 Lichang Yang1
Lichang Yang1 Manman Xu
Manman Xu- 1Xuzhou Affiliated Hospital of Nanjing University of Chinese Medicine, Xuzhou, China
- 2Department of Geriatrics, Guang' anmen Hospital, China Academy of Chinese Medical Sciences, Beijing, China
Immunosenescence, the age-associated decline of the immune system, is pivotal in fostering drug resistance within the tumor microenvironment (TME). The accumulation of senescent immune cells and the release of pro-inflammatory senescence-associated secretory phenotype (SASP) factors create a milieu that supports tumor survival and undermines therapeutic efficacy. Traditional cancer treatments often fail to address this underlying issue, leading to suboptimal outcomes. This article proposes an innovative strategy to overcome immunosenescence-induced drug resistance through the nanoparticle-mediated delivery of herbal-derived natural products (HDNPs), which possess senolytic and immunomodulatory properties capable of clearing senescent cells and rejuvenating immune function. Nanoparticle delivery systems enhance these compounds’ stability, bioavailability, and targeted delivery to the TME and senescent immune cells. By harnessing the synergistic effects of HDNPs and nanotechnology, this approach offers a novel and multifaceted solution to drug resistance in cancer therapy. It holds the potential to restore immune surveillance, reduce pro-survival signaling in cancer cells, and enhance the efficacy of conventional treatments. This paradigm shift emphasizes the importance of addressing immunosenescence as a therapeutic target and paves the way for more effective and personalized cancer interventions.
1 Introduction
Cancer remains a leading cause of mortality globally, with millions of new cases diagnosed each year. Despite significant advancements in oncology, the efficacy of cancer treatments is often hindered by the development of drug resistance, which is a major obstacle to successful therapy (1). Drug resistance can be intrinsic or acquired and involves complex mechanisms that allow cancer cells to evade the cytotoxic effects of chemotherapeutic agents (2). Key mechanisms of drug resistance include increased drug efflux via overexpression of transporter proteins, alterations in drug targets, activation of DNA repair pathways, evasion of apoptosis, and metabolic reprogramming (3). The tumor microenvironment (TME) also plays a crucial role by providing a protective niche that supports cancer cell survival and contributes to resistance (4). The heterogeneity of tumors adds another layer of complexity, as different cell populations within the same tumor may respond variably to treatment (5). This intratumoral diversity facilitates the selection of resistant clones under therapeutic pressure, leading to treatment failure and disease progression. Emerging evidence suggests that immunosenescence—the gradual deterioration of the immune system associated with aging—significantly contributes to cancer progression and drug resistance (6). Immunosenescence affects innate and adaptive immunity, leading to reduced immunosurveillance and an increased incidence of malignancies in older people (7). Within the TME, immunosenescent cells exhibit altered functionality, including decreased proliferation, impaired cytokine production, and reduced cytotoxic activity. These changes can create an immunosuppressive environment that fosters tumor growth and diminishes the efficacy of chemotherapy and immunotherapy. Moreover, senescent immune cells can secrete various pro-inflammatory cytokines, chemokines, and proteases collectively known as the senescence-associated secretory phenotype (SASP) (8). The SASP can promote tumor cell proliferation, angiogenesis, and metastasis while inhibiting apoptosis, thereby contributing to drug resistance. Understanding the interplay between immunosenescence and drug resistance is crucial for developing novel therapeutic strategies that can enhance treatment efficacy, particularly in aging populations where cancer incidence is highest (9).
Immunosenescence is characterized by accumulated aged immune cells that have reached replicative senescence due to telomere shortening or have become senescent in response to stress and DNA damage (10). These cells display altered surface markers, such as increased expression of inhibitory receptors (e.g., PD-1, CTLA-4) and decreased levels of costimulatory molecules. Functionally, they exhibit diminished proliferative capacity, reduced cytokine production, and impaired ability to eliminate tumor cells (11). In the TME, immunosenescent cells contribute to drug resistance through several mechanisms. Firstly, senescent immune cells secrete SASP factors that promote tumor cell survival and proliferation. Interleukin-6 (IL-6) and interleukin-8 (IL-8) can activate survival pathways in cancer cells, leading to resistance against apoptosis induced by chemotherapy (12). Secondly, the reduced cytotoxicity of senescent T cells and natural killer (NK) cells diminishes the immune system’s ability to recognize and eliminate cancer cells, allowing resistant clones to expand (13). Thirdly, immunosenescence is associated with an increase in regulatory T cells (Tregs) and myeloid-derived suppressor cells (MDSCs), which further suppress anti-tumor immune responses and contribute to a permissive environment for tumor growth (14). In addition, senescent immune cells can influence other components of the TME, such as fibroblasts and endothelial cells, enhancing angiogenesis and extracellular matrix remodeling that facilitate tumor progression and metastasis (15). These factors collectively create a TME that supports cancer cell survival and proliferation while undermining the effectiveness of therapeutic agents.
Herbal-derived natural products (HDNPs) have garnered significant interest due to their diverse bioactive compounds with immunomodulatory, anti-inflammatory, and anti-cancer properties. Compounds such as quercetin, fisetin, and epigallocatechin gallate (EGCG) have demonstrated the ability to modulate immune cell function, reduce SASP factor secretion, and induce apoptosis in senescent cells (16). These HDNPs can target multiple signaling pathways involved in immunosenescence and drug resistance. Quercetin has been shown to inhibit the NF-κB pathway, reducing the production of pro-inflammatory cytokines (17). Fisetin can activate the Nrf2 pathway, enhancing the antioxidant capacity of cells and reducing oxidative stress (18). EGCG has been reported to modulate epigenetic modifications, restoring the function of aging immune cells (19). Despite their therapeutic potential, HDNPs often face challenges such as poor solubility, low bioavailability, and rapid metabolism, which limit their clinical application. Nanoparticle-based delivery systems offer a promising solution to these issues by enhancing the stability and bioavailability of HDNPs (20). Nanoparticles can be engineered to deliver HDNPs specifically to the TME and senescent immune cells, minimizing systemic toxicity and improving therapeutic efficacy. Features such as controlled release, protection from degradation, and surface modification with targeting ligands enable precise delivery and sustained therapeutic action (21). Moreover, nanoparticles can cross biological barriers and accumulate in tumors via the enhanced permeability and retention (EPR) effect, further increasing the concentration of HDNPs at the desired site (21). This targeted approach enhances the anti-cancer effects of HDNPs and allows for the modulation of immunosenescence within the TME, potentially reversing drug resistance mechanisms.
This article explores the innovative approach of utilizing nanoparticles to deliver HDNPs that modulate immunosenescence-induced drug resistance in cancer therapy. By integrating insights from oncology, immunology, and nanotechnology, we seek to comprehensively analyze how nanoparticle-mediated HDNP delivery can rejuvenate aging immune cells, suppress SASP factors, and enhance anti-tumor immunity. This strategy holds promise for overcoming drug resistance and improving therapeutic outcomes in cancer patients, particularly the older.
2 Immunosenescence-induced drug resistance in cancer
Immunosenescence refers to the gradual deterioration of the immune system associated with aging, leading to an increased susceptibility to infections, diseases, and cancer progression (22). Within the TME, immunosenescence is characterized by the accumulation of senescent immune cells, including T cells, NK cells, and dendritic cells (DCs). These senescent cells exhibit altered phenotypes and diminished functional capacities, such as reduced proliferative ability and impaired cytokine production. The accumulation of senescent immune cells is driven by persistent antigen exposure, telomere shortening, and oxidative stress (23). Repetitive stimulation of T cells by tumor antigens can induce a senescent state, which in turn upregulates the expression of inhibitory receptors like PD-1 and TIM-3, leading to T cell exhaustion (24). This immunosenescent profile contributes to a diminished immune response against tumor cells. Furthermore, senescent immune cells secrete various pro-inflammatory cytokines, chemokines, growth factors, and proteases collectively known as SASP (25). The SASP can have paradoxical effects within the TME. While initially intended to recruit immune cells for clearance of senescent cells, the chronic presence of SASP factors can promote tumorigenesis (25). SASP components such as IL-6, IL-8, and vascular endothelial growth factor (VEGF) enhance tumor cell proliferation, angiogenesis, and metastasis (25). Additionally, matrix metalloproteinases (MMPs) secreted by senescent cells degrade the extracellular matrix, facilitating cancer cell invasion (26). The SASP thus creates a pro-tumorigenic environment that supports cancer progression. The altered cytokine milieu from immunosenescence fosters an environment conducive to drug resistance. Pro-inflammatory cytokines like IL-6 and tumor necrosis factor-alpha (TNF-α) activate survival signaling pathways in cancer cells, such as the STAT3 and NF-κB pathways (27). Activation of these pathways upregulates anti-apoptotic proteins (Bcl-2, Bcl-xL) and drug efflux transporters (P-glycoprotein), reducing the efficacy of chemotherapeutic agents (28). Furthermore, SASP factors can induce epithelial-to-mesenchymal transition (EMT) in cancer cells, associated with increased resistance to chemotherapy and targeted therapies (29). EMT endows cancer cells with stem cell-like properties, enhancing their survival ability in the presence of anticancer drugs.
On the other hand, immunosenescence impairs the immune system’s ability to recognize and eliminate cancer cells, contributing to immune evasion and therapeutic resistance (30). Senescent T cells exhibit decreased cytotoxic activity and reduced interferon-gamma (IFN-γ) production, a critical cytokine for anti-tumor immunity. Similarly, senescent NK cells show diminished expression of activating receptors and impaired degranulation capacity (30). This compromised immune surveillance allows tumor cells to proliferate unchecked and develop mechanisms to resist therapy. Reduced immune pressure can enable the expansion of cancer cell clones with mutations conferring drug resistance. Additionally, the immunosuppressive environment can inhibit the effectiveness of immunotherapies designed to reactivate immune responses against tumors (31). Recent research has highlighted the direct involvement of immunosenescent cells in promoting drug resistance. A study demonstrated that senescent CD8+ T cells in the TME secrete IL-10, suppressing the effector T cells’ function and contributing to resistance against anti-PD-1 therapy in melanoma models (32). The depletion of senescent T cells restored responsiveness to immunotherapy, indicating their role in therapeutic failure. Another study by Di Mitri et al. showed that senescent stromal cells in prostate cancer release SASP factors that activate the IL-6/STAT3 pathway in tumor cells, leading to resistance to chemotherapy (33). Blocking IL-6 signaling sensitized cancer cells to treatment, further establishing the link between immunosenescence and drug resistance.
Clinical data also support the association between immunosenescence and poor treatment outcomes in cancer patients. Older patients often exhibit reduced responses to chemotherapy and immunotherapy compared to younger individuals (34). This disparity is partly attributed to age-related declines in immune function, including decreased T cell diversity and function. A retrospective analysis by Elias et al. found that older patients with non-small cell lung cancer had lower overall survival rates following immunotherapy, correlating with markers of immunosenescence (35). Similarly, studies have shown that high levels of senescent immune cells in peripheral blood are associated with decreased progression-free survival in patients undergoing targeted therapies (36). These findings underscore the impact of immunosenescence on therapeutic efficacy and highlight the need for interventions that address immune aging to overcome drug resistance (Figure 1).
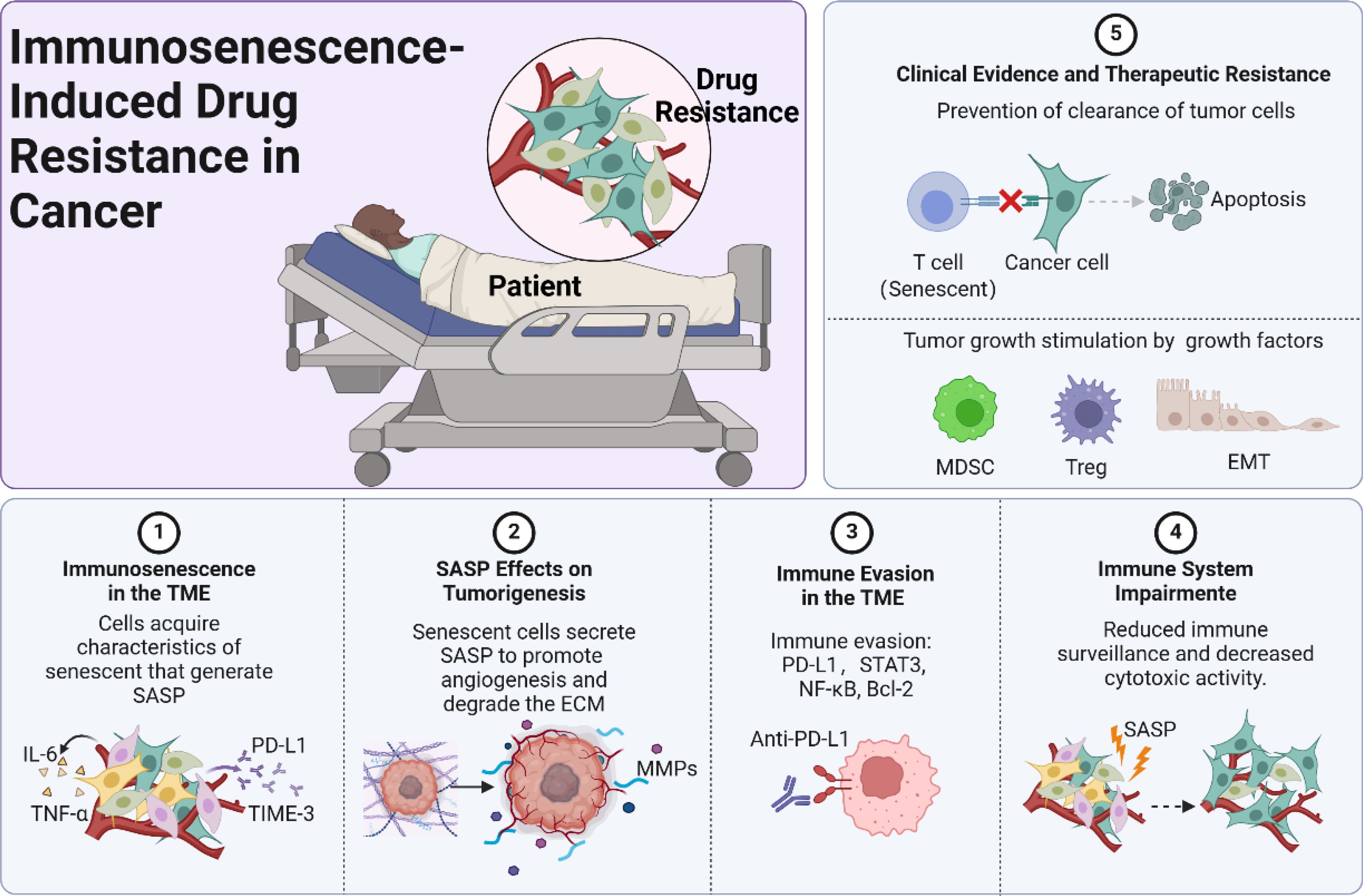
Figure 1. Immunosenescence-induced drug resistance in cancer.
3 Herbal-derived natural products targeting immunosenescence
3.1 HDNPs with anti-immunosenescence properties
HDNPs possess significant potential to mitigate immunosenescence and reverse drug resistance in cancer therapy. Compounds such as quercetin, fisetin, EGCG, resveratrol, curcumin, and others have demonstrated unique capabilities to modulate immune aging mechanisms within the TME (Figure 2).
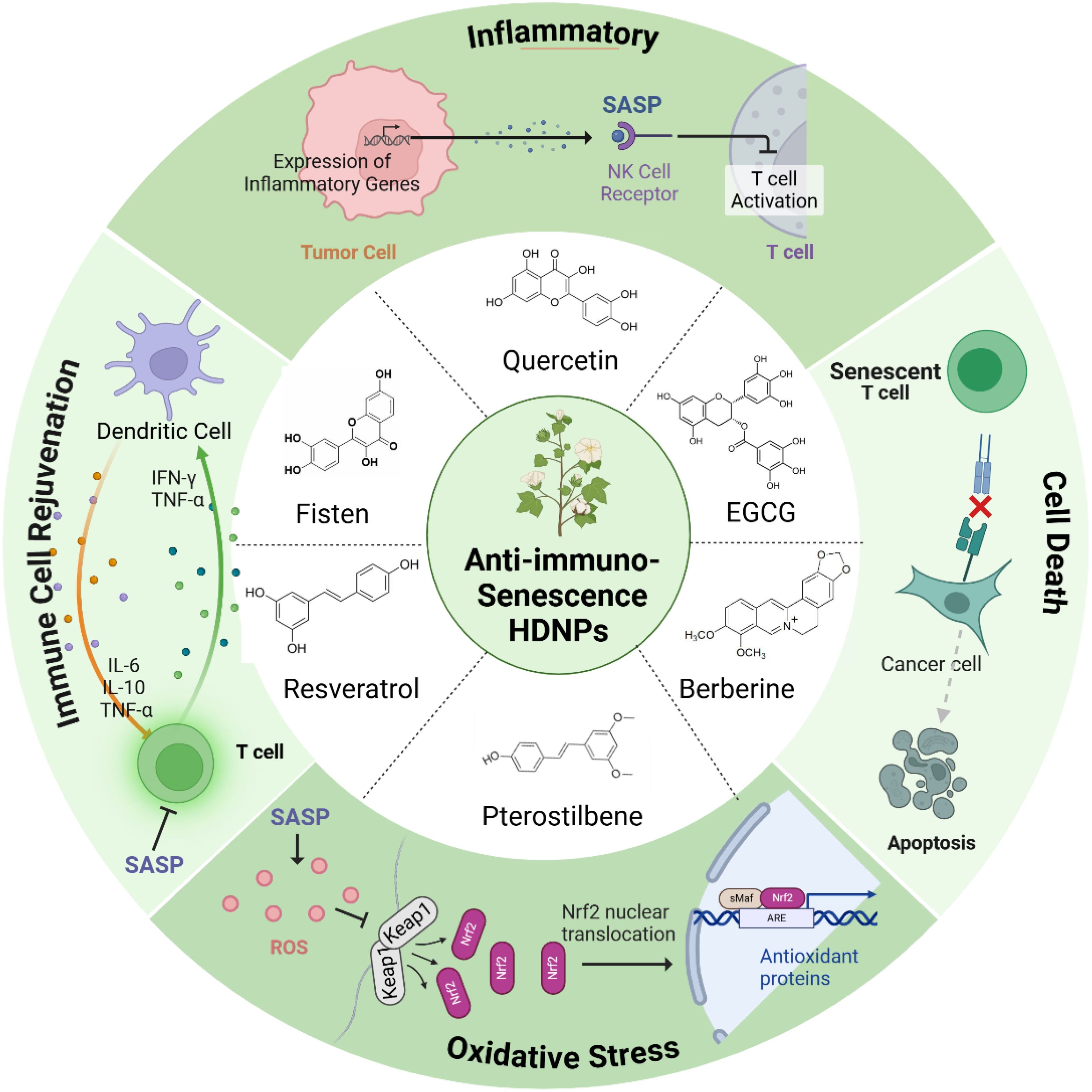
Figure 2. HDNPs with anti-immunosenescence properties and their mechanisms of action.
Quercetin, abundant in fruits and vegetables, selectively induces apoptosis in senescent cells by targeting anti-apoptotic pathways unique to these cells, thus reducing the tumor-supportive milieu created by senescent cells (37, 38). Fisetin, primarily found in strawberries and apples, effectively decreases the secretion of SASP factors such as IL-6 and IL-8, reducing inflammation and rejuvenating the proliferative capacity of cytotoxic immune cells including T cells and NK cells (39, 40). EGCG from green tea modulates immune signaling pathways via suppression of NF-κB and AP-1, thus attenuating pro-inflammatory gene expression and fostering immune homeostasis through Treg cell modulation and anti-inflammatory differentiation of naïve T cells (41, 42). Resveratrol, found in grapes and berries, enhances immune function and delays immune cell senescence by activating longevity-associated pathways such as SIRT1 and AMPK, thereby reinforcing antitumor immune surveillance (43). Curcumin, derived from turmeric, exhibits anti-inflammatory properties by suppressing NF-κB signaling, reducing SASP-related inflammation (44). Ginsenosides from Panax ginseng demonstrate immunomodulatory effects that combat immune aging, potentially rejuvenating T-cell and NK-cell functions (45). Similarly, Astragaloside IV promotes telomerase activity, delaying cellular senescence and enhancing immune cell functionality (46). Berberine suppresses pro-inflammatory SASP factors through inhibition of the NF-κB pathway, while sulforaphane induces apoptosis specifically in senescent cells, reducing their accumulation (47, 48). Genistein, an isoflavone in soy products, further contributes by inhibiting inflammatory signaling pathways critical in SASP regulation (49) (Table 1).
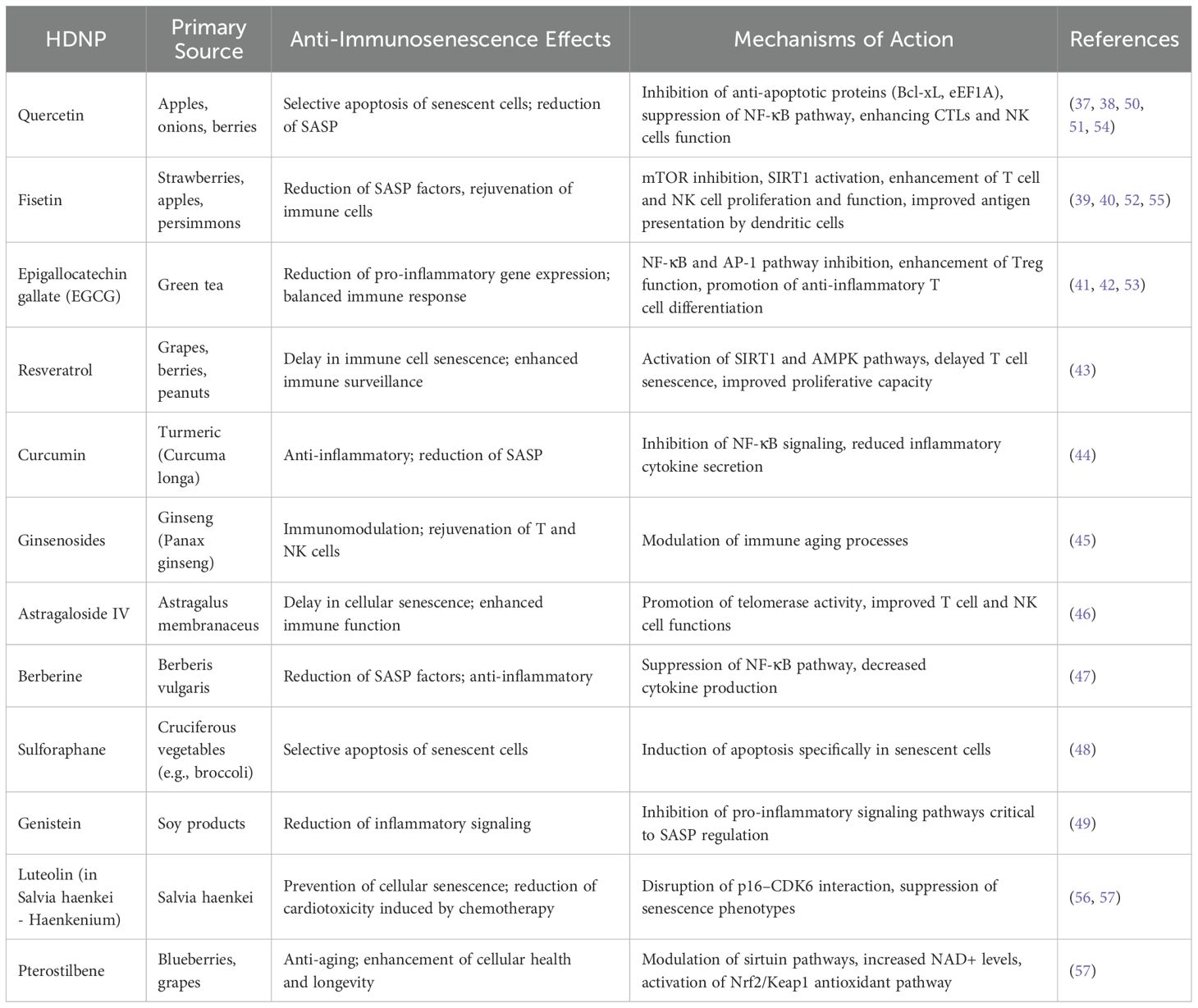
Table 1. Mechanistic insights into the effects of HDNPs on immunosenescence in the TME.
3.2 Mechanisms of action
HDNPs counter immunosenescence through three primary mechanisms: senolytic activity, inflammation modulation, and immune cell rejuvenation.
As senolytics, quercetin and fisetin selectively induce apoptosis in senescent cells via inhibition of anti-apoptotic proteins such as Bcl-xL and eEF1A, significantly reducing SASP burden and tumor-promoting inflammation (50, 51). Fisetin’s ability to modulate mTOR and activate SIRT1 pathways further reduces secretion of pro-inflammatory cytokines IL-6 and IL-8, thus limiting inflammatory signaling in the TME (52). EGCG similarly decreases NF-κB activation, curtailing chronic inflammation and hampering tumor survival pathways (53). In immune rejuvenation, quercetin enhances cytotoxic functions of CTLs and NK cells, significantly improving their tumor-eliminating capacity (54). Fisetin boosts antigen presentation capabilities by increasing co-stimulatory molecules on dendritic cells, facilitating enhanced T cell activation (55). EGCG promotes balanced immune responses by expanding Treg populations that mitigate excessive inflammation without compromising antitumor immunity (41). Additionally, Zumerle et al. demonstrated that a polyphenol-rich natural extract from Salvia haenkei (Haenkenium, HK) can delay aging in mice by lowering systemic senescence markers such as p16 and p27 (56). Luteolin in HK disrupts the p16–CDK6 interaction, preventing cell cycle arrest, suppressing senescence phenotypes, and mitigating doxorubicin-induced senescence and cardiotoxicity—thereby highlighting HK’s potential to reduce side effects and confer anti-aging benefits in cancer therapy (57). Additionally, pterostilbene exerts anti-aging and anti-senescent effects by modulating sirtuin pathways and enhancing NAD+ levels. This potent antioxidant—chemically similar to resveratrol but more bioavailable—can boost NAD+ up to 90% in 30 days when combined with nicotinamide riboside (NR), underscoring its promise for improving cellular health and longevity (57). Moreover, pterostilbene activates the Nrf2/Keap1 pathway, fortifying the cell’s antioxidant defenses and thereby improving resilience to oxidative stress—hallmarks of aging-related cellular deterioration (57) (Figure 2).
Despite these promising biological effects, HDNPs face substantial pharmacokinetic barriers including poor bioavailability, rapid systemic clearance, and potential off-target toxicity at high concentrations. To overcome these limitations, nanoparticle-based delivery systems offer significant advantages by enhancing stability, bioavailability, and targeted release of HDNPs to senescent cells within the TME. Functionalized nanoparticles provide precision delivery, improving therapeutic efficacy while minimizing systemic toxicity and adverse effects (20, 58–61) (Table 1).
4 Nanoparticle-mediated delivery of HDNPs
4.1 Improved stability and solubility
Incorporating HDNPs into nanoparticle delivery systems offers several advantages that enhance their therapeutic potential against immunosenescence-induced drug resistance in cancer therapy. Many HDNPs, such as quercetin and fisetin, possess poor water solubility and are unstable under physiological conditions, which limits their bioavailability and therapeutic efficacy. Encapsulating these compounds in nanoparticles, such as liposomes, polymeric nanoparticles, polymeric micelles, nanogels, carbon nanotubes, and gold nanoparticles, can significantly enhance their solubility and protect them from degradation (Figure 3). Nanoparticles can shield HDNPs from enzymatic metabolism and chemical hydrolysis, prolonging their systemic circulation time and increasing the likelihood of reaching target sites (20).
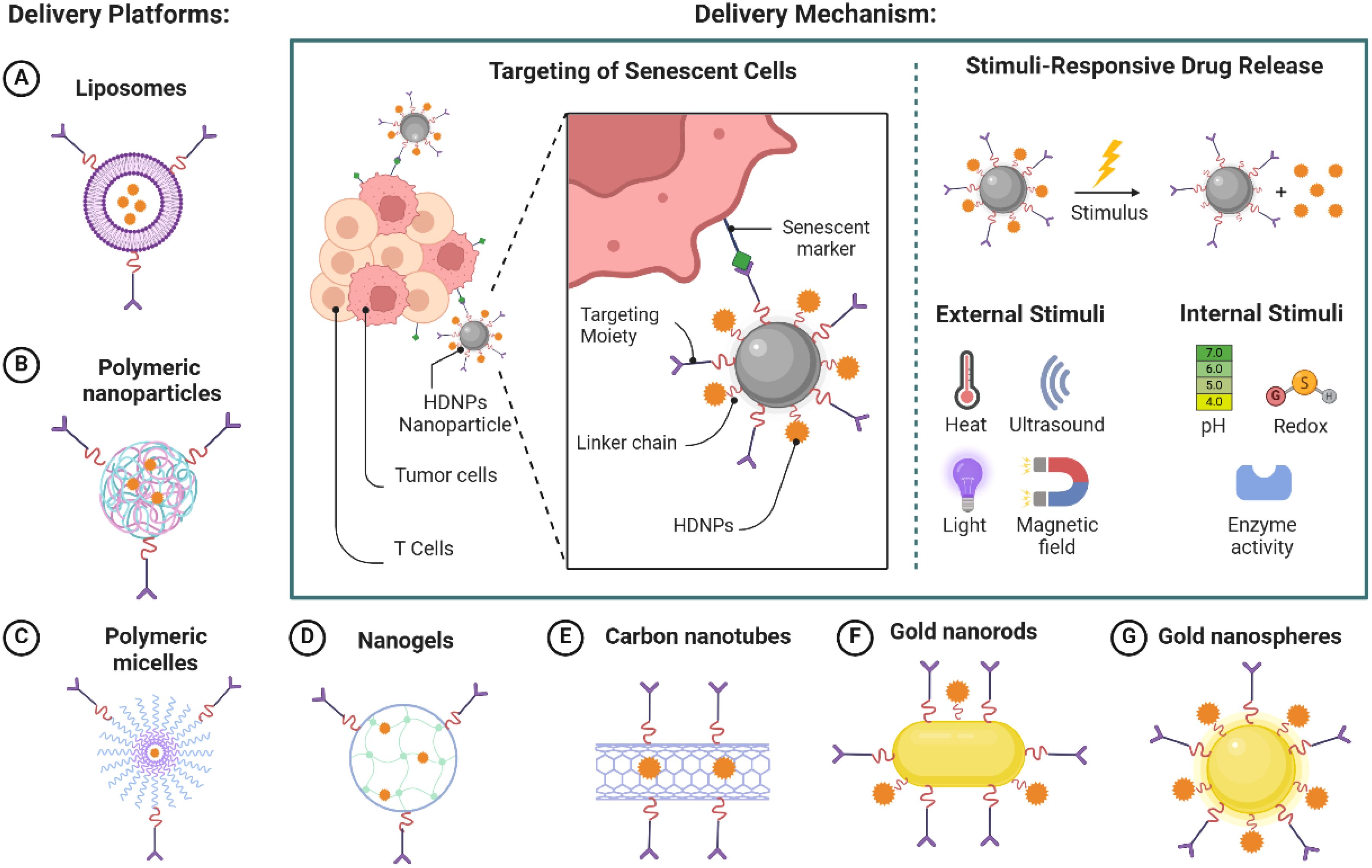
Figure 3. Nanoparticle-mediated delivery of HDNPs targeting senescent cells.
Nanoparticles can be engineered for targeted delivery to the TME and senescent immune cells. Selective accumulation at the desired site is achieved by modifying the surface of nanoparticles with specific ligands or antibodies that recognize senescence-associated markers. Although senescence-associated β-galactosidase (SA-β-gal) is commonly used, it lacks specificity due to its association with general lysosomal activity. Therefore, more robust targeting strategies integrate multiple markers—such as p16INK4A, p21Cip1, γ-H2AX, and SASP-related surface proteins—to more accurately identify senescent cells. This targeted approach enhances the local concentration of HDNPs, maximizing therapeutic effects while minimizing off-target toxicity.
4.2 Controlled release profiles
Nanoparticle delivery systems can be designed to release controlled HDNPs that respond to specific stimuli within the TME. Controlled release mechanisms, such as pH-sensitive or enzyme-responsive systems, ensure that HDNPs are released at the site of action over an extended period (62). This maintains effective therapeutic concentrations and reduces systemic exposure and side effects, improving patient compliance and treatment outcomes. To effectively modulate immunosenescence, nanoparticles must be strategically designed to deliver HDNPs specifically to senescent immune cells within the TME. Senescent cells express unique biomarkers that can be exploited for targeted drug delivery (63). Nanoparticles can be functionalized with ligands or antibodies that specifically bind to the markers, enhancing uptake by senescent cells (64). This targeted delivery ensures that HDNPs exert their senolytic effects primarily on senescent cells, reducing the burden of the senescent cell population within the TME.
The TME exhibits distinct characteristics, such as acidic pH, hypoxia, and elevated levels of certain enzymes, which can be utilized to trigger the release of HDNPs from nanoparticles (65). Designing stimuli-responsive nanoparticles allows for: I) PH-sensitive release: Nanoparticles remain stable at physiological pH but release their payload in the acidic environment of the TME (pH ~6.5) or within endosomal/lysosomal compartments (pH ~5.0) after cellular uptake (66). II) Enzyme-responsive release: Nanoparticles degrade and release HDNPs in the presence of enzymes overexpressed in the TME, such as MMPs (67). These design strategies ensure that HDNPs are released precisely where needed, enhancing efficacy and reducing systemic toxicity (Figure 3).
4.3 Examples of nanoparticle-HDNP systems
Liposomes are versatile nanocarriers composed of phospholipid bilayers capable of encapsulating hydrophilic and hydrophobic compounds. Quercetin-loaded liposomes have been developed to target senescent fibroblasts within the TME (68). Recent studies have demonstrated that liposomes can be engineered with different surface properties, such as cationic, anionic, and zwitterionic characteristics, to optimize their interaction with cellular membranes and improve their therapeutic efficacy (69). Research indicates that quercetin and rutin, when encapsulated in cationic liposomes, can inhibit the toxic effects of empty liposomes, thereby enhancing cell viability (69). This targeted approach aims to deliver quercetin more effectively and exploit the unique properties of the TME, where senescent cells often reside, to improve therapeutic outcomes in cancer treatment. Resveratrol-loaded nanoparticles have been engineered to enhance targeting and uptake in immunosenescent cells within the TME, specifically by modulating nanoparticle surface chemistry and functional groups to bind receptors upregulated in senescent cells (70). Mechanistically, these nanoparticles utilize ligand-mediated binding to specific markers such as SA-β-gal or CD38, which are highly expressed in immunosenescent cells. While SA-β-gal is commonly employed, CD38 is increasingly recognized as an immunosenescence marker due to its elevated expression in aged immune cells, where it contributes to NAD+ degradation, cellular metabolic dysregulation, and impaired immune function (71). Once internalized, the resveratrol payload exerts senolytic and senostatic effects, inhibiting pro-inflammatory cytokine release (SASP factors) and restoring immune functionality through SIRT1 activation, downregulation of NF-κB signaling, and attenuation of oxidative stress (70). This approach selectively targets immunosenescent cells and reduces chronic inflammation in the TME, potentially enhancing immune cell infiltration and improving therapeutic responses to cancer immunotherapy (Table 2).
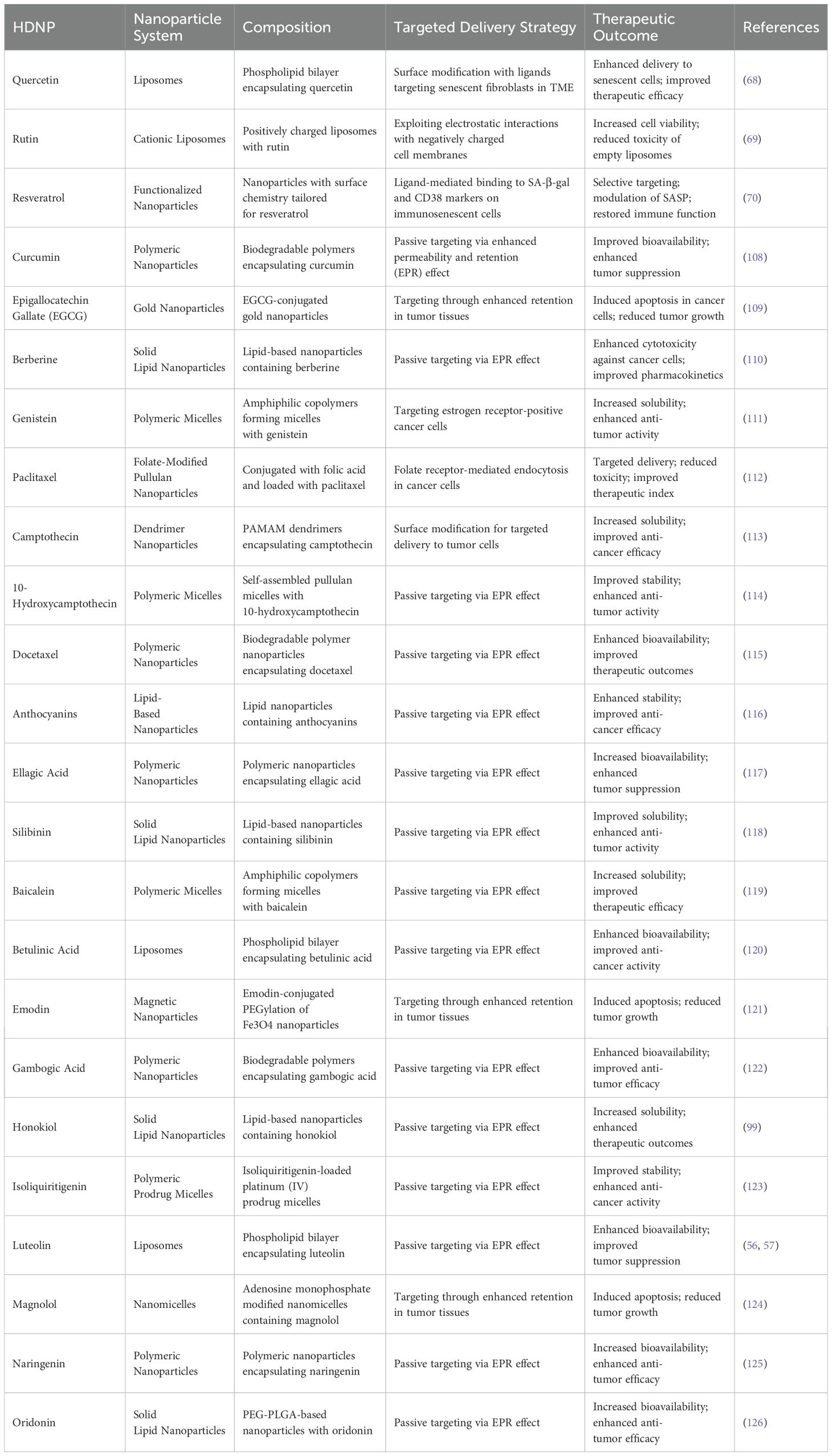
Table 2. Summary of nanoparticle-HDNP systems in cncer therapy.
Not only that but innovative nanoparticle designs for HDNP delivery are also needed. For example, the development of nanoparticles that respond to multiple TME stimuli (e.g., pH, redox conditions) to achieve precise spatiotemporal release of HDNPs (72). Besides, incorporating imaging agents into HDNP-loaded nanoparticles for theranostic applications allows simultaneous cancer therapy and monitoring (73). Meanwhile, the combination therapies are also important (74). Designing nanoparticles co-loaded with HDNPs and chemotherapeutic agents to achieve synergistic effects in overcoming drug resistance. Nanoparticles loaded with HDNPs offers a promising strategy to modulate immunosenescence-induced drug resistance in cancer therapy. By improving the pharmacokinetic profiles of HDNPs and enabling targeted delivery to senescent cells within the TME, nanoparticles enhance therapeutic efficacy while minimizing systemic side effects (Table 2). Beyond their application in HDNP-based therapies, nanoparticle-mediated delivery systems represent a broadly applicable platform for enhancing the pharmacological performance of diverse therapeutic agents. Their versatility, biocompatibility, and capacity for targeted delivery make them attractive not only for cancer treatment but also for addressing a wide range of diseases globally. These advantages highlight the potential of nanoparticle systems to serve as a foundational strategy in the development of next-generation therapeutics.
5 Overcoming drug resistance by targeting immunosenescence with HDNP-loaded nanoparticles
5.1 Modulation of pro-survival signaling and SASP factors
Immunosenescence—the gradual decline of the immune system due to aging—plays a pivotal role in fostering an immunosuppressive TME, which significantly contributes to cancer progression and the development of drug resistance. This process involves the accumulation of senescent immune cells that secrete the SASP. The SASP not only promotes tumor growth but also enhances the survival and proliferation of cancer cells, thereby impeding the efficacy of various therapeutic interventions (15).
HDNPs encapsulated within nanoparticles have emerged as a promising strategy to counteract immunosenescence and its associated drug resistance. These HDNP-loaded nanoparticles can selectively target and eliminate senescent cells within the TME, thereby reducing SASP-mediated pro-survival signals that contribute to therapeutic resistance. Quercetin-loaded nanoparticles have demonstrated the ability to inhibit the NF-κB signaling pathway, leading to decreased secretion of IL-6 and IL-8—cytokines implicated in cancer cell survival and chemoresistance (75). Similarly, fisetin modulates the mTOR pathway and activates SIRT1, significantly diminishing SASP secretion and associated pro-survival signaling (39). Encapsulating these HDNPs within nanoparticles enhances their bioavailability and delivery precision, specifically concentrating their effects in the TME, leading to a more profound reduction in pro-survival signals and effectively reversing drug resistance.
Additionally, SASP factors directly inhibit apoptosis and enhance cell survival through various signaling cascades. HDNP-loaded nanoparticles can restore apoptotic signaling pathways in cancer cells compromised by SASP-driven resistance. EGCG delivered via nanoparticles has shown enhanced intracellular uptake, significantly activating apoptotic mechanisms by promoting caspase activation, upregulating pro-apoptotic proteins (e.g., Bax), and downregulating anti-apoptotic proteins (e.g., Bcl-2) (76). By overcoming bioavailability limitations, nanoparticle delivery ensures therapeutic concentrations within cancer cells, countering the anti-apoptotic effects mediated by SASP factors. (Figure 4).
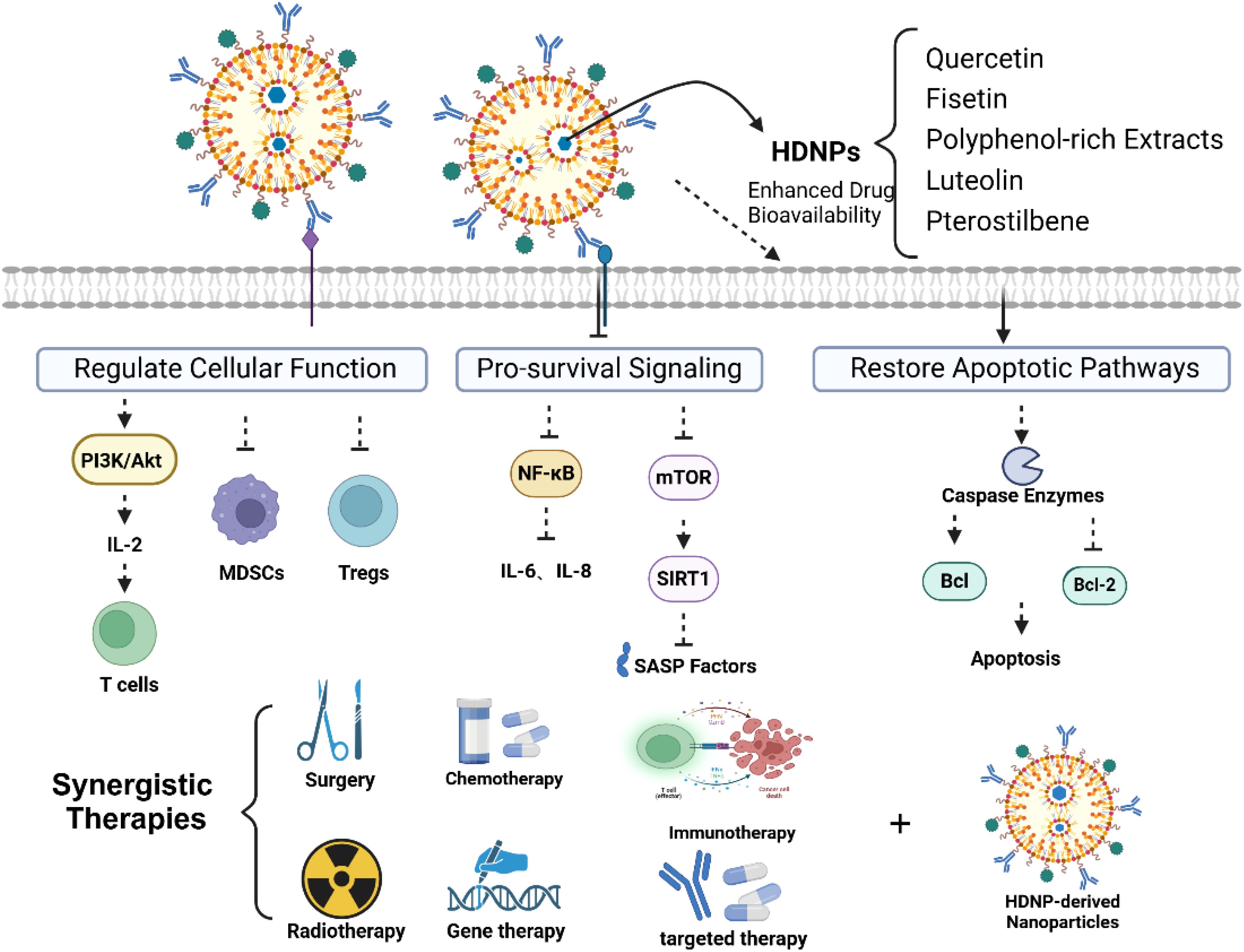
Figure 4. Overcoming drug resistance through modulation of immunosenescence.
5.2 Rejuvenation of immune cell function and reversal of immunosuppression
The TME characterized by immunosenescence exhibits diminished CTL functionality alongside increased populations of immunosuppressive cells such as MDSCs and Tregs, leading to compromised anti-tumor immunity (77). HDNP-loaded nanoparticles represent an innovative therapeutic strategy with substantial potential for reversing these immune deficits associated with cellular senescence, thus rejuvenating immune cell function (Figure 4).
Fisetin-loaded nanoparticles demonstrate notable efficacy in revitalizing senescent CTLs. By encapsulating fisetin within targeted nanoparticles, studies have shown significant improvements in the proliferation, cytotoxicity, and effector functions of CTLs, reversing signs of immunosenescence through modulation of key signaling pathways, including PI3K/Akt, and by enhancing IL-2 secretion (78, 79). These nanoparticles ensure precise delivery to aged, dysfunctional CTLs, restoring their capacity to effectively identify and eliminate tumor cells, thereby strengthening immune surveillance compromised by aging.
Quercetin-loaded nanoparticles also play a critical role in targeting immunosenescent, immunosuppressive cell populations within the TME. Research demonstrates that these nanoparticles effectively reduce both the expansion and suppressive activities of senescent MDSCs and Tregs by inhibiting immunosuppressive signaling pathways such as STAT3 and NF-κB (80). By selectively delivering quercetin to senescent immunosuppressive cells, nanoparticles enhance treatment specificity, significantly mitigating off-target effects and improving therapeutic outcomes.
Additionally, curcumin-loaded nanoparticles have been extensively investigated for their potential in reversing immunosenescence-associated immune suppression. Curcumin nanoparticles effectively reduce senescence-driven accumulation and functionality of Tregs and MDSCs, decrease levels of pro-inflammatory SASP cytokines such as IL-6 and TGF-β, and simultaneously rejuvenate effector T cell responses by modulating the inflammatory cytokine profile within the aged TME (81, 82). Further studies highlight the promising role of resveratrol-loaded nanoparticles in counteracting immunosenescence-associated immunosuppression. These nanoparticles notably reduce the recruitment and function of aged MDSCs, simultaneously promoting CTL infiltration and activation through mechanisms involving SIRT1 activation and inhibition of NF-κB signaling pathways, which are commonly dysregulated in senescent immune cells (83, 84). EGCG-loaded nanoparticles derived from green tea exhibit remarkable efficacy in overcoming immune senescence. They modulate the aged immune environment by reducing immunosenescence-induced immunosuppressive factors and promoting T cell-mediated immunity, thereby restoring a favorable milieu for immune cell activation and tumor suppression (85, 86).
Moreover, integrating multiple HDNP-loaded nanoparticles into combinational therapies represents a promising strategy to reverse comprehensive immunosenescence within the TME. Nanoparticles co-loaded with HDNPs and immune checkpoint inhibitors demonstrate enhanced therapeutic efficacy by simultaneously reversing immunosenescence-induced suppression and rejuvenating overall immune function. Nanoparticles co-delivering curcumin and anti-PD-1 antibodies exhibit synergistic effects in preclinical models, significantly improving T cell infiltration and reversing tumor-associated immune aging and dysfunction (87). Collectively, these nanoparticle-mediated HDNP strategies underscore the significant therapeutic potential in rejuvenating senescent immune cell functionality, reversing age-associated immunosuppression, and effectively overcoming immunosenescence-driven drug resistance in cancer therapy, highlighting their importance for further clinical translation.
5.3 Synergistic integration of HDNP-loaded nanoparticles with conventional therapies
Integrating HDNP-loaded nanoparticles with standard cancer treatments such as chemotherapy, radiotherapy, and immunotherapy represents a powerful strategy to counteract immunosenescence-driven drug resistance. These nanoparticles can potentiate conventional therapies by reducing immunosuppressive signaling, enhancing apoptosis in cancer cells, and restoring immune surveillance (Figure 4). The co-delivery of quercetin and doxorubicin via nanoparticles has resulted in notable synergistic effects. Quercetin inhibits drug efflux mechanisms and survival pathways, thereby significantly enhancing doxorubicin’s cytotoxicity even in resistant cancer cells (88).
Furthermore, combining HDNP-loaded nanoparticles with conventional chemotherapeutics enables dose reductions, minimizing adverse effects without compromising treatment efficacy. Studies involving curcumin nanoparticles combined with paclitaxel showed enhanced anti-tumor efficacy at lower paclitaxel doses compared to higher doses of paclitaxel alone, demonstrating the potential of nanoparticle delivery systems in enhancing therapeutic efficiency and safety profiles (89). Future research should prioritize the development of multifunctional, personalized HDNP nanoparticle systems capable of addressing multiple immunosenescence mechanisms simultaneously, optimizing clinical outcomes in cancer treatment.
6 Preclinical evidence and case studies
6.1 In vitro
Several in vitro studies have demonstrated that nanoparticle-mediated delivery of HDNPs effectively reduces senescence markers in cancer cell cultures. Quercetin-loaded nanoparticles have decreased the expression of senescence-associated markers in senescent tumor cells. In a study by Lewinska et al., quercetin encapsulated in nanoparticles significantly reduced senescence markers in human fibroblast cultures compared to free quercetin, indicating enhanced senolytic activity (90). Similarly, fisetin-loaded nanoparticles have been reported to suppress senescence markers in prostate cancer cells. Researchers formulated fisetin nanoparticles using PLGA (poly(lactic-co-glycolic acid)) and demonstrated that the nanoparticles reduced the expression of SASP factors such as IL-6 and IL-8 more effectively than fisetin alone (91). This suggests that nanoparticle delivery enhances the ability of HDNPs to modulate immunosenescence at the cellular level.
Nanoparticle-HDNP formulations have exhibited enhanced cytotoxic effects against drug-resistant cancer cell lines. EGCG-loaded nanoparticles were tested against multidrug-resistant breast cancer cells. Research showed that the nanoparticles increased intracellular accumulation of EGCG, leading to higher apoptosis rates in resistant cells than in free EGCG (92). The nanoparticles overcame efflux pump-mediated drug resistance by facilitating EGCG entry into cancer cells. Additionally, quercetin nanoparticles have been found to overcome resistance in ovarian cancer cells resistant to paclitaxel. Wang et al. demonstrated that quercetin nanoparticles inhibited cell proliferation and induced apoptosis more effectively than free quercetin, suggesting that nanoparticle delivery can enhance the cytotoxicity of HDNPs against resistant cancer cells (93). The study highlighted the potential of nanoparticles to sensitize resistant cancer cells to conventional therapies.
6.2 In vivo
In vivo, studies have provided evidence that nanoparticle-mediated delivery of HDNPs can delay tumor growth and reduce drug resistance. Studies investigated the effects of quercetin-loaded nanoparticles in mice bearing drug-resistant lung tumors. The treatment resulted in significant tumor growth inhibition and decreased expression of drug resistance proteins compared to controls (94). The nanoparticles improved the bioavailability of quercetin and facilitated its accumulation in tumor tissues. Similarly, Gera et al. have developed a nano-photo composite consisting of phytochemical extract (BRM270) and estimated its cytotoxic potential against HepG2 human hepatoma cancer cells. The toxicity potential of nano-photo composite against HepG2 cells was higher than free BRM270. The nano-photo composite also shows a reduction in cell growth compared to doxorubicin as it reduced concentration to 12 μg/mL. The BRM270 extract was found to downregulate specific proteins that are overexpressed during chemotherapy in HepG2 cells. The BRM270 inhibits the regulation of MMP9, BCL2, and IL 6, which are related to the potential induced apoptosis and cell proliferation (95). The enhanced anti-tumor activity was attributed to improved delivery and sustained release of drugs at the tumor site.
It is necessary to note that enhanced infiltration of functional immune cells into the tumor site is essential. Nanoparticle-HDNP formulations have been shown to modulate the tumor immune microenvironment. Treating EGCG-loaded nanoparticles in a mouse model of colon cancer led to increased infiltration of CD8+ cytotoxic T lymphocytes and decreased numbers of immunosuppressive MDSCs in tumor tissues (96). This shift enhanced anti-tumor immunity and contributed to tumor regression. Moreover, a study found quercetin nanoparticles to enhance NK cell activity within tumors (97). The nanoparticles improved the cytotoxic function of NK cells, contributing to tumor regression. These findings suggest nanoparticle-mediated HDNP delivery can rejuvenate immune cell functions suppressed by immunosenescence.
6.3 Comparative analysis
Comparative studies have consistently shown nanoparticle-mediated HDNP delivery is more effective than HDNPs administered alone. In the study, mice treated with quercetin nanoparticles exhibited more excellent tumor suppression than those treated with free quercetin, highlighting the enhanced efficacy of nanoparticle delivery (98). The nanoparticles improved pharmacokinetics and facilitated targeted delivery to tumors. Similarly, another research demonstrated that fisetin nanoparticles had superior anti-tumor effects than free fisetin in melanoma-bearing mice (99). The enhanced efficacy was attributed to increased bioavailability, targeted delivery, and sustained release provided by the nanoparticle system.
Preclinical models have shown that nanoparticle-HDNP treatments can improve survival rates and quality of life indicators compared to the HDNP monomer groups. In the study by Granja et al., mice treated with EGCG nanoparticles had prolonged survival and better physical condition than controls, indicating improved quality of life (92). The treatment was associated with reduced tumor burden and minimal toxicity. Furthermore, the study by Park et al. reported that quercetin nanoparticle treatment not only extended survival but also reduced systemic toxicity, as evidenced by stable body weight and normal organ histology in treated animals (100). These findings suggest nanoparticle-mediated HDNP delivery can enhance therapeutic efficacy while minimizing adverse effects, potentially translating into better clinical outcomes.
7 Clinical translation prospects and limitations
While nanoparticle-mediated delivery of HDNPs offers substantial therapeutic potential, several critical limitations and challenges must be thoroughly addressed to ensure safe and effective clinical translation. One significant challenge is ensuring the safety, biocompatibility, and controlled biodistribution of nanoparticle systems. Nanoparticles can interact unpredictably with biological systems, leading to potential toxicity, immunogenic reactions, and unintended distribution to non-target tissues (101). The physicochemical properties of nanoparticles, such as size, shape, surface charge, and composition, can influence their interaction with cells and tissues. For instance, smaller nanoparticles may penetrate tissues more deeply but pose a higher risk of crossing biological barriers and accumulating in non-target organs (101). Moreover, the long-term effects of nanoparticles in the body are not fully understood. Some materials used in nanoparticle formulations may be non-biodegradable or degrade into toxic byproducts. Thus, rigorous long-term toxicity assessments and biodegradation studies are essential before clinical applications. Furthermore, challenges in reproducibility and consistency during nanoparticle manufacturing and scaling up production processes pose significant barriers to clinical translation. The complexity of manufacturing consistent nanoparticle batches with precise physicochemical properties can complicate regulatory approval and clinical deployment.
The regulatory landscape for combination therapies that involve natural products and advanced drug delivery systems is complex. Herbal-derived compounds are often classified as dietary supplements or herbal medicines with different regulatory requirements than conventional pharmaceuticals. When combined with nanoparticles, HDNPs may fall under the category of combination products, necessitating compliance with regulations for drugs and medical devices (102). Regulatory agencies, such as the U.S. Food and Drug Administration (FDA) and the European Medicines Agency (EMA), require rigorous documentation of safety, efficacy, and quality for approval (103). Challenges include the standardization of natural product extracts, batch-to-batch consistency, and the complexity of nanoparticle characterization. Additionally, the lack of clear guidelines for nanoparticle-based natural product therapies can impede regulatory approval (21). Collaborative efforts between regulatory bodies, researchers, and industry stakeholders are needed to develop comprehensive frameworks that facilitate the clinical translation of these innovative therapies.
Although several clinical trials have explored HDNPs and nanoparticle systems in cancer treatment, clinical studies specifically targeting immunosenescence-induced drug resistance remain limited. For example, a Phase I clinical trial evaluated the safety and pharmacokinetics of nano-encapsulated curcumin (a polyphenol derived from turmeric) in patients with advanced pancreatic cancer, demonstrating acceptable safety profiles and some therapeutic benefits (104). Another trial studied the efficacy of green tea polyphenols in prostate cancer patients, showing reduced levels of prostate-specific antigen (PSA) and indicating potential anti-cancer activity (105). Advances in understanding the role of immunosenescence in cancer progression and drug resistance provide a rationale for designing clinical trials that assess the efficacy of nanoparticle-HDNP systems in modulating immune aging and improving therapeutic outcomes. In a recent study, researchers developed a self-assembling nanoparticle using the natural anticancer agent honokiol (HK) (Phase I trial CTR20170822), achieving 100% drug loading and enhanced tumor targeting via the EPR effect while improving p53-selective antitumor immunity with excellent stability and minimal toxicity (106, 107). Nonetheless, focused clinical trials assessing the efficacy and safety of nanoparticle-HDNP systems in modulating immunosenescence and overcoming drug resistance are critically needed.
Advancing clinical research should prioritize studies that explicitly target aging-related immune dysfunction. Clinical trials should emphasize elderly patient populations who are more susceptible to immunosenescence-driven drug resistance. Future research could explore combinational therapeutic approaches by integrating nanoparticle-HDNP systems with established chemotherapies or immunotherapies to potentiate clinical outcomes and reduce adverse effects. Moreover, incorporating validated biomarkers of immunosenescence, such as senescence-associated genes and immune cell profiles, could facilitate personalized treatment approaches and precise monitoring of therapeutic responses. Stratifying patients based on their immunosenescence status may further optimize therapeutic efficacy, providing essential insights into resistance mechanisms and enabling more effective patient-specific therapeutic strategies.
8 Conclusions
This article delves into the complex interplay between immunosenescence and drug resistance in cancer treatment. The immune aging process, characterized by altered cytokine profiles and weakened immune surveillance, plays a critical role in the emergence of drug resistance. HDNPs have shown promise in addressing immunosenescence by triggering apoptosis in senescent cells, reducing the secretion of SASP factors, and boosting the functionality of immune cells. Enhancing the stability, solubility, and targeted delivery of HDNPs to the TME and senescent immune cells via nanoparticles has amplified their therapeutic potential. These advancements have demonstrated an ability to decrease senescence markers, improve cytotoxic effects on resistant cancer cell lines, delay tumor progression, and increase the infiltration of functional immune cells into tumors. Although preclinical studies show great potential, more research is necessary to comprehensively understand the mechanisms through which immunosenescence promotes drug resistance and how HDNP-loaded nanoparticles can modulate these pathways. Further investigation into additional senolytic HDNPs and the refinement of nanoparticle formulations could lead to more potent therapeutic options. In conclusion, nanoparticle-mediated HDNP delivery offers an innovative and promising approach to overcoming drug resistance caused by immunosenescence in cancer therapy. By addressing the core mechanisms of immune aging, this strategy can potentially improve treatment efficacy and enhance the quality of life for cancer patients.
Author contributions
LY: Funding acquisition, Writing – original draft. MX: Conceptualization, Funding acquisition, Writing – original draft, Writing – review & editing. JY: Supervision, Project administration, Writing – review & editing. WW: Supervision, Project administration, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was funded by the National Natural Science Foundation of China (82204922); the China Academy of Traditional Chinese Medicine Outstanding Young Scientific and Technological Talents Cultivation Special Programme (ZZ18-YQ-022); the Postgraduate Research & Practice Innovation Program of Jiangsu Province (SJCX2309690); and the TCM Science and Technology Development Plan of Jiangsu Province: Clinical Study of the Five Elements Theory of TCM in the Metastasis, Prevention, and Treatment of Primary Lung Cancer (MS2022064).
Acknowledgments
Figures were created with BioRender software, biorender.com.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Holohan C, Van Schaeybroeck S, Longley DB, Johnston PG. Cancer drug resistance: an evolving paradigm. Nat Rev Cancer. (2013) 13:714–26.
3. Wang L, Wang X, Zhu X, Zhong L, Jiang Q, Wang Y, et al. Drug resistance in ovarian cancer: from mechanism to clinical trial. Mol Cancer. (2024) 23:66.
4. Picco N, Milne A, Maini P, Anderson A. The role of environmentally mediated drug resistance in facilitating the spatial distribution of residual disease. (2024). doi: 10.21203/rs.3.rs-4526427/v1
5. Bhat GR, Sethi I, Sadida HQ, Rah B, Mir R, Algehainy N, et al. Cancer cell plasticity: From cellular, molecular, and genetic mechanisms to tumor heterogeneity and drug resistance. Cancer Metastasis Rev. (2024) 43:197–228. doi: 10.1007/s10555-024-10172-z
6. Pawelec G. Immunosenescence and cancer. Biogerontology. (2017) 18:717–21. doi: 10.1007/s10522-017-9682-z
7. Pereira BI, Akbar AN. Convergence of innate and adaptive immunity during human aging. Front Immunol. (2016) 7:445. doi: 10.3389/fimmu.2016.00445
8. Fane M, Weeraratna AT. How the ageing microenvironment influences tumour progression. Nat Rev Cancer. (2020) 20:89–106. doi: 10.1038/s41568-019-0222-9
9. Liu Z, Liang Q, Ren Y, Guo C, Ge X, Wang L, et al. Immunosenescence: molecular mechanisms and diseases. Signal Transduct Targeted Ther. (2023) 8:200. doi: 10.1038/s41392-023-01451-2
10. Rodriguez JE, Naigeon M, Goldschmidt V, Roulleaux Dugage M, Seknazi L, Danlos FX, et al. Immunosenescence, inflammaging, and cancer immunotherapy efficacy. Expert Rev Anticancer Ther. (2022) 22:915–26. doi: 10.1080/14737140.2022.2098718
11. Jagger A, Shimojima Y, Goronzy JJ, Weyand CM. Regulatory T cells and the immune aging process: a mini-review. Gerontology. (2014) 60:130–7. doi: 10.1159/000355303
12. Zhang J-W, Zhang D, Yu B-P. Senescent cells in cancer therapy: why and how to remove them. Cancer Lett. (2021) 520:68–79. doi: 10.1016/j.canlet.2021.07.002
13. Fulop T, Larbi A, Dupuis G, Le Page A, Frost EH, Cohen AA, et al. Immunosenescence and inflamm-aging as two sides of the same coin: friends or foes? Front Immunol. (2018) 8:1960. doi: 10.3389/fimmu.2017.01960
14. Zhang T, Wen R, Fan H, Yu Y, Jia H, Peng Z, et al. Impact and potential value of immunosenescence on solid gastrointestinal tumors. Front Immunol. (2024) 15:1375730. doi: 10.3389/fimmu.2024.1375730
15. Faget DV, Ren Q, Stewart SA. Unmasking senescence: context-dependent effects of SASP in cancer. Nat Rev Cancer. (2019) 19:439–53. doi: 10.1038/s41568-019-0156-2
16. Vaiserman A, Koliada A, Lushchak O. Phyto-nanotechnology in anti-aging medicine. Aging (Albany NY). (2021) 13:10818. doi: 10.18632/aging.203026
17. Jiang M, Lv Z, Huang Y, Cheng Z, Meng Z, Yang T, et al. Quercetin alleviates lipopolysaccharide-induced inflammatory response in bovine mammary epithelial cells by suppressing TLR4/NF-κB signaling pathway. Front Vet Sci. (2022) 9:915726. doi: 10.3389/fvets.2022.915726
18. Maher P. Modulation of multiple pathways involved in the maintenance of neuronal function during aging by fisetin. Genes Nutr. (2009) 4:297–307. doi: 10.1007/s12263-009-0142-5
19. Mereles D, Hunstein W. Epigallocatechin-3-gallate (EGCG) for clinical trials: more pitfalls than promises? Int J Mol Sci. (2011) 12:5592–603. doi: 10.3390/ijms12095592
20. Mohammed FS, Omay SB, Sheth KN, Zhou J. Nanoparticle-based drug delivery for the treatment of traumatic brain injury. Expert Opin Drug Deliv. (2023) 20:55–73. doi: 10.1080/17425247.2023.2152001
21. Anselmo AC, Mitragotri S. Nanoparticles in the clinic: An update. Bioeng Trans Med. (2019) 4:e10143. doi: 10.1002/btm2.10143
22. Fülöp T, Larbi A, Witkowski JM. Human inflammaging. Gerontology. (2019) 65:495–504. doi: 10.1159/000497375
23. Saavedra D, Añé-Kourí AL, Barzilai N, Caruso C, Cho K-H, Fontana L, et al. Aging and chronic inflammation: highlights from a multidisciplinary workshop. Immun Ageing. (2023) 20:25. doi: 10.1186/s12979-023-00352-w
24. Zhao J, Wang Z, Tian Y, Ning J, Ye H. In T cell exhaustion and senescence for ovarian cancer immunotherapy. Semin Cancer Biol. (2024) 104:1–15. doi: 10.1016/j.semcancer.2024.07.001
25. Wang B, Han J, Elisseeff JH, Demaria M. The senescence-associated secretory phenotype and its physiological and pathological implications. Nat Rev Mol Cell Biol. (2024) 12:1–21. doi: 10.1038/s41580-024-00727-x
26. Mebratu YA, Soni S, Rosas L, Rojas M, Horowitz JC, Nho R. The aged extracellular matrix and the profibrotic role of senescence-associated secretory phenotype. Am J Physiology-Cell Physiol. (2023) 325:C565–79.
27. Sun S-C. The non-canonical NF-κB pathway in immunity and inflammation. Nat Rev Immunol. (2017) 17:545–58.
28. Shahar N, Larisch S. Inhibiting the inhibitors: Targeting anti-apoptotic proteins in cancer and therapy resistance. Drug Resist Updates. (2020) 52:100712.
29. Chambers CR, Ritchie S, Pereira BA, Timpson P. Overcoming the senescence-associated secretory phenotype (SASP): a complex mechanism of resistance in the treatment of cancer. Mol Oncol. (2021) 15:3242–55. doi: 10.1002/1878-0261.13042
31. Hazeldine J, Lord JM. The impact of ageing on natural killer cell function and potential consequences for health in older adults. Ageing Res Rev. (2013) 12:1069–78. doi: 10.1016/j.arr.2013.04.003
32. Liu X, Si F, Bagley D, Ma F, Zhang Y, Tao Y, et al. Blockades of effector T cell senescence and exhaustion synergistically enhance antitumor immunity and immunotherapy. J Immunother Cancer. (2022) 10:e005020. doi: 10.1136/jitc-2022-005020
33. Di Mitri D, Alimonti A. Non-cell-autonomous regulation of cellular senescence in cancer. Trends Cell Biol. (2016) 26:215–26. doi: 10.1016/j.tcb.2015.10.005
34. Tagliamento M, Frelaut M, Baldini C, Naigeon M, Nencioni A, Chaput N, et al. The use of immunotherapy in older patients with advanced non-small cell lung cancer. Cancer Treat Rev. (2022) 106:102394. doi: 10.1016/j.ctrv.2022.102394
35. Elias R, Karantanos T, Sira E, Hartshorn KL. Immunotherapy comes of age: immune aging & checkpoint inhibitors. J Geriatric Oncol. (2017) 8:229–35. doi: 10.1016/j.jgo.2017.02.001
36. Derhovanessian E, Solana R, Larbi A, Pawelec G. Immunity, ageing and cancer. Immun Ageing. (2008) 5:1–16. doi: 10.1186/1742-4933-5-11
37. Aghababaei F, Hadidi M. Recent advances in potential health benefits of quercetin. Pharmaceuticals. (2023) 16:1020. doi: 10.3390/ph16071020
38. Wang Y, Chang J, Liu X, Zhang X, Zhang S, Zhang X, et al. Discovery of piperlongumine as a potential novel lead for the development of senolytic agents. Aging (Albany NY). (2016) 8:2915. doi: 10.18632/aging.101100
39. Yousefzadeh MJ, Zhu Y, McGowan SJ, Angelini L, Fuhrmann-Stroissnigg H, Xu M, et al. Fisetin is a senotherapeutic that extends health and lifespan. EBioMedicine. (2018) 36:18–28. doi: 10.1016/j.ebiom.2018.09.015
40. Sundarraj K, Raghunath A, Perumal E. A review on the chemotherapeutic potential of fisetin: In vitro evidences. Biomed Pharmacother. (2018) 97:928–40. doi: 10.1016/j.biopha.2017.10.164
41. Almatroodi SA, Almatroudi A, Khan AA, Alhumaydhi FA, Alsahli MA, Rahmani AH. Potential therapeutic targets of epigallocatechin gallate (EGCG), the most abundant catechin in green tea, and its role in the therapy of various types of cancer. Molecules. (2020) 25:3146. doi: 10.3390/molecules25143146
42. Rameshrad M, Razavi BM, Hosseinzadeh H. Protective effects of green tea and its main constituents against natural and chemical toxins: A comprehensive review. Food Chem Toxicol. (2017) 100:115–37. doi: 10.1016/j.fct.2016.11.035
43. Xia N, Daiber A, Förstermann U, Li H. Antioxidant effects of resveratrol in the cardiovascular system. Br J Pharmacol. (2017) 174:1633–46. doi: 10.1111/bph.v174.12
44. Bielak-Zmijewska A, Grabowska W, Ciolko A, Bojko A, Mosieniak G, Bijoch Ł, et al. The role of curcumin in the modulation of ageing. Int J Mol Sci. (2019) 20:1239. doi: 10.3390/ijms20051239
45. Zhao H, Han Z, Li G, Zhang S, Luo Y. Therapeutic potential and cellular mechanisms of Panax notoginseng on prevention of aging and cell senescence-associated diseases. Aging Dis. (2017) 8:721. doi: 10.14336/AD.2017.0724
46. Hong H, Xiao J, Guo Q, Du J, Jiang Z, Lu S, et al. Cycloastragenol and Astragaloside IV activate telomerase and protect nucleus pulposus cells against high glucose−induced senescence and apoptosis. Exp Ther Med. (2021) 22:1–11. doi: 10.3892/etm.2021.10761
47. Mc Cubrey JA, Lertpiriyapong K, Steelman LS, Abrams SL, Yang LV, Murata RM, et al. Effects of resveratrol, curcumin, berberine and other nutraceuticals on aging, cancer development, cancer stem cells and microRNAs. Aging. (2017) 9:1477–536. doi: 10.18632/aging.v9i6
48. Santín-Márquez R, Alarcón-Aguilar A, López-Diazguerrero NE, Chondrogianni N, Königsberg M. Sulforaphane-role in aging and neurodegeneration. Geroscience. (2019) 41:655–70.
49. Mas-Bargues C, Borras C, Vina J. The multimodal action of genistein in Alzheimer’s and other age-related diseases. Free Radical Biol Med. (2022) 183:127–37. doi: 10.1016/j.freeradbiomed.2022.03.021
50. Zhu Y, Doornebal EJ, Pirtskhalava T, Giorgadze N, Wentworth M, Fuhrmann-Stroissnigg H, et al. New agents that target senescent cells: the flavone, fisetin, and the BCL-XL inhibitors, A1331852 and A1155463. Aging (Albany NY). (2017) 9:955. doi: 10.18632/aging.101202
51. Fuhrmann-Stroissnigg H, Ling YY, Zhao J, McGowan SJ, Zhu Y, Brooks RW, et al. Identification of HSP90 inhibitors as a novel class of senolytics. Nat Commun. (2017) 8:422. doi: 10.1038/s41467-017-00314-z
52. Rauf A, Abu-Izneid T, Imran M, Hemeg HA, Bashir K, Aljohani AS, et al. Therapeutic potential and molecular mechanisms of the multitargeted flavonoid fisetin. Curr Topics Med Chem. (2023) 23:2075–96. doi: 10.2174/1568026623666230710162217
53. Yang Y, Han X, Chen Y, Wu J, Li M, Yang H, et al. EGCG induces pro-inflammatory response in macrophages to prevent bacterial infection through the 67LR/p38/JNK signaling pathway. J Agric Food Chem. (2021) 69:5638–51. doi: 10.1021/acs.jafc.1c01353
54. Moody R, Wilson K, Jaworowski A, Plebanski M. Natural compounds with potential to modulate cancer therapies and self-reactive immune cells. Cancers. (2020) 12:673. doi: 10.3390/cancers12030673
55. Feng G, Jiang Z-y, Sun B, Fu J, Li T-z. Fisetin alleviates lipopolysaccharide-induced acute lung injury via TLR4-mediated NF-κB signaling pathway in rats. Inflammation. (2016) 39:148–57. doi: 10.1007/s10753-015-0233-y
56. Zumerle S, Sarill M, Saponaro M, Colucci M, Contu L, Lazzarini E, et al. Targeting senescence induced by age or chemotherapy with a polyphenol-rich natural extract improves longevity and healthspan in mice. Nat Aging. (2024) 4:1231–48. doi: 10.1038/s43587-024-00663-7
57. Dellinger RW, Santos SR, Morris M, Evans M, Alminana D, Guarente L, et al. (nicotinamide riboside and pterostilbene) increases NAD+ levels in humans safely and sustainably: a randomized, double-blind, placebo-controlled study. NPJ Aging Mech Dis. (2017) 3:17. doi: 10.1038/s41514-017-0016-9
58. Huang M-C, Hsueh TY, Cheng Y-Y, Lin L-C, Tsai T-H. Pharmacokinetics and biliary excretion of fisetin in rats. J Agric Food Chem. (2018) 66:6300–7. doi: 10.1021/acs.jafc.8b00917
59. Chen X, Yin OQ, Zuo Z, Chow MS. Pharmacokinetics and modeling of quercetin and metabolites. Pharm Res. (2005) 22:892–901. doi: 10.1007/s11095-005-4584-1
60. Hu J, Webster D, Cao J, Shao A. The safety of green tea and green tea extract consumption in adults–results of a systematic review. Regul Toxicol Pharmacol. (2018) 95:412–33. doi: 10.1016/j.yrtph.2018.03.019
61. Mazzanti G, Di Sotto A, Vitalone A. Hepatotoxicity of green tea: an update. Arch Toxicol. (2015) 89:1175–91. doi: 10.1007/s00204-015-1521-x
62. Negut I, Bita B. Polymersomes as innovative, stimuli-responsive platforms for cancer therapy. Pharmaceutics. (2024) 16:463. doi: 10.3390/pharmaceutics16040463
63. de Mera-Rodríguez JA, Álvarez-Hernán G, Gañán Y, Martín-Partido G, Rodríguez-León J, Francisco-Morcillo J. Is senescence-associated β-galactosidase a reliable in vivo marker of cellular senescence during embryonic development? Front Cell Dev Biol. (2021) 9:623175.
64. Song S, Lam EW-F, Tchkonia T, Kirkland JL, Sun Y. Senescent cells: emerging targets for human aging and age-related diseases. Trends Biochem Sci. (2020) 45:578–92. doi: 10.1016/j.tibs.2020.03.008
65. Lee H, Dey D, Kim K, Kim S, Kim E, Kang S, et al. Hypoxia-responsive nanomedicine to overcome tumor microenvironment-mediated resistance to chemo-photodynamic therapy. Mater Today Adv. (2022) 14:100218. doi: 10.1016/j.mtadv.2022.100218
66. Li Z, Huang J, Wu J. pH-Sensitive nanogels for drug delivery in cancer therapy. Biomater Sci. (2021) 9:574–89. doi: 10.1039/D0BM01729A
67. Kuang T, Liu Y, Gong T, Peng X, Hu X, Yu Z. Enzyme-responsive nanoparticles for anticancer drug delivery. Curr Nanosci. (2016) 12:38–46. doi: 10.2174/1573413711666150624170518
68. Shi Y, Zhang Y, Zhang Y, Yao J, Guo J, Xu X, et al. Advances in nanotherapy for targeting senescent cells. Int J Nanomed. (2024) 19:8797–813. doi: 10.2147/IJN.S469110
69. Bonechi C, Donati A, Tamasi G, Leone G, Consumi M, Rossi C, et al. Protective effect of quercetin and rutin encapsulated liposomes on induced oxidative stress. Biophys Chem. (2018) 233:55–63. doi: 10.1016/j.bpc.2017.11.003
70. Kureel SK, Blair B, Sheetz MP. Recent advancement in elimination strategies and potential rejuvenation targets of senescence. Adv Biol. (2024) 8:2300461. doi: 10.1002/adbi.202300461
71. Suryadevara V, Hudgins AD, Rajesh A, Pappalardo A, Karpova A, Dey AK, et al. SenNet recommendations for detecting senescent cells in different tissues. Nat Rev Mol Cell Biol. (2024) 25:1–23. doi: 10.1038/s41580-024-00738-8
72. Zhang J, Lin Y, Lin Z, Wei Q, Qian J, Ruan R, et al. Stimuli-responsive nanoparticles for controlled drug delivery in synergistic cancer immunotherapy. Adv Sci. (2022) 9:2103444. doi: 10.1002/advs.202103444
73. Torchilin VP. Multifunctional, stimuli-sensitive nanoparticulate systems for drug delivery. Nat Rev Drug Discovery. (2014) 13:813–27. doi: 10.1038/nrd4333
74. Shen S, Li T, Fan J, Shao Q, Dong H, Xu X, et al. Lipid-polymer hybrid nanoparticle with cell-distinct drug release for treatment of stemness-derived resistant tumor. Acta Pharm Sin B. (2023) 13:1262–73. doi: 10.1016/j.apsb.2022.11.009
75. Fan T, Du Y, Zhang M, Zhu AR, Zhang J. Senolytics cocktail dasatinib and quercetin alleviate human umbilical vein endothelial cell senescence via the TRAF6-MAPK-NF-κB axis in a YTHDF2-dependent manner. Gerontology. (2022) 68:920–34. doi: 10.1159/000522656
76. Singh BN, Shankar S, Srivastava RK. Green tea catechin, epigallocatechin-3-gallate (EGCG): mechanisms, perspectives and clinical applications. Biochem Pharmacol. (2011) 82:1807–21. doi: 10.1016/j.bcp.2011.07.093
77. Rentschler M, Braumüller H, Briquez PS, Wieder T. Cytokine-induced senescence in the tumor microenvironment and its effects on anti-tumor immune responses. Cancers. (2022) 14:1364. doi: 10.3390/cancers14061364
78. Afroze N, Pramodh S, Shafarin J, Bajbouj K, Hamad M, Sundaram MK, et al. Fisetin deters cell proliferation, induces apoptosis, alleviates oxidative stress and inflammation in human cancer cells, HeLa. Int J Mol Sci. (2022) 23:1707. doi: 10.3390/ijms23031707
79. Stevens Barrón J, Chapa González C, Álvarez Parrilla E, de la Rosa L. Nanoparticle-mediated delivery of flavonoids: impact on proinflammatory cytokine production: A systematic review. Biomolecules. (2023) 13:1158. doi: 10.3390/biom13071158
80. Liao Y, Xie X, Zhang C, Zhong H, Shan L, Yu P, et al. Quercetin exerts anti-tumor immune mechanism by regulating IL-6/JAK2/STAT3 signaling pathway to deplete Treg cells. Toxicon. (2024) 243:107747. doi: 10.1016/j.toxicon.2024.107747
81. Serati-Nouri H, Rasoulpoor S, Pourpirali R, Sadeghi-Soureh S, Esmaeilizadeh N, Dadashpour M, et al. In vitro expansion of human adipose-derived stem cells with delayed senescence through dual stage release of curcumin from mesoporous silica nanoparticles/electrospun nanofibers. Life Sci. (2021) 285:119947. doi: 10.1016/j.lfs.2021.119947
82. Guo Z, Zhang Y, Gong Y, Li G, Pan J, Dou D, et al. Antibody functionalized curcuma-derived extracellular vesicles loaded with doxorubicin overcome therapy-induced senescence and enhance chemotherapy. J Controlled Release. (2025) 379:377–89. doi: 10.1016/j.jconrel.2025.01.029
83. Budi HS, Farhood B. Tumor microenvironment remodeling in oral cancer: Application of plant derived-natural products and nanomaterials. Environ Res. (2023) 233:116432. doi: 10.1016/j.envres.2023.116432
84. Hieber C, Grabbe S, Bros M. Counteracting immunosenescence—Which therapeutic strategies are promising? Biomolecules. (2023) 13:1085.
85. Han X, Zhang G, Wu X, Xu S, Liu J, Wang K, et al. Microfluidics-enabled fluorinated assembly of EGCG-ligands-siTOX nanoparticles for synergetic tumor cells and exhausted t cells regulation in cancer immunotherapy. J Nanobiotechnol. (2024) 22:90. doi: 10.1186/s12951-024-02328-4
86. Payne A, Nahashon S, Taka E, Adinew GM, Soliman KF. Epigallocatechin-3-Gallate (EGCG): New therapeutic perspectives for neuroprotection, aging, and neuroinflammation for the modern age. Biomolecules. (2022) 12:371. doi: 10.3390/biom12030371
87. Wan X, Wang D. Curcumin: epigenetic modulation and tumor immunity in antitumor therapy. Planta Med. (2025). doi: 10.1055/a-2499-1140. [Epub ahead of print]
88. Liang Y, Liu Z-Y, Wang P-Y, Li Y-J, Wang R-R, Xie S-Y. Nanoplatform-based natural products co-delivery system to surmount cancer multidrug-resistant. J Controlled Release. (2021) 336:396–409. doi: 10.1016/j.jconrel.2021.06.034
89. Pi C, Zhao W, Zeng M, Yuan J, Shen H, Li K, et al. Anti-lung cancer effect of paclitaxel solid lipid nanoparticles delivery system with curcumin as co-loading partner in vitro and in vivo. Drug Deliv. (2022) 29:1878–91. doi: 10.1080/10717544.2022.2086938
90. Lewinska A, Adamczyk-Grochala J, Bloniarz D, Olszowka J, Kulpa-Greszta M, Litwinienko G, et al. AMPK-mediated senolytic and senostatic activity of quercetin surface functionalized Fe3O4 nanoparticles during oxidant-induced senescence in human fibroblasts. Redox Biol. (2020) 28:101337. doi: 10.1016/j.redox.2019.101337
91. Syed DN, M Adhami V, Imran Khan M, Mukhtar H. Inhibition of Akt/mTOR signaling by the dietary flavonoid fisetin. Anti-Cancer Agents Med Chem. (2013) 13:995–1001. doi: 10.2174/18715206113139990129
92. Granja A, Pinheiro M, Reis S. Epigallocatechin gallate nanodelivery systems for cancer therapy. Nutrients. (2016) 8:307. doi: 10.3390/nu8050307
93. Wang Y, Yu H, Wang S, Gai C, Cui X, Xu Z, et al. Targeted delivery of quercetin by nanoparticles based on chitosan sensitizing paclitaxel-resistant lung cancer cells to paclitaxel. Mater Sci Eng: C. (2021) 119:111442. doi: 10.1016/j.msec.2020.111442
94. Caro C, Pourmadadi M, Eshaghi MM, Rahmani E, Shojaei S, Paiva-Santos AC, et al. Nanomaterials loaded with Quercetin as an advanced tool for cancer treatment. J Drug Deliv Sci Technol. (2022) 78:103938. doi: 10.1016/j.jddst.2022.103938
95. Gera M, Kim N, Ghosh M, Sharma N, Chandimali N, Koh H, et al. Synthesis and evaluation of the antiproliferative efficacy of BRM270 phytocomposite nanoparticles against human hepatoma cancer cell lines. Mater Sci Eng: C. (2019) 97:166–76. doi: 10.1016/j.msec.2018.11.055
96. Talib WH, Awajan D, Alqudah A, Alsawwaf R, Althunibat R, Abu AlRoos M, et al. Targeting cancer hallmarks with epigallocatechin gallate (EGCG): mechanistic basis and therapeutic targets. Molecules. (2024) 29:1373. doi: 10.3390/molecules29061373
97. Jatal R. Novel Therapeutic strategies that interfere with senescence escape for the treatment of drug resistance in colorectal and breast cancer. Université d’Angers; Universidade de Santiago de Compostela (2021).
98. Baksi R, Singh DP, Borse SP, Rana R, Sharma V, Nivsarkar M. In vitro and in vivo anticancer efficacy potential of Quercetin loaded polymeric nanoparticles. Biomed Pharmacother. (2018) 106:1513–26. doi: 10.1016/j.biopha.2018.07.106
99. Danciu C, Soica C, Antal D, Alexa E, Pavel IZ, Ghiulai R, et al. Natural compounds in the chemoprevention of Malignant melanoma. Anti-Cancer Agents Med Chem. (018) 18:631–44. doi: 10.2174/1871520617666171121142522
100. Pinheiro RG, Pinheiro M, Neves AR. Nanotechnology innovations to enhance the therapeutic efficacy of quercetin. Nanomaterials. (2021) 11:2658. doi: 10.3390/nano11102658
101. Hussain Z, Thu HE, Haider M, Khan S, Sohail M, Hussain F, et al. A review of imperative concerns against clinical translation of nanomaterials: Unwanted biological interactions of nanomaterials cause serious nanotoxicity. J Drug Deliv Sci Technol. (2020) 59:101867. doi: 10.1016/j.jddst.2020.101867
103. Wagner V, Dullaart A, Bock A-K, Zweck A. The emerging nanomedicine landscape. Nat Biotechnol. (2006) 24:1211–7. doi: 10.1038/nbt1006-1211
104. Kanai M, Otsuka Y, Otsuka K, Sato M, Nishimura T, Mori Y, et al. A phase I study investigating the safety and pharmacokinetics of highly bioavailable curcumin (Theracurmin®) in cancer patients. Cancer Chemother Pharmacol. (2013) 71:1521–30. doi: 10.1007/s00280-013-2151-8
105. Thomas R, Williams M, Sharma H, Chaudry A, Bellamy P. A double-blind, placebo-controlled randomised trial evaluating the effect of a polyphenol-rich whole food supplement on PSA progression in men with prostate cancer—the UK NCRN Pomi-T study. Prostate Cancer Prostatic Dis. (2014) 17:180–6. doi: 10.1038/pcan.2014.6
106. Ji H, Wang W, Li X, Han X, Zhang X, Wang J, et al. Natural small molecules enabled efficient immunotherapy through supramolecular self-assembly in P53-mutated colorectal cancer. ACS Appl Mater Interfaces. (2022) 14:2464–77. doi: 10.1021/acsami.1c16737
107. Sasia C, Borgonetti V, Mancini C, Lori G, Arbiser JL, Taddei ML, et al. The neolignan honokiol and its synthetic derivative honokiol hexafluoro reduce neuroinflammation and cellular senescence in microglia cells. Cells. (2024) 13:1652. doi: 10.3390/cells13191652
108. Qiu N, Du X, Ji J, Zhai G. A review of stimuli-responsive polymeric micelles for tumor-targeted delivery of curcumin. Drug Dev Ind Pharm. (2021) 47:839–56. doi: 10.1080/03639045.2021.1934869
109. Yan X, Zhang X, McClements DJ, Zou L, Liu X, Liu F. Co-encapsulation of epigallocatechin gallate (EGCG) and curcumin by two proteins-based nanoparticles: Role of EGCG. J Agric Food Chem. (2019) 67:13228–36. doi: 10.1021/acs.jafc.9b04415
110. Jawed Khan M, Hafeez A, Aftab Siddiqui M. Nanocarrier based delivery of berberine: a critical review on pharmaceutical and preclinical characteristics of the bioactive. Curr Pharm Biotechnol. (2023) 24:1449–64.
111. Tyagi N, Song YH, De R. Recent progress on biocompatible nanocarrier-based genistein delivery systems in cancer therapy. J Drug Targeting. (2019) 27:394–407. doi: 10.1080/1061186X.2018.1514040
112. Bahmani A, Taghvaei A, Firozian F, Chehardoli G. Folic acid as an exploiter of natural endocytosis pathways in drug delivery. Chem Methodol. (2024) 8:96–122.
113. Behera A, Padhi S. Passive and active targeting strategies for the delivery of the camptothecin anticancer drug: a review. Environ Chem Lett. (2020) 18:1557–67. doi: 10.1007/s10311-020-01022-9
114. Varshosaz J, Taymouri S, Ghassami E. Supramolecular self-assembled nanogels a new platform for anticancer drug delivery. Curr Pharm Design. (2017) 23:5242–60.
115. Raspantini GL, Luiz MT, Abriata JP, de Oliveira Eloy J, Vaidergorn MM, da Silva Emery F, et al. PCL-TPGS polymeric nanoparticles for docetaxel delivery to prostate cancer: Development, physicochemical and biological characterization. Colloids Surfaces A: Physicochem Eng Aspects. (2021) 627:127144. doi: 10.1016/j.colsurfa.2021.127144
116. Zhong H, Luo X, Abdullah, Liu X, Hussain M, Guan R. Nano-targeted delivery system: a promising strategy of anthocyanin encapsulation for treating intestinal inflammation. Crit Rev Food Sci Nutr. (2025), 1–22. doi: 10.1080/10408398.2025.2458741
117. Suri S, Mirza MA, Anwer MK, Alshetaili AS, Alshahrani SM, Ahmed FJ, et al. Development of NIPAAm-PEG acrylate polymeric nanoparticles for co-delivery of paclitaxel with ellagic acid for the treatment of breast cancer. J Polymer Eng. (2019) 39,:271–8. doi: 10.1515/polyeng-2018-0169
118. Jafari S, Bakhshaei A, Eskandani M, Molavi O. Silibinin-loaded nanostructured lipid carriers for growth inhibition of cisplatin-resistant ovarian cancer cells. ASSAY Drug Dev Technol. (2022) 20,:339–48. doi: 10.1089/adt.2022.060
119. Srivastava S, Kumar A, Yadav PK, Kumar M, Mathew J, Pandey AC, et al. Formulation and performance evaluation of polymeric mixed micelles encapsulated with baicalein for breast cancer treatment. Drug Dev Ind Pharm. (2021) 47,:1512–22. doi: 10.1080/03639045.2021.2007394
120. Wang K, Shang J, Tao C, Huang M, Wei D, Yang L, et al. Advancements in betulinic acid-loaded nanoformulations for enhanced anti-tumor therapy. Int J Nanomed. (2024) 19:14075–103. doi: 10.2147/IJN.S493489
121. Ren S, Song L, Tian Y, Zhu L, Guo K, Zhang H, et al. Emodin-conjugated PEGylation of Fe3O4 nanoparticles for FI/MRI dual-modal imaging and therapy in pancreatic cancer. Int J Nanomed. (2021) 16:7463–78. doi: 10.2147/IJN.S335588
122. Wang Y, Liang X, Tong R, Yang J, Yang Y, Zhong J, et al. Gambogic acid-loaded polymeric micelles for improved therapeutic effect in breast cancer. J Biomed Nanotechnol. (2018) 14:1695–704. doi: 10.1166/jbn.2018.2626
123. Yang X, Qiu Y, Fang F, Cao Z, Fu H, Chen X, et al. Isoliquiritigenin-loaded platinum (IV) prodrug micelles induce sustained endoplasmic reticulum stress for promoting cisplatin chemosensitivity in ovarian cancer. ACS Mater Lett. (2024) 6:5275–84. doi: 10.1021/acsmaterialslett.4c01192
124. Zheng Y, Deng J, Li L, Cao J, Xie Q, Fu Y. Adenosine monophosphate modified nanomicelles for precise delivery of magnolol enhances chemosensitivity in cancer therapy. J Drug Deliv Sci Technol. (2025) 104:106559. doi: 10.1016/j.jddst.2024.106559
125. Yıldırım M, Acet Ö, Yetkin D, Acet BÖ, Karakoc V, Odabası M. Anti-cancer activity of naringenin loaded smart polymeric nanoparticles in breast cancer. J Drug Deliv Sci Technol. (2022) 74:103552. doi: 10.1016/j.jddst.2022.103552
Keywords: nanoparticles, herbal-derived natural products, immunosenescence, drug resistance, cancer therapy, tumor microenvironment
Citation: Yang L, Wu W, Yang J and Xu M (2025) Nanoparticle-mediated delivery of herbal-derived natural products to modulate immunosenescence-induced drug resistance in cancer therapy: a comprehensive review. Front. Oncol. 15:1567896. doi: 10.3389/fonc.2025.1567896
Received: 28 January 2025; Accepted: 28 March 2025;
Published: 28 April 2025.
Edited by:
Arumugam Jayakumar, University of Texas MD Anderson Cancer Center, United StatesReviewed by:
Anna Calabrò, University of Palermo, ItalyAnkush Paladhi, Indian Institute of Technology Bombay, India
Copyright © 2025 Yang, Wu, Yang and Xu. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Wei Wu, d3cyMjM4NDFAMTYzLmNvbQ==; Manman Xu, eHVtbWpvdXJuYWxAMTYzLmNvbQ==; Jing Yang, TWF5b3V5b3U2Njg4QDE2My5jb20=