 Laura Rey-Vargas1,2
Laura Rey-Vargas1,2 Lina María Bejarano-Rivera1
Lina María Bejarano-Rivera1 Patricia López-Correa3
Patricia López-Correa3 Diego Felipe Ballen-Lozano4
Diego Felipe Ballen-Lozano4 Silvia J. Serrano-Gómez1,5*
Silvia J. Serrano-Gómez1,5*- 1Cancer Biology Research Group, National Cancer Institute, Bogotá, Colombia
- 2Doctoral Program in Biological Sciences, Pontificia Universidad Javeriana, Bogotá, Colombia
- 3Department of Pathology, National Cancer Institute, Bogotá, Colombia
- 4Clinical Oncology Unit, National Cancer Institute, Bogotá, Colombia
- 5Research Support and Follow-Up Group, National Cancer Institute, Bogotá, Colombia
Background: HER2-positive breast tumors are clinically important breast cancer subtypes with an overall unfavorable prognosis, but also with current optimal treatment options that have significantly improved the patients’ survival. Several epidemiological registries have reported varying prevalence rates of HER2-positive breast tumors among population groups. In this review, we describe the prevalence of HER2-positive breast tumors by ethnicity, with a special focus on Asian and Latina women, along with genetic variants located in or near ERBB2 that might affect its protein expression.
Methods: We conducted a literature search for studies reporting differences in HER2-positive breast tumor prevalence among populations and HER2/ERBB2 molecular features based on genomic background or ancestry.
Results: Overall, Asian and Latina women tend to have higher proportions of HER2-amplified tumors, compared to non-Hispanic white (NHW) women. Additionally, higher Indigenous American ancestry is associated with an increased likelihood of HER2-positive tumors and elevated ERBB2 expression. We also describe reported differences in the genotype of several genetic variants in ERBB2 or nearby genomic regions according to HER2 expression, and mention variants in other genes that may also be associated.
Conclusions: This literature review contributes to a better understanding of the underlying biology of HER2 expression in breast tumors, and the possible mechanisms that explain the differences in the distribution of HER2-positive subtypes among various population groups.
1 Introduction
Breast cancer is currently the most common malignancy diagnosed in women worldwide (46.8 per 100,000) and the leading cause of cancer mortality among women (12.6 per 100,000) (1). At the molecular level, breast cancer is a heterogeneous disease (2). Four major intrinsic subtypes have been described: luminal A, luminal B, human epidermal growth factor receptor 2 (HER2)-enriched, and basal-like. In the clinical setting, these subtypes can be identified by immunohistochemistry (IHC) techniques, mainly based on the expression of hormone receptors (HR) (estrogen (ER) and progesterone (PR) receptors) and the HER2 protein (3, 4). Each subtype has a different prognosis; patients with luminal A tumors have the best clinical outcome, whereas those with triple negative (TN) tumors or HER2 expression (whether classified as luminal/HER2+ or HER2-enriched subtypes), often present unfavorable outcomes as a consequence of these tumors’ aggressive phenotype (e.g., higher proliferation index and less differentiated tumors) (5). However, the clinical outcome of patients with HER2-positive tumors have significantly improved over the past years with the use of anti-HER2 therapy based on monoclonal antibodies such as trastuzumab, and tyrosine kinase (TK) inhibitors such as lapatinib (5, 6).
The prevalence of breast tumors with HER2 overexpression in the overall population ranges between 15% - 20% (4, 7), nonetheless, these percentages may vary according to ethnicity. Population-based studies have reported a higher proportion of HER2-amplified tumors in Asian and Latina women compared to non-Hispanic white (NHW) women (8–12). These variations can be partly related to the differences in the presentation of several reproductive and lifestyle factors between these population groups (13–15). However, in the past few years, various studies have stated that genetic-related factors can also contribute to HER2 expression in breast tumors (16–18). It has been described that the presence of genetic variants, such as single nucleotide polymorphisms (SNPs) in ERBB2, might influence the expression and activity of the HER2 protein (18–20). It is possible that these genetic variations are population-specific events (i.e., genetic variants whose allele frequencies are significantly higher in one population compared to others) that could contribute in part to the differences in the prevalence of HER2-positive breast tumors among population groups (21). Due to the clinical implications of the HER2 expression in breast tumors (6, 22), the aim of this review was to describe the prevalence of HER2-positive breast tumors by ethnicity, along with genetic variants located at ERBB2 or nearby regions, that might affect its protein expression, with a special focus on reports for Asian and Latina women. We expect that this review will contribute to a better understanding of the underlying biology of HER2 expression in breast tumors, and the possible mechanisms that could explain the differences in the distribution of HER2-positive subtypes among various population groups.
2 Material and methods
The literature search was conducted in accordance with the Preferred Reporting Items for Systematic Reviews and Meta-Analyses (PRISMA) guidelines (Figure 1). The literature search was conducted using PubMed (NIH). We included original articles in English that assessed differences in HER2-positive breast tumors by ethnicity. This initial search included the Medical Subject Headings (MeSH) “breast cancer”, “HER2 subtype”, “populations” OR “race” OR ethnicity”, and “incidence”. To refine the search, the terms “review” and “trial” were excluded. 89 publications were retrieved from this search, and 14 articles that explicitly reported breast cancer subtype distribution among several ethnic groups were included. Studies where HER2 subtype prevalence was described among one single population were excluded as these articles do not allow comparisons among several groups. An additional search was conducted for articles that assessed genetic ancestry association with ERBB2/HER2 expression and/or its molecular features (amplification status, copy number variations, etc), using the MeSH words “genetic ancestry” “ERBB2”, “HER2 expression” and “breast cancer”, excluding the word “review”. A total of 16 publications were retrieved and 7 of these were included. Studies that evaluated the distribution of genetic variants or SNPs in ERBB2 according to HER2 expression were also included. The MeSH words used were “HER2”, “ERBB2”, “SNPs OR polymorphisms”, and “breast cancer”; the terms “review” and “trial” were excluded. We focused mainly on SNPs located at ERBB2 or nearby regions. 56 publications were retrieved, of which 5 were included. We did not limit the search by date. In total, 26 articles were selected for our review.
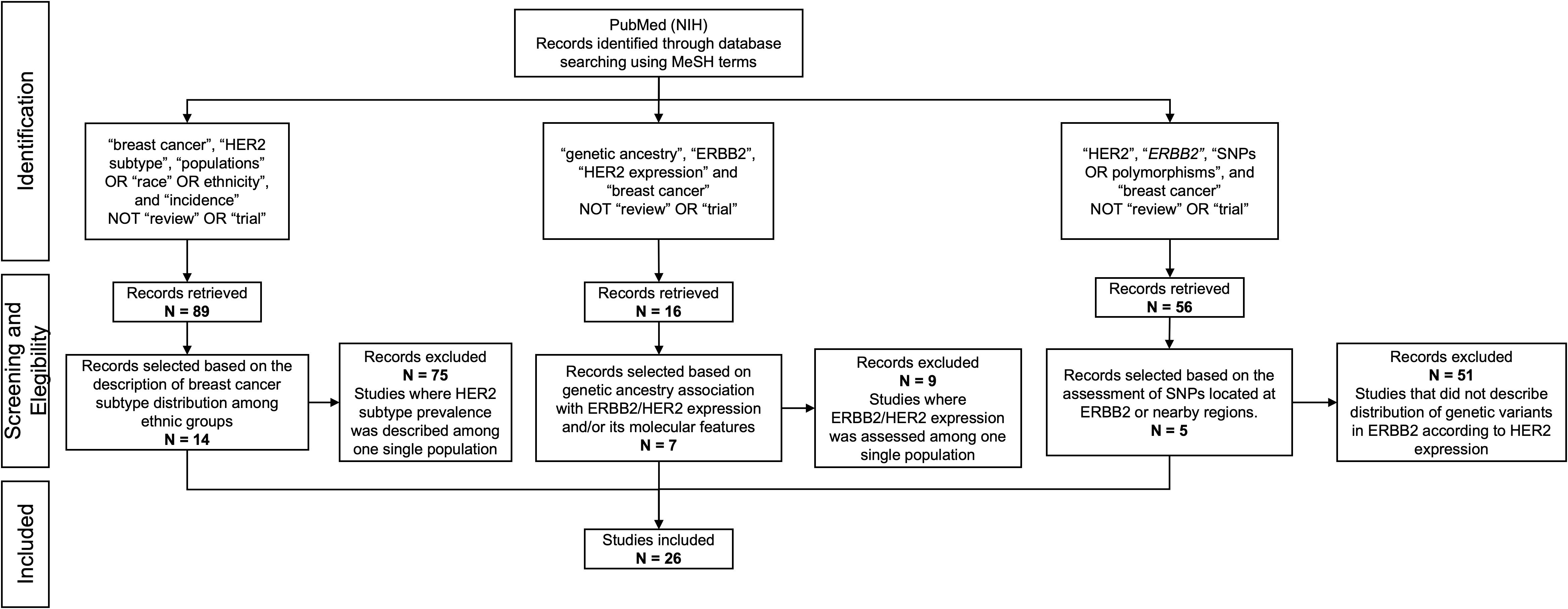
Figure 1. PRISMA flow diagram showing the database searches, the number of articles screened for eligibility, and the final number of full-text articles included.
3 Results
3.1 Prevalence of HER2-positive subtypes (luminal/HER2 and HER2-enriched) by ethnic group
After the molecular characterization of breast tumors published by Perou et al. (23), differences in the distribution of intrinsic subtypes by ethnic groups have been widely reported (8, 24). It is well known that African American (AA) women are more likely to develop TN breast cancer, whereas NHW women have higher odds for luminal-like subtypes (25). Regarding HER2-positive tumors, its distribution among populations is less clear (Tables 1, 2).
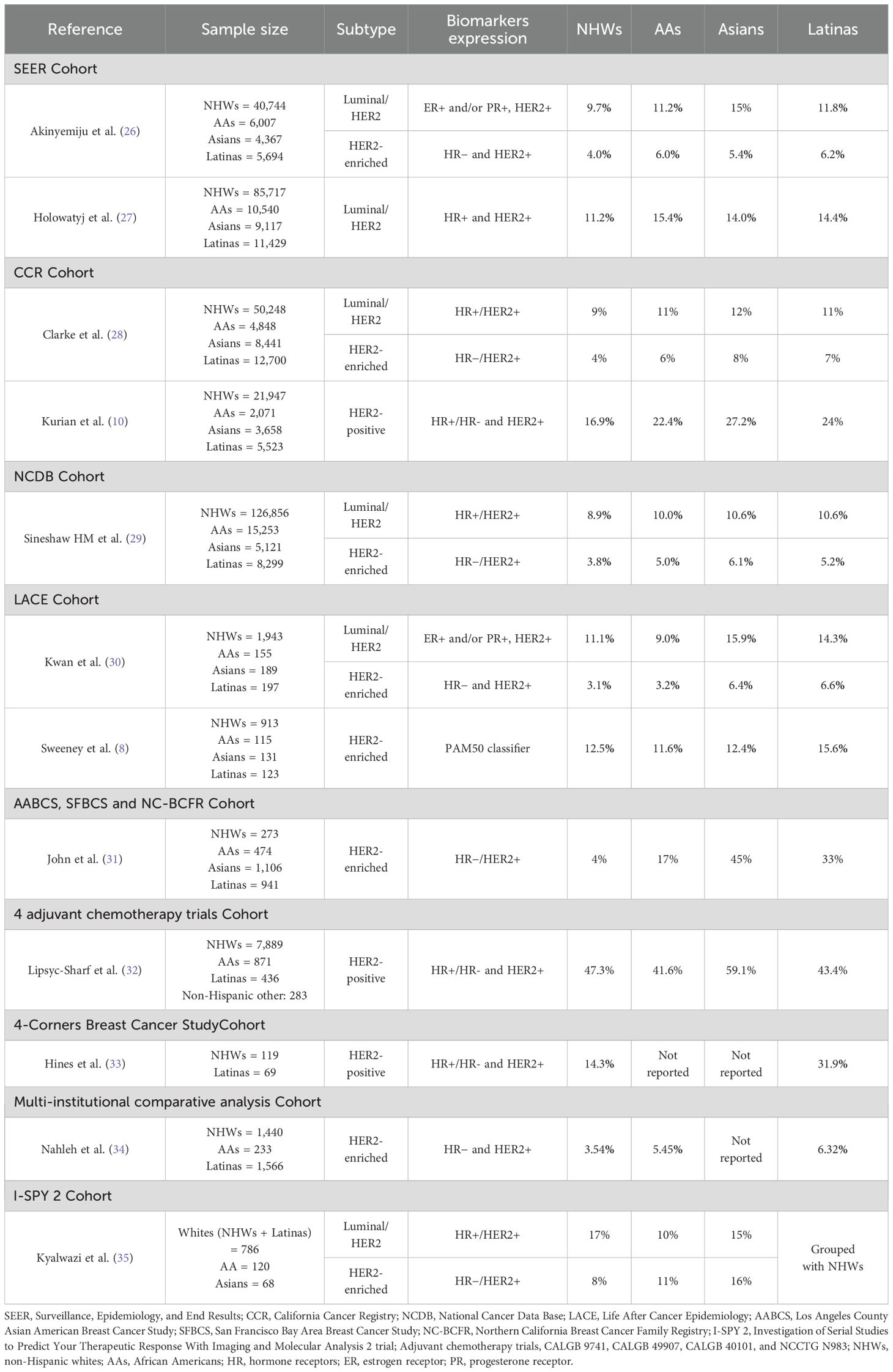
Table 1. Prevalence of HER2-positive breast tumors among different ethnic groups.
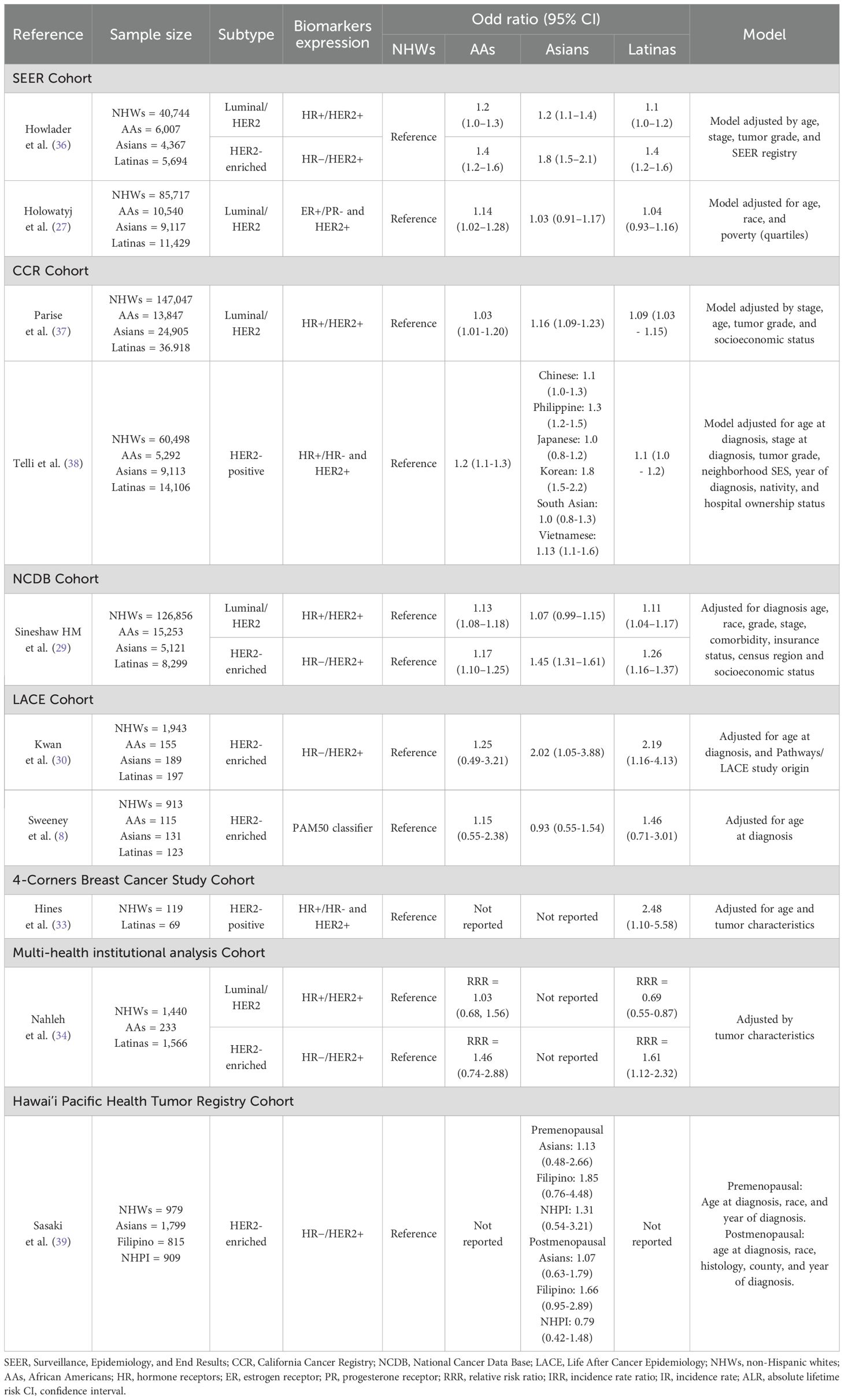
Table 2. Differences in the odds for HER2-positive subtypes (luminal/HER2 or HER2-enriched) according to population groups.
3.1.1 Overall trends from population and hospital-based registries
The Surveillance, Epidemiology, and End Results (SEER) registry, a large dataset that collected clinical and pathological information on cancer patients diagnosed in the United States (U.S) from 2010 to 2015, is estimated to cover approximately 97% of the incident cancers within the catchment area zones (40). In that sense, it gathers patients from different ethnic groups and represents a valuable resource to study breast cancer subtype distribution among several populations. SEER-based studies have consistently reported a higher prevalence (Table 1) and greater odds (Table 2) for HER2-positive subtypes in Asian, AA and Latina women. Akinyemiju et al. (26) showed a higher proportion of the luminal/HER2 subtype in Asian (15%), Latina (11.8%), and AA women (11.2%), compared to NHWs (9.7%). Moreover, specifically for the HER2-enriched subtype, they reported that Latinas presented the highest prevalence among all population groups (6.2%), followed by the AAs (6.0%), Asians (5.4%) and NHWs (4.0%). In the same way, Howlader et al. (36) reported a higher risk for HER2-positive subtypes in Latinas, either for the luminal/HER2 (odd ratio (OR)=1.1, 95% confidence interval (CI), 1.0–1.2) or the HER2-enriched subtype (OR=1.4, 95% CI, 1.2–1.6), compared to NHWs, in a model adjusted by age, stage, tumor grade, and SEER registry. Similar results were found in Asian and AA women for the luminal/HER2 (Asians: OR=1.2, 95% CI, 1.1–1.4; AAs: OR=1.2, 95% CI, 1.0–1.3) and the HER2-enriched subtypes (Asians: OR=1.8, 95% CI, 1.5–2.1; AAs: OR=1.4, 95% CI, 1.2–1.6). Furthermore, Holowatyj et al. (27) also reported a higher prevalence of the luminal/HER2 subtype in AAs (15.4%), Latinas (14.4%), and Asians (14.1%), compared with NHW women (11.2%), along with a statistically significant association between AA ethnicity and the luminal/HER2 subtype (OR=1.14, 95% CI, 1.02–1.28); they also found a tendency in Latinas and Asian women for having this subtype, using NHWs as the reference group (Latinas: OR=1.04, 95% CI, 0.93–1.16; Asians: OR= 1.03, 95% CI, 0.91–1.17).
Data from the National Cancer Data Base (NCDB) (29), a hospital-based but much larger U.S national cancer registry that covers approximately 70% of all newly diagnosed cancer cases in the country (41), included 260,174 breast cancer patients and consistently revealed that Latinas and AAs are more likely to develop luminal/HER2 tumors (Latinas: OR=1.11, 95% CI, 1.04-1.17; AAs: OR=1.13, 95% CI, 1.08–1.18) and the HER2-enriched subtype (Latinas: OR=1.26; 95% CI 1.16–1.37; AAs: OR=1.17, 95% CI, 1.10–1.25), compared to NHWs. This was also observed for Asian women, who presented the highest odds for HER2-enriched tumors among all populations (OR=1.45, 95% CI, 1.31–1.61). These findings in Asians were replicated by John et al. (31), in a cohort derived from three population-based studies—the Los Angeles County Asian American Breast Cancer Study (AABCS), the San Francisco Bay Area Breast Cancer Study (SFBCS), and the Northern California Breast Cancer Family Registry (NC-BCFR). Their analysis revealed that the highest prevalence of HER2-enriched tumors was observed in Asian women (45%), followed by Latinas (33%), whereas the prevalence among NHWs was reported at only 4%.
Reports on differences in HER2-positive breast tumors among ethnic groups have also been published using U.S state-wide cancer-based studies, like the California Cancer Registry (CCR), which gathers high quality data from all cancer patients diagnosed in 48 of California’s 58 counties. Clarke et al. (28) found a higher proportion of HER2-positive subtypes (either HER2-enriched or luminal/HER2) in Asians, Latinas, and AAs compared to NHWs (20%, 18% and 17% vs. 13%, respectively). In the same way, Telli et al. (38) also reported a higher likelihood of having either luminal/HER2 or HER2-enriched breast tumors in AAs and Latina women from the CCR (AAs: OR= 1.2, 95% CI, 1.1-1.3; Latinas: OR= 1.1, 95% CI, 1.0-1.2). Correspondingly, Parise et al. (37) reported higher odds for luminal/HER2 tumors in Asian and Latina women compared to NHWs, after adjusting for stage, age, tumor grade, and socioeconomic status (Asians: OR=1.16, 95% CI, 1.09-1.23; Latinas: OR=1.09, 95% CI, 1.03-1.15).
These results in Asians, AAs and Latinas have even been replicated in women from different parts of the U.S. Kwan et al. (30) conducted a study in 2,544 women from the Life After Cancer Epidemiology (LACE) study, which includes patients from the Kaiser Permanente Northern California Cancer Registry (KPNCAL) and the Utah cancer registry (UCR). They reported that Asian and Latina women had the highest prevalence of luminal/HER2 (Asian: 15.9%, Latina: 14.3%, NHWs: 11.1%, AAs: 9.0%; p=0.18) and HER2-enriched subtypes (Asian: 6.4%, Latina: 6.6%, NHWs: 3.1%, AAs: 3.2%; p=0.03) among all population groups; and also presented the highest odds for HER2-positive tumors (Asians: OR=2.02, 95% CI, 1.05-3.88; Latinas: OR=2.19, 95% CI, 1.16-4.16).
These findings consistently demonstrate a clear association between HER2-positive tumors and AA, Asian, and Latina ethnicities, suggesting the presence of disparities in the distribution of breast cancer subtypes among minority groups in the U.S. However, it is important to note that breast tumor subtype classification is often conducted based on the IHC expression of ER, PR and HER2 biomarkers (42). This approach is subjected to multiple limitations such as the variability of the staining and scoring by the pathologist, and cut-off points to define positive or negative cases, particularly for ER and PR (43). In contrast, gene expression-based assays account for a better approach for subtype classification. A study conducted by Sweeney et al. (8) based on the LACE registry described the overall distribution of breast cancer subtypes in relation to clinicopathologic categories among 1,319 women from different ethnic groups, applying the PAM50 classifier. The results showed that Latinas tend to present with non-luminal A subtypes, including HER2-enriched tumors (OR=1.46, 95% CI, 0.71–3.01), compared to NHWs. The same tendency was observed for AAs (OR=1.15, 95% CI, 0.55-2.38) but not for the Asian group (OR=0.93, 95% CI, 0.55-1.54), which may be explained by the small sample size of this group in the study and/or differences in the method used to define breast cancer subtypes. Gene expression-based technologies, such as PAM50, reflect in a better way the tumors biology, therefore, it is still necessary to keep exploring differences in breast cancer subtype classification among ethnic groups using molecular classifiers.
Most of the population-based research presented above includes large datasets of Caucasian patients, while minority groups such as Asians and Latinas are consistently underrepresented. This poses significant challenges for epidemiological analyses, particularly in terms of statistical power and the ability to draw conclusions that accurately reflect the realities of these populations. The same issue extends to genomic research, where limited diversity can obscure population-specific genetic variants (44). Therefore, improving the inclusion of these groups is essential to identify relevant biomarkers, refine informed therapeutic strategies, and ultimately reduce health disparities.
3.1.2 Focused reports on the distribution of HER2-positive breast cancer in Asian and Latina women
In light of these findings, particular attention has been given to the Asian ethnic group to further investigate disparities among its subpopulations. For instance, Parise et al. (37) specifically focused on Asian ethnicities and reported a strong association between HER2-positive tumors and the Korean self-reported population (OR = 1.63, 95% CI: 1.38–1.99). Similarly, another CCR-based study who paid special attention to Asian also showed that of all population groups, Koreans present the highest prevalence (36%) and odds for HER2-overexpressing tumors (OR= 1.8, 95% CI, 1.5-2.2), followed by Philippines (OR= 1.3, 95% CI, 1.2-1.5) and Vietnamese (OR= 1.3, 95% CI, 1.1-1.6) (38). Comparable results were reported by Sasaki et al. (39) for premenopausal breast cancer patients across different Asian ethnicities, with ORs of 1.13 (95% CI: 0.48–2.66) for Asians, 1.85 (95% CI: 0.76–4.48) for Filipinos, and 1.31 (95% CI: 0.54–3.21) for Native Hawaiian or Pacific Islander women. These findings underscore the importance of disaggregated analyses among Asian subpopulations to identify significant variations in the prevalence and odds of HER2-positive breast cancer, differences that may otherwise be overlooked when these groups are studied collectively.
However, some reports fail to account for the significant heterogeneity within Asian subpopulations, leading to analyses that combine Asians with other ethnic groups. This was the case for a large study based on four adjuvant chemotherapy trials (CALGB 9741, CALGB 49907, CALGB 40101, and NCCTG N9831), which included 9,479 breast cancer patients (32). Among this population, only 2.98% corresponded to Asians and other ethnicities, such as American Indian or Alaska Native, Native Hawaiian, or Other Pacific Islanders. As a result, these women were grouped and analyzed collectively as “non-Hispanic other”. In this category, the prevalence of HER2-positive subtypes was notably higher (59%) compared to NHWs (47.3%), AAs (41.6%), and Latinas (43.4%), leaving unclear the potential disparities that might exist within the ethnicities included in the “non-Hispanic other” group (Table 1).
Grouping multiple ethnic populations into a single broad category results in a heterogeneous cohort. While this approach may increase the overall sample size and facilitate statistical analysis, it limits the ability to uncover meaningful differences or disparities that may exist among the distinct ethnic subgroups included within the aggregated category.
A similar scenario was observed for Latinas in a recent study based on the Investigation of Serial Studies to Predict Your Therapeutic Response with Imaging and Molecular Analysis 2 (I-SPY 2) (35). The study included 990 breast cancer patients from various self-identified ethnicities across different U.S. territories. However, due to the low representation of Latina patients (12% of the cohort), they were primarily grouped with the White women cohort. As a result, the distribution of the luminal/HER2 subtype revealed a higher prevalence among the White cohort (17%) compared to the Asian (15%) and African American (10%) cohorts. This highlights the challenges in drawing conclusions and comparing results across studies due to the mixed classification of patients within ethnic groups. It also reinforces the ongoing issue of minority underrepresentation in epidemiological research.
Although there is still an issue of underrepresentation of Latinas in the SEER and other population- and hospital-based studies (45, 46), recent efforts reflect a trend toward creating more representative population registries, with a particular focus on minorities. For instance, Nahleh et al. (34) conducted a multi-health institutional analysis where 45.5% (1,566/3,441) of the cases were U.S Latinas from Texas and California; other populations groups included were NHWs (1,440: 41.8%) and AAs (233: 6.7%). They evaluated differences in tumor characteristics among ethnic groups and found that, compared to NHWs, Latinas have a higher relative risk ratio (RRR) for HER2-enriched tumors (RRR=1.61, 95% CI, 1.12-2.32, p=0.01) after having adjusted by age at diagnosis, histological subtype, chemotherapy, and surgery. Interestingly, the association between luminal/HER2 tumors and Latin ethnicity was in the oppositive direction (RRR=0.69, 95% CI, 0.55-0.87, p=0.001), which might be related to the protective association for luminal breast tumors found in the Latinas group, compared to the NHW women (RRR= 0.76, 95% CI, 0.64-0.9, p=0.002). On the other hand, no statistically significant associations were found in AA breast cancer patients for neither of the HER2-overexpressing tumors (luminal/HER2 = 1.03, 95% CI, 0.68-1.56; p= 0.862HER2-enriched=1.46, 95% CI, 0.74-2.88, p= 0.273).
Specifically for Latinas, it is important to highlight that often in these epidemiological studies the prevalence of HER2-positive subtypes might be underestimated, given that HER2 status is less likely to be tested in this population group (47), and a higher proportion of these cases are excluded due to the lack of complete information. Even with these limitations, studies with small sample sizes have found similar results. For instance, a 4-Corners Breast Cancer-based study that only included 285 women (NHWs=119 and Latinas=69) reported a higher prevalence of HER2-positive tumors in Latinas compared to NHWs (31.9% vs 14.3%, p<0.001, respectively), and a higher likelihood of having HER2-positive tumors for this population group (OR=2.48, 95% CI, 1.10-5.58) in a model adjusted for age and tumor characteristics (33).
Other factors related to lifestyle (such as diet, exercise, alcohol consumption, tobacco use, among others) and reproductive behaviors (including contraceptive use, parity, and breastfeeding) have been explored in the search to elucidate potential contributors to health disparities in breast cancer (48, 49) as well as for each intrinsic subtype (50, 51), and are describe extensively elsewhere. Among these, in relation to HER2-positive tumors, it has been shown that having a family history of cancer, higher breast density, and obesity (body mass index >30) increases the odds of having this particular subtype (51). As these risk factors vary by ethnic group, it is possible that differences in their prevalence among populations may help explain the variation in the distribution of HER2-positive tumors described above (52).
Other reports have focused not only on the expression of HER2 but on the molecular features of its codifying gene, ERBB2, and its relationship with biological contributors that help explain differences in its distribution among ethnic groups. Therefore, we provide a description of the studies so far that have assessed ERBB2 by ethnicity, and also, by genetic ancestry, as a more accurate definition of people’s ancestral origin.
3.2 Molecular features of ERBB2 by ethnicity and genetic ancestry
Few studies have reported differences in ERBB2 gene expression and its molecular features by population groups. Kan et al. (53) conducted a study to evaluate molecular differences between an Asian cohort of women from Korea (n=187) with breast cancer, and a group of Caucasians (n=745) and AAs (n=158) from The Cancer Genome Atlas (TCGA) database, and showed a higher frequency of somatic alterations (mutations and copy number variations (CNVs)) in ERBB2 in the Asian cohort (20% vs. 9.1%, respectively), along with a higher proportion of HER2-enriched (8% vs 2.9%, respectively) and luminal/HER2 subtypes (14.4% vs. 10.4%, respectively) in Asian women, compared to the TCGA group (Caucasians + AAs). Similarly, Pan et al. (12) reported a higher level of amplifications in the ERBB2 region (17q12) in a cohort of 560 Asian women from Malaysia, along with a higher prevalence of the HER2-enriched molecular subtype assessed by PAM50 in Asian women compared to a group of Caucasians from the TCGA (23.3% vs 9.9%, respectively).
In terms of gene expression, Grunda et al. (54) reported a higher expression of the ERBB2 gene in Caucasians when compared to AA breast cancer patients (ERBB2 mean fold change (FC) in Caucasians: 1.61 vs AAs: 0.63, p=0.012), and other genes associated with prognosis (ESR1 and GATA3), disease progression (HSPB1 and SERPINA3) and response to chemotherapy (CLDN7 and DLC1). Even though the epidemiological studies described above usually report higher prevalence of HER2-positive tumors among AAs compared to NHWs, it is possible that Grunda’s et al. (54) conflicting results are related to the small sample size of the study (AAs= 11, Caucasians=11). It is also worth mentioning that the later study only assessed gene expression, whereas prevalence studies reported above are based on HER2 protein expression. Therefore, it is possible that several post-transcriptional mechanisms might be exerting an effect on ERBB2/HER2 expression, this way contributing to the reported differences between these population groups (55, 56).
Self-reported ethnicity comprehends a heterogeneous group with mixed ancestries (57). This has drawn attention to the importance of analyzing breast cancer molecular features by genomic estimated race or genetic ancestry. This estimation is based on the analysis of ancestry-informative markers (AIMs), which capture differences in allele frequencies across major continental populations. This approach allows ancestry to be quantified as an objective and continuous variable, enabling researchers to assess how varying proportions of European, Indigenous American (IA), African, and Asian ancestry may correlate with disease phenotypes (44).
This approach was applied in a recent study by Miyashita et al. (58), which assessed the genetic ancestry of 3,433 breast cancer patients from the Tempus Database. The study differentiated African ancestry patients (>20% African, <10% IA ancestry, and a combined African plus European likelihood >70%) from European ancestry patients (>80% European and <10% IA ancestry). Interestingly, the study found a higher enrichment of several hallmark and oncogenic gene signature sets in the European cohort compared to African ancestry patients, including the ERBB2 gene set ERBB2_UP.V1_UP, which refers to a group of genes defined in the Gene Set Enrichment Analysis (GSEA) database that are consistently upregulated in the context of ERBB2 pathway activation. However, this finding was only observed in stage IV tumors with an HR+/HER2- subtype. Although the enrichment of this gene set does not necessarily indicate that the ERBB2 gene itself is expressed, it reflects the upregulation of genes associated with ERBB2 activation and signaling. In this context, these findings strongly suggest the presence of important differences in the tumor biology of breast cancer patients, potentially driven by their genetic ancestry.
A study conducted in Latinas from Colombia by Serrano-Gomez et al. (16) assessed the association between genetic ancestry and gene expression in 42 women with luminal breast tumors. When patients were stratified according to the average fraction of IA ancestry (low IA <36%, high IA ≥ 36%), the high IA ancestry group showed a higher expression of ERBB2, along with other genes located at the HER2 amplicon, such as GRB7 and MIEN1. In line with these results, Marker et al. (21) reported an association between the HER2-enriched subtype and the IA ancestry fraction in a cohort of 1,312 Peruvian women with breast cancer. They reported that the odds of presenting a HER2-enriched subtype increased by a factor of 1.30 per every 10% increase in IA ancestry (p=0.004) after adjusting for age at diagnosis, African ancestry fraction and height. This association was replicated in other sets of patients from Mexico (OR =1.20; 95% CI, 0.90–1.59) and Colombia (OR = 1.28; 95% CI, 1.03–1.60). In the same way, a separate study in Colombian women that included breast cancer patients from different health institutions around the country also reported a suggestive association between the IA ancestry and both HER2-enriched (OR =1.18; 95% CI, 0.50 - 2.67) and luminal B/HER2 subtypes (OR =1.61; 95% CI, 0.84 - 3.09), after adjusting for health institution, age at diagnosis, and clinical stage (59). This positions genetic ancestry as a potentially valuable tool in the clinical setting, as women with a higher contribution of IA ancestry may exhibit more frequent ERBB2 overexpression, potentially making them more responsive to HER2-low targeted therapies, such as antibody-drug conjugates (ADCs) like T-DM1 and T-DXd.
Even though only a few studies have assessed ERBB2 differential expression between population groups, the results gathered so far suggest that there are differences in gene expression by genetic ancestry that may account for differences in the biology, prognosis, and outcomes of breast cancer between population groups (52). For instance, epidemiological studies comparing population groups have reported that Latina women are more likely to present clinical characteristics associated with poor prognosis, such as being diagnosed at advanced stages (60). In line with this, a higher risk of breast cancer mortality has also been reported for Latina women compared to NHW women (61), which may ultimately be related to a higher frequency of more aggressive subtypes, like HER2-positive tumors. It is possible that variations in the distribution of the HER2-positive subtypes by ethnicity could be partly related to the presence of genetic variants.
3.3 HER2 overexpression according to genetic variants in ERBB2
Genetic variants are defined as specific changes in a genomic region that are partly responsible for phenotype differences reported between population groups, such as differences in susceptibility to various diseases, including cancer (62). Genetic variants or SNPs in ERBB2 have been associated with breast cancer risk (63–65), therapy response and resistance to anti-HER2 treatments (66, 67). Given that most of the genetic variants reported so far in ERBB2 are located at the transmembrane domain coding region, their main effect is related to changes in HER2 protein activity (68). In addition to this, other studies have found differences in HER2 expression according to the ERBB2 SNPs genotype (Table 3).
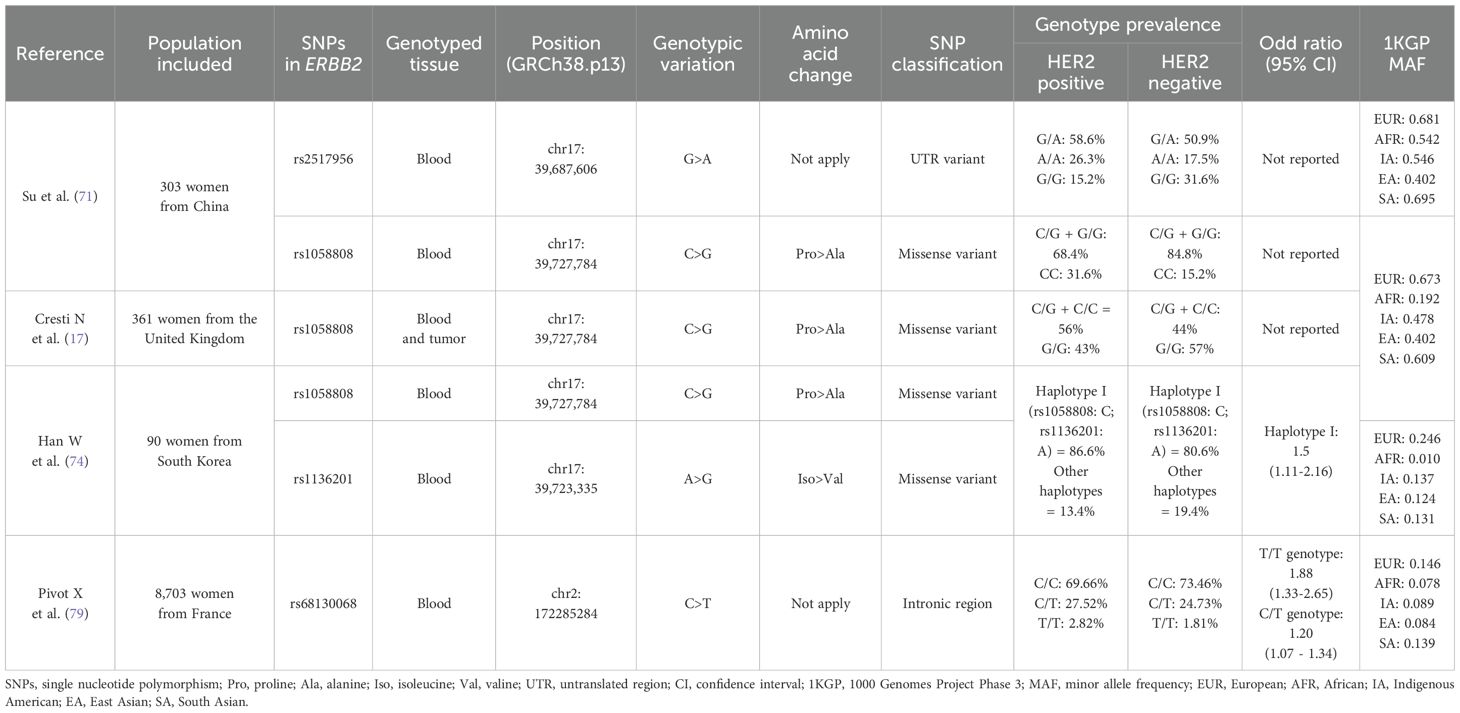
Table 3. Differences in ERBB2 SNPs genotype according to HER2-expression, and risk of HER2-positive tumors, among various population groups.
Su et al. (71) evaluated the expression of the HER2 protein according to the SNP rs1058808 genotype, a genetic variant (C>G) located at the ERBB2 gene residue 1170 that encodes for either a proline (C allele) or alanine (G allele) at the C-terminal region of the HER2 protein (69). This study that included 303 Chinese women reported that patients with the C/G and G/G germline genotypes had a higher frequency of HER2-positive tumors, compared to patients with the C/C genotype (C/G: 58.6% and G/G: 26.3% vs. C/C: 15.2%, p = 0.007). This difference was also statistically significant under the dominant model (C/G + G/G: 84.8% vs. C/C: 15.2%, p = 0.003). Nonetheless, opposite results were reported in a European population, in a study that included 361 breast cancer patients from the United Kingdom. They analyzed germline and tumor genotype of numerous SNPs, including the rs1058808 and HER2 expression by IHC and found a significantly higher proportion of HER2 positive tumors in the proline carriers (either C/G or C/C), compared to the alanine carriers (G/G genotype) (56% vs. 43%, respectively, p=0.015) (17). These contradictory results might be partly explained by the differences in the rs1058808-G minor allele frequency (MAF) between both populations (1000Genomes project (1KGP): Asians G=0.4018 vs Europeans G=0.6730) (72), coupled with the flip-flop phenomenon, where the same allele confers risk in one population but is protective in another (73), although it is also possible that sample size differences between both studies might contribute as well to these conflicting results. On the other hand, these studies only assessed the differences in the proportion of HER2 positive cases according to rs1058808 genotype, therefore, it is likely that other biological factors, meaning additional genetic variants beyond rs1058808, might also contribute to the expression of the HER2 protein in these diverse populations.
The same genetic variant (rs1058808) was also evaluated in another study by Han W et al. (74) in 90 breast cancer women from South Korea. The association with HER2 expression was assessed for the rs1058808 SNP and at the same time, for five other SNPs at the ERBB2 gene, all together as a haplotype. They reported that patients carrying the germline haplotype configuration I, which includes the rs1058808-C allele, were 1.5 times more likely to develop tumors with HER2 overexpression, compared to patients without this specific genotype (OR=1.5; 95% CI, 1.11-2.16, p= 0.009). This haplotype also included the rs1136201 SNP (I655V), a genetic variation (A>G) that leads to an isoleucine (I)-to-valine (V) substitution at codon 655 within the transmembrane domain of the ERBB2 gene (63). This haplotype containing the rs1058808-C and rs1136201-A alleles, was found associated with HER2 overexpression among Korean breast cancer patients (74). According to the 1KGP, both SNPs alleles are considerably more frequent among East Asians, compared to other populations such as Europeans (East Asians: rs1058808-C=0.571 and rs1136201-A=0.8760 vs. Europeans: rs1058808-C= 0.33232and rs1136201-A=0.7545) (72, 75). In that sense, these SNPs genotypes might contribute to the higher prevalence of the HER2-positive tumors reported above among Asian populations when compared to Europeans. Several meta-analyses have reported an association between the rs1136201 genotype and breast cancer risk (63, 76), nonetheless, the impact of this genetic variation on the expression of its protein remains unclear.
Other types of genetic variants that localize at non-coding gene sequences have been reported to affect gene expression (70, 77). One of these genetic variants, the rs2517956, a 2 Kb upstream variant (G>A) of the ERBB2 gene (78), was genotyped in 303 breast cancer patients from China, and evaluated according to the HER2 protein expression by IHC. This study found a higher proportion of HER2 positive tumors among patients with the A/A germline genotype, compared to homozygous G/G breast cancer patients (26.3% vs 15.2%, respectively, p=0.008) (1). However, the highest proportion of HER2-positive tumors was actually found among cases with the heterozygous A/G genotype (58.6%), suggesting this association might have an alternative underlying mechanism to the additive model. Additionally, it is worth noting that the overall rs2517956 allele’s frequency reported on the 1KGP (78) is relatively uniform across population, suggesting that this variant is unlikely to contribute to differences in HER2 expression in breast tumors among ethnic groups.
An important effort to elucidate potential genetic variants that confer risk for HER2-positive breast tumors was made by the French National Cancer Institute through a case-case Genome-Wide Association Study (GWAS) in over 8,703 women (79). They identified a SNP (rs68130068) located at chromosome 2 within an intronic region, which was potentially associated with HER2-positive breast tumors. Even though this SNP genotype (C>T) does not affect the sequence of any nearby gene, they reported that patients with the homozygous genotype for the minor allele (T/T) were 88% more likely to have HER2-positive breast tumors (OR=1.88; 95% CI, 1.33-2.65, p=0.00033), compared to patients with the C/C genotype. Heterozygous (C/T) patients also showed higher risk for HER2-positive tumors when compared to the C/C group (OR=1.20; 95% CI, 1.07-1.34, p=0.0013). Nonetheless, when the genome-wide association was tested, the rs68130068 SNP only reached a borderline level of significance (p=3.6x10-6).
Even though the mechanism by which the rs68130068 variant and others that are located at non-codifying genome sequences might contribute to the tumor phenotype is not entirely understood, it has been hypothesized that these SNPs regulate gene expression through several allele-specific mechanisms according to the distance from the regulated gene (80). SNPs that map close are referred to as cis- expression quantitative trait loci (eQTLs), whereas SNPs that map far from the regulated genes are referred to as trans-eQTLs (81). In that sense, it is possible that these eQTLs, whose allele frequencies vary across specific populations or ancestry groups, may influence the transcription of ERBB2 or other genes upregulated in HER2-positive tumors. This could provide a potential biological mechanism underlying the disparities observed between populations for this particular breast cancer subtype.
Other studies have also reported differences in HER2 expression in breast tumors according to the genotype of SNPs located at other genes with roles related to gene regulation (FOXP3, NOTCH3) (82, 83), hormone signaling (ESR1 and CYP19A1) (84–86) and proliferation/survival control (WISP-1, CASP8, KRAS, TGFBR2, VEGF-A and CCND3) (87–92). Possibly, the impact of these SNPs on the HER2 expression arises from their regulatory effects on the gene in which they are located, indirectly influencing the expression of other genes and proteins, such as ERBB2/HER2. In this context, differences in the genotype frequencies of these SNPs across populations may partially explain the disparities reported in HER2-positive breast tumors among ethnic groups. Further research is needed to deepen our understanding of health disparities between populations and to shed light on the underlying biology of HER2 expression in breast tumors.
4 Limitations and perspectives
This review certainly has some limitations, primarily related to topics that were not addressed in depth. For instance, while we reviewed differences in HER2-positive breast tumors across population groups, such differences may also influence patient outcomes and contribute to disparities in survival rates among these groups. However, these issues have already been thoroughly explored in other publications (52, 93). Similarly, although we describe studies that have analyzed genetic ancestry and its potential influence on the prevalence of HER2-positive tumors, the underlying genetic mechanisms are not discussed in detail, as this remains an area of ongoing investigation. Nevertheless, we hypothesize that this may be related to the presence of certain SNPs whose genotype frequencies vary across populations and have been associated with HER2 expression. Still, many other genetic variants, within different genes and genomic regions, may also be involved. As this is a broad and evolving topic, further systematic reviews are needed to consolidate and analyze the growing body of literature concerning the molecular epidemiology of HER2-positive breast cancer.
5 Conclusions
We reviewed studies that reported differences in HER2-positive breast cancer subtypes among population groups and consistently found a significantly higher prevalence of these tumors in Latina and Asian women. In some reports, this was also observed in AA women. Furthermore, studies that analyzed breast cancer patients according to genetic ancestry showed that a higher IA ancestry fraction is associated with ERBB2/HER2 expression. These results suggest that variations in the distribution of HER2-positive subtypes by ethnicity could be partly related to differences in allele frequencies of certain genetic variants among populations. We reviewed SNPs that can be found directly at the ERBB2 gene sequence, either affecting the protein structure or its transcriptional regulation. However, other reports have described SNPs located at independent genes that, due to their biological functions, may also affect HER2 expression in an indirect manner. Further studies with larger sample sizes are still needed to elucidate if differences in some of the aforementioned SNPs genotypes can actually contribute to a higher risk of developing HER2-positive breast cancer.
Author contributions
LR-V: Writing – review & editing, Writing – original draft, Methodology, Investigation, Conceptualization, Data curation. LB-R: Writing – original draft, Methodology, Investigation, Conceptualization. PL-C: Writing – review & editing. DB-L: Funding acquisition, Resources, Writing – review & editing. SS-G: Data curation, Supervision, Conceptualization, Funding acquisition, Resources, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was funded and funded by the Colombian National Cancer Institute (H1905030037-14), the Science and Technology Colombian Ministry - MINCIENCIAS (Programa de Becas de Excelencia Doctoral del Bicentenario 2do corte), and the Pontificia Universidad Javeriana, Bogotá D.C, Colombia.
Acknowledgments
We acknowledge the Colombian National Cancer Institute, the Science and Technology Colombian Ministry – MINCIENCIAS, and the Pontificia Universidad Javeriana, Bogotá D.C, Colombia, for supporting this work.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Bray F, Laversanne M, Hyuna S, Ferlay J, Siegel RL, Soerjomataram I, et al. Global cancer statistics 2022: GLOBOCAN estimates of incidence and mortality worldwide for 36 cancers in 185 countries. CA Cancer J Clin. (2024) 74:229–63. doi: 10.3322/CAAC.21834
2. Turashvili G and Brogi E. Tumor heterogeneity in breast cancer. Front Med (Lausanne). (2017) 4:227. doi: 10.3389/fmed.2017.00227
3. Harbeck N and Gnant M. Breast cancer. Lancet. (2017) 389:1134–50. doi: 10.1016/S0140-6736(16)31891-8
4. Prat A, Pineda E, Adamo B, Galván P, Fernández A, Gaba L, et al. Clinical implications of the intrinsic molecular subtypes of breast cancer. Breast. (2015) 24:S26–35. doi: 10.1016/j.breast.2015.07.008
5. Prat A, Carey LA, Adamo B, Vidal M, Tabernero J, Cortés J, et al. Molecular features and survival outcomes of the intrinsic subtypes within HER2-positive breast cancer. J Natl Cancer Inst. (2014) 106(8):dju152. doi: 10.1093/jnci/dju152
6. Waks AG and Winer EP. Breast cancer treatment: A review. JAMA - J Am Med Assoc. (2019) 321:288–300. doi: 10.1001/jama.2018.19323
7. Godoy-Ortiz A, Sanchez-Muñoz A, Parrado MRC, Álvarez M, Ribelles N, Dominguez AR, et al. Deciphering HER2 breast cancer disease: Biological and clinical implications. Front Oncol. (2019) 9:1124. doi: 10.3389/fonc.2019.01124
8. Sweeney C, Bernard PS, Factor RE, Kwan ML, Habel LA, Quesenberry CP, et al. Intrinsic subtypes from PAM50 gene expression assay in a population-based breast cancer cohort: differences by age, race, and tumor characteristics. Cancer Epidemiol Biomarkers Prev. (2014) 23:714–24. doi: 10.1158/1055-9965.EPI-13-1023
9. Chen L and Li CI. Racial disparities in breast cancer diagnosis and treatment by hormone receptor and HER2 status. Cancer Epidemiol Biomarkers Prev. (2015) 24:1666–72. doi: 10.1158/1055-9965.EPI-15-0293
10. Kurian AW, Fish K, Shema SJ, and Clarke CA. Lifetime risks of specific breast cancer subtypes among women in four racial/ethnic groups. Breast Cancer Res. (2010) 12:R99. doi: 10.1186/bcr2780
11. Parise CA, Bauer KR, and Caggiano V. Variation in breast cancer subtypes with age and race/ethnicity. Crit Rev Oncol Hematol. (2010) 76:44–52. doi: 10.1016/j.critrevonc.2009.09.002
12. Pan JW, Zabidi MMA, Ng PS, Meng MY, Hasan SN, Sandey B, et al. The molecular landscape of Asian breast cancers reveals clinically relevant population-specific differences. Nat Commun. (2020) 11:1–12. doi: 10.1038/s41467-020-20173-5
13. Martinez ME, Wertheim BC, Natarajan L, Schwab R, Bondy M, Daneri-Navarro A, et al. Reproductive factors, heterogeneity, and breast tumor subtypes in women of Mexican descent. Cancer Epidemiol Biomarkers Prev. (2013) 22:1853–61. doi: 10.1158/1055-9965.EPI-13-0560
14. Cruz GI, Martínez ME, Natarajan L, Wertheim BC, Gago-Dominguez M, Bondy M, et al. Hypothesized role of pregnancy hormones on HER2+ breast tumor development. Breast Cancer Res Treat. (2014) 137:237–46. doi: 10.1007/s10549-012-2313-0.HYPOTHESIZED
15. Yang XR, Sherman ME, Rimm DL, Lissowska J, Brinton LA, Peplonska B, et al. Differences in risk factors for breast cancer molecular subtypes in a population-based study. Cancer Epide-miol Biomarkers Prev. (2007) 16:439–82. doi: 10.1158/1055-9965.EPI-06-0806
16. Serrano-Gómez SJ, Sanabria-Salas MC, Garay J, Baddoo MC, Hernández-Suarez G, Mejía JC, et al. Ancestry as a potential modifier of gene expression in breast tumors from Colombian women. PloS One. (2017) 12:1–21. doi: 10.1371/journal.pone.0183179
17. Cresti N, Lee J, Rourke E, Televantou D, Jamieson D, Verrill M, et al. Genetic variants in the HER2 gene: Influence on HER2 overexpression and loss of heterozygosity in breast cancer. Eur J Cancer. (2016) 55:27–37. doi: 10.1016/j.ejca.2015.10.066
18. Bose R, Kavuri SM, Searleman AC, Shen W, Shen D, Koboldt DC, et al. Activating HER2 mutations in HER2 gene amplification negative breast cancer. Cancer Discov. (2013) 3:224–37. doi: 10.1158/2159-8290.CD-12-0349
19. Furrer D, Lemieux J, Côté MA, Provencher L, Laflamme C, Barabé F, et al. Evaluation of human epidermal growth factor receptor 2 (HER2) single nucleotide polymorphisms (SNPs) in normal and breast tumor tissues and their link with breast cancer prognostic factors. Breast. (2016) 30:191–6. doi: 10.1016/j.breast.2016.09.014
20. Davis MB, Liu X, Wang S, Reeves J, Khramtsov A, Huo D, et al. Expression and sub-cellular localization of an epigenetic regulator, co-activator arginine methyltransferase 1 (CARM1), is associated with specific breast cancer subtypes and ethnicity. Mol Cancer. (2013) 12:1–17. doi: 10.1186/1476-4598-12-40
21. Marker KM, Zavala VA, Vidaurre T, Lott PC, Vásquez JN, Casavilca-Zambrano S, et al. Human epidermal growth factor receptor 2-positive breast cancer is associated with indigenous american ancestry in latin american women. Cancer Res. (2020) 80(9):1893–901. doi: 10.1158/0008-5472.CAN-19-3659
22. Prat A, Pascual T, and Adamo B. Intrinsic molecular subtypes of HER2+ breast cancer. Oncotarget. (2017) 8:73362–3. doi: 10.18632/oncotarget.20629
23. Perou CM, Sørlie T, Eisen MB, van de Rijn M, Jeffrey SS, Rees CA, et al. Molecular portraits of human breast tumours. Nature. (2000) 406:747–52. doi: 10.1038/35021093
24. Kohler BA, Sherman RL, Howlader N, Jemal A, Ryerson AB, Henry KA, et al. Annual report to the nation on the status of cancer, 1975-2011, featuring incidence of breast cancer subtypes by race/ethnicity, poverty, and state. JNCI: J Natl Cancer Institute. (2015) 107:djv048. doi: 10.1093/jnci/djv048
25. Troester MA, Sun X, Allott EH, Geradts J, Cohen SM, Tse C-K, et al. Racial differences in PAM50 subtypes in the carolina breast cancer study. J Natl Cancer Inst. (2017) 110(2):176–82. doi: 10.1093/jnci/djx135
26. Akinyemiju TF, Pisu M, Waterbor JW, and Altekruse SF. Socioeconomic status and incidence of breast cancer by hormone receptor subtype. Springerplus. (2015) 4:508. doi: 10.1186/s40064-015-1282-2
27. Holowatyj AN, Ruterbusch JJ, Ratnam M, Gorski DH, and Cote ML. HER2 status and disparities in luminal breast cancers. Cancer Med. (2016) 5:2109–16. doi: 10.1002/CAM4.757
28. Clarke CA, Keegan THM, Yang J, Press DJ, Kurian AW, Patel AH, et al. Age-Specific incidence of Breast cancer Subtypes: Understanding the Black-White crossover. J Natl Cancer Inst. (2012) 104:1094–101. doi: 10.1093/jnci/djs264
29. Sineshaw HM, Gaudet M, Ward EM, Flanders WD, Desantis C, Lin CC, et al. Association of race/ethnicity, socioeconomic status, and breast cancer subtypes in the National Cancer Data Base (2010-2011). Breast Cancer Res Treat. (2014) 145:753–63. doi: 10.1007/S10549-014-2976-9
30. Kwan ML, Kushi LH, Weltzien E, Maring B, Kutner SE, Fulton RS, et al. Epidemiology of breast cancer subtypes in two prospective cohort studies of breast cancer survivors. Breast Cancer Res. (2009) 11:1–13. doi: 10.1186/BCR2261
31. John EM, Koo J, Phipps AI, Longacre TA, Kurian AW, Ingles SA, et al. Reproductive characteristics, menopausal status, race and ethnicity, and risk of breast cancer subtypes defined by ER, PR and HER2 status: the Breast Cancer Etiology in Minorities study. Breast Cancer Res. (2024) 26:1–29. doi: 10.1186/S13058-024-01834-5/TABLES/7
32. Lipsyc-Sharf M, Ballman KV, Campbell JD, Muss HB, Perez EA, Shulman LN, et al. Age, body mass index, tumor subtype, and racial and ethnic disparities in breast cancer survival. JAMA Netw Open. (2023) 6:E2339584. doi: 10.1001/JAMANETWORKOPEN.2023.39584
33. Hines LM, Risendal B, Byers T, Mengshol S, Lowery J, and Singh M. Ethnic disparities in breast tumor phenotypic subtypes in hispanic and non-hispanic white women. J Womens Health. (2011) 20:1543–50. doi: 10.1089/jwh.2010.2558
34. Nahleh Z, Otoukesh S, Mirshahidi HR, Nguyen AL, Nagaraj G, Botrus G, et al. Disparities in breast cancer: a multi-institutional comparative analysis focusing on American Hispanics. Cancer Med. (2018) 7:2710–7. doi: 10.1002/cam4.1509
35. Kyalwazi B, Yau C, Campbell MJ, Yoshimatsu TF, Chien AJ, Wallace AM, et al. Race, gene expression signatures, and clinical outcomes of patients with high-risk early breast cancer. JAMA Netw Open. (2023) 6:E2349646. doi: 10.1001/JAMANETWORKOPEN.2023.49646
36. Howlader N, Altekruse SF, Li CI, Chen VW, Clarke CA, Ries LAG, et al. US incidence of Breast cancer Subtypes Defined by Joint Hormone receptor and Her2 Status. J Natl Cancer Inst. (2014) 106(5):dju055. doi: 10.1093/jnci/dju055
37. Parise C and Caggiano V. Disparities in the risk of the ER/PR/HER2 breast cancer subtypes among Asian Americans in California. Cancer Epidemiol. (2014) 38:556–62. doi: 10.1016/j.canep.2014.08.001
38. Telli ML, Chang ET, Kurian AW, Keegan THM, Mcclure LA, Lichtensztajn D, et al. Asian ethnicity and breast cancer subtypes: a study from the California Cancer Registry. (2011) 127(2):471–8. doi: 10.1007/s10549-010-1173-8
39. Sasaki T, Liyanage A, Bansil S, Silva A, Pagano I, Hidalgo EY, et al. Differences in breast cancer subtypes among racial/ethnic groups. Cancers (Basel). (2024) 16(20):3462. doi: 10.3390/CANCERS16203462
40. Cronin KA, Ries LAG, and Edwards BK. The surveillance, epidemiology, and end results (SEER) program of the national cancer institute. Cancer. (2014) 120 Suppl 23:3755–7. doi: 10.1002/CNCR.29049
41. Feig B. Comprehensive databases: A cautionary note. Ann Surg Oncol. (2013) 20:1756–8. doi: 10.1245/S10434-013-2904-Y/METRICS
42. Goldhirsch A, Winer EP, Coates AS, Gelber RD, Piccart-Gebhart M, Thürlimann B, et al. Personalizing the treatment of women with early breast cancer: highlights of the St Gallen International Expert Consensus on the Primary Therapy of Early Breast Cancer 2013. Ann Oncol. (2013) 24:2206–23. doi: 10.1093/annonc/mdt303
43. Kim SW, Roh J, and Park CS. Immunohistochemistry for pathologists: Protocols, pitfalls, and tips. J Pathol Transl Med. (2016) 50:411–8. doi: 10.4132/jptm.2016.08.08
44. Lee KK, Rishishwar L, Ban D, Nagar SD, Mariño-Ramírez L, McDonald JF, et al. Association of genetic ancestry and molecular signatures with cancer survival disparities: A pan-cancer analysis. Cancer Res. (2022) 82:1222–33. doi: 10.1158/0008-5472.CAN-21-2105/675684/AM/ASSOCIATION-OF-GENETIC-ANCESTRY-AND-MOLECULAR
45. Loree JM, Anand S, Dasari A, Unger JM, Gothwal A, Ellis LM, et al. Disparity of race reporting and representation in clinical trials leading to cancer drug approvals from 2008 to 2018. JAMA Oncol. (2019) 5:e191870–e191870. doi: 10.1001/JAMAONCOL.2019.1870
46. Ford JG, Howerton MW, Lai GY, Gary TL, Bolen S, Gibbons MC, et al. Barriers to recruiting underrepresented populations to cancer clinical trials: a systematic review. Cancer. (2008) 112:228–42. doi: 10.1002/CNCR.23157
47. Jo Lund M, Butler EN, Hair BY, Ward KC, Andrews JH, Oprea-Ilies G, et al. Age/race differences in HER2 testing and in incidence rates for breast cancer triple subtypes A population-based study and first report. Cancer. (2010) 116:2549–59. doi: 10.1002/cncr.25016
48. Anderson KN, Schwab RB, and Martinez ME. Reproductive risk factors and breast cancer subtypes: a review of the literature. Breast Cancer Res Treat. (2014) 144:1–10. doi: 10.1007/s10549-014-2852-7
49. Lester SP, Kaur AS, and Vegunta S. Association between lifestyle changes, mammographic breast density, and breast cancer. Oncologist. (2022) 27:548–54. doi: 10.1093/ONCOLO/OYAC084
50. Rey-Vargas L, Sanabria-Salas MC, Fejerman L, and Serrano-Gómez SJ. Risk factors for triple-negative breast cancer among latina women. Cancer Epidemiol Biomarkers Prev. (2019) 28:1771–83. doi: 10.1158/1055-9965.epi-19-0035
51. McCarthy AM, Friebel-Klingner T, Ehsan S, He W, Welch M, Chen J, et al. Relationship of established risk factors with breast cancer subtypes. Cancer Med. (2021) 10:6456–67. doi: 10.1002/CAM4.4158
52. Yap YS. Outcomes in breast cancer—does ethnicity matter? ESMO Open. (2023) 8:101564. doi: 10.1016/J.ESMOOP.2023.101564
53. Kan Z, Ding Y, Kim J, Jung HH, Chung W, Lal S, et al. Multi-omics profiling of younger Asian breast cancers reveals distinctive molecular signatures. Nat Commun. (2018) 9(1):1725. doi: 10.1038/s41467-018-04129-4
54. Grunda JM, Steg AD, He Q, Steciuk MR, Byan-Parker S, Johnson MR, et al. Differential expression of breast cancer-associated genes between stage- and age-matched tumor specimens from African- and Caucasian-American Women diagnosed with breast cancer. BMC Res Notes. (2012) 5:248. doi: 10.1186/1756-0500-5-248
55. Jewer M, Findlay SD, and Postovit L-M. Post-transcriptional regulation in cancer progression: Microenvironmental control of alternative splicing and translation. J Cell Commun Signal. (2012) 6:233. doi: 10.1007/S12079-012-0179-X
56. Tan S, Ding K, Chong QY, Zhao J, Liu Y, Shao Y, et al. Post-transcriptional regulation of ERBB2 by miR26a/b and HuR confers resistance to tamoxifen in estrogen receptor-positive breast cancer cells. J Biol Chem. (2017) 292:13551–64. doi: 10.1074/JBC.M117.780973
57. Mersha TB and Abebe T. Self-reported race/ethnicity in the age of genomic research: its potential impact on understanding health disparities. Hum Genomics. (2012) 9(1):1. doi: 10.1186/s40246-014-0023-x
58. Miyashita M, Bell JSK, Wenric S, Karaesmen E, Rhead B, Kase M, et al. Molecular profiling of a real-world breast cancer cohort with genetically inferred ancestries reveals actionable tumor biology differences between European ancestry and African ancestry patient populations. Breast Cancer Res. (2023) 25(1):58. doi: 10.1186/S13058-023-01627-2
59. Rey-Vargas L, Bejarano-Rivera LM, Mejia-Henao JC, Sua LF, Bastidas-Andrade JF, Ossa CA, et al. Association of genetic ancestry with HER2, GRB7 AND estrogen receptor expression among Colombian women with breast cancer. Front Oncol. (2022) 12:989761/BIBTEX. doi: 10.3389/FONC.2022.989761/BIBTEX
60. Azin A, Tahmasebi H, Brar A, Azin S, Ko G, Covelli A, et al. Racial, ethnic and socioeconomic disparities in diagnosis, treatment, and survival of patients with breast cancer. Am J Surg. (2023) 225:154–61. doi: 10.1016/J.AMJSURG.2022.07.003
61. Loon Ooi S, Elena Martinez M, Li CI, and Author C. Disparities in breast cancer characteristics and outcomes by race/ethnicity. Breast Cancer Res Treat. (2011) 127:729–38. doi: 10.1007/s10549-010-1191-6
62. Talseth-Palmer BA and Scott RJ. Genetic variation and its role in Malignancy. Int J Biomed Sci. (2011) 7:158–71.
63. Krishna BM, Chaudhary S, Panda AK, Mishra DR, and Mishra SK. Her2 Ile655Val polymorphism and its association with breast cancer risk: An updated meta-analysis of case-control studies. Sci Rep. (2018) 8(1):7427. doi: 10.1038/s41598-018-25769-y
64. Ma Y, Yang J, Zhang P, Liu Z, Yang Z, and Qin H. Lack of association between HER2 codon 655 polymorphism and breast cancer susceptibility: Meta-analysis of 22 studies involving 19,341 subjects. Breast Cancer Res Treat. (2011) 125:237–41. doi: 10.1007/s10549-010-0965-1
65. Wang H, Liu L, Lang Z, Guo S, Gong H, Guan H, et al. Polymorphisms of ERBB2 and breast cancer risk: A meta-analysis of 26 studies involving 35,088 subjects. J Surg Oncol. (2013) 108:337–41. doi: 10.1002/jso.23386
66. Toomey S, Madden SF, Furney SJ, Fan Y, McCormack M, Stapleton C, et al. The impact of ERBB-family germline single nucleotide polymorphisms on survival response to adjuvant trastuzumab treatment in HER2-positive breast cancer. Oncotarget. (2016) 7:75518–25. doi: 10.18632/oncotarget.12782
67. Stanton SE, Ward MM, Christos P, Sanford R, Lam C, Cobham MV, et al. Pro1170 Ala polymorphism in HER2-neu is associated with risk of trastuzumab cardiotoxicity. BMC Cancer (2015) 15:267. doi: 10.1186/s12885-015-1298-6
68. Gaibar M, Beltrán L, Romero-Lorca A, Fernández-Santander A, Novillo A, and Selli C. Somatic mutations in HER2 and implications for current treatment paradigms in HER2-positive breast cancer. J Oncol. (2020) 2020:6375956. doi: 10.1155/2020/6375956
69. Si P, Xu Y, Ouyang T, Li J, Wang T, Fan Z, et al. HER2 Pro1170Ala polymorphism is associated with decreased survival rate in HER2-negative breast cancer. Oncol Lett. (2017) 13:3793–8. doi: 10.3892/ol.2017.5866
70. Lonsdale J, Thomas J, Salvatore M, Phillips R, Lo E, Shad S, et al. The genotype-tissue expression (GTEx) project. Nat Genet. (2013) 45:580–5. doi: 10.1038/ng.2653
71. Su Y, Jiang Y, Sun S, Yin H, Shan M, Tao W, et al. Effects of HER2 genetic polymorphisms on its protein expression in breast cancer. Cancer Epidemiol. (2015) 39:1123–7. doi: 10.1016/j.canep.2015.08.011
72. NCBI. National center for biotechnology information: reference SNP (refSNP) cluster report rs1058808 . Available online at: https://www.ncbi.nlm.nih.gov/snp/rs1058808frequency_tab (Accessed March 14, 2021).
73. Wang S, Qian F, Zheng Y, Ogundiran T, Zheng W, Blot W, et al. Genetic variants demonstrating flip-flop phenomenon and breast cancer risk prediction among women of African ancestry. Breast Cancer Res Treat. (2020) 168:834–43. doi: 10.1007/s10549-017-4638-1.Genetic
74. Han W, Kang D, Lee JE, Park IA, Choi J-Y, Lee K-M, et al. A haplotype analysis of HER-2 gene polymorphisms: association with breast cancer risk, HER-2 protein expression in the tumor, and disease recurrence in korea. Clin Cancer Res. (2005) 11:4775–8. doi: 10.1158/1078-0432.CCR-04-2208
75. NCBI. National center for biotechnology information: reference SNP (refSNP) cluster report rs1136201 . Available online at: https://www.ncbi.nlm.nih.gov/snp/rs1136201frequency_tab (Accessed March 14, 2021).
76. Chen W, Yang H, Tang WR, Feng SJ, and Wei YL. Updated meta-analysis on HER2 polymorphisms and risk of breast cancer: Evidence from 32 studies. Asian Pacific J Cancer Prev. (2014) 15:9643–7. doi: 10.7314/APJCP.2014.15.22.9643
77. Gamazon ER, Segrè AV, van de Bunt M, Wen X, Xi HS, Hormozdiari F, et al. Using an atlas of gene regulation across 44 human tissues to inform complex disease- and trait-associated variation. Nat Genet. (2018) 50:956–67. doi: 10.1038/s41588-018-0154-4
78. NCBI. National center for biotechnology information: reference SNP (refSNP) cluster report rs2517956 . Available online at: https://www.ncbi.nlm.nih.gov/snp/rs2517956variant_details (Accessed March 18, 2021).
79. Pivot X, Romieu G, Fumoleau P, Rios M, Bonnefoi H, Bachelot T, et al. Constitutional variants are not associated with HER2-positive breast cancer: results from the SIGNAL/PHARE clinical cohort. NPJ Breast Cancer. (2017) 3:1–5. doi: 10.1038/s41523-017-0005-y
80. Rivandi M, Martens JWM, and Hollestelle A. Elucidating the underlying functional mechanisms of breast cancer susceptibility through post-GWAS analyses. Front Genet. (2018) 9:280. doi: 10.3389/fgene.2018.00280
81. Zhang L, Yu Y, Shi T, Kou M, Sun J, Xu T, et al. Genome-wide analysis of expression quantitative trait loci (eQTLs) reveals the regulatory architecture of gene expression variation in the storage roots of sweet potato. Horticulture Res. (2020) 7:1–12. doi: 10.1038/s41438-020-0314-4
82. Zheng J, Deng J, Jiang L, Yang L, You Y, Hu M, et al. Heterozygous genetic variations of FOXP3 in xp11.23 elevate breast cancer risk in chinese population via skewed X-chromosome inactivation. Hum Mutat. (2013) 34:619–28. doi: 10.1002/humu.22284
83. Hian C, Chan T, Munusamy P, Loke SY, Koh GL, Sern E, et al. Identification of novel breast cancer risk loci. Cancer Res. (2017) 77(19):5428–37. doi: 10.1158/0008-5472.CAN-17-0992
84. Reba A, Kallel I, Rebai M, Khabir A, and Farid NR. Genetic polymorphisms in the egfr (R521K) and estrogen receptor (T594T) genes, EGFR and erbb-2 protein expression, and breast cancer risk in Tunisia. J BioMed Biotechnol. (2009) 2009:753683. doi: 10.1155/2009/753683
85. Dunning AM, Michailidou K, Kuchenbaecker KB, Thompson D, French JD, Beesley J, et al. Breast cancer risk variants at 6q25 display different phenotype associations and regulate ESR1, RMND1 and CCDC170. Nat Genet. (2016) 48:374–86. doi: 10.1038/ng.3521
86. Fasching PA, Loehberg CR, Strissel PL, Lux MP, Bani MR, Schrauder M, et al. Single nucleotide polymorphisms of the aromatase gene (CYP19A1), HER2/neu status, and prognosis in breast cancer patients. Breast Cancer Res Treat. (2008) 112:89–98. doi: 10.1007/s10549-007-9822-2
87. Wang Y, Yang SH, Hsu PW, Chien SY, Wang CQ, Su CM, et al. Impact of WNT1-inducible signaling pathway protein-1 (WISP-1) genetic polymorphisms and clinical aspects of breast cancer. Medicine. (2019) 98:e17854. doi: 10.1097/MD.0000000000017854
88. Vahednia E, Shandiz FH, Bagherabad MB, Moezzi A, Afzaljavan F, Tajbakhsh A, et al. The Impact of CASP8 rs10931936 and rs1045485 Polymorphisms as well as the Haplotypes on Breast Cancer Risk: A Case-Control Study. Clin Breast Cancer. (2019) 19:e563–77. doi: 10.1016/j.clbc.2019.02.011
89. Cerne J-Z, Stegel V, Gersak K, and Novakovic S. KRAS rs61764370 is associated with HER2-overexpressed and poorly-differentiated breast cancer in hormone replacement therapy users: a case control study. BMC Cancer. (2012) 12:105. doi: 10.1186/1471-2407-12-105
90. Zhang B, Li Y, Li L, Chen M, Zhang C, Zuo X-B, et al. Association study of susceptibility loci with specific breast cancer subtypes in Chinese women. Breast Cancer Res Treat. (2014) 146:503–14. doi: 10.1007/s10549-014-3041-4
91. Rahoui J, Sbitti Y, Touil N, Laraqui A, Ibrahimi A, Rhrab B, et al. The single nucleotide polymorphism +936 C/T VEGF is associated with human epidermal growth factor receptor 2 expression in Moroccan breast cancer women. Med Oncol. (2014) 31:1–5. doi: 10.1007/S12032-014-0336-6
92. Justenhoven C, Pierl CB, Haas S, Fischer H-P, Hamann U, Baisch C, et al. Polymorphic loci of E2F2, CCND1 and CCND3 are associated with HER2 status of breast tumors. Int J Cancer. (2009) 124:2077–81. doi: 10.1002/IJC.24198
Keywords: breast neoplasms, HER2 gene, American Native Continental Ancestry, single nucleoside polymorphism, ethnic group
Citation: Rey-Vargas L, Bejarano-Rivera LM, López-Correa P, Ballen-Lozano DF and Serrano-Gómez SJ (2025) Differences in the distribution of HER2-positive breast tumors according to ethnicity and genetic variants in ERBB2: a special focus on Asian and Latina women. Front. Oncol. 15:1635681. doi: 10.3389/fonc.2025.1635681
Received: 26 May 2025; Accepted: 10 July 2025;
Published: 24 July 2025.
Edited by:
Jorge Melendez-Zajgla, National Institute of Genomic Medicine (INMEGEN), MexicoReviewed by:
Bertha Rueda-Zarazua, National Autonomous University of Mexico, MexicoHilmaris Centeno-Girona, Comprehensive Cancer Center, University of Puerto Rico, Puerto Rico
Copyright © 2025 Rey-Vargas, Bejarano-Rivera, López-Correa, Ballen-Lozano and Serrano-Gómez. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Silvia J. Serrano-Gómez, c2lsdmlhanNlcnJhbm9AZ21haWwuY29t