Andreu V, Roberts KG, Harvey RC, Yang W, Cheng C, Pei D, et al. Inherited GATA3 variants are associated with Ph-like childhood acute lymphoblastic leukemia and risk of relapse. Nat Genet. (2013) 45:1494–8. doi: 10.1038/ng.2803
7. Harvey RC, Mullighan CG, Chen IM, Wharton W, Mikhail FM, Carroll AJ, et al. Rearrangement of CRLF2 is associated with mutation of JAK kinases, alteration of IKZF1, Hispanic/Latino ethnicity, and a poor outcome in pediatric B-progenitor acute lymphoblastic leukemia. Blood. (2010) 115:5312–21. doi: 10.1182/blood-2009-09-245944
8. Raca G, Abdel-Azim H, Yue F, Broach J, Payne JL, Reeves ME, et al. Increased Incidence of IKZF1 deletions and IGH-CRLF2 translocations in B-ALL of Hispanic/Latino children-a novel health disparity. Leukemia. (2021) 35:2399–402. doi: 10.1038/s41375-021-01133-4
9. Martínez-Anaya D, Moreno-Lorenzana D, Reyes-León A, Juárez-Figueroa U, Dean M, Aguilar-Hernández MM, et al. Characterization of philadelphia-like pre-B acute lymphoblastic leukemia: experiences in Mexican pediatric patients. Int J Mol Sci. (2022) 23:9587. doi: 10.3390/ijms23179587
10. Mittelman SD, Kim J, Raca G, Li G, Oberley MJ, Orgel E. Increased prevalence of CRLF2 rearrangements in obesity-associated acute lymphoblastic leukemia. Blood. (2021) 138:199–202. doi: 10.1182/blood.2021011106
11. Yang H, Zhang H, Luan Y, Liu T, Yang W, Roberts KG, et al. Noncoding genetic variation in GATA3 increases acute lymphoblastic leukemia risk through local and global changes in chromatin conformation. Nat Genet. (2022) 54:170–9. doi: 10.1038/s41588-021-00993-x
12. Juárez-Velázquez MDR, Moreno-Lorenzana DL, Martínez Anaya DA, Hernández Monterde EA, Aguilar-Hernández MM, Reyes-León A, et al. High occurrence of CRLF2 abnormalities in Mexican children with B-cell acute lymphoblastic leukemia. Cytokine. (2022) 155:155896. doi: 10.1016/j.cyto.2022.155896
13. Palmi C, Vendramini E, Silvestri D, Longinotti G, Frison D, Cario G, et al. Poor prognosis for P2RY8-CRLF2 fusion but not for CRLF2 over-expression in children with intermediate risk B-cell precursor acute lymphoblastic leukemia. Leukemia. (2012) 26:2245–53. doi: 10.1038/leu.2012.101
14. Paredes-Aguilera R, Rivera-Luna R, Martínez-Ávalos A. Leucemia aguda linfoblástica. In: Rivera-Luna R, editor. Protocolos Técnicos Cáncer en Niños. Editores de Textos Mexicanos, México City, México (2010). p. 9–60.
15. Dyer SC, Austine-Orimoloye O, Azov AG, Barba M, Barnes I, Barrera-Enriquez VP, et al. Ensembl 2025. Nucleic Acids Res. (2025) 53:D948–57. doi: 10.1093/nar/gkae1071
16. Migita NA, Jotta PY, do Nascimento NP, Vasconcelos VS, Centoducatte GL, Massirer KB, et al. Classification and genetics of pediatric B-other acute lymphoblastic leukemia by targeted RNA sequencing. Blood Adv. (2023) 7:2957–71. doi: 10.1182/bloodadvances.2022009179
17. Pena SDJ, Santos FR, Tarazona-Santos E. Genetic admixture in Brazil. Am J Med Genet C Semin Med Genet. (2020) 184:928–38. doi: 10.1002/ajmg.c.v184.4
18. Perez-Andreu V, Roberts KG, Xu H, Smith C, Zhang H, Yang W, et al. A genome-wide association study of susceptibility to acute lymphoblastic leukemia in adolescents and young adults. Blood. (2015) 125:680–6. doi: 10.1182/blood-2014-09-595744
19. Rivera-Luna R, Perez-Vera P, Galvan-Diaz C, Velasco-Hidalgo L, Olaya-Vargas A, Cardenas-Cardos R, et al. Triple-hit explanation for the worse prognosis of pediatric acute lymphoblastic leukemia among Mexican and Hispanic children. Front Oncol. (2022) 12:1072811. doi: 10.3389/fonc.2022.1072811
20. Madzio J, Pastorczak A, Sedek L, Braun M, Taha J, Wypyszczak K, et al. GATA3 germline variant is associated with CRLF2 expression and predicts outcome in pediatric B-cell precursor acute lymphoblastic leukemia. Genes Chromosomes Cancer. (2019) 58:619–26. doi: 10.1002/gcc.22748
21. Li C, Liang W, He Y, Zhao X, Qian J, Li Z, et al. Inherited GATA3 variant associated with positive minimal residual disease in childhood B-cell acute lymphoblastic leukemia via asparaginase resistance. Clin Transl Med. (2021) 11:e507. doi: 10.1002/ctm2.v11.8
22. García-Guzmán AD. Asociación y valor predictivo del ángulo de fase con la presencia de desnutrición en pacientes pediátricos hospitalizados con diagnóstico oncológico. México City (México: Escuela de Dietética y Nutrición (2023).
Keywords: pre-B acute lymphoblastic leukemia, Mexicans, children, overweight, obesity, CRLF2 overexpression, SNPs in GATA3
Citation: Reyes-León A, Castro-Vargas E, Juárez-Velázquez MdR, Martínez Anaya D, Salas-Labadía C, Moreno-Lorenzana D, Galván-Díaz CA, López-Santiago N, García-Padilla E, García-Guzmán AD, Medina-Vera I and Pérez-Vera P (2025) GATA3 germline variants in childhood pre-B acute lymphoblastic leukemia: association with CRLF2 overexpression and overweight in Mexican patients. Front. Oncol. 15:1533756. doi: 10.3389/fonc.2025.1533756
Received: 24 November 2024; Accepted: 10 March 2025;
Published: 12 May 2025.
Edited by:
Sharon R. Pine, University of Colorado Anschutz Medical Campus, United StatesReviewed by:
Azza Mahmoud Kamel, Cairo University, EgyptAlexandra Kovach, Children’s Hospital of Los Angeles, United States
Copyright © 2025 Reyes-León, Castro-Vargas, Juárez-Velázquez, Martínez Anaya, Salas-Labadía, Moreno-Lorenzana, Galván-Díaz, López-Santiago, García-Padilla, García-Guzmán, Medina-Vera and Pérez-Vera. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Patricia Pérez-Vera, cHBlcmV6dmVyYUB5YWhvby5jb20=