 Jiayi Luo1†
Jiayi Luo1† Lijiao Wu
Lijiao Wu Jingqi Zhang
Jingqi Zhang Xiangjin Wang
Xiangjin Wang- 1Shanghai Shuguang Hospital affiliated to Shanghai University of Traditional Chinese Medicine, Shanghai, China
- 2Hospital of Chengdu University of Traditional Chinese Medicine, Chengdu, China
- 3College of Medicine and Life Sciences, Chengdu University of Traditional Chinese Medicine, Chengdu, China
- 4College of lntegrated Traditional Chinese and Western Medicine, Hunan University of Chinese Medicine, Changsha, China
The prevalence of bone and joint diseases is projected to increase owing to rapidly aging populations, sedentary lifestyles, and unhealthy diets. This poses a significant challenge for the global healthcare system. In recent years, natural herbal medicines have been used to treat various types of orthopedic diseases, opening new frontiers in new drug research. Epimedium is a traditional Chinese herb with a long history of medicinal use. It is commonly used to treat osteoporosis, joint disorders, cardiovascular diseases, sexual dysfunction and aging. The primary active component of Epimedium is icariin (ICA), an isoprenylated flavonoid. Recent studies have demonstrated its significant positive effects on bone metabolism and remodeling, including promoting osteoblast proliferation and mineralization, reducing osteoclast activity, and inhibiting inflammation and oxidative stress. Thus, ICA represents a potential compound for treating bone and joint diseases. However, the specific mechanisms underlying these effects have not yet been fully elucidated. This paper focuses on the latest advances in the use of ICA for the treatment of skeletal and joint diseases, covering a range of conditions including osteoporosis, osteoarthritis, rheumatoid arthritis, intervertebral disc degeneration and fractures. Building on this, we have systematically integrated its multi-target pharmacological mechanism network for the first time, elucidating its multi-pathway synergistic effects mediated by regulating the balance of the bone microenvironment. In summary, as a multi-target natural compound, ICA demonstrates significant translational medicine potential in the comprehensive treatment of bone and joint diseases, providing a critical theoretical foundation for the development of novel therapeutic strategies.
1 Introduction
Epimedium, the largest herbaceous genus in the Berberidaceae family, comprises approximately 62 species (Zhuang et al., 2023). These plants have a discontinuous distribution from Algeria in North Africa to East Asia, where the majority of the species (approximately 52) have undergone continuous evolution (Wang J. et al., 2022; Zhang et al., 2022b). Certain species of Epimedium have been used in traditional Asian medicine and have been shown to have significant therapeutic activity (Liang et al., 2025). Epimedium was first recorded in Shennong Ben Cao Jing and is recognized as a “medium” herb in the most famous Chinese medical text, Ben Cao Gang Mu (Zhang et al., 2022a).
Epimedium dispels wind and disperses cold, tonifies the kidneys, and strengthens tendons (Ma H. et al., 2011). Traditional medicine posits that tonifying kidney yang can nourish kidney essence, thereby promoting bone marrow production and nourishing bones to strengthen tendons and bones, as well as alleviate joint stiffness and pain (e.g., rheumatic pain and impaired joint flexion/extension). Additionally, according to traditional Chinese medical theory, “obstruction of meridians by wind, cold, and dampness is the root cause of arthralgia.” Possessing wind-cold-dispelling properties, Epimedium can eliminate wind-cold-dampness pathogens invading the tendons and bones, dredge qi and blood, and thus synergize with its effects of tonifying the kidney and strengthening the tendons. Owing to its therapeutic effects on the bones and kidneys, it is used as a traditional medicine for treating osteoporosis, joint disorders, cardiovascular disorders, sexual dysfunction, as well as slowing aging in many countries and regions (Guo et al., 2022; Wang et al., 2025). Therefore, Epimedium has great potential for research and development.
Previous studies have identified more than 260 compounds in epimedium, including flavonoids, lignans, violacein, phenolic glycosides, and other compounds representing various categories of secondary metabolites (Wu et al., 2003). The most well-known and phytochemically characteristic of these compounds are flavonoids, with the most prominent components being isoprenylated flavonoid glycosides (Lin et al., 2023). As the flavonoid component with the highest content and most extensive research, icariin (ICA) has been identified as the primary material basis mediating the core pharmacological effects of Epimedium crude extracts, including anti-inflammatory activity, promotion of bone formation, and inhibition of bone resorption (Ma H. et al., 2011). In addition, the Chinese Pharmacopoeia also designates ICA as a chemical marker for the quality control of Epimedium (Zhou et al., 2015). ICA has been shown to possess several pharmacological effects, including osteoprotective, anti-osteoporotic, anti-inflammatory and antioxidant capabilities, as well as enhancing cardiovascular function, neuroprotection, and regulating the immune system (Ming et al., 2013; Buhrmann et al., 2020; He et al., 2020; Yan et al., 2023). Intensive research on the effects of ICA on bone damage has attracted attention to its potential as a therapeutic agent for the treatment of bone and joint diseases.
Bone and joint diseases are a major societal burden affecting people worldwide (Imagama et al., 2020). Generally, they arise from abnormal metabolism and cell death in the skeletal system, including osteoblasts, osteoclasts, chondrocytes, and bone marrow mesenchymal stem cells (BMMSCs) (Zeng et al., 2022). A precise balance between bone resorption and formation is crucial for bone homeostasis (Li et al., 2022). Imbalance in bone homeostasis is one of the major causes of the development of certain bone diseases, including osteoporosis (OP), osteoarthritis (OA), rheumatoid arthritis (RA), intervertebral disc degeneration (IVDD), and delayed healing/nonunion fractures (Wang S. et al., 2020; Zeng et al., 2022). In addition, trauma, substance abuse, cartilage degeneration, hormonal imbalance, and aging are potential risk factors for the development of bone disease (Cao et al., 2017; Luo et al., 2018). Currently, the standard treatments for bone diseases include surgery-based revascularization, joint replacement, oral pharmacological agents, rehabilitation in non-surgical programs, and alternative therapies (McCormick et al., 2014; Tonomura et al., 2020). Although these strategies are frequently updated with advances in medicine and play a crucial role in disease prevention and treatment, they have some drawbacks, such as irreversible complications, high costs, limitations in long-term use, and side effects (Ruderman, 2012; Sugiyama et al., 2015). Side effects have been reported with long-term use of anti-osteoporotic drugs, such as bisphosphonates, which can cause upper gastrointestinal bleeding, hypersensitivity reactions, hypocalcemia and myalgia (Papapetrou, 2009); and use of teriparatide for more than 2 years may increase the risk of developing osteosarcoma (Ma et al., 2004). In recent years, natural Chinese medicines have been used to treat various orthopedic conditions owing to their excellent efficacy, high safety and few side effects (Mukwaya et al., 2014). ICA, a natural compound extracted from traditional Chinese medicine (TCM), plays an important role in the treatment of bone and joint diseases. ICA possesses osteoinductive potential for bone tissue engineering and contributes to the regulation of various signaling pathways, including those involved in anti-osteoporosis, osteogenesis, anti-fracture, chondrogenesis, angiogenesis and anti-inflammation (Zhang et al., 2014). Numerous animal studies have confirmed the osteoprotective effects of ICA in vitro and in vivo. One study found that ICA promoted osteogenic differentiation in vitro and alleviated osteoporosis in vivo by inhibiting the Notch signaling pathway (Xu Y. et al., 2019). Another in vivo study showed that ICA treatment reduced articular cartilage damage in OA rats by inhibiting cartilage extracellular matrix (ECM) degradation and chondrocyte iron death (Xiao et al., 2024). Angiogenesis also plays an important role in bone regeneration, and it has been reported that ICA can enhance angiogenesis through endothelial cell migration, proliferation and renal therapy in vivo (Chung et al., 2008). Additionally, ICA can be stabilized for local release using biomaterials, making it a promising drug candidate for the treatment of bone and joint disorders (Zhao et al., 2010).
Although existing studies have confirmed the multifaceted roles of ICA in skeletal and joint diseases, its mechanisms of action remain fragmented: on the one hand, the signaling pathways regulated by ICA in different diseases lack cross-disease systemic associations; on the other hand, the interactions between emerging mechanisms such as oxidative stress and autophagy and classical osteogenic/osteoclastic pathways remain unclear, and research linking drug toxicity to clinical translation is relatively weak. Based on this, this review systematically integrates the cross-disease regulatory networks of ICA in various diseases such as OP and OA, breaking through the limitations of single-disease mechanism studies; it focuses on elucidating the interactions between emerging mechanisms such as oxidative stress and autophagy and classical signaling pathways; simultaneously, it offers forward-looking insights into drug toxicity assessment and clinical translation strategies, providing a more comprehensive theoretical foundation for the full-chain development of ICA from basic research to clinical application—an area previously under-explored in systematic research.
2 Epimedium and its active ingredient–icariin
Herbal Epimedium is prepared from the dried leaves of the Epimedium medicinal plant. Epimedium has been used in traditional Chinese medicine for over 2000 years, and is commonly used to treat bone fractures, joint ailments, chronic diseases and aging (Gao et al., 2022). The primary pharmacologically active compounds in Epimedium are isopentenyl flavonol glycosides, with various sugar groups at the 3-OH and 7-OH positions. Among these compounds, epimedin A, B, and C exhibit certain biological activities (Shen et al., 2007a). However, ICA is currently the most extensively studied and mechanistically well-defined core active monomer, which can be extracted from the dried stems and leaves of E. sagittatum Maxim., E. pubescens Maxim., E. brevicornu Maxim., E. koreanum Nakai, and E. wushanense T. S. Ying (Liu et al., 2023).
ICA (C33H40O15, molecular weight: 676.66) (Yang et al., 2019) is an isoprenyl flavonoid that appears as a yellowish powder (Figure 1). It is composed of a glucose group at C-3, a methoxyl group at C-4, a prenyl group at the C-8 position and a rhamnosyl group at C-7 (He et al., 2020). This structure endows ICA with diverse pharmacological activities. Recent research has focused on its bone-protective, anti-inflammatory, anti-cancer, and immunomodulatory effects (Li Y. et al., 2024), demonstrating significant potential particularly in the treatment of bone and joint diseases.
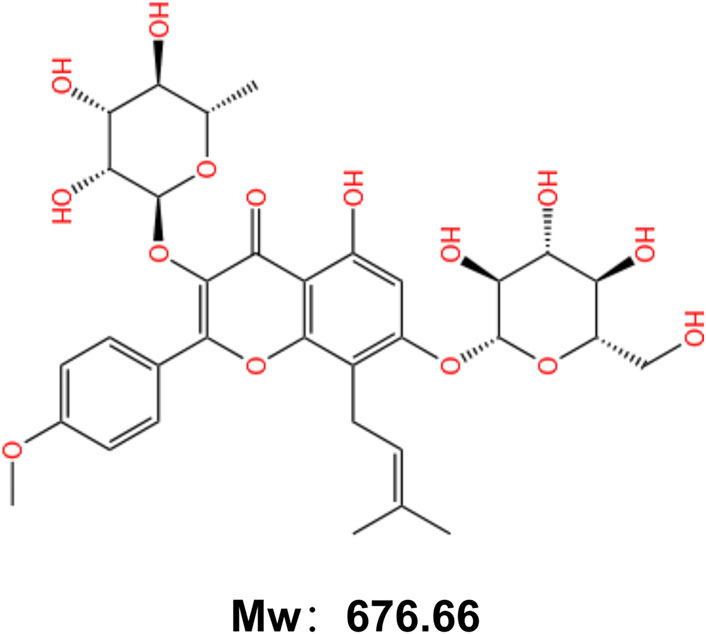
Figure 1. Chemical structure of Icariin.
Currently, multiple studies have been conducted to provide a global perspective on the pharmacokinetics of ICA. The pharmacokinetic properties of ICA are closely linked to the method of administration. Xu et al. investigated the absorption behavior of ICA in rats following oral or intramuscular injection and discovered that the half-life of orally administered ICA was 3.149–2.364 h (Xu et al., 2017). Intravenously administered ICA exhibited a rapid decrease in blood concentration, with a half-life of 0.562–0.200 h. They also investigated the metabolic pattern of ICA and found that plasma and tissue concentrations of ICA remained very low at all collection sites, suggesting that epimedium glycosides are poorly absorbed after oral administration. In addition, the concentrations were much higher in the liver, lung and reproductive organs than in other tissues, suggesting that the liver, lung and reproductive organs are the main targets of ICA in rats. Ye et al. found that ICA is difficult to cross the blood-brain barrier (Lika et al., 1999), but peripheral administration of ICA has also been reported to have significant central nervous system pharmacological activity (Jin et al., 2014; Li F. et al., 2015). In addition, the distribution of ICA varied with concentration and sex. For instance, at moderate concentrations, ICA is present in the testicles of male rats and at higher concentrations in the uterus and ovaries of female rats (Xu et al., 2017). Further, it has been found that ICA metabolizes to Epimedium and desmethyl Epimedium in humans (Shen et al., 2007b), whereas it is only metabolized to desmethyl Epimedium in SD rats, suggesting species-dependent metabolism (Wong et al., 2009). Therefore, a more thorough metabolic profile of ICA in humans needs to be established. For orally administered drugs, the most important pharmacokinetic property is their bioavailability (Tan et al., 2021). Notably, like other flavonoids, ICA and its derivatives have low oral bioavailability (∼12%), which may be due to the low water solubility and membrane permeability of ICA and the slow dissolution rate in biological fluids (Jin et al., 2019). Several methods have been developed to improve the oral bioavailability of ICA. In the most successful study (Szabó et al., 2022), a 6.57-fold increase in bioavailability was achieved using phospholipid complex formation. In addition, the simultaneous application of several different techniques may result in a synergistic effect that exponentially increases the bioavailability of insoluble flavonoids. These cutting-edge studies open up the possibility of the therapeutic potential of ICA in bone and joint diseases.
Positive advances in the pharmacological activities of ICA have been made in recent years in relation to the skeletal system, cancer, nervous system, immune system and cardiovascular system (Li C. et al., 2015). A series of studies have demonstrated multiple mechanisms through which ICA treats bone and joint diseases, including the induction of bone formation, inhibition of bone resorption and angiogenesis (Wang et al., 2018). For example, Wang et al. found that ICA and five other flavonoids enhance extracellular matrix mineralization, promote osteoblast proliferation and differentiation, and facilitate the osteogenic differentiation of bone marrow mesenchymal stem cells (BMSCs) (Wang et al., 2024). Additionally, Li et al. confirmed that icariin not only inhibits osteoclast formation but also suppresses their bone resorption capacity (Li F. et al., 2024). Although clinical trials remain limited, existing research has revealed that ICA can treat bone and joint diseases through multiple pathways. However, these molecular mechanisms require further comprehensive summarization (Figure 2).
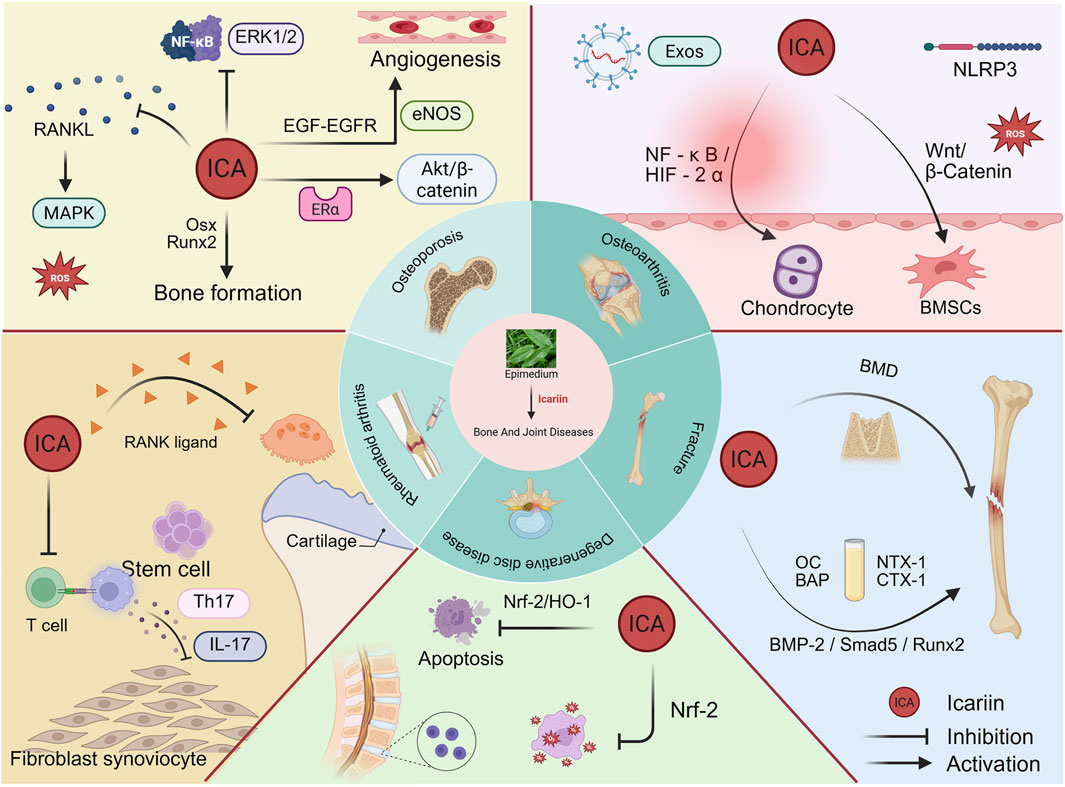
Figure 2. Therapeutic potential of Icariin in bone and joint disorders.
3 Molecular mechanism of icariin in treating bone and joint diseases
3.1 Osteoporosis
Osteoporosis (OP) is a chronic metabolic bone disease characterized by reduced bone density and microarchitecture, resulting in increased bone fragility and fracture risk (Li et al., 2023). OP affects over 200 million people worldwide, making it one of the most prevalent diseases among the elderly (Jia et al., 2012; Ballane et al., 2017). OP causes over 8.9 million osteoporotic fractures annually worldwide (Böcker et al., 2022), resulting in a significant socioeconomic burden. The primary treatments for OP include humanized receptor activator of nuclear factor-κΒ ligand (RANKL) monoclonal antibodies, bisphosphonates, selective estrogen receptor modulators and calcitonin (Liu et al., 2020). However, these therapeutic strategies are associated with significant adverse effects, including jaw osteonecrosis, hypercalcemia, breast cancer and cardiovascular disease (Fleisch, 2003; Barrett-Connor et al., 2006). The pathogenesis of OP mainly includes the loss of bone homeostasis, with the maintenance of bone homeostasis dependent on the appropriate balance between osteoblasts differentiated from BMMSCs and osteoclasts produced by the fusion of mononuclear macrophages (Song et al., 2022). Various transcription factors, such as osterix (Osx), runt-related protein 2 (Runx2), Wingless and INT-1 signals have been shown to regulate osteoblast differentiation and function (Long, 2011). Similarly, osteoclast differentiation and function are regulated by factors, such as RANKL, macrophage colony-stimulating factor (M-CSF), cytokines and αVβ 3 integrins (Teitelbaum, 2000). Additionally, numerous loci related to OP that regulate bone homeostasis have been identified. These include the low-density lipoprotein receptor-related protein 5 (LRP5), estrogen receptor α (ESR1) and osteoprotegerin (OPG) (Richards et al., 2008).
In addition, ICA inhibits osteoclastogenesis to treat OP by regulating RANKL-induced signaling pathways. RANKL primarily controls the function and survival of mature osteoclasts (Takayanagi et al., 2002). It binds to its receptor RANK, which is expressed on the surface of osteoclast precursor cells, whereas OPG serves as a protective factor for inducing osteoclast differentiation (Nakashima et al., 2012). Xu et al. discovered that ICA inhibited RANKL-induced phosphorylation of three MAPKs in RAW264.7 cells, indicating that ICA exerts an inhibitory effect on osteoclast differentiation by regulating MAPK phosphorylation in mature osteoclasts (Xu Q. et al., 2019). ICA administration also significantly restored serum calcium levels and OPG/RANKL ratios in osteoblasts, acting as a bone preservative in simulating microgravity-induced bone loss (He J. et al., 2018).In addition, ICA inhibits osteoclast differentiation by downregulating TRAF6, and inhibiting the activation of ERK1/2 and nuclear factor-κB (NF-κB), thereby effectively inhibiting RANKL-induced expression of nuclear factor of activated T cell cytosolic 1 (NFATc1) and c-Fos transcription factors, which are key factors regulating osteoclast-specific genes (Kim et al., 2018).
The decreased bone mass and increased bone marrow adipose tissue in OP is closely related to the dynamic balance between the differentiation of BMMSCs into adipocytes and osteoblasts. Inhibition of adipogenic differentiation of BMMSCs and promotion of osteogenic differentiation are important indices to validate the treatment of OP (Justesen et al., 2001). Several studies have shown that the maturation and mineralization of osteoblasts and osteogenic differentiation of BMMSCs during bone formation can be improved and enhanced by ICA treatment (Huang et al., 2007). Mechanistically, ICA exerts these effects through activation of the Wnt/β-catenin pathway, which is attributed to the potent anti-apoptotic activity of ICA (Chen et al., 2016). In addition, Zhang et al. demonstrated that mouse MC3T3 - E1 and RAW264.7 cells could be co-cultured in vitro, and ICA treatment increased the osteogenic activity of these cells (Zhang et al., 2017). ICA also promotes bone formation by inducing the expression of Osx, Runx2, alkaline phosphatase (ALP), type I collagen, and other proosteoblast genes to promote bone formation (Ma H. P. et al., 2011). In addition, ICA has been shown to increase the expression of Runx2 and Bmp2, while reducing the expression of CCAAT/enhancer binding protein alpha (C/EBPa) and peroxisome proliferator-activated receptor gamma (Pparγ). This suggests that ICA stimulates osteoblast differentiation and mineralization and inhibits osteoblast fatty degeneration, thereby increasing the number of osteoblasts that differentiate into mature osteocytes (Zhang et al., 2016). Ye et al. found that ICA promotes the proliferation and osteogenic differentiation of rat adipose stem cells through the Rho A-TAZ pathway at an optimal concentration of 10–7 M (Ye et al., 2017).
Another way that ICA acts as an antiosteoporotic agent is by preventing estrogen deficiency. Estrogen is used clinically for bone conditions because it can regulate bone formation and resorption in bone homeostasis. Estrogen receptor α (Erα) mediates the activity of estrogen in safeguarding cortical bone mass, which is crucial for osteoblast differentiation (Manolagas et al., 2013). Decreased estrogen concentration in postmenopausal women often reduces the amount and function of ERα, potentially resulting in osteoporosis (Song et al., 2008). Zhao et al. reported that ICA activates the Akt/β-catenin pathway by upregulating ERα expression through phytoestrogen administration in de-ovulated rats (Zhao et al., 2019). This promotes osteoblast proliferation and differentiation, leading to improved locomotor skeletal muscle response. ICA also promotes transcription of the downstream osteogenic gene osteocalcin (Ocn) through STAT3, thereby preventing estrogen deficiency-induced alveolar bone loss (Xu et al., 2020). In an in vivo study, Xu et al. elucidated a novel mechanism by which ICA promotes osteogenic differentiation and bone formation by inhibiting the Notch signaling pathway (Xu Y. et al., 2019). In addition, ICA rapidly induces extracellular signaling to promote the activation of extracellular signal-regulated kinase (ERK) and c-Jun N-terminal kinase (JNK), which effectively promote osteoblast proliferation (Song et al., 2013). Interestingly, the use of the estrogen receptor antagonist, ICI 182, 780, weakened ICA-mediated proliferation and mineralization of osteoblasts, indicating that ICA may function through the ER.
Oxidative stress plays a vital role in decreasing bone density and strength. Low oxygen levels or hypoxia result in an increased generation of reactive oxygen species (ROS) (Miura and Tanno, 2010), which can cause damage to various cellular components, such as proteins, lipids and DNA (Kim and Park, 2003). The antioxidant properties of ICA diminish hypoxia-induced oxidative stress and apoptosis in primary osteoblasts by reducing ROS production and intracellular malondialdehyde (MDA) levels and increasing superoxide dismutase activity. Additionally, it inhibits hypoxia-induced reduction in osteoblast differentiation and mineralization in a dose-dependent manner (Ma et al., 2014). Furthermore, ICA scavenges ROS and maintains osteoblast mitochondrial and primary ciliary homeostasis in diabetes and iron overload-induced OP. ICA also reverses iron overload-induced reductions in the expression of Runx2, alkaline phosphatase and bone-bridging proteins, and significantly prevents bone loss in mice with iron-overloaded (Jing et al., 2019; Liu J. et al., 2022).
The vascular system exerts a pivotal role in bone tissue growth, repair, and remodeling by facilitating nutrient supply and waste removal (Wang and Yeung, 2017). The local blood supply provides essential cells and growth factors for bone regeneration. Notably, specialized cell types within the local vasculature, such as H-type vascular endothelial cells, have been shown to form a complex communication network with various cells in the bone microenvironment (Yang et al., 2026). Studies have demonstrated that H-type vessels can both secrete osteogenic factors and establish physical interactions with bone progenitor cells, thereby directly mediating the “angiogenesis-osteogenesis coupling” process (Kusumbe et al., 2014). ICA has been demonstrated to promote osteogenesis and angiogenesis. In porcine aortic endothelial cells, ICA upregulates the expression of endothelial nitric oxide synthase (eNOS) by activating the EGF-EGFR pathway, thereby modulating endothelial cell function (Liu et al., 2011). Cheng et al. reported that ICA prevents bone loss in ovariectomized rats through activating angiogenesis and inhibiting inflammatory responses (Cheng et al., 2014). Additionally, ICA has been widely utilized in bone defect repair, drug-eluting scaffold development, and biomimetic scaffold fabrication. Zhao et al. constructed a biomimetic scaffold incorporating in situ-encapsulated PLGA@icariin microspheres, which regulates the immune microenvironment and activates the vascular-bone regeneration cascade via the STAT3 signaling pathway (Zhao et al., 2025). Collectively, these studies highlight the potential of ICA as a therapeutic agent with vascular endothelial growth factor (VEGF)-like activity.
In addition to those mentioned above, there are many signaling pathways involved in the treatment of OP. Huang et al. found that ICA could alleviate glucocorticoid (GC)-induced OP by regulating the balance of the EphB4/Ephrin-B2 axis, thereby promoting OP recovery (Huang et al., 2020). In addition, the gut microbiota-bone axis has been recognized as a key mediator of bone homeostasis (Ding et al., 2020). ICA has been shown to prevent bone loss by modulating gut microbiota (GM) and regulating metabolite changes. Potential mechanisms include GM-regulated serum biomarkers RANK, RANKL, OPG and tartrate-resistant acid phosphatase (TRACP), and changes in fecal metabolites such as bile acids, amino acids and fatty acids (Wang et al., 2022c).
3.2 Osteoarthritis
Cartilage is an avascular tissue and the absence of blood vessels is considered a key feature of permanent cartilage homeostasis. The intact cartilage and its microenvironment provide balanced support for joint stability (Goldring and Goldring, 2010). Osteoarthritis (OA) is a chronic bone and joint disease characterized by degeneration and destruction of articular cartilage and osteophytes in weight-bearing areas (Zhu et al., 2022). Primary clinical symptoms include pain, swelling and deformation of the knee joint (Martel-Pelletier et al., 2016). Throughout the progression of OA, pathologies affecting the cartilage include oxidative stress, inflammation, apoptosis, cartilage matrix loss and autophagy.
Cellular inflammation and ECM degradation are increasingly recognized as key drivers of cartilage damage in OA. Chondrocytes respond to inflammatory factors in joint tissues and produce proinflammatory factors (Goldring and Marcu, 2009). Interleukin-1β (IL-1β) and tumor necrosis factor-α (TNF-α), catabolic cytokines involved in ECM degradation, promote protease production in chondrocytes, leading to cartilage joint destruction (Chen et al., 2015). A previous study showed that ICA can protect chondrocytes from inflammatory damage by inhibiting the activation of the NF-κB/HIF-2α signaling pathway in a mouse model of bone defect (Wang P. et al., 2020). ICA reduces the expressions of HIF-2α, MMP9 and ADMTS5, while promoting those of chondrocyte-specific markers SOX9, AGG and Col2α, in turn inhibiting cartilage degradation and increasing chondrocyte viability. NO and ROS are key in the progression of OA and can interact and derive a series of highly oxidized free radicals that can lead to cell damage and apoptosis (Ansari et al., 2020; Jiang et al., 2023). Zuo et al. reported that Nrf2, a key redox transcription factor, plays an important role in inhibiting oxidative stress and demonstrated that ICA can reduce excessive ROS production and increase the expression of Nrf2 and downstream antioxidant enzymes to achieve cartilage-protection (Zuo et al., 2019).
Inflammation, oxidative stress and cell-death mechanisms (apoptosis, pyroptosis and autophagy) are closely connected. In OA, inflammation, oxidative stress and autophagy are correlated with chondrocyte apoptosis (Hwang and Kim, 2015). Wang et al. found that ICA induces the upregulation of CYTOR in IncRNA to inhibit the apoptosis of chondrocytes (Wang et al., 2021). In contrast to apoptosis, pyroptosis is pro-inflammatory (Wang et al., 2022b). Pyroptosis is primarily mediated by the NLRP3 inflammasome. On one hand, it accelerates the release of pro-inflammatory cytokines and expands the local inflammatory response; on the other hand, the formation of caspase-1 and gasdermin (GSDM) D-N leads to cell swelling and dissolution, which play a cytotoxic role (Wang et al., 2022b). In a lipopolysaccharide (LPS)-induced OA rat model, ICA was shown to reduce LPS-induced pyroptosis and inhibit collagen formation by arresting the NLRP3 inflammasome-mediated caspase-1 signaling pathway, effectively controlling the local inflammatory response (Zu et al., 2019).
Autophagy is a self-protection mechanism that uses autophagosomes and lysosomes to eliminate damaged organelles and boost cellular adaptation and survival (Li et al., 2019). Consequently, targeted therapy focusing on autophagy is a popular technique in the treatment of OA (Duan et al., 2020). The mammalian target of rapamycin (mTOR) negatively regulates autophagy and is regulated by upstream phosphatidylinositol 3-kinase (PI3K)/AKT and AMP-activated protein kinase (AMPK) signaling (Sun et al., 2020). Tang et al. discovered that ICA activates autophagy and reduces apoptosis in human chondrocytes in a dose-dependent manner. Animal experiments further demonstrated that ICA mediates PI3K/AKT/mTOR signaling, thereby regulating chondrocyte autophagy to alleviate OA (Tang et al., 2021) (Figure 3).
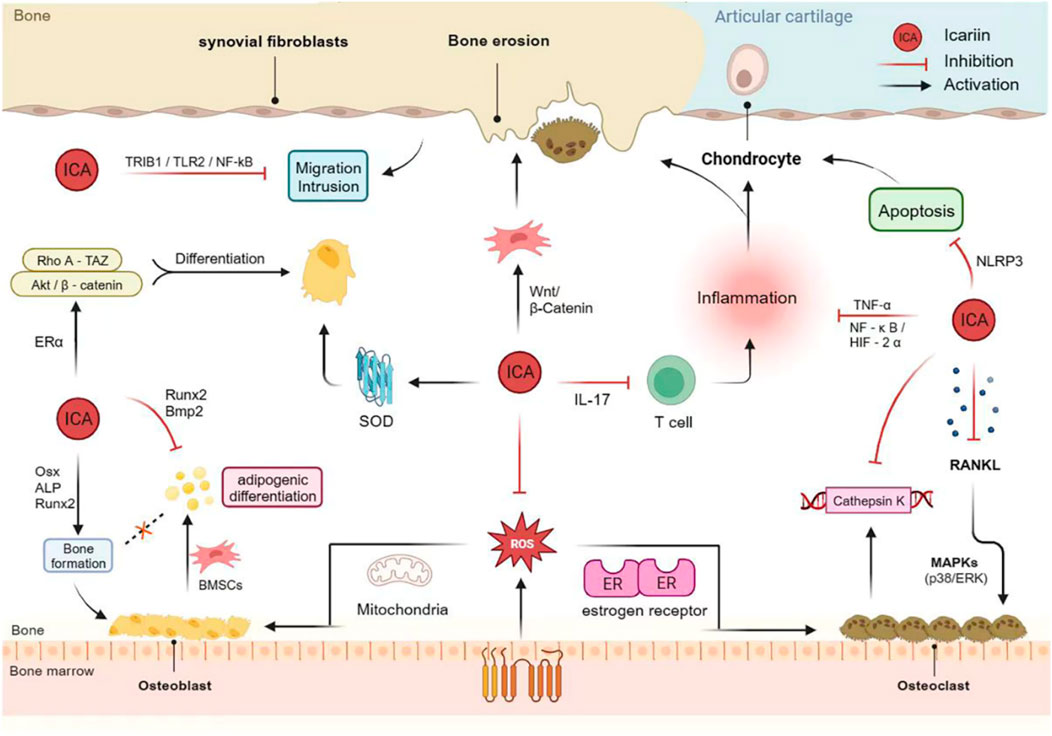
Figure 3. Mechanism of action of Icariin in bone protection and bone damage.
With the rise of regenerative medicine, stem cell and cytokine therapies have become viable options for treating OA (Zhu et al., 2022). Of these, BMMSCs are the most extensively researched and favored source for autologous cell therapy to promote cartilage repair (Ding et al., 2021). Zhu et al. reported that hydrogels loaded with different concentrations of ICA could differentially promote the proliferation and chondrogenic differentiation of BMMSCs through the activation of Wnt/β-catenin signaling in vitro (Zhu et al., 2022). Another in vivo study showed that intra-articular injections of ICA and BMSCs in female New Zealand rabbits enhanced the BMP2/Smad signaling pathway, which promoted repair of cartilage damage (Jiao et al., 2022). Furthermore, ICA enhances the tolerance of BMMSCs to ischemia and hypoxia, thereby promoting recovery from cartilage damage (Tang et al., 2022). In response to possible growth factor side effects, such as TGF-β stimulating BMMSC differentiation, ICA has been shown to improve the ability of TGF-β to promote BMMSC differentiation into juvenile bone. Interestingly, ICA did not increase the hypertrophic response or fibrotic cartilage formation (Wang et al., 2014).
In addition to the aforementioned factors, iron death is closely linked to OA. Luo and Zhang found that treatment of LPS-induced human synoviocytes with ICA decreased iron content, MDA and other factors associated with iron metabolism, while counteracting the increase in iron content and the imbalance between the oxidative and antioxidant systems induced by the GPX4 inhibitor RSL3 (Luo and Zhang, 2021). Moreover, some medical scientists have turned their attention to exosomes (Exos) (Watanabe et al., 2021); however, studies on the treatment of OA using ICA through modulation of Exos are scarce and deserve more attention in the future.
3.3 Rheumatoid arthritis
Rheumatoid arthritis (RA) is an autoimmune inflammatory disease of the joints affecting approximately 1% of the global population. It is characterized by synovial hyperplasia, mononuclear cell infiltration, and the erosion of cartilage and bone (Nagashima et al., 2012). The development of RA is intricate, involving dysfunctional immune cells and synovial fibroblasts, along with unusual expression of multiple autoantibodies and cytokines (McInnes et al., 2016).
Autoimmune diseases often result in inflammatory responses and subsequent tissue damage owing to humoral and cellular immune-mediated immune complex deposition and cytokine production (Khan et al., 2022). The STAT family of signal transducers and activators regulates cell proliferation, apoptosis, angiogenesis and immune response (Shahmarvand et al., 2018). Research has demonstrated that ICA alleviates rheumatoid inflammation and joint destruction by inhibiting the expression of osteoclast markers, such as β3 integrin, cathepsin K and MMP-9. In addition, it reduces the number of Th17 cells and inhibits STAT3 activation-mediated IL-17 production (Chi et al., 2014). Furthermore, it markedly increases mRNA and protein expression of Fas and the activities of caspase8 and caspase3 in CD4 + lymphocytes and decreases mRNA and protein expression of B-cell lymphoma-2 (Wang H. et al., 2017). Previous studies have shown that microRNAs (miRNAs) play an important role in the pathogenesis of RA, and miR-223-3p has been suggested as one of the possible biomarkers of RA (Ogando et al., 2016). ICA inhibits proliferation and inflammatory cytokine secretion by modulating miR-223-33p/NLRP3 signaling, which promotes apoptosis (Wu et al., 2020). Thus, ICA treats RA by modulating immune responses and inflammatory factors.
The hyperproliferation of fibroblast-like synoviocytes (FLS) and synovial angiogenesis are typical pathological features of RA (Lefèvre et al., 2009). Therefore, the inhibition of FLS proliferation is one way to treat RA. p21 is a cell cycle protein-dependent kinase inhibitor that regulates cell cycle progression in response to various stimuli (Karimian et al., 2016). Pu et al. found that ICA upregulates the expression of p21 and downregulates the expression of cytokinins B and E, suggesting that ICA inhibits cell proliferation by interfering with the cell cycle progression of FLS (Pu et al., 2021). ICA also inhibits TNF-α-induced inflammatory response and RA-FLS by regulating the TRIB1/TLR2/NF-κB pathway (Wu et al., 2022). In addition, ICA inhibits iron death by activating the Xc-/GPX4 axis, thereby protecting synovial cells from death (Luo and Zhang, 2021). However, increased invasion and migration of FLS due to inflammatory stimulation of the synovium in patients with RA also accelerates the destruction of articular cartilage and bone (Bottini and Firestein, 2013). Moreover, ICA considerably inhibits the migratory ability of FLS but has a lesser effect on their invasive ability (Pu et al., 2021).
Cathepsin K is expressed in osteoclasts and synovial fibroblasts (Salminen-Mankonen et al., 2007). Overexpression of cathepsin K in mice leads to synovial inflammation and increased cartilage and bone destruction with age, which is a characteristic of RA (Maffia et al., 2004). ICA inhibits the collagenase activity of cathepsin K, resulting in a reduction in clinical inflammation, fibrin deposition, synovial inflammation and cartilage degradation by 11%–25% (Sun et al., 2013).
3.4 Degenerative disc disease
Low back pain (LBP) is a common orthopedic condition worldwide that can result in disability and affect daily life. Intervertebral disc degeneration (IVDD) is the primary cause of LBP (Vergroesen et al., 2015). The intervertebral disc (IVD) comprises of the upper and lower cartilaginous endplates (CEPs), nucleus pulposus (NP) and outer annulus fibrosus (Stergar et al., 2019). Despite ongoing research, the pathogenesis of IVDD remains poorly understood and involves a multitude of effector molecules and complex signaling networks (Risbud and Shapiro, 2014). Current treatment options for IVDD include pain management using therapeutic agents and surgery. However, these methods do not achieved the desired therapeutic effects (Sharifi et al., 2015).
Degeneration and calcification of the CEPs can significantly affect the biomechanics and nutrient supply of the IVD, which is a crucial factor in the onset and progression of IVDD (Ashinsky et al., 2020). The CEPs are a layer of hydrated cartilaginous tissue, with chondrocytes as the exclusive cell type (Wang et al., 2016). Various risk factors, including mechanical overload, injury, instability, chemical or genetic disorders and smoking may contribute to the inflammation of CEPs and oxidative stress in chondrocytes, resulting in reduced cell viability and CEP degeneration (Vergroesen et al., 2015; Dowdell et al., 2017). Furthermore, oxidative stress may cause mitochondrial dysfunction and morphological disruption, activating the mitochondrial apoptotic pathway and leading to apoptosis of endplate chondrocytes (He R. et al., 2018). Mitochondrial autophagy is crucial for maintaining mitochondrial homeostasis as it removes damaged mitochondria from the cell and decreases mitochondrial ROS production (Song et al., 2021). In addition to mitochondria-dependent apoptosis, iron death, which is characterized by mitochondrial shrinkage, enhanced mitochondrial membrane density, lipid peroxidation and the involvement of a unique set of genes, decreases cell viability in the NP and is thought to be involved in IVDD (Yang et al., 2021).
Recent mouse experiments have shown (Shao et al., 2022) that the anti-inflammatory and antioxidant activities of ICA prevent CEP degeneration and calcification in the pathology of IVDD. The Nrf-2/HO-1 pathway, activated by ICA, facilitates Parkin-mediated mitosis and inhibits iron-related apoptosis in chondrocytes, which in turn alleviates redox imbalance and mitochondrial dysfunction, ultimately improving cell survival. Hua et al. demonstrated that ICA has a protective effect against H2O2-induced oxidative damage and mitochondria-mediated apoptosis in human NP cells through modulating the Nrf-2 signaling pathway (Hua et al., 2020). Although targeting Nrf2 to regulate lipid peroxidation and iron death is a potential intervention strategy, the specific mechanisms involved require further investigation.
The activation of endogenous stem cells for the restoration and reconstruction of IVDD is considered a superior biological therapy (Huang et al., 2017). ICA enhances the proliferation and growth of neural stem cells in development by regulating cell cycle-related genes and proteins (Fu et al., 2018). Zhang et al. found that ICA can increase the expression of chemotactic cytokines, such as IGF-1, TGF-β, SDF-1 and CCL-5, promoting the migration of intervertebral disc region-derived stem cells to repair degenerated IVDs (Zhang Z. et al., 2022). ICA is a novel and potent long-acting pain reliever that inhibits the overexpression of CINC-1/IL-8; at 100 mg/kg/d, ICA demonstrated analgesic effectiveness similar to that of celecoxib, but with longer-lasting efficacy (Li et al., 2020).
3.5 Fracture
Fractures are among the most common types of bone and joint injuries. Most patients with fractures heal naturally rather than by accelerated healing, which is an extremely slow process of bone remodeling (Dimitriou et al., 2011). Bone remodeling includes both bone resorption and formation, which control the remodeling or replacement of bone after an injury, such as a fracture, as well as microinjuries that occur during normal activity (Wei and Cooper, 2023). During bone resorption, active osteoclasts secrete acids and enzymes to break down and resorb the mineralized bone matrix, reducing the amount of bone in the region. During bone formation, osteoblasts continue to differentiate to form new bone tissue (Frost, 2004). Therefore, the search for drugs or interventions that regulate bone remodeling is essential.
Research has shown that ICA is a potential bone-inducing compound for bone tissue engineering (Zhao et al., 2010), and oral administration of ICA promotes bone formation by inhibiting bone resorption and increases peak bone mineral density (BMD) and bone mass (Yang et al., 2014). Bone formation markers (such as osteocalcin (OC) and bone-specific alkaline phosphatase (BAP)) and bone resorption markers, including telopeptides of type 1 collagen (C-terminal: CTX-1 and N-terminal: NTX-1), and serum band 5 tartrate-resistant acid phosphatase (TRACP-5) are low in the blood under normal conditions (Froberg et al., 1999). A study showed that the higher the levels of OC in the bones and blood of rats modeled for femur fractures, the faster the fracture healed (Aydin et al., 2014). Zhang et al. found that ICA upregulated serum levels of OC, BAP, NTX-1 and CTX-1, and activated the BMP-2/Smad5/Runx2 signaling pathway to accelerate fracture healing in relatively young and aged rats (Zhang et al., 2021). In vivo experiments showed (Cao et al., 2017) that post-fracture administration of ICA accelerated mineralization and osteogenesis and attenuated BMD loss in osteopenic rats. ICA also promotes MC3T3-E1 cell growth and regulates osteoblast differentiation (Lai et al., 2018). In a rat model of femoral fracture, ICA treatment significantly improved bone mineral density and histopathological outcomes at both the early and late fracture stages (Gürbüz et al., 2019).
A recent study created a titanium dioxide nanotube-polydopamine-ICA (TNT-DP-ICA) drug delivery platform and evaluated its in vitro and in vivo biological properties. They found that TNT-DP-ICA showed reduced inflammatory response in vitro, and after 1 month and 3 months of implantation in fracture rats, the fibrous capsule around the implant decreased, the thickness of newly formed bone tissue increased, and the osseointegration process was significantly improved (Negrescu et al., 2022). Huang et al. designed an antibiotic release system comprising ICA, vancomycin (VA) and calcium phosphate cement to treat infectious bone disease in patients with open fractures (Huang et al., 2013). They discovered that combining ICA with VA could overcome the negative effect of bacteria on bone healing and achieve adequate local drug concentrations. Although further studies are necessary to confirm the therapeutic effectiveness of this strategy for infectious bone diseases, this discovery suggests the potential use of ICA in combination with antibiotics to treat this condition.
3.6 Other diseases
In addition to the five aforementioned diseases, ICA exerts therapeutic effects in other bone and joint disorders. One study investigated the role of ICA in a rat model of monosodium urate (MSU)-induced gouty arthritis. MSU increased the degree of ankle swelling, promoted inflammatory cell infiltration, and elevated cytokine levels in rat synovial tissues, all of which were alleviated by ICA (Cao, 2021). Ankylosing spondylitis (AS) is an inflammatory rheumatic disease characterized by spinal inflammation and new bone formation (Kim and Lee, 2023). ICA was found to induce apoptosis of CD4+ T cells in patients with AS, thereby exerting anti-rheumatic activity (Wang H. et al., 2017). Additionally, ICA inhibits the expression of fibroblast-specific transcription factors Cbfa1 and Osx via the cytokine BMP-2, thus suppressing fibroblast ossification (Jia et al., 2014). Emerging evidence indicates that ICA may regulate bone metabolism and remodel the tumor microenvironment. Osteosarcoma (OS) is the most common primary skeletal malignancy. Ren et al. demonstrated that ICA inhibits the Wnt/β-catenin signaling pathway and downregulates the expression of downstream phosphorylated p-GSK3β, β-catenin, c-Myc, and cyclin D1, thereby suppressing OS cell proliferation and promoting apoptosis (Ren et al., 2018). ICA also enhances OS sensitivity to the antitumor antibiotic Adriamycin by downregulating multidrug resistance protein 1 (MDR1) and the PI3K/AKT pathways, which may provide novel insights into OS chemotherapy (Wang et al., 2015). Moreover, ICA has been identified as a promising immunomodulator for metastatic prostate cancer (PCa) bone metastasis, as it significantly reduces PCa metastasis-induced bone destruction in vivo by inhibiting osteoclastogenesis through downregulation of the TAM/CCL5 pathway (Chen et al., 2023). Recent advances in novel drug delivery systems have highlighted the potential of ICA-loaded biomaterials in bone tissue engineering due to their osteogenic properties. Liu et al. investigated the effect of ICA on the inhibition of peri-implantitis-associated bacteria and developed a calcium phosphate cement composite loaded with ICA-gelatin microspheres, which not only promoted osteoinductivity and bone formation, but also alleviated inflammation (Liu N. et al., 2022). Avascular necrosis of the femoral head is a common hip disease (Ahmed et al., 2023); 3D-printed porous Ti6Al4V reconstruction rods loaded with ICA can effectively promote osteogenesis and neovascularization and interfere with further development of the necrosis (Lei et al., 2023).
4 Current status of clinical studies
The efficacy of ICA in bone and joint disease has been demonstrated in numerous clinical trials (Table 1). ICA has been shown to reduce the incidence of osteoporosis in postmenopausal women. The results of a 24-month randomized, double-blind, placebo-controlled clinical trial demonstrated that phytoestrogenic flavonoids (ICA, genistein and daidzein) derived from Epimedium exerted a beneficial effect on preventing bone loss in late postmenopausal women without inducing significant endometrial hyperplasia (Zhang et al., 2007). In particular, long-term administration (up to 12–24 months) of ICA products led to a time-dependent improvement in lumbar spine and femoral neck BMD. However, the effect of ICA on maintaining BMD was relatively modest (Zhang et al., 2007). In another randomized, double-blind, controlled clinical trial of OP patients (360 cases in the Epimedium total flavonoid capsule group and 120 cases in the Gusongbao capsule group), the overall efficacy rates of the main symptoms in the Epimedium total flavonoid capsule group and the control group were 90.83% and 75.00%, respectively, and the rates of BMD improvement were 47.38% and 34.23%. This suggests that Epimedium Total Flavonoid Capsule may increase BMD and improve major symptoms in OP patients (Min et al., 2013). The reported adverse events included rash, constipation, diarrhea, palpitations, tinnitus and gastrointestinal dysfunction, with an incidence of 6.67% (Min et al., 2013). Although ICA has been evaluated in rodent models of bone loss, to date, no studies have reported its efficacy in non-rodent models (Wang et al., 2018). Therefore, further large-scale clinical trials and novel drug delivery systems are needed to explore the efficacy of ICA and its derivatives in bone and joint disorders. Notably, various materials have been developed as scaffolds for ICA delivery, including chitosan/nanohydroxyapatite, porous PHBV (poly(3-hydroxybutyrate-co-3-hydroxyvalerate)) and gelatin/hyaluronic acid composite microspheres (Fan et al., 2012; Xia et al., 2013; Yan et al., 2016), though additional investigations are still required.
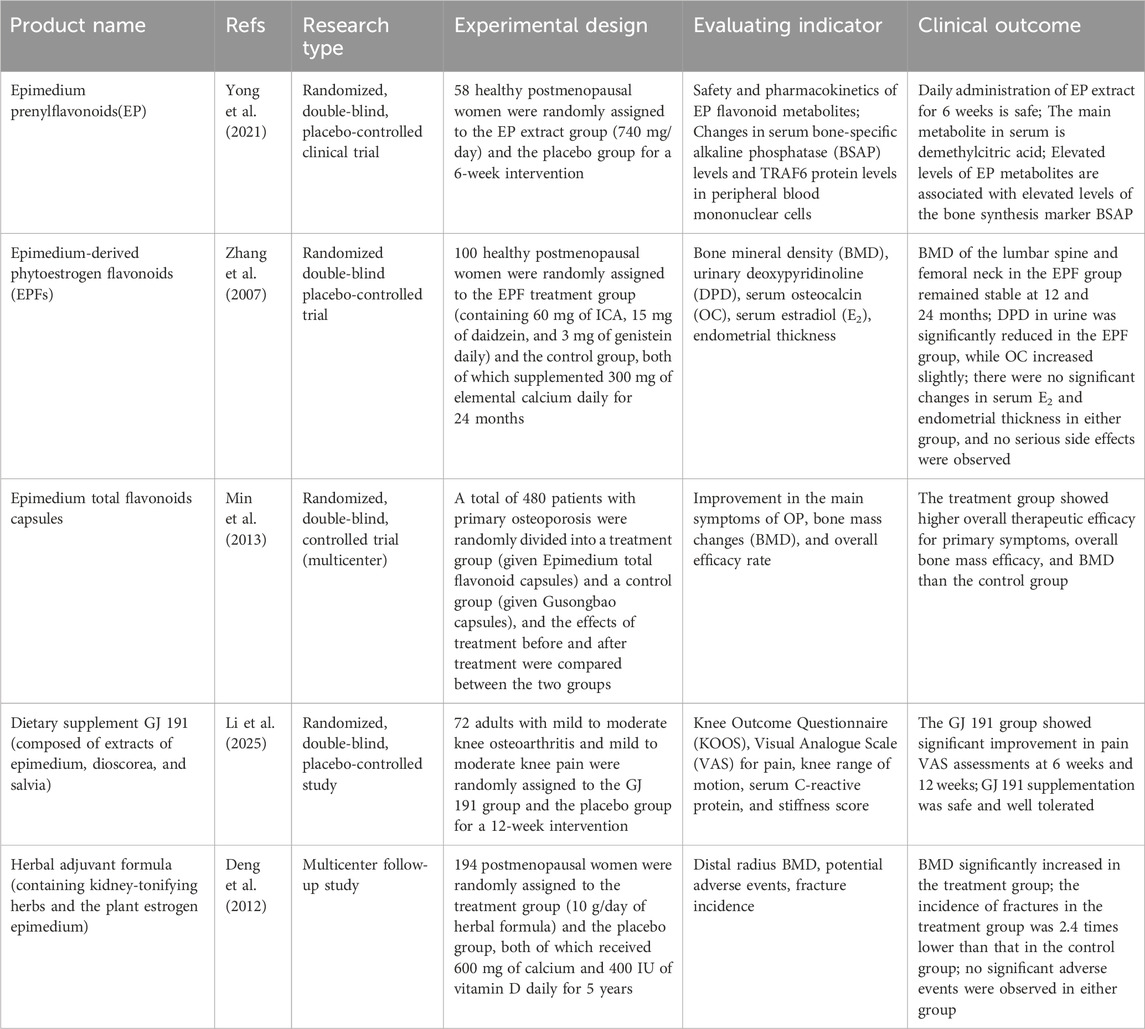
Table 1. Current clinical studies and results of ICA.
5 Toxic side effects
Epimedium, as a Chinese herbal medicine, has been insufficiently studied regarding its systemic toxicity and adverse reactions. Sui et al. evaluated the safety profile of Epimedium aqueous extract through experiments including the mouse bone marrow micronucleus test, the Ames test, and the TK gene mutation test. Results showed that Epimedium is non-mutagenic, with an LD50 exceeding 80 g/kg; its IC50 values in Chinese hamster ovary cells and lung cells were 55.4 and 19.53 mg/mL, respectively, and all toxicity tests yielded negative results (Sui et al., 2006). Recently, a Chinese herbal formula containing 70% Epimedium was reported to cause acute liver failure: two female osteoporosis patients exhibited significant elevations in alanine aminotransferase and aspartate aminotransferase after several months of administration (Pei and Cao, 2009; Zheng et al., 2014). Cheng et al. also found that vomiting, nausea, and decreased locomotion were observed in mice after 3 days of Epimedium administration (Cheng and Cai, 2000). Although epimedium is associated with adverse reactions, the toxic side effects of ICA are negligible. Studies have shown that when NIH mice were gavaged with Epimedium total flavonoids at 36 g/kg/day for 7 days (1440 times the standard human dose), no mortality was observed (Dongmei, 2007). In a normal Wistar rat model, long-term toxicity of Epimedium total flavonoids was evaluated via gavage of the extract at 410 g/kg/day for 12 weeks; no significant differences in any analyzed blood parameters or major pathological changes were found in treated rats (Li et al., 2008). In addition, Zhong et al. evaluated the toxicity changes of Epimedium flavonoids in zebrafish and showed that ICA had negligible toxicity to zebrafish embryos (Zhong et al., 2019). However, other studies have found that ICA can cause developmental toxicity by disrupting thyroid development and hormone synthesis, but this is time- and dose-dependent (Wu et al., 2023). One study showed that ICA protected DNA from excessive oxidative stress in a model of AAPH (2,2′-azobis(2-amidinopropane) dihydrochloride)-induced DNA oxidative damage (Zhao et al., 2007). Notably, the concentrations used (10–5 to 10–4 M) exhibited clear cytotoxicity, suggesting controversy regarding the extent of ICA-mediated DNA protection against oxidative damage in cellular and animal models. Furthermore, ICA induces osteogenic differentiation of human BMMSCs in a dose-dependent manner, with an optimal in vitro concentration range of 10–9 to 10–5 M; concentrations exceeding 10–5 M are cytotoxic (Fan et al., 2011). In summary, no known warnings or contraindications for the use of Epimedium species currently exist, and the optimal dosage remains unclear. Even fewer toxicological studies on ICA have been reported, but at least in the context of bone and joint disorders, ICA has not been found to exhibit significant toxic side effects.
In summary, we have explored the therapeutic role of ICA for various bone and joint diseases as much as possible, but the efficacy and safety of ICA in these diseases are still lacking analysis. To be sure, ICA as a potential drug for bone tissue engineering has been shown to treat bone and joint diseases through multiple pathways and targets, and the related clinical symptoms have been improved with clear efficacy. Nevertheless, head-to-head comparisons of ICA’s efficacy and safety across these diseases are scarce, necessitating more in-depth investigations. In addition, given that ICA is frequently used in combination with other herbs and drugs, understanding its potential drug-drug interactions is critical. These interactions involve not only phase I P450 enzyme genes and drug transporters but also phase II metabolic genes (Li et al., 2012; Zhou et al., 2012; Zhu et al., 2012). In humans, the aromatase cytochrome P450 (CYP19A1) catalyzes the production of estrogen from C19 androgens (Simpson et al., 2002). Aromatase activity is a key factor in bone development and mineralization and is essential for estrogen production in bone (Ribot et al., 2006). Phosphodiesterase 5 (PDE5) inhibitors have been shown to increase aromatase expression and estrogen biosynthesis in human adipocytes and ovarian granulosa cells (Li et al., 2017).
Notably, the PDE5 inhibitor sildenafil and ICA analogues promote osteoblast differentiation by activating the cyclic adenosine monophosphate (cAMP)/protein kinase G (PKG)/Src homology 2 domain-containing tyrosine phosphatase 2 (SHP2) pathway, thereby stimulating aromatase expression (Wisanwattana et al., 2021), providing new insights into the role of estrogen biosynthesis in the treatment of OP. In addition, a related study showed that ICA decreased Cyp2e1 enzyme activity in mice (Min et al., 2009) but induced Cyp3a enzyme activity in rats. Recently, ICA has been shown to inhibit UDP-glucuronosyltransferases, particularly Ugt1 family enzymes, in vitro. Xu et al. found that ICA had no inhibitory effect on the expression of P450 and Ugts genes in vivo, but that ICA induced the expression of Cyp4a14 in the liver of mice and resulted in a slight increase in the expression of Ugt2b1, Ugt2b5 and Ugt2b36 (Xu et al., 2014). These studies exemplify the potential drug-drug interactions of ICA, but the mechanisms involved are still poorly understood, and more high-quality clinical studies of natural herbal medicines with anabolic and anti-catabolic effects are needed in the future.
6 Discussion
Epimedium is a medicinal plant used in various herbal formulas and modern proprietary Chinese medicinal products. Recent studies have indicated that ICA is the most abundant constituent of Epimedium and possesses a wide range of pharmacological activities, including anti-osteoporotic, immunomodulatory, antioxidant, osteoprotective and anti-inflammatory effects. Owing to these properties, ICA has been identified as a potentially beneficial compound for bone protection and treatment of bone damage. This study reviewed the molecular mechanisms of ICA in the treatment of bone and joint diseases. Reducing the expression of inflammatory mediators, preventing oxidative stress and apoptosis, inhibiting FLS proliferation and migration, promoting osteoblast differentiation, and decreasing osteoclast activity are important mechanisms for the treatment of bone and joint diseases using ICA (Table 2). The relevant major pathways and transforming factors include RANKL, IL-1β, TNF-α, NF-κB/HIF-2α pathway, PI3K/Akt pathway, Rho A- TAZ pathway, BMP-2/Smad5/Runx2 pathway, ERα, Nrf2, AMPK and STAT3. Research has shown that ICA, a natural drug, has numerous advantages over surgical and nonsurgical interventions, such as having multiple mechanisms and targets, affordability, low toxicity and minimal side effects.
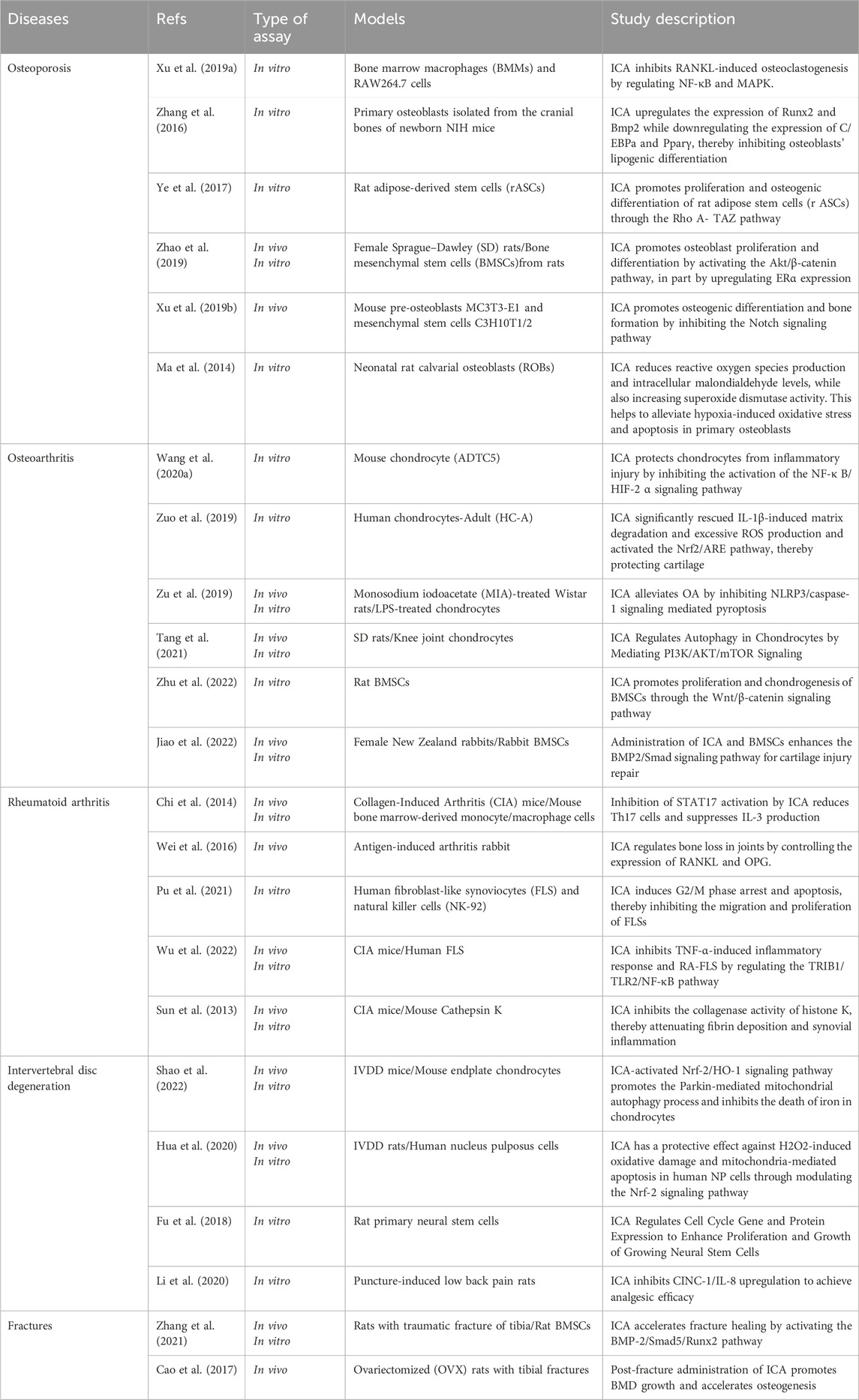
Table 2. Literature Reports on icariin in the treatment of bone and joint diseases.
However, many challenges remain regarding the application of ICA in bone and joint diseases. Currently, relevant studies are mostly conducted in cellular and animal models and human clinical trials are scarce, suggesting that the clinical translation of ICA is not imminent. According to the biopharmaceutical classification system, ICA is classified as a class IV drug with low solubility, permeability and bioavailability (Dong et al., 2021). These unfavorable physicochemical and pharmacokinetic factors severely limit its clinical application. Therefore, there is an urgent need to improve the bioavailability and clinical efficacy of ICA. ICA promotes bone formation and bone protection by acting on a variety of proteins or genes; thus, it is essential to further investigate its structure and possible direct targets in cells to promote targeted therapy for various bone diseases. In terms of toxicology, there is a lack of robust evidence from in vivo or in vitro studies on the chronic, acute and long-term toxicity of ICA, making it difficult to predict the safety and adverse events associated with its use in bone and joint disorders. Larger sample sizes are needed for preclinical and clinical systematic investigation of the side effects and efficacy of ICA. Because ICA and its various derivatives have varying degrees of potency in osteogenesis, work remains to be done to determine which compounds have the strongest osteogenic activity, which is challenging at this time because the full details of their mechanisms of action are unknown and comparative studies are lacking.
This article outlines the potential of plant-derived ICA in the treatment of bone and joint disorders; however, it has several limitations. First, although ICA has shown promising results in cellular and animal experiments for treating bone diseases, only a small number of relevant clinical trials have been retrieved, and evidence from many of these trials is lacking to demonstrate the safety and therapeutic potential of the molecule in patients with bone diseases. Second, most studies cited in this review had small sample sizes, which may have introduced variability and limited the generalizability of their findings. Therefore, large-scale randomized controlled trials are needed to address these limitations and strengthen the evidence base. Third, many gaps need to be addressed in the development of novel drug delivery systems for the treatment of bone and joint diseases. It has been reported that ICA-nanocarrier coupling has good results in vitro; however, in vivo experiments are needed for further validation (Wang Q. et al., 2017; Zhang et al., 2019). Future research needs to focus on the following aspects: (1) in-depth study of the mechanism of ICA in the treatment of orthopedic diseases, especially involving certain hotspots, such as intestinal flora, iron death, Exos and stem cells; (2) optimizing the drug properties, dosage and dosing regimen of ICA to improve its stability and efficacy in vivo while minimizing toxicity and side effects; (3) exploring the relationship between ICA and the common therapeutic strategies for bone diseases (drugs and surgery) when used in combination; (4) Further develop novel drug delivery systems such as nanoparticles and liposomes using ICA as a carrier, or combine ICA with other materials, including polymers and hydrogels, to improve its bioavailability and promote clinical application; (5) To identify the direct molecular targets of ICA for the treatment of bone diseases by using cyberpharmacology or experimental validation to enhance its targeted therapeutic effect.
7 Conclusion
In recent years, Chinese medicine has demonstrated unique efficacy in the treatment of bone diseases. This paper presents a thorough review of the mechanisms of action of ICA in different types of bone diseases. Existing research provides scientific evidence that ICA, a natural compound, may serve as a promising supplement in the prevention and treatment of various bone and joint diseases, attributed to its diverse pharmacological activities, including anti-inflammatory, antioxidative, osteogenic, anti-osteoclastic, and anti-apoptotic effects. Although ICA has beneficial effects on bone health, few studies have extended beyond animal models, and a large number of laboratory studies and clinical trials are needed to confirm its efficacy. Further research may establish ICA as a promising drug candidate for the treatment of bone and joint diseases.
Author contributions
JL: Project administration, Writing – original draft, Writing – review and editing, Conceptualization, Data curation, Methodology. LW: Conceptualization, Investigation, Methodology, Writing – original draft, Writing – review and editing. YX: Formal Analysis, Writing – review and editing. JZ: Validation, Writing – review and editing. XZ: Validation, Writing – review and editing. XW: Formal Analysis, Supervision, Writing – original draft, Writing – review and editing.
Funding
The author(s) declare that no financial support was received for the research and/or publication of this article.
Acknowledgements
The author would like to thank Xiaoyu Du for his valuable feedback on this article. The image is created by biorender.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Ahmed, A., Mahon, J., and Karkuri, A. (2023). Readability of online information on core decompression of the hip for avascular necrosis. Cureus 15 (12), e50298. doi:10.7759/cureus.50298
Ansari, M. Y., Ahmad, N., and Haqqi, T. M. (2020). Oxidative stress and inflammation in osteoarthritis pathogenesis: role of polyphenols. Biomed. Pharmacother. 129, 110452. doi:10.1016/j.biopha.2020.110452
Ashinsky, B. G., Bonnevie, E. D., Mandalapu, S. A., Pickup, S., Wang, C., Han, L., et al. (2020). Intervertebral disc degeneration is associated with aberrant endplate remodeling and reduced small molecule transport. J. Bone Min. Res. 35 (8), 1572–1581. doi:10.1002/jbmr.4009
Aydin, A., Halici, Z., Akoz, A., Karaman, A., Ferah, I., Bayir, Y., et al. (2014). Treatment with α-lipoic acid enhances the bone healing after femoral fracture model of rats. Naunyn Schmiedeb. Arch. Pharmacol. 387 (11), 1025–1036. doi:10.1007/s00210-014-1021-1
Ballane, G., Cauley, J. A., Luckey, M. M., and El-Hajj Fuleihan, G. (2017). Worldwide prevalence and incidence of osteoporotic vertebral fractures. Osteoporos. Int. 28 (5), 1531–1542. doi:10.1007/s00198-017-3909-3
Barrett-Connor, E., Mosca, L., Collins, P., Geiger, M. J., Grady, D., Kornitzer, M., et al. (2006). Effects of raloxifene on cardiovascular events and breast cancer in postmenopausal women. N. Engl. J. Med. 355 (2), 125–137. doi:10.1056/NEJMoa062462
Böcker, W., Doobaree, I. U., Khachatryan, A., Dornstauder, E., Bartsch, R., Worth, G., et al. (2022). Fractures in untreated patients with osteoporosis in Germany: an InGef healthcare insurance database analysis. Osteoporos. Int. 33 (1), 77–86. doi:10.1007/s00198-021-06051-w
Bottini, N., and Firestein, G. S. (2013). Duality of fibroblast-like synoviocytes in RA: passive responders and imprinted aggressors. Nat. Rev. Rheumatol. 9 (1), 24–33. doi:10.1038/nrrheum.2012.190
Buhrmann, C., Honarvar, A., Setayeshmehr, M., Karbasi, S., Shakibaei, M., and Valiani, A. (2020). Herbal remedies as potential in cartilage tissue engineering: an overview of new therapeutic approaches and strategies. Molecules 25 (13), 3075. doi:10.3390/molecules25133075
Cao, Y. (2021). Icariin alleviates MSU-Induced rat GA models through NF-κB/NALP3 pathway. Cell Biochem. Funct. 39 (3), 357–366. doi:10.1002/cbf.3598
Cao, H., Zhang, Y., Qian, W., Guo, X. P., Sun, C., Zhang, L., et al. (2017). Effect of icariin on fracture healing in an ovariectomized rat model of osteoporosis. Exp. Ther. Med. 13 (5), 2399–2404. doi:10.3892/etm.2017.4233
Chen, C., Xie, J., Rajappa, R., Deng, L., Fredberg, J., and Yang, L. (2015). Interleukin-1β and tumor necrosis factor-α increase stiffness and impair contractile function of articular chondrocytes. Acta Biochim. Biophys. Sin. (Shanghai) 47 (2), 121–129. doi:10.1093/abbs/gmu116
Chen, G., Wang, C., Wang, J., Yin, S., Gao, H., Xiang, L. U., et al. (2016). Antiosteoporotic effect of icariin in ovariectomized rats is mediated via the Wnt/β-catenin pathway. Exp. Ther. Med. 12 (1), 279–287. doi:10.3892/etm.2016.3333
Chen, C., Wang, S., Wang, N., Zheng, Y., Zhou, J., Hong, M., et al. (2023). Icariin inhibits prostate cancer bone metastasis and destruction via suppressing TAM/CCL5-mediated osteoclastogenesis. Phytomedicine 120, 155076. doi:10.1016/j.phymed.2023.155076
Cheng, J., and Cai, H. (2000). Adverse reactions to zhuangguguanjie wan and cause analysis. Adverse Drug React. J. 1, 15–19.
Cheng, K., Chen, K. M., Ge, B. F., Zhen, P., Gao, Y. H., and Ma, H. P. (2014). Comparison research with icariin and genistein by anti-inflammatory reaction and angiogenesis pathway to inhibit bone loss on ovariectomized rats. Zhong Yao Cai 37 (4), 627–631.
Chi, L., Gao, W., Shu, X., and Lu, X. (2014). A natural flavonoid glucoside, icariin, regulates Th17 and alleviates rheumatoid arthritis in a murine model. Mediat. Inflamm. 2014, 392062. doi:10.1155/2014/392062
Chung, B. H., Kim, J. D., Kim, C. K., Kim, J. H., Won, M. H., Lee, H. S., et al. (2008). Icariin stimulates angiogenesis by activating the MEK/ERK- and PI3K/Akt/eNOS-dependent signal pathways in human endothelial cells. Biochem. Biophys. Res. Commun. 376 (2), 404–408. doi:10.1016/j.bbrc.2008.09.001
Deng, W. M., Zhang, P., Huang, H., Shen, Y. G., Yang, Q. H., Cui, W. L., et al. (2012). Five-year follow-up study of a kidney-tonifying herbal fufang for prevention of postmenopausal osteoporosis and fragility fractures. J. Bone Min. Metab. 30 (5), 517–524. doi:10.1007/s00774-012-0351-7
Dimitriou, R., Jones, E., McGonagle, D., and Giannoudis, P. V. (2011). Bone regeneration: current concepts and future directions. BMC Med. 9, 66. doi:10.1186/1741-7015-9-66
Ding, K., Hua, F., and Ding, W. (2020). Gut microbiome and osteoporosis. Aging Dis. 11 (2), 438–447. doi:10.14336/ad.2019.0523
Ding, N., Li, E., Ouyang, X., Guo, J., and Wei, B. (2021). The therapeutic potential of bone marrow mesenchymal stem cells for articular cartilage regeneration in osteoarthritis. Curr. Stem Cell Res. Ther. 16 (7), 840–847. doi:10.2174/1574888x16666210127130044
Dong, M., Wu, S., Xu, H., Yu, X., Wang, L., Bai, H., et al. (2021). FBS-derived exosomes as a natural nano-scale carrier for icariin promote osteoblast proliferation. Front. Bioeng. Biotechnol. 9, 615920. doi:10.3389/fbioe.2021.615920
Dongmei, L. (2007). Experimental study on acute toxicity with total flavonoids of epimedium in mice. China Pharm.
Dowdell, J., Erwin, M., Choma, T., Vaccaro, A., Iatridis, J., and Cho, S. K. (2017). Intervertebral disk degeneration and repair. Neurosurgery 80 (3s), S46–s54. doi:10.1093/neuros/nyw078
Duan, R., Xie, H., and Liu, Z. Z. (2020). The role of autophagy in osteoarthritis. Front. Cell Dev. Biol. 8, 608388. doi:10.3389/fcell.2020.608388
Fan, J. J., Cao, L. G., Wu, T., Wang, D. X., Jin, D., Jiang, S., et al. (2011). The dose-effect of icariin on the proliferation and osteogenic differentiation of human bone mesenchymal stem cells. Molecules 16 (12), 10123–10133. doi:10.3390/molecules161210123
Fan, J., Bi, L., Wu, T., Cao, L., Wang, D., Nan, K., et al. (2012). A combined chitosan/nano-size hydroxyapatite system for the controlled release of icariin. J. Mater Sci. Mater Med. 23 (2), 399–407. doi:10.1007/s10856-011-4491-4
Fleisch, H. (2003). Bisphosphonates in osteoporosis. Eur. Spine J. 12 (2 Suppl. ment), S142–S146. doi:10.1007/s00586-003-0622-z
Froberg, M. K., Garg, U. C., Stroncek, D. F., Geis, M., McCullough, J., and Brown, D. M. (1999). Changes in serum osteocalcin and bone-specific alkaline phosphatase are associated with bone pain in donors receiving granulocyte-colony-stimulating factor for peripheral blood stem and progenitor cell collection. Transfusion 39 (4), 410–414. doi:10.1046/j.1537-2995.1999.39499235675.x
Frost, H. M. (2004). A 2003 update of bone physiology and Wolff's law for clinicians. Angle Orthod. 74 (1), 3–15. doi:10.1043/0003-3219(2004)074<0003:Auobpa>2.0.Co;2
Fu, X., Li, S., Zhou, S., Wu, Q., Jin, F., and Shi, J. (2018). Stimulatory effect of icariin on the proliferation of neural stem cells from rat hippocampus. BMC Complement. Altern. Med. 18 (1), 34. doi:10.1186/s12906-018-2095-y
Gao, Y., Shi, W., Tu, C., Li, P., Zhao, G., Xiao, X., et al. (2022). Immunostimulatory activity and structure-activity relationship of epimedin B from Epimedium brevicornu maxim. Front. Pharmacol. 13, 1015846. doi:10.3389/fphar.2022.1015846
Goldring, M. B., and Goldring, S. R. (2010). Articular cartilage and subchondral bone in the pathogenesis of osteoarthritis. Ann. N. Y. Acad. Sci. 1192, 230–237. doi:10.1111/j.1749-6632.2009.05240.x
Goldring, M. B., and Marcu, K. B. (2009). Cartilage homeostasis in health and rheumatic diseases. Arthritis Res. Ther. 11 (3), 224. doi:10.1186/ar2592
Guo, M., Pang, X., Xu, Y., Jiang, W., Liao, B., Yu, J., et al. (2022). Plastid genome data provide new insights into the phylogeny and evolution of the genus epimedium. J. Adv. Res. 36, 175–185. doi:10.1016/j.jare.2021.06.020
Gürbüz, K., Yerer, M. B., Gürbüz, P., and Halıcı, M. (2019). Icariin promotes early and late stages of fracture healing in rats. Eklem Hast. Cerrahisi 30 (3), 282–288. doi:10.5606/ehc.2019.66796
He, J. P., Feng, X., Wang, J. F., Shi, W. G., Li, H., Danilchenko, S., et al. (2018a). Icariin prevents bone loss by inhibiting bone resorption and stabilizing bone biological apatite in a hindlimb suspension rodent model. Acta Pharmacol. Sin. 39 (11), 1760–1767. doi:10.1038/s41401-018-0040-8
He, R., Cui, M., Lin, H., Zhao, L., Wang, J., Chen, S., et al. (2018b). Melatonin resists oxidative stress-induced apoptosis in nucleus pulposus cells. Life Sci. 199, 122–130. doi:10.1016/j.lfs.2018.03.020
He, C., Wang, Z., and Shi, J. (2020). Pharmacological effects of icariin. Adv. Pharmacol. 87, 179–203. doi:10.1016/bs.apha.2019.10.004
Hua, W., Li, S., Luo, R., Wu, X., Zhang, Y., Liao, Z., et al. (2020). Icariin protects human nucleus pulposus cells from hydrogen peroxide-induced mitochondria-mediated apoptosis by activating nuclear factor erythroid 2-related factor 2. Biochim. Biophys. Acta Mol. Basis Dis. 1866 (1), 165575. doi:10.1016/j.bbadis.2019.165575
Huang, J., Yuan, L., Wang, X., Zhang, T. L., and Wang, K. (2007). Icaritin and its glycosides enhance osteoblastic, but suppress osteoclastic, differentiation and activity in vitro. Life Sci. 81 (10), 832–840. doi:10.1016/j.lfs.2007.07.015
Huang, J. G., Pang, L., Chen, Z. R., and Tan, X. P. (2013). Dual-delivery of vancomycin and icariin from an injectable calcium phosphate cement-release system for controlling infection and improving bone healing. Mol. Med. Rep. 8 (4), 1221–1227. doi:10.3892/mmr.2013.1624
Huang, Z., Zhang, L., Feng, X., Chen, T., and Bi, S. (2017). A new in vivo method to retard progression of intervertebral disc degeneration through stimulation of endogenous stem cells with simvastatin. Med. Hypotheses 101, 65–66. doi:10.1016/j.mehy.2017.02.014
Huang, M., Wang, Y., and Peng, R. (2020). Icariin alleviates glucocorticoid-induced osteoporosis through EphB4/Ephrin-B2 axis. Evid. Based Complement. Altern. Med. 2020, 2982480. doi:10.1155/2020/2982480
Hwang, H. S., and Kim, H. A. (2015). Chondrocyte apoptosis in the pathogenesis of osteoarthritis. Int. J. Mol. Sci. 16 (11), 26035–26054. doi:10.3390/ijms161125943
Imagama, S., Ando, K., Kobayashi, K., Seki, T., Hamada, T., Machino, M., et al. (2020). Impact of comorbidity rates of lumbar spondylosis, knee osteoarthritis, and osteoporosis on physical QOL and risk factors for poor physical QOL in middle-aged and elderly people. Mod. Rheumatol. 30 (2), 402–409. doi:10.1080/14397595.2019.1601839
Jia, M., Nie, Y., Cao, D. P., Xue, Y. Y., Wang, J. S., Zhao, L., et al. (2012). Potential antiosteoporotic agents from plants: a comprehensive review. Evid. Based Complement. Altern. Med. 2012, 364604. doi:10.1155/2012/364604
Jia, C., Liu, H., Li, M., Wu, Z., and Feng, X. (2014). Effects of icariin on cytokine-induced ankylosing spondylitis with fibroblastic osteogenesis and its molecular mechanism. Int. J. Clin. Exp. Pathol. 7 (12), 9104–9109.
Jiang, H., Ji, P., Shang, X., and Zhou, Y. (2023). Connection between osteoarthritis and nitric oxide: from pathophysiology to therapeutic target. Molecules 28 (4), 1683. doi:10.3390/molecules28041683
Jiao, F., Tang, W., Wang, J., Liu, D., Zhang, H., and Tang, D. (2022). Icariin promotes the repair of bone marrow mesenchymal stem cells in rabbit knee cartilage defects via the BMP/smad pathway. Ann. Transl. Med. 10 (12), 691. doi:10.21037/atm-22-2515
Jin, F., Gong, Q. H., Xu, Y. S., Wang, L. N., Jin, H., Li, F., et al. (2014). Icariin, a phosphodiesterase-5 inhibitor, improves learning and memory in APP/PS1 transgenic mice by stimulation of NO/cGMP signalling. Int. J. Neuropsychopharmacol. 17 (6), 871–881. doi:10.1017/s1461145713001533
Jin, J., Wang, H., Hua, X., Chen, D., Huang, C., and Chen, Z. (2019). An outline for the pharmacological effect of icariin in the nervous system. Eur. J. Pharmacol. 842, 20–32. doi:10.1016/j.ejphar.2018.10.006
Jing, X., Du, T., Chen, K., Guo, J., Xiang, W., Yao, X., et al. (2019). Icariin protects against iron overload-induced bone loss via suppressing oxidative stress. J. Cell Physiol. 234 (7), 10123–10137. doi:10.1002/jcp.27678
Justesen, J., Stenderup, K., Ebbesen, E. N., Mosekilde, L., Steiniche, T., and Kassem, M. (2001). Adipocyte tissue volume in bone marrow is increased with aging and in patients with osteoporosis. Biogerontology 2 (3), 165–171. doi:10.1023/a:1011513223894
Karimian, A., Ahmadi, Y., and Yousefi, B. (2016). Multiple functions of p21 in cell cycle, apoptosis and transcriptional regulation after DNA damage. DNA Repair (Amst) 42, 63–71. doi:10.1016/j.dnarep.2016.04.008
Khan, A., Khan, S. U., Khan, A., Shal, B., Rehman, S. U., Rehman, S. U., et al. (2022). Anti-inflammatory and anti-rheumatic potential of selective plant compounds by targeting TLR-4/AP-1 signaling: a comprehensive molecular docking and simulation approaches. Molecules 27 (13), 4319. doi:10.3390/molecules27134319
Kim, S. H., and Lee, S. H. (2023). Updates on ankylosing spondylitis: pathogenesis and therapeutic agents. J. Rheum. Dis. 30 (4), 220–233. doi:10.4078/jrd.2023.0041
Kim, J. Y., and Park, J. H. (2003). ROS-Dependent caspase-9 activation in hypoxic cell death. FEBS Lett. 549 (1-3), 94–98. doi:10.1016/s0014-5793(03)00795-6
Kim, B., Lee, K. Y., and Park, B. (2018). Icariin abrogates osteoclast formation through the regulation of the RANKL-mediated TRAF6/NF-κB/ERK signaling pathway in Raw264.7 cells. Phytomedicine 51, 181–190. doi:10.1016/j.phymed.2018.06.020
Kusumbe, A. P., Ramasamy, S. K., and Adams, R. H. (2014). Coupling of angiogenesis and osteogenesis by a specific vessel subtype in bone. Nature 507 (7492), 323–328. doi:10.1038/nature13145
Lai, Y., Cao, H., Wang, X., Chen, S., Zhang, M., Wang, N., et al. (2018). Porous composite scaffold incorporating osteogenic phytomolecule icariin for promoting skeletal regeneration in challenging osteonecrotic bone in rabbits. Biomaterials 153, 1–13. doi:10.1016/j.biomaterials.2017.10.025
Lefèvre, S., Knedla, A., Tennie, C., Kampmann, A., Wunrau, C., Dinser, R., et al. (2009). Synovial fibroblasts spread rheumatoid arthritis to unaffected joints. Nat. Med. 15 (12), 1414–1420. doi:10.1038/nm.2050
Lei, H., Zhou, Z., Liu, L., Gao, C., Su, Z., Tan, Z., et al. (2023). Icariin-loaded 3D-printed porous Ti6Al4V reconstruction rods for the treatment of necrotic femoral heads. Acta Biomater. 169, 625–640. doi:10.1016/j.actbio.2023.07.057
Li, D. M., Yin, X. F., Liu, J. H., and Cai, D. W. (2008). Experimental study on long term toxicity of total flavonoids of epimedium.
Li, L., Hu, H., Xu, S., Zhou, Q., and Zeng, S. (2012). Roles of UDP-glucuronosyltransferases in phytochemical metabolism of herbal medicines and the associated herb-drug interactions. Curr. Drug Metab. 13 (5), 615–623. doi:10.2174/1389200211209050615
Li, C., Li, Q., Mei, Q., and Lu, T. (2015a). Pharmacological effects and pharmacokinetic properties of icariin, the major bioactive component in herba epimedii. Life Sci. 126, 57–68. doi:10.1016/j.lfs.2015.01.006
Li, F., Dong, H. X., Gong, Q. H., Wu, Q., Jin, F., and Shi, J. S. (2015b). Icariin decreases both APP and Aβ levels and increases neurogenesis in the brain of Tg2576 mice. Neuroscience 304, 29–35. doi:10.1016/j.neuroscience.2015.06.010
Li, F., Du, B. W., Lu, D. F., Wu, W. X., Wongkrajang, K., Wang, L., et al. (2017). Flavonoid glycosides isolated from Epimedium brevicornum and their estrogen biosynthesis-promoting effects. Sci. Rep. 7 (1), 7760. doi:10.1038/s41598-017-08203-7
Li, Y., Wu, Y., Jiang, K., Han, W., Zhang, J., Xie, L., et al. (2019). Mangiferin prevents TBHP-induced apoptosis and ECM degradation in mouse osteoarthritic chondrocytes via restoring autophagy and ameliorates murine osteoarthritis. Oxid. Med. Cell Longev. 2019, 8783197. doi:10.1155/2019/8783197
Li, J., Luo, M., Wang, S., Jin, G., Han, Z., Ma, Y., et al. (2020). Icariin ameliorates lower back pain in rats via suppressing the secretion of cytokine-induced neutrophil Chemoatractant-1. Biomed. Res. Int. 2020, 4670604. doi:10.1155/2020/4670604
Li, Z., Li, D., Su, H., Xue, H., Tan, G., and Xu, Z. (2022). Autophagy: an important target for natural products in the treatment of bone metabolic diseases. Front. Pharmacol. 13, 999017. doi:10.3389/fphar.2022.999017
Li, Z., Li, D., Chen, R., Gao, S., Xu, Z., and Li, N. (2023). Cell death regulation: a new way for natural products to treat osteoporosis. Pharmacol. Res. 187, 106635. doi:10.1016/j.phrs.2022.106635
Li, F., Zhu, F., Wang, S., Hu, H., Zhang, D., He, Z., et al. (2024a). Icariin alleviates cisplatin-induced premature ovarian failure by inhibiting ferroptosis through activation of the Nrf2/ARE pathway. Sci. Rep. 14 (1), 17318. doi:10.1038/s41598-024-67557-x
Li, Y., Wei, Z. F., and Su, L. (2024b). Anti-aging effects of icariin and the underlying mechanisms: a mini-review. Aging Med. Milt. 7 (1), 90–95. doi:10.1002/agm2.12284
Li, L., Wang, Y., Wang, S., Zong, J., Zhang, Z., Zou, S., et al. (2025). A randomized, double-blind, placebo-controlled study investigating the safety and efficacy of a herbal formulation on knee joint function in adults with knee osteoarthritis. J. Integr. Complement. Med. 31 (1), 54–63. doi:10.1089/jicm.2023.0124
Liang, J., Liu, J., Sun, W., Li, P., Sun, H., Cao, W., et al. (2025). Study on the screening of anti-osteoporosis components in processed epimedium-based on spectrum-effect analysis. Chem. Biodivers., e02936. doi:10.1002/cbdv.202402936
Lin, Y., Chen, W. W., Ding, B., Guo, M., Liang, M., Pang, H., et al. (2023). Highly efficient bioconversion of icariin to icaritin by whole-cell catalysis. Microb. Cell Fact. 22 (1), 64. doi:10.1186/s12934-023-02068-4
Liu, T., Qin, X. C., Li, W. R., Zhou, F., Li, G. Y., Xin, H., et al. (2011). Effects of icariin and icariside II on eNOS expression and NOS activity in porcine aorta endothelial cells. Beijing Da Xue Xue Bao Yi Xue Ban. 43 (4), 500–504.
Liu, Z., Zhang, M., Shen, Z., Ke, J., Zhang, D., and Yin, F. (2020). Efficacy and safety of 18 anti-osteoporotic drugs in the treatment of patients with osteoporosis caused by glucocorticoid: a network meta-analysis of randomized controlled trials. PLoS One 15 (12), e0243851. doi:10.1371/journal.pone.0243851
Liu, J., Cheng, Q., Wu, X., Zhu, H., Deng, X., Wang, M., et al. (2022a). Icariin treatment rescues diabetes induced bone loss via scavenging ROS and activating primary Cilia/Gli2/Osteocalcin signaling pathway. Cells 11 (24), 4091. doi:10.3390/cells11244091
Liu, N., Huang, S., Guo, F., Zhai, S., Wang, D., Li, F., et al. (2022b). Calcium phosphate cement with icariin-loaded gelatin microspheres as a local drug delivery system for bone regeneration. Biomed. Eng. Online 21 (1), 89. doi:10.1186/s12938-022-01052-0
Liu, Y., Yang, H., Xiong, J., Zhao, J., Guo, M., Chen, J., et al. (2023). Icariin as an emerging candidate drug for anticancer treatment: current status and perspective. Biomed. Pharmacother. 157, 113991. doi:10.1016/j.biopha.2022.113991
Long, F. (2011). Building strong bones: molecular regulation of the osteoblast lineage. Nat. Rev. Mol. Cell Biol. 13 (1), 27–38. doi:10.1038/nrm3254
Luo, H., and Zhang, R. (2021). Icariin enhances cell survival in lipopolysaccharide-induced synoviocytes by suppressing ferroptosis via the Xc-/GPX4 axis. Exp. Ther. Med. 21 (1), 72. doi:10.3892/etm.2020.9504
Luo, Y., Zhang, Y., and Huang, Y. (2018). Icariin reduces cartilage degeneration in a mouse model of osteoarthritis and is associated with the changes in expression of Indian hedgehog and parathyroid hormone-related protein. Med. Sci. Monit. 24, 6695–6706. doi:10.12659/msm.910983
Ma, Y., Sato, M., Vahle, J., Long, G., Sandusky, G., and Westmore, M. (2004). Bone neoplasms in F344 rats given teriparatide [rhPTH(1-34)] are dependent on duration of treatment and dose. Toxicol. Pathol. 32 (4), 426–438. doi:10.1080/01926230490462138
Ma, H., He, X., Yang, Y., Li, M., Hao, D., and Jia, Z. (2011a). The genus epimedium: an ethnopharmacological and phytochemical review. J. Ethnopharmacol. 134 (3), 519–541. doi:10.1016/j.jep.2011.01.001
Ma, H. P., Ming, L. G., Ge, B. F., Zhai, Y. K., Song, P., Xian, C. J., et al. (2011b). Icariin is more potent than genistein in promoting osteoblast differentiation and mineralization in vitro. J. Cell Biochem. 112 (3), 916–923. doi:10.1002/jcb.23007
Ma, H. P., Ma, X. N., Ge, B. F., Zhen, P., Zhou, J., Gao, Y. H., et al. (2014). Icariin attenuates hypoxia-induced oxidative stress and apoptosis in osteoblasts and preserves their osteogenic differentiation potential in vitro. Cell Prolif. 47 (6), 527–539. doi:10.1111/cpr.12147
Maffia, P., Brewer, J. M., Gracie, J. A., Ianaro, A., Leung, B. P., Mitchell, P. J., et al. (2004). Inducing experimental arthritis and breaking self-tolerance to joint-specific antigens with trackable, ovalbumin-specific T cells. J. Immunol. 173 (1), 151–156. doi:10.4049/jimmunol.173.1.151
Manolagas, S. C., O'Brien, C. A., and Almeida, M. (2013). The role of estrogen and androgen receptors in bone health and disease. Nat. Rev. Endocrinol. 9 (12), 699–712. doi:10.1038/nrendo.2013.179
Martel-Pelletier, J., Barr, A. J., Cicuttini, F. M., Conaghan, P. G., Cooper, C., Goldring, M. B., et al. (2016). Osteoarthritis. Nat. Rev. Dis. Prim. 2, 16072. doi:10.1038/nrdp.2016.72
McCormick, F., Harris, J. D., Abrams, G. D., Frank, R., Gupta, A., Hussey, K., et al. (2014). Trends in the surgical treatment of articular cartilage lesions in the United States: an analysis of a large private-payer database over a period of 8 years. Arthroscopy 30 (2), 222–226. doi:10.1016/j.arthro.2013.11.001
McInnes, I. B., Buckley, C. D., and Isaacs, J. D. (2016). Cytokines in rheumatoid arthritis - shaping the immunological landscape. Nat. Rev. Rheumatol. 12 (1), 63–68. doi:10.1038/nrrheum.2015.171
Min, W., An-Sheng, S., Qin, W. U., Li-Mei, Y. U., Xiao-Long, X., Qi-Hai, G., et al. (2009). Effects icariin on the contents of liver microsomal cytochrome P450 and the activities of CYP2E1. West China J. Pharm. Sci.
Min, L. U., Linhua, W., and Yiwen, L. (2013). Treatment of primary osteoporosis with epimedium total flavone capsule: a multicenter clinical observation on 360 cases. Chin. J. Osteoporos. 19 (3), 279–274.
Ming, L. G., Chen, K. M., and Xian, C. J. (2013). Functions and action mechanisms of flavonoids genistein and icariin in regulating bone remodeling. J. Cell Physiol. 228 (3), 513–521. doi:10.1002/jcp.24158
Miura, T., and Tanno, M. (2010). Mitochondria and GSK-3beta in cardioprotection against ischemia/reperfusion injury. Cardiovasc Drugs Ther. 24 (3), 255–263. doi:10.1007/s10557-010-6234-z
Mukwaya, E., Xu, F., Wong, M. S., and Zhang, Y. (2014). Chinese herbal medicine for bone health. Pharm. Biol. 52 (9), 1223–1228. doi:10.3109/13880209.2014.884606
Nagashima, M., Takahashi, H., Shimane, K., Nagase, Y., and Wauke, K. (2012). Osteogenesis and osteoclast inhibition in rheumatoid arthritis patients treated with bisphosphonates alone or in combination with pitavastatin over an 18-month follow-up after more than 4 years of treatment with bisphosphonates. Arthritis Res. Ther. 14 (5), R224. doi:10.1186/ar4063
Nakashima, T., Hayashi, M., and Takayanagi, H. (2012). New insights into osteoclastogenic signaling mechanisms. Trends Endocrinol. Metab. 23 (11), 582–590. doi:10.1016/j.tem.2012.05.005
Negrescu, A. M., Mitran, V., Draghicescu, W., Popescu, S., Pirvu, C., Ionascu, I., et al. (2022). TiO(2) nanotubes functionalized with icariin for an attenuated in vitro immune response and improved in vivo osseointegration. J. Funct. Biomater. 13 (2), 43. doi:10.3390/jfb13020043
Ogando, J., Tardáguila, M., Díaz-Alderete, A., Usategui, A., Miranda-Ramos, V., Martínez-Herrera, D. J., et al. (2016). Notch-regulated miR-223 targets the aryl hydrocarbon receptor pathway and increases cytokine production in macrophages from rheumatoid arthritis patients. Sci. Rep. 6, 20223. doi:10.1038/srep20223
Papapetrou, P. D. (2009). Bisphosphonate-associated adverse events. Hormones 8 (2), 96–110. doi:10.14310/horm.2002.1226
Pei, D., and Cao, Y. (2009). Acute drug-induced liver injury caused by xianlinggubao in one patient. Chin. Hepatol. 14, 457–458.
Pu, L., Meng, Q., Li, S., Liu, B., and Li, F. (2021). Icariin arrests cell cycle progression and induces cell apoptosis through the mitochondrial pathway in human fibroblast-like synoviocytes. Eur. J. Pharmacol. 912, 174585. doi:10.1016/j.ejphar.2021.174585
Ren, Y., Zhu, F., and Liu, Z. (2018). Inhibitory effect of icariin on osteosarcoma cell proliferation via the Wnt/β-catenin signaling pathway. Oncol. Lett. 16 (2), 1405–1410. doi:10.3892/ol.2018.8821
Ribot, C., Trémollieres, F., and Pouillés, J. M. (2006). Aromatase and regulation of bone remodeling. Jt. Bone Spine 73 (1), 37–42. doi:10.1016/j.jbspin.2005.02.005
Richards, J. B., Rivadeneira, F., Inouye, M., Pastinen, T. M., Soranzo, N., Wilson, S. G., et al. (2008). Bone mineral density, osteoporosis, and osteoporotic fractures: a genome-wide association study. Lancet 371 (9623), 1505–1512. doi:10.1016/s0140-6736(08)60599-1
Risbud, M. V., and Shapiro, I. M. (2014). Role of cytokines in intervertebral disc degeneration: pain and disc content. Nat. Rev. Rheumatol. 10 (1), 44–56. doi:10.1038/nrrheum.2013.160
Ruderman, E. M. (2012). Overview of safety of non-biologic and biologic DMARDs. Rheumatol. Oxf. 51 (Suppl. 6), vi37–vi43. doi:10.1093/rheumatology/kes283
Salminen-Mankonen, H. J., Morko, J., and Vuorio, E. (2007). Role of cathepsin K in normal joints and in the development of arthritis. Curr. Drug Targets 8 (2), 315–323. doi:10.2174/138945007779940188
Shahmarvand, N., Nagy, A., Shahryari, J., and Ohgami, R. S. (2018). Mutations in the signal transducer and activator of transcription family of genes in cancer. Cancer Sci. 109 (4), 926–933. doi:10.1111/cas.13525
Shao, Y., Sun, L., Yang, G., Wang, W., Liu, X., Du, T., et al. (2022). Icariin protects vertebral endplate chondrocytes against apoptosis and degeneration via activating Nrf-2/HO-1 pathway. Front. Pharmacol. 13, 937502. doi:10.3389/fphar.2022.937502
Sharifi, S., Bulstra, S. K., Grijpma, D. W., and Kuijer, R. (2015). Treatment of the degenerated intervertebral disc; closure, repair and regeneration of the annulus fibrosus. J. Tissue Eng. Regen. Med. 9 (10), 1120–1132. doi:10.1002/term.1866
Shen, P., Guo, B. L., Gong, Y., Hong, D. Y., Hong, Y., and Yong, E. L. (2007a). Taxonomic, genetic, chemical and estrogenic characteristics of epimedium species. Phytochemistry 68 (10), 1448–1458. doi:10.1016/j.phytochem.2007.03.001
Shen, P., Wong, S. P., and Yong, E. L. (2007b). Sensitive and rapid method to quantify icaritin and desmethylicaritin in human serum using gas chromatography-mass spectrometry. J. Chromatogr. B Anal. Technol. Biomed. Life Sci. 857 (1), 47–52. doi:10.1016/j.jchromb.2007.06.026
Simpson, E. R., Clyne, C., Rubin, G., Boon, W. C., Robertson, K., Britt, K., et al. (2002). Aromatase--a brief overview. Annu. Rev. Physiol. 64, 93–127. doi:10.1146/annurev.physiol.64.081601.142703
Song, C., Wang, J., Song, Q., Li, X., Chen, Z., Ma, Q., et al. (2008). Simvastatin induces estrogen receptor-alpha (ER-alpha) in murine bone marrow stromal cells. J. Bone Min. Metab. 26 (3), 213–217. doi:10.1007/s00774-007-0820-6
Song, L., Zhao, J., Zhang, X., Li, H., and Zhou, Y. (2013). Icariin induces osteoblast proliferation, differentiation and mineralization through estrogen receptor-mediated ERK and JNK signal activation. Eur. J. Pharmacol. 714 (1-3), 15–22. doi:10.1016/j.ejphar.2013.05.039
Song, Y., Lu, S., Geng, W., Feng, X., Luo, R., Li, G., et al. (2021). Mitochondrial quality control in intervertebral disc degeneration. Exp. Mol. Med. 53 (7), 1124–1133. doi:10.1038/s12276-021-00650-7
Song, S., Guo, Y., Yang, Y., and Fu, D. (2022). Advances in pathogenesis and therapeutic strategies for osteoporosis. Pharmacol. Ther. 237, 108168. doi:10.1016/j.pharmthera.2022.108168
Stergar, J., Gradisnik, L., Velnar, T., and Maver, U. (2019). Intervertebral disc tissue engineering: a brief review. Bosn. J. Basic Med. Sci. 19 (2), 130–137. doi:10.17305/bjbms.2019.3778
Sugiyama, T., Kim, Y. T., and Oda, H. (2015). Osteoporosis therapy: a novel insight from natural homeostatic system in the skeleton. Osteoporos. Int. 26 (2), 443–447. doi:10.1007/s00198-014-2923-y
Sui, H., Gao, P., and Xu, H. (2006). The safety evaluation of herba epimedii water extract. Carcinogenesis,Teratogenesis & Mutagen. 18 (6), 439–442.
Sun, P., Liu, Y., Deng, X., Yu, C., Dai, N., Yuan, X., et al. (2013). An inhibitor of cathepsin K, icariin suppresses cartilage and bone degradation in mice of collagen-induced arthritis. Phytomedicine 20 (11), 975–979. doi:10.1016/j.phymed.2013.04.019
Sun, K., Luo, J., Guo, J., Yao, X., Jing, X., and Guo, F. (2020). The PI3K/AKT/mTOR signaling pathway in osteoarthritis: a narrative review. Osteoarthr. Cartil. 28 (4), 400–409. doi:10.1016/j.joca.2020.02.027
Szabó, R., Rácz, C. P., and Dulf, F. V. (2022). Bioavailability improvement strategies for icariin and its derivates: a review. Int. J. Mol. Sci. 23 (14), 7519. doi:10.3390/ijms23147519
Takayanagi, H., Kim, S., Matsuo, K., Suzuki, H., Suzuki, T., Sato, K., et al. (2002). RANKL maintains bone homeostasis through c-Fos-dependent induction of interferon-beta. Nature 416 (6882), 744–749. doi:10.1038/416744a
Tan, O. J., Loo, H. L., Thiagarajah, G., Palanisamy, U. D., and Sundralingam, U. (2021). Improving oral bioavailability of medicinal herbal compounds through lipid-based formulations - a scoping review. Phytomedicine 90, 153651. doi:10.1016/j.phymed.2021.153651
Tang, Y., Li, Y., Xin, D., Chen, L., Xiong, Z., and Yu, X. (2021). Icariin alleviates osteoarthritis by regulating autophagy of chondrocytes by mediating PI3K/AKT/mTOR signaling. Bioengineered 12 (1), 2984–2999. doi:10.1080/21655979.2021.1943602
Tang, W., Zhang, H., Liu, D., and Jiao, F. (2022). Icariin accelerates cartilage defect repair by promoting chondrogenic differentiation of BMSCs under conditions of oxygen-glucose deprivation. J. Cell Mol. Med. 26 (1), 202–215. doi:10.1111/jcmm.17073
Teitelbaum, S. L. (2000). Bone resorption by osteoclasts. Science 289 (5484), 1504–1508. doi:10.1126/science.289.5484.1504
Tonomura, H., Nagae, M., Takatori, R., Ishibashi, H., Itsuji, T., and Takahashi, K. (2020). The potential role of hepatocyte growth factor in degenerative disorders of the synovial joint and spine. Int. J. Mol. Sci. 21 (22), 8717. doi:10.3390/ijms21228717
Vergroesen, P. P., Kingma, I., Emanuel, K. S., Hoogendoorn, R. J., Welting, T. J., van Royen, B. J., et al. (2015). Mechanics and biology in intervertebral disc degeneration: a vicious circle. Osteoarthr. Cartil. 23 (7), 1057–1070. doi:10.1016/j.joca.2015.03.028
Wang, W., and Yeung, K. W. K. (2017). Bone grafts and biomaterials substitutes for bone defect repair: a review. Bioact. Mater 2 (4), 224–247. doi:10.1016/j.bioactmat.2017.05.007
Wang, Z. C., Sun, H. J., Li, K. H., Fu, C., and Liu, M. Z. (2014). Icariin promotes directed chondrogenic differentiation of bone marrow mesenchymal stem cells but not hypertrophy in vitro. Exp. Ther. Med. 8 (5), 1528–1534. doi:10.3892/etm.2014.1950
Wang, Z., Yang, L., Xia, Y., Guo, C., and Kong, L. (2015). Icariin enhances cytotoxicity of doxorubicin in human multidrug-resistant osteosarcoma cells by inhibition of ABCB1 and down-regulation of the PI3K/Akt pathway. Biol. Pharm. Bull. 38 (2), 277–284. doi:10.1248/bpb.b14-00663
Wang, F., Cai, F., Shi, R., Wang, X. H., and Wu, X. T. (2016). Aging and age related stresses: a senescence mechanism of intervertebral disc degeneration. Osteoarthr. Cartil. 24 (3), 398–408. doi:10.1016/j.joca.2015.09.019
Wang, H., Jiang, Q., and Feng, X. (2017a). Effect of icariin on apoptosis and expression of fas, fas ligand, B cell lymphoma, and Bcl-2-associated X protein in CD4+ T lymphocytes from patients with ankylosing spondylitis. J. Tradit. Chin. Med. 37 (2), 207–213. doi:10.1016/s0254-6272(17)30046-8
Wang, Q., Cao, L., Liu, Y., Zheng, A., Wu, J., Jiang, X., et al. (2017b). Evaluation of synergistic osteogenesis between icariin and BMP2 through a micro/meso hierarchical porous delivery system. Int. J. Nanomedicine 12, 7721–7735. doi:10.2147/ijn.S141052
Wang, Z., Wang, D., Yang, D., Zhen, W., Zhang, J., and Peng, S. (2018). The effect of icariin on bone metabolism and its potential clinical application. Osteoporos. Int. 29 (3), 535–544. doi:10.1007/s00198-017-4255-1
Wang, P., Meng, Q., Wang, W., Zhang, S., Xiong, X., Qin, S., et al. (2020a). Icariin inhibits the inflammation through down-regulating NF-κB/HIF-2α signal pathways in chondrocytes. Biosci. Rep. 40 (11). doi:10.1042/bsr20203107
Wang, S., Deng, Z., Ma, Y., Jin, J., Qi, F., Li, S., et al. (2020b). The role of autophagy and mitophagy in bone metabolic disorders. Int. J. Biol. Sci. 16 (14), 2675–2691. doi:10.7150/ijbs.46627
Wang, G., Zhang, L., Shen, H., Hao, Q., Fu, S., and Liu, X. (2021). Up-regulation of long non-coding RNA CYTOR induced by icariin promotes the viability and inhibits the apoptosis of chondrocytes. BMC Complement. Med. Ther. 21 (1), 152. doi:10.1186/s12906-021-03322-1
Wang, J., Huang, R., Liang, Q., and Zhang, Y. (2022a). The complete chloroplast genome of epimedium muhuangense (berberidaceae). Mitochondrial DNA B Resour. 7 (5), 870–872. doi:10.1080/23802359.2022.2071650
Wang, S., Wang, H., Feng, C., Li, C., Li, Z., He, J., et al. (2022b). The regulatory role and therapeutic application of pyroptosis in musculoskeletal diseases. Cell Death Discov. 8 (1), 492. doi:10.1038/s41420-022-01282-0
Wang, S., Wang, S., Wang, X., Xu, Y., Zhang, X., Han, Y., et al. (2022c). Effects of icariin on modulating gut microbiota and regulating metabolite alterations to prevent bone loss in ovariectomized rat model. Front. Endocrinol. (Lausanne) 13, 874849. doi:10.3389/fendo.2022.874849
Wang, W., Sun, D. F., Dong, Z., and Zhang, W. L. (2024). Icariin suppresses osteogenic differentiation and promotes bone regeneration in porphyromonas gingivalis-infected conditions through EphA2-RhoA signaling pathway. Int. Immunopharmacol. 143 (Pt 1), 113302. doi:10.1016/j.intimp.2024.113302
Wang, S., Wang, H., Zhang, Y., Yin, G., Zhang, X., and Zhang, F. (2025). Epimedium and its chemical constituents in cancer treatment: a comprehensive review of traditional applications, antitumor effects, pharmacokinetics, delivery systems, and toxicology. J. Ethnopharmacol. 347, 119738. doi:10.1016/j.jep.2025.119738
Watanabe, Y., Tsuchiya, A., and Terai, S. (2021). The development of mesenchymal stem cell therapy in the present, and the perspective of cell-free therapy in the future. Clin. Mol. Hepatol. 27 (1), 70–80. doi:10.3350/cmh.2020.0194
Wei, X., and Cooper, D. M. L. (2023). The various meanings and uses of bone “remodeling” in biological anthropology: a review. Am. J. Biol. Anthropol. 182 (2), 318–329. doi:10.1002/ajpa.24825
Wei, C. C., Ping, D. Q., You, F. T., Qiang, C. Y., and Tao, C. (2016). Icariin prevents cartilage and bone degradation in experimental models of arthritis. Mediat. Inflamm. 2016, 9529630. doi:10.1155/2016/9529630
Wisanwattana, W., Wongkrajang, K., Cao, D. Y., Shi, X. K., Zhang, Z. H., Zhou, Z. Y., et al. (2021). Inhibition of phosphodiesterase 5 promotes the aromatase-mediated estrogen biosynthesis in osteoblastic cells by activation of cGMP/PKG/SHP2 pathway. Front. Endocrinol. (Lausanne) 12, 636784. doi:10.3389/fendo.2021.636784
Wong, S. P., Shen, P., Lee, L., Li, J., and Yong, E. L. (2009). Pharmacokinetics of prenylflavonoids and correlations with the dynamics of estrogen action in sera following ingestion of a standardized epimedium extract. J. Pharm. Biomed. Anal. 50 (2), 216–223. doi:10.1016/j.jpba.2009.04.022
Wu, H., Lien, E. J., and Lien, L. L. (2003). Chemical and pharmacological investigations of epimedium species: a survey. Prog. Drug Res. 60, 1–57. doi:10.1007/978-3-0348-8012-1_1
Wu, Z. M., Luo, J., Shi, X. D., Zhang, S. X., Zhu, X. B., and Guo, J. (2020). Icariin alleviates rheumatoid arthritis via regulating miR-223-3p/NLRP3 signalling axis. Autoimmunity 53 (8), 450–458. doi:10.1080/08916934.2020.1836488
Wu, Z. M., Xiang, Y. R., Zhu, X. B., Shi, X. D., Chen, S., Wan, X., et al. (2022). Icariin represses the inflammatory responses and survival of rheumatoid arthritis fibroblast-like synoviocytes by regulating the TRIB1/TLR2/NF-kB pathway. Int. Immunopharmacol. 110, 108991. doi:10.1016/j.intimp.2022.108991
Wu, M., Zheng, N., Zhan, X., He, J., Xiao, M., Zuo, Z., et al. (2023). Icariin induces developmental toxicity via thyroid hormone disruption in zebrafish larvae. Food Chem. Toxicol. 182, 114155. doi:10.1016/j.fct.2023.114155
Xia, L., Li, Y., Zhou, Z., Dai, Y., Liu, H., and Liu, H. (2013). Icariin delivery porous PHBV scaffolds for promoting osteoblast expansion in vitro. Mater Sci. Eng. C Mater Biol. Appl. 33 (6), 3545–3552. doi:10.1016/j.msec.2013.04.050
Xiao, J., Luo, C., Li, A., Cai, F., Wang, Y., Pan, X., et al. (2024). Icariin inhibits chondrocyte ferroptosis and alleviates osteoarthritis by enhancing the SLC7A11/GPX4 signaling. Int. Immunopharmacol. 133, 112010. doi:10.1016/j.intimp.2024.112010
Xu, S. F., Jin, T., Lu, Y. F., Li, L. S., Liu, J., and Shi, J. S. (2014). Effect of icariin on UDP-Glucuronosyltransferases in mouse liver. Planta Med. 80 (5), 387–392. doi:10.1055/s-0034-1368265
Xu, S., Yu, J., Zhan, J., Yang, L., Guo, L., and Xu, Y. (2017). Pharmacokinetics, tissue distribution, and metabolism study of icariin in rat. Biomed. Res. Int. 2017, 4684962. doi:10.1155/2017/4684962
Xu, Q., Chen, G., Liu, X., Dai, M., and Zhang, B. (2019a). Icariin inhibits RANKL-induced osteoclastogenesis via modulation of the NF-κB and MAPK signaling pathways. Biochem. Biophys. Res. Commun. 508 (3), 902–906. doi:10.1016/j.bbrc.2018.11.201
Xu, Y., Li, L., Tang, Y., Yang, J., Jin, Y., and Ma, C. (2019b). Icariin promotes osteogenic differentiation by suppressing notch signaling. Eur. J. Pharmacol. 865, 172794. doi:10.1016/j.ejphar.2019.172794
Xu, H., Zhou, S., Qu, R., Yang, Y., Gong, X., Hong, Y., et al. (2020). Icariin prevents oestrogen deficiency-induced alveolar bone loss through promoting osteogenesis via STAT3. Cell Prolif. 53 (2), e12743. doi:10.1111/cpr.12743
Yan, H., Zhou, Z., Huang, T., Peng, C., Liu, Q., Zhou, H., et al. (2016). Controlled release in vitro of icariin from gelatin/hyaluronic acid composite microspheres. Polym. Bull. 73 (4), 1055–1066. doi:10.1007/s00289-015-1534-x
Yan, F., Liu, J., Chen, M. X., Zhang, Y., Wei, S. J., Jin, H., et al. (2023). Icariin ameliorates memory deficits through regulating brain insulin signaling and glucose transporters in 3×Tg-AD mice. Neural Regen. Res. 18 (1), 183–188. doi:10.4103/1673-5374.344840
Yang, L., Yu, Z., Qu, H., and Li, M. (2014). Comparative effects of hispidulin, genistein, and icariin with estrogen on bone tissue in ovariectomized rats. Cell Biochem. Biophys. 70 (1), 485–490. doi:10.1007/s12013-014-9945-0
Yang, A., Yu, C., Lu, Q., Li, H., Li, Z., and He, C. (2019). Mechanism of action of icariin in bone marrow mesenchymal stem cells. Stem Cells Int. 2019, 5747298. doi:10.1155/2019/5747298
Yang, R. Z., Xu, W. N., Zheng, H. L., Zheng, X. F., Li, B., Jiang, L. S., et al. (2021). Involvement of oxidative stress-induced annulus fibrosus cell and nucleus pulposus cell ferroptosis in intervertebral disc degeneration pathogenesis. J. Cell Physiol. 236 (4), 2725–2739. doi:10.1002/jcp.30039
Yang, Q., Liu, S., Liu, H., Liu, Y., He, Z., Zheng, Z., et al. (2026). A new paradigm in bone tissue biomaterials: enhanced osteogenesis-angiogenic coupling by targeting H-type blood vessels. Biomaterials 324, 123423. doi:10.1016/j.biomaterials.2025.123423
Ye, Y., Jing, X., Li, N., Wu, Y., Li, B., and Xu, T. (2017). Icariin promotes proliferation and osteogenic differentiation of rat adipose-derived stem cells by activating the RhoA-TAZ signaling pathway. Biomed. Pharmacother. 88, 384–394. doi:10.1016/j.biopha.2017.01.075
Yong, E. L., Cheong, W. F., Huang, Z., Thu, W. P. P., Cazenave-Gassiot, A., Seng, K. Y., et al. (2021). Randomized, double-blind, placebo-controlled trial to examine the safety, pharmacokinetics and effects of epimedium prenylflavonoids, on bone specific alkaline phosphatase and the osteoclast adaptor protein TRAF6 in post-menopausal women. Phytomedicine 91, 153680. doi:10.1016/j.phymed.2021.153680
Zeng, Z., Zhou, X., Wang, Y., Cao, H., Guo, J., Wang, P., et al. (2022). Mitophagy-A new target of bone disease. Biomolecules 12 (10), 1420. doi:10.3390/biom12101420
Zhang, G., Qin, L., and Shi, Y. (2007). Epimedium-derived phytoestrogen flavonoids exert beneficial effect on preventing bone loss in late postmenopausal women: a 24-month randomized, double-blind and placebo-controlled trial. J. Bone Min. Res. 22 (7), 1072–1079. doi:10.1359/jbmr.070405
Zhang, X., Liu, T., Huang, Y., Wismeijer, D., and Liu, Y. (2014). Icariin: does it have an osteoinductive potential for bone tissue engineering? Phytother. Res. 28 (4), 498–509. doi:10.1002/ptr.5027
Zhang, D., Fong, C., Jia, Z., Cui, L., Yao, X., and Yang, M. (2016). Icariin stimulates differentiation and suppresses adipocytic transdifferentiation of primary osteoblasts through estrogen receptor-mediated pathway. Calcif. Tissue Int. 99 (2), 187–198. doi:10.1007/s00223-016-0138-2
Zhang, S., Feng, P., Mo, G., Li, D., Li, Y., Mo, L., et al. (2017). Icariin influences adipogenic differentiation of stem cells affected by osteoblast-osteoclast co-culture and clinical research adipogenic. Biomed. Pharmacother. 88, 436–442. doi:10.1016/j.biopha.2017.01.050
Zhang, X., Lin, X., Liu, T., Deng, L., Huang, Y., and Liu, Y. (2019). Osteogenic enhancement between icariin and bone morphogenetic protein 2: a potential osteogenic compound for bone tissue engineering. Front. Pharmacol. 10, 201. doi:10.3389/fphar.2019.00201
Zhang, X., Chen, Y., Zhang, C., Zhang, X., Xia, T., Han, J., et al. (2021). Effects of icariin on the fracture healing in young and old rats and its mechanism. Pharm. Biol. 59 (1), 1245–1255. doi:10.1080/13880209.2021.1972121
Zhang, Y., Li, J., Wang, Y., and Liang, Q. (2022a). Taxonomy of epimedium (berberidaceae) with special reference to Chinese species. Chin. Herb. Med. 14 (1), 20–35. doi:10.1016/j.chmed.2021.12.001
Zhang, Y., Zhang, C., Li, Z., Zeng, C., Xue, Z., Li, E., et al. (2022b). New 8-prenylated quercetin glycosides from the flowers of Epimedium acuminatum and their testosterone production-promoting activities. Front. Chem. 10, 1014110. doi:10.3389/fchem.2022.1014110
Zhang, Z., Qin, F., Feng, Y., Zhang, S., Xie, C., Huang, H., et al. (2022c). Icariin regulates stem cell migration for endogenous repair of intervertebral disc degeneration by increasing the expression of chemotactic cytokines. BMC Complement. Med. Ther. 22 (1), 63. doi:10.1186/s12906-022-03544-x
Zhao, F., Tang, Y. Z., and Liu, Z. Q. (2007). Protective effect of icariin on DNA against radical-induced oxidative damage. J. Pharm. Pharmacol. 59 (12), 1729–1732. doi:10.1211/jpp.59.12.0016
Zhao, J., Ohba, S., Komiyama, Y., Shinkai, M., Chung, U. I., and Nagamune, T. (2010). Icariin: a potential osteoinductive compound for bone tissue engineering. Tissue Eng. Part A 16 (1), 233–243. doi:10.1089/ten.TEA.2009.0165
Zhao, R., Bu, W., and Chen, Y. (2019). Icariin treatment enhanced the skeletal response to exercise in estrogen-deficient rats. Int. J. Environ. Res. Public Health 16 (19), 3779. doi:10.3390/ijerph16193779
Zhao, F., Chen, F., Song, T., Tian, L., Guo, H., Li, D., et al. (2025). One-step strategy for fabricating icariin-encapsulated biomimetic scaffold: orchestrating immune, angiogenic, and osteogenic Cascade for enhanced bone regeneration. Bioact. Mater 52, 271–286. doi:10.1016/j.bioactmat.2025.06.001
Zheng, M., Zhou, Y., and Lv, X. (2014). A case of liver failure caused by combination of xianling gubao capsule and diclofenac sodium sustained-release capsules. Chin. J. Pharmacovigil. 11, 507–508.
Zhong, R., Chen, Y., Ling, J., Xia, Z., Zhan, Y., Sun, E., et al. (2019). The toxicity and metabolism properties of herba epimedii flavonoids on laval and adult zebrafish. Evid. Based Complement. Altern. Med. 2019, 3745051. doi:10.1155/2019/3745051
Zhou, X., Chan, K., and Yeung, J. H. (2012). Herb-drug interactions with danshen (salvia miltiorrhiza): a review on the role of cytochrome P450 enzymes. Drug Metabol. Drug Interact. 27 (1), 9–18. doi:10.1515/dmdi-2011-0038
Zhou, Z., Luo, J., Wang, J., Li, L., and Kong, L. (2015). Simultaneous enrichment and separation of flavonoids from herba epimedii by macroporous resins coupled with preparative chromatographic method. Nat. Prod. Res. 29 (2), 185–188. doi:10.1080/14786419.2014.964704
Zhu, H. M., Qin, L., Garnero, P., Genant, H. K., Zhang, G., Dai, K., et al. (2012). The first multicenter and randomized clinical trial of herbal fufang for treatment of postmenopausal osteoporosis. Osteoporos. Int. 23 (4), 1317–1327. doi:10.1007/s00198-011-1577-2
Zhu, Y., Ye, L., Cai, X., Li, Z., Fan, Y., and Yang, F. (2022). Icariin-loaded hydrogel regulates bone marrow mesenchymal stem cell chondrogenic differentiation and promotes cartilage repair in osteoarthritis. Front. Bioeng. Biotechnol. 10, 755260. doi:10.3389/fbioe.2022.755260
Zhuang, W., Sun, N., Gu, C., Liu, S., Zheng, Y., Wang, H., et al. (2023). A literature review on epimedium, a medicinal plant with promising slow aging properties. Heliyon 9 (11), e21226. doi:10.1016/j.heliyon.2023.e21226
Zu, Y., Mu, Y., Li, Q., Zhang, S. T., and Yan, H. J. (2019). Icariin alleviates osteoarthritis by inhibiting NLRP3-mediated pyroptosis. J. Orthop. Surg. Res. 14 (1), 307. doi:10.1186/s13018-019-1307-6
Zuo, S., Zou, W., Wu, R. M., Yang, J., Fan, J. N., Zhao, X. K., et al. (2019). Icariin alleviates IL-1β-Induced matrix degradation by activating the Nrf2/ARE pathway in human chondrocytes. Drug Des. Devel Ther. 13, 3949–3961. doi:10.2147/dddt.S203094
Glossary
ICA Icariin
BMMSCs Bone marrow mesenchymal stem cells
OP Osteoporosis
OA Osteoarthritis
RA Rheumatoid arthritis
IVDD Intervertebral disc degeneration
TCM Traditional Chinese medicine
ECM Extracellular matrix
RANKL Receptor activator of nuclear factor-κΒ ligand
Osx Osterix
Runx2 Runt-related protein 2
LRP5 Lipoprotein receptor-related protein 5
ESR1 Estrogen receptor α
OPG Osteoprotegerin
NF-κB Nuclear factor-κB
ALP Alkaline phosphatase
Ocn Osteocalcin
ROS Reactive oxygen species
TRACP Tartrate-resistant acid phosphatase
IL-1β Interleukin-1β
TNF-α Tumor necrosis factor-α
mTOR Mammalian target of rapamycin
PI3K Phosphatidylinositol 3-kinase
AMPK AMP-activated protein kinase
FLS Fibroblast-like synoviocytes
LBP Low back pain
CEPs Cartilaginous endplates
NP Nucleus pulposus
BMD Bone mineral density
OC Osteocalcin
OS Osteosarcoma
MDR1 Multidrug resistance protein 1
PCa Prostate cancer
PDE5 Phosphodiesterase 5
Keywords: icariin, natural flavonoids, Osteometabolic balance, inflammation, oxidativestress, mechanisms
Citation: Luo J, Wu L, Xu Y, Zhang J, Zhao X and Wang X (2025) The therapeutic potential of icariin in the intervention of bone and joint diseases through multiple pathways: a narrative review. Front. Pharmacol. 16:1606912. doi: 10.3389/fphar.2025.1606912
Received: 06 April 2025; Accepted: 27 October 2025;
Published: 07 November 2025.
Edited by:
Rui Liu, Chinese Academy of Medical Sciences and Peking Union Medical College, ChinaReviewed by:
Min-Cong He, Guangzhou University of Chinese Medicine, ChinaLiang Kang, First Affiliated Hospital of Anhui Medical University, China
Copyright © 2025 Luo, Wu, Xu, Zhang, Zhao and Wang. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Xiangjin Wang, NzkwMjY1OTIwQHFxLmNvbQ==
†These authors have contributed equally to this work