 Samuel Trujano-Camacho1
Samuel Trujano-Camacho1 Carlos Contreras-Romero2
Carlos Contreras-Romero2 Verónica García-Castillo1
Verónica García-Castillo1 David Sánchez-Marín3
David Sánchez-Marín3 Mercedes Olvera-Valencia4
Mercedes Olvera-Valencia4 Mauricio Rodríguez-Dorantes5
Mauricio Rodríguez-Dorantes5 Oscar Peralta-Zaragoza6
Oscar Peralta-Zaragoza6 David Cantú de León7
David Cantú de León7 Eduardo López-Urrutia1
Eduardo López-Urrutia1 Carlos Pérez-Plasencia1*
Carlos Pérez-Plasencia1*- 1Laboratorio de Genómica, Unidad de Biomedicina, Facultad de Estudios Superiores-Iztacala (FES-IZTACALA), Universidad Nacional Autónoma de México (UNAM), Tlalnepantla, Mexico
- 2Laboratorio de Diagnóstico Molecular Genelab, Cuahutémoc, Mexico
- 3Posgrado en Ciencias Biológicas, Facultad de Medicina, Universidad Nacional Autónoma de México (UNAM), Coyoacan, Mexico
- 4Laboratorio de Virus y Cáncer, Dirección de Investigación, Instituto Nacional de Cancerología, México City, Mexico
- 5Laboratorio de Oncogenómica, Instituto Nacional de Medicina Genómica, Tlalpan, Mexico
- 6Dirección de Infecciones Crónicas y Cáncer, Centro de Investigación Sobre Enfermedades Infecciosas, Instituto Nacional de Salud Pública, Cuernavaca, Mexico
- 7Dirección de Investigación, Instituto Nacional de Cancerología, México City, Mexico
Cervical cancer (CC) remains among the top causes of death for women worldwide, especially in low-income countries, where screening strategies are less widespread. Treatment strategies are mainly based on DNA-damaging agents, though resistance mechanisms still pose a substantial challenge. Among the cellular components that mediate treatment resistance, long non-coding RNAs (lncRNAs) stand out because of their broad regulatory effects. They are involved in virtually all drug resistance mechanisms, such as drug efflux, DNA repair, evasion of cell death, and aberrant epigenetic modifications. Although resistance mechanisms are fundamentally similar in most cancers, the underlying regulatory networks vary substantially. Here, we review the literature for lncRNAs involved in treatment resistance mechanisms in general, and then focus on lncRNAs that mediate resistance in CC. We found a broad area of opportunity in lncRNA research in resistant CC, as the lncRNAs involved are still to be described. These master regulators are promising candidates for response markers and therapeutic targets. May this compilation serve as the basis for further descriptions of the regulatory roles of lncRNA in CC treatment resistance.
1 Introduction
Cervical cancer (CC) is the fourth most common cancer among women worldwide and the third leading cause of cancer-related mortality. In 2022 alone, 662,044 cases and 348,709 deaths from cervical cancer occurred, with mortality exceeding 50% in many cases. While most cases are detected in high-income countries, the number of deaths is disproportionately high in low-income countries, where CC ranks as the second most deadly cancer for women (1).
Human Papillomavirus (HPV) infection is accepted as the main etiology of CC. HPV virions can reach any of the different epithelial regions that comprise the uterine cervix: the endocervix, lined by columnar glandular epithelium, the ectocervix, lined by stratified squamous epithelium, or the transformation zone formed by metaplastic epithelium (2), giving rise to two histological variants: squamous cell carcinoma (85%) and adenocarcinoma (15%) (3).
Current pharmacological treatment strategies for CC include platinum-based chemoradiation therapy (CCRT) and brachytherapy. For advanced or recurrent cases, a platinum-based chemotherapy doublet, often combining cisplatin with paclitaxel, ifosfamide, gemcitabine, topotecan, or vinorelbine, to mention a few, is the first-line treatment. However, the response rate for early-to-intermediate stages (IIA-IIIB) is only 68%, and 5-year overall survival (OS) and progression-free survival (PFS) rates remain low, at 68% and 66%, respectively (4). Several groups are currently working to optimize these effects through novel drug delivery strategies of already approved drugs. These strategies include delivery of the recently approved pembrolizumab through nanoparticles of nanopolymers (5). Nonetheless, resistance to therapy –whether intrinsic or acquired– remains a critical challenge, undermining treatment efficacy and contributing to progressive, advanced, or metastatic disease.
Therapy resistance in CC, as in other cancers, arises through molecular mechanisms involving genetic and epigenetic alterations. Cancer stem cells (CSCs) play a prominent role, alongside mechanisms such as enhanced drug efflux, increased DNA repair, evasion of apoptosis, autophagy, epithelial-mesenchymal transition (EMT), and aberrant epigenetic modifications (6).
Recent research highlights the involvement of long noncoding RNAs (lncRNAs) in these processes, positioning them as pivotal players in therapy resistance. LncRNAs are RNA molecules longer than 200 bp that, while not typically translated into proteins, fold into complex secondary structures, enabling interactions with DNA, RNA, or proteins. These interactions allow lncRNAs to act as molecular decoys, signaling mediators, guides, and scaffolds for protein complexes (7). Deregulated lncRNAs have been shown to influence every hallmark of cancer, promoting malignant transformation and tumor progression. Multi-omics studies have further demonstrated that lncRNAs contribute directly to a cancerous phenotype by interacting with various macromolecules and molecular complexes (8).
This review focuses on the role of lncRNAs in mediating drug resistance in CC. First, we explore their involvement in known resistance mechanisms such as drug efflux, DNA repair, and epigenetic modifications. Then, we analyze reported associations between specific treatments and lncRNAs, emphasizing their potential as biomarkers for predicting therapeutic response and guiding treatment strategies. By understanding their molecular and clinical significance, we aim to underscore the promise of lncRNAs as targets to overcome resistance and improve outcomes for CC patients. Many of these lncRNAs affecting resistance mechanisms have yet to be characterized in the context of cervical cancer, highlighting the need for further research and raising important new questions for future investigation in this field (Figure 1).
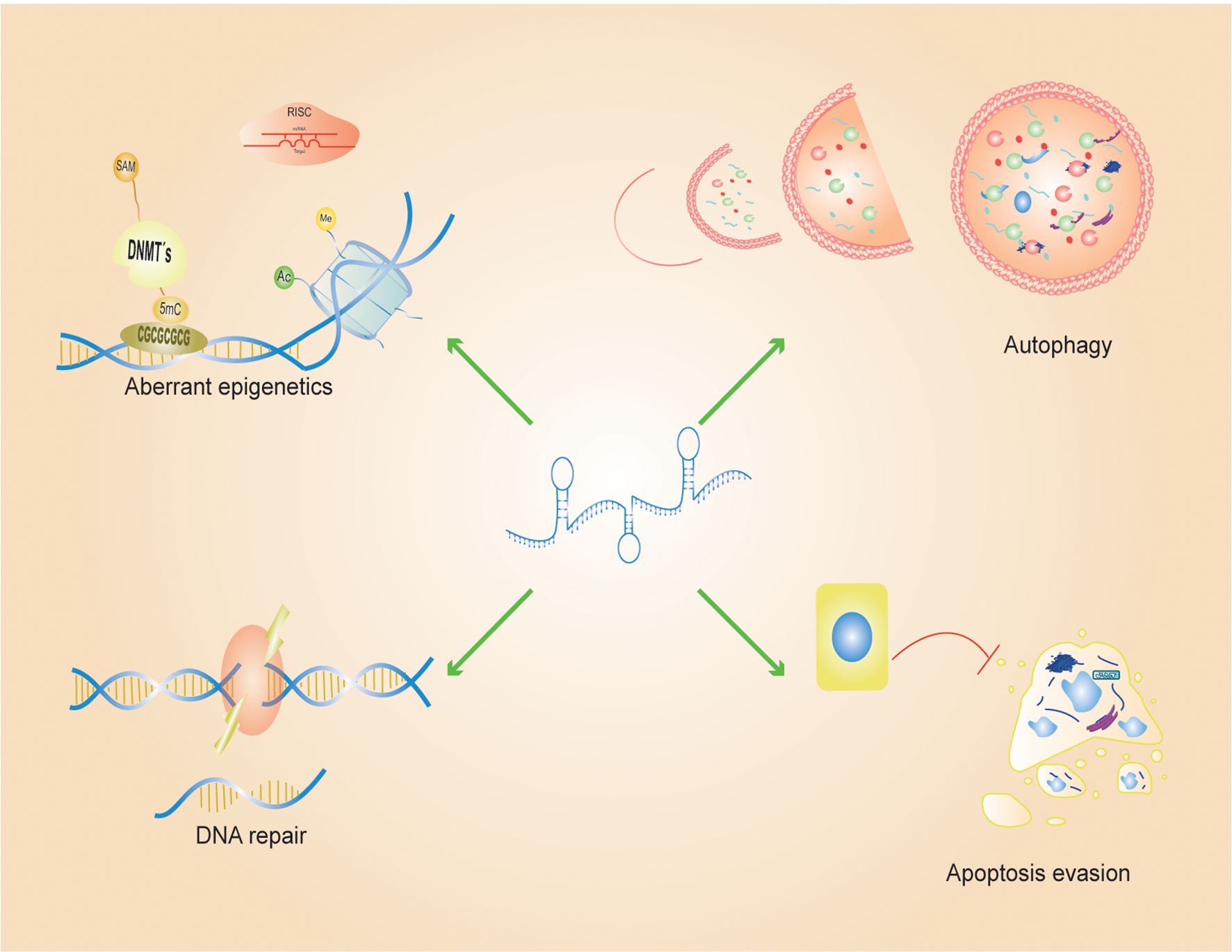
Figure 1. LncRNA-mediated resistance processes: Aberrant epigenetic processes, autophagy, DNA repair, evasion of apoptosis and epithelial-mesenchymal transition are some of the main processes mediated by LncRNAs in cancer. To promote a chemoresistant phenotype, long non-coding RNAs (LncRNAs) modulate gene expression through diverse epigenetic mechanisms, including DNA methylation, histone modifications, and functioning as competing endogenous RNAs (ceRNAs) that sequester microRNAs (miRNAs). Beyond epigenetic regulation, LncRNAs bolster cellular resistance to therapy-induced cytotoxicity by activating autophagy which enhance tumor cell survival and metastatic potential. Furthermore, LncRNAs play a pivotal role in preserving genomic stability by facilitating the repair of single- and double-strand DNA breaks and by interfering with apoptotic signaling pathways, thereby enabling cancer cells to evade programmed cell death and sustain malignant progression.
2 lncRNAs involved in resistance mechanisms
The sensitivity of each tumor to different chemotherapy drugs and the determinants of their specificity depend on various factors, including the tumor microenvironment and dynamic changes that activate specific mechanisms, such as those mentioned above (9). In this regard, several studies have described the involvement of lncRNAs in resistance mechanisms. Although those reviewed below were not necessarily performed in cervical cancer (CC), they demonstrate that long noncoding RNAs are implicated in drug resistance and sensitivity in cancer cells (10). Understanding how lncRNAs modulate drug response is the basis for new avenues to address resistance to anticancer therapies.
2.1 Reduced absorption and increased drug efflux
One of the main resistance mechanisms is reduced drug uptake and increased drug efflux, which are primarily mediated through an increase in the ATP-binding cassette (ABC) superfamily transporters (11). There are 48 known transporters, classified into seven subfamilies (ABC A to G); 30 of them contribute to the development of drug resistance in tumors, most notably P-glycoprotein (P-gp) and breast cancer resistance protein (BCRP/ABCG2) (12). Several lncRNAs play crucial roles in regulating these drug transporters.
While much of the evidence comes from studies in other cancer types, these findings provide valuable insights for cervical cancer (CC). For example, the lncRNA HOTAIR has been studied in gastric cancer, where its overexpression regulates the activity of P-glycoprotein (P-gp), promoting the efflux of drugs such as doxorubicin and paclitaxel (13). MALAT1 plays a significant role in lung and gastric cancer, increasing resistance to cisplatin. Its ability to modulate ABC transporters and signaling pathways suggests a direct impact on treatment efficacy (14). Considering that both lncRNAs are overexpressed and associated with a poor prognosis in CC (15, 16) and that they promote resistance to drugs used in CC treatment regimens, we can begin to explore the implications of lncRNAs in specific tumors and as resistance mechanisms shared among tumors dependent on the presence of specific regulators, such as MALAT1 and HOTAIR. Similarly, reviewing other lncRNAs involved in similar resistance mechanisms can potentially lead us to new, unexplored CC targets. The lncRNA MRUL (multidrug resistance-related and upregulated lncRNA), for example, regulates ABCB1 expression in gastric cancer, thereby promoting multidrug resistance (17). These findings highlight the direct role of specific lncRNAs in modulating ATP-binding cassette (ABC) transporters and driving the multidrug-resistant phenotype in various cancer types.
2.2 Increased DNA repair and apoptosis evasion
Various genotoxic treatments damage DNA by breaking covalent bonds between nucleotides, disrupting genome replication and transcription. The repair machinery can intervene by correcting the damage at specific checkpoints through a signaling cascade or by triggering cell death (18). The DNA damage response induces apoptosis through the so-called intrinsic pathway, characterized by membrane permeabilization and release of pro-apoptotic factors such as cytochrome C or apoptotic protease activation factor 1 (Apaf-1) into the cytosol. Resistance mechanisms usually involve deregulation of proteins involved in the intrinsic apoptosis pathway, including members of the Bcl-2 family and the p53 tumor suppressor, overexpression of inhibitors of apoptosis proteins (IAPs), or overactivation of signaling pathways such as PI3K/AKT, even in the presence of DNA damage (19).
LncRNAs play a key role in DNA damage repair and apoptosis avoidance through regulating the ATM, ATR, and p53 pathways (20). Homologous recombination (HR) stands out among the DNA repair mechanisms and is the most studied in cancer. It is regulated by ATM, ATR, BRCA1-2, and RAF5, to name a few (21). The lncRNAs NEAT1, ANRIL, and ScaRNA2 perform regulatory roles in HR by stabilizing ATM and ATR, thus increasing the rate of DNA repair and conferring resistance to cancer cells (22, 23).
In this context, NEAT1 and ANRIL are markedly overexpressed in cervical cancer tissues and cell lines, correlating with more severe clinical features and reduced patient survival (24–26). Suppressing NEAT1 expression hampers cervical cancer glycolytic activity, which has been widely associated with high PD-L1 expression, as well as with the inhibition of apoptosis, lactate production, and immunosuppressive processes (27), whereas its upregulation produces the opposite effect. NEAT1 contributes to cervical cancer progression through multiple pathways, including the NEAT1/miR-133a/SOX4 axis, and the WNT/β-catenin/PDK1 signaling cascade axis (28, 29) both pathways are widely associated with the evasion of apoptosis, which is achieved through the transcriptional activation of genes such as survivin (30, 31). Meanwhile, ANRIL inhibition reduces the apoptosis evasion capabilities of CC cells (32).
Similarly, lncRNAs such as APAF1-binding lncRNA (ABL) or LINC00942 have been implicated in drug resistance processes by promoting the escape from apoptosis. ABL competes directly with cytochrome C to interact with apoptotic protease-activating factor 1 (APAF1), thereby preventing apoptosome formation (33). LINC00942, in turn, regulates c-Myc mRNA stabilization through the LINC00942/MSI2/c-Myc axis, preventing apoptosis as well (34).
2.3 Autophagy
Autophagy is a normal cell death process; however, cancer cells resort to autophagy to draw energy from macromolecules, thereby favoring growth and proliferation. Cervical cancer cell lines have shown resistance to drugs such as paclitaxel and cisplatin when autophagy is active, while autophagy-inhibiting drugs such as 3-methyladenine or chloroquine can restore drug cytotoxicity (35). Paclitaxel resistance is achieved through autophagy by activating HIF1α and the Warburg effect, while HIF2a promotes cisplatin drug resistance (36, 37). In addition, autophagy related to drug resistance is induced by elements and pathways dysregulated in cervical cancer, including PI3K/Akt/mTOR pathway, Beclin1, Bcl-2, Ras, p53, DUSP1, heparinase, HMGB1, and noncoding RNAs (38).
Along these lines, lncRNAs, such as LincRNA-p21 and HOTAIR, have positive feedback mechanisms that activate HIF1a transcription in CC (39, 40). This suggests that they could play an important role in resistance processes mediated by autophagy, though this has not yet been fully described.
Similarly, other lncRNAs, such as PVT1 in pancreatic cancer and EIF3J-DT and CRNDE in gastric cancer, have been associated with autophagy processes and resistance to gemcitabine and 5-fluorouracil (5-FU), respectively. Their high expression in CC could lead us to resistance mechanisms that have been little explored in this type of cancer (41–43).
2.4 Aberrant epigenetics
Given that chromatin structures strongly influence transcriptional activity, epigenetic events play a fundamental role, not only in tumor development and progression, but also in the generation of resistance to antitumor drugs (44). The cancer epigenome is characterized by changes in DNA methylation and histone modification, which allow tumoral progression (45). Several genes are hypomethylated and hypermethylated in cervical cancer, such as RASSF1A, TFPI2, MEG3, KLF4, IFN-γ, RAD51L3, and XRCC2 (46). Changes in DNA methylation lead to increased drug resistance through the upregulation of the PI3K/AKT, Wnt/β-catenin, Notch, and NF-κB signaling pathways. For instance, in cervical cancer, methylation of the Casp8A2 gene is sufficient to increase drug resistance; on the contrary, hypermethylation of WRN confers cervical cancer cells high sensitivity to the topoisomerase I inhibitor (CPT-11) (47).
The balance between histone deacetylases (HDACs) and histone acetyltransferases (HATs) also regulates gene transcription. For instance, in cervical cancer, MOF, a HAT enzyme, inhibits replication stress induced by cisplatin (48), while HDAC6 is involved in cell proliferation and motility (49).
HOTAIR is one such lncRNA; it may function as a protein scaffold for the Polycomb Repressive Complex 2 (PRC2), which modifies histone residues and binds DNMT3b to methylate promoter regions (50). On the other hand, lncRNA H19 modulates the activity of ALDH and activates the WNT/β-catenin pathway by interacting with miR-141, causing chemoresistance in colorectal tumors (51). H19 also interacts with let-7, upregulating LAT2 and OLR1 and leading to increased GSTM3TV2 levels in gemcitabine-resistant pancreatic tumor tissues (52). In ovarian cancer, NEAT1 knockdown releases miR-770-5p from sponging, which decreases PARP expression. Elevated PARP levels synergize with platinum compounds, improving the efficiency of chemotherapeutic approaches (53).
3 Long noncoding RNAs associated with therapy response
Only a handful of studies have associated lncRNAs with drug resistance in cervical cancer (CC), making it challenging to elucidate the mechanisms underlying drug resistance (54). Considering the therapeutic schemes (55) and the reported roles of lncRNAs in response to them, it is important to note that the expression profiles of lncRNAs promoting chemoresistance is similar between cervical cancer (CC) and other gynecological cancers. This allows us to gain insight on the possible role of lncRNAs that have not yet been described in CC, as the involved mechanisms are likely similar. This in-depth analysis could lead to new proposals for establishing markers based on the relationship between lncRNA expression levels and their involvement in drug-dependent resistance. (Table 1).
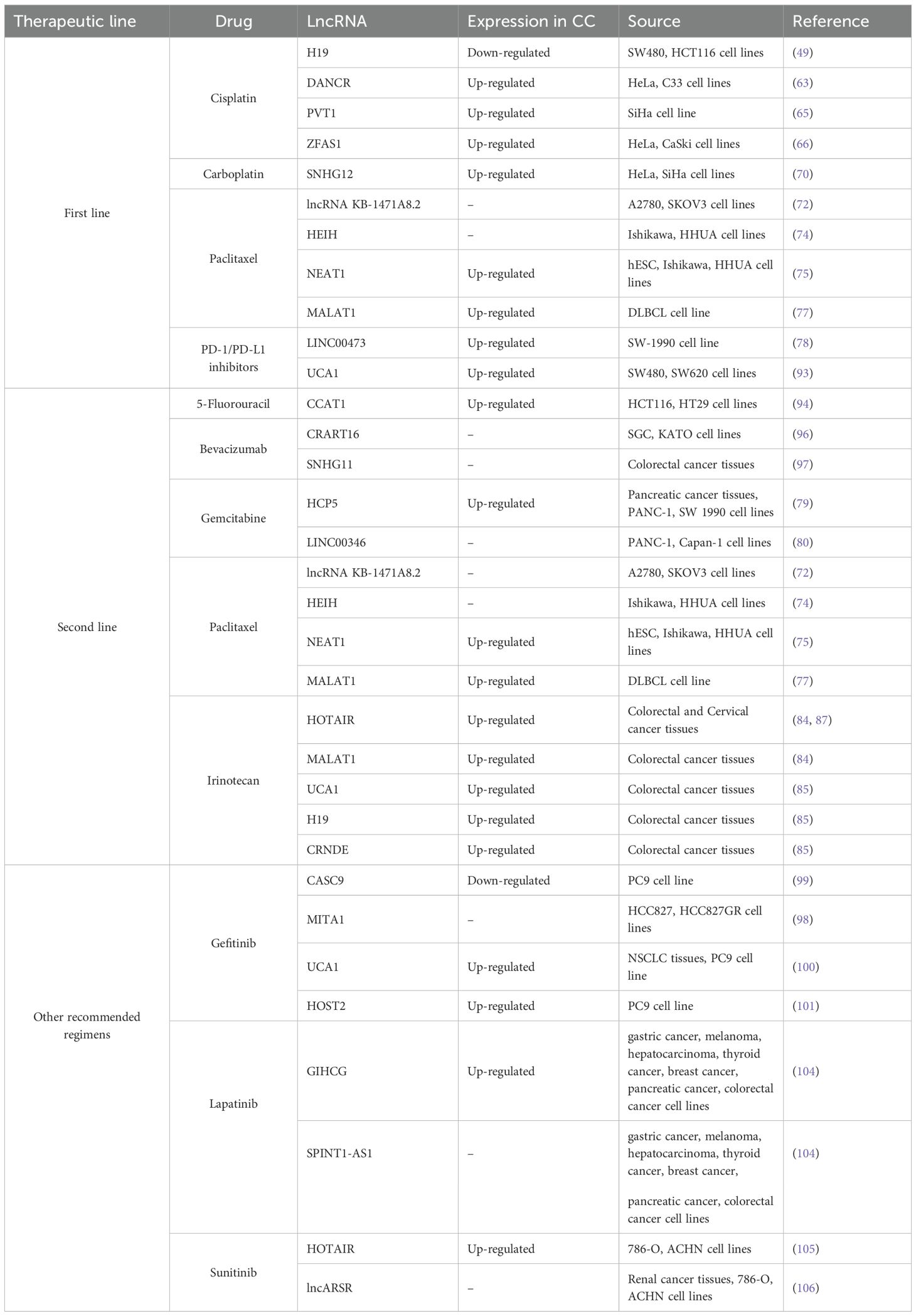
Table 1. LncRNAs are involved chemoresistance in CC.
3.1 First-line therapy
The first treatments for CC patients are radiosensitizers, mainly platinum compounds such as Cisplatin and Carboplatin; these are often used in combination with microtubule polymerization inhibitors like Paclitaxel or topoisomerase-1 inhibitors such as Topotecan (55). Though in many cases, patient therapies must be reformulated due to the development of mechanisms that promote or enable resistance to drugs and first-line therapies (56) (Table 1).
The main mechanisms by which tumor cells become resistant to first-line platinum drugs are drug efflux, DNA repair, and evasion of cell death (57). An enhanced DNA damage response bypasses the effect of cisplatin therapy in several ways. For instance, the heterodimer ERCC1-ERCC4 removes DNA bulky lesions through Nucleotide Excision Repair (NER) (58). Thus, ERCC1 is negatively correlated with responsiveness to cisplatin, which makes it a prognostic factor (59). The DNA mismatch repair pathway is also implicated, through the overexpression of MSH2 and downregulation of PMS2 (60, 61).
Furthermore, cisplatin induces apoptosis to achieve its antitumoral effect; however, in CC cells, the mechanisms leading to apoptosis are dysregulated, and cancer cells escape death. This dysregulation includes overexpression of antiapoptotic proteins such as BCL-2, Bcl-XL, and Bag-1; and downregulation of caspase activators, which decreases caspases in patients with drug resistance (62). Cisplatin-induced apoptosis requires an active MAPK, while deactivation of other pathways due to mutation or loss of P53 and NK-κβ reduces chemotherapy efficacy (60, 63, 64).
3.1.1 Cisplatin
It has been proven that some lncRNAs are directly implicated in cisplatin resistance in CC. For example, DANCR binds to miR-665, increasing the nuclear distribution of SMAD by overexpression of TGFBR1 (65). GAS5 sponges miR-21, enhancing the expression of phosphorylated STAT3 and E2F3; this decreases TIMP3 and PDCD4, leading to a tumoral cisplatin-resistant phenotype (66). PVT1 overexpression is associated with poor prognosis. It binds to and stabilizes MYC and Nuclein, contributing to cervical cancer development and its inhibition has also been strongly associated with sensitivity to cisplatin treatment (67). ZFAS1 has been reported to be upregulated in CC tissues. Its increased expression indicates poor prognosis, and its silencing enhances cisplatin sensitivity; however, the mechanism by which this effect occurs is unknown (68).
Several studies point to UCA1 promoting cisplatin resistance in cervical cancer-derived cells through regulation of caspase-3, p21, CDK and survivin expression (69). Recently another mechanism was demonstrated in which UCA1 acts as a sponge for miR-195-5p, targeting IKBKB promoting cisplatin resistance (70).
3.1.2 Carboplatin
At the time of writing, direct involvement of lncRNAs in carboplatin resistance in CC has not been reported; however, several lncRNAs have been linked to carboplatin resistance in other gynecologic cancers. For example, deregulation of the IncRNA PVT1 has been deemed necessary for resistance in ovarian cancer (71). LncRNAs such as H19 and SNHG12, which are highly relevant in CC, have been associated with carboplatin resistance in ovarian cancer through the regulation of epigenetic mechanisms (72, 73).
3.1.3 PD-1/PD-L1 inhibitors
Drugs targeted at programmed cell death protein-1 (PD-1) and programmed cell death ligand (PD-L1) are important immune checkpoint inhibitors (ICIs). PD-L1 is overexpressed in tumor cells or untransformed cells within the tumor microenvironment, inhibiting cytotoxic T cells by binding to the PD-1 receptor on activated T cells. PD-1 and PD-L1 inhibitors block this interaction, preventing cancer cells from evading the immune system and acting as ICIs by reactivating the T-cell-mediated tumor cell death (74). In CC, the specific therapy for PD-L1-positive is relatively new (55), so the resistance mechanism to this therapy is still to be studied. Some groups have reported a positive correlation between lncRNAs and PD-L1 expression. For example, MALAT1 is a ceRNA for miR-195 in lymphoma (75) and, in pancreatic cancer, LINC00473 sponges miR-195-5p (76); both increasing PD-L1 expression and both lncRNAs are overexpressed and are strongly associated with tumor development and poor prognosis in cervical cancer (The long noncoding RNA LINC00473, a target of microRNA 34a, promotes tumorigenesis by inhibiting ILF2 degradation in cervical cancer LncRNA MALAT1 Accelerates Cervical Carcinoma Proliferation by Suppressing miR-124 Expression in Cervical Tumor Cells). However, at the time of writing, no study correlates or characterizes any lncRNA and the development of resistance to PD-L1.
3.2 Second-line therapy
The second line of treatment comprises drugs such as Paclitaxel, Gemcitabine, Fluorouracil, and Irinotecan. These are used in patients whose progression continues even after first-line treatment and after 3–6 months of progression-free survival (55).
3.2.1 Paclitaxel
LncRNAs are also associated with resistance to drugs used in combination with cisplatin in gynecologic cancers. The expression of the lncRNA KB-1471A8.2 can inhibit resistance to Paclitaxel in ovarian cancer, decreasing migration and invasion while increasing apoptosis. Overexpressing KB-1471A8.2 suppresses CDK4 expression, decreasing the proportion of cells in S phase and increasing the proportion in G0/G1 phase (77). On the other hand, lncRNA HEIH binds the EZH2 protein and participates in chromatin remodeling in hepatocellular carcinoma (78). It also enhances Paclitaxel tolerance in endometrial cancer through the activation of the Mitogen-activated protein kinase (MAPK) pathway proteins such as p38, C-Jun, and C-fos; although the consequential resistance mechanism is still unclear. (79). So, HEIH and KB-1471A8.2 are apparently antagonistic, but both lead to Paclitaxel resistance.
NEAT1 is overexpressed in CC, where it plays a key role in carcinogenesis and tumoral progression; this lncRNA also regulates cellular processes such as proliferation, migration, and invasion (25). NEAT1 has also been related to paclitaxel resistance in ovarian cancer, where it sponges miR-194, leading to ZEB1 overexpression. This protein is an Epithelial-Mesenchyme Transition (EMT)-inducing transcription factor. Thus, NEAT1 overexpression leads to enhanced tumor growth and low apoptosis levels in Paclitaxel-resistant ovarian cancer cells (80).
3.2.2 Gemcitabine
LncRNA-mediated gemcitabine resistance mechanisms have been mostly described in pancreatic cancer through miRNA regulation. At least two lncRNAs promote proliferation, migration, invasion, and apoptosis evasion: HCP5 through the miR-214-3p/HDGF axis and it is also overexpressed in cervical cancer cell lines (81), and LINC00346 through the miR-188-3p/BRD4 axis It could represent an unexplored target in CC (82).
Elevated expression of AFAP1-AS1 in cervical cancer is associated with poor patient prognosis. Moreover, individuals in the gemcitabine-resistant group exhibited significantly higher levels of AFAP1-AS1 compared to those in the gemcitabine-sensitive group. Functionally, AFAP1-AS1 enhances tumor growth and contributes to gemcitabine resistance in cervical cancer cells. Mechanistically, AFAP1-AS1 regulates the expression of epidermal growth factor receptor (EGFR) by acting as a molecular sponge for miR-7-5p (83).
Notably, DLG1-AS1 expression was markedly elevated in gemcitabine-resistant HeLa/GEM and SiHa/GEM cervical cancer cells. Silencing DLG1-AS1 significantly reduced the viability and proliferation of these resistant cells. In vivo, tumors in nude mice with DLG1-AS1 knockdown showed reduced volume following gemcitabine treatment. DLG1-AS1 targets miR-16-5p, which regulates HDGF expression. Inhibition of miR-16-5p reversed the effects of DLG1-AS1 knockdown in gemcitabine-resistant cervical cancer cells (84). In fact, DLG1-AS1 has been identified as the most significantly upregulated lncRNA in cervical cancer tissues, and its elevated expression was associated with poor patient prognosis. Silencing DLG1-AS1 inhibited the proliferation of cervical cancer cells. Further analysis demonstrated that DLG1-AS1 acts as a competing endogenous RNA (ceRNA), binding to miR-107 and thereby preventing it from repressing its target gene, ZHX1 (85).
3.2.3 Irinotecan
Little has been described regarding the involvement of lncRNAs in the processes leading to Irinotecan resistance. However, key lncRNAs have been detected, including CRNDE, H19, UCA1, HOTAIR, and MALAT1, which also play a key role in the development of CC. In addition, the resistance mediated by MALAT1 and HOTAIR (As mentioned above, both are important in CC) has been extensively linked to polymorphisms detected in both, enabling their use as potential biomarkers of irinotecan sensitivity in colorectal cancer (86, 87).
Clinically, elevated HOTAIR expression in cervical cancer tissues correlates with advanced tumor stage, lymph node metastasis, and decreased overall survival (88). Its potential as a prognostic biomarker is increasingly recognized, as it reflects tumor biological aggressiveness and clinical outcome (15). HOTAIR plays a crucial role in therapeutic resistance, since its overexpression has been linked to reduced sensitivity to both chemotherapy and radiotherapy (89). HOTAIR enhances DNA damage repair, inhibits apoptosis, and promotes survival signaling pathways, thereby diminishing the efficacy of cytotoxic treatments. Targeting HOTAIR or its downstream pathways may offer a promising strategy to overcome resistance and improve therapeutic responses in cervical cancer patients (reviewed in 90).
3.2.4 5-Fluorouracile
We have already mentioned before the role of NEAT1 in Paclitaxel resistance, however, has also been reported as a precursor to generate cells resistant to other chemotherapy drugs. The study performed by Shao in 2021 showed the direct correlation between NEAT1 expression and 5-Fu) resistance in CC. the knocking down of NEAT1 by shRNA promoted the sensitivity of 5-Fu in CaSki cells due to the overexpression of miR-34a, this miRNA regulates the glycolysis through targeting LDHA, NEAT1 once again work as ceRNA to miR-34a inhibiting the interaction with its target mRNA and leading to 5-FU resistance in CC (91).
Substantially overexpressed in CC tissues, the lncRNA UCA1 can regulate proliferation, migration, and invasion by sponging miR-145 (92). Conversely, mir-23b-3p functions as a tumor suppressor, reducing the proliferation, migration, and invasion by targeting c-Met (93). The interaction between these ncRNAs has been characterized, albeit only in colorectal cancer. UCA1 sponges miR-23b-3p, leading to overexpression of the EMT-promoting ZNF281, which results in 5-Fu resistance (94). Another promising lncRNA is CCAT1, which promotes proliferation and invasion in cervical cancer but enhances 5-Fu sensitivity in colorectal cancer (95). This represents another instance of a resistance mechanism involving lncRNAs that are overexpressed in CC, which remain insufficiently characterized.
3.2.5 Other second-line regimes
In addition to the drugs mentioned above, recent studies based on data from The Cancer Genome Atlas have revealed new drug targets. These molecules regulate signaling pathways that drive tumor progression. As a result, several drugs in clinical stages I-II have been identified as therapy candidates for CC, including therapies against VEGF (Bevacizumab), EGFR (Gefitinib), HER2 (Lapatinib), and tyrosine kinase receptors (Sunitinib) (96).
Despite the selectivity of targeted therapies, several lncRNAs have been reported to orchestrate resistance mechanisms to such drugs in various types of cancer. For example, the lncRNAs SNHG11 and CRART16 mediate resistance to bevacizumab through regulation of the miR-1207-5p/ABCC1 axis in colorectal cancer and miR-122-5p/FOS in gastric cancer, respectively (97, 98).
Gefitinib resistance has been associated with the expression of several lncRNAs, mainly in small cell lung cancer (NSCLC). The lncRNA MITA1, for example, favors an increase in cell viability and induction of autophagy in the presence of Gefitinib (99). Similarly, epigenetic regulations of lncRNAs such as UCA1, which is also overexpressed in CC, have also been associated with Gefitinib resistance through interaction with EZH2 (100–102). The lncRNA HOST2 favors Gefitinib resistance by sponging miR-621 (103), while CCAT1 allows resistance to this drug by repressing miR-218 in NSCLC (104), and the overexpression of these two lncRNAs promotes cell proliferation and migration (105, 106).
The extensive participation of lncRNAs in various biological processes has also allowed the recognition of lncRNAs as biomarkers of drug sensitivity or as indicators of resistance to therapies such as lapatinib. LncRNAs such as SPINT1-AS1 in the NCI-N87 and MCF7 cell lines and GAS5 in the SKBR-3/Tr cell line have been described as lapatinib sensitivity indicators (107). Overexpression of the lncRNA GIHCG has been also reported as a biomarker of lapatinib resistance (108).
LncRNAs also mediate resistance to sunitinib. HOTAIR regulates miR-17-5p, favoring autophagy in renal cancer (109); and the lncARSR/AXL/c-MET and lncRNECVSR/ERβ/Hif2 axes regulate cell proliferation, leading to sunitinib resistance in renal cell carcinoma (110, 111).
3.3 Signaling pathway inhibitors
On the other hand, there are reports of lncRNAs regulating signaling pathways such as ERK/MAPK, Wnt/β-catenin, and the PI3k/AKT as, have been related to lncRNAs, some of the most relevant that favor tumor development and drug resistance in CC (112). This has led to the development of drugs specifically targeting effectors of these signaling pathways. Within the ERK/MAPK pathway, relevance has been given to ERK inhibitors in BRAF or RAS mutations, highlighting ulixertinib, which is in the early clinical phases (113). In the PI3K/AKT pathway, several inhibitors targeting PI3K and mTOR proteins have been developed whose relevance has reached even clinical stages, including buparlisib (phase II clinical trial) and gedatosilb (phase Ib clinical trial) (114). Finally, for the Wnt/β-catenin pathway, the focus has been on the development of inhibitors targeting β-catenin, inhibitors of tankinase, and inhibitors of β-catenin/TCF interaction (CRT3/iCRT5/iCRT14/BC21/BC23/HI-B1), to mention a few (115).
Although the development of the signaling pathway inhibitors has advanced substantially, very little has been described about the resistance mechanisms themselves. Recent studies have allowed us to glimpse into new mechanisms by which lncRNAs favor resistance to specific inhibitors. HOTAIR, for example, has been highlighted as an important regulator of the Wnt/β-catenin and PI3K/AKT pathways even in the presence of the inhibitor ICRT14; knockdown of HOTAIR optimized inhibition and led to cell death in the HeLa cell line (40, 116).
4 Concluding remarks
After reviewing the available literature on lncRNAs and drug resistance in CC, it is evident that only a few lncRNAs are directly associated with chemoresistance to some therapies. The most widely described mechanism is the competitive binding of lncRNAs to miRNAs, which prevents the downregulation of their mRNA targets. This ceRNA effect results in the alteration of cellular processes, such as EMT, migration, invasion, and proliferation, ultimately leading to chemoresistance to standard treatments. However, when compared to other gynecological cancers, such as ovarian and endometrial, in which the same drugs are employed, cervical cancer remains poorly explored (Figure 2). Interestingly, the expression profiles of lncRNAs that promote chemoresistance are similar in CC and other gynecological cancers, providing insight into the mechanisms that may underlie the development of chemoresistance to therapies used in CC. The study of lncRNAs and the drug resistance mechanisms they participate in is a promising tool for developing better therapeutic schemes that will undoubtedly expand in the following years, resulting in more favorable clinical outcomes for patients with CC.
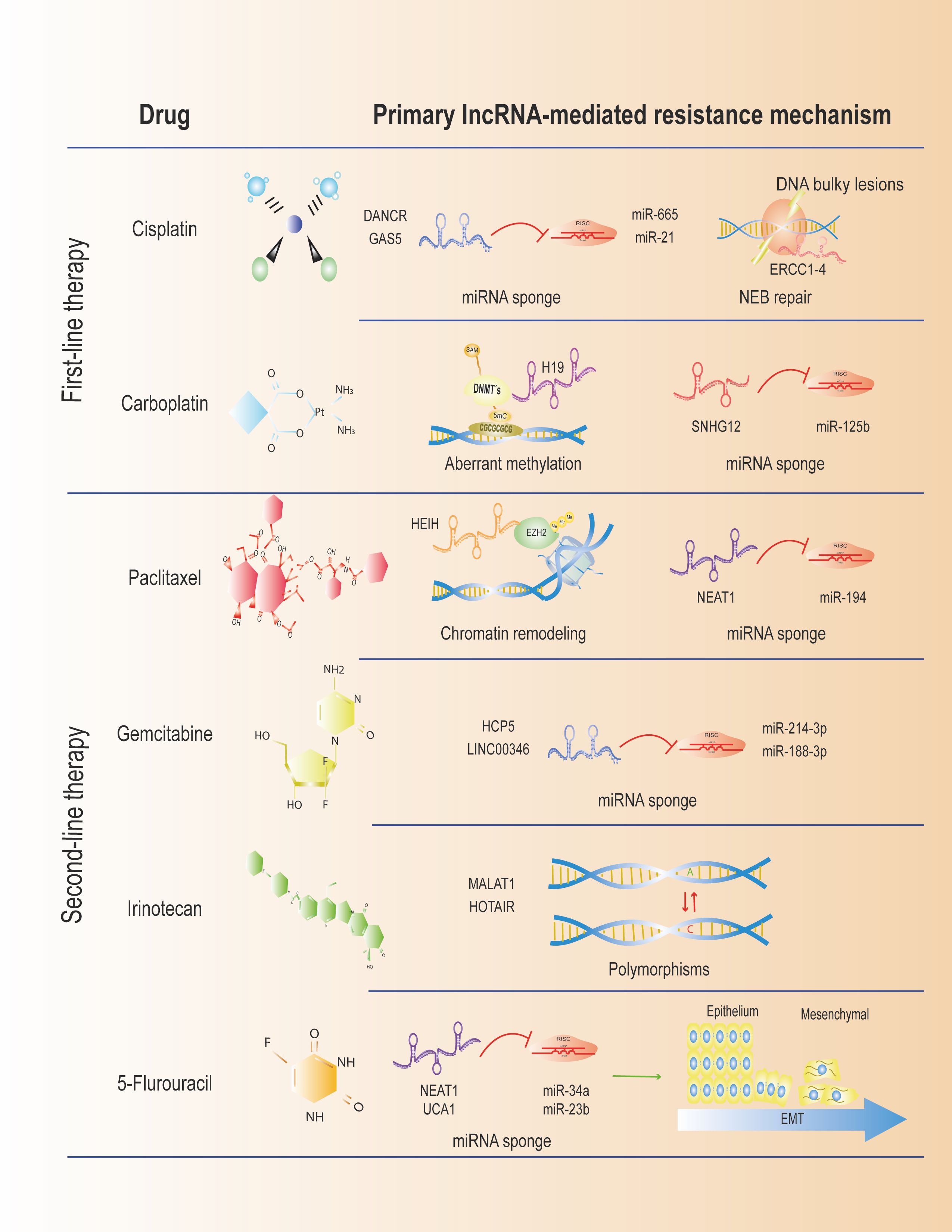
Figure 2. Treatment regimens for CC associated with LncRNAs. We show some described lncRNAs mediating resistance in the first- and second-line therapies.
Author contributions
ST-C: Writing – original draft, Conceptualization, Investigation. CC-R: Investigation, Writing – original draft, Conceptualization. VG-C: Writing – original draft, Investigation. DS-M: Conceptualization, Investigation, Writing – original draft. MO-V: Investigation, Writing – original draft. MR-D: Writing – original draft, Investigation. OP-Z: Writing – original draft, Investigation. DL: Writing – review & editing, Investigation. EL-U: Writing – review & editing. CP-P: Conceptualization, Investigation, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The reviewer MA declared a shared parent affiliation with the author(s) MO-V and DL to the handling editor at time of review.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Wu J, Jin Q, Zhang Y, Ji Y, Li J, Liu X, et al. Global burden of cervical cancer: current estimates, temporal trend and future projections based on the GLOBOCAN 2022. J Natl Cancer Cent. (2025) 5:322–9. doi: 10.1016/j.jncc.2024.11.006
2. Balasubramaniam SD, Balakrishnan V, Oon CE, and Kaur G. Key molecular events in cervical cancer development. Medicina. (2019) 55:384. doi: 10.3390/medicina55070384
3. Kaur H, Silverman PM, Iyer RB, Verschraegen CF, Eifel PJ, and Charnsangavej C. Diagnosis, staging, and surveillance of cervical carcinoma. Am J Roentgenol. (2003) 180:1621–31. doi: 10.2214/ajr.180.6.1801621
4. Gennigens C, Jerusalem G, Lapaille L, Cuypere MD, Streel S, Kridelka F, et al. Recurrent or primary metastatic cervical cancer: current and future treatments. ESMO Open. (2022) 7:100579. doi: 10.1016/j.esmoop.2022.100579
5. Agarwal N, Mishra PK, Das S, Kumar A, Sharma M, Panwar D, et al. Emerging Trends in cervical cancer Treatment: Transitioning from traditional to innovative delivery strategies. Int J Pharm. (2025) 681:125878. doi: 10.1016/j.ijpharm.2025.125878
6. Khan SU, Fatima K, Aisha S, and Malik F. Unveiling the mechanisms and challenges of cancer drug resistance. Cell Commun Signal. (2024) 22:109. doi: 10.1186/s12964-023-01302-1
7. Mattick JS, Amaral PP, Carninci P, Carpenter S, Chang HY, Chen L-L, et al. Long non-coding RNAs: definitions, functions, challenges and recommendations. Nat Rev Mol Cell Biol. (2023) 24:430–47. doi: 10.1038/s41580-022-00566-8
8. Yan H and Bu P. Non-coding RNA in cancer. Essays Biochem. (2021) 65:625–39. doi: 10.1042/ebc20200032
9. Tilsed CM, Fisher SA, Nowak AK, Lake RA, and Lesterhuis WJ. Cancer chemotherapy: insights into cellular and tumor microenvironmental mechanisms of action. Front Oncol. (2022) 12:960317. doi: 10.3389/fonc.2022.960317
10. Liu K, Gao L, Ma X, Huang J-J, Chen J, Zeng L, et al. Long non-coding RNAs regulate drug resistance in cancer. Mol Cancer. (2020) 19:54. doi: 10.1186/s12943-020-01162-0
11. Dean M, Moitra K, and Allikmets R. The human ATP-binding cassette (ABC) transporter superfamily. Hum Mutat. (2022) 43:1162–82. doi: 10.1002/humu.24418
12. Xue X and Liang X-J. Overcoming drug efflux-based multidrug resistance in cancer with nanotechnology. Chin J Cancer. (2012) 31:100–9. doi: 10.5732/cjc.011.10326
13. Wang H, Qin R, Guan A, Yao Y, Huang Y, Jia H, et al. HOTAIR enhanced paclitaxel and doxorubicin resistance in gastric cancer cells partly through inhibiting miR-217 expression. J Cell Biochem. (2018) 119:7226–34. doi: 10.1002/jcb.26901
14. Fang Z, Chen W, Yuan Z, Liu X, and Jiang H. LncRNA-MALAT1 contributes to the cisplatin-resistance of lung cancer by upregulating MRP1 and MDR1 via STAT3 activation. BioMed Pharmacother. (2018) 101:536–42. doi: 10.1016/j.biopha.2018.02.130
15. Huang L, Liao L-M, Liu A-W, Wu J-B, Cheng X-L, Lin J-X, et al. Overexpression of long noncoding RNA HOTAIR predicts a poor prognosis in patients with cervical cancer. Arch Gynecol Obstet. (2014) 290:717–23. doi: 10.1007/s00404-014-3236-2
16. Liang T, Wang Y, Jiao Y, Cong S, Jiang X, Dong L, et al. LncRNA MALAT1 accelerates cervical carcinoma proliferation by suppressing miR-124 expression in cervical tumor cells. J Oncol. (2021) 2021:8836078. doi: 10.1155/2021/8836078
17. Wang Y, Zhang D, Wu K, Zhao Q, Nie Y, and Fan D. Long noncoding RNA MRUL promotes ABCB1 expression in multidrug-resistant gastric cancer cell sublines. Mol Cell Biol. (2014) 34:3182–93. doi: 10.1128/mcb.01580-13
18. Alhmoud JF, Woolley JF, Moustafa A-EA, and Malki MI. DNA damage/repair management in cancers. Cancers. (2020) 12:1050. doi: 10.3390/cancers12041050
19. Neophytou CM, Trougakos IP, Erin N, and Papageorgis P. Apoptosis deregulation and the development of cancer multi-drug resistance. Cancers. (2021) 13:4363. doi: 10.3390/cancers13174363
20. Su M, Wang H, Wang W, Wang Y, Ouyang L, Pan C, et al. LncRNAs in DNA damage response and repair in cancer cells. Acta Bioch Bioph Sin. (2018) 50:433–9. doi: 10.1093/abbs/gmy022
21. Cerbinskaite A, Mukhopadhyay A, Plummer ER, Curtin NJ, and Edmondson RJ. Defective homologous recombination in human cancers. Cancer Treat Rev. (2012) 38:89–100. doi: 10.1016/j.ctrv.2011.04.015
22. Liu L, Chen Y, Huang Y, Cao K, Liu T, Shen H, et al. Long non-coding RNA ANRIL promotes homologous recombination-mediated DNA repair by maintaining ATR protein stability to enhance cancer resistance. Mol Cancer. (2021) 20:94. doi: 10.1186/s12943-021-01382-y
23. Chen Y, Shen H, Liu T, Cao K, Wan Z, Du Z, et al. ATR-binding lncRNA ScaRNA2 promotes cancer resistance through facilitating efficient DNA end resection during homologous recombination repair. J Exp Clin Cancer Res. (2023) 42:256. doi: 10.1186/s13046-023-02829-4
24. Wang L and Zhu H. Long non-coding nuclear paraspeckle assembly transcript 1 acts as prognosis biomarker and increases cell growth and invasion in cervical cancer by sequestering microRNA-101. Mol Med Rep. (2018) 17:2771–7. doi: 10.3892/mmr.2017.8186
25. Guo H-M, Yang S-H, Zhao S-Z, Li L, Yan M-T, and Fan M-C. LncRNA NEAT1 regulates cervical carcinoma proliferation and invasion by targeting AKT/PI3K. Eur Rev Med Pharmacol Sci. (2018) 22:4090–7. doi: 10.26355/eurrev_201807_15400
26. Zhang D, Sun G, Zhang H, Tian J, and Li Y. Long non-coding RNA ANRIL indicates a poor prognosis of cervical cancer and promotes carcinogenesis via PI3K/Akt pathways. BioMed Pharmacother. (2017) 85:511–6. doi: 10.1016/j.biopha.2016.11.058
27. Zhou D, Duan Z, Li Z, Ge F, Wei R, and Kong L. The significance of glycolysis in tumor progression and its relationship with the tumor microenvironment. Front Pharmacol. (2022) 13:1091779. doi: 10.3389/fphar.2022.1091779
28. Farzaneh M, Dari MAG, Anbiyaiee A, Najafi S, Dayer D, Salehi AM, et al. Emerging roles of the long non-coding RNA NEAT1 in gynecologic cancers. J Cell Commun Signal. (2023) 17:531–47. doi: 10.1007/s12079-023-00746-x
29. Cao L, Abudureheman W, Shen G, Ouyang Y, Yang W, Zhao Q, et al. LncRNA NEAT1 regulation of the miR-101-3p/RAC1 axis affects cervical cancer aerobic glycolysis and progression. Sci Rep. (2025) 15:17436. doi: 10.1038/s41598-025-01698-5
30. Wang Y, Cao Z, Liu F, and Ou Y. Clinical significance of activated Wnt/β-catenin signaling in apoptosis inhibition of oral cancer. Open Life Sci. (2021) 16:1045–52. doi: 10.1515/biol-2021-0104
31. Zhang S, Xiong X, and Sun Y. Functional characterization of SOX2 as an anticancer target. Signal Transduct Target Ther. (2020) 5:135. doi: 10.1038/s41392-020-00242-3
32. Zhang W-Y, Liu Y-J, He Y, and Chen P. Down-regulation of long non-coding RNA ANRIL inhibits the proliferation, migration and invasion of cervical cancer cells. Cancer biomark. (2018) 23:243–53. doi: 10.3233/cbm-181467
33. Wang Q, Chen C, Xu X, Shu C, Cao C, Wang Z, et al. APAF1-binding long noncoding RNA promotes tumor growth and multidrug resistance in gastric cancer by blocking apoptosome assembly. Adv Sci. (2022) 9:2201889. doi: 10.1002/advs.202201889
34. Zhu Y, Zhou B, Hu X, Ying S, Zhou Q, Xu W, et al. LncRNA LINC00942 promotes chemoresistance in gastric cancer by suppressing MSI2 degradation to enhance c-Myc mRNA stability. Clin Transl Med. (2022) 12:e703. doi: 10.1002/ctm2.703
35. Pandey S and Chandravati C. Autophagy in cervical cancer: an emerging therapeutic target. Asian Pac J Cancer Prev. (2012) 13:4867–71. doi: 10.7314/apjcp.2012.13.10.4867
36. Peng X, Gong F, Chen Y, Jiang Y, Liu J, Yu M, et al. Autophagy promotes paclitaxel resistance of cervical cancer cells: involvement of Warburg effect activated hypoxia-induced factor 1-α-mediated signaling. Cell Death Dis. (2014) 5:e1367–7. doi: 10.1038/cddis.2014.297
37. Jiang L, Xia Y, Zhong T, Zhang H, Jin Q, Li F, et al. HIF2A overexpression reduces cisplatin sensitivity in cervical cancer by inducing excessive autophagy. Transl Cancer Res. (2019) 9:75–84. doi: 10.21037/tcr.2019.11.17
38. Zhou J, Kang Y, Chen L, Wang H, Liu J, Zeng S, et al. The drug-resistance mechanisms of five platinum-based antitumor agents. Front Pharmacol. (2020) 11:343. doi: 10.3389/fphar.2020.00343
39. Yang F, Zhang H, Mei Y, and Wu M. Reciprocal regulation of HIF-1α and lincRNA-p21 modulates the warburg effect. Mol Cell. (2014) 53:88–100. doi: 10.1016/j.molcel.2013.11.004
40. Trujano-Camacho S, León DC, Pérez-Yepez E, Contreras-Romero C, Coronel-Hernandez J, Millan-Catalan O, et al. HOTAIR promotes the hyperactivation of PI3K/akt and wnt/β-catenin signaling pathways via PTEN hypermethylation in cervical cancer. Cells. (2024) 13:1484. doi: 10.3390/cells13171484
41. Zhou C, Yi C, Yi Y, Qin W, Yan Y, Dong X, et al. LncRNA PVT1 promotes gemcitabine resistance of pancreatic cancer via activating Wnt/β-catenin and autophagy pathway through modulating the miR-619-5p/Pygo2 and miR-619-5p/ATG14 axes. Mol Cancer. (2020) 19:118. doi: 10.1186/s12943-020-01237-y
42. Zhang F, Wang H, Yu J, Yao X, Yang S, Li W, et al. LncRNA CRNDE attenuates chemoresistance in gastric cancer via SRSF6-regulated alternative splicing of PICALM. Mol Cancer. (2021) 20:6. doi: 10.1186/s12943-020-01299-y
43. Luo Y, Zheng S, Wu Q, Wu J, Zhou R, Wang C, et al. (lncRNA) EIF3J-DT induces chemoresistance of gastric cancer via autophagy activation. Autophagy. (2021) 17:4083–101. doi: 10.1080/15548627.2021.1901204
44. Aziz MH and Ahmad A. Epigenetic basis of cancer drug resistance. Cancer Drug Resist. (2020) 3:113–6. doi: 10.20517/cdr.2020.06
45. Egger G, Liang G, Aparicio A, and Jones PA. Epigenetics in human disease and prospects for epigenetic therapy. Nature. (2004) 429:457–63. doi: 10.1038/nature02625
46. Masuda K, Banno K, Yanokura M, Tsuji K, Kobayashi Y, Kisu I, et al. Association of epigenetic inactivation of the WRN gene with anticancer drug sensitivity in cervical cancer cells. Oncol Rep. (2012) 28:1146–52. doi: 10.3892/or.2012.1912
47. Chen C-C, Lee K-D, Pai M-Y, Chu P-Y, Hsu C-C, Chiu C-C, et al. Changes in DNA methylation are associated with the development of drug resistance in cervical cancer cells. Cancer Cell Int. (2015) 15:98. doi: 10.1186/s12935-015-0248-3
48. Singh DK, Pandita RK, Singh M, Chakraborty S, Hambarde S, Ramnarain D, et al. MOF suppresses replication stress and contributes to resolution of stalled replication forks. Mol Cell Biol. (2018) 38:e00484–17. doi: 10.1128/mcb.00484-17
49. Salemi LM, Maitland MER, Yefet ER, and Schild-Poulter C. Inhibition of HDAC6 activity through interaction with RanBPM and its associated CTLH complex. BMC Cancer. (2017) 17:460. doi: 10.1186/s12885-017-3430-2
50. Portoso M, Ragazzini R, Brenčič Ž, Moiani A, Michaud A, Vassilev I, et al. PRC2 is dispensable for HOTAIR-mediated transcriptional repression. EMBO J. (2017) 36:981–94. doi: 10.15252/embj.201695335
51. Ren J, Ding L, Zhang D, Shi G, Xu Q, Shen S, et al. Carcinoma-associated fibroblasts promote the stemness and chemoresistance of colorectal cancer by transferring exosomal lncRNA H19. Theranostics. (2018) 8:3932–48. doi: 10.7150/thno.25541
52. Xiong G, Liu C, Yang G, Feng M, Xu J, Zhao F, et al. Long noncoding RNA GSTM3TV2 upregulates LAT2 and OLR1 by competitively sponging let-7 to promote gemcitabine resistance in pancreatic cancer. J Hematol Oncol. (2019) 12:97. doi: 10.1186/s13045-019-0777-7
53. Zhu M, Yang L, and Wang X. NEAT1 knockdown suppresses the cisplatin resistance in ovarian cancer by regulating miR-770-5p/PARP1 axis. Cancer Manag Res. (2020) 12:7277–89. doi: 10.2147/cmar.s257311
54. Wen X, Liu S, Sheng J, and Cui M. Recent advances in the contribution of noncoding RNAs to cisplatin resistance in cervical cancer. PeerJ. (2020) 8:e9234. doi: 10.7717/peerj.9234
55. Abu-Rustum NR, Yashar CM, Bean S, Bradley K, Campos SM, Chon HS, et al. NCCN guidelines insights: cervical cancer, version 1.2020: featured updates to the NCCN guidelines. J Natl Compr Cancer Netw. (2020) 18:660–6. doi: 10.6004/jnccn.2020.0027
56. Housman G, Byler S, Heerboth S, Lapinska K, Longacre M, Snyder N, et al. Drug resistance in cancer: an overview. Cancers. (2014) 6:1769–92. doi: 10.3390/cancers6031769
57. Chen S-H and Chang J-Y. New insights into mechanisms of cisplatin resistance: from tumor cell to microenvironment. Int J Mol Sci. (2019) 20:4136. doi: 10.3390/ijms20174136
58. Galluzzi L, Senovilla L, Vitale I, Michels J, Martins I, Kepp O, et al. Molecular mechanisms of cisplatin resistance. Oncogene. (2012) 31:1869–83. doi: 10.1038/onc.2011.384
59. Bellmunt J, Paz-Ares L, Cuello M, Cecere FL, Albiol S, Guillem V, et al. Gene expression of ERCC1 as a novel prognostic marker in advanced bladder cancer patients receiving cisplatin-based chemotherapy. Ann Oncol. (2007) 18:522–8. doi: 10.1093/annonc/mdl435
60. Siddik ZH. Cisplatin: mode of cytotoxic action and molecular basis of resistance. Oncogene. (2003) 22:7265–79. doi: 10.1038/sj.onc.1206933
61. Zhang Y, Shu YM, Wang SF, Da BH, Wang ZH, and Li HB. Stabilization of mismatch repair gene PMS2 by glycogen synthase kinase 3β is implicated in the treatment of cervical carcinoma. BMC Cancer. (2010) 10:58. doi: 10.1186/1471-2407-10-58
62. Brozovic A and Osmak M. Activation of mitogen-activated protein kinases by cisplatin and their role in cisplatin-resistance. Cancer Lett. (2007) 251:1–16. doi: 10.1016/j.canlet.2006.10.007
63. Brozovic A, Fritz G, Christmann M, Zisowsky J, Jaehde U, Osmak M, et al. Long-term activation of SAPK/JNK, p38 kinase and fas-L expression by cisplatin is attenuated in human carcinoma cells that acquired drug resistance. Int J Cancer. (2004) 112:974–85. doi: 10.1002/ijc.20522
64. Venkatraman M, Anto RJ, Nair A, Varghese M, and Karunagaran D. Biological and chemical inhibitors of NF-κB sensitize SiHa cells to cisplatin-induced apoptosis. Mol Carcinog. (2005) 44:51–9. doi: 10.1002/mc.20116
65. Cao L, Jin H, Zheng Y, Mao Y, Fu Z, Li X, et al. DANCR-mediated microRNA-665 regulates proliferation and metastasis of cervical cancer through the ERK/SMAD pathway. Cancer Sci. (2019) 110:913–25. doi: 10.1111/cas.13921
66. Yao T, Lu R, Zhang J, Fang X, Fan L, Huang C, et al. Growth arrest-specific 5 attenuates cisplatin-induced apoptosis in cervical cancer by regulating STAT3 signaling via miR-21. J Cell Physiol. (2019) 234:9605–15. doi: 10.1002/jcp.27647
67. Iden M, Fye S, Li K, Chowdhury T, Ramchandran R, and Rader JS. The lncRNA PVT1 contributes to the cervical cancer phenotype and associates with poor patient prognosis. PloS One. (2016) 11:e0156274. doi: 10.1371/journal.pone.0156274
68. Feng L-L, Shen F-R, Zhou J-H, and Chen Y-G. Expression of the lncRNA ZFAS1 in cervical cancer and its correlation with prognosis and chemosensitivity. Gene. (2019) 696:105–12. doi: 10.1016/j.gene.2019.01.025
69. Wang B, Huang Z, Gao R, Zeng Z, Yang W, Sun Y, et al. Expression of long noncoding RNA urothelial cancer associated 1 promotes cisplatin resistance in cervical cancer. Cancer Biotherapy Radiopharm. (2017) 32:101–10. doi: 10.1089/cbr.2016.2156
70. Wang B, Li L, Wu Z, Qian X, Yu W, and Huang Z. Long noncoding RNA UCA1 knockdown inhibits cisplatin-resistant cervical cancer tumorigenesis via the miR-195-5p/IKBKB axis. Acta Biochim Biophys Sin. (2025) 57:1492–506. doi: 10.3724/abbs.2025032
71. Liu E, Liu Z, and Zhou Y. Carboplatin-docetaxel-induced activity against ovarian cancer is dependent on up-regulated lncRNA PVT1. Int J Clin Exp Pathol. (2015) 8:3803–10.
72. Jin X, Chen X, Zhang Z, Hu W, Ou R, Li S, et al. Long noncoding RNA SNHG12 promotes the progression of cervical cancer via modulating miR-125b/STAT3 axis. J Cell Physiol. (2019) 234:6624–32. doi: 10.1002/jcp.27403
73. Roychowdhury A, Samadder S, Das P, Mazumder DI, Chatterjee A, Addya S, et al. Deregulation of H19 is associated with cervical carcinoma. Genomics. (2020) 112:961–70. doi: 10.1016/j.ygeno.2019.06.012
74. Ai L, Chen J, Yan H, He Q, Luo P, Xu Z, et al. Research status and outlook of PD-1/PD-L1 inhibitors for cancer therapy. Drug Des Dev Ther. (2020) 14:3625–49. doi: 10.2147/dddt.s267433
75. Wang Q-M, Lian G-Y, Song Y, Huang Y-F, and Gong Y. LncRNA MALAT1 promotes tumorigenesis and immune escape of diffuse large B cell lymphoma by sponging miR-195. Life Sci. (2019) 231:116335. doi: 10.1016/j.lfs.2019.03.040
76. Zhou W, Zhang M, Liu C, Kang Y, Wang J, and Yang X. Long noncoding RNA LINC00473 drives the progression of pancreatic cancer via upregulating programmed death-ligand 1 by sponging microRNA-195-5p. J Cell Physiol. (2019) 234:23176–89. doi: 10.1002/jcp.28884
77. Zhang M, Liu S, Fu C, Wang X, Zhang M, Liu G, et al. LncRNA KB-1471A8.2 overexpression suppresses cell proliferation and migration and antagonizes the paclitaxel resistance of ovarian cancer cells. Cancer Biotherapy Radiopharm. (2019) 34:316–24. doi: 10.1089/cbr.2018.2698
78. Yang F, Zhang L, Huo X, Yuan J, Xu D, Yuan S, et al. Long noncoding RNA high expression in hepatocellular carcinoma facilitates tumor growth through enhancer of zeste homolog 2 in humans. Hepatology. (2011) 54:1679–89. doi: 10.1002/hep.24563
79. Guo J-L, Tang T, Li J-H, Yang Y-H, Zhang L, and Quan Y. LncRNA HEIH enhances paclitaxel-tolerance of endometrial cancer cells via activation of MAPK signaling pathway. Pathol Oncol Res. (2020) 26:1757–66. doi: 10.1007/s12253-019-00718-w
80. An J, Lv W, and Zhang Y. LncRNA NEAT1 contributes to paclitaxel resistance of ovarian cancer cells by regulating ZEB1 expression via miR-194. OncoTargets Ther. (2017) 10:5377–90. doi: 10.2147/ott.s147586
81. Liu Y, Wang J, Dong L, Xia L, Zhu H, Li Z, et al. Long noncoding RNA HCP5 regulates pancreatic cancer gemcitabine (GEM) resistance by sponging hsa-miR-214-3p to target HDGF. OncoTargets Ther. (2019) 12:8207–16. doi: 10.2147/ott.s222703
82. Shi W, Zhang C, Ning Z, Hua Y, Li Y, Chen L, et al. Long non-coding RNA LINC00346 promotes pancreatic cancer growth and gemcitabine resistance by sponging miR-188-3p to derepress BRD4 expression. J Exp Clin Cancer Res. (2019) 38:60. doi: 10.1186/s13046-019-1055-9
83. Wang C, Zhang T, and Zhang C. LncRNA AFAP1-AS1 exhibits oncogenic characteristics and promotes gemcitabine-resistance of cervical cancer cells through miR-7-5p/EGFR axis. Oncol Res. (2024) 32:1867–79. doi: 10.32604/or.2024.044547
84. Zou M, Cheng X, and Liu R. lncRNA DLG1-AS1 promotes cervical cancer cell gemcitabine resistance by regulating miR-16-5p/HDGF. J Obstet Gynaecol Res. (2022) 48:1836–47. doi: 10.1111/jog.15245
85. Rui X, Xu Y, Huang Y, Ji L, and Jiang X. lncRNA DLG1-AS1 promotes cell proliferation by competitively binding with miR-107 and up-regulating ZHX1 expression in cervical cancer. Cell Physiol Biochem. (2018) 49:1792–803. doi: 10.1159/000493625
86. Lampropoulou D-I, Aravantinos G, Katifelis H, Lazaris F, Laschos K, Theodosopoulos T, et al. Long non-coding RNA polymorphisms and prediction of response to chemotherapy based on irinotecan in patients with metastatic colorectal cancer. Cancer biomark. (2019), 25:1–9. doi: 10.3233/cbm-182383
87. Sun F, Liang W, and Qian J. The identification of CRNDE, H19, UCA1 and HOTAIR as the key lncRNAs involved in oxaliplatin or irinotecan resistance in the chemotherapy of colorectal cancer based on integrative bioinformatics analysis. Mol Med Rep. (2019) 20:3583–96. doi: 10.3892/mmr.2019.10588
88. Kim HJ, Lee DW, Yim GW, Nam EJ, Kim S, Kim SW, et al. Long non-coding RNA HOTAIR is associated with human cervical cancer progression. Int J Oncol. (2014) 46:521–30. doi: 10.3892/ijo.2014.2758
89. Zhang W, Wu Q, Liu Y, Wang X, Ma C, and Zhu W. LncRNA HOTAIR Promotes Chemoresistance by Facilitating Epithelial to Mesenchymal Transition through miR-29b/PTEN/PI3K Signaling in Cervical Cancer. Cells Tissues Organs. (2022) 211:16–29. doi: 10.1159/000519844
90. Zhou Y-H, Cui Y-H, Wang T, and Luo Y. Chapter Four Long non-coding RNA HOTAIR in cervical cancer: Molecular marker, mechanistic insight, and therapeutic target. Adv Clin Chem. (2020) 97:117–40. doi: 10.1016/bs.acc.2019.12.004
91. Shao X, Zheng X, Ma D, Liu Y, and Liu G. Inhibition of lncRNA-NEAT1 sensitizes 5-Fu resistant cervical cancer cells through de-repressing the microRNA-34a/LDHA axis. Biosci Rep. (2021) 41:BSR20200533. doi: 10.1042/bsr20200533
92. Wei H, Qiu Y-Q, Zeng Q-S, Wang S-F, and Yi C-J. LncRNA UCA1 regulates proliferation, migration and invasion of cervical cancer cells by targeting miR-145. Eur Rev Med Pharmacol Sci. (2020) 24:3555–64. doi: 10.26355/eurrev_202004_20816
93. Campos-Viguri GE, Peralta-Zaragoza O, Jiménez-Wences H, Longinos-González AE, Castañón-Sánchez CA, Ramírez-Carrillo M, et al. MiR-23b-3p reduces the proliferation, migration and invasion of cervical cancer cell lines via the reduction of c-Met expression. Sci Rep. (2020) 10:3256. doi: 10.1038/s41598-020-60143-x
94. Xian Z, Hu B, Wang T, Zeng J, Cai J, Zou Q, et al. lncRNA UCA1 contributes to 5-fluorouracil resistance of colorectal cancer cells through miR-23b-3p/ZNF281 axis. OncoTargets Ther. (2020) 13:7571–83. doi: 10.2147/ott.s258727
95. Yang C, Pan Y, and Deng SP. Downregulation of lncRNA CCAT1 enhances 5-fluorouracil sensitivity in human colon cancer cells. BMC Mol Cell Biol. (2019) 20:9. doi: 10.1186/s12860-019-0188-1
96. Vora C and Gupta S. Targeted therapy in cervical cancer. ESMO Open. (2018) 3:e000462. doi: 10.1136/esmoopen-2018-000462
97. Zhang J, Pang X, Lei L, Zhang J, Zhang X, Chen Z, et al. LncRNA CRART16/miR-122-5p/FOS axis promotes angiogenesis of gastric cancer by upregulating VEGFD expression. Aging (Albany NY). (2022) 14:4137–57. doi: 10.18632/aging.204078
98. Huang W, Zhang H, Tian Y, Li Y, Li J, Zhong X, et al. LncRNA SNHG11 enhances bevacizumab resistance in colorectal cancer by mediating miR-1207-5p/ABCC1 axis. Anti-Cancer Drugs. (2022) 33:575–86. doi: 10.1097/cad.0000000000001289
99. Hu J, Dong S, Pei Y, Wang J, Zhang J, and Wei X. LncRNA MITA1 promotes gefitinib resistance by inducing autophagy in lung cancer cells. Biochem Biophys Res Commun. (2021) 551:21–6. doi: 10.1016/j.bbrc.2021.02.130
100. Chen Z, Chen Q, Cheng Z, Gu J, Feng W, Lei T, et al. Long non-coding RNA CASC9 promotes gefitinib resistance in NSCLC by epigenetic repression of DUSP1. Cell Death Dis. (2020) 11:858. doi: 10.1038/s41419-020-03047-y
101. Xu T, Yan S, Wang M, Jiang L, Ma P, Lu B, et al. LncRNA UCA1 induces acquired resistance to gefitinib by epigenetically silencing CDKN1A expression in non-small-cell lung cancer. Front Oncol. (2020) 10:656. doi: 10.3389/fonc.2020.00656
102. Yang T-J, Wang L, Zhang Y, Zheng J-D, and Liu L. LncRNA UCA1 regulates cervical cancer survival and EMT occurrence by targeting miR-155. Eur Rev Med Pharmacol Sci. (2020) 24:9869–79. doi: 10.26355/eurrev_202010_23197
103. Chen Z-Y, Liu H-Y, Jiang N, and Yuan J-M. LncRNA HOST2 enhances gefitinib-resistance in non-small cell lung cancer by down-regulating miRNA-621. Eur Rev Med Pharmacol Sci. (2019) 23:9939–46. doi: 10.26355/eurrev_201911_19560
104. Jin X, Liu X, Zhang Z, and Guan Y. lncRNA CCAT1 acts as a microRNA-218 sponge to increase gefitinib resistance in NSCLC by targeting HOXA1. Mol Ther - Nucleic Acids. (2020) 19:1266–75. doi: 10.1016/j.omtn.2020.01.006
105. Zhang Y, Jia L-G, Wang P, Li J, Tian F, Chu Z-P, et al. The expression and significance of lncRNA HOST2 and microRNA let-7b in HPV-positive cervical cancer tissues and cell lines. Eur Rev Med Pharmacol Sci. (2019) 23:2380–90. doi: 10.26355/eurrev_201903_17384
106. Shen H, Wang L, Xiong J, Ren C, Gao C, Ding W, et al. Long non-coding RNA CCAT1 promotes cervical cancer cell proliferation and invasion by regulating the miR-181a-5p/MMP14 axis. Cell Cycle. (2019) 18:1110–21. doi: 10.1080/15384101.2019.1609829
107. Li W, Zhai L, Wang H, Liu C, Zhang J, Chen W, et al. Downregulation of LncRNA GAS5 causes trastuzumab resistance in breast cancer. Oncotarget. (2016) 7:27778–86. doi: 10.18632/oncotarget.8413
108. Xiang Z, Song S, Zhu Z, Sun W, Gifts JE, Sun S, et al. LncRNAs GIHCG and SPINT1-AS1 are crucial factors for pan-cancer cells sensitivity to lapatinib. Front Genet. (2019) 10:25. doi: 10.3389/fgene.2019.00025
109. Li D, Li C, Chen Y, Teng L, Cao Y, Wang W, et al. LncRNA HOTAIR induces sunitinib resistance in renal cancer by acting as a competing endogenous RNA to regulate autophagy of renal cells. Cancer Cell Int. (2020) 20:338. doi: 10.1186/s12935-020-01419-0
110. Qu L, Ding J, Chen C, Wu Z-J, Liu B, Gao Y, et al. Exosome-transmitted lncARSR promotes sunitinib resistance in renal cancer by acting as a competing endogenous RNA. Cancer Cell. (2016) 29:653–68. doi: 10.1016/j.ccell.2016.03.004
111. He M, Yang H, Shi H, Hu Y, Chang C, Liu S, et al. Sunitinib increases the cancer stem cells and vasculogenic mimicry formation via modulating the lncRNA-ECVSR/ERβ/Hif2-α signaling. Cancer Lett. (2022) 524:15–28. doi: 10.1016/j.canlet.2021.08.028
112. Manzo-Merino J, Contreras-Paredes A, Vázquez-Ulloa E, Rocha-Zavaleta L, Fuentes-Gonzalez AM, and Lizano M. The role of signaling pathways in cervical cancer and molecular therapeutic targets. Arch Med Res. (2014) 45:525–39. doi: 10.1016/j.arcmed.2014.10.008
113. Smalley I and Smalley KSM. ERK inhibition: A new front in the war against MAPK pathway–driven cancers? Cancer Discov. (2018) 8:140–2. doi: 10.1158/2159-8290.cd-17-1355
114. Mishra R, Patel H, Alanazi S, Kilroy MK, and Garrett JT. PI3K inhibitors in cancer: clinical implications and adverse effects. Int J Mol Sci. (2021) 22:3464. doi: 10.3390/ijms22073464
115. Wang B, Li X, Liu L, and Wang M. β-Catenin: oncogenic role and therapeutic target in cervical cancer. Biol Res. (2020) 53:33. doi: 10.1186/s40659-020-00301-7
116. Trujano-Camacho S, León DC, Delgado-Waldo I, Coronel-Hernández J, Millan-Catalan O, Hernández-Sotelo D, et al. Inhibition of wnt-β-catenin signaling by ICRT14 drug depends of post-transcriptional regulation by HOTAIR in human cervical cancer heLa cells. Front Oncol. (2021) 11:729228. doi: 10.3389/fonc.2021.729228
Keywords: lncRNAs, non-coding RNAs, chemoresistance, cancer, cervical cancer
Citation: Trujano-Camacho S, Contreras-Romero C, García-Castillo V, Sánchez-Marín D, Olvera-Valencia M, Rodríguez-Dorantes M, Peralta-Zaragoza O, de León DC, López-Urrutia E and Pérez-Plasencia C (2025) Long noncoding RNAs involved in therapeutic response: implications for cervical cancer drug resistance. Front. Oncol. 15:1644104. doi: 10.3389/fonc.2025.1644104
Received: 09 June 2025; Accepted: 30 September 2025;
Published: 21 October 2025.
Edited by:
Sharon R Pine, University of Colorado Anschutz Medical Campus, United StatesReviewed by:
Rebeca Debora Martinez-Contreras, Meritorious Autonomous University of Puebla, MexicoMarco Alonso Andonegui, Subdirection of Basic Investigation, National Institute of Cancerology (INCAN), Mexico
Copyright © 2025 Trujano-Camacho, Contreras-Romero, García-Castillo, Sánchez-Marín, Olvera-Valencia, Rodríguez-Dorantes, Peralta-Zaragoza, de León, López-Urrutia and Pérez-Plasencia. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Carlos Pérez-Plasencia, Y2FybG9zLnBwbGFzQGdtYWlsLmNvbQ==