 Klaudia Skóra1,2*
Klaudia Skóra1,2* Damian Strojny2,3,4
Damian Strojny2,3,4 Dawid Sobański2,5
Dawid Sobański2,5 Rafał Staszkiewicz2,6,7
Rafał Staszkiewicz2,6,7 Kamil Bryś7Paweł Gogol2,8,9,10Krzysztof Bereza11,12Michalina Masternak13
Kamil Bryś7Paweł Gogol2,8,9,10Krzysztof Bereza11,12Michalina Masternak13 Beniamin Oskar Grabarek2,14
Beniamin Oskar Grabarek2,14- 1Department of Neurological Rehabilitation, District Hospital of St. Padre Pio in Sędziszów Małopolski, Sędziszów Małopolski, Poland
- 2Collegium Medicum, WSB University, Dąbrowa Górnicza, Poland
- 3Department of Neurology, New Medical Techniques Specialist Hospital of St. Family in Rudna Mała, Rzeszow, Poland
- 4Institute of Health Care, National Academy of Applied Sciences in Przemyśl, Przemyśl, Poland
- 5Department of Neurosurgery, St. Raphael Hospital, Krakow, Poland
- 6Department of Neurosurgery, 5th Military Clinical Hospital with the SP ZOZ Polyclinic in Krakow, Krakow, Poland
- 7Department of Neurosurgery, Faculty of Medicine in Zabrze, Academy of Silesia in Katowice, Katowice, Poland
- 8Department of Anesthesiology and Intensive Care, Our Lady of Perpetual Help Hospital in Wołomin, Wołomin, Poland
- 9Department of Trauma and Orthopedic Surgery, Our Lady of Perpetual Help Hospital in Wołomin, Wołomin, Poland
- 10Pain Treatment Clinic, Our Lady of Perpetual Help Hospital in Wołomin, Wołomin, Poland
- 11Department of Mother and Child Health, Faculty of Health Sciences, Institute of Nursing and Midwifery, Jagiellonian University Medical College, Krakow, Poland
- 12Department of Brachytherapy, Jagiellonian University Medical College, Krakow, Poland
- 13Silesian Center for Rehabilitation and Manual Therapy Revita in Mysłowice, Mysłowice, Poland
- 14Faculty of Medicine and Health Sciences, Andrzej Frycz Modrzewski University, Cracow, Poland
Background: Astrocytic tumors, particularly glioblastomas, are aggressive brain malignancies with poor prognosis. Transforming growth factor-beta (TGF-β) isoforms—TGF-β-1, TGF-β-2, and TGF-β-3—play critical roles in glioma progression, yet their isoform-specific expression patterns and regulatory mechanisms remain incompletely defined. This study aimed to evaluate the differential expression of TGF-β isoforms and their regulation by epigenetic mechanisms and microRNAs (miRNAs) across astrocytic tumor grades.
Methods: Sixty-five astrocytic tumor samples (WHO grades 2-4) were analyzed. Gene and protein expression of TGF-β-1, -2, and -3 were assessed using reverse transcription quantitative polymerase chain reaction (RT-qPCR), enzyme-linked immunosorbent assay (ELISA), and immunohistochemistry (IHC). Promoter methylation was analyzed using methylation-specific PCR (MSPCR). Differentially expressed regulatory miRNAs were identified by microarray and in silico target prediction. Survival associations were evaluated by Kaplan–Meier and Cox regression analyses.
Results: TGF-β-1 and TGF-β-3 were significantly upregulated in high-grade astrocytomas (p < 0.05), whereas TGF-β-2 showed no consistent changes. TGF-β-3 expression strongly correlated with poor survival (Exp(B) = 1.02644, p < 0.0001), while TGF-β-1 showed a weaker, non-significant association. Among regulatory miRNAs, hsa-miR-2278 (targeting TGF-β-3) was upregulated and significantly associated with worse survival (Exp(B) = 1.437, p = 0.008), while hsa-miR-3196 (targeting TGF-β-1) was downregulated and trended toward better prognosis (Exp(B) = 0.8897, p = 0.076).
Conclusion: TGF-β-3 is a potent prognostic biomarker in astrocytic tumors and a promising candidate for targeted therapeutic intervention. Regulatory miRNAs such as hsa-miR-2278 and hsa-miR-3196 may serve as molecular modulators of TGF-β signaling and potential adjuncts in personalized glioma therapy. These findings warrant further investigation into miRNA-based therapeutics targeting the TGF-β axis in high-grade gliomas.
1 Introduction
Astrocytic tumors, originating from astrocytes, are among the most prevalent brain tumors and are classified into four grades based on histological characteristics and severity (1–3). Despite advances in clinical research, the prognosis for these tumors remains poor. Patients with low-grade gliomas (LGGs) (grades II and III) have a median survival of 5–10 years, whereas those with high-grade gliomas (grade IV) typically survive only 1–2 years (4). Glioblastoma multiforme (GBM), the most common and aggressive grade IV astrocytic tumor, is associated with a particularly poor prognosis (5, 6).
The integration of molecular biology into neuro-oncology has significantly refined the classification of brain tumors. The World Health Organization (WHO) revised its classification system in 2016, incorporating genomic profiling and epigenetic changes (7). The latest 2021 WHO classification further underscores the increasing importance of molecular diagnostics, introducing a distinction between grade IV astrocytoma and glioblastoma, both of which were previously grouped under the same category (8–10). As part of this refinement, some tumors previously classified as grade 3 astrocytomas have been redefined as glioblastoma grade 4 based on their molecular features. These advancements highlight the critical role of molecular profiling in diagnosing and classifying astrocytic tumors (8–10).
Molecular biomarkers play a vital role in determining prognosis and guiding treatment strategies (11). Key markers include isocitrate dehydrogenase (IDH1/2) mutations, MGMT promoter methylation, 1p/19q co-deletion, and epidermal growth factor receptor (EGFR) amplification. Among these, IDH mutations are particularly significant, as they are associated with improved survival across all glioma grades. For example, patients with IDH-mutated grade IV astrocytomas have a median survival of approximately 31 months, whereas those with IDH wild-type tumors have a shorter median survival of 15 months (12–14).
Transforming growth factor-beta (TGFβ) is a multifunctional cytokine family that includes six isoforms, three of which—TGFβ1, TGFβ2, and TGFβ3—are present in humans. These isoforms share considerable sequence similarity (71–79%) but are encoded by distinct genes. Their biological activity is highly dependent on the relative expression of each isoform, influencing various cellular processes (15). TGFβ plays a crucial role in both physiological and pathological conditions by regulating key cellular functions such as growth, differentiation, inflammation, and tissue repair (16). As an anti-inflammatory cytokine, TGFβ is secreted by immune cells following injury, modulating immune responses and promoting healing (17). Studies by Cekanaviciute et al. have shown that in response to Toxoplasma gondii infection, TGFβ signaling is activated in astrocytes, which helps control neuroinflammation. Conversely, inhibition of TGFβ signaling leads to excessive immune infiltration, increased secretion of pro-inflammatory cytokines and chemokines, and neuronal damage (18). Beyond its immunoregulatory role, TGFβ is integral to nervous system function. Among its isoforms, TGFβ1 is the most abundant and is particularly involved in astrocyte-mediated scar formation following brain injury (19). Experimental studies have demonstrated that TGFβ upregulates neurocan, a chondroitin sulfate proteoglycan that contributes to glial scar development (20). TGFβ signaling is implicated in multiple cancers, including lung, breast, pancreatic, colorectal, and melanoma (21). It has also been extensively studied in gliomas, yet the specific interactions between TGFβ1, TGFβ2, and TGFβ3 isoforms in astrocytic tumors remain poorly understood (22, 23).
TGFβ is also a key player in tumorigenesis, particularly in gliomas. Its role in cancer is complex and often described as the “TGFβ paradox.” (22, 24). In early tumor development, TGFβ functions as a tumor suppressor by inhibiting cell proliferation, inducing differentiation, promoting apoptosis and autophagy, and limiting angiogenesis and inflammation (25). However, in advanced cancer stages, TGFβ facilitates tumor progression by promoting extracellular matrix remodeling, enhancing angiogenesis, and creating an immunosuppressive tumor microenvironment that enables metastasis (24, 26, 27). Clinically, this dual role is significant—elevated TGFβ levels correlate with better prognosis in early-stage tumors but are linked to increased aggressiveness, invasiveness, and poorer outcomes in advanced malignancies. This underscores the potential of TGFβ as both a biomarker and a therapeutic target (28–30).
A study by Naik et al. explored the relationship between epigenetic factors and TGF-β signaling pathways (31). The authors concluded that dynamic epigenetic modifications are essential for determining cancer cell behavior, influencing tumor microenvironment interactions, and affecting the overall carcinogenesis process. Their analysis revealed complex regulatory networks in tumors, involving long non-coding RNAs (lncRNAs), microRNAs (miRNAs), and post-translational histone modifications, all of which are closely linked to TGF-β signaling (31). Similarly, Ding et al. demonstrated that epigenetic regulation of TGF-β, including chromatin remodeling, non-coding RNA regulation, DNA methylation, and histone modifications, not only contributes to tumor cell formation and growth but also affects the response to radiotherapy (32). This study highlights that understanding the impact of epigenetics is a key aspect in both the diagnosis and treatment of cancer patients.
Methylation profiling has emerged as an essential tool in brain tumor classification, complementing histological evaluation for more accurate tumor identification. The integration of histopathology and molecular techniques is expected to enhance prognostic accuracy and improve patient management. In some cases, molecular alterations can justify classifying a tumor as malignant, even if histological features suggest a lower grade. This shift reflects the growing reliance on molecular markers in neuro-oncology and the potential for personalized therapeutic strategies (33).
MicroRNAs (miRNAs) are short, non-coding RNA molecules that play a crucial role in gene expression regulation by blocking or degrading mRNA (34). MiRNAs regulate gene expression by targeting mRNA and binding directly to complementary sites in the 3′ untranslated region (3′ UTR) of the target mRNA (35). As a result, miRNAs can regulate multiple mRNA targets involved in various biological processes, including DNA damage repair, apoptosis, proliferation, cell cycle regulation, senescence, invasiveness, and angiogenesis (36). Experimental studies confirm that several miRNAs act as modulators of TGF-β signaling at multiple levels by targeting ligands, receptors, R-Smad, co-Smad, I-Smad, and non-Smad pathway components, as well as downstream targets of TGF-β signaling (37).
Therefore, the study aimed was to evaluate variances in the expression patterns of TGFβ1–3 in astrocytic tumors with respect to the degree of malignancy.
2 Materials and methods
2.1 Patient selection and clinical sampling
This study involved the collection of astrocytic brain tumor specimens from 65 patients undergoing surgical resection. The procedures were conducted at two neurosurgical centers in Krakow, Poland: the Department of Neurosurgery at the 5th Military Clinical Hospital with the SP ZOZ Polyclinic and the Department of Neurosurgery at Szpital św. Rafała. Of the 65 patients included in the study, 35 were female and 30 were male, with an average age of approximately 56 to 59 years across different malignancy grades. Among female patients, 10 had G2 tumors, 7 had G3 tumors, and 18 had G4 tumors. In the male group, 7 had G2 tumors, 5 had G3 tumors, and 18 had G4 tumors. To ensure standardized preoperative conditions, all participants followed a fasting regimen, consuming their final meal at 6 PM the evening before surgery. This was in accordance with the hospitals’ nutritional policy, which schedules the last meal at this time to prepare patients for surgery the following morning. All procedures were performed between 8 AM and 11 AM, and only elective surgeries were included in the study.
Eligibility for participation was determined based on predefined criteria. Patients qualified for the study if they were scheduled for astrocytic tumor resection at one of the designated hospitals and provided informed consent. Individuals with additional neoplastic conditions were excluded.
The inclusion criteria were (1): histopathologically confirmed astrocytic brain tumor (G2–G4) (2); planned elective surgical resection (3); age ≥18 years; and (4) signed informed consent.
The exclusion criteria were (1): presence of other primary or metastatic tumors (2); emergency surgical intervention (3); incomplete neuroimaging; and (4) insufficient tissue quality for molecular analysis.
The preliminary diagnosis of astrocytic tumors was established through contrast-enhanced computed tomography (CT) and further confirmed using magnetic resonance imaging (MRI). The MRI protocol included T1- and T2-weighted sequences, fluid-attenuated inversion recovery (FLAIR) sequences, and, when necessary, diffusion tensor imaging. In cases where the tumor was located near eloquent brain regions, additional imaging techniques such as functional MRI and diffusion MRI tractography were employed to assist with neuronavigation.
Surgical resection aimed for maximal tumor removal while minimizing damage to adjacent healthy tissue. Intraoperative techniques included neuronavigation, fluorescence-guided surgery using 5-aminolevulinic acid (5-ALA) for grade IV tumors, and direct cortical stimulation for lesions in proximity to the sensorimotor cortex. The definitive diagnosis was established through histopathological evaluation, classifying tumor malignancy according to the World Health Organization (WHO) grading system (8–10).
2.2 Extraction of total ribonucleic acid
Tissue samples were homogenized using a T18 Digital Ultra-Turrax handheld rotor-stator homogenizer (IKA Poland Ltd., Warsaw, Poland). Total RNA was extracted with TRIzol reagent (Invitrogen Life Technologies, Carlsbad, CA, USA), following the manufacturer’s protocol. To further purify the RNA and remove contaminants, the RNeasy mini kit (QIAGEN, Hilden, Germany) was employed. Additionally, DNase I treatment (Fermentas International Inc., Burlington, ON, Canada) was applied to eliminate any residual genomic DNA.
RNA quality was assessed through 1% agarose gel electrophoresis with ethidium bromide staining (0.5 mg/mL) to verify integrity. RNA concentration was determined by measuring absorbance at 260 nm, ensuring accurate quantification of sample yield and purity.
2.3 Microarray analysis of gene expression
A comparative analysis of circadian clock-related gene expression in tumor tissues versus control tissues was conducted using the HG-U 133_A2 microarray platform (Affymetrix, Santa Clara, CA, USA) and the GeneChip™ 3′ IVT PLUS reagent kit (Affymetrix; Catalog Number 902416). The study strictly adhered to the manufacturer’s protocols and methodologies established in previous research (38).
2.4 MicroRNA profiling and target prediction
To investigate the role of circadian clock-related microRNAs (miRNAs) and their influence on gene expression, a microarray analysis was performed using the GeneChip miRNA 2.0 Array (Affymetrix). This commercial platform ensures high precision and reliability in detecting differentially expressed miRNAs between tumor and control tissues. The microarray profiling process was conducted in strict accordance with the manufacturer’s instructions to maintain standardization and reproducibility. Identified miRNAs that demonstrated differential expression were further analyzed using two widely recognized databases—TargetScan (http://www.targetscan.org/) (39) and miRanda (http://mirdb.org)— (40) to predict their interactions with messenger RNAs (mRNAs). Predicted targets with a confidence score exceeding 80 were considered highly reliable, indicating strong miRNA–mRNA interactions. Conversely, predictions with scores below 60 required further validation to confirm their authenticity (40, 41).
2.5 Validation of gene expression by quantitative reverse-transcription polymerase chain reaction
To validate the microarray data, qRT-PCR was performed on selected genes using the SensiFast SYBR No-ROX One-Step kit (Bioline, London, UK). The thermal cycling protocol included reverse transcription at 45°C, polymerase activation at 95°C for 2 minutes, followed by 40 cycles of denaturation at 95°C for 5 seconds, annealing at 60°C for 10 seconds, and elongation at 72°C for 5 seconds. Gene expression levels were analyzed using the 2−ΔΔCt method, where a fold change of 1 represented the control group, values greater than 1 indicated overexpression, and values below 1 signified gene silencing. To ensure accuracy and consistency, glyceraldehyde 3-phosphate dehydrogenase (GAPDH) was used as an internal control for normalization. Detailed primer sequences are provided in Table 1 for reference.

Table 1. Nucleotide sequence of the primers used in RT-qPCR for TGF-β-1–3 and GAPDH.
2.6 DNA Methylation analysis by methylation-specific PCR
CpG island locations within the selected gene sequences were identified using the MethPrimer program (http://www.urogene.org/cgi-bin/methprimer/methprimer.cgi; accessed January 19, 2025) (42). Primer design followed strict criteria, including a CpG island length exceeding 100 nucleotides, a GC content above 50%, and an observed-to-expected ratio greater than 0.6 (Table 2).
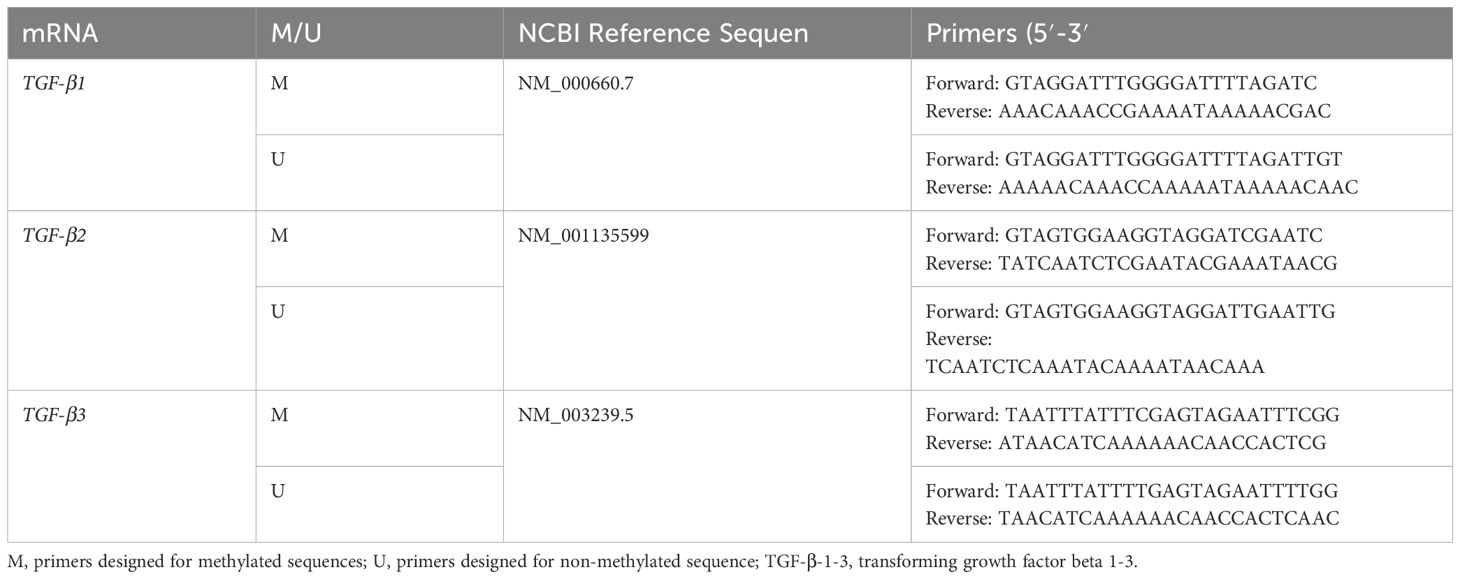
Table 2. Characteristics of primers designed for the MSP.
To assess DNA methylation status, sodium bisulfite conversion was performed according to the manufacturer’s recommendations, followed by sample purification. Methylation-specific PCR (MSP) was carried out using the QuantiTect SYBR Green PCR Kit (Qiagen GmbH, Hilden, Germany) with designed primers. The thermal cycling protocol consisted of an initial denaturation at 95°C for 5 minutes, followed by 40 cycles of 30 seconds each at 94°C (denaturation), 65°C (annealing), and 72°C (elongation).
For further analysis, the methylation status of specific genes, including irisin, ghrelin, and titin, was evaluated by electrophoresis of PCR products on a 1% agarose gel containing ethidium bromide (0.5 µg/mL) in 1x TBE buffer at 120 V. Fragment sizes were assessed using the pBR322/HaeIII size marker. To confirm the specificity of amplification, control samples consisting of methylated and non-methylated DNA were included, utilizing the EpiTect Control DNA set (Qiagen GmbH, Hilden, Germany).
2.7 Protein quantification by enzyme-linked immunosorbent assay and western blot
The expression levels of TGF-β-1, TGF-β-2, and TGF-β-3 were quantified using both enzyme-linked immunosorbent assay (ELISA) and Western blot analysis following electrophoretic separation in polyacrylamide gel. Polyclonal antibodies specific to each TGF-β isoform were used according to the manufacturer’s protocols: anti-TGF-β-1 (bs-0086R, STI, Poznań, Poland; dilution 1:1000), anti-TGF-β-2 (bs-20412R, STI, Poznań, Poland; dilution 1:1000), and anti-TGF-β-3 (bs-0099R, STI, Poznań, Poland; dilution 1:1000). Glyceraldehyde-3-phosphate dehydrogenase (GAPDH) (sc-47724, Santa Cruz Biotechnology, Dallas, TX, USA; dilution 1:500) was used as a loading control. For signal detection, a horseradish peroxidase (HRP)-conjugated goat anti-rabbit IgG secondary antibody (Bio-Rad, Milan, Italy; catalog number 1706515; dilution 1:3000) was applied.
Absorbance measurements for ELISA were performed at 540 nm using an M200PRO microplate reader (Tecan, Männedorf, Switzerland). A detailed description of the ELISA and Western blot procedures is available in our previous publications (38).
To validate assay performance, recombinant TGF-β proteins were used as positive controls for each isoform. These included recombinant TGF-β-1 (MBS2122438), TGF-β-2 (MBS2153787), and TGF-β-3 (MBS2086791), all purchased from MyBioSource Inc. (San Diego, CA, USA). A detailed description of the ELISA and Western blot protocols can be found in previous studies (38).
2.8 Immunohistochemical detection of TGF-β isoforms
Tissue specimens were sectioned into 8.0 µm-thick slices using a microtome (Leica Microsystems, Germany). The subsequent processing steps, including dehydration, antigen retrieval, antibody incubations, and staining, were performed according to the manufacturer’s instructions provided in the manuals for the DAB Substrate Kit (Peroxidase, HRP; Vector Laboratories, Newark, California, USA) and the IHC-Paraffin Protocol (IHC-P; Abcam plc, Cambridge, UK).
Immunohistochemical reactions were observed and documented using a Nikon Coolpix fluorescent optical system. The cellular localization and quantification of the selected proteins were analyzed through computer-assisted image analysis using ImageJ software. Images were captured from three slides at 200× magnification.
The optical density of DAB reaction products was assessed in the regions where immunohistochemical reactions occurred in response to the selected proteins. This analysis was performed using the IHC-Profiler plug-in within ImageJ software. Additionally, the average percentage of the DAB-stained area was calculated relative to the background values in each field.
2.9 Statistical analysis
Statistical analyses were conducted using StatPlus and the Transcriptome Analysis Console (Affymetrix). The Shapiro–Wilk test assessed data normality (p < 0.05). For group comparisons, one-way ANOVA with Benjamini–Hochberg correction and Scheffe’s post hoc test (p < 0.05) were applied. Pairwise comparisons were performed using Student’s t-test. Survival analysis was conducted using the Kaplan-Meier method, with differences assessed by the log-rank test. Cox proportional hazards regression evaluated the impact of TGF-β-1 and TGF-β-3, and selected miRNAs expression on survival. All tests were two-tailed, with p < 0.05 considered statistically significant.
2.10 Sample size analysis
According to national statistics, approximately 3,270 primary brain tumors were diagnosed in Poland between 2020 and 2021 (43). Given our study duration and access to clinical material, we enrolled 65 patients. Assuming a maximum variance (fraction size = 0.5) and a 95% confidence level, this sample yields a maximum margin of error of approximately 12%.
3 Results
3.1 Microarray and RTqPCR expression analysis of TGF-β-1–3 in G3/G4 astrocytic tumor samples compared to G2 samples
Microarray and RT-qPCR analyses revealed distinct expression patterns of TGF-β isoforms across tumor grades. Microarray data (Table 3) showed that TGF-β-1 expression was elevated 1.43-fold in G3 tumors compared to G2 (p < 0.05) and 1.54-fold in G4 tumors compared to G2 (p < 0.05). TGF-β-3 expression showed even greater upregulation, with a 2.76-fold increase in G3 (p < 0.05) and a 3.45-fold increase in G4 samples relative to G2 (p < 0.05). In contrast, TGF-β-2 expression remained relatively stable, with fold changes of 1.10 in G3 and 1.17 in G4, showing no statistically significant differences (p>0.05).
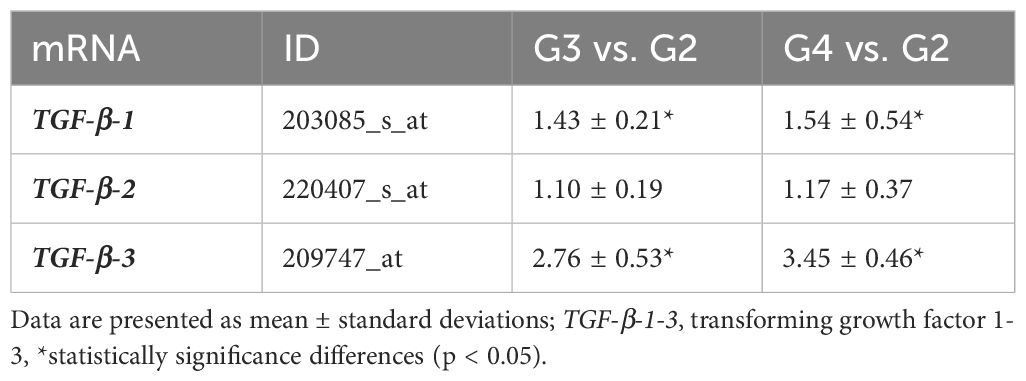
Table 3. Microarray profile of TGF-β-1–3 expression in G3 and G4 astrocytic tumor samples compared to G2 samples.
RT-qPCR validation (Figure 1) confirmed these findings, with TGF-β-1 mRNA levels significantly elevated in G3 and G4 tumors (p < 0.05). Similarly, TGF-β-3 mRNA levels increased significantly in higher grades, while TGF-β-2 mRNA expression did not differ significantly across grades.
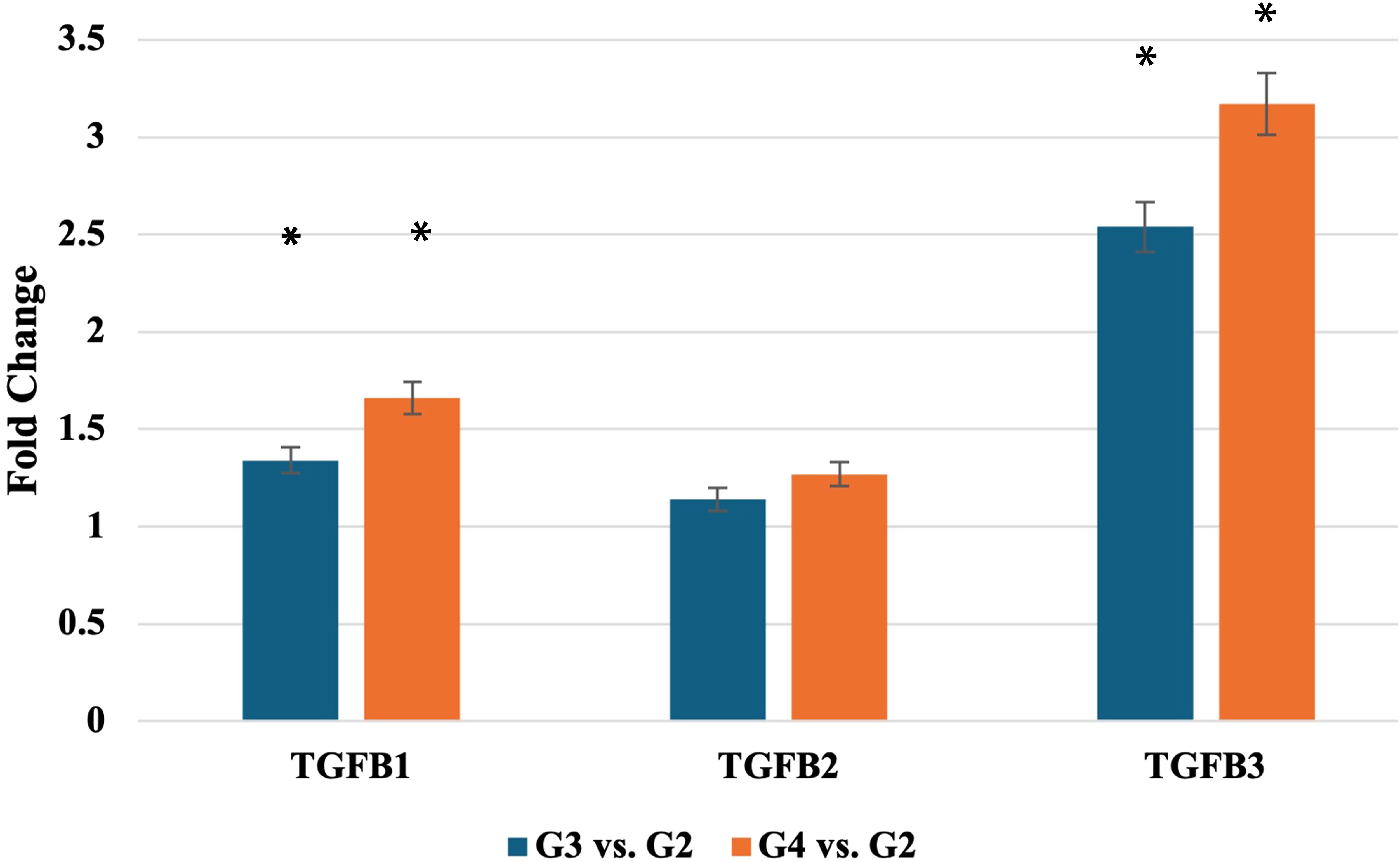
Figure 1. Expression profile of TGF-β-1–3 mRNA obtained via RTqPCR. TGF-β-1-3, transforming growth factor 1-3; *statistically significance differences (p < 0.05).
These grade-associated increases in TGF-β-1 and TGF-β-3 mRNA expression suggest a role in tumor progression. To identify upstream regulators potentially driving these changes, we next analyzed candidate microRNAs with predicted binding to the TGF-β isoforms.
3.2 Quantification of TGF-β-1–3 protein concentration in G2, G3, and G4 astrocytic tumors by ELISA and western blot analysis
The ELISA-based quantification of TGF-β-1–3 protein levels in G2, G3, and G4 astrocytic tumors revealed significant differences between tumor grades (Table 4). TGF-β-1 concentration increased from 723.87 ± 123.13 pg/ml in G2 to 956.13 ± 87.65 pg/ml in G4, while TGF-β-3 levels rose from 876.23 ± 76.87 pg/ml in G2 to 1108.81 ± 156.73 pg/ml in G4. In contrast, TGF-β-2 remained below the detection limit in all samples.
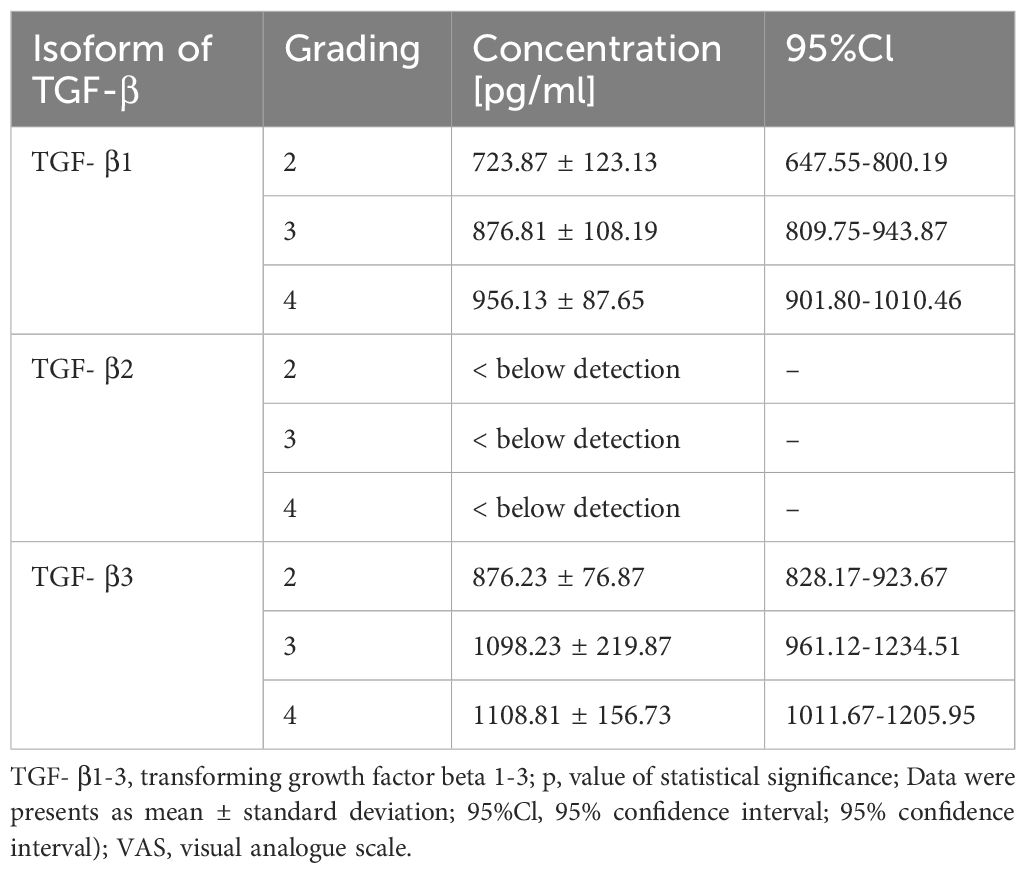
Table 4. Quantification of TGF-β-1–3 Protein Concentration in G2, G3, and G4 Astrocytic Tumors by ELISA.
Statistical analysis revealed a significant main effect (p < 0.05), with post hoc testing confirming significant differences between G2 and G4 for both TGF-β-1 and TGF-β-3.
Figure 2 presents a representative electropherogram, confirming the specificity of the reaction and the integrity of the samples. The presence of TGF-β-1 was validated by detecting a 44 kDa band, while TGF-β-3 was confirmed by a 47 kDa band. The sample quality was further verified by the presence of GAPDH at 37 kDa, serving as a loading control. Western blot quantification showed that TGF-β-1 levels, normalized to GAPD, were 0.34 ± 0.11 in G2 samples, 0.45 ± 0.18 in G3, and 0.47 ± 0.12 in G4, with no significant differences observed (p > 0.05). Notably, TGF-β-2 was not detected in any of the analyzed samples. In contrast, TGF-β-3 expression, also normalized to GAPDH, was significantly elevated across tumor grades, increasing from 2.17 ± 0.42 in G2 to 4.54 ± 0.65 in G3 and 5.15 ± 1.09 in G4 (p < 0.05).
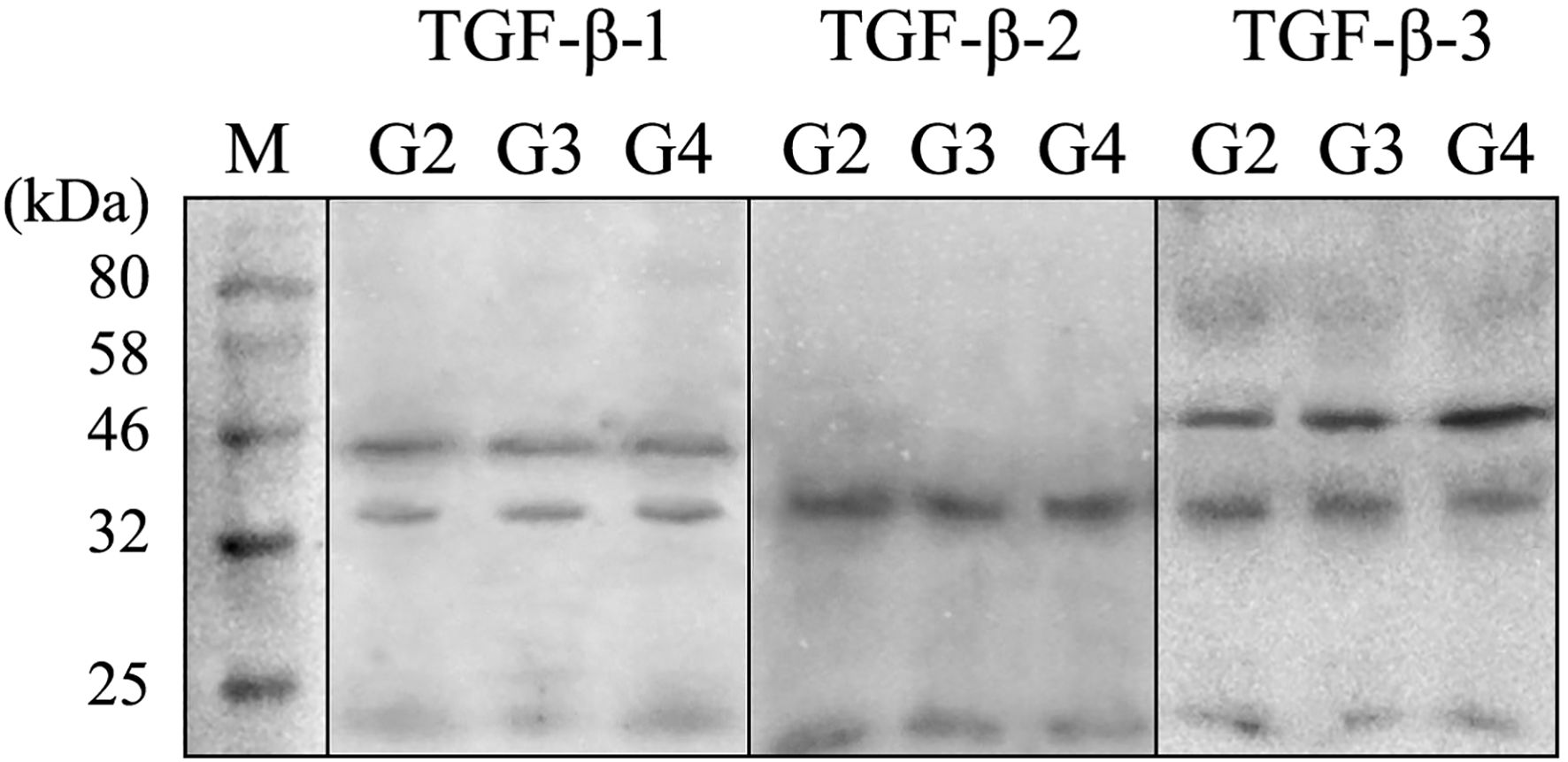
Figure 2. Example of electrophoretic separation of TGF- β1–3 in the G2, G3, and G4 astrocytic tumor samples. TGF β-1 and - 3, transforming growth factor beta 1 and 3; kDa, kilo Daltons; M, size marker (New England Biolabs Marker).
3.3 TGF-β-1 and TGF-β-3 expression profile determined by IHC
IHC staining showed elevated optical density for TGF-β-1 in G3 (137.81 ± 8.76% vs. G2) and G4 (178.01 ± 12.19% vs. G2) samples (p < 0.05). TGF-β-3 optical density increased even more markedly: 195.03 ± 14.56% in G3 and 276.19 ± 19.76% in G4 relative to G2 (p < 0.05). No TGF-β-2 signal was detected in any group. The results were presented in the Figure 3.
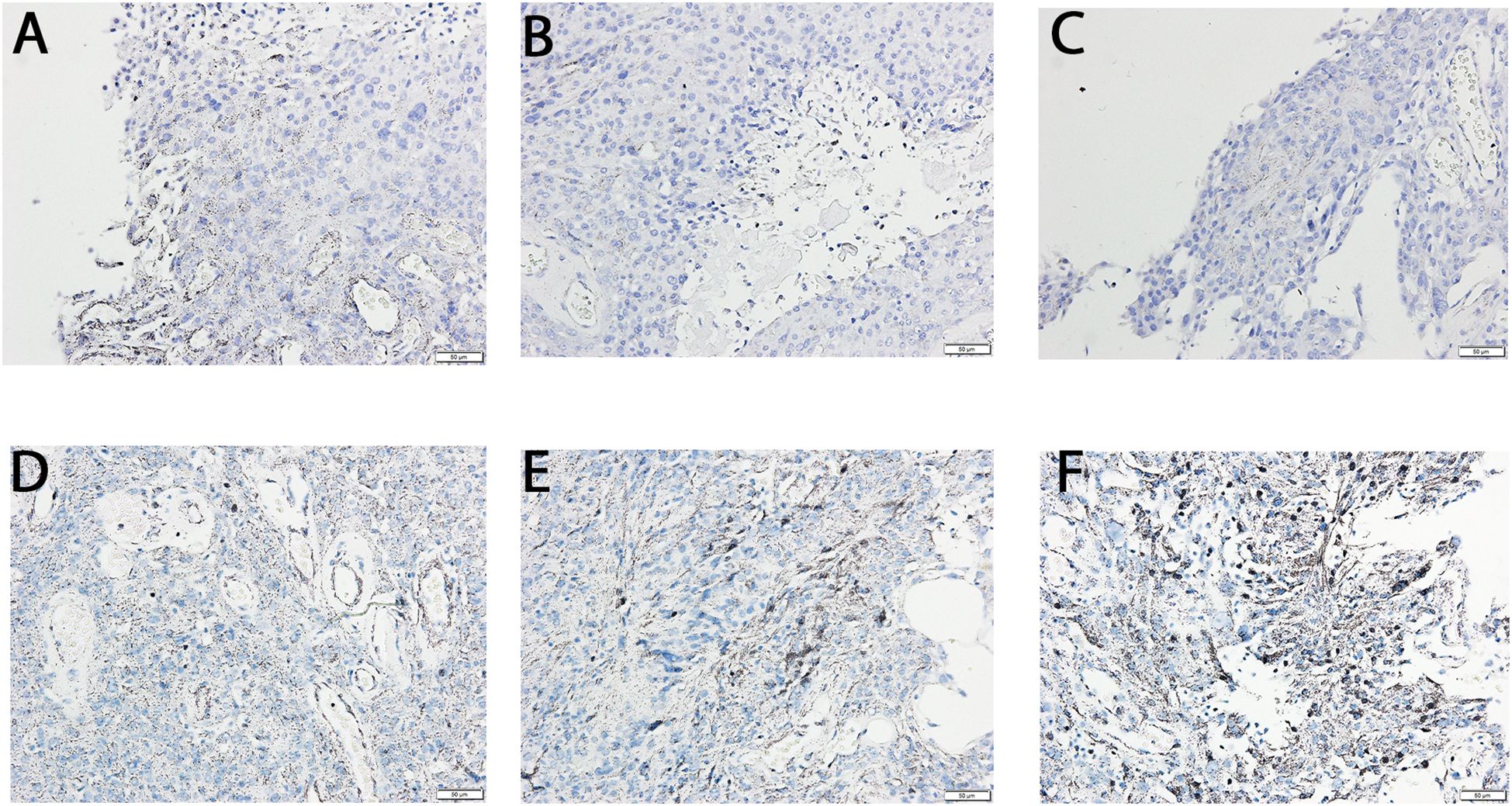
Figure 3. Immunochemical expression of TGF-β -1and -3 in the G2, G3, and G4 astrocytic tumor samples. (A) TGF β -1 in G2 samples; (B) TGF β -1 in G3 samples; (C) TGF β -1 in G4 samples; (D) TGF β -3 in G2 samples; E, TGF β -3 in G3 samples; F, TGF β -3 in G4 samples.
3.4 Predicted miRNA-mediated regulation of TGF-β-1–3 expression
To explore post-transcriptional regulatory mechanisms, we analyzed differentially expressed miRNAs across tumor grades and predicted their targets (Table 5). miR-3196 was markedly downregulated in both G3 and G4 astrocytomas and was predicted to target TGF-β-1 (target score: 80). Conversely, miR-466, miR-141-3p, and miR-200a-3p were upregulated and predicted to regulate TGF-β-2 (target scores: 99, 95, and 95, respectively). miR-2278, moderately upregulated in higher-grade tumors, showed a strong predictive association with TGF-β-3 regulation (target score: 92).
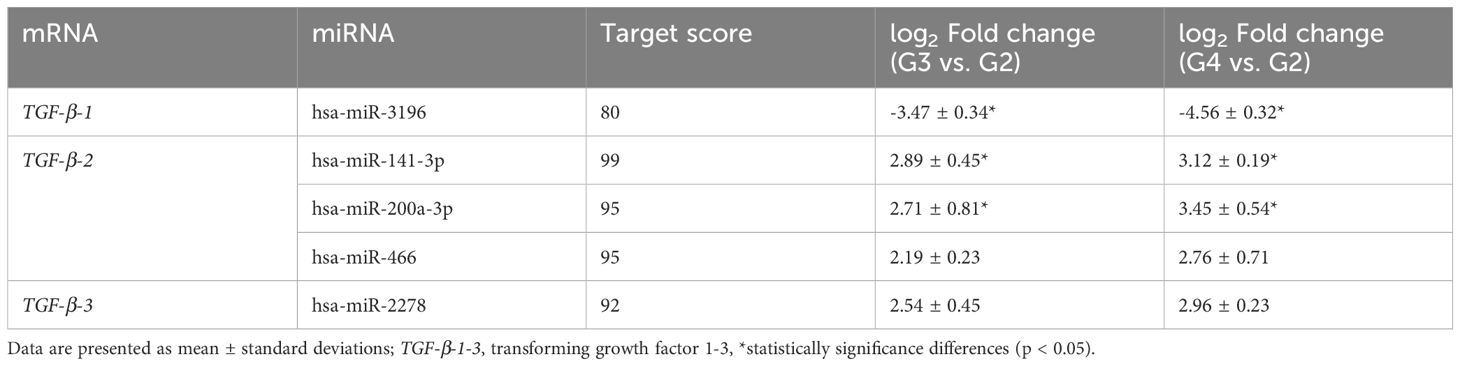
Table 5. Differential expression profile of miRNAs predicted to regulate TGF-β isoforms in G3 and G4 astrocytic tumor samples compared to G2.
The predicted interactions between these miRNAs and TGF-β isoforms highlight a potential post-transcriptional mechanism of regulation. To further investigate additional layers of control, we examined the methylation status of these genes across tumor grades.
3.5 Epigenetic studies revealed differential DNA methylation patterns of TGF-β isoforms in G2, G3 and G4 tumours
To explore the epigenetic regulation of TGF-β- isoforms in glioma, the DNA methylation patterns of TGF-β–1–3 were analyzed by MSPCR. MSPCR revealed variable epigenetic regulation of TGF-β isoforms by tumor grade. In G2 tumors, partial methylation was observed for TGF-β-1 and TGF-β-3, while TGF-β-2 was unmethylated. In G3 samples, most TGF-β-1 and TGF-β-2 genes were methylated, whereas TGF-β-3 exhibited low methylation. In G4 tumors, TGF-β-2 was highly methylated in nearly all cases, while TGF-β-1 and TGF-β-3 showed mixed methylation statuses (Figure 4).
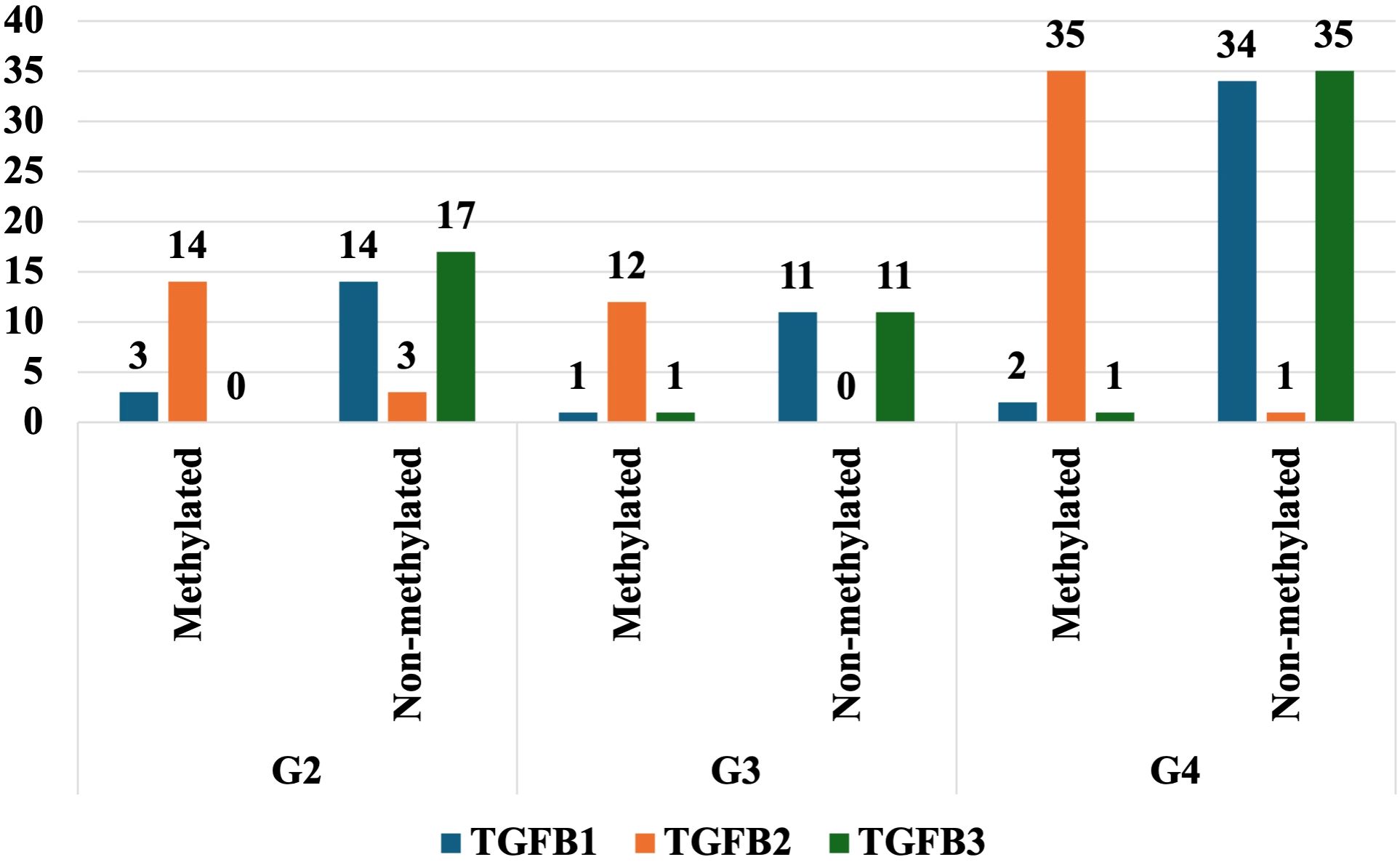
Figure 4. The degree of methylation of selected genes in the G2, G3, and G4 astrocytic tumor samples.
3.6 Kaplan-Meier survival analysis and cox proportional hazards model for TGF-β-1 and TGF-β-3 in astrocytic tumors
Kaplan-Meier survival analysis was conducted to evaluate the prognostic impact of TGF-β-1 and TGF-β-3 expression on patient survival, revealing that higher expression levels of both isoforms were associated with poorer survival outcomes (Figure 5). Cox regression analysis for TGF-β-1 (Exp(B) = 1.00225, p = 0.08054) indicated that patients with higher TGF-β-1 expression exhibited reduced survival, but the effect was not statistically significant (p > 0.05), suggesting that while a trend was observed, additional validation may be required to confirm its prognostic relevance. In contrast, TGF-β-3 expression (Exp(B) = 1.02644, p < 0.0001) was strongly associated with decreased survival, with each unit increase in expression raising the risk of death by 2.6%. This effect was highly statistically significant (p < 0.0001), indicating that TGF-β-3 could serve as a valuable prognostic biomarker in astrocytic tumors. Overall, these findings suggest that TGF-β-3 expression plays a crucial role in tumor progression and patient survival, whereas TGF-β-1 exhibits a weaker, albeit negative, influence on survival outcomes.
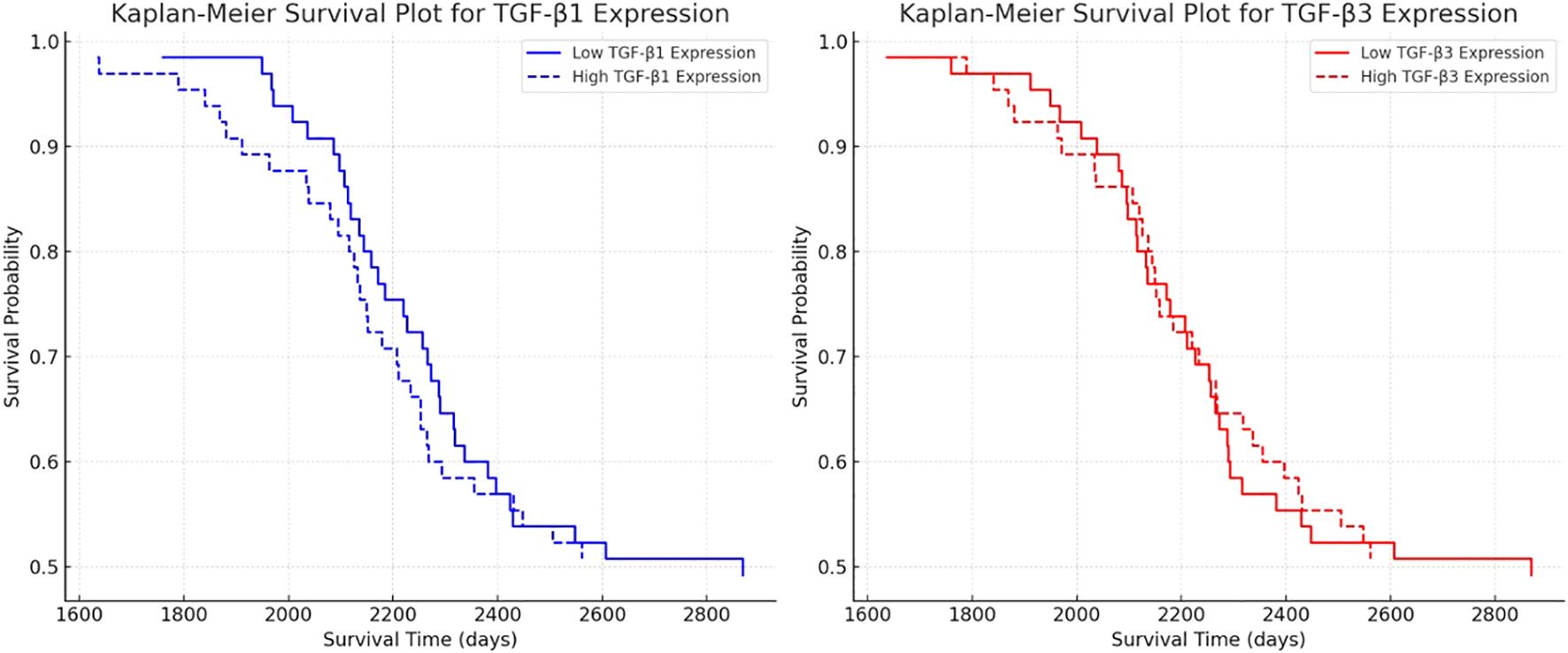
Figure 5. Kaplan-Meier survival analysis for TGF-β-1 and TGF-β-3 expression in astrocytic tumors.
In turn, Kaplan-Meier survival analysis was conducted to evaluate the prognostic impact of hsa-miR-3196, hsa-miR-141-3p, hsa-miR-200a-3p, hsa-miR-466, and hsa-miR-2278 expression on patient survival, revealing that higher expression of hsa-miR-2278 and lower expression of hsa-miR-3196 were associated with poorer survival outcomes (Figure 6). Cox regression analysis for hsa-miR-3196 (Exp(B) = 0.8897, p = 0.076) indicated that patients with reduced expression exhibited decreased survival, though the effect did not reach statistical significance (p > 0.05), suggesting a negative survival trend that warrants further investigation. In contrast, hsa-miR-2278 (Exp(B) = 1.437, p = 0.008) was significantly associated with increased mortality risk, with each unit increase in expression elevating the risk of death by approximately 43.7%. This effect was statistically significant (p < 0.01), supporting the potential of hsa-miR-2278 as a prognostic biomarker in astrocytic tumors. The remaining miRNAs—hsa-miR-141-3p, hsa-miR-200a-3p, and hsa-miR-466—showed no significant impact on survival, indicating limited prognostic value in this cohort.
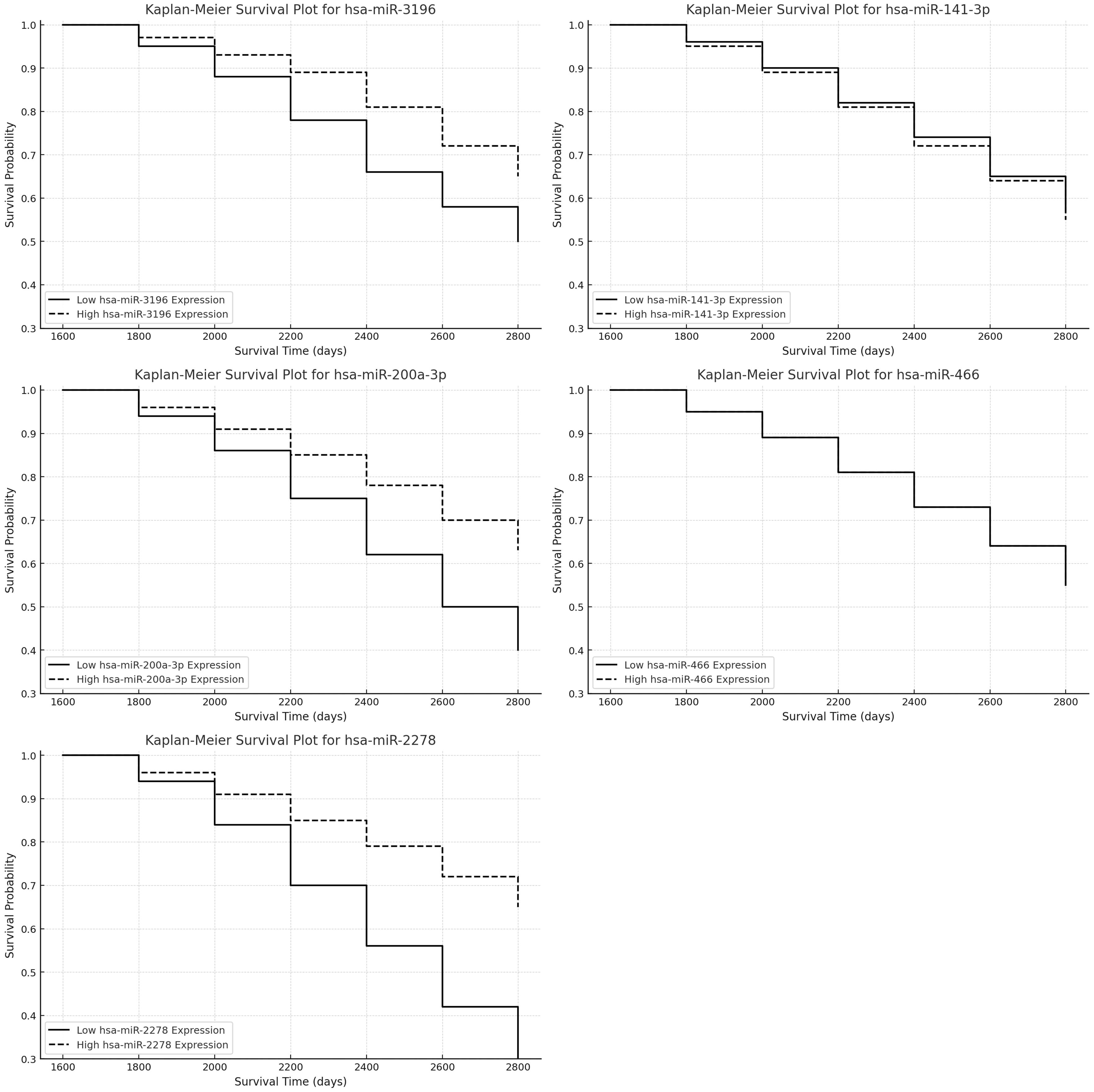
Figure 6. Kaplan-Meier survival analysis for selected miRNAs expression in astrocytic tumors.
4 Discussion
Despite increasing recognition of TGF-β’s role in glioma biology, the distinct contributions of its isoforms (TGF-β-1, -β2, and -β3) across tumor grades remain unclear. Our study addresses this critical gap by integrating expression profiling, DNA methylation, and miRNA regulation to elucidate the isoform-specific dynamics and prognostic significance of TGF-β in astrocytic tumors. Our results demonstrate significantly increased expression of TGF-β-1 and TGF-β-3 in G3 and G4 tumors compared to grade II (G2), while TGF-β-2 levels remained statistically unchanged. These findings align with prior studies reporting upregulation of TGF-β isoforms in high-grade gliomas. According to other researchers, including Frei et al., there is a strong correlation between TGF-β isoform expression and glioma progression [51]. Their study analyzed 64 newly diagnosed and 16 recurrent astrocytic tumors, assessing TGF-β-1 to TGF-β-3 expression. Among the isoforms, TGF-β-1 mRNA was the most abundant, while TGF-β-3 mRNA was the least. Survival analysis indicated that patients with high mRNA expression of TGF-β-2 or pSmad1/5/8 protein had worse outcomes compared to other study participants (44).
Kurowska et al. investigated 43 astrocytic tumor tissue sections across various disease stages, quantitatively assessing TGF-β isoform mRNA levels and the expression of genes associated with TGF-β signaling (22). Their microarray analysis showed a statistically significant increase in TGF-β-1 and TGF-β-2 expression in G3/G4 tumors compared to G2, whereas RT-qPCR validation confirmed this increase only for TGF-β-2. The authors suggested that quantifying TGF-β-2 mRNA could be a valuable diagnostic tool in the future (22). This finding differs from our study results, which indicate that TGF-β-3 is a strong prognostic biomarker in astrocytic tumors, significantly correlating with poor survival outcomes. TGF-β-1 showed an increasing trend in high-grade tumors but lacked statistical significance in survival analysis. These results highlight TGF-β-3 as a potential therapeutic target in astrocytic tumors. Differences in study outcomes may arise from methodological variations, patient cohort composition and size, or tumor microenvironment-specific factors. The roles of individual TGF-β isoforms in gliomas may be more complex than previously thought, warranting further investigation into the regulatory mechanisms of these cytokines in tumor progression.
A study by Battle et al. demonstrated that genetic and epigenetic changes can alter transcriptional circuits in cancer cells, leading to a shift in TGF-β signaling from protective to tumor-promoting effects (45). This study highlights that TGF-β-mediated immune suppression is context-dependent. Battle et al. state that in tissues exposed to continuous antigenic stimulation, such as the gastrointestinal tract, TGF-β suppresses inflammation and adaptive immune responses, thereby reducing tumor development. However, in advanced cancer stages, TGF-β signaling enables immune evasion in various tumor types and contributes to disease progression (45). This effect allows transformed tumor cells to evade immune system-mediated destruction, including by T cells and natural killer (NK) cells. Understanding TGF-β’s role in cancer development provides hope for identifying specific TGF-β pathway inhibitors to improve cancer prognosis.
Our study demonstrates the significant impact of epigenetic factors on the regulation of TGF-β-1–3 expression in astrocytic tumors. We observed different methylation patterns across various tumor grades. In G2 tumors, methylation was detected for TGF-β-1 and TGF-β-3 transcripts, whereas TGF-β-2 remained unmethylated. In G3 astrocytic tumors, most samples showed methylation for TGF-β-1 and TGF-β-2, whereas TGF-β-3 exhibited minimal methylation. In G4 tumors, nearly all samples were methylated for TGF-β-2, whereas TGF-β-1 and TGF-β-3 displayed a mixture of methylated and unmethylated cases. These observed methylation trends suggest potential epigenetic regulation of TGF-β expression in astrocytic tumor progression.
In our study, we identified several miRNAs that may influence the regulation of TGF-β expression. MiR-3196 was found to be significantly downregulated in G3 and G4 tumors, with predictive analysis indicating its potential role in regulating TGF-β-1 expression. Notably, hsa-miR-466, hsa-miR-141-3p, and hsa-miR-200a-3p were upregulated in both tumor grades, with high predictive scores for regulating TGF-β-2. Similarly, hsa-miR-2278, which showed moderate upregulation, demonstrated a strong predictive association with TGF-β-3 regulation.
Qi-Qi et al. highlight the role of miR-141, which functions as either a tumor suppressor in different cancers, regulating tumor cell proliferation, apoptosis, invasion, and metastasis through various signaling pathways, including the phosphatidylinositol-3-kinase (PI3K)/protein kinase B (AKT) pathway and the constitutive activation of nuclear factor-κB (NF-κB). Their validation of the target gene and pathway analysis provided insights into the role of this miRNA in different tissues (46). Their review also presents new findings suggesting that miR-141 could serve as a non-invasive biomarker and therapeutic target for several types of cancer.
A meta-analysis by Peng et al. on the miR-200 family demonstrated that these regulatory epigenetic molecules modulate physiological and pathological processes by targeting multiple genes, ultimately influencing glioma cell proliferation and invasion, as well as therapeutic response and prognosis (47). In turn, the study by Chen et al. reported that grade IV glioma tissues exhibited significantly lower levels of miR-200a compared to grade II–III gliomas. Glioma patients were categorized into high- and low-miR-200a expression groups based on the median expression value. Their analysis showed that low miR-200a expression was significantly associated with an advanced clinical stage of glioma. Additionally, miR-200a levels were markedly reduced in several common glioma cell lines compared to healthy astrocytic cells (48). The research by Bian et al. demonstrated that miR-141 was significantly downregulated in glioma tissues and cell lines compared to normal brain tissues, with its expression correlating with pathological tumor grade. Forced expression of miR-141 in glioma cells significantly inhibited cell proliferation, migration, and invasion, whereas miR-141 silencing had the opposite effect (49).
In summary, our study revealed a complex and isoform-specific relationship between promoter methylation, microRNA expression, and the regulation of TGF-β isoforms in astrocytic tumors. Notably, TGF-β-2 exhibited substantial promoter methylation in glioblastoma (G4) samples but did not show consistent downregulation at the mRNA or protein level, suggesting that promoter methylation alone is insufficient to repress its expression in this context. This finding implies the involvement of compensatory or overriding regulatory mechanisms such as miRNA-mediated translational control. In contrast, TGF-β-3 expression was significantly upregulated in high-grade tumors despite exhibiting relatively low promoter methylation, indicating methylation-independent transcriptional activation. Our integrated analysis of differentially expressed miRNAs further supports this regulatory complexity. Specifically, the downregulation of miR-3196, predicted to target TGF-β-1, and upregulation of miR-2278, predicted to regulate TGF-β-3, were both associated with increased expression of their respective protein targets. Meanwhile, miRNAs predicted to target TGF-β-2 (e.g., hsa-miR-141-3p and hsa-miR-200a-3p) were found to be upregulated in higher-grade tumors, potentially contributing to post-transcriptional repression of TGF-β-2, thereby explaining the discrepancy between methylation and expression profiles. These findings are consistent with the concept of mutual regulation between miRNAs and DNA methylation in cancer epigenetics, as described by Wang et al. (50). In this model, miRNAs can influence the activity of DNA methyltransferases (DNMTs), while their own expression is in turn modulated by promoter methylation. Thus, in gliomas, the interplay between miRNA expression and DNA methylation may create a dynamic regulatory network governing the expression of TGF-β isoforms. This dual-layered control appears to be grade-dependent, emphasizing the importance of considering both epigenetic mechanisms in understanding glioma progression and in identifying therapeutic targets (31, 50–54).
Furthermore, in our study, we assessed the correlation between TGF-β-1 and TGF-β-3 expression and five-year survival. Our survival analysis indicated that high TGF-β-3 expression was significantly associated with poor prognosis, whereas the effect of TGF-β-1 on survival was weaker and did not reach statistical significance. This finding aligns with previous reports suggesting that TGF-β plays a complex role in astrocytic tumor progression, acting as a tumor suppressor in the early stages and a promoter of aggressiveness in advanced cases (55, 56).
Elevated TGF-β-3 expression in high-grade tumors may contribute to a more invasive tumor phenotype through several mechanisms. Firstly, TGF-β is a key regulator of epithelial-to-mesenchymal transition (EMT), which enhances the migratory and invasive capabilities of cancer cells. In gliomas, this process facilitates tumor spread within the brain, leading to poorer prognosis (57). secondly, TGF-β exerts immunosuppressive effects by inhibiting T and NK cell activity and promoting the M2 phenotype of suppressor cells in the tumor microenvironment, thereby reducing the efficacy of the body’s immune response (24, 26, 27).
Moreover, our Cox regression analysis revealed that each unit increase in TGF-β-3 expression elevated the risk of patient mortality by 2.6% (Exp(B) = 1.02644, p < 0.0001), highlighting its strong prognostic significance. While high TGF-β-1 expression also trended toward worse survival outcomes, this association did not reach statistical significance (Exp(B) = 1.00225, p = 0.08054). It is possible that TGF-β-1 plays a more complex role, with its net effect on glioma progression depending on molecular context and interactions with other signaling pathways.
The strong correlation between elevated TGF-β-3 expression in high-grade tumors and poor patient survival suggests its potential use as a biomarker for risk stratification and therapeutic decision-making. Current treatment strategies for high-grade gliomas primarily involve surgical resection, radiotherapy, and temozolomide-based chemotherapy. Unfortunately, these approaches have limited efficacy, and gliomas often develop resistance to treatment (58). Increased TGF-β-3 expression may contribute to this resistance by promoting immunosuppression and angiogenesis in the tumor microenvironment, suggesting that targeted inhibition of TGF-β-3 could enhance the effectiveness of existing therapies.
Although pharmacologic inhibition of the TGF-β pathway has long been explored in glioma, clinical applications have been hampered by systemic toxicity and the pleiotropic, context-dependent nature of TGF-β signaling (59, 60). Direct inhibition of TGF-β can disrupt its tumor-suppressive functions in early-stage disease while only partially suppressing its pro-oncogenic effects in advanced stages. In this context, the microRNAs identified in our study—particularly miR-2278 and miR-3196—may provide a more selective and biologically nuanced means of modulating the TGF-β pathway. Because miRNAs act post-transcriptionally and, in a tissue,-specific manner, they may allow for isoform- and context-selective regulation with fewer off-target effects compared to systemic inhibitors (61–63). These miRNAs not only reflect key aspects of tumor biology but also represent promising therapeutic targets for overcoming TGF-β–driven progression and resistance in astrocytic tumors.
In addition to gene-level analysis, we evaluated the prognostic impact of selected regulatory microRNAs predicted to target TGF-β isoforms. Kaplan-Meier survival analysis revealed that high expression of hsa-miR-2278 was significantly associated with worse patient outcomes, with a 43.7% increase in mortality risk per unit expression (Exp(B) = 1.437, p = 0.008). This finding is particularly relevant given that miR-2278 is predicted to regulate TGF-β-3, suggesting a mechanistic link between its upregulation in high-grade gliomas and the enhanced expression of this profibrotic cytokine. Conversely, hsa-miR-3196 exhibited a trend toward better survival in the high-expression group (Exp(B) = 0.8897, p = 0.076), consistent with its downregulation in high-grade tumors and putative targeting of TGF-β-1. Other analyzed miRNAs (hsa-miR-141-3p, hsa-miR-200a-3p, and hsa-miR-466) did not show statistically significant associations with survival. These results suggest that miR-2278 and miR-3196 may influence glioma progression via post-transcriptional regulation of TGF-β isoforms and may serve as potential biomarkers for patient stratification.
The upregulation of TGF-β-3 in high-grade gliomas may drive a more aggressive phenotype via several mechanisms. Current treatment strategies for high-grade gliomas primarily involve surgical resection, radiotherapy, and temozolomide-based chemotherapy. Unfortunately, these approaches have limited efficacy, and gliomas often develop resistance to treatment (58). Increased TGF-β-3 expression may contribute to this resistance by promoting immunosuppression and angiogenesis in the tumor microenvironment, suggesting that targeted inhibition of TGF-β-3 could enhance the effectiveness of existing therapies. Similarly, modulation of TGF-β-regulating miRNAs, such as miR-2278 and miR-3196, may offer novel therapeutic avenues for influencing tumor behavior and overcoming resistance mechanisms.
This study highlights the differential expression and epigenetic regulation of TGF-β isoforms in astrocytic tumors, demonstrating that TGF-β-1 and TGF-β-3 exhibit increased expression in high-grade tumors, whereas TGF-β-2 remains largely unchanged. Notably, TGF-β-3 emerged as a strong prognostic biomarker, significantly correlating with poorer survival, whereas TGF-β-1 showed an increasing trend in high-grade tumors but lacked statistical significance in survival analysis. The observed methylation patterns and miRNA interactions suggest a complex epigenetic regulation of TGF-β signaling in tumor progression, further emphasizing its role in glioma pathophysiology. These findings underscore the potential of TGF-β-3 as a therapeutic target, highlighting the need for further studies to explore targeted interventions aimed at modulating its expression and activity. While this study provides important molecular insights, future research integrating functional assays and larger patient cohorts is necessary to validate these results and advance precision medicine approaches in astrocytic tumor management.
4.1 Limitations of the study
While this study offers meaningful insights into the differential expression and epigenetic regulation of TGF-β isoforms and their associated microRNAs across astrocytic tumor grades, several limitations should be considered. First, the modest sample size (n = 65) from a limited geographical region may restrict the generalizability of the findings and introduces the potential for selection bias. Although TGF-β-3 emerged as a strong prognostic marker, the novelty of the study is tempered by existing literature extensively documenting the role of TGF-β signaling in glioma (22, 23, 64–70). Moreover, although predictive bioinformatics analyses identified candidate miRNAs and methylation patterns potentially regulating TGF-β isoforms, the lack of functional validation through in vitro assays (e.g., miRNA modulation, dual-luciferase reporter assays, or SMAD phosphorylation studies) limits the mechanistic interpretation. Similarly, interactions between treatment history, genetic markers such as IDH mutation, and the expression of TGF-β isoforms were not explored. Despite these limitations, the study provides a comprehensive, multi-level molecular profile of TGF-β isoforms in glioma and identifies TGF-β-3 and miR-2278 as promising prognostic indicators. Future studies should incorporate functional experiments, broader patient cohorts, and multivariate analyses to confirm these findings and support their clinical relevance.
Data availability statement
The original contributions presented in the study are publicly available. This data can be found here: https://doi.org/10.6084/m9.figshare.29304938.v1.
Ethics statement
Ethical considerations were a primary focus in this study, strictly following the guidelines of the 2013 Declaration of Helsinki related to human experimentation. The Bioethical Committee of the Regional Medical Chamber in Krakow granted approval, with reference number 247/KBL/OIL/2021, and the State Academy of Applied Sciences in Przemyśl (approval number: 9/2024). The studies were conducted in accordance with the local legislation and institutional requirements. Written informed consent for participation in this study was provided by the participants’ legal guardians/next of kin. Written informed consent was obtained from the individual(s) for the publication of any potentially identifiable images or data included in this article.
Author contributions
KS: Conceptualization, Data curation, Writing – original draft, Writing – review & editing. DSt: Conceptualization, Writing – original draft. DSo: Methodology, Writing – review & editing. RS: Methodology, Writing – review & editing. KB: Writing – original draft. PG: Writing – original draft. KB: Writing – original draft. MM: Resources, Writing – original draft. BG: Conceptualization, Data curation, Project administration, Software, Supervision, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Cano-Valdez AM and Sevilla-Lizcano DB. Pathological classification of brain tumors. In: Monroy-Sosa A, Chakravarthi SS, de la Garza-Salazar JG, Meneses Garcia A, and Kassam AB, editors. Principles of neuro-oncology. Springer International Publishing, Cham (2021). p. 75–105. doi: 10.1007/978-3-030-54879-7_5
2. Pienkowski T, Kowalczyk T, Kretowski A, and Ciborowski M. A review of gliomas-related proteins. Characteristics of potential biomarkers. Am J Cancer Res. (2021) 11:3425.
3. Machnik G, Bułdak Ł, Zapletal-Pudełko K, Grabarek BO, Staszkiewicz R, Sobański D, et al. The impact of wound-healing assay, phorbol myristate acetate (PMA) stimulation and siRNA-mediated FURIN gene silencing on endogenous retroviral ERVW-1 expression level in U87-MG astrocytoma cells. Adv Med Sci. (2024) 69:113–24. doi: 10.1016/j.advms.2024.02.007
4. Chen Z and Hambardzumyan D. Immune microenvironment in glioblastoma subtypes. Front Immunol. (2018) 9:1004. doi: 10.3389/fimmu.2018.01004
5. Hanif F, Muzaffar K, Perveen K, Malhi SM, and Simjee SU. Glioblastoma Multiforme: A Review of its Epidemiology and Pathogenesis through Clinical Presentation and Treatment. Asian Pac J Cancer Prev. (2017) 18:3–9. doi: 10.22034/APJCP.2017.18.1.3
6. Ostrom QT, Cioffi G, Gittleman H, Patil N, Waite K, Kruchko C, et al. CBTRUS statistical report: primary brain and other central nervous system tumors diagnosed in the United States in 2012–2016. Neuro-Oncology. (2019) 21:v1–v100. doi: 10.1093/neuonc/noz150
7. Reifenberger G, Wirsching H-G, Knobbe-Thomsen CB, and Weller M. Advances in the molecular genetics of gliomas - implications for classification and therapy. Nat Rev Clin Oncol. (2017) 14:434–52. doi: 10.1038/nrclinonc.2016.204
8. Bale TA and Rosenblum MK. The 2021 WHO Classification of Tumors of the Central Nervous System: An update on pediatric low-grade gliomas and glioneuronal tumors. Brain Pathol. (2022) 32:e13060. doi: 10.1111/bpa.13060
9. Horbinski C, Berger T, Packer RJ, and Wen PY. Clinical implications of the 2021 edition of the WHO classification of central nervous system tumours. Nat Rev Neurol. (2022) 18:515–29. doi: 10.1038/s41582-022-00679-w
10. Louis DN, Perry A, Wesseling P, Brat DJ, Cree IA, Figarella-Branger D, et al. The 2021 WHO classification of tumors of the central nervous system: a summary. Neuro Oncol. (2021) 23:1231–51. doi: 10.1093/neuonc/noab106
11. Sokolenko AP and Imyanitov EN. Molecular diagnostics in clinical oncology. Front Mol Biosci. (2018) 5:76. doi: 10.3389/fmolb.2018.00076
12. Ceccarelli M, Barthel FP, Malta TM, Sabedot TS, Salama SR, Murray BA, et al. Molecular profiling reveals biologically discrete subsets and pathways of progression in diffuse glioma. Cell. (2016) 164:550–63. doi: 10.1016/j.cell.2015.12.028
13. Radke J, Koch A, Pritsch F, Schumann E, Misch M, Hempt C, et al. Predictive MGMT status in a homogeneous cohort of IDH wildtype glioblastoma patients. Acta Neuropathologica Commun. (2019) 7:89. doi: 10.1186/s40478-019-0745-z
14. Labreche K, Kinnersley B, Berzero G, Di Stefano AL, Rahimian A, Detrait I, et al. Diffuse gliomas classified by 1p/19q co-deletion, TERT promoter and IDH mutation status are associated with specific genetic risk loci. Acta Neuropathol. (2018) 135:743–55. doi: 10.1007/s00401-018-1825-z
15. Huynh LK, Hipolito CJ, and ten Dijke P. A perspective on the development of TGF-β Inhibitors for cancer treatment. Biomolecules. (2019) 9:743. doi: 10.3390/biom9110743
16. Chen S, Liu S, Ma K, Zhao L, Lin H, and Shao Z. TGF-β signaling in intervertebral disc health and disease. Osteoarthritis Cartilage. (2019) 27:1109–17. doi: 10.1016/j.joca.2019.05.005
17. Zhao H, Wu L, Yan G, Chen Y, Zhou M, Wu Y, et al. Inflammation and tumor progression: signaling pathways and targeted intervention. Signal transduction targeted Ther. (2021) 6:263. doi: 10.1038/s41392-021-00658-5
18. Cekanaviciute E, Dietrich HK, Axtell RC, Williams AM, Egusquiza R, Wai KM, et al. Astrocytic TGF-β Signaling limits inflammation and reduces neuronal damage during central nervous system toxoplasma infection. J Immunol. (2014) 193:139–49. doi: 10.4049/jimmunol.1303284
19. Cheng T, Xu Z, and Ma X. The role of astrocytes in neuropathic pain. Front Mol Neurosci. (2022) 15:1007889. doi: 10.3389/fnmol.2022.1007889
20. Li S, Gu X, and Yi S. The regulatory effects of transforming growth factor-β on nerve regeneration. Cell Transplant. (2017) 26:381–94. doi: 10.3727/096368916X693824
21. Fabregat I, Fernando J, Mainez J, and Sancho P. TGF-beta signaling in cancer treatment. Curr Pharm design. (2014) 20:2934–47. doi: 10.2174/13816128113199990591
22. Kurowska N, Strzalka-Mrozik B, Madej M, Pająk K, Kruszniewska-Rajs C, Kaspera W, et al. Differences in the expression patterns of TGFβ Isoforms and associated genes in astrocytic brain tumors. Cancers (Basel). (2022) 14:1876. doi: 10.3390/cancers14081876
23. Chen Q, Fu W, Tang X, Wang L, Niu Q, Wang S, et al. ADP-Ribosylation Factor Like GTPase 4C (ARL4C) augments stem-like traits of glioblastoma cells by upregulating ALDH1A3. J Cancer. (2021) 12:818–26. doi: 10.7150/jca.45052
24. Birch JL, Coull BJ, Spender LC, Watt C, Willison A, Syed N, et al. Multifaceted transforming growth factor-beta (TGFβ) signalling in glioblastoma. Cell Signalling. (2020) 72:109638. doi: 10.1016/j.cellsig.2020.109638
25. Peng D, Fu M, Wang M, Wei Y, and Wei X. Targeting TGF-β signal transduction for fibrosis and cancer therapy. Mol Cancer. (2022) 21:104. doi: 10.1186/s12943-022-01569-x
26. Cantelli G, Crosas-Molist E, Georgouli M, and Sanz-Moreno V. TGFB-induced transcription in cancer. Semin Cancer Biol. (2017) 42:60–9. doi: 10.1016/j.semcancer.2016.08.009
27. Eble JA and Niland S. The extracellular matrix in tumor progression and metastasis. Clin Exp Metastasis. (2019) 36:171–98. doi: 10.1007/s10585-019-09966-1
28. Garg P, Pareek S, Kulkarni P, Horne D, Salgia R, and Singhal SS. Exploring the potential of tgfβ As A diagnostic marker and therapeutic target against cancer. Biochem Pharmacol. (2024), 116646.
29. Sheikh K, Amjad M, Irfan M, Anjum S, Majeed T, Riaz M, et al. Exploring TGF-β Signaling in cancer progression: prospects and therapeutic strategies. OTT. (2025) 18:233–62. doi: 10.2147/OTT.S493643
30. Danielpour D. Advances and challenges in targeting tgf-β isoforms for therapeutic intervention of cancer: A mechanism-based perspective. Pharmaceuticals. (2024) 17:533. doi: 10.3390/ph17040533
31. Naik A and Thakur N. Epigenetic regulation of TGF-β and vice versa in cancers – A review on recent developments. Biochim Biophys Acta (BBA) - Rev Cancer. (2024) 1879:189219. doi: 10.1016/j.bbcan.2024.189219
32. Ding Y, Zhou G, and Hu W. Epigenetic regulation of TGF-β pathway and its role in radiation response. Int J Radiat Biol. (2024) 100:834–48. doi: 10.1080/09553002.2024.2327395
33. Park JW, Lee K, Kim EE, Kim S-I, and Park S-H. Brain tumor classification by methylation profile. J Korean Med Sci. (2023) 38:e356. doi: 10.3346/jkms.2023.38.e356
34. Ahmed SP, Castresana JS, and Shahi MH. Glioblastoma and miRNAs. Cancers. (2021) 13:1581. doi: 10.3390/cancers13071581
35. Wang M, Wu Q, Fang M, Huang W, and Zhu H. miR-152-3p sensitizes glioblastoma cells towards cisplatin via regulation of SOS1 [Retraction. OTT. (2023) 16:477–8. doi: 10.2147/OTT.S427670
36. Bahreyni-Toossi M-T, Dolat E, Khanbabaei H, Zafari N, and Azimian H. microRNAs: Potential glioblastoma radiosensitizer by targeting radiation-related molecular pathways. Mutat Research/Fundamental Mol Mech Mutagenesis. (2019) 816–818:111679. doi: 10.1016/j.mrfmmm.2019.111679
37. Butz H, Rácz K, Hunyady L, and Patócs A. Crosstalk between TGF-β signaling and the microRNA machinery. Trends Pharmacol Sci. (2012) 33:382–93. doi: 10.1016/j.tips.2012.04.003
38. Staszkiewicz R, Sobański D, Pulka W, Gładysz D, Gadzieliński M, Strojny D, et al. Variances in the expression profile of circadian clock-related genes in astrocytic brain tumors. Cancers. (2024) 16:2335. doi: 10.3390/cancers16132335
39. Agarwal V, Bell GW, Nam J-W, and Bartel DP. Predicting effective microRNA target sites in mammalian mRNAs. eLife. (2015) 4:e05005. doi: 10.7554/eLife.05005
40. Chen Y and Wang X. miRDB: an online database for prediction of functional microRNA targets. Nucleic Acids Res. (2020) 48:D127–31. doi: 10.1093/nar/gkz757
41. Liu W and Wang X. Prediction of functional microRNA targets by integrative modeling of microRNA binding and target expression data. Genome Biol. (2019) 20:18. doi: 10.1186/s13059-019-1629-z
42. Li L-C and Dahiya R. MethPrimer: designing primers for methylation PCRs. Bioinformatics. (2002) 18:1427–31. doi: 10.1093/bioinformatics/18.11.1427
43. Raporty | Krajowy rejestr nowotworów . Available online at: http://onkologia.org.pl/pl/raporty (Accessed May 5, 2025).
44. Frei K, Gramatzki D, Tritschler I, Schroeder JJ, Espinoza L, Rushing EJ, et al. Transforming growth factor-β pathway activity in glioblastoma. Oncotarget. (2015) 6:5963–77. doi: 10.18632/oncotarget.3467
45. Batlle E and Massagué J. Transforming growth factor-β Signaling in immunity and cancer. Immunity. (2019) 50:924–40. doi: 10.1016/j.immuni.2019.03.024
46. Luo Q, Tian Y, Qu G, Kun-Huang, and Luo S. Functional mechanism and clinical implications of miR-141 in human cancers. Cell Signalling. (2022) 95:110354. doi: 10.1016/j.cellsig.2022.110354
47. Peng L, Fu J, and Ming Y. The miR-200 family: multiple effects on gliomas. Cancer Manag Res. (2018) 10:1987–92. doi: 10.2147/CMAR.S160945
48. Chen X, Liu K, Yang P, Kuang W, Huang H, Tu E, et al. microRNA−200a functions as a tumor suppressor by targeting FOXA1 in glioma. Exp Ther Med. (2018) 7:221–9. doi: 10.3892/etm.2018.6895
49. Bian E-B, Ma C-C, He X-J, Wang C, Zong G, Wang H-L, et al. Epigenetic modification of miR-141 regulates SKA2 by an endogenous ‘sponge’ HOTAIR in glioma. Oncotarget. (2016) 7:30610–25. doi: 10.18632/oncotarget.8895
50. Wang S, Wu W, and Claret FX. Mutual regulation of microRNAs and DNA methylation in human cancers. Epigenetics. (2017) 12:187–97. doi: 10.1080/15592294.2016.1273308
51. Fuso A, Raia T, Orticello M, and Lucarelli M. The complex interplay between DNA methylation and miRNAs in gene expression regulation. Biochimie. (2020) 173:12–6. doi: 10.1016/j.biochi.2020.02.006
52. Han L, Witmer PDW, Casey E, Valle D, and Sukumar S. DNA methylation regulates microRNA expression. Cancer Biol Ther. (2007) 6:1290–4. doi: 10.4161/cbt.6.8.4486
53. He X-X, Kuang S-Z, Liao J-Z, Xu C-R, Chang Y, Wu Y-L, et al. The regulation of microRNA expression by DNA methylation in hepatocellular carcinoma. Mol Biosyst. (2015) 11:532–9. doi: 10.1039/C4MB00563E
54. Zmarzły N, Wojdas E, Skubis A, Sikora B, and Mazurek U. DNA methylation: gene expression regulation. Folia Biologica et Oecologica (2017) 12:1–10. doi: 10.1515/fobio-2016-0001
55. Haque S and Morris JC. Transforming growth factor-β: A therapeutic target for cancer. Hum Vaccin Immunother. (2017) 13:1741–50. doi: 10.1080/21645515.2017.1327107
56. Seoane J and Gomis RR. TGF-β Family signaling in tumor suppression and cancer progression. Cold Spring Harb Perspect Biol. (2017) 9:a022277. doi: 10.1101/cshperspect.a022277
57. Miyazono K, Katsuno Y, Koinuma D, Ehata S, and Morikawa M. Intracellular and extracellular TGF-β signaling in cancer: some recent topics. Front Med. (2018) 12:387–411. doi: 10.1007/s11684-018-0646-8
58. Weller M, van den Bent M, Tonn JC, Stupp R, Preusser M, Cohen-Jonathan-Moyal E, et al. European Association for Neuro-Oncology (EANO) guideline on the diagnosis and treatment of adult astrocytic and oligodendroglial gliomas. Lancet Oncol. (2017) 18:e315–29. doi: 10.1016/S1470-2045(17)30194-8
59. Kim B-G, Malek E, Choi SH, Ignatz-Hoover JJ, and Driscoll JJ. Novel therapies emerging in oncology to target the TGF-β pathway. J Hematol Oncol. (2021) 14:1–20. doi: 10.1186/s13045-021-01053-x
60. Tschernia NP and Gulley JL. Tumor in the crossfire: inhibiting TGF-β to enhance cancer immunotherapy. BioDrugs. (2022) 36:153–80. doi: 10.1007/s40259-022-00521-1
61. Zhang S, Cheng Z, Wang Y, and Han T. The risks of miRNA therapeutics: in a drug target perspective. Drug design Dev Ther. (2021) 15:721–33. doi: 10.2147/DDDT.S288859
62. Fu Z, Wang L, Li S, Chen F, Au-Yeung KK-W, and Shi C. MicroRNA as an important target for anticancer drug development. Front Pharmacol. (2021) 12:736323. doi: 10.3389/fphar.2021.736323
63. Menon A, Abd-Aziz N, Khalid K, Poh CL, and Naidu R. miRNA: a promising therapeutic target in cancer. Int J Mol Sci. (2022) 23:11502. doi: 10.3390/ijms231911502
64. Duan W-W, Yang L-T, Liu J, Dai Z-Y, Wang Z-Y, Zhang H, et al. A TGF-β signaling-related lncRNA signature for prediction of glioma prognosis, immune microenvironment, and immunotherapy response. CNS Neurosci Ther. (2024) 30:e14489. doi: 10.1111/cns.14489
65. Chao M, Liu N, Sun Z, Jiang Y, Jiang T, Xv M, et al. TGF-β signaling promotes glioma progression through stabilizing Sox9. Front Immunol. (2021) 11:592080. doi: 10.3389/fimmu.2020.592080
66. Tu Y, Xu L, Xu J, Bao Z, Tian W, Ye Y, et al. Loss of deubiquitylase USP2 triggers development of glioblastoma via TGF-β signaling. Oncogene. (2022) 41:2597–608. doi: 10.1038/s41388-022-02275-0
67. Burghardt I, Ventura E, Weiss T, Schroeder JJ, Seystahl K, Zielasek C, et al. Endoglin and TGF-β signaling in glioblastoma. Cell Tissue Res. (2021) 384:613–24. doi: 10.1007/s00441-020-03323-5
68. Hernández-Vega AM and Camacho-Arroyo I. Crosstalk between 17β-estradiol and TGF-β signaling modulates glioblastoma progression. Brain Sci. (2021) 11:564.
Keywords: transforming growth factor beta 1-3, astrocytic tumors, survival analysis, micro RNA, molecular marker
Citation: Skóra K, Strojny D, Sobański D, Staszkiewicz R, Bryś K, Gogol P, Bereza K, Masternak M and Grabarek BO (2025) Multi-omics profiling of TGF-β isoforms and regulatory miRNAs in astrocytic tumors reveals TGF-β-3 as a prognostic biomarker. Front. Oncol. 15:1592685. doi: 10.3389/fonc.2025.1592685
Received: 21 March 2025; Accepted: 30 May 2025;
Published: 20 June 2025.
Edited by:
Liam Chen, University of Minnesota, United StatesReviewed by:
Bakhya Shree, Institute of Cancer Research (ICR), United KingdomTamilanban T, Mahsa University, Malaysia
Copyright © 2025 Skóra, Strojny, Sobański, Staszkiewicz, Bryś, Gogol, Bereza, Masternak and Grabarek. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Klaudia Skóra, a2xhdWRpYXNrb3JhMjZAZ21haWwuY29t