 Fernando Pasteran1*
Fernando Pasteran1* Juan Manuel De Mendieta1Natalia Pujato2Gina Dotta3†Lisandro J. González3,4Mabel Rizzo5Alejandra Fernández6Paola Ceriana1Lucia Maccari1Melina Rapoport1
Juan Manuel De Mendieta1Natalia Pujato2Gina Dotta3†Lisandro J. González3,4Mabel Rizzo5Alejandra Fernández6Paola Ceriana1Lucia Maccari1Melina Rapoport1 Sonia Gómez1Celeste Lucero1María Alejandra Menocal1Ezequiel Albornoz1Denise De Belder7Marcelo Radisic2
Sonia Gómez1Celeste Lucero1María Alejandra Menocal1Ezequiel Albornoz1Denise De Belder7Marcelo Radisic2 Alejandro J. Vila3,4Alejandra Corso1
Alejandro J. Vila3,4Alejandra Corso1- 1Servicio Antimicrobianos, National Reference Laboratory for Antimicrobial Resistance, Instituto Nacional de Enfermedades Infecciosas, ANLIS “Dr. Carlos Malbrán”, Buenos Aires, Argentina
- 2Instituto de Trasplante de Alta Complejidad, Buenos Aires, Argentina
- 3Instituto de Biología Molecular y Celular de Rosario, CONICET, Universidad Nacional de Rosario, Rosario, Argentina
- 4Facultad de Ciencias Bioquímicas y Farmacéuticas, Universidad Nacional de Rosario, Rosario, Argentina
- 5Laboratorio de Microbiología, Hospital Interzonal General de Agudos “Dr. José Penna”, Bahía Blanca, Argentina
- 6Laboratorio de Microbiología “Dr. Rapela”, Buenos Aires, Argentina
- 7Unidad Operativa Centro Nacional de Genómica y Bioinformática, Instituto Nacional de Enfermedades Infecciosas, ANLIS “Dr. Carlos Malbrán”, Buenos Aires, Argentina
Introduction: The spread pan-drug resistant pathogens pose a critical challenge to current therapies, resulting in high mortality and necessitating alternative approaches.
Methods: We report pan-drug resistant Klebsiella pneumoniae isolates from five patients in a single hospital, including resistance to cefiderocol and cefepime-zidebactam in one isolate.
Results: Whole-genome sequencing identified blaNDM-5 and blaCTX-M-15 genes in all isolates, explaining carbapenemase and extended-spectrum β- lactamase phenotypes, with blaKPC-2 in one isolate. A novel sulfhydryl variable β-lactamase (SHV) variant, blaSHV-231, was present in all isolates under a strong promoter. Two isolates exhibited a non-synonymous mutation in fstI encoding PBP3, the primary target of aztreonam in Gram-negative bacteria. Genomic and phenotypic characterization guided successful compassionate treatment using aztreonam, ceftazidime-avibactam, and amoxicillin-clavulanate at maximum doses.
Discussion: Dissection of the roles of the substitutions present in blaSHV-231 revealed that this variant was responsible for the reduced susceptibility to aztreonam-avibactam, at the expense of a higher susceptibility to clavulanate. Targeted therapy can be successful upon dissection of unexpected mechanisms of resistance that enhance the contribution of endemic β-lactamase.
1 Introduction
Antimicrobial resistance is a critical global threat to human health. The emergence of multidrug resistance in Gram-negative pathogens has outpaced the development, distribution and availability of new antimicrobial agents (Bonomo et al., 2024). Particularly concerning is the recent rise of pan-drug-resistant (PDR) Gram-negative bacteria, which are non-susceptible to all conventional antimicrobial agents (Karakonstantis et al., 2019). PDR infections, predominantly involving Klebsiella pneumoniae, Acinetobacter baumannii and Pseudomonas aeruginosa isolates in critically ill patients, are associated with high mortality rates and pose a significant public health threat (Karakonstantis et al., 2019).
The recent development of new antimicrobials to combat carbapenemase-producing organisms, such as ceftazidime-avibactam, alone or in combination with aztreonam, imipenem-relebactam, meropenem-vaborbactam and cefiderocol has provided some relief for treating challenging strains (Yahav et al., 2020; Soman et al., 2024; Wang et al., 2024). However, there are reports of resistance events to last-generation drugs, and the restricted accessibility to some of these drugs in most low- and middle-income countries excludes them from the current consensus definitions of PDR (Magiorakos et al., 2012; Yahav et al., 2020).
Metallo-β-lactamase (MBL) producers, among the most widely disseminated carbapenemase globally, are effectively treated using cefiderocol or a combination therapy of ceftazidime-avibactam and aztreonam (Tamma et al., 2024; Wise et al., 2024). In this combination, aztreonam evades hydrolysis by MBLs, while avibactam neutralizes frequently co-produced extended-spectrum β-lactamases (ESBLs). This allows aztreonam to effectively reach its target site, the penicillin-binding protein 3 (PBP3) (Georgopapadakou et al., 1982; Yahav et al., 2020).
Carbapenemase producers are endemic in Argentina, with blaNDM and blaKPC being responsible for 88% of carbapenemase-producing Enterobacterales infections (Echegorry et al., 2024). NDM producers in Argentina are typically susceptible to aztreonam plus ceftazidime-avibactam (Echegorry et al., 2024). Only one previously documented outbreak of PDR Enterobacterales had occurred in Argentina, before the local approval of ceftazidime-avibactam, driven by expansion of an NDM-1 plus OXA-163-producing Providencia stuartii clone (Macchi et al., 2017).
Here, we report the emergence of closely related NDM-producing K. pneumoniae clinical isolates that initially exhibited resistance to all available antibiotics, including aztreonam-avibactam and subsequently evolved to resistance to both, cefiderocol and cefepime-zidebactam. Whole-genome sequencing (WGS) revealed the presence of multiple β-lactamases and other critical mutations in the resistome. This resistance phenotype led us to a successful compassionate treatment using maximum doses of aztreonam, ceftazidime-avibactam, and amoxicillin-clavulanate. These isolates demonstrated notable resistance to aztreonam-avibactam, which we attribute to the novel ESBL, SHV-231, responsible of a reduced susceptibility to avibactam at the expense of a higher susceptibility to clavulanate. This shows that the identification of the resistance mechanisms helps dealing with challenging PDR infections by resorting to alternative compassionate therapies.
1.1 Clinical case presentation
Five cases of PDR K. pneumoniae were identified in a single institution in Buenos Aires City between August 2022 and January 2023. All patients had undergone kidney transplants.
Patient 1: a 38-year-old male with type 1 diabetes developed a urinary tract infection (UTI) from NDM-producing K. pneumoniae after a renal-pancreatic transplant. Initial treatment with ceftazidime-avibactam and aztreonam was effective, but an NDM-producing K. pneumoniae strain with a PDR phenotype (Kpn-1-M28162) appeared in August 2022, requiring further medical attention. Compassionate treatment with ceftazidime-avibactam and aztreonam infusion, and oral amoxicillin-clavulanate (7 days) led to improved symptoms (the rationale for this triple antibiotic scheme is explained in further sections).
Patient 2: a 38-year-old male developed a UTI from an MBL-producing K. pneumoniae strain (PDR phenotype, Kpn-2-M28195) shortly after kidney transplant. Treatment with ceftazidime-avibactam and aztreonam infusion, and oral amoxicillin-clavulanate led to improved symptoms. In February 2023, gastrointestinal colonization by the same PDR germ was confirmed.
Patient 3: a 54-year-old male with chronic renal failure received a kidney transplant in August 2022. Recurrent UTIs following transplantation, including an episode in November caused by an MBL-producing K. pneumoniae strain (PDR phenotype, Kpn-3-M28196), was successfully managed with ceftazidime-avibactam and aztreonam. In December, a new UTI episode due to PDR K. pneumoniae isolate was compassionate treated with ceftazidime-avibactam and aztreonam infusion, and oral amoxicillin-clavulanate for 7 days that led to improved symptoms. Subsequent urine cultures were negative, and the patient showed favorable progress, leading to discharge.
Patient 4: a 42-year-old male with end-stage renal disease and on hemodialysis since 2016 faced multiple UTIs after a kidney transplant between April and September 2022 due to MBL-producing K. pneumoniae, treated with ceftazidime-avibactam and aztreonam or colistin plus rifampin. UTIs recurred in September, October, and November 2022 (PDR phenotype Kpn-4-M28206). The patient received compassionate treatment with ceftazidime-avibactam and aztreonam, and oral amoxicillin-clavulanate for 10 days in October and 4 weeks in December. Throughout, the patient remained afebrile with stable renal function, asymptomatic, and was ultimately discharged.
Patient 5: a 34-year-old female with type 1 diabetes and long-term hemodialysis had a combined kidney-pancreas transplant in late 2022. In February 2023, she presented with a UTI caused by a PDR K. pneumoniae strain (Kpn-5-M28413). Treatment was initiated with ceftazidime-avibactam and aztreonam, and amoxicillin-clavulanate and resulted in clinical cure. Her condition further deteriorated due to surgical complications, developing candidemia, coagulopathy, and an upper gastrointestinal hemorrhage, ultimately leading to her death in late February 2023.
2 Materials and methods
2.1 Bacterial identification and susceptibility testing
Bacterial identification was performed by MALDI-TOF (Bruker, United States) (Clinical and Laboratory Standards Institute, 2017). Susceptibility testing was conducted through agar dilution, disc diffusion and broth microdilution (BMD) using either commercial panels (Sensititre, Thermo-Fisher) or an in-house BMD method, employing either cation-adjusted Mueller-Hinton (CAMHB, Difco, United States) or iron-depleted CAMHB for cefiderocol (Clinical and Laboratory Standards Institute, 2024a; Clinical and Laboratory Standards Institute, 2024b). The panel of agents tested included: ampicillin, cefazolin, cefotaxime, ceftazidime, cefepime, aztreonam, cefoxitin, ertapenem, meropenem, imipenem, amoxicillin-clavulanate, ampicillin-sulbactam, piperacillin-tazobactam, ceftolozane-tazobactam, ceftazidime-avibactam, aztreonam-ceftazidime-avibactam, aztreonam-avibactam, aztreonam-relebactam, imipenem-relebactam, cefepime-zidebactam, gentamicin, amikacin, ciprofloxacin, trimethoprim-sulfamethoxazole, minocycline, tigecycline, chloramphenicol, rifampin, nitrofurantoin, fosfomycin, colistin and cefiderocol.
MICs were interpreted based on the latest CLSI criteria, except for aztreonam-avibactam, colistin and fosfomycin, which followed EUCAST guidelines, and tigecycline, interpreted with FDA standards (U.S. Food and Drug Administration, 2025; Clinical and Laboratory Standards Institute, 2024c; European Committee on Antimicrobial Susceptibility Testing, 2024).
To assess pertinent resistance mechanisms, susceptibility tests were conducted for selected agents by supplementing agar dilution plates with dipicolinic acid (Sigma, United States) at a final concentration of 1,000 μM, serving as an MBL inhibitor (Bahr et al., 2021). Aztreonam potentiation tests involved the same procedure by agar dilution, testing aztreonam alone or with avibactam, relebactam, clavulanic acid, and/or rifampin at a fixed concentration of 4 mg/L, and/or ceftazidime, ertapenem and fosfomycin at a fixed concentration of 10 mg/L, aligned with concentrations defined by PK/PD (Majumdar et al., 2002; Rodríguez-Gascón and Canut-Blasco, 2019; Clinical and Laboratory Standards Institute, 2024c). Zidebactam combinations were tested in a fixed ratio of 1:1 (Clinical and Laboratory Standards Institute, 2024c). Potentiation was defined as a 2-fold decrease in the MIC value of aztreonam in the presence of the inhibitor and/or accompanying antimicrobial agent.
2.2 Molecular characterization of the isolates
Initial characterization of resistance mechanisms was carried out using multiplex PCR to detect the most prevalent carbapenemase genes, including blaKPC, blaNDM, blaVIM, blaIMP, and blaOXA-48-like. Additional analyses included multiplex PCR for ESBLs (blaCTX-M, blaPER) and plasmid-mediated AmpC β-lactamases (blaCMY) (Faccone et al., 2023).
2.3 Conjugation assays
Biparental conjugation assays were conducted on solid media using Kpn-2 and Kpn-4 aztreonam-avibactam-resistant isolates. Escherichia coli J53 (azide-resistant and susceptible to aztreonam-avibactam) served as the recipient strain. Donor and recipient strains were mixed in a 3:1 ratio on tryptic soy agar plates and incubated at 35°C for 18 h. The conjugation mix was then resuspended in 1 mL of saline, and transconjugants were selected on tryptic soy agar plates containing 200 mg/L azide and 1 mg/L aztreonam plus 4 mg/L avibactam.
2.4 WGS and bioinformatic tools
Genomic DNA was extracted using the QIACube connect system with QIAamp DNA Blood Mini Kit (Qiagen, Germantown, MD, United States). Short-read sequencing was performed using the Nextera XT DNA library preparation kit and the Illumina MiSeq technologies (Illumina, San Diego, CA, United States) to generate 250 bp paired-end reads performed at the Unidad Operativa Centro Nacional de Genómica y Bioinformática ANLIS Malbrán. Quality control and trimming of the reads were performed by FastQC v0.11.5 and Trim Galore v0.6.3, respectively. Reads were de novo assembled using Unicycler v0.4.8-beta and taxonomy was confirmed with Kraken2 v2.1.2. AMRfinderPlus v3.11.17 was used to identify resistance genes and Prokka v.1.14 for annotation (Seemann, 2014; Wick et al., 2017; Feldgarden et al., 2020). Additionally, other bioinformatics tools were used to search for or compare specific genes, genomic regions, or amino acid sequences, like BLASTn and UniProt (Bateman et al., 2021). The confirmation of nucleotide sequences for blaSHV, blaNDM, and blaKPC was conducted through in-house PCR using Sanger technology ABI PRISM 3100 (Applied Biosystems, United States). The PCR reaction was assembled with 0.2 mM of dNTPs, 1.5 mM of MgCl2, 1X buffer, and 1.5 U of Taq polymerase (Invitrogen, United States) in a final volume of 50 µL. The primer concentration for blaSHV, blaNDM, and blaKPC was set at 0.2 µM. The specific primers utilized were as follows: SHV-Fa [5′-GCCCGGTTATTCTTATTTGTCGC-3′] and SHV-Ra [5′-TCTTTCCGATGCCGCCGCCAGTCA] for blaSHV, NDMin-F [5′-CTATTTACTAGGCCTCGCATT] and NDMin-R [5′-ATAAAACGCCTCTGTCACAT] for blaNDM, and KPC-F [5′-AACAAGGAATATCGTTGATG-3′] and KPC-R [5′-AGATGATTTTCAGAGCCTTA-3′] for blaKPC. These primer sets were designed to yield amplicons of 1,016, 936, and 916 bp, respectively.
2.5 Phylogenetic analysis
The genetic relationships among isolates were assessed by constructing a pairwise single nucleotide polymorphism (SNP) distance matrix and a phylogenetic tree based on a SNPs core genome alignment. The core genome alignment was first generated using Roary v3.12.0, concatenating all genes present in 99% of the samples. Subsequently, snp-sites v2.5.1 was used to create a core genome alignment consisting solely of SNPs, and snp-dist v0.8.2 was employed to generate the SNP difference matrix. Prevouisly reported SNP cutoffs (less than 21 SNPs) was used to define hospital clusters (David et al., 2019). A maximum likelihood phylogenetic tree was generated using RAxML v8.2.11, employing the GTR model with gamma-distributed site heterogeneity (GTRGAMMA) and 1,000 bootstrap replicates, based on the SNPs core alignment. The resulting tree was visualized using MEGA v11.0.13. Reference isolates include the following NCBI bio projects: NZ_JARERZ010000001, CP006923, CP006918, GCF_000316245.2, AUSMDU00008079, GCF_000599925.1 (Deleo et al., 2014; Venturini et al., 2019). Core genome MLST (cgMLST) analysis was also studied with the PathogenWatch tool based on the allelic profile of 629 conserved genes.
2.6 Cloning of blaSHV allelic variants and susceptibility testing of recombinant clones
The blaSHV variants were engineered by incorporating a Strep Tag sequence at the C-terminal end for immunodetection and cloned into the pMBLe vector, enabling protein expression through IPTG-controlled. E. coli DH5α cells were transformed with these vectors using the chemical method and cells were selected with 50 mg/L gentamicin. PCR amplification and Sanger sequencing confirmed the integrity of the synthesized genes. The susceptibility of E. coli expressing various blaSHV alleles was tested in 100 µM IPTG by measuring the MICs of different antibiotics (González et al., 2016).
2.7 Structural modeling of PBP3 mutation
To visualize the potential structural impact of the A413V substitution in PBP3, we performed in silico analysis using the PyMOL Molecular Graphics System, Version 3.0 (Schrödinger, LLC). The wild-type and mutant amino acid sequences were modeled based on available structural templates, and the A413V mutation was introduced manually.
2.8 PDR definition
The strains were classified as PDR according to the consensus definition established by Latin American countries in coordination with Pan American Health Organization (PAHO/WHO), which defines PDR in Enterobacterales as phenotypic resistance to penicillins, cephalosporins, monobactams, carbapenems, legacy β-lactam/β-lactamase inhibitor combinations, aminoglycosides, fosfomycin, lipopeptides, tetracyclines/glycylcyclines, quinolones and folate pathway inhibitors (Jiménez Pearson et al., 2019). This definition was subsequently expanded at the national level by the Argentine Ministry of Health to include newer antimicrobials incorporated into the national formulary, such as ceftolozane-tazobactam, ceftazidime-avibactam, aztreonam-avibactam, and imipenem-relebactam (https://www.argentina.gob.ar/sites/default/files/2021/08/instructivo_pandrogoresistencia-16092024.pdf). As cefiderocol is not currently listed in the national vademecum, it was initially excluded from the local definition of PDR.
3 Results
3.1 Identification of PDR K. pneumoniae and antimicrobial susceptibility
The five isolates were identified as K. pneumoniae by MALDI-TOF. MIC determination confirmed the PDR phenotype in all isolates, including, ceftazidime-avibactam (MIC, >256 mg/L), aztreonam-avibactam or aztreonam-relebactam (32 - >256 mg/L, see details in other section), imipenem-relebactam (>32 mg/L), tigecycline (>2 mg/L), fosfomycin (>64 mg/L), and colistin (>8 mg/L). Susceptibility to cefiderocol was demonstrated in Kpn-1 to Kpn-4 strains (MICs 1–2 mg/L), on the contrary Kpn-5 rendered resistance (>8 mg/L). Cefepime-zidebactam demonstrated promising activity against Kpn-1 and Kpn-2 (8 mg/L). However, Kpn-3 and Kpn-4 exhibited a marked increase in MIC values (32 mg/L), and Kpn-5 showed a high level of resistance to this combination (>64 mg/L). The carbapenem phenotype was abolished by the metal chelator dipicolinic acid, revealing the presence of an MBL, except for Kpn-3, indicating additional mechanisms.
3.2 PCR and WGS analysis
Initial characterization of resistance mechanisms by PCR revealed the presence of blaNDM and blaCTX-M in all isolates, while blaKPC was detected exclusively in Kpn-3.
WGS confirmed the taxonomic identity of the K. pneumoniae isolates (5,431,038–5,542,594 bp), classified as clonal group 258 (CG258) with capsular type K107/O1/O2V2 (wzi 154). The isolates were genetically related with 31–89 SNPs between them (Table 1). This polymorphism exceeded the threshold typically used to define outbreak clusters, suggesting the circulation and local evolution of a closely-related strains within the institution. Phylogenetic analysis based on maximum likelihood revealed that the isolates belong to Clade II of the ST258 lineage (Supplementary Figure S1). According to cgMLST, all five isolates were close to core genome sequence type 562. No virulence loci, including rmpADC/rmpA2, were detected in any isolate.
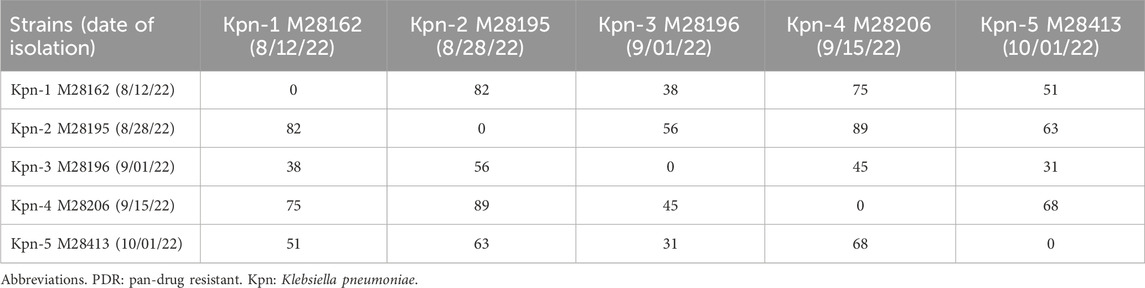
Table 1. Number of core genome single nucleotide polymorphisms between K. pneumoniae PDR isolates.
WGS confirmed the presence of blaNDM-5 and blaCTX-M-15 in all strains, and blaKPC-2 in Kpn-3 (Figure 1). A novel allelic variant of the blaSHV gene (blaSHV-231) was present in all isolates (Accession No. OP951208). blaSHV-231 exhibited two non-synonymous substitutions: K230R and A233G compared to blaSHV-5 (Figure 2), and its upstream promoter sequence harbored a single-nucleotide change in the +10 region. In Kpn-1, blaSHV-231 was located on a 150 kb chromosomal contig, as indicated by its genomic context, which included housekeeping and regulatory genes such as ftrA, duf4177, aac, lacI, lacZ, emrB, ermD, glpR, adh, fruK, fbaA, ptsH, and gntP. In the remaining strains, blaSHV-231 was found on shorter contigs (3,214–3,995 bp), whose gene surroundings similarly support a chromosomal location. Resistance to aztreonam-avibactam was not transferable in biparental conjugation assays after 48 h of incubation, supporting the chromosomal nature of this resistance determinant. Two isolates (Kpn-2 and Kpn-5) harbored a chromosomal gene duplication event, carrying both, blaSHV-231 and the ancestral blaSHV-1 allele.
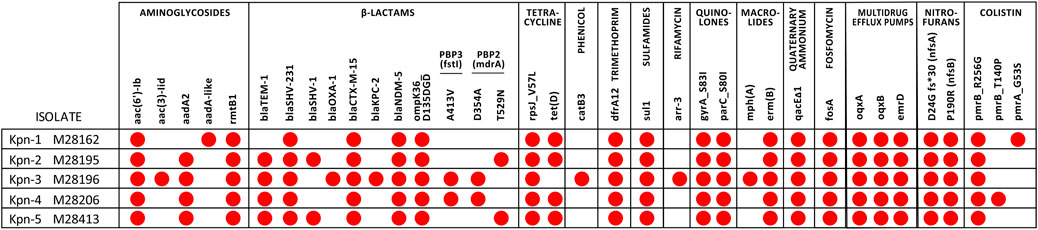
Figure 1. Resistance genes and/or chromosomal mutations in pan-drug resistant isolates obtained by WGS. Red dots indicate the presence of the indicated gene or mutation.

Figure 2. Alignment of the amino acid sequences of selected SHV variants. Conserved residues of the ß-lactamase are denoted in blue while mutations are highlighted in white.
All isolates possessed IncFIB(pQil), ColRNAI, and Col (pHAD28) plasmid replicons. IncFII was present in all strains except Kpn-1, and IncM1 was exclusive to Kpn-3. IncFIB(K) was found in Kpn-2 and Kpn-3. Although blaKPC-2 was not co-located with a plasmid replicon in the assembly, the exclusive presence of an IncM1 plasmid in Kpn-3—currently associated with blaKPC dissemination in Argentina—suggests a plasmid-borne origin.
All isolates contained multiple resistance genes spanning various antibiotic families. Notably, modifications in outer membrane porins, such as OmpK36/OmpC, and the colistin-associated pmrB gene mutation (R256G and T246A) were present (Figure 1). The isolates displayed no difference in iron metabolism cirA, fepA, fhuA, iroN, btuB, ExbB, ExbD, TonB genes. Seven unique mutations, including one in rnpA, an RNA degradation protein, were found in the cefiderocol-resistant isolate Kpn-5.
Core genome analysis revealed polymorphisms in PBP3 (A413V) for Kpn-3 and Kpn-4 and PBP2 (D354A in Kpn-3 and Kpn-4, T529N in Kpn-2 and Kpn-5). The A413 V mutation in PBP3 is predicted to alter the structural loop adjacent to the active site, potentially diminishing the binding affinity of aztreonam (The PyMOL Molecular Graphics System, Version 3.0 Schrödinger, LLC). No evidence of the amino acid insertions in the β-lactam binding pocket of PBP3 commonly associated with impaired affinity for aztreonam were observed (Periasamy et al., 2020; Sato et al., 2020).
3.3 Evaluation of alternatives treatment regimens: synergistic combinations for PDR infections
Due to the lack of effective standard therapies, alternative regimens were investigated to provide compassionate-use treatments for the affected patients. Aztreonam paired with avibactam, relebactam, or clavulanic acid showed limited effectiveness in isolates Kpn-1, Kpn-2, and Kpn-5 (MICs 32–64 mg/L), while Kpn-3 and Kpn-4 maintained resistance at MICs >256 mg/L (Figure 3). The rationale for drug selection in this compassionate treatment was based on a strategic combination of β-lactamase inhibitors with aztreonam to overcome complex resistance mechanisms. Avibactam was employed primarily to inhibit additional β-lactamases known to compromise the activity of monobactams, such as blaCTX-M (present in Kpn-1 to Kpn-5) and blaKPC (identified in Kpn-3). To address the presence of an ESBL variant with mutations suggesting an inhibitor resistance profile, such as blaSHV-231, a range of combinations were evaluated—including both legacy and newer-generation β-lactamase inhibitors, as well as non-classical options like rifampin, fosfomycin or colistin, which may enhance aztreonam activity through synergistic effects. The combination of clavulanate and avibactam outperformed other combinations with aztreonam, including dual β-lactam, fosfomycin, rifampicin, or colistin therapies. The combination of these two inhibitors lowered aztreonam MICs to 1–8 mg/L for all isolates, except Kpn-2 and Kpn-3 (MICs 32 mg/L), and resulted in successful treatment (Figure 3).
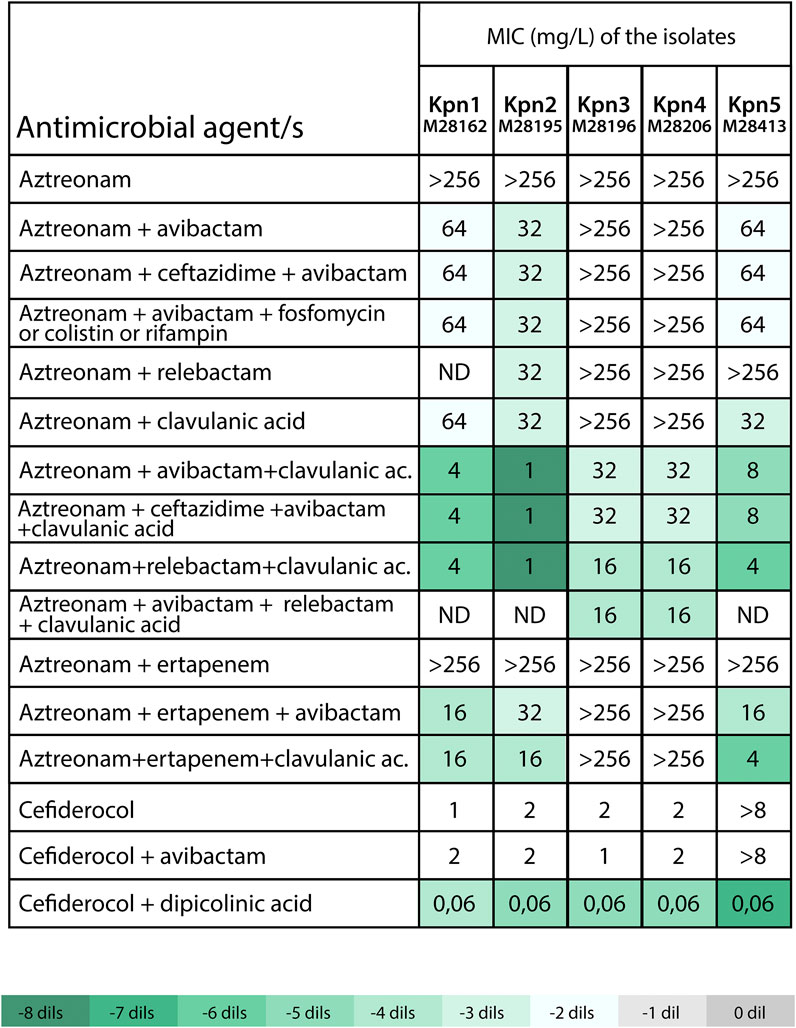
Figure 3. Susceptibility to selected antimicrobial agents alone or in combinations against PDR K. pneumoniae isolates. Avibactam, relebactam, clavulanic acid and rifampin were tested at fixed concentration of 4 mg/L, ceftazidime at fixed concentration of 8 mg/L, ertapenem and fosfomycin at fixed concentration of 10 mg/L and dipicolinic acid at fixed concentration of 1,000 µM. ND: not determined. Abbreviations. XDR: pan-drug resistant. Kpn: Klebsiella pneumoniae. The color-coded scale in the table indicates the log2 (dils.) difference between MICs for β-lactam with the inhibitor in relation to MICs for β-lactam alone (aztreonam or cefiderocol) for each assessed clinical strain, following the provided reference in Table.
Kpn-5 was the only isolate resistant to cefiderocol. Interestingly, in vitro treatment with dipicolinic acid restored susceptibility to cefiderocol, highlighting the role of NDM-5 in this resistance phenotype. To further investigate the mechanism underlying this resistance, we conducted an in silico analysis of the blaNDM-5 promoter region, alongside a comparative analysis of the relative gene copies of the blaNDM-5 contig versus the constitutive gene gyrA. The promoter region of blaNDM-5 was found to be conserved and identical to previously reported sequences (Li et al., 2022). However, gene copies analysis revealed a stark difference between blaNDM-5 and other isolates, with the copies of blaNDM-5 in Kpn-5 being significantly higher (5.0 vs. 0.3–1.06 in other isolates) (Supplementary Table S1). This finding reinforces the hypothesis that overexpression of blaNDM-5 is a key driver of cefiderocol resistance in the absence of mutations in the previously mentioned iron transporter genes. This overexpression is also likely responsible for the high level of resistance to cefepime-zidebactam observed in Kpn-5.
3.4 SHV-231 β-lactamase is responsible for the decreased susceptibility to aztreonam-avibactam
Since aztreonam resistance has been also associated with the co-production of other ESBLs (Mendes et al., 2021; Findlay et al., 2023) we assessed the potential impact of the blaSHV-231 variant on the inhibitor resistance phenotype. In follow-up experiments to validate our hypothesis and the clinical response, we cloned the blaSHV-231 gene into an expression vector in E. coli, as well as different combinations of the substitutions present in blaSHV-231 to dissect their contributions to the resistance phenotype and determined the susceptibility profile of the different variants (Figure 4). blaSHV-231 carries the two non-synonymous mutations characteristic of blaSHV-5 (G234S and E235K) responsible of an extended-spectrum phenotype (Billot-Klein et al., 1990). The alteration at K230 stands out because is a highly conserved residue (Dubois et al., 2008; Papp-Wallace et al., 2015). The K230R substitution (blaSHV-84) not only reduced susceptibility to clavulanic acid, as previously reported (Manageiro et al., 2010) but presented an expanded inhibitor resistance profile that also included avibactam (Figure 4). Substitution A233G alone, instead, affected inhibition by clavulanic acid but not avibactam. The inclusion of K230R in blaSHV-5 made it more susceptible to aztreonam (MIC 1 mg/L vs. 8 mg/L for blaSHV-5), but did not affect ceftazidime resistance (MIC 8 mg/L). Instead, addition of both A233G and K230R into blaSHV-5 (i.e., blaSHV-231) led to a recovery of aztreonam resistance (MIC 8 mg/L) and a reduced susceptibility to inhibition by avibactam, with only a two-fold decrease in aztreonam-avibactam MIC compared to a six-fold decrease for blaSHV-5. Notably, the protective efficacy of all tested inhibitors for aztreonam diminished in blaSHV-231, despite avibactam was more adversely impacted than clavulanic acid. When both inhibitors were evaluated together, the MIC of aztreonam against blaSHV-231 reached levels comparable to those observed for the blaSHV-5 allele (Figure 4).
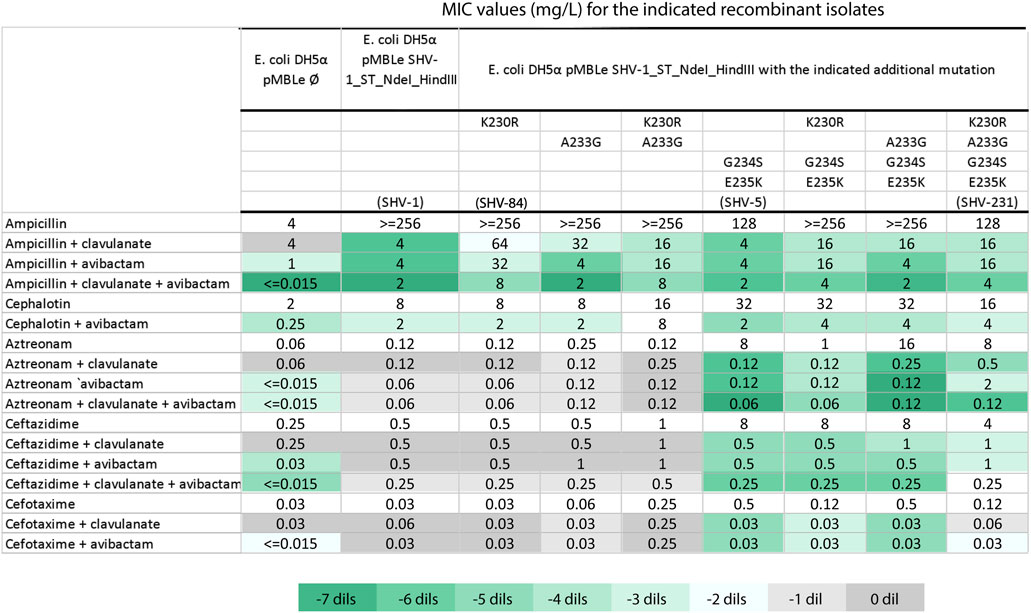
Figure 4. Susceptibility testing of recombinant E. coli strains Minimum Inhibitory Concentrations (MICs) were determined for E. coli DH5α recombinants carrying either the empty pMBL plasmid or blaSHV-1 with the specified mutations. The mutation combinations resulting in blaSHV-1, blaSHV-5, blaSHV-84 and blaSHV-231 are denoted in parentheses. Clavulanate and avibactam were held at a fixed final concentration of 4 mg/L. The color-coded scale in the table indicates the log2 difference (dils.) between MICs for β-lactam with the inhibitor in relation to MICs for β-lactam alone for each assessed recombinant strain, following the provided reference in Table.
4 Discussion
The increasing incidence of PDR Gram-negative bacilli poses a significant clinical challenge, particularly when involving immunocompromised patients (Zowawi et al., 2015; Avgoulea et al., 2018; Lázaro-Perona et al., 2018; Longo et al., 2019). Here we report the spread of PDR NDM-producing K. pneumoniae isolates in Buenos Aires. Since its detection in Argentina in 2014, NDM has become endemic, with the COVID-19 pandemic exacerbating resistance, particularly through dual carbapenemase producers (Pasteran et al., 2014; Faccone et al., 2023; Echegorry et al., 2024).
Despite in vitro susceptibility to cefiderocol in four of the five isolates, this antibiotic was not considered clinically actionable, as it is not available in Argentina. While the Latin American consensus definition of PDR (Jiménez Pearson et al., 2019) reflects regional realities, alternative frameworks—such as the European consensus by Magiorakos et al. (2012)—define PDR as non-susceptibility to all agents across all antimicrobial categories. In our view, relying solely on definitions that presume universal drug availability may inadvertently exclude the clinical realities faced by low and middle-income countries. A more inclusive and context-sensitive approach to defining PDR may be warranted, ideally through consensus processes that involve equitable participation of all member states.
All five clinical isolates were identified as belonging to Clade II of the high-risk clonal group CG258, a major K. pneumoniae lineage known for acquiring carbapenemase genes and contributing to global dissemination. Historically, Clade II has been closely associated with the spread of blaKPC-producing strains in North and South America (Chen et al., 2014; Lai et al., 2019). Supporting this, a recent multicenter study in Argentina reported K. pneumoniae as the predominant host of blaNDM, with 70% of cases linked to CG258 (Echegorry et al., 2024).
The precise origin of these PDR strains remains elusive. Several factors indicate the local emergence of PDR microorganisms at the institutional level as the index case had previously undergone treatment with ceftazidime-avibactam plus aztreonam in the months leading up to the recovery from the initial strain. Regrettably, surveillance practices, limited to carbapenemase detection, may have enabled the spread of these strains. Following identification of these strains displaying aztreonam-avibactam resistance, the ultimate line drug in Argentina, measures taken for infection control were reinforced, leading to containment of the PDR strains by the end of June 2023.
All five K. pneumoniae strains contained between 22 (Kpn-4) and 29 (Kpn-2) antibiotic resistance genes across different drug families. All isolates carried multiple β-lactamases, including blaNDM-5 and blaCTX-M-15, aminoglycoside methylases and other modifying enzymes and several core genome target mutations. Amino acid substitutions R256G and T246A in PmrB validated colistin resistance and have been linked with CG258 in South America (Longo et al., 2019). The V57I mutation in rpsJ, associated with tigecycline resistance outside the AcrAB-TolC system, is also linked to this clonal complex (Villa et al., 2014). The loss of major porin OpmK36 could have increased MICs to β-lactams and generated cross-resistance to other drugs (Tsai et al., 2011). This phenomenon has been frequently observed in the CG258, which multiples reports across South America (David et al., 2019).
4.1 Clinical management through compassionate treatment
In the absence of dedicated trials for treating PDR infections, clinical management relies on case reports and limited studies, often outside the renal transplant context (Karakonstantis et al., 2019; Sivasankar et al., 2023). Recognizing the urgent need for treatment alternatives, we exploited genomic insights to guide compassionate therapy. Given the abundance of resistance genes that confer resistance to conventional drugs, we focused on aztreonam by using locally available legacy and newer-generation β-lactamase inhibitors. Patients received treatments involving maximum doses of aztreonam, ceftazidime-avibactam, and oral amoxicillin-clavulanate, leading to clinical cure. Although the triple regimen exhibited higher MIC values, therapeutic levels in the urine were achievable (Ramsey and MacGowan, 2016). However, one patient required an extended treatment course to prevent relapse.
4.2 Genomic path to pan-β-resistance
The observed resistance to aztreonam-avibactam stemmed from mutations in PBP2 and PBP3, and a novel SHV-231 β-lactamase. blaSHV-231 had two nonsynonymous mutations (K230 R and A233G) compared to blaSHV-5 (Billot-Klein et al., 1990), maintaining the ESBL phenotype while reducing susceptibility to aztreonam-avibactam, a rare trait in this β-lactamase, potentiated by overexpression due to its promoter region. This evolutionary perspective was anticipated by Winkler and coworkers (Winkler et al., 2015). Using an E. coli expression model, we confirmed that the triple combination therapy reduced MIC values in clones harboring blaSHV-231, although the observed MICs were significantly lower than those detected in the corresponding clinical isolates. This discrepancy is likely attributable to the limited expression of SHV variants in the laboratory host strains. In the clinical isolates, WGS revealed that the blaSHV-231 was linked to a strong P-S promoter (where the C in the second position of the −10 region is substituted by an A). This mutation has been shown to elicit a 200-fold enhancement of β-lactamase expression, leading to a significant increase in MICs (Leung et al., 1997; Rice et al., 2000; Kamruzzaman et al., 2015). Therefore, we expect high expression levels of blaSHV-231 in all clinical isolates. Multiple alignments on contemporary blaSHV promoter-associated sequences of K. pneumoniae strains using the NCBI database, revealed that out of 1,000 cases reviewed, only 139 contained a P-S promoter. On this basis, we propose that the PDR isolates may have been subjected to substantial selection pressure—potentially driven by multiple prior treatment regimens—leading to the observed promoter mutation. We therefore attribute the pan-β-lactam resistant phenotype to the overexpression of blaSHV-231 combined with the presence of NDM.
Isolates Kpn-3 and Kpn-4 showed higher resistance to aztreonam-avibactam due to PBP3 and PBP2 mutations. PBP3 lacks the canonical YRIK, YRIM, and YRIP insertions, displaying only the A413V substitution, previously reported in combination with these insertions (Periasamy et al., 2020; Sato et al., 2020). Residue A413 is located within a conserved amino acid loop (positions 402–420) shared across multiple classes of PBPs and is positioned opposite the active site. The substitution of alanine with a bulkier valine—bearing an additional methyl group oriented toward the active site—may introduce steric hindrance that interferes with substrate binding, contributing to ceftazidime and aztreonam resistance (Sauvage et al., 2014; Zhang et al., 2017). PBP2 substitutions, particularly D354A, correlated with high resistance levels to ceftazidime-avibactam (Livermore et al., 2018), even after MBL inhibition, since MICs remained at 32 mg/L. This suggest a role of PBP2 as a secondary target for diazabicyclooctanes, although the effect of avibactam is weaker than analogs nacubactam and zidebactam (Morinaka et al., 2015; Livermore et al., 2017). This PBP2 alteration might also contribute to higher resistance to cefepime-zidebactam in these two isolates. In summary, we propose that PBP3 and PBP2 mutations with the synergistic effect of the overexpression of blaSHV-231, hindered the resensitization of aztreonam even after the introduction of multiple inhibitors in Kpn-3 and Kpn-4. These mutations can be attributed to prior exposure to aztreonam plus ceftazidime-avibactam of these patients. Moreover, patient #4 underwent two previous treatment regimens with this combination.
All five K. pneumoniae isolates shared core plasmid replicons—IncFIB(pQil), ColRNAI, and Col (pHAD28)—commonly found in high-risk, multidrug-resistant clones. IncFII, present in four isolates, aligns with its known role in spreading ESBL and carbapenemase genes, particularly in CG258 lineages (Rozwandowicz et al., 2018). IncM1, detected only in Kpn-3, has been recently linked to carbapenemase mobilization, while IncFIB(K) in Kpn-2 and Kpn-3 may contribute to the convergence of resistance and virulence traits (Martin et al., 2023). These results highlight a conserved plasmid backbone with additional isolate-specific replicons, reflecting local evolution and adaptation.
Four initial isolates displayed susceptibility to cefiderocol, while the latest isolate (Kpn-5) acquired resistance by overexpression of blaNDM-5 without previous treatment with this drug. This finding reinforces the hypothesis that overexpression of NDM-5 is a key driver of cefiderocol resistance in the absence of mutations in the previously mentioned iron transporter genes (Simner et al., 2023). This overexpression is also likely responsible for the high level of resistance to cefepime-zidebactam observed in Kpn-5.
The emergence of pan-β-lactam resistance poses severe clinical concerns. Most recently approved drugs for Gram-negative bacilli are β-lactams, except eravacycline and plazomicin (World Health Organization, 2024), making this resistance particularly alarming. Of greater concern, we note that these isolates present resistance to not-yet-approved drugs, like cefepime-zidebactam, further limiting the therapeutic options against NDM-producing strains. These cases demonstrate that personalized combination therapies can effectively manage PDR infections.
5 Concluding remarks
The pan-β-lactam-resistant phenotype observed in five clinical isolates highlights the adaptability of K. pneumoniae to the evolutionary pressures in clinical environments. The emergence of a new SHV variant gaining legacy and newer-generation β-lactamase inhibitors resistance while preserving ESBL activity, enhanced by mutations in PBPs provides a picture of this adaptation. This evolutionary convergence occurs in a CG258 clone, capable of withstanding selective pressures and forging new evolutionary pathways. This genomic adaptability emphasizes the need of comprehensive genomic surveillance to curb the spread high-risk clones. This study emphasizes the importance of timely genomic and phenotypic characterization of locally circulating pathogens to develop more tailored therapeutic strategies.
Data availability statement
The datasets generated for this study can be found in the GenBank database under Bioproject PRJNA1062347 (https://www.ncbi.nlm.nih.gov/bioproject/?term=PRJNA1062347) and the following accession numbers (BioSample Accession): Kpn-1-M28162, JAYMDN000000000 (SAMN39296142); Kpn-2-M28195, SAMN39296143 (JAYMDO000000000); Kpn-3-M28196, SAMN39296144 (JAYMDP000000000); Kpn-4-M28206, SAMN39296145 (JAYMDQ000000000); Kpn-5-M28413, SAMN39296146 (JAYMDR000000000).
Ethics statement
Ethical approval was not required for the study involving humans in accordance with the local legislation and institutional requirements. Written informed consent to participate in this study was not required from the participants or the participants’ legal guardians/next of kin in accordance with the national legislation and the institutional requirements. Written informed consent was not obtained from the individual(s) for the publication of any potentially identifiable images or data included in this article because retrospective, anonymized, non-interventional study.
Author contributions
FP: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review and editing. JM: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Software, Validation, Visualization, Writing – original draft, Writing – review and editing. NP: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Validation, Visualization, Writing – review and editing. GD: Conceptualization, Data curation, Formal Analysis, Investigation, Methodology, Validation, Visualization, Writing – review and editing. LG: Conceptualization, Formal Analysis, Investigation, Supervision, Validation, Writing – original draft, Writing – review and editing. MR: Formal Analysis, Investigation, Methodology, Visualization, Writing – review and editing. AF: Formal Analysis, Investigation, Methodology, Visualization, Writing – review and editing. PC: Investigation, Supervision, Validation, Visualization, Writing – review and editing. LM: Formal Analysis, Investigation, Methodology, Visualization, Writing – review and editing. MR: Formal Analysis, Investigation, Methodology, Visualization, Writing – review and editing. SG: Formal Analysis, Supervision, Validation, Visualization, Writing – original draft, Writing – review and editing. CL: Formal Analysis, Investigation, Methodology, Visualization, Writing – review and editing. MM: Investigation, Methodology, Visualization, Writing – review and editing. EA: Investigation, Methodology, Visualization, Writing – review and editing. DD: Formal Analysis, Methodology, Validation, Writing – review and editing. MR: Conceptualization, Formal Analysis, Supervision, Validation, Visualization, Writing – review and editing. AV: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review and editing. AC: Conceptualization, Data curation, Formal Analysis, Funding acquisition, Investigation, Methodology, Project administration, Resources, Software, Supervision, Validation, Visualization, Writing – original draft, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. Work at the NRL was supported by regular federal budget at ANLIS “Dr. C. Malbrán”. Research at IBR was supported by grants from Agencia I+D+I (PICT-2020-00031 to A.J.V.) and MinCyT (REPARA to A.J.V.).
Acknowledgments
We dedicate this work to the memory of our dear colleague and friend Gina Dotta, whose support and inspiration continue to guide us.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The authors declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2025.1570278/full#supplementary-material
Abbreviations
XDR, extensively drug-resistant (XDR); MBL, metallo-β-lactamase; ESBL, extended-spectrum β-lactamases; PBP, penicillin-binding protein; WGS, Whole-genome sequencing; UTI, urinary tract infection; CG, clonal group.
References
Avgoulea, K., Di Pilato, V., Zarkotou, O., Sennati, S., Politi, L., Cannatelli, A., et al. (2018). Characterization of extensively drug-resistant or pandrug-resistant sequence type 147 and 101 OXA-48-producing Klebsiella pneumoniae causing bloodstream infections in patients in an intensive care unit. Antimicrob. Agents Chemother. 62. doi:10.1128/AAC.02457-17
Bahr, G., González, L. J., and Vila, A. J. (2021). Metallo-β-lactamases in the age of multidrug resistance: from structure and mechanism to evolution, dissemination, and inhibitor design. Chem. Rev. 121, 7957–8094. doi:10.1021/acs.chemrev.1c00138
Bateman, A., Martin, M.-J., Orchard, S., Magrane, M., Agivetova, R., Ahmad, S., et al. (2021). UniProt: the universal protein knowledgebase in 2021. Nucleic Acids Res. 49, D480–D489. doi:10.1093/nar/gkaa1100
Billot-Klein, D., Gutmann, L., and Collatz, E. (1990). Nucleotide sequence of the SHV-5 beta-lactamase gene of a Klebsiella pneumoniae plasmid. Antimicrob. Agents Chemother. 34, 2439–2441. doi:10.1128/AAC.34.12.2439
Bonomo, R. A., Perez, F., Hujer, A. M., Hujer, K. M., and Vila, A. J. (2024). The real crisis in antimicrobial resistance: failure to anticipate and respond. Clin. Infect. Dis. 78 (6), 1429–1433. doi:10.1093/cid/ciad758
Chen, L., Mathema, B., Pitout, J. D., DeLeo, F. R., and Kreiswirth, B. N. (2014). Epidemic Klebsiella pneumoniae ST258 is a hybrid strain. mBio 24 (3), e01355–14. doi:10.1128/mBio.01355-14
Clinical and Laboratory Standards Institute (2024c). M100-S34 Performance standards for antimicrobial susceptibility testing thirty-four informational supplement. Wayne, PA: Clinical and Laboratory Standards Institute.
Clinical and Laboratory Standards Institute (2017). M58 - methods for the identification of cultured microorganisms using matrix-assisted laser desorption/ionization time-of-flight mass spectrometry. 1st Edition. Wayne, PA: Clinical and Laboratory Standards Institute.
Clinical and Laboratory Standards Institute (2024a). M02 - performance standards for antimicrobial disk susceptibility tests. 14th Edition. Wayne, PA: Clinical and Laboratory Standards Institute.
Clinical and Laboratory Standards Institute (2024b). M07 - clinical and Laboratory Standards Institute. Methods for dilution antimicrobial susceptibility tests for bacteria that grow aerobically M07. 12th Edition. Wayne, PA: Clinical and Laboratory Standards Institute.
David, S., Reuter, S., Harris, S. R., Glasner, C., Feltwell, T., Argimon, S., et al. (2019). Epidemic of carbapenem-resistant Klebsiella pneumoniae in Europe is driven by nosocomial spread. Nat. Microbiol. 4, 1919–1929. doi:10.1038/s41564-019-0492-8
Deleo, F. R., Chen, L., Porcella, S. F., Martens, C. A., Kobayashi, S. D., Porter, A. R., et al. (2014). Molecular dissection of the evolution of carbapenem-resistant multilocus sequence type 258 Klebsiella pneumoniae. Proc. Natl. Acad. Sci. U. S. A. 1 (111), 4988–4993. doi:10.1073/pnas.1321364111
Dubois, V., Poirel, L., Demarthe, F., Arpin, C., Coulange, L., Minarini, L. A. R., et al. (2008). Molecular and biochemical characterization of SHV-56, a novel inhibitor-resistant β-lactamase from Klebsiella pneumoniae. Antimicrob. Agents Chemother. 52, 3792–3794. doi:10.1128/AAC.00387-08
Echegorry, M., Marchetti, P., Sanchez, C., Olivieri, L., Faccone, D., Martino, F., et al. (2024). National multicenter study on the prevalence of carbapenemase-producing enterobacteriaceae in the post-COVID-19 era in Argentina: the RECAPT-AR study. Antibiot. (Basel) 27 (12), 1139. doi:10.3390/antibiotics13121139
European Committee on Antimicrobial Susceptibility Testing (EUCAST) (2024). Clinical breakpoints - breakpoints and guidance, v14.
Faccone, D., Gomez, S. A., de Mendieta, J. M., Sanz, M. B., Echegorry, M., Albornoz, E., et al. (2023). Emergence of hyper-epidemic clones of Enterobacterales clinical isolates Co-producing KPC and metallo-beta-lactamases during the COVID-19 pandemic. Pathogens 12, 479. doi:10.3390/pathogens12030479
Feldgarden, M., Brover, V., Haft, D. H., Prasad, A. B., Slotta, D. J., Tolstoy, I., et al. (2020). Validating the AMRFinder tool and resistance gene database by using antimicrobial resistance genotype-phenotype correlations in a collection of isolates. Antimicrob. Agents Chemother. 22, e00483–19. doi:10.1128/AAC.00483-19
Findlay, J., Poirel, L., Bouvier, M., Gaia, V., and Nordmann, P. (2023). Resistance to ceftazidime-avibactam in a KPC-2–producing Klebsiella pneumoniae caused by the extended-spectrum beta-lactamase VEB-25. Eur. J. Clin. Microbiol. Infect. Dis. 42, 639–644. doi:10.1007/s10096-023-04582-0
Georgopapadakou, N. H., Smith, S. A., and Sykes, R. B. (1982). Mode of action of azthreonam. Antimicrob. Agents Chemother. 21, 950–956. doi:10.1128/AAC.21.6.950
González, L. J., Bahr, G., Nakashige, T. G., Nolan, E. M., Bonomo, R. A., and Vila, A. J. (2016). Membrane anchoring stabilizes and favors secretion of New Delhi metallo-β-lactamase. Nat. Chem. Biol. 12, 516–522. doi:10.1038/nchembio.2083
Jiménez-Pearson, M. A., Galas, M., Corso, A., Hormazábal, J. C., Duarte-Valderrama, C., Salgado-Marcano, N., et al. (2019). Latin American consensus to define, categorize, and report multidrug-resistant, extensively drug-resistant, or pandrug-resistant pathogens. Rev. Panam. Salud Publica 22, 43. doi:10.26633/RPSP.2019.65
Kamruzzaman, M., Patterson, J. D., Shoma, S., Ginn, A. N., Partridge, S. R., and Iredell, J. R. (2015). Relative strengths of promoters provided by common mobile genetic elements associated with resistance gene expression in gram-negative bacteria. Antimicrob. Agents Chemother. 59, 5088–5091. doi:10.1128/AAC.00420-15
Karakonstantis, S., Kritsotakis, E. I., and Gikas, A. (2019). Pandrug-resistant Gram-negative bacteria: a systematic review of current epidemiology, prognosis and treatment options. J. Antimicrob. Chemother. 75, 271–282. doi:10.1093/jac/dkz401
Lai, Y.-C., Lu, M.-C., and Hsueh, P.-R. (2019). Hypervirulence and carbapenem resistance: two distinct evolutionary directions that led high-risk Klebsiella pneumoniae clones to epidemic success. Expert Rev. Mol. diagn. 19, 825–837. doi:10.1080/14737159.2019.1649145
Lázaro-Perona, F., Sotillo, A., Troyano-Hernáez, P., Gómez-Gil, R., de la Vega-Bueno, Á., and Mingorance, J. (2018). Genomic path to pandrug resistance in a clinical isolate of Klebsiella pneumoniae. Int. J. Antimicrob. Agents 52, 713–718. doi:10.1016/j.ijantimicag.2018.08.012
Leung, M., Shannon, K., and French, G. (1997). Rarity of transferable beta-lactamase production by Klebsiella species. J. Antimicrob. Chemother. 39, 737–745. doi:10.1093/jac/39.6.737
Li, Y., Zhang, R., and Wang, S. (2022). A natural novel mutation in the blaNDM-5 promoter reducing carbapenems resistance in a clinical Escherichia coli strain. Microbiolology Spectr. 10, e0118321. doi:10.1128/spectrum.01183-21
Livermore, D. M., Mushtaq, S., Doumith, M., Jamrozy, D., Nichols, W. W., and Woodford, N. (2018). Selection of mutants with resistance or diminished susceptibility to ceftazidime/avibactam from ESBL- and AmpC-producing Enterobacteriaceae. J. Antimicrob. Chemother. 73, 3336–3345. doi:10.1093/jac/dky363
Livermore, D. M., Mushtaq, S., Warner, M., Vickers, A., and Woodford, N. (2017). In vitro activity of cefepime/zidebactam (WCK 5222) against Gram-negative bacteria. J. Antimicrob. Chemother. 72, 1373–1385. doi:10.1093/jac/dkw593
Longo, L. G. A., de Sousa, V. S., Kraychete, G. B., Justo-da-Silva, L. H., Rocha, J. A., Superti, S. V., et al. (2019). Colistin resistance emerges in pandrug-resistant Klebsiella pneumoniae epidemic clones in Rio de Janeiro, Brazil. Int. J. Antimicrob. Agents 54, 579–586. doi:10.1016/j.ijantimicag.2019.08.017
Macchi, A., Harispe, M. E., Pasteran, F., and Corso, A. (2017). The post-antibiotic era has arrived: interventions to contain an outbreak of pan-drug- resistant (PDR) NDM-1 and OXA-48-like dual producer Providencia stuartii (PST) in an acute care hospital in Argentina. Vienna: European Society of Clinical Microbiology and Infectious Disease.
Magiorakos, A.-P., Srinivasan, A., Carey, R. B., Carmeli, Y., Falagas, M. E., Giske, C. G., et al. (2012). Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: an international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 18, 268–281. doi:10.1111/j.1469-0691.2011.03570.x
Majumdar, A. K., Musson, D. G., Birk, K. L., Kitchen, C. J., Holland, S., McCrea, J., et al. (2002). Pharmacokinetics of ertapenem in healthy young volunteers. Antimicrob. Agents Chemother. 46, 3506–3511. doi:10.1128/AAC.46.11.3506-3511.2002
Manageiro, V., Ferreira, E., Albuquerque, L., Bonnet, R., and Canica, M. (2010). Biochemical study of a new inhibitor-resistant β-lactamase, SHV-84, produced by a clinical Escherichia coli strain. Antimicrob. Agents Chemother. 54, 2271–2272. doi:10.1128/AAC.01442-09
Martin, M. J., Stribling, W., Ong, A. C., Maybank, R., Kwak, Y. I., Rosado-Mendez, J. A., et al. (2023). A panel of diverse Klebsiella pneumoniae clinical isolates for research and development. Microb. Genom 9 (5), mgen000967. doi:10.1099/mgen.0.000967
Mendes, R. E., Doyle, T. B., Streit, J. M., Arhin, F. F., Sader, H. S., and Castanheira, M. (2021). Investigation of mechanisms responsible for decreased susceptibility of aztreonam/avibactam activity in clinical isolates of Enterobacterales collected in Europe, Asia and Latin America in 2019. J. Antimicrob. Chemother. 76, 2833–2838. doi:10.1093/jac/dkab279
Morinaka, A., Tsutsumi, Y., Yamada, M., Suzuki, K., Watanabe, T., Abe, T., et al. (2015). OP0595, a new diazabicyclooctane: mode of action as a serine β-lactamase inhibitor, antibiotic and β-lactam ‘enhancer. J. Antimicrob. Chemother. 70, 2779–2786. doi:10.1093/jac/dkv166
Papp-Wallace, K. M., Winkler, M. L., Taracila, M. A., and Bonomo, R. A. (2015). Variants of β-lactamase KPC-2 that are resistant to inhibition by avibactam. Antimicrob. Agents Chemother. 59, 3710–3717. doi:10.1128/AAC.04406-14
Pasteran, F., Meo, A., Gomez, S., Derdoy, L., Albronoz, E., Faccone, D., et al. (2014). Emergence of genetically related NDM-1-producing Providencia rettgeri strains in Argentina. J. Glob. Antimicrob. Resist. 2 (4), 344–345. doi:10.1016/j.jgar.2014.07.003
Periasamy, H., Joshi, P., Palwe, S., Shrivastava, R., Bhagwat, S., and Patel, M. (2020). High prevalence of Escherichia coli clinical isolates in India harbouring four amino acid inserts in PBP3 adversely impacting activity of aztreonam/avibactam. J. Antimicrob. Chemother. 75, 1650–1651. doi:10.1093/jac/dkaa021
Ramsey, C., and MacGowan, A. P. (2016). A review of the pharmacokinetics and pharmacodynamics of aztreonam. J. Antimicrob. Chemother. 71, 2704–2712. doi:10.1093/jac/dkw231
Rice, L. B., Carias, L. L., Hujer, A. M., Bonafede, M., Hutton, R., Hoyen, C., et al. (2000). High-level expression of chromosomally encoded SHV-1 β-lactamase and an outer membrane protein change confer resistance to ceftazidime and piperacillin- tazobactam in a clinical isolate of Klebsiella pneumoniae. Antimicrob. Agents Chemother. 44, 362–367. doi:10.1128/AAC.44.2.362-367.2000
Rodríguez-Gascón, A., and Canut-Blasco, A. (2019). Deciphering pharmacokinetics and pharmacodynamics of fosfomycin. Rev. Esp. Quimioter. 32 (Suppl. 1), 19–24.
Rozwandowicz, M., Brouwer, M. S., Fischer, J., Wagenaar, J. A., Gonzalez-Zorn, B., Guerra, B., et al. (2018). Plasmids carrying antimicrobial resistance genes in Enterobacteriaceae. J. Antimicrob. Chemother. 73 (5), 1121–1137. doi:10.1093/jac/dkx488
Sato, T., Ito, A., Ishioka, Y., Matsumoto, S., Rokushima, M., Kazmierczak, K. M., et al. (2020). Escherichia coli strains possessing a four amino acid YRIN insertion in PBP3 identified as part of the SIDERO-WT-2014 surveillance study. JAC Antimicrob. Resist 2, dlaa081. doi:10.1093/jacamr/dlaa081
Sauvage, E., Derouaux, A., Fraipont, C., Joris, M., Herman, R., Rocaboy, M., et al. (2014). Crystal structure of penicillin-binding protein 3 (PBP3) from Escherichia coli. PLoS One 9, e98042. doi:10.1371/journal.pone.0098042
Seemann, T. (2014). Prokka: rapid prokaryotic genome annotation. Bioinformatics 30, 2068–2069. doi:10.1093/bioinformatics/btu153
Simner, P. J., Bergman, Y., Conzemius, R., Jacobs, E., Tekle, T., Beisken, S., et al. (2023). An NDM-producing Escherichia coli clinical isolate exhibiting resistance to cefiderocol and the combination of ceftazidime-avibactam and aztreonam: another step toward pan-β-lactam resistance. Open Forum Infect. Dis. 10, ofad276. doi:10.1093/ofid/ofad276
Sivasankar, S., Premnath, M. A., Boppe, A., Grobusch, M. P., and Jeyaraj, S. (2023). Screening of MMV pandemic response and pathogen box compounds against pan-drug-resistant Klebsiella pneumoniae to identify potent inhibitory compounds. New Microbes New Infect. 55, 101193. doi:10.1016/j.nmni.2023.101193
Soman, R., Sirsat, R., Sunavala, A., Punatar, N., Mehta, J., Rodrigues, C., et al. (2024). Successful treatment of sino-pulmonary infection & skull base osteomyelitis caused by New Delhi metallo-β-lactamase-producing Pseudomonas aeruginosa in a renal transplant recipient by using an investigational antibiotic cefepime/zidebactam (WCK 5222). Eur. J. Clin. Microbiol. Infect. Dis. doi:10.1007/s10096-024-04791-1
Tamma, P. D., Heil, E. L., Justo, J. A., Mathers, A. J., Satlin, M. J., and Bonomo, R. A. (2024). Infectious diseases society of America 2024 guidance on the treatment of antimicrobial-resistant gram-negative infections. Clin. Infect. Dis., ciae403. doi:10.1093/cid/ciae403
Tsai, Y.-K., Fung, C.-P., Lin, J.-C., Chen, J.-H., Chang, F.-Y., Chen, T.-L., et al. (2011). Klebsiella pneumoniae outer membrane porins OmpK35 and OmpK36 play roles in both antimicrobial resistance and virulence. Antimicrob. Agents Chemother. 55, 1485–1493. doi:10.1128/AAC.01275-10
U.S. Food and Drug Administration (2025). Antibacterial susceptibility test interpretive criteria. Available online at: https://www.fda.gov/drugs/development-resources/antibacterial-susceptibility-test-interpretive-criteria (Accessed January 24, 2025).
Venturini, C., Zakour, N. B., Bowring, B., Morales, S., Cole, R., Kovach, Z., et al. (2019). K. pneumoniae ST258 genomic variability and bacteriophage susceptibility. bioRxiv. doi:10.1101/628339
Villa, L., Feudi, C., Fortini, D., García-Fernández, A., and Carattoli, A. (2014). Genomics of KPC-producing Klebsiella pneumoniae sequence type 512 clone highlights the role of RamR and ribosomal S10 protein mutations in conferring tigecycline resistance. Antimicrob. Agents Chemother. 58, 1707–1712. doi:10.1128/AAC.01803-13
Wang, L., Zhu, J., Chen, L., and Du, H. (2024). Cefiderocol: clinical application and emergence of resistance. Drug Resist. Updat. 72, 101034. doi:10.1016/j.drup.2023.101034
Wick, R. R., Judd, L. M., Gorrie, C. L., and Holt, K. E. (2017). Unicycler: resolving bacterial genome assemblies from short and long sequencing reads. PLoS Comput. Biol. 13, e1005595. doi:10.1371/journal.pcbi.1005595
Winkler, M. L., Papp-Wallace, K. M., Taracila, M. A., and Bonomo, R. A. (2015). Avibactam and inhibitor-resistant SHV β-lactamases. Antimicrob. Agents Chemother. 59, 3700–3709. doi:10.1128/AAC.04405-14
Wise, M. G., Karlowsky, J. A., Mohamed, N., Hermsen, E. D., Kamat, S., Townsend, A., et al. (2024). Global trends in carbapenem- and difficult-to-treat-resistance among World Health Organization priority bacterial pathogens: ATLAS surveillance program 2018-2022. J. Glob. Antimicrob. Resist. 37, 168–175. doi:10.1016/j.jgar.2024.03.020
World Health Organization (2024). Antibacterial agents in clinical and preclinical development: an overview. Geneva: WHO.
Yahav, D., Giske, C. G., Grāmatniece, A., Abodakpi, H., Tam, V. H., and Leibovici, L. (2020). New β-Lactam–β-Lactamase inhibitor combinations. Clin. Microbiol. Rev. 34. doi:10.1128/CMR.00115-20
Zhang, Y., Kashikar, A., Brown, C. A., Denys, G., and Bush, K. (2017). Unusual Escherichia coli PBP 3 insertion sequence identified from a collection of carbapenem-resistant enterobacteriaceae tested in vitro with a combination of ceftazidime-ceftaroline-or aztreonam-avibactam. Antimicrob. Agents Chemother. 61. doi:10.1128/AAC.00389-17
Keywords: multi-drug resistance, emerging pathogens, Klebsiella pneumoniae, pan-drug resistance, aztreonam-avibactam, NDM carbapenemase, cefiderocol, cefepime-zidebactam
Citation: Pasteran F, Manuel De Mendieta J, Pujato N, Dotta G, González LJ, Rizzo M, Fernández A, Ceriana P, Maccari L, Rapoport M, Gómez S, Lucero C, Menocal MA, Albornoz E, De Belder D, Radisic M, Vila AJ and Corso A (2025) From genomics to treatment: overcoming pan-drug-resistant Klebsiella pneumoniae in clinical settings. Front. Pharmacol. 16:1570278. doi: 10.3389/fphar.2025.1570278
Received: 03 February 2025; Accepted: 15 May 2025;
Published: 30 May 2025.
Edited by:
Ramona Iseppi, University of Modena and Reggio Emilia, ItalyReviewed by:
Shangxin Yang, University of California, Los Angeles, United StatesPedro Teixeira, National Institute of Health Dr. Ricardo Jorge, Portugal
Copyright © 2025 Pasteran, Manuel De Mendieta, Pujato, Dotta, González, Rizzo, Fernández, Ceriana, Maccari, Rapoport, Gómez, Lucero, Menocal, Albornoz, De Belder, Radisic, Vila and Corso. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Fernando Pasteran, ZnBhc3RlcmFuQGdtYWlsLmNvbQ==
†Deceased