 Alfred Lee Edgar1
Alfred Lee Edgar1 Luis Felipe Dias Lopes2*
Luis Felipe Dias Lopes2* Eduarda Grando Lopes3
Eduarda Grando Lopes3 Izabella Danezi Felin4
Izabella Danezi Felin4 Carlos Roberto Felin5
Carlos Roberto Felin5 João Francisco Pollo Gaspary6
João Francisco Pollo Gaspary6- 1Research and Development Department, ElastroCrete, LLC, Veyo, UT, United States
- 2Department of Administrative Sciences, Federal University of Santa Maria, Santa Maria, Brazil
- 3Veterinary Medicine Course, Federal University of Santa Maria, Santa Maria, Brazil
- 4Department of Pathology, Federal University of Santa Maria, Santa Maria, Brazil
- 5Oncocenter Clinic, Santa Maria, Brazil
- 6Instituto AuBento – Center for Teaching, Clinical Practice, and Research in Orthomolecular and Translational Health Innovation, Santa Maria, Brazil
Background: The tumor microenvironment (TME) is characterized by a reversed pH gradient—acidic extracellular and alkaline intracellular conditions—arising from mitochondrial dysfunction, metabolic reprogramming, and dysregulated proton transport. These alterations establish a permissive niche for tumor progression, immune evasion, and resistance to therapy. Although the TME is increasingly recognized as a key determinant of cancer behavior, effective and targeted strategies for its bioenergetic reprogramming remain scarce.
Objectives: This study introduces and evaluates Eigen/Zundel Complexes-Rich Water (EZC-Rich Water) as a novel hydrated proton supplementation strategy capable of targeting Warburg-induced proton dysregulation and restoring mitochondrial function, while stabilizing electrochemical membrane dynamics within the TME.
Methods: A structured translational research design was implemented, combining Work Breakdown Structure (WBS), Open Innovation, and Design Thinking methodologies. This approach enabled the identification of Fundamental Points of View (FPV’s)—physiological targets underlying TME dysfunction—and Critical Success Factors (CSF’s)—mechanistic requirements for therapeutic efficacy. Multicriteria decision analysis was applied to integrate findings from oncology, bioenergetics, and physical chemistry, linking hydrated proton supplementation to improved zeta potential, electrosmotic flow, mitochondrial coupling, and redox regulation.
Results: Integrative analyses demonstrated that EZC-Rich Water delivers metastable hydrated proton clusters (H9O4+ and H5O2+) that support selective and efficient proton transfer via the Grotthuss mechanism. This supplementation facilitates compartmentalized acid–base modulation without inducing systemic acidosis, aligning with prioritized FPV’s and validated CSF’s. The proposed strategy shows translational potential to correct pH inversion, optimize oxidative phosphorylation, and restore bioenergetic integrity in the TME.
Conclusion: Hydrated proton supplementation through EZC-Rich Water represents an innovative bioenergetic intervention with potential to reprogram the tumor microenvironment. By targeting core metabolic dysfunctions such as the Warburg effect and mitochondrial uncoupling, this clinically adaptable and low-risk strategy introduces a new paradigm in nutritional oncology. Further preclinical and clinical studies are warranted to validate its efficacy, safety, and translational applicability in oncology and related metabolic disorders.
Systematic review registration: https://www.crd.york.ac.uk/prospero/display_record.php?, identifier CRD420251065137; https://www.crd.york.ac.uk/prospero/display_record.php?, identifier CRD420251022205.
1 Introduction
The tumor microenvironment (TME) represents a highly dynamic and pathophysiologically distinct compartment that sustains cancer progression through metabolic reprogramming, immune evasion, and resistance to therapy (1, 2). A defining hallmark of the TME is the inversion of the normal acid-base gradient, resulting in acidic extracellular pH (pHe) and alkaline intracellular pH (pHi) (3, 4). This dysregulated pH landscape enhances glycolytic flux (the Warburg effect)—a metabolic reprogramming whereby cancer cells preferentially convert glucose to lactate via aerobic glycolysis even in the presence of sufficient oxygen, diverting pyruvate away from mitochondrial oxidative phosphorylation. This shift supports anabolic growth, contributes to extracellular acidification, and promotes invasive phenotypes, while also impairing drug delivery by altering transmembrane electrochemical potentials and protonation-dependent drug activity (5–8).
Cancer cells achieve this pH inversion through coordinated upregulation of proton pumps (H+-ATPases), Na+/H+ exchangers, and carbonic anhydrases—particularly CAIX under hypoxic conditions (9–14). These adaptations are intimately linked to mitochondrial dysfunction, which disrupts proton-coupled oxidative phosphorylation, impairs redox balance, and further reinforces extracellular acidification via lactate and proton export (15–17). The resulting bioenergetic instability not only compromises the effectiveness of conventional therapies but also fosters a microenvironment that is inherently resistant to immune and pharmacological interventions (1, 4, 6, 8).
Despite advances in molecular oncology, clinically effective and targeted strategies for microenvironmental modulation remain limited, especially in the face of therapeutic resistance and metabolic plasticity (18–21). This translational gap underscores the urgent need for innovative approaches capable of restoring acid-base and redox homeostasis within the TME—a recognized driver of tumor progression and poor therapeutic response.
In a previous study, Gaspary et al. (2024) (15) demonstrated that controlled, hormetic administration of carbon dioxide could partially restore mitochondrial and acid-base homeostasis by enhancing carbonic acid availability and modulating proton gradients. These findings provided translational insights into employing physicochemical agents to modulate the biophysical parameters of the tumor microenvironment (TME), thereby opening novel investigative pathways for adjunctive cancer therapies focused on targeted pH regulation.
Building on these translational foundations, the present study introduces Eigen/Zundel Complexes-Rich Water (EZC-Rich Water) as a next-generation nutritional intervention, conceptually developed through a structured innovation framework integrating Open Innovation (22), Design Thinking (23), and multicriteria analysis (24). EZC-Rich Water is designed to modulate the tumor microenvironment by delivering bioavailable hydrated proton complexes—Eigen (H9O4+) and Zundel (H5O2+) (25)—which play central roles in proton transport through the Grotthuss mechanism, supporting localized acid-base modulation and enhancing mitochondrial proton motive force (26, 27). In contrast to conventional acidified solutions, EZC-Rich Water offers a structured and targeted proton delivery system, minimizing disruption to systemic electrolyte balance and reducing the risk of cytotoxic acidification (25, 28, 29).
This paper presents a structured translational synthesis of bioenergetic evidence supporting the use of EZC-Rich Water to reprogram the TME, restore mitochondrial efficiency, optimize redox potential, and stabilize cellular membrane dynamics. By bridging fundamental proton chemistry with cancer metabolism in a systematically integrated framework, we propose hydrated proton supplementation as a novel, clinically feasible, and mechanistically sophisticated adjunct to contemporary oncological strategies. This approach is positioned to address both the biochemical vulnerabilities of the TME and the pressing translational need for innovation in cancer therapy. This approach directly targets the metabolic signature of cancer, notably the Warburg effect and mitochondrial dysfunction, offering a promising pathway for TME reprogramming.
2 Methods
This study followed a translational research design structured through the Work Breakdown Structure (WBS) methodology (30, 31), allowing systematic decomposition of the research question into interdependent and chronologically ordered Work Packages (WP’s). Each WP was conceived to explore a specific layer of evidence required to assess the bioenergetic and clinical feasibility of hydrated proton supplementation with Eigen/Zundel Complexes-Rich Water (EZC-Rich Water) in modulating tumor microenvironment dynamics. The WBS framework was supported by principles of Open Innovation (22) and Design Thinking (23), facilitating conceptual triangulation and hypothesis refinement across bioenergetic, metabolic, and oncological domains. The research project was organized into six interconnected WPs, as summarized below.
2.1 WP1 – strategic project governance and methodological oversight
This initial work package ensured alignment with translational standards and methodological consistency across all stages of the research. A supervisory panel with expertise in bioenergetics, oncology, and biomedical engineering monitored the implementation of each WP, ensuring compliance with PRISMA guidelines in systematic reviews (32) and maintaining documentation according to PROSPERO standards where applicable. Iterative review cycles enabled adjustments in the scope and terminology across WPs as new evidence emerged. The structured methodological approach, outlining the sequential and integrative nature of all Work Packages (WP’s), is depicted in Figure 1.
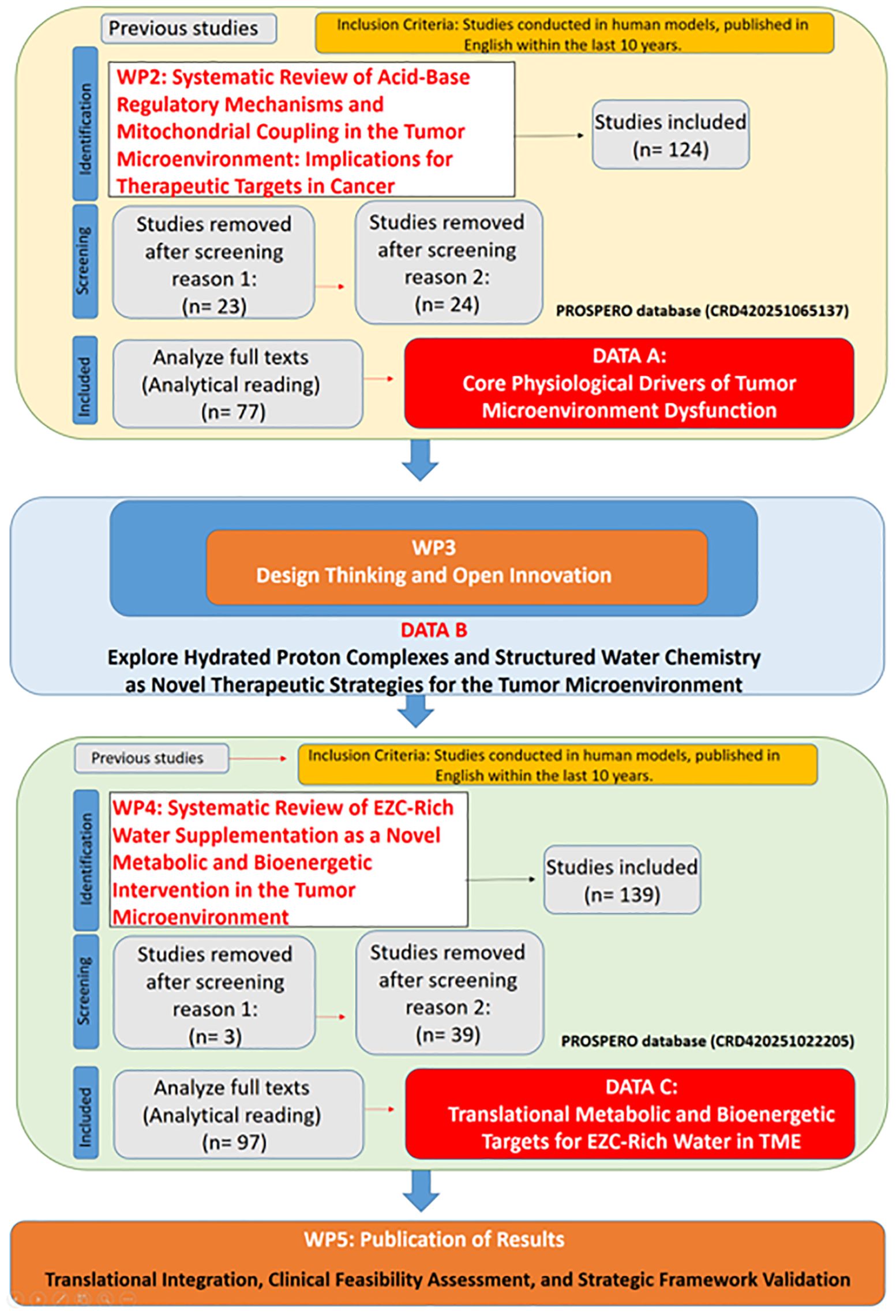
Figure 1. Structured methodological framework for translational research on hydrated proton complexes and bioenergetic modulation in the tumor microenvironment.
2.2 WP2 – systematic review of electrochemical and metabolic regulation in the tumor microenvironment: implications for therapeutic targeting
A targeted systematic review was conducted and formally registered in the PROSPERO database (CRD420251065137) to map the core physiological mechanisms underlying pH inversion and bioenergetic dysregulation within the tumor microenvironment (TME), emphasizing intracellular alkalinization, extracellular acidification, mitochondrial coupling failure, and metal ion-related modulation. Searches were performed in major databases using the following indexed terms: “tumor microenvironment” AND “pH gradient” (20 results); “tumor microenvironment” AND “ion transporters” (13 results); “tumor microenvironment” AND “Warburg effect” AND “therapeutic target” (15 results); “tumor microenvironment” AND “carbonic anhydrase” AND “therapeutic target” (7 results); “tumor microenvironment” AND “metalloproteases” AND “extracellular matrix” (23 results); “tumor microenvironment” AND modulation AND “metal ions” (10 results); “tumor microenvironment” AND “anti-cancer properties” (32 results); and “tumor microenvironment” AND “zeta potential” AND “antitumor effect” (4 results). Filters applied included studies conducted in human models, published in English within the last 10 years. After title and abstract screening, a total of 124 studies were identified. Following full-text review and methodological assessment, 77 studies were ultimately included, prioritizing mechanistic, physiological, and translational findings (Figure 2).
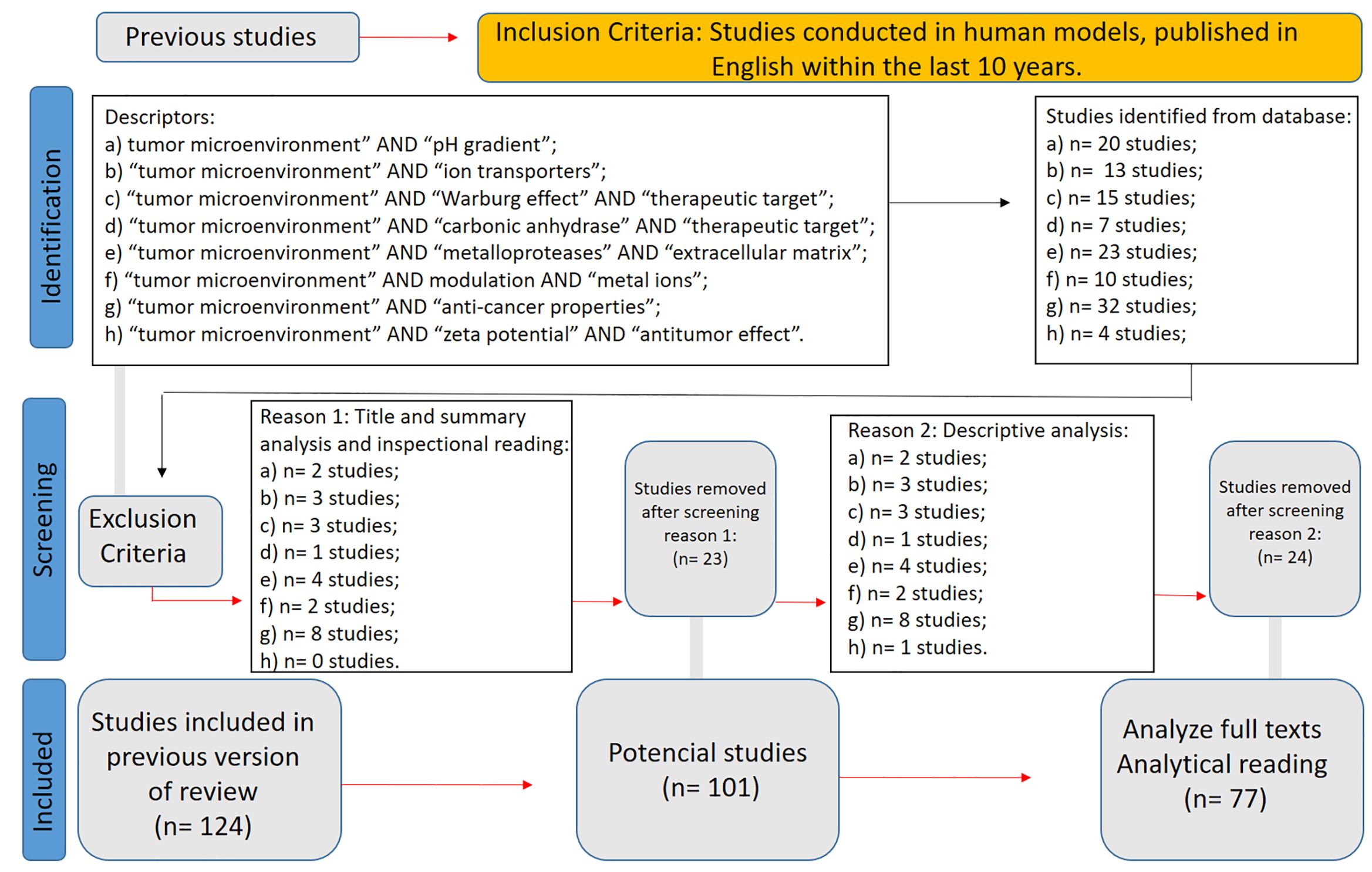
Figure 2. WP2 PRISMA flow diagram.
Based on the findings from WP2, this stage applied a multicriteria analysis adapted from the decision-support framework proposed by Bana e Costa et al. (1999) (24) to systematically identify Fundamental Points of View (FPV’s), representing essential physiological targets relevant for innovative cancer interventions, and Critical Success Factors (CSF’s), defined as mechanistic drivers crucial for effectively modulating tumor pH and mitochondrial bioenergetics. These FPV’s and CSF’s were further refined through an integrative literature review encompassing studies on mitochondrial membrane potential, zeta potential, intracellular redox regulation, and proton-coupled transport mechanisms.
2.3 WP3 – design thinking and open innovation for hydrated proton complexes and structured water chemistry as novel therapeutic strategies for the tumor microenvironment
Based on the multicriteria analysis data provided by WP2, WP3 was structured to systematically explore novel translational opportunities by integrating Open Innovation (22) and Design Thinking (23) frameworks. This WP applied a structured methodological framework to systematically explore the therapeutic potential of hydrated proton complexes, specifically Eigen and Zundel configurations, and structured water chemistry as novel modulators of the tumor microenvironment (TME). An in-depth scientific and translational review was conducted to elucidate the role of these complexes in enhancing hydrated proton mobility via the Grotthuss mechanism, emphasizing their distinct physicochemical characteristics compared to conventional acidified water. Key molecular properties, including proton hopping efficiency, electroconductivity, stability of the zeta potential, and interactions at biological membrane interfaces, were critically analyzed to assess the viability of their therapeutic application through structured water supplementation. Experimental insights were carefully drawn from recent advancements in physical chemistry, electrochemistry, dielectric biology, and translational biomedical research.
The translational rationale developed through WP3 informed a subsequent targeted systematic review (WP4) to assess specifically the bioenergetic and metabolic evidence supporting EZC-Rich Water as an innovative nutritional adjunct in oncology.
2.4 WP4 – systematic review of EZC-Rich water supplementation as a metabolic and bioenergetic intervention in the tumor microenvironment
A second systematic review was registered in the PROSPERO database (CRD420251022205) to assess translational evidence supporting the use of structured water enriched with Eigen/Zundel complexes as a nutritional strategy for human health optimization. Indexed descriptors included “hydronium”, “structured water”, “hydrated proton”, “zeta potential”, and “bioenergetics”. Inclusion criteria prioritized peer-reviewed human or in vitro studies reporting effects on mitochondrial function, pH modulation, membrane permeability, and systemic redox balance.
WP5 was designed to systematically investigate the translational relevance of structured water supplementation enriched with hydrated proton complexes—specifically Eigen (H9O4+) and Zundel (H5O2+) structures—focusing on their capacity to modulate human bioenergetic, membrane electrochemical, and acid-base dynamics. A systematic literature review was conducted and registered under the PROSPERO database (CRD420251022205), following PRISMA guidelines (32).
The search strategy targeted descriptors such as “hydronium,” “Eigen complex,” “Zundel complex,” “hydrated proton,” “structured water,” “proton mobility,” and “zeta potential,” across PubMed, Scopus, and LILACS databases. Studies were included if they reported effects on pH modulation, mitochondrial function, membrane permeability, electrosmotic dynamics, or systemic bioenergetic regulation. Only peer-reviewed articles in English published after 2004 were considered.
From an initial pool of 139 identified records, 42 studies were excluded after title and abstract screening. Of the 97 articles subjected to full-text review, all were retained for final qualitative synthesis. The detailed literature selection process for this systematic review, conducted according to PRISMA guidelines, is illustrated in Figure 3.
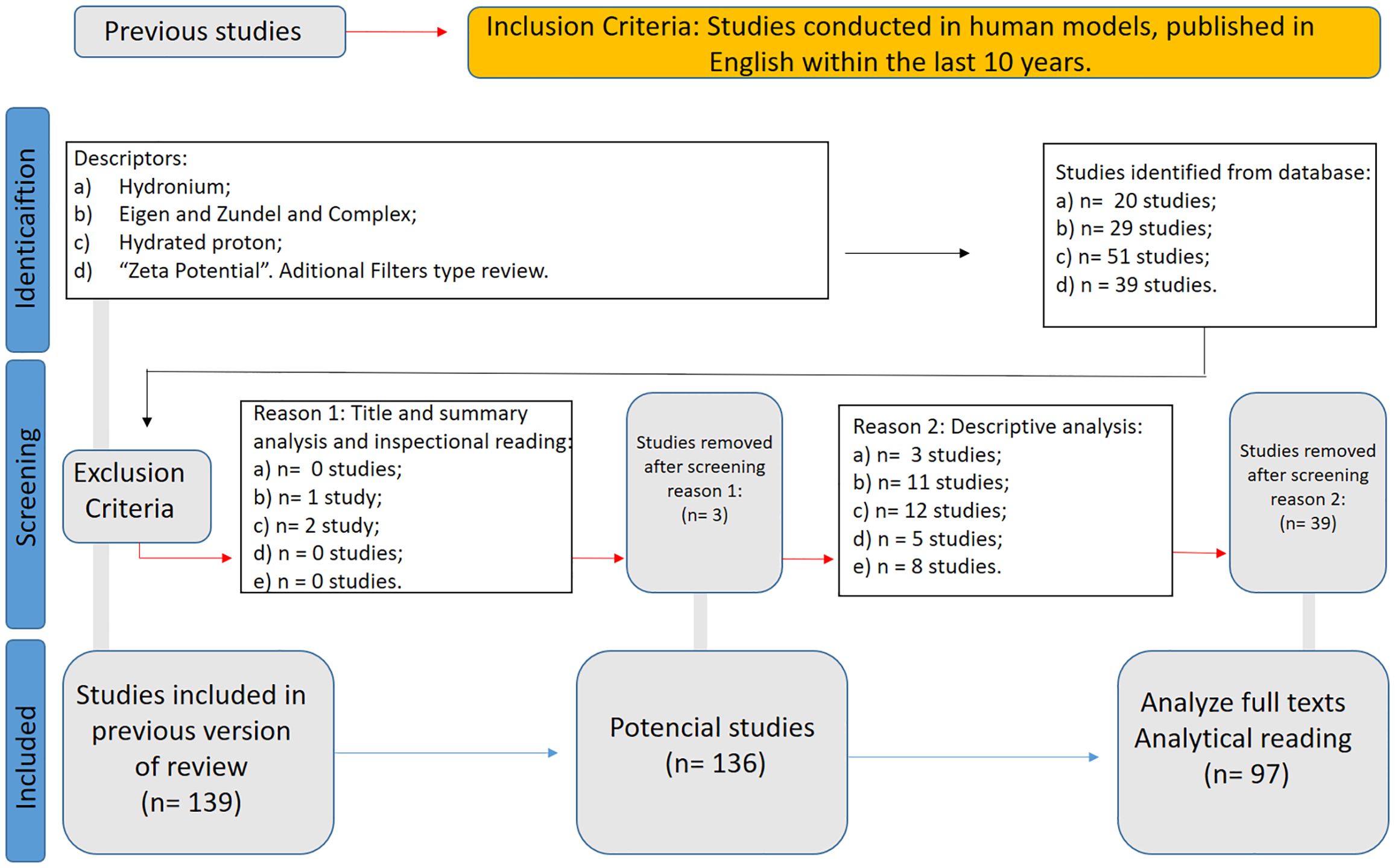
Figure 3. WP4 PRISMA flow diagram.
Finally, the comprehensive data and insights obtained from WP2 to WP4 were synthesized and critically evaluated in WP5, establishing a robust translational foundation for future experimental validation and publication.
2.5 WP5 – translational integration, clinical feasibility assessment, and strategic framework validation
This final stage synthesized and integrated findings from all previous WPs, employing structured methodological frameworks including SMART (Specific, Measurable, Achievable, Relevant, Time-bound) (33) and FINER (Feasible, Interesting, Novel, Ethical, Relevant) criteria (34). The therapeutic and translational potential of EZC-Rich Water was systematically assessed as a hydrated proton supplementation strategy for adjunctive cancer treatment, evaluating hypothesis plausibility, potential routes of administration, biomarker targets (e.g., zeta potential modulation, mitochondrial efficiency, redox homeostasis), and safety considerations. Concrete translational objectives, measurable biochemical and physiological outcomes, and the technological achievability of proposed interventions were critically analyzed. Relevance to current unmet needs in nutritional and metabolic oncology was highlighted, as was the novelty and ethical viability of the proposed supplementation strategy. Finally, this WP provided clear recommendations for future experimental validation (in vitro, in vivo, clinical trials) and structured a robust scientific rationale suitable for peer-reviewed publications.
3 Results
The Work Breakdown Structure (WBS) methodology (30, 31) allowed for the progressive consolidation of translational evidence across six interconnected Work Packages (WP2 to WP5). This structure enabled the integration of findings from oncology, bioenergetics, and water chemistry, guiding the formulation of a coherent hypothesis: that supplementation with water enriched in hydrated proton complexes—specifically Eigen and Zundel structures—may serve as a strategic adjunct for modulating tumor microenvironment dynamics, particularly through pH gradient correction, mitochondrial optimization, and redox regulation.
Each WP generated distinct insights that collectively reinforce the biological plausibility and therapeutic potential of EZC-Rich Water in the context of cancer bioenergetics. The sections below detail the findings from each WP sequentially.
3.1 WP2 – mechanistic drivers of electrochemical and bioenergetic imbalance in the tumor microenvironment
The WP2 comprised a targeted systematic review designed to elucidate the primary physiological and molecular mechanisms underpinning acid-base dysregulation within the tumor microenvironment (TME). These studies collectively underscored the presence of a reversed pH gradient as a hallmark of cancerous tissues. Specifically, extracellular pH (pHe) typically ranged between 6.2 and 6.9, contrasting markedly with the alkaline intracellular pH (pHi) values observed between 7.12 and 7.65. This inversion contrasts sharply with physiological norms in healthy tissues, where intracellular pH ranges from 7.0 to 7.2, and extracellular pH from 7.3 to 7.4. Such pathological acidification of the extracellular milieu is closely associated with tumor invasiveness, apoptosis resistance, therapeutic failure, genomic instability, and immune evasion (1, 3, 4, 35–37).
Central mechanisms identified as driving this abnormal pH landscape include the upregulation of proton transporters such as H+-ATPases, carbonic anhydrase IX (CAIX) (9–11), and Na+/H+ exchangers (37–39), alongside metabolic adaptations exemplified by the Warburg effect (19, 20), promoting aerobic glycolysis and subsequent lactic acid production. Furthermore, conditions of hypoxia (15), inflammatory signaling cascades, and impaired mitochondrial oxidative phosphorylation (16–18) were consistently highlighted as upstream modulators reinforcing these disturbances and driving the reversal of the pH gradient (40).
Critically, heterogeneity in pH regulation emerged as a recurring theme, with numerous studies documenting pronounced inter- and intra-tumoral variability. This variability, characterized by distinct microregional pH profiles, was posited as contributing significantly to tumor aggressiveness and therapeutic resistance (40–42).
Based on this comprehensive synthesis, a structured set of Fundamental Points of View (FPV’s) was formalized to encapsulate the principal acid-base and bioenergetic abnormalities characteristic of the TME (Table 1). The final structure of the Fundamental Points of View (FPV’s) emerged from an iterative and integrative synthesis of recent systematic reviews, experimental studies, and high-impact position papers on the tumor microenvironment. Each FPV represents a critical and translationally actionable axis identified across multiple literature filters and thematic searches.
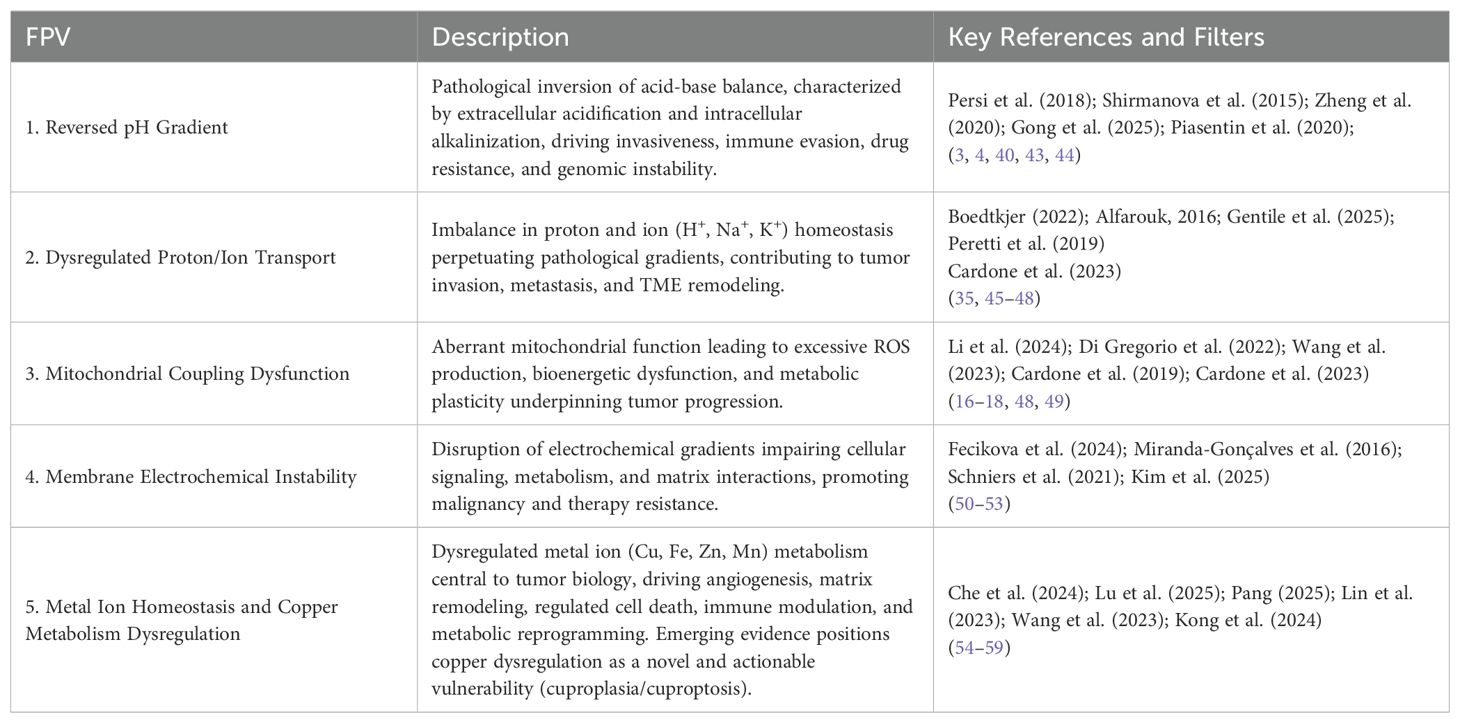
Table 1. Fundamental Points of View (FPV’s) associated with tumor microenvironment dysregulation.
Subsequent systematic searches (totaling 77 additional studies) robustly validated these FPV’s, particularly reinforcing the centrality of reversed pH gradients, mitochondrial dysfunction, dysregulated ion transport, and copper metabolism dysregulation. Although no additional FPV’s emerged, recent literature emphasized novel downstream effectors, such as the pH-sensitive receptor GPR4 (60) and the ion channel TRPC4 (61), further underscoring the complexity and potential therapeutic opportunities inherent within the acid-base and metabolic landscape of tumors.
Targeted analyses using specific descriptors further strengthened these conclusions. Studies filtered under “tumor microenvironment” and “ion transporters” consistently highlighted proton and ion transport dysregulation, supporting the FPV framework while elucidating complex multi-protein transport assemblies integral to cancer survival and therapy resistance. Literature focusing on carbonic anhydrase isoforms (9–11, 50) underscored their critical role as key enzymatic regulators driving extracellular acidification and intracellular alkalinization, thus confirming their central position within the identified FPVs.
Additionally, systematic examination of metalloproteases within the TME illuminated their essential roles in extracellular matrix remodeling, invasion, and immune modulation (62–65). Notably, emerging data suggests copper’s indirect but pivotal role in regulating matrix metalloproteinase (MMP) activity through redox modulation and oxidative stress pathways (66, 67), reinforcing copper metabolism dysregulation as an integrative and actionable FPV.
The refined FPVs not only clarify core physiological vulnerabilities within the TME but also provide a coherent translational framework. This enabled the structured alignment of each FPV to corresponding intervention strategies involving Eigen/Zundel Complexes-Rich Water (EZC-Rich Water), facilitating hypothesis-driven research into its therapeutic potential.
Recent cutting-edge evidence positions copper metabolism as a critical nexus in cancer bioenergetics (68–70), notably influencing mitochondrial respiration and regulated cell death pathways through modulation of enzymes like cytochrome c oxidase (CcO)—the terminal complex of the mitochondrial electron transport chain, responsible for facilitating the transfer of electrons to oxygen and contributing directly to the generation of proton motive force for ATP synthesis (71). Intriguingly, studies (72–74) indicate that CcO activity may be enhanced by local availability of protonated water clusters (Eigen/Zundel configurations), suggesting that targeted supplementation with EZC-Rich Water could exploit copper-dependent metabolic vulnerabilities to selectively induce regulated cell death mechanisms such as cuproptosis (58) and ferroptosis (75).
In addition to the identification of FPVs, WP2 also aimed to extract and validate Critical Success Factors (CSF’s) necessary for effective modulation of the tumor microenvironment (TME). Using a multicriteria decision-support framework adapted from Bana e Costa et al. (1999) (24), the selected FPVs were systematically confronted with biological and physicochemical variables identified in the literature, prioritizing those factors capable of influencing pH dynamics, mitochondrial function, and membrane stability.
A total of eight condensed CSF’s were validated, each representing a key biological mechanisms implicated in tumor progression and potential therapeutic modulation via proton dynamics. These CSF’s were categorized based on their functional alignment with the FPV’s previously identified, thus consolidating a structured translational framework linking the acid-base, mitochondrial, and electrochemical dysregulation observed in cancer to possible intervention targets. The validated CSF’s are summarized in Table 2.
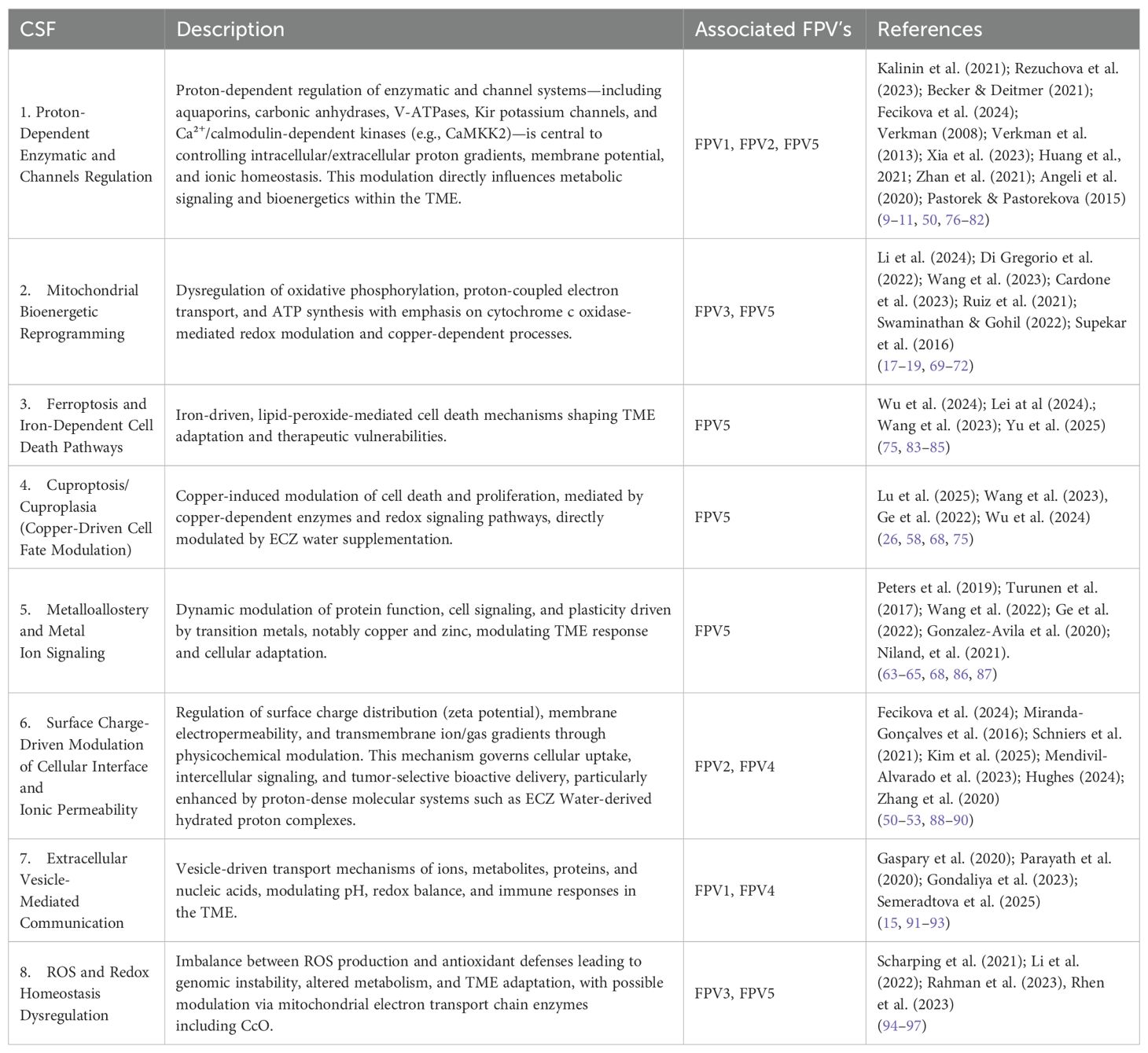
Table 2. Validated Critical Success Factors (CSF’s) and their association with Fundamental Points of View (FPV’s).
All the CSF’s identified and incorporated into our translational framework are grounded in a robust and up-to-date scientific literature. Most of these mechanisms—such as ion channel regulation, mitochondrial dysfunction, and redox imbalance—are widely recognized as consolidated drivers of tumor microenvironment (TME) dysregulation, with extensive experimental and clinical validation (98, 99).
It is important to note, however, that the inclusion of copper-driven metalloallostery as a CSF reflects a rapidly evolving understanding of cancer biology. Although this concept is relatively recent, it has gained considerable scientific traction as a key modulator of metabolic and signaling plasticity in cancer, as highlighted in recent reviews (26, 58, 68, 75). Its integration into our framework is intended to capture this emerging biological dimension, which is likely to shape future therapeutic strategies targeting TME vulnerabilities.
Likewise, the explicit integration of cytochrome c oxidase (CcO) activity within the CSF of mitochondrial bioenergetic reprogramming is supported by a growing body of evidence underscoring (72–74, 100) its pivotal role in mitochondrial respiration (71), cell death regulation (101, 102), and copper metabolism (68). CcO thus serves both as a mechanistic link and a potential therapeutic target within the TME.
In summary, WP2 provided a robust and integrative characterization of acid-base dysregulation, mitochondrial bioenergetic dysfunction, and metal homeostasis alterations within the tumor microenvironment. While most identified CSF’s are already well-established in cancer biology, this framework deliberately incorporates emerging yet scientifically substantiated mechanisms, thus positioning the research at the frontier of translational innovation. The systematic synthesis and formalization of FPVs and CSFs presented here offer a comprehensive foundation for assessing the therapeutic potential of EZC-Rich Water supplementation, clearly identifying actionable physiological targets for next-generation oncology interventions.
3.2 WP3 – translational potential of hydrated proton complexes and structured water chemistry for tumor microenvironment modulation
Considering the critical vulnerabilities identified in WP2—namely reversed pH gradients (FPV1), dysregulated proton and ion transport (FPV2), mitochondrial coupling dysfunction (FPV3), membrane electrochemical instability (FPV4), and alterations in metal ion homeostasis, particularly copper metabolism (FPV5)—WP3 was designed to explore physicochemical strategies capable of selectively addressing these targets through hydrated proton complexes. Hydrated proton complexes, specifically Eigen (H9O4+) and Zundel (H5O2+) clusters, emerged as promising translational candidates due to their distinctive capacity to modulate localized proton dynamics, stabilize membrane interactions, and maintain electrochemical balance without inducing systemic acidification or electrolyte disturbances (25, 28, 29). Therefore, WP3 aimed to systematically investigate the structural, biophysical, and translational properties of these proton complexes, emphasizing their unique solvation structures, enhanced proton mobility, and controlled interactions at biological membrane interfaces, in contrast to conventional diluted acid solutions containing free hydrogen ions (H+). Unlike conventional acid-based approaches, these protonated water clusters exert influence at the intersection of biophysical integrity, electrochemical signaling, and mitochondrial metabolism—thereby enabling a multidimensional modulation of tumor physiology.
Scientific literature indicates that proton conduction through water occurs predominantly via the Grotthuss mechanism, where protons are rapidly transferred along hydrogen-bonded water networks through a series of concerted molecular rearrangements (26, 27). Eigen and Zundel complexes are transient intermediates of this mechanism, facilitating highly mobile and directionally coordinated proton transfer, crucial for maintaining localized pH gradients in biological systems.
The translational relevance of hydrated proton complexes is rooted in three key properties. First, the enhanced proton mobility enabled by structured hydration promotes superior charge transport compared to free protons, allowing effective modulation of localized pH gradients without inducing abrupt systemic acidification (25, 29). Second, hydrated proton clusters exhibit high membrane interface stability, favorably interacting with biological membranes and modulating electrochemical gradients essential for mitochondrial bioenergetics and cellular signaling (103, 104). Third, their metastable organization reduces cytotoxic potential by minimizing non-specific acidification events and preserving electrolyte homeostasis, an important safety advantage for translational applications (28).
Recent advances have highlighted the translational impact of hydrated proton complexes, particularly in the context of copper metabolism within tumor cells. Copper acts as a vital cofactor for cytochrome c oxidase (CcO)—the terminal enzyme of the mitochondrial respiratory chain—which orchestrates ATP synthesis, redox homeostasis, and cell fate decisions. In tumors, where copper dysregulation and mitochondrial suppression are often observed, CcO activity is highly dependent on local proton availability, particularly through Eigen and Zundel cluster dynamics at its catalytic interface (72).
Thus, supplementation with EZC-Rich Water may synergize with the copper-dependent machinery of cancer cell mitochondria, restoring or enhancing CcO function and potentially tipping the balance towards increased mitochondrial activity, redox regulation, and induction of autophagy or cell death in metabolically vulnerable tumors. This convergence of proton transport, copper metabolism, and mitochondrial regulation provides a strong rationale for targeting the TME with strategies that bridge physical chemistry and metabolic oncology.
These findings provide a robust physicochemical basis for proposing Eigen/Zundel Complexes-Rich Water (EZC-Rich Water) as a clinically adaptable supplementation strategy aimed at reprogramming the tumor microenvironment. Instead of acting through systemic pH depression, EZC-Rich Water is hypothesized to promote localized proton enrichment at membrane interfaces and interstitial spaces, potentially stabilizing mitochondrial proton motive force, optimizing ATP synthesis, and restoring peritumoral electrochemical balance. A synthesis of these translational insights is presented in Table 3.
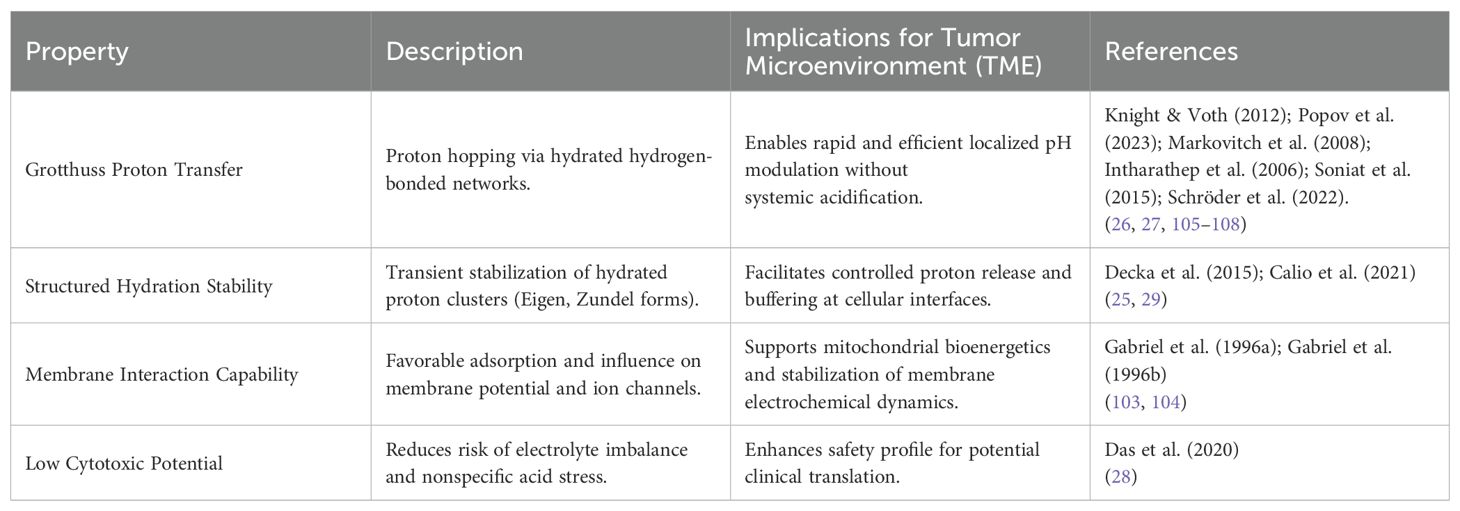
Table 3. Translational properties of Eigen/Zundel complexes relevant to tumor microenvironment modulation.
This approach is conceptually aligned with recent advances in physical oncology and nanomedicine, which recognize the tumor microenvironment’s electrochemical and mechanical aberrancies as therapeutic entry points. As reviewed by Nicolas-Boluda et al. (2018) (109), nanomaterials capable of modulating surface charge, tissue stiffness, and interstitial conductivity are increasingly explored to overcome drug resistance and enhance therapeutic penetration — a framework within which ECZ Water, rich in protonated water clusters, may function as a tunable, bioactive nano-agent.
Collectively, these findings establish a compelling physicochemical and translational rationale for the use of EZC-Rich Water in oncology. By enabling localized proton enrichment without systemic acid-base disruption, this strategy aims to restore peritumoral electrochemical stability, re-enable mitochondrial respiratory activity, and selectively sensitize metabolically vulnerable tumors to therapeutic modulation. Building on the physicochemical rationale presented above, WP4 systematically investigates the evidence supporting the translational use of EZC-Rich Water, with particular attention to its biomedical differentiation from conventional acidic solutions.
3.3 WP4 – evidence synthesis on EZC-Rich water supplementation
In continuity with the foundational FPVs mapped in WP2, WP4 reexamines their relationship with hydrated proton complexes under a translational lens and WP4 was initiated to systematically investigate the translational relevance of structured water supplementation enriched with hydrated proton complexes, specifically Eigen (H9O4+) and Zundel (H5O2+) forms. A systematic literature review (SLR) was registered under PROSPERO (CRD420251022205), following PRISMA guidelines (32), to address the central research question: Could EZC-Rich Water exhibit therapeutic potential, particularly in modulating the cellular microenvironment and inducing transient systemic pH variations?
The initial step involved defining the specific physicochemical properties of EZC-Rich Water and establishing a clear differentiation from conventional acidified water, forming the basis for subsequent multi-criteria analyses. Table 4 presents a detailed comparative analysis.
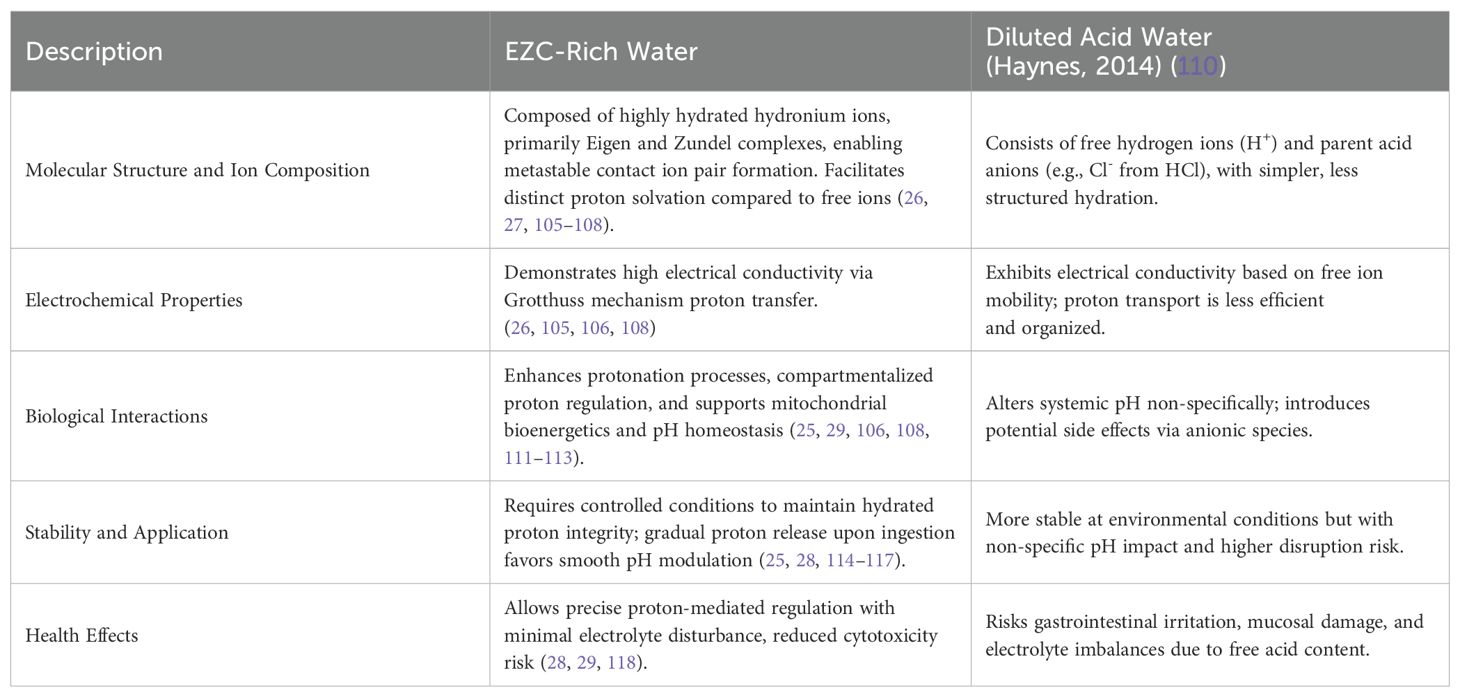
Table 4. Comparison between Eigen/Zundel complex-rich water and diluted acid water.
EZC-Rich Water can be synthesized through electrochemical methods. According to Alkhadra et al. (2022) (119), water electrolysis using direct current (DC) across ion-exchange membranes such as Nafion generates both hydronium (H3O+) and hydroxide (OH-) ions. The highly dynamic proton mobility via the Grotthuss mechanism poses challenges for stabilization, requiring immediate consumption after generation or low-temperature synthesis techniques to preserve the structured hydrated proton integrity (Markovitch et al., 2008 (105); Knight & Voth, 2012 (26); Schröder et al., 2022 (108)).
Alternative strategies, such as low-temperature chemical synthesis (Wurzbarger, 1986) (114), aim to extend the lifetime of hydrated proton complexes, potentially improving the translational viability of EZC-Rich Water for biomedical applications. Moreover, administration routes—including oral ingestion, transdermal absorption, or intravenous delivery—are under consideration to optimize proton gradient modulation for therapeutic effect without systemic acid-base disturbances.
To ensure conceptual coherence between the vulnerabilities mapped in the tumor microenvironment (FPVs identified in WP2) and the hypothesized modulatory actions of Eigen/Zundel Complexes-Rich Water, a comparative framework was developed to align each fundamental physiological target with its corresponding mechanism of intervention. This approach clarifies how the proposed supplementation may directly address each core axis of tumor dysregulation (see Table 5). By mapping one-to-one (or one-to-many) relationships, the framework underscores the translational logic of the strategy and offers a transparent rationale for the therapeutic potential of EZC-Rich Water in oncology.
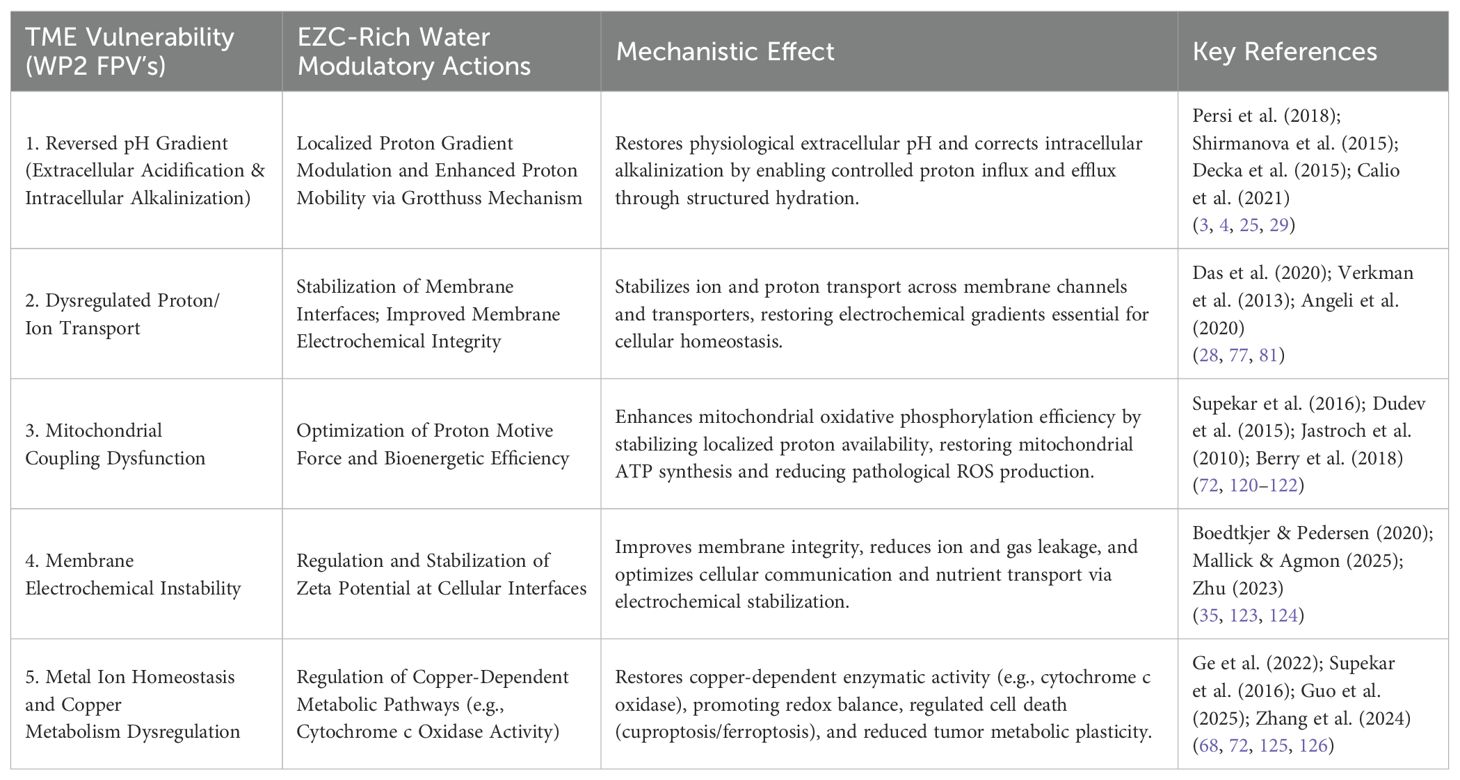
Table 5. Comparative mapping table: alignment of tumor microenvironment FPVs with modulatory actions of EZC-rich water.
To strengthen the translational foundation of the proposed intervention, WP4 reexamined the Critical Success Factors (CSF’s) identified in WP2 in light of the physicochemical properties of EZC-Rich Water. This comparative analysis demonstrates how hydrated proton complexes may interact with and modulate the core biological mechanisms represented by each CSF. Table 6 presents this functional alignment, highlighting the potential contributions of EZC-Rich Water across distinct dimensions of tumor microenvironment dysregulation.
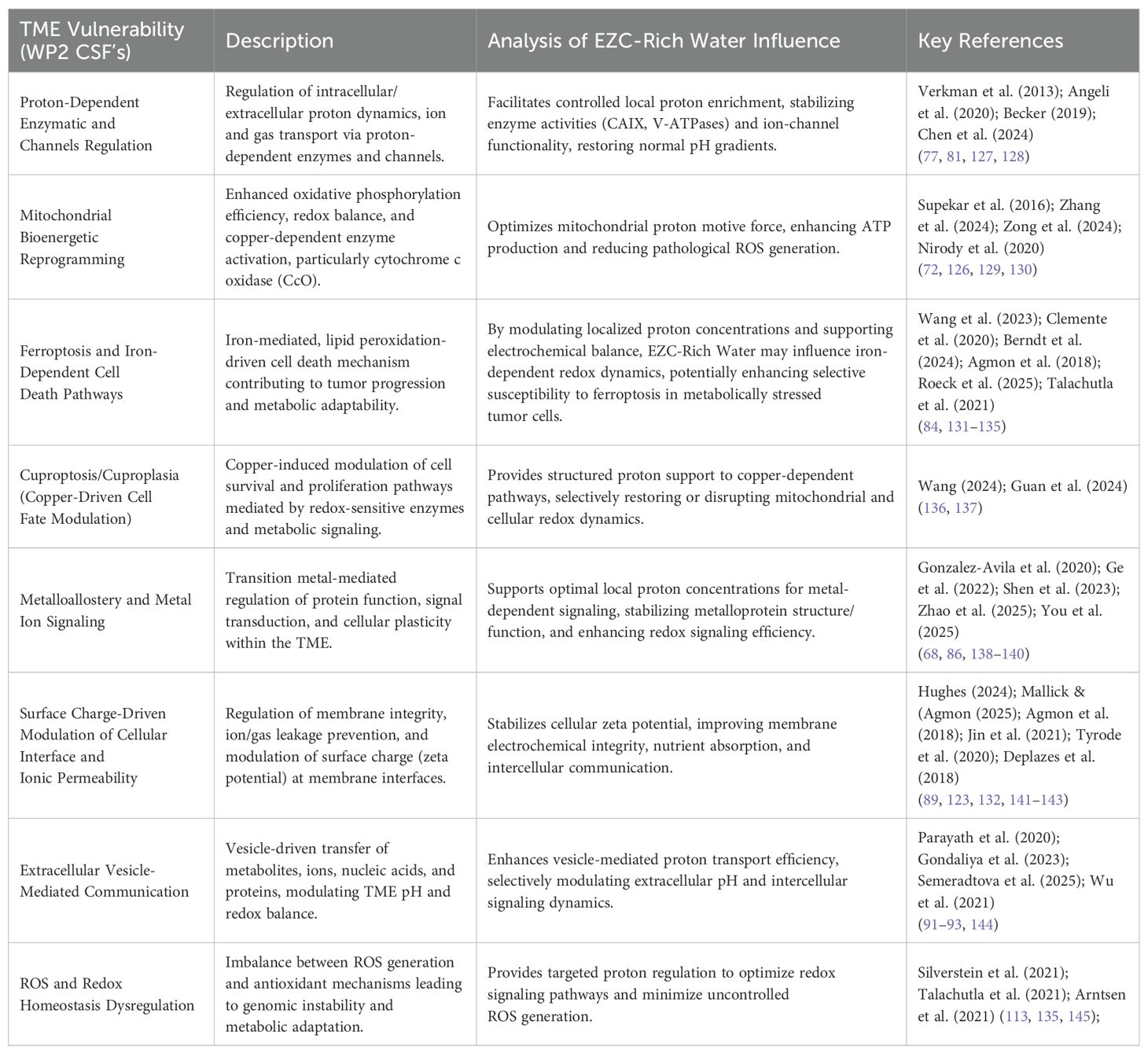
Table 6. Critical Success Factors (CSF’s) and potential contributions of EZC-rich water.
The refined alignment of FPVs and CSFs further highlights the innovative translational potential of EZC-Rich Water, establishing a clear and mechanistically sound pathway for subsequent preclinical and clinical validation studies in cancer bioenergetics. This integrative matrix demonstrates that EZC-Rich Water acts simultaneously on multiple physiological targets linked to cancer progression. By delivering hydrated proton complexes capable of modulating membrane dynamics, buffering pH gradients, and enhancing mitochondrial proton motive force, this supplementation strategy presents a mechanistically grounded and translationally feasible intervention.
While the refined alignment of FPVs and CSFs underscores the mechanistic rationale and translational feasibility of EZC-Rich Water, it is important to acknowledge that much of the current evidence supporting its potential derives from indirect experimental models, physicochemical simulations, and foundational studies in bioenergetics and interfacial proton dynamics. Direct validation of hydrated proton complexes in clinical or biomedical laboratory settings remains limited, reflecting the novelty of this approach rather than a lack of scientific plausibility. As such, the framework proposed here should be viewed as a theoretically robust and mechanistically informed starting point, warranting targeted experimental efforts to substantiate its therapeutic applications in cancer and beyond. Together with the conceptual synthesis illustrated in Figure 4, this framework positions EZC-Rich Water supplementation as a biophysical strategy with clinical relevance, potentially applicable in oncology and other bioenergetic dysfunction scenarios.
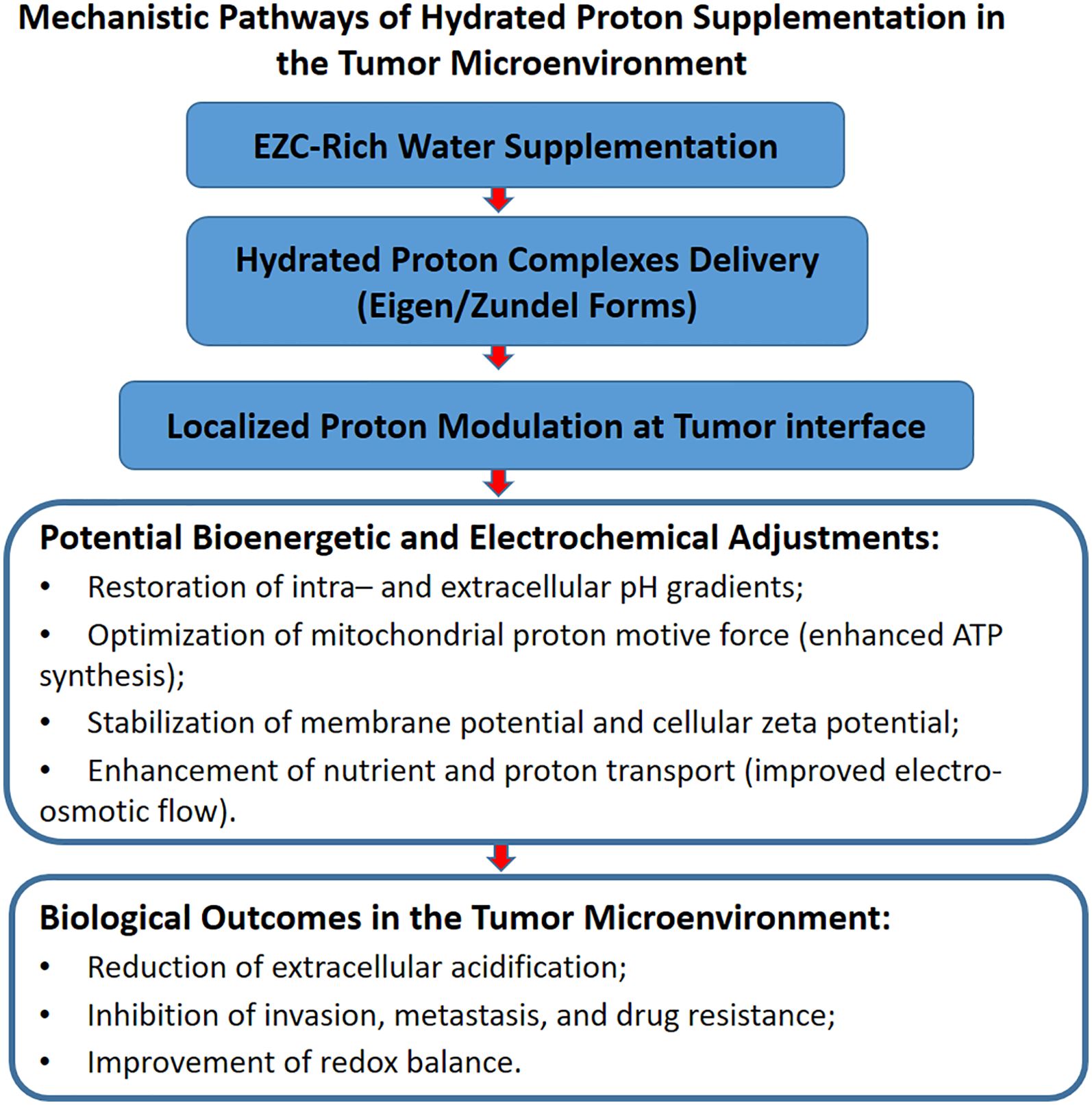
Figure 4. Structured supplementation with EZC-Rich Water facilitates targeted modulation of tumor microenvironment pH and bioenergetics, driving therapeutic outcomes.
Analysis of the selected studies revealed that EZC-Rich Water supplementation exerts measurable impacts on five key FPVs, reinforcing its translational potential. First, it promotes modulation of intra- and extracellular pH gradients, facilitating the reversion of pathological pH inversion commonly observed in tumor microenvironments. Second, it optimizes mitochondrial ATP synthesis by enhancing proton motive force and improving overall energy conversion efficiency. Third, it regulates cellular membrane permeability, favoring more efficient nutrient transport, ion flux regulation, and stabilization of electrochemical gradients. Fourth, it enhances electrosmotic flow, supporting proton and molecule transport dynamics within tissue microenvironments. Finally, EZC-Rich Water stabilizes cellular zeta potential, contributing to the maintenance of membrane integrity and the optimization of cellular communication processes.
In addition to functional evidence, chemical analysis of EZC-Rich Water demonstrated its physicochemical differentiation from conventional diluted acidic solutions. Unlike free hydrogen ion-containing waters, EZC-Rich Water retains a metastable, structured organization of hydrated proton clusters (Eigen and Zundel complexes), promoting selective and regulated proton interactions without introducing disruptive ionic loads (25, 29, 105).
Thus, WP4 provided a solid translational foundation, demonstrating that EZC-Rich Water possesses distinct physicochemical and biological properties capable of addressing key bioenergetic vulnerabilities within the tumor microenvironment. Collectively, these findings underscore EZC-Rich Water’s innovative translational potential as a targeted therapeutic adjunct, and clearly justify subsequent experimental validation—ranging from preclinical models to eventual clinical trials—to establish its efficacy and safety profile in oncological applications. Beyond its oncological relevance, the experimental insights gained in WP4 also laid the groundwork for advancing hydrated proton supplementation from a theoretical construct into a broader translational research pathway, opening new perspectives for adjunctive interventions in metabolic disorders and mitochondrial dysfunction syndromes. These insights consolidate the translational foundation for future experimental validation and clinical exploration of EZC-Rich Water.
3.4 WP5 – integration of findings and formalization of translational hypothesis
Building on the mechanistic coherence established through WP2 to WP4, WP5 formalizes a structured translational hypothesis: that supplementation with Eigen/Zundel Complexes-Rich Water (EZC-Rich Water) may strategically modulate critical vulnerabilities within the tumor microenvironment (TME) by correcting localized pH gradients, enhancing mitochondrial bioenergetics, and stabilizing membrane electrochemical dynamics. Conceptually, this approach integrates recent advances in structured water chemistry, dielectric biology, and proton-mediated redox signaling, distinctly differing from conventional methods of systemic acidification or generalized antioxidant supplementation. Instead, EZC-Rich Water delivers metastable hydrated proton complexes (Eigen and Zundel forms), acting primarily through localized proton enrichment at cellular interfaces and mitochondrial membranes.
This translational proposition specifically addresses the FPVs previously identified as core vulnerabilities in the TME—reversed pH gradients, dysregulated ion transport, mitochondrial coupling dysfunction, membrane electrochemical instability, and metal ion homeostasis dysregulation. By providing a structured and controlled proton delivery mechanism, EZC-Rich Water may not only normalize aberrant extracellular acidification and intracellular alkalinization but also optimize mitochondrial proton gradients essential for ATP synthesis and redox balance. Moreover, this intervention could stabilize cellular membranes through modulation of zeta potential, further enhancing nutrient transport, intercellular communication, and overall metabolic resilience.
A critical component of this integrative hypothesis is the intersection between proton dynamics and copper metabolism. EZC-Rich Water supplementation may uniquely facilitate the optimal function of copper-dependent mitochondrial enzymes such as cytochrome c oxidase (CcO). By stabilizing localized proton availability at catalytic sites, EZC-Rich Water may enhance CcO activity, thus potentially triggering controlled mitochondrial reprogramming and regulated cell death pathways, including cuproptosis, autophagy, or ferroptosis (Figure 4).
To rigorously evaluate the translational feasibility and clinical viability of this proposal, we applied structured methodological criteria frameworks—SMART (Specific, Measurable, Achievable, Relevant, Time-bound) (33) and FINER (Feasible, Interesting, Novel, Ethical, Relevant) (34). This critical evaluation confirmed that EZC-Rich Water supplementation meets key parameters essential for advancing translational oncology research. Table 7 summarizes this structured feasibility assessment, clearly outlining how the proposed therapeutic strategy satisfies essential scientific, practical, and ethical standards.
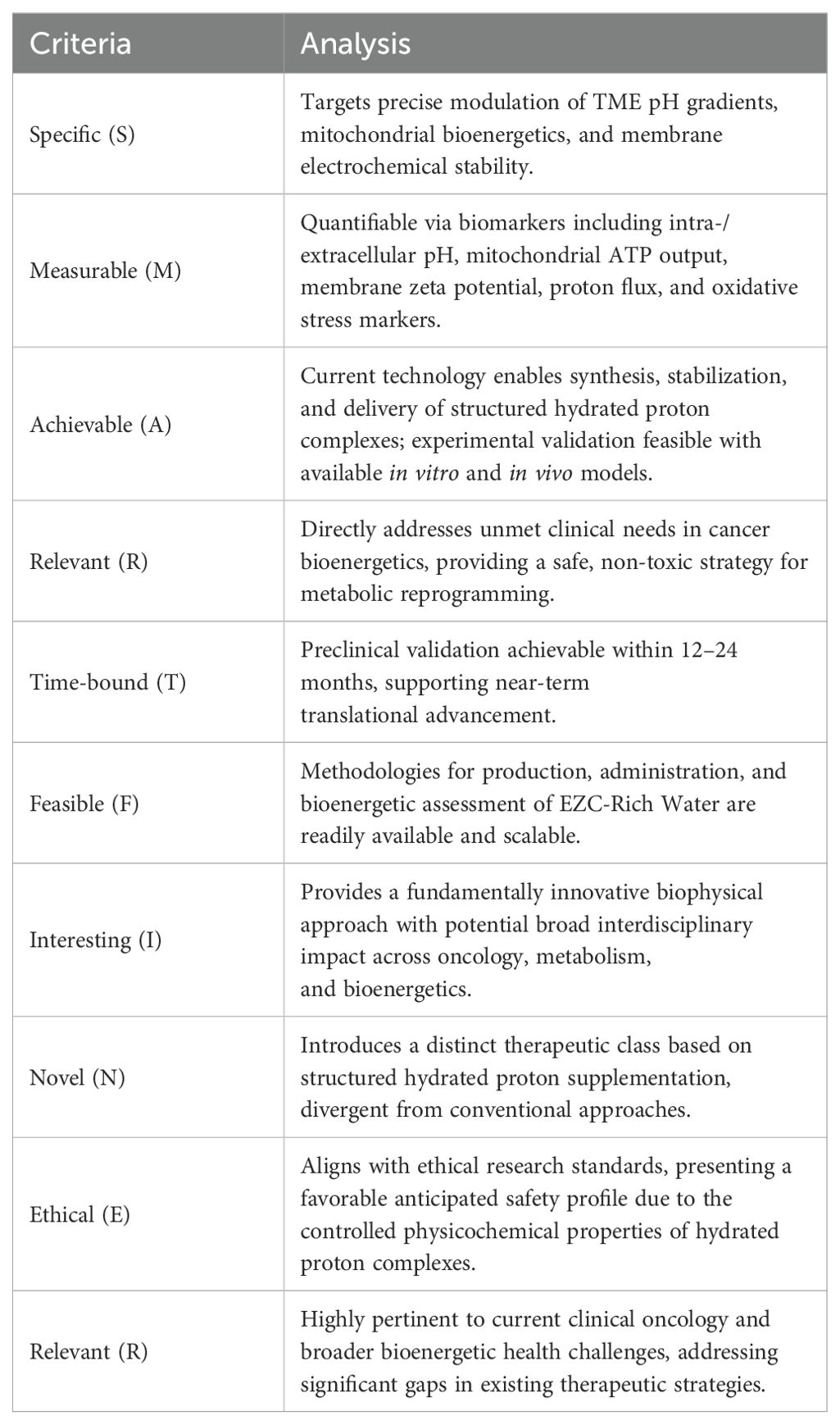
Table 7. Outcome of SMART and FINER evaluation for EZC-rich water supplementation hypothesis.
Collectively, WP5 affirms that the supplementation with EZC-Rich Water possesses substantial scientific credibility and practical feasibility. By clearly delineating the bioenergetic and physicochemical rationale, as well as the strategic alignment between Fundamental Points of View (FPV’s) and validated Critical Success Factors (CSF’s), this synthesis lays a solid translational foundation. Therefore, EZC-Rich Water represents a scientifically robust, ethically sound, and clinically relevant adjunctive intervention strategy, warranting targeted preclinical and clinical validation efforts in oncology and broader bioenergetic optimization contexts.
By integrating these mechanistic insights into a coherent translational framework, this study robustly supports the hypothesis that structured hydrated proton supplementation can act as a low-risk, multi-targeted therapeutic strategy, addressing multiple bioenergetic and metabolic vulnerabilities simultaneously. The comprehensive synthesis provided here lays a firm foundation for further exploration of clinical implications, translational relevance, and innovative therapeutic applications, as explored in the following discussion.
4 Discussion
This study presents EZC-Rich Water supplementation as a translational bioenergetic intervention grounded in the regulation of proton gradients and the restoration of acid–base and metabolic homeostasis within the tumor microenvironment, particularly targeting the Warburg effect and mitochondrial dysfunction. Although still in the early stages of clinical exploration, the mechanistic rationale and experimental integration proposed here provide a coherent foundation for targeted preclinical exploration. By bridging quantum-level proton dynamics with systemic physiological regulation, this approach introduces a new class of bioenergetic strategies that extend beyond traditional pharmacological or metabolic interventions. Quantum-level proton transport dynamics, referring to the coherent, rapid proton transfers via hydrogen-bonded water networks described by the Grotthuss mechanism, have emerging translational implications for biomedical interventions (27, 145–148). Recent studies reinforce the presence of Grotthuss-type proton transfer within biological systems, including active enzyme sites (Zlobin et al., 2024 (147)) and voltage-gated proton channels (Morgan & DeCoursey, 2003 (148)), suggesting that structured hydrated proton movement is not limited to bulk water but also plays a role in bioenergetic and redox regulation. These insights provide a strong molecular rationale for exploring translational applications of proton-rich supplementation strategies, such as EZC-Rich Water.
Hydrated proton complexes—Eigen and Zundel structures—possess a unique capacity for facilitating localized proton mobility via the Grotthuss mechanism. Their application in aqueous supplementation form enables compartmentalized acid-base modulation without systemic acidosis, preserving intracellular signaling fidelity, mitochondrial efficiency, and membrane integrity. These structured protons, when delivered via EZC-Rich Water, may act on key bioenergetic axes, modulating mitochondrial oxidative phosphorylation, supporting ATP synthesis, and reducing reactive oxygen species generation (Lee & Nandab, 2020 (149); Aklima et al., 2021 (150); Bertholet et al., 2022 (151)).
The evidence consolidated across the Work Packages suggests that transient and localized proton adjustments, as promoted by EZC-Rich Water, can positively influence zeta potential, membrane fluidity, redox resilience, and electrosmotic flow—all of which are severely impaired in cancerous tissues. These effects are mechanistically plausible and conceptually aligned with previous theoretical explorations regarding carbon dioxide’s hormetic potential in the tumor microenvironment. Previous research conducted by the same group (Gaspary et al., 2024 (15)) have demonstrated that CO2’s conversion to carbonic acid may assist in buffering extracellular acidity, potentially enhancing immunotherapeutic responses and modulating key enzymatic systems such as carbonic anhydrase IX (Ronca et al., 2024 (152)), heme oxygenase (Surh et al., 2020 (153)), and matrix metalloproteinases (Niland et al., 2021 (87)).
By integrating these perspectives, it becomes evident that modulating local proton dynamics—whether via CO2 pathways or via structured water supplementation—offers a convergent route to reprogram the biochemical landscape of malignant tissues. Both approaches—EZC-Rich Water and controlled CO2 exposure—appear capable of influencing critical elements of the tumor ecosystem, including pH regulation, mitochondrial metabolism, immune surveillance, and cellular communication.
The distinction between these two modalities lies primarily in delivery and control. EZC-Rich Water offers a targeted, non-gaseous, and potentially safer method of introducing hydrated protons in a metastable and bioavailable form. Meanwhile, CO2, though physiologically potent, poses challenges in delivery standardization and long-term safety, particularly under chronic or high-concentration exposure conditions (15). Nonetheless, the theoretical convergence highlights the growing recognition of proton gradients as dynamic modulators of cellular behavior and disease progression.
Despite these promising mechanistic foundations, it must be acknowledged that these hypotheses remain at an early experimental stage. Observational case reports from the AuBento Institute suggest preliminary clinical benefits following CO2-modulation protocols, such as PSA reduction and tumor regression (15); however, rigorous, controlled clinical trials are necessary to substantiate both mechanisms (Chesbrough, 2003 (22); Flessa & Huebner, 2021 (154). Additionally, the absence of adequate in vitro models capable of replicating systemic adaptive processes highlights the complexity inherent in studying such bioenergetic interventions.
From a translational standpoint, one of the most pressing challenges is the refinement of dosing protocols, definition of optimal proton concentrations, and assessment of long-term systemic impacts. Whether administered orally, transdermally, or intravenously, EZC-Rich Water demands comprehensive profiling of bioavailability, pharmacokinetics, systemic distribution, and potential toxicity under standardized preclinical conditions. These evaluations must also consider compatibility with existing therapies and establish stable, replicable dosage regimens through longitudinal studies. Similarly, CO2-based interventions must delineate precise thresholds between beneficial hormetic exposure and harmful systemic acidosis, while identifying reliable biomarkers of therapeutic response and physiological resilience (15). Despite the robust theoretical rationale developed thus far, clinical applicability remains speculative in the absence of dose standardization, physicochemical stability assessments, and safety data in human models. These are not minor concerns, but critical gatekeepers for advancing toward empirical validation. As such, rigorously designed experimental studies are imperative to move beyond exploratory plausibility and establish a solid foundation for clinical translation.
The unifying principle behind both strategies is the recognition that precise modulation of proton gradients—rather than direct cellular cytotoxicity—may represent a fundamentally new paradigm in oncology and bioenergetic medicine. Instead of targeting the tumor directly, the approach focuses on reprogramming the biological terrain to create conditions unfavorable for malignancy and conducive to systemic resilience.
Emerging evidence strongly supports the pivotal role of proton fluxes in mitochondrial bioenergetics, oxidative phosphorylation efficiency, and redox homeostasis. Hydronium ions, inherently present in aqueous environments, play critical roles in intracellular signaling and energy metabolism. Localized adjustments in proton distribution, as hypothesized with EZC-Rich Water supplementation, could facilitate ion transport across membranes, stabilize mitochondrial function, and optimize redox balance, offering a finely tuned mechanism to enhance cellular adaptation and resilience (Lee & Nandab, 2020; Aklima et al., 2021; Bertholet et al., 2022) (149–151).
In support of this hypothesis, experimental findings indicate that hydronium ions can modulate phospholipid bilayer properties, impacting membrane conductivity, ion flux, and signal transduction (Deplazes et al., 2018) (143). These phenomena reinforce the concept that structured hydration-mediated proton delivery could exert tangible bioenergetic benefits at the membrane interface level.
Although WP5 provided a structured feasibility analysis of the proposed intervention using SMART and FINER frameworks, it also served to consolidate the key mechanistic dimensions supporting the clinical plausibility of EZC-Rich Water supplementation. Based on the evidence synthesized throughout the WPs, Table 8 highlights how transient proton gradient modulation may influence key physiological systems across local, cellular, and systemic levels.
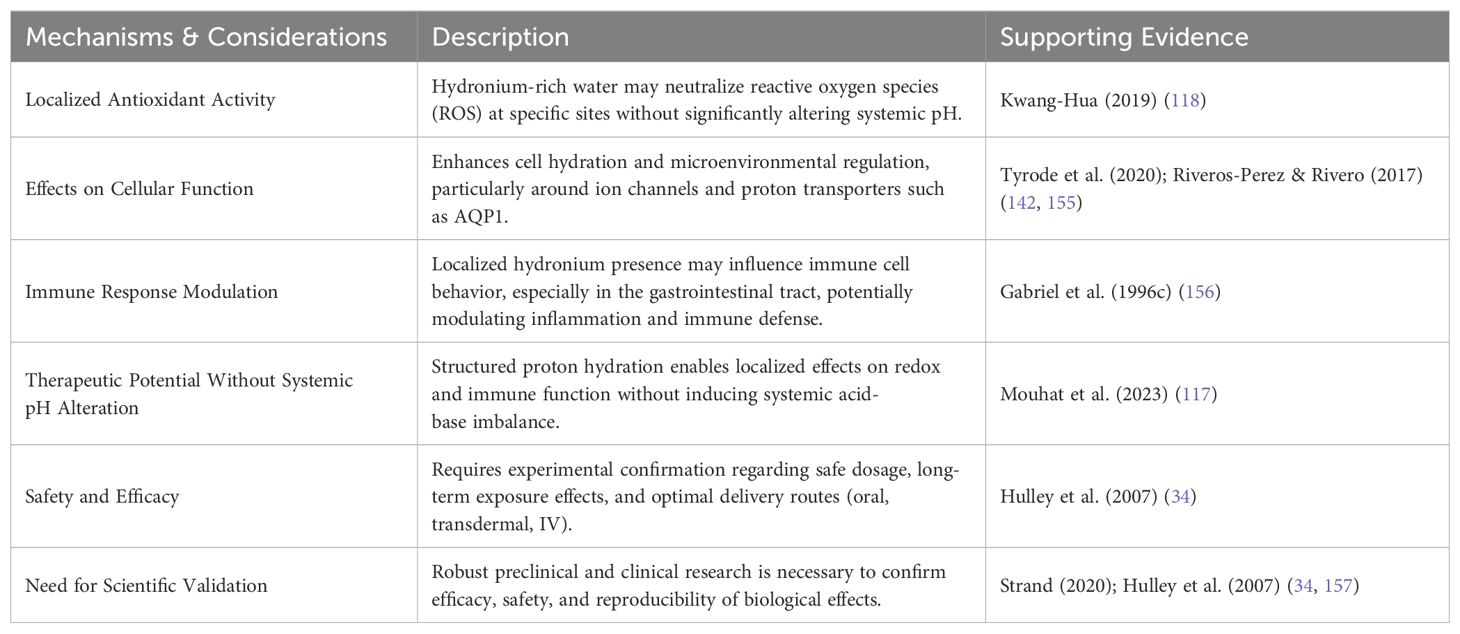
Table 8. Local, cellular, and therapeutic considerations of EZC-rich water supplementation.
Taken together, the findings from WP1 to WP5 construct a robust translational framework supporting the therapeutic potential of EZC-Rich Water supplementation. By targeting the central dysfunctions of the tumor microenvironment — pH inversion, mitochondrial inefficiency, electrochemical instability — this approach introduces a novel, biophysically grounded, and biologically coherent adjunctive strategy. The integration of hydrated proton complexes within a clinically adaptable format, combined with their low cytotoxicity and precise biochemical targeting, advances the field of bioenergetic modulation in oncology.
These mechanisms are not exclusive to tumor regulation but probably extend to general homeostatic processes such as membrane stability, redox signaling, immune responses, and targeted pH buffering. This reinforces the hypothesis that hydrated proton supplementation may act not merely as an acidifying agent, but as a bioenergetic modulator with multidimensional therapeutic potential. As illustrated in Figure 5, the transient and localized adjustment of proton gradients — the mechanistic basis of the proposed intervention — may influence multiple physiological systems simultaneously. These include aquaporin channel regulation, cytochrome c oxidase activity, cellular immune responses, central nervous system homeostasis, and efficient proton transfer. This multi-domain responsiveness reinforces the broader systemic relevance of the intervention beyond cancer alone, with implications in neurobiology, immunology, and metabolic disease.
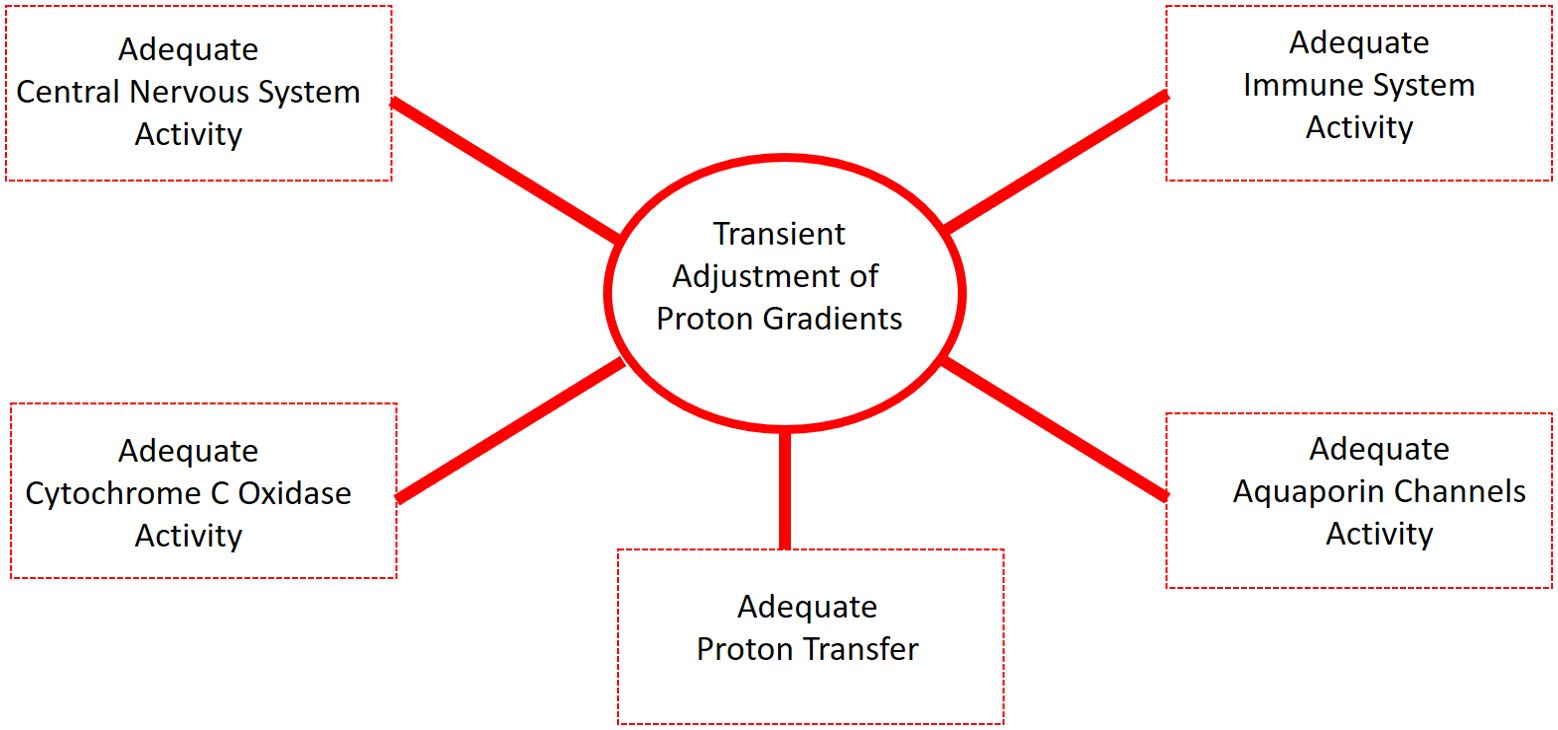
Figure 5. Conceptual representation of the systemic physiological outcomes mediated by the transient adjustment of proton gradients, central to the biological action of EZC-Rich Water supplementation.
Interestingly, the mechanistic rationale behind EZC-Rich Water supplementation converges with other non-pharmacological strategies such as controlled CO2 exposure. Both approaches act by subtly influencing interfacial proton dynamics—particularly at membrane boundaries—affecting localized electrochemical conditions such as charge polarization and redox signaling, without inducing systemic acid-base disruption. This convergence reinforces the symbolic unity of the intervention: metabolic restoration through precise interfacial modulation rather than global biochemical interference, offering a conceptually elegant and minimally invasive approach to reprogram tumor metabolism.
The hypothesis built throughout the structured research stages now calls for a critical examination in light of current scientific literature. In the following section, we explore the implications, strengths, limitations, and future directions for hydrated proton supplementation — contextualizing its bioenergetic logic, its translational novelty, and its potential clinical applications in cancer therapy and beyond.
By consolidating these mechanisms into an integrative translational summary, this multifaceted and interdisciplinary research provides robust support for the hypothesis that hydrated proton supplementation could serve as a low-risk, multi-targeted strategy—particularly effective in reversing the Warburg effect, restoring mitochondrial function, and optimizing broader biological processes in both pathological and physiological contexts. These insights establish a solid foundation for broader reflections on implications, clinical relevance, and future research directions explored throughout the discussion. Moreover, while the current theoretical framework robustly outlines the translational potential of EZC-Rich Water supplementation in oncology, additional validation across multiple experimental models will be essential to ensure reproducibility and generalizability of the findings. Future studies should specifically consider randomized controlled trials designed to evaluate key bioenergetic and clinical biomarkers outlined in this research, such as mitochondrial ATP output, proton flux dynamics, membrane electrochemical stability, and redox homeostasis. Furthermore, extending the application of EZC-Rich Water supplementation beyond oncology to other metabolic, inflammatory, and neurodegenerative diseases could provide broader validation of its systemic therapeutic potential, reinforcing its interdisciplinary significance and enhancing its translational impact across diverse clinical domains.
These results also support the emergence of a novel therapeutic paradigm—one that integrates biophysical insight, metabolic precision, and physiological modulation. EZC-Rich Water may be representative of a new class of adjunctive strategies that operate at the intersection of structured hydration chemistry, mitochondrial bioenergetics, and tumor interface dynamics. As such, this supplementation model may transcend its immediate oncological scope and signal broader applications across regenerative, metabolic, and redox-centered medicine. Although this study is framed within the oncological landscape, the underlying principles of hydrated proton dynamics—particularly in mitochondrial function and membrane electrochemical stability—suggest translational potential in metabolic disorders, chronic inflammation, and neurodegenerative syndromes. These applications should be explored in parallel to preclinical validation in cancer models.
Ultimately, while the theoretical framework is well-grounded, translational credibility will only be achieved through rigorous experimental validation across preclinical and clinical models—ensuring reproducibility, safety, and real-world efficacy.
5 Conclusions
This study introduces Eigen/Zundel Complexes-Rich Water (EZC-Rich Water) as a mechanistically grounded and translationally feasible approach for tumor microenvironment (TME) modulation. By addressing proton gradient dysregulation—a central hallmark of cancer bioenergetics—this supplementation strategy offers a novel route to reprogram pathological pH inversion, mitochondrial dysfunction, and membrane electrochemical instability.
Unlike conventional acidified solutions, EZC-Rich Water leverages metastable hydrated proton clusters—Eigen (H9O4+) and Zundel (H5O2+)—to enhance proton mobility and interface stability without systemic acidosis. The integrative mapping between Fundamental Physiological Vulnerabilities (FPVs) and Critical Success Factors (CSFs) strengthens its therapeutic plausibility, particularly in contexts marked by the Warburg effect, ion transport imbalance, and copper-dependent mitochondrial suppression.
The findings also highlight mechanistic convergence with other bioenergetic interventions—such as controlled CO2 exposure—reinforcing the emerging importance of interfacial proton dynamics in translational oncology and metabolic research.
Despite its conceptual rigor, the clinical viability of EZC-Rich Water remains dependent on empirical validation. Key priorities for future research include: standardization of proton concentrations, stability profiling, pharmacokinetic evaluation, and safety assessment through preclinical and human studies. Randomized controlled trials should be designed to monitor specific biomarkers of mitochondrial function, redox modulation, and peritumoral pH regulation, providing a robust foundation for regulatory advancement.
In summary, EZC-Rich Water supplementation emerges as a promising adjunctive strategy within a new class of biophysically informed therapeutics. While current evidence supports its theoretical and mechanistic potential, its definitive translational value will depend on sustained interdisciplinary efforts and rigorous experimental corroboration. If validated, this intervention may contribute meaningfully to the evolving landscape of personalized and metabolically targeted medicine.
Data availability statement
The original contributions presented in the study are included in the article. Further inquiries can be directed to the corresponding author.
Author contributions
ALE: Conceptualization, Writing – original draft, Writing – review & editing, Formal analysis, Methodology, Investigation. LFDL: Writing – original draft, Writing – review & editing, Formal analysis, Methodology, Investigation, Supervision. EGL: Methodology, Formal analysis, Writing – review & editing. IPDF: Methodology, Formal analysis, Writing – review & editing. CRF: Methodology, Formal analysis, Writing – review & editing. JFPG: Conceptualization, Writing – original draft, Writing – review & editing, Formal analysis, Methodology, Investigation, Supervision, Visualization.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Conflict of interest
Author ALE is the co-founder of ElastoCrete, LLC, a company specializing in decorative and structural concrete systems, which is entirely unrelated to healthcare or to the subject matter of this manuscript.
The remaining authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. In preparing this manuscript, the authors used ChatGPT (OpenAI, 2024 (158)) as a continuous support tool for translation and language refinement, with the aim of improving clarity, consistency, and adherence to academic tone. Given that the authors are non-native English speakers, this language model assisted in shaping the full text in a fluent and professional manner. The presence of generative AI is therefore embedded throughout the manuscript. Nevertheless, all content has been carefully reviewed and edited by the authors, who take full responsibility for the integrity, accuracy, and scientific validity of the manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Webb BA, Chimenti M, Jacobson MP, and Barber DL. Dysregulated pH: A perfect storm for cancer progression. Nat Rev Cancer. (2011) 11:671–7. doi: 10.1038/nrc3110
2. Corbet C and Feron O. Cancer cell metabolism and mitochondria: Nutrient plasticity for TCA cycle fueling. Biochim Biophys Acta Rev Cancer. (2017) 1868:7–15. doi: 10.1016/j.bbcan.2017.01.002
3. Persi E, Duran-Frigola M, Damaghi M, Roush WR, Aloy P, Cleveland JL, et al. Systems analysis of intracellular pH vulnerabilities for cancer therapy. Nat Commun. (2018) 9:2997. doi: 10.1038/s41467-018-05261-x
4. Shirmanova MV, Druzhkova IN, Lukina MM, Matlashov ME, Belousov VV, Snopova LB, et al. Intracellular pH imaging in cancer cells in vitro and tumors in vivo using the new genetically encoded sensor SypHer2. Biochim Biophys Acta. (2015) 1850:1905–11. doi: 10.1016/j.bbagen.2015.05.001
5. Robey IF. Examining the relationship between diet-induced acidosis and cancer. Nutr Metab (Lond). (2012) 9:72. doi: 10.1186/1743-7075-9-72
6. Bogdanov A, Bogdanov A, Chubenko V, Volkov N, Moiseenko F, and Moiseyenko V. Tumor acidity: From hallmark of cancer to target of treatment. Front Oncol. (2022) 12:979154. doi: 10.3389/fonc.2022.979154
7. Tafech A and Stéphanou A. On the importance of acidity in cancer cells and therapy. Biol (Basel). (2024) 13:225. doi: 10.3390/biology13040225
8. Barba I, Carrillo-Bosch L, and Seoane J. Targeting the warburg effect in cancer: where do we stand? Int J Mol Sci. (2024) 25:3142. doi: 10.3390/ijms25063142
9. Kalinin S, Malkova A, Sharonova T, Sharoyko V, Bunev A, Supuran CT, et al. Carbonic anhydrase IX inhibitors as candidates for combination therapy of solid tumors. Int J Mol Sci. (2021) 22:13405. doi: 10.3390/ijms222413405
10. Rezuchova I, Bartosova M, Belvoncikova P, Takacova M, Zatovicova M, Jelenska L, et al. Carbonic anhydrase IX in tumor tissue and plasma of breast cancer patients: reliable biomarker of hypoxia and prognosis. Int J Mol Sci. (2023) 24:4325. doi: 10.3390/ijms24054325
11. Becker HM and Deitmer JW. Proton transport in cancer cells: the role of carbonic anhydrases. Int J Mol Sci. (2021) 22:3171. doi: 10.3390/ijms22063171
12. Rahman MA, Yadab MK, and Ali MM. Emerging role of extracellular pH in tumor microenvironment as a therapeutic target for cancer immunotherapy. Cells. (2024) 13:1924. doi: 10.3390/cells13221924
13. Pukkanasut P, Jaskula-Sztul R, Gomora JC, and Velu SE. Therapeutic targeting of voltage-gated sodium channel NaV1.7 for cancer metastasis. Front Pharmacol. (2024) 15:1416705. doi: 10.3389/fphar.2024.1416705
14. Kobliakov VA. Role of proton pumps in tumorigenesis. Biochem (Mosc). (2017) 82:401–12. doi: 10.1134/S0006297917040010
15. Gaspary JFP, Edgar L, Lopes LFD, Rosa CB, and Siluk JCM. Translational insights into the hormetic potential of carbon dioxide: from physiological mechanisms to innovative adjunct therapeutic potential for cancer. Front Physiol. (2024) 15:1415037. doi: 10.3389/fphys.2024.1415037
16. Li Z, Zhang W, Guo S, Qi G, Huang J, Gao J, et al. A review of advances in mitochondrial research in cancer. Cancer Control. (2024) 31:10732748241299072. doi: 10.1177/10732748241299072
17. Di Gregorio J, Petricca S, Iorio R, Toniato E, and Flati V. Mitochondrial and metabolic alterations in cancer cells. Eur J Cell Biol. (2022) 101:151225. doi: 10.1016/j.ejcb.2022.151225
18. Wang SF, Tseng LM, and Lee HC. Role of mitochondrial alterations in human cancer progression and cancer immunity. J BioMed Sci. (2023) 30:61. doi: 10.1186/s12929-023-00956-w
19. Bader JE, Voss K, and Rathmell JC. Targeting metabolism to improve the tumor microenvironment for cancer immunotherapy. Mol Cell. (2020) 78:1019–33. doi: 10.1016/j.molcel.2020.05.034
20. Vaupel P, Schmidberger H, and Mayer A. The Warburg effect: essential part of metabolic reprogramming and central contributor to cancer progression. Int J Radiat Biol. (2019) 95:912–9. doi: 10.1080/09553002.2019.1589653
21. Liberti MV and Locasale JW. The warburg effect: how does it benefit cancer cells? Trends Biochem Sci. (2016) 41:211–8. doi: 10.1016/j.tibs.2015.12.001
22. Chesbrough H. Open innovation: The new imperative for creating and profiting from technology. Boston (MA: Harvard Business School Publishing (2003).
24. Bana e Costa CA, Ensslin L, Corrêa EC, and Vansnick JC. Decision support systems in action: Integrated application in a multicriteria decision aid process. Eur J Oper Res. (1999) 113:315–20. doi: 10.1016/S0377-2217(98)00219-7
25. Decka D, Schwaab G, and Havenith M. A THz/FTIR fingerprint of the solvated proton: Evidence for Eigen structure and Zundel dynamics. Phys Chem Chem Phys. (2015) 17:11898–907. doi: 10.1039/c5cp01035g
26. Knight C and Voth GA. The curious case of the hydrated proton. Acc Chem Res. (2012) 45:101–9. doi: 10.1021/ar200140h
27. Popov I, Zhu Z, Young-Gonzales AR, Sacci RL, Mamontov E, Gainaru C, et al. Search for a Grotthuss mechanism through the observation of proton transfer. Commun Chem. (2023) 6:77. doi: 10.1038/s42004-023-00878-6
28. Das S, Imoto S, Sun S, Nagata Y, Backus EHG, and Bonn M. Nature of excess hydrated proton at the water-air interface. J Am Chem Soc. (2020) 142:945–52. doi: 10.1021/jacs.9b10807
29. Calio PB, Li C, and Voth GA. Resolving the structural debate for the hydrated excess proton in water. J Am Chem Soc. (2021) 143:18672–83. doi: 10.1021/jacs.1c08552
30. Project Management Institute. A guide to the project management body of knowledge. 7th. Newtown Square (PA: PMI (2021).
31. Project Management Institute. Practice standard for work breakdown structures. 3rd. Newtown Square (PA: PMI (2019).
32. Page MJ, Moher D, Bossuyt PM, Boutron I, Hoffmann TC, Mulrow CD, et al. PRISMA 2020 explanation and elaboration: Updated guidance and exemplars for reporting systematic reviews. BMJ. (2021) 372:n160. doi: 10.1136/bmj.n160
34. Hulley SB, Cummings SR, Browner WS, Grady DG, and Newman TB. Designing clinical research. 3rd. Philadelphia (PA: Lippincott Williams & Wilkins (2007).
35. Boedtkjer E and Pedersen SF. The acidic tumor microenvironment as a driver of cancer. Annu Rev Physiol. (2020) 82:103–26. doi: 10.1146/annurev-physiol-021119-034627
36. Czowski BJ, Romero-Moreno R, Trull KJ, and White KA. Cancer and pH dynamics: transcriptional regulation, proteostasis, and the need for new molecular tools. Cancers (Basel). (2020) 12:2760. doi: 10.3390/cancers12102760
37. Liu Y, White KA, and Barber DL. Intracellular pH regulates cancer and stem cell behaviors: A protein dynamics perspective. Front Oncol. (2020) 10:1401. doi: 10.3389/fonc.2020.01401
38. Spugnini EP, Sonveaux P, Stock C, Perez-Sayans M, De Milito A, Avnet S, et al. Proton channels and exchangers in cancer. Biochim Biophys Acta. (2015) 1848(10 Pt B):2715–26. doi: 10.1016/j.bbamem.2014.10.015
39. Boedtkjer E. Ion channels, transporters, and sensors interact with the acidic tumor microenvironment to modify cancer progression. Rev Physiol Biochem Pharmacol. (2022) 182:39–84. doi: 10.1007/112_2021_63
40. Zheng T, Jäättelä M, and Liu B. pH gradient reversal fuels cancer progression. Int J Biochem Cell Biol. (2020) 125:105796. doi: 10.1016/j.biocel.2020.105796
41. Korenchan DE and Flavell RR. Spatiotemporal pH heterogeneity as a promoter of cancer progression and therapeutic resistance. Cancers (Basel). (2019) 11:1026. doi: 10.3390/cancers11071026
42. Proietto M, Crippa M, Damiani C, Pasquale V, Sacco E, Vanoni M, et al. Tumor heterogeneity: preclinical models, emerging technologies, and future applications. Front Oncol. (2023) 13:1164535. doi: 10.3389/fonc.2023.1164535
43. Gong Q, Song X, Tong Y, Huo L, Zhao X, Han Y, et al. Recent advances of anti-tumor nano-strategies via overturning pH gradient: alkalization and acidification. J Nanobiotechnol. (2025) 23:42. doi: 10.1186/s12951-025-03134-2
44. Piasentin N, Milotti E, and Chignola R. The control of acidity in tumor cells: a biophysical model. Sci Rep. (2020) 10:13613. doi: 10.1038/s41598-020-70396-1
45. Alfarouk KO. Tumor metabolism, cancer cell transporters, and microenvironmental resistance. J Enzyme Inhib Med Chem. (2016) 31:859–66. doi: 10.3109/14756366.2016.1140753
46. Gentile R, Feudi D, Sallicandro L, and Biagini A. Can the tumor microenvironment alter ion channels? Unraveling their role in cancer. Cancers (Basel). (2025) 17:1244. doi: 10.3390/cancers17071244
47. Peretti M, Badaoui M, Girault A, Van Gulick L, Mabille MP, Tebbakha R, et al. Original association of ion transporters mediates the ECM-induced breast cancer cell survival: Kv10.1-Orai1-SPCA2 partnership. Sci Rep. (2019) 9:1175. doi: 10.1038/s41598-018-37602-7
48. Cardone RA, Alfarouk KO, Elliott RL, Alqahtani SS, Ahmed SBM, Aljarbou Begum HM, et al. Intracellular and microenvironmental regulation of mitochondrial membrane potential in cancer cells. WIREs Mech Dis. (2023) 15:e1595. doi: 10.1002/wsbm.1595
49. Cardone RA, Alfarouk KO, Elliott RL, Alqahtani SS, Ahmed SBM, Aljarbou AN, et al. The role of sodium hydrogen exchanger 1 in dysregulation of proton dynamics and reprogramming of cancer metabolism as a sequela. Int J Mol Sci. (2019) 20:3694. doi: 10.3390/ijms20153694
50. Fecikova S, Csaderova L, Belvoncikova P, Puzderova B, Bernatova K, Talac T, et al. Can hypoxia marker carbonic anhydrase IX serve as a potential new diagnostic marker and therapeutic target of non-small cell lung cancer? Neoplasma. (2024) 71:123–42. doi: 10.4149/neo_2024_231219N652
51. Miranda-Gonçalves V, Granja S, Martinho O, Honavar M, Pojo M, Costa BM, et al. Hypoxia-mediated upregulation of MCT1 expression supports the glycolytic phenotype of glioblastomas. Oncotarget. (2016) 7:46335–43. doi: 10.18632/oncotarget.10114
52. Schniers BK, Rajasekaran D, Korac K, Sniegowski T, Ganapathy V, and Bhutia YD. PEPT1 is essential for the growth of pancreatic cancer cells: a viable drug target. Biochem J. (2021) 478:3757–74. doi: 10.1042/BCJ20210377
53. Kim EY, Abides J, Keller CR, Martinez SR, and Li W. Tumor microenvironment lactate: is it a cancer progression marker, immunosuppressant, and therapeutic target? Molecules. (2025) 30:1763. doi: 10.3390/molecules30081763
54. Che R, Wang Q, Li M, Shen J, and Ji J. Quantitative proteomics of tissue-infiltrating T cells from CRC patients identified lipocalin-2 induces T-cell apoptosis and promotes tumor cell proliferation by iron efflux. Mol Cell Proteomics. (2024) 23:100691. doi: 10.1016/j.mcpro.2023.100691
55. Lu K, Wijaya CS, Yao Q, Jin H, and Feng L. Cuproplasia and cuproptosis, two sides of the coin. Cancer Commun (Lond). (2025) 45:505–24. doi: 10.1002/cac2.70001
56. Pang Z. Copper metabolism in hepatocellular carcinoma: from molecular mechanisms to therapeutic opportunities. Front Mol Biosci. (2025) 12:1578693. doi: 10.3389/fmolb.2025.1578693
57. Lin Z, He Y, Wu Z, Yuan Y, Li X, and Luo W. Comprehensive analysis of copper-metabolism-related genes about prognosis and immune microenvironment in osteosarcoma. Sci Rep. (2023) 13:15059. doi: 10.1038/s41598-023-42053-w
58. Wang Z, Jin D, Zhou S, Dong N, Ji Y, An P, et al. Regulatory roles of copper metabolism and cuproptosis in human cancers. Front Oncol. (2023) 13:1123420. doi: 10.3389/fonc.2023.1123420
59. Kong L, Liu M, Yang H, Yan P, Luo Y, Xiang S, et al. Expression of copper metabolism-related genes is associated with the tumor immune microenvironment and predicts the prognosis of hepatocellular carcinoma. Transl Cancer Res. (2024) 13:2251–65. doi: 10.21037/tcr-23-1890
60. Stolwijk JA, Wallner S, Heider J, Kurz B, Pütz L, Michaelis S, et al. GPR4 in the pH-dependent migration of melanoma cells in the tumor microenvironment. Exp Dermatol. (2023) 32:479–90. doi: 10.1111/exd.14735
61. Kurz B, Michael HP, Förch A, Wallner S, Zeman F, Decking SM, et al. Expression of pH-sensitive TRPC4 in common skin tumors. Int J Mol Sci. (2023) 24:1037. doi: 10.3390/ijms24021037
62. Marozzi M, Parnigoni A, Negri A, Viola M, Vigetti D, Passi A, et al. Inflammation, extracellular matrix remodeling, and proteostasis in tumor microenvironment. Int J Mol Sci. (2021) 22:8102. doi: 10.3390/ijms22158102
63. Peters F and Becker-Pauly C. Role of meprin metalloproteases in metastasis and tumor microenvironment. Cancer Metastasis Rev. (2019) 38:347–56. doi: 10.1007/s10555-019-09805-5
64. Turunen SP, Tatti-Bugaeva O, and Lehti K. Membrane-type matrix metalloproteases as diverse effectors of cancer progression. Biochim Biophys Acta Mol Cell Res. (2017) 1864(11 Pt A):1974–1988. doi: 10.1016/j.bbamcr.2017.04.002
65. Wang Q, Wang K, Tan X, Li Z, and Wang H. Immunomodulatory role of metalloproteases in cancers: Current progress and future trends. Front Immunol. (2022) 13:1064033. doi: 10.3389/fimmu.2022.1064033
66. Ding P, Zhang X, Jin S, Duan B, Chu P, Zhang Y, et al. CD147 functions as the signaling receptor for extracellular divalent copper in hepatocellular carcinoma cells. Oncotarget. (2017) 8:51151–63. doi: 10.18632/oncotarget.17712
67. Zhang Y, Mo Y, Yuan J, Zhang Y, Mo L, and Zhang Q. MMP-3 activation is involved in copper oxide nanoparticle-induced epithelial-mesenchymal transition in human lung epithelial cells. Nanotoxicology. (2021) 15:1380–402. doi: 10.1080/17435390.2022.2030822
68. Ge EJ, Bush AI, Casini A, Cobine PA, Cross JR, DeNicola GM, et al. Connecting copper and cancer: from transition metal signalling to metalloplasia. Nat Rev Cancer. (2022) 22:102–13. doi: 10.1038/s41568-021-00417-2
69. Tsvetkov P, Coy S, Petrova B, Dreishpoon M, Verma A, Abdusamad M, et al. Copper induces cell death by targeting lipoylated TCA cycle proteins. Science. (2022) 375:1254–61. doi: 10.1126/science.abf0529
70. Ruiz LM, Libedinsky A, and Elorza AA. Role of copper on mitochondrial function and metabolism. Front Mol Biosci. (2021) 8:711227. doi: 10.3389/fmolb.2021.711227
71. Swaminathan AB and Gohil VM. The role of COA6 in the mitochondrial copper delivery pathway to cytochrome c oxidase. Biomolecules. (2022) 12:125. doi: 10.3390/biom12010125
72. Supekar S, Gamiz-Hernandez AP, and Kaila VR. A protonated water cluster as a transient proton-loading site in cytochrome c oxidase. . Angew Chem Int Ed Engl. (2016) 55:11940–4. doi: 10.1002/anie.201603606
73. Saura P, Riepl D, Frey DM, Wikström M, and Kaila VRI. Electric fields control water-gated proton transfer in cytochrome c oxidase. Proc Natl Acad Sci U S A. (2022) 119:e2207761119. doi: 10.1073/pnas.2207761119
74. Gorbikova E, Samsonov SA, and Kalendar R. Probing the proton-loading site of cytochrome C oxidase using time-resolved fourier transform infrared spectroscopy. Molecules. (2020) 25:3393. doi: 10.3390/molecules25153393
75. Wu X, Bai Z, Wang H, Wang H, Hou D, Xu Y, et al. CRISPR-Cas9 gene editing strengthens cuproptosis/chemodynamic/ferroptosis synergistic cancer therapy. Acta Pharm Sin B. (2024) 14:4059–72. doi: 10.1016/j.apsb.2024.05.029
76. Verkman AS, Hara-Chikuma M, and Papadopoulos MC. Aquaporins–new players in cancer biology. J Mol Med (Berl). (2008) 86:523–9. doi: 10.1007/s00109-008-0303-9
78. Xia C, Liu C, Ren S, Cai Y, Zhang Q, and Xia C. Potassium channels, tumorigenesis and targeted drugs. BioMed Pharmacother. (2023) 162:114673. doi: 10.1016/j.biopha.2023.114673
79. Huang W, Liu Y, Luz A, Berrong M, Meyer JN, Zou Y, et al. Calcium/calmodulin dependent protein kinase kinase 2 regulates the expansion of tumor-induced myeloid-derived suppressor cells. Front Immunol. (2021) 12:754083. doi: 10.3389/fimmu.2021.754083
80. Zhan Q, Jeon J, Li Y, Huang Y, Xiong J, Wang Q, et al. CAMK2/CaMKII activates MLKL in short-term starvation to facilitate autophagic flux. Autophagy. (2022) 18:726–44. doi: 10.1080/15548627.2021.1954348
81. Angeli A, Carta F, Nocentini A, Winum JY, Zalubovskis R, Akdemir A, et al. Carbonic anhydrase inhibitors targeting metabolism and tumor microenvironment. Metabolites. (2020) 10:412. doi: 10.3390/metabo10100412
82. Pastorek J and Pastorekova S. Hypoxia-induced carbonic anhydrase IX as a target for cancer therapy: from biology to clinical use. Semin Cancer Biol. (2015) 31:52–64. doi: 10.1016/j.semcancer.2014.08.002
83. Lei G, Zhuang L, and Gan B. The roles of ferroptosis in cancer: Tumor suppression, tumor microenvironment, and therapeutic interventions. Cancer Cell. (2024) 42:513–34. doi: 10.1016/j.ccell.2024.03.011
84. Wang S, Guo Q, Xu R, Lin P, Deng G, and Xia X. Combination of ferroptosis and pyroptosis dual induction by triptolide nano-MOFs for immunotherapy of Melanoma. J Nanobiotechnol. (2023) 21:383. doi: 10.1186/s12951-023-02146-0
85. Yu Z, Lepoitevin M, and Serre C. Iron-MOFs for biomedical applications. Adv Healthc Mater. (2025) 14:e2402630. doi: 10.1002/adhm.202402630
86. Gonzalez-Avila G, Sommer B, García-Hernández AA, and Ramos C. Matrix metalloproteinases’ role in tumor microenvironment. Adv Exp Med Biol. (2020) 1245:97–131. doi: 10.1007/978-3-030-40146-7_5
87. Niland S, Riscanevo AX, and Eble JA. Matrix metalloproteinases shape the tumor microenvironment in cancer progression. Int J Mol Sci. (2022) 23:146. doi: 10.3390/ijms23010146
88. Mendivil-Alvarado H, Limon-Miro AT, Carvajal-Millan E, Lizardi-Mendoza J, Mercado-Lara A, Coronado-Alvarado CD, et al. Extracellular vesicles and their zeta potential as future markers associated with nutrition and molecular biomarkers in breast cancer. Int J Mol Sci. (2023) 24:6810. doi: 10.3390/ijms24076810
89. Hughes MP. The cellular zeta potential: cell electrophysiology beyond the membrane. Integr Biol (Camb). (2024) 16:zyae003. doi: 10.1093/intbio/zyae003
90. Zhang M, Li T, Zhu J, Tuo B, and Liu X. Physiological and pathophysiological role of ion channels and transporters in the colorectum and colorectal cancer. J Cell Mol Med. (2020) 24:9486–94. doi: 10.1111/jcmm.15600
91. Parayath NN, Padmakumar S, and Amiji MM. Extracellular vesicle-mediated nucleic acid transfer and reprogramming in the tumor microenvironment. Cancer Lett. (2020) 482:33–43. doi: 10.1016/j.canlet.2020.04.009
92. Gondaliya P, Sayyed AA, Driscoll J, Patel K, and Patel T. Extracellular vesicle RNA signaling in the liver tumor microenvironment. Cancer Lett. (2023) 558:216089. doi: 10.1016/j.canlet.2023.216089
93. Semeradtova A, Liegertova M, Herma R, Capkova M, Brignole C, and Del Zotto G. Extracellular vesicles in cancer’s communication: messages we can read and how to answer. Mol Cancer. (2025) 24:86. doi: 10.1186/s12943-025-02282-1
94. Scharping NE, Rivadeneira DB, Menk AV, Vignali PDA, Ford BR, Rittenhouse NL, et al. Mitochondrial stress induced by continuous stimulation under hypoxia rapidly drives T cell exhaustion. Nat Immunol. (2021) 22:205–15. doi: 10.1038/s41590-020-00834-9
95. Li Y, Zhao L, and Li XF. Hypoxia and the tumor microenvironment. Technol Cancer Res Treat. (2021) 20:15330338211036304. doi: 10.1177/15330338211036304
96. Rahman MA, Ahmed KR, Haque F, Park MN, and Kim B. Recent advances in cellular signaling interplay between redox metabolism and autophagy modulation in cancer: an overview of molecular mechanisms and therapeutic interventions. Antioxidants (Basel). (2023) 12:428. doi: 10.3390/antiox12020428
97. Ren Y, Wang R, Weng S, Xu H, Zhang Y, Chen S, et al. Multifaceted role of redox pattern in the tumor immune microenvironment regarding autophagy and apoptosis. Mol Cancer. (2023) 22:130. doi: 10.1186/s12943-023-01831-w
98. Hanahan D. Hallmarks of cancer: new dimensions. Cancer Discov. (2022) 12:31–46. doi: 10.1158/2159-8290.CD-21-1059
99. Jain RK. Normalizing tumor microenvironment to treat cancer: bench to bedside to biomarkers. J Clin Oncol. (2013) 31:2205–18. doi: 10.1200/JCO.2012.46.3653
100. Farhana A, Alsrhani A, Khan YS, and Rasheed Z. Cancer bioenergetics and tumor microenvironments—Enhancing chemotherapeutics and targeting resistant niches through nanosystems. Cancers (Basel). (2023) 15:3836. doi: 10.3390/cancers15153836
101. Lei T, Rui Y, Xiaoshuang Z, Jinglan Z, and Jihong Z. Mitochondria transcription and cancer. Cell Death Discov. (2024) 10:168. doi: 10.1038/s41420-024-01926-3
102. Pham L, Arroum T, Wan J, Pavelich L, Bell J, Morse PT, et al. Regulation of mitochondrial oxidative phosphorylation through tight control of cytochrome c oxidase in health and disease—Implications for ischemia/reperfusion injury, inflammatory diseases, diabetes, and cancer. Redox Biol. (2024) 78:103426. doi: 10.1016/j.redox.2024.103426
103. Gabriel C, Gabriel S, and Corthout E. The dielectric properties of biological tissues: I. Literature survey. Phys Med Biol. (1996) 41:2231–49. doi: 10.1088/0031-9155/41/11/001
104. Gabriel S, Lau RW, and Gabriel C. The dielectric properties of biological tissues: II. Measurements in the frequency range 10 Hz to 20 GHz. Phys Med Biol. (1996) 41:2251–69. doi: 10.1088/0031-9155/41/11/002
105. Markovitch O, Chen H, Izvekov S, Paesani F, Voth GA, and Agmon N. Special pair dance and partner selection: Elementary steps in proton transport in liquid water. J Phys Chem B. (2008) 112:9456–66. doi: 10.1021/jp804018y
106. Intharathep P, Tongraar A, and Sagarik K. Ab initio QM/MM dynamics of H3O+ in water. J Comput Chem. (2006) 27:1723–32. doi: 10.1002/jcc.20503
107. Soniat M, Kumar R, and Rick SW. Hydrated proton and hydroxide charge transfer at the liquid/vapor interface of water. J Chem Phys. (2015) 143:044702. doi: 10.1063/1.4926831
108. Schröder M, Gatti F, Lauvergnat D, Meyer HD, and Vendrell O. The coupling of the hydrated proton to its first solvation shell. Nat Commun. (2022) 13:6170. doi: 10.1038/s41467-022-33650-w
109. Nicolas-Boluda A, Silva AKA, Fournel S, and Gazeau F. Physical oncology: New targets for nanomedicine. Biomaterials. (2018) 150:87–99. doi: 10.1016/j.biomaterials.2017.10.014
110. Haynes WM. CRC handbook of chemistry and physics. 95th. Boca Raton (FL: CRC Press (2014). doi: 10.1201/b17118
111. Cukrowski I, Lange JH, Groenewald F, and Raubenheimer HG. Gold(I) hydrides as proton acceptors in dihydrogen bond formation. ChemPhysChem. (2017) 18:2288–94. doi: 10.1002/cphc.201700383
112. Oksanen E, Chen JC, and Fisher SZ. Neutron crystallography for the study of hydrogen bonds in macromolecules. Molecules. (2017) 22:596. doi: 10.3390/molecules22040596
113. Silverstein TP. The proton in biochemistry: Impacts on bioenergetics, biophysical chemistry, and bioorganic chemistry. Front Mol Biosci. (2021) 8:764099. doi: 10.3389/fmolb.2021.764099
114. Wurzbarger SR. High molecular weight ionized water. U.S. Patent No. 4,613,416. Washington (DC: U.S. Patent and Trademark Office (1986).
115. Schran C and Marx D. Quantum nature of the hydrogen bond from ambient conditions down to ultra-low temperatures. Phys Chem Chem Phys. (2019) 21:24967–75. doi: 10.1039/c9cp04795f
116. Teschke O, de Castro JR, Valente Filho JF, and Soares DM. Hydrated excess proton Raman spectral densities probed in floating water bridges. ACS Omega. (2018) 3:13977–83. doi: 10.1021/acsomega.8b02285
117. Mouhat F, Peria M, Morresi T, Vuilleumier R, Saitta AM, and Casula M. Thermal dependence of the hydrated proton and optimal proton transfer in the protonated water hexamer. Nat Commun. (2023) 14:6930. doi: 10.1038/s41467-023-42366-4
118. Kwang-Hua CW. Temperature-dependent viscosity dominated transport control through AQP1 water channel. J Theor Biol. (2019) 480:92–8. doi: 10.1016/j.jtbi.2019.08.006
119. Alkhadra MA, Su X, Suss ME, Tian H, Guyes EN, Shocron AN, et al. Electrochemical methods for water purification, ion separations, and energy conversion. Chem Rev. (2022) 122:13547–635. doi: 10.1021/acs.chemrev.1c00396
120. Dudev T, Musset B, Morgan D, Cherny VV, Smith SM, Mazmanian K, et al. Selectivity mechanism of the voltage-gated proton channel, HV1. Sci Rep. (2015) 5:10320. doi: 10.1038/srep10320
121. Jastroch M, Divakaruni AS, Mookerjee S, Treberg JR, and Brand MD. Mitochondrial proton and electron leaks. Essays Biochem. (2010) 47:53–67. doi: 10.1042/bse0470053
122. Berry BJ, Trewin AJ, Amitrano AM, Kim M, and Wojtovich AP. Use the protonmotive force: mitochondrial uncoupling and reactive oxygen species. J Mol Biol. (2018) 430:3873–91. doi: 10.1016/j.jmb.2018.03.025
123. Mallick S and Agmon N. Multi-proton dynamics near membrane-water interface. Nat Commun. (2025) 16:3276. doi: 10.1038/s41467-025-58167-w
124. Zhu Y. Production, characteristics, and biological effects of protonated small water clusters. Proc SPIE. (2023) 25:127580E-1–9. doi: 10.1117/12.3005480
125. Guo Z, Chen D, Yao L, Sun Y, Li D, Le J, et al. The molecular mechanism and therapeutic landscape of copper and cuproptosis in cancer. Signal Transduct Target Ther. (2025) 10:149. doi: 10.1038/s41392-025-02192-0
126. Zhang X, Tang B, Luo J, Yang Y, Weng Q, Fang S, et al. Cuproptosis, ferroptosis and PANoptosis in tumor immune microenvironment remodeling and immunotherapy: culprits or new hope. Mol Cancer. (2024) 23:255. doi: 10.1186/s12943-024-02130-8
127. Becker HM. Carbonic anhydrase IX and acid transport in cancer. Br J Cancer. (2020) 122:157–67. doi: 10.1038/s41416-019-0642-z
128. Chen T, Lin X, Lu S, and Li B. V-ATPase in cancer: mechanistic insights and therapeutic potentials. Cell Commun Signal. (2024) 22:613. doi: 10.1186/s12964-024-01998-9
129. Zong Y, Li H, Liao P, Chen L, Pan Y, Zheng Y, et al. Mitochondrial dysfunction: mechanisms and advances in therapy. Signal Transduct Target Ther. (2024) 9:124. doi: 10.1038/s41392-024-01839-8
130. Nirody JA, Budin I, and Rangamani P. ATP synthase: Evolution, energetics, and membrane interactions. J Gen Physiol. (2020) 152:e201912475. doi: 10.1085/jgp.201912475
131. Clemente LP, Rabenau M, Tang S, Stanka J, Cors E, et al. Dynasore blocks ferroptosis through combined modulation of iron uptake and inhibition of mitochondrial respiration. Cells. (2020) 9:2259. doi: 10.3390/cells9102259
132. Berndt C, Alborzinia H, Amen VS, Ayton S, Barayev U, Bartelt A, et al. Ferroptosis in health and disease. Redox Biol. (2024) 75:103211. doi: 10.1016/j.redox.2024.103211
133. Agmon E, Solon J, Bassereau P, and Stockwell BR. Modeling the effects of lipid peroxidation during ferroptosis on membrane properties. Sci Rep. (2018) 8:5155. doi: 10.1038/s41598-018-23408-0
134. Roeck BF, Lotfipour Nasudivar S, Vorndran MRH, Schueller L, Yapici FI, et al. Ferroptosis spreads to neighboring cells via plasma membrane contacts. Nat Commun. (2025) 16:2951. doi: 10.1038/s41467-025-58175-w
135. Talachutla S, Bhat S, Duster AW, and Lin H. Improved indicator algorithms for tracking a hydrated proton as a local structural defect in grotthuss diffusion in aqueous solutions. Chem Phys Lett. (2021) 784:139121. doi: 10.1016/j.cplett.2021.139121
136. Wang Z. Cu in cancer metabolism. Nat Cell Biol. (2024) 26:2013. doi: 10.1038/s41556-024-01578-6
137. Guan M, Cheng K, Xie XT, Li Y, Ma MW, Zhang B, et al. Regulating copper homeostasis of tumor cells to promote cuproptosis for enhancing breast cancer immunotherapy. Nat Commun. (2024) 15:10060. doi: 10.1038/s41467-024-54469-7
138. Shen F, Fang Y, Wu Y, Zhou M, Shen J, and Fan X. Metal ions and nanometallic materials in antitumor immunity: Function, application, and perspective. J Nanobiotechnol. (2023) 21:20. doi: 10.1186/s12951-023-01771-z
139. Zhao L, Gui Y, Cai J, and Deng X. Biometallic ions and derivatives: a new direction for cancer immunotherapy. Mol Cancer. (2025) 24:17. doi: 10.1186/s12943-025-02225-w
140. You Y, Guo Z, Wolter T, and Hu Q. Intracellular metal ion-based chemistry for programmed cell death. Chem Soc Rev. (2025) 54:1552–82. doi: 10.1039/D4CS00930D
141. Jin J, Dai S, Li X, Liu Y, and Lu H. pH-switchable latexes based on the nonionic amphiphilic diblock copolymer with a chargeable end-group on the core-forming block. Langmuir. (2021) 37:6123–31. doi: 10.1021/acs.langmuir
142. Tyrode E, Sengupta S, and Sthoer A. Identifying Eigen-like hydrated protons at negatively charged interfaces. Nat Commun. (2020) 11:493. doi: 10.1038/s41467-020-14370-5
143. Deplazes E, Poger D, Cornell B, and Cranfield CG. The effect of H3O+ on the membrane morphology and hydrogen bonding of a phospholipid bilayer. Biophys Rev. (2018) 10:1371–5. doi: 10.1007/s12551-018-0454-z
144. Wu Q, Zhang H, Sun S, Wang L, and Sun S. Extracellular vesicles and immunogenic stress in cancer. Cell Death Dis. (2021) 12:894. doi: 10.1038/s41419-021-04171-z
145. Arntsen C, Chen C, Calio PB, Li C, and Voth GA. The hopping mechanism of the hydrated excess proton and its contribution to proton diffusion in water. J Chem Phys. (2021) 154:194506. doi: 10.1063/5.0040758
146. Miyake T and Rolandi M. Grotthuss mechanisms: from proton transport in proton wires to bioprotonic devices. J Phys Condens Matter. (2016) 28:23001. doi: 10.1088/0953-8984/28/2/023001
147. Zlobin A, Smirnov I, and Golovin A. Dynamic interchange between two protonation states is characteristic of active sites of cholinesterases. Protein Sci. (2024) 33:e5100. doi: 10.1002/pro.5100
148. Morgan D and DeCoursey TE. Diversity of voltage gated proton channels. Front Biosci. (2003) 8:s1266–79. doi: 10.2741/1191
149. Lee IS and Nandab SS. Proton transfer reaction in water: Hydronium ion formation. Adv J Chem A. (2020) 3:255–8. doi: 10.33945/SAMI/AJCA.2020.3.2
150. Aklima J, Onojima T, Kimura S, Umiuchi K, Shibata T, Kuraoka Y, et al. Effects of matrix pH on spontaneous transient depolarization and reactive oxygen species production in mitochondria. Front Cell Dev Biol. (2021) 9:692776. doi: 10.3389/fcell.2021.692776
151. Bertholet AM, Natale AM, Bisignano P, Suzuki J, Fedorenko A, Hamilton J, et al. Mitochondrial uncouplers induce proton leak by activating AAC and UCP1. Nature. (2022) 606:180–7. doi: 10.1038/s41586-022-04747-5
152. Ronca R and Supuran CT. Carbonic anhydrase IX: An atypical target for innovative therapies in cancer. Biochim Biophys Acta Rev Cancer. (2024) 1879:189120. doi: 10.1016/j.bbcan.2024.189120
153. Surh YJ, Chung HT, Na HK, Dulak J, and Stec DE. Progress in heme oxygenase research. Arch Biochem Biophys. (2020) 685:108321. doi: 10.1016/j.abb.2020.108321
154. Flessa S and Huebner C. Innovations in health care—A conceptual framework. Int J Environ Res Public Health. (2021) 18:10026. doi: 10.3390/ijerph181910026
155. Riveros-Perez E and Rivero R. Water in the human body: An anesthesiologist’s perspective on the connection between physicochemical properties of water and physiologic relevance. Ann Med Surg (Lond). (2017) 27:1–8. doi: 10.1016/j.amsu.2017.12.007
156. Gabriel S, Lau RW, and Gabriel C. The dielectric properties of biological tissues: III. Parametric models for the dielectric spectrum of tissues. Phys Med Biol. (1996) 41:2271–93. doi: 10.1088/0031-9155/41/11/003
157. Strand D. Everyday characterizations of translational research: Researchers’ own use of terminology and models in medical research and practice. Palgrave Commun. (2020) 6:10. doi: 10.1057/s41599-020-0489-1
158. OpenAI. ChatGPT-4o (March 9 version) [Large language model]. San Francisco, CA, United States: OpenAI (2025). Available online at: https://openai.com (Accessed August 15, 2025).
Keywords: tumor microenvironment, hydrated proton complexes, mitochondrial dysfunction, Warburg effect, redox signaling, bioenergetic reprogramming
Citation: Edgar AL, Lopes LFD, Lopes EG, Felin ID, Felin CR and Gaspary JFP (2025) Hydrated proton complexes supplementation for tumor microenvironment reprogramming: a bioenergetic strategy targeting the Warburg effect and mitochondrial dysfunction. Front. Oncol. 15:1647054. doi: 10.3389/fonc.2025.1647054
Received: 14 June 2025; Accepted: 04 August 2025;
Published: 05 September 2025.
Edited by:
Egidio Iorio, National Institute of Health (ISS), ItalyReviewed by:
Aleš Dvořák, Charles University, CzechiaRosa Vona, National Institute of Health (ISS), Italy
Copyright © 2025 Edgar, Lopes, Lopes, Felin, Felin and Gaspary. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Luis Felipe Dias Lopes, bHVpcy5sb3Blc0B1ZnNtLmJy