 Małgorzata Kołtun-Jasion1*
Małgorzata Kołtun-Jasion1* Karolina Czerwiec1
Karolina Czerwiec1 Andrzej Parzonko1Aleksandra Bakiera1
Andrzej Parzonko1Aleksandra Bakiera1 Marcin Ożarowski2
Marcin Ożarowski2 Anna Karolina Kiss1
Anna Karolina Kiss1- 1Department of Pharmaceutical Biology, Faculty of Pharmacy, Medical University of Warsaw, Warsaw, Poland
- 2Institute of Natural Fibres and Medicinal Plants, National Research Institute, Poznań, Poland
Introduction: Rhodiola rosea L., mainly known within the medicinal plant industry as golden root, Arctic root, or rose root, derives its name from its economic significance, distinctive morphology, and restricted geographical distribution. Extracts from Rhodiola rosea roots/rhizomes are widely used across Europe and Asia as ingredients of traditional herbal medicines and dietary supplements, with numerous claims regarding their adaptogenic effects. With the growing demand for pharmaceutical products that relieve stress-related fatigue and exhaustion, driven by technological advancements and increasing psychophysical challenges, R. rosea has become a highly sought-after resource. However, this heightened demand has also increased the risk of adulteration and the proliferation of low-quality products on the market. The reproducible efficacy and quality of R. rosea preparations are largely dependent on the variable content of key active compounds, such as rosavin, which directly influence product quality. The rapid expansion of the dietary supplement market, coupled with insufficient quality verification of products entering the market, underscores the need for rigorous identification and quality assessment of these products.
Methods: This study aimed to perform a phytochemical analysis of 13 dietary supplements claiming to contain R. rosea using HPTLC and LC-MS techniques and to correlate these findings with their anti-inflammatory activity in an LPS-stimulated BV2 microglial cell model, in vitro.
Results: Our study indicates that nearly 60% of the tested preparations did not contain the declared amount of Rhodiola rosea roots/rhizomes or the characteristic marker compounds associated with this species. Furthermore, rosavin was detected in only 9 out of the 13 analyzed products, with 4 of these containing only trace amounts of this marker compound. Misidentification of R. rosea was most frequently observed among tablet and capsule formulations, whereas products in the form of cut raw material exhibited the highest quality. Moreover, rosavin significantly and dose-dependently inhibited the secretion of interleukin-6 (IL-6) and tumor necrosis factor-alpha (TNF-α) in lipopolysaccharide (LPS)-stimulated microglial cells.
Discussion: The identification of R. rosea in only 40% of the preparations underlines that rigorous control and standardisation of herbal supplements are crucial to understanding their therapeutic activity and preventing adulteration.
1 Introduction
Rhodiola rosea L., [Crassulaceae, syn. Sedum rhodiola Vill] is affiliated with a group of perennial shrubs commonly associated with subarctic and alpine areas of eastern Siberia. Currently, Rhodiola rosea is found in hard-to-reach rocky terrains, sea cliffs and mountains of the northern hemisphere, ranging from the low Arctic to the high temperate zones of Asia, Europe, and North America (Scandinavia, Iceland, Greenland, Alaska or Canada), making the world supply from wild populations on an industrial scale extremely limited (Blomkvist et al., 2009).
To date, 74 accepted species have been identified within the Rhodiola genus. Many of them are used in national systems of traditional medicine, including Rhodiola heterodonta (Hook.f. and Thomson) Boriss, Rhodiola quadrifida (Pall.) Fisch and Mey, Rhodiola semenovii (Regel and Herder) Boriss, Rhodiola kirilowii (Regel) Maxim., and Rhodiola crenulata (Hook.f. and Thomson) H.Ohba. Of all these species, R. rosea is the one most studied for its phytochemical properties as well as its use in medicine and therapy. R. rosea is one of the main representatives of the group of plant adaptogens used for medicinal purposes linked to stress and age-related impairments of cognitive functions as a tonic and immunomodulating therapeutic agent (European Food Safety Authority (EFSA), 2012). The most valuable parts of the plant used in therapy are R. rosea roots and rhizomes, of a golden metallic colour, harvested after at least a 5-year vegetative period (Galambosi, 2006).
R. rosea has a long history of use as a medicinal plant, appearing in reports from Asian and European countries. The therapeutic use of Rhodiola rosea roots traces back to Tibetan Traditional Medicine (TTM), which has influenced its more recent use in Traditional Chinese medicine (TCM), where it was employed to treat altitude sickness, fatigue, and mood disorders, as well as headaches, diarrhoea, skin conditions and swelling, typically in the form of infusions and tinctures (Tao et al., 2019).
The European Medicines Agency classifies the dry extract (DER 1.5–5:1) obtained from R. rosea roots and rhizomes as a traditional herbal medicinal product used for the prevention and temporary relief of fatigue and sensation of psycho-physical weakness (European Medicines Agency, 2012). Currently, in the European Union, R. rosea preparations are used as active ingredients in registered traditional herbal medicinal products (HMP) and as ingredients in dietary supplements, suggesting potential benefits on memory, learning, and attention. Supplements containing Rhodiola are recognised as beneficial for managing physical and mental stress in the European Food Safety Authority’s (EFSA) consolidated list of Article 13 health claims (European Food Safety Authority, 2012). Moreover, R. rosea root extracts can be found in “functional foods”, sports and energy drinks, alcoholic beverages, as well as the cosmetic industry (European Commission, 2020; Lamadrid et al., 2019).
The multidirectional properties of R. rosea - concerning its antibacterial, anxiolytic, antioxidant, immunomodulating, cardio and neuroprotective, as well as nootropic effects are strongly correlated with the rich phytochemical composition of its underground organs (Lee et al., 2013; Zhang et al., 2022). Chemical investigation on this plant revealed approximately 150 chemical entities, encompassing phenylethanoids (i.e., salidroside with its aglycone–tyrosol and its derivatives), phenylpropanoids (i.e., rosin, rosavin, rosarin), and flavonoids characteristic of this species (herbacetin, gossypetin and kaempferol derivatives), as well as cyanogenic glycosides, gallic acid derivatives and proanthocyanidin constituents among others (Alperth et al., 2019). Although it is still not entirely clear which compounds are responsible for the adaptogenic effects of the raw material, the observed effects have been considered to be mainly based on phenylethane and phenylpropane derivatives. The latest edition of the European Pharmacopoeia (11th edition, 2023) emphasises the quality assessment of R. rosea-based products by considering the content of marker compounds, salidroside and rosavins, in a ratio not lower than 1:3. In turn, the United States Pharmacopeia guidelines assess the quality mark for the raw material as containing not less than 0.3% of the phenylpropanoid glycosides rosarin, rosavin and rosin (these three compounds are also collectively referred to as ‘rosavins’) calculated as rosavin, and not less than 0.08% of salidroside, calculated on a dry weight basis (United States Pharmacopeial Convention, 2017; Ezra Bejar and John, 2017). Salidroside has been identified in several species of the Rhodiola genus, including R. crenulata, R. rosea, R. sachalinensis, and R. kirilowii, while rosavin has been isolated specifically from the roots of R. rosea and R. sachalinensis (in smaller amounts) (Booker et al., 2016; Nakamura et al., 2008). Despite the presence of salidroside in most species of the Rhodiola genus, its relatively low content in R. rosea does not allow for its unequivocal recognition as a marker compound for this species. Consequently, monitoring the presence of rosavin, as a distinctive chemotaxonomic marker, seems to be considered synonymous with the high quality of the tested preparations.
The rising demand for R. rosea has been paralleled by an increase in dietary supplements claiming its inclusion. Considering the economic advantage of producing dietary supplements over medicinal products, as well as the lack of precise guidelines for analysing their composition, proper quality control of R. rosea preparations poses a significant global challenge within the pharmaceutical industry. The plant’s limited native range, coupled with its difficult accessibility, prolonged growth period, and morphological similarities within the Rhodiola genus, has led to frequent adulteration with other species (Nikolaichuk et al., 2021). Furthermore, the adaptogenic properties attributed to R. rosea, together with the growing number of individuals seeking relief from excessive psycho-physical stress, fatigue, and exhaustion, have rendered R. rosea an endangered species in many regions. The global demand for adaptogenic medicinal plants such as R. rosea and its key compounds, salidroside and rosavins, has also driven an increase in preclinical and clinical studies aimed at uncovering new applications for R. rosea in modern phytotherapy, which has further depleted the natural resources of the raw material (Hung et al., 2011; Lu et al., 2022).
The aim of our study was to assess the phytochemical analysis of commercial products of the underground parts of R. rosea and/or their potential contaminants. Herein, HPTLC and LC-MS fingerprints describing the phytochemical composition of 13 preparations containing R. rosea unprocessed roots or their extracts were analysed, with a particular emphasis on assessing the rosavin presence within these products. The traditional use of R. rosea is well-documented for its adaptogenic and stimulating effects. As the search for natural substances that help the human body adapt to challenges from technological advancements and modern working conditions continues, we analysed the qualitative composition of the tested extracts, including rosavin, and correlated these findings with their anti-inflammatory effects in a BV2 microglial cell model. Incorporating the potential impact of the tested samples on inflammatory responses within a microglial model may provide a valuable tool for additional quality control of adaptogenic plant materials.
2 Materials and methods
2.1 Plant material used for the study
The plant material, including dietary supplements available in pharmacies and herbal stores, was purchased commercially. The preparations claiming to contain extracts from the roots and rhizomes of Rhodiola rosea were obtained as standarised/non–standarised extracts in the form of tablets (3) or capsules (5). The herbal products consisting of cut, unprocessed roots/rhizomes (4) or powdered raw material (1) were also analyzed. Reference material (Rhodiola rosea roots, Crassulaceae) was obtained from the Institute of Natural Fibres and Medicinal Plants crops controlled cultivation.
Detailed information on the examined preparations is provided in the Supplementary Material (Supplementary Table S1). A voucher specimen of the tested dietary supplements and plant material has been deposited in the Plant Collection of the Department of Pharmaceutical Biology at the Medical University of Warsaw (no. 80/2024). The materials used for the study are summarised in the table below (Table 1).
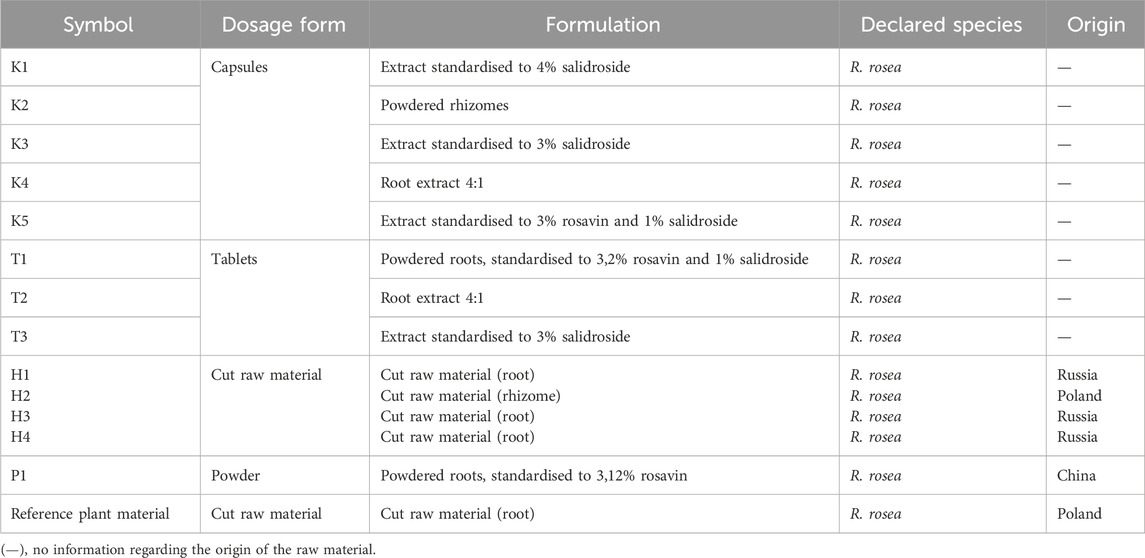
Table 1. Source of plant material used in the study.
2.2 Preparation of extracts from investigated samples for phytochemical analysis
The contents of tablets, capsules, sachets, as well as cut roots and rhizomes, were finely ground. Extracts were prepared using 60% ethanol (Avantor Performance Materials Poland S.A., Gliwice, Poland) as the solvent, in accordance with the guidelines of the European Medicines Agency (European Medicines Agency, 2012). Considering the diverse chemical groups present in R. rosea, infusions were also prepared from each of the examined dietary supplements as the most widely used and most easily available form of natural raw materials administration.
Infusions were prepared by adding boiling water to the plant material at a ratio of 1:10 and left under cover for 15 min according to the method recommended by the European Pharmacopoeia. Ethanolic extracts at a concentration of 60% were prepared by adding a solvent to the finely ground and crushed material (1:5 ratio) and heating it at 95°C for 1 hour. After filtration, ethanol was evaporated under reduced pressure. The aqueous residues (from infusions and ethanolic extracts) were lyophilised to obtain dry powders.
Lyophilisates were dissolved in methanol:water (8:2, v/v) to reach a final concentration of 10 mg/mL. The reference substance (rosavin, Sigma–Aldrich Chemie GmbH, Steinheim, Germany) was dissolved in pure methanol to obtain a stock solution at a concentration of 1 mg/mL.
2.3 HPTLC apparatus and chromatographic conditions
The High-performance thin-layer chromatography (HPTLC) analysis was performed using a TLC system provided by Camag (Muttenz, Switzerland). The setup included a Linomat 5 sample applicator fitted with a 100 µL Hamilton syringe, an Automatic Developing Chamber 2 (ADC2), a Scanner 4, an Automatic Derivatizer, a Visualizer 2, and a Plate Heater III. Device operation and image analysis were managed using the WinCATS 1.4.2 software (Camag).
Silica gel 60 F254-coated HPTLC plates, glass-backed, measuring 20 × 10 cm (Merck, Darmstadt, Germany), served as the stationary phase. Before use, the plates were prewashed with methanol, dried for 10 min at ambient temperature, and subsequently activated at 80°C for 30 min. Extract samples (10 mg/mL) and standard solution samples (1 mg/mL) were applied in 10 µL volumes as 6 mm bands, with 12 mm spacing between tracks and distances of 12 mm and 10 mm from the side and lower plate edges, respectively.
A consistent application rate of 150 nL/s was maintained. The plates were developed using a mobile phase comprising ethyl acetate, methanol, water, and formic acid in the ratio of 77:13:10:2 (v/v/v/v). Plate development took place in an automatic developing chamber, which was presaturated with 10 mL of the mobile phase at a controlled room temperature of 23°C ± 2°C and a relative humidity of 40% ± 5%. The plate development process involved the following parameters: humidity control for 5 min, tank saturation for 5 min, plate preconditioning for 5 min, a mobile phase migration distance of 80 mm, and a plate drying time of 10 min. Spot visualisation was performed under white light as well as UV light at 254 nm and 366 nm. For derivatisation, plates were treated with a 1% solution of vanillin in sulfuric acid solution (Loba Feinchemie, Apolda, Germany). The derivatisation process was carried out using a CAMAG Automatic Derivatizer (set to level 3). Post–spraying, the plates were imaged again under white light and UV light at 254 nm and 366 nm.
2.4 UHPLC–DAD–ESI–MS/MS analysis
The Ultra-High Performance Liquid Chromatography – Diode Array Detection – Electrospray Ionization – Tandem Mass Spectrometry (UHPLC–DAD–ESI–MS/MS) analysis was performed on a UHPLC–3000 RS system (Dionex, Germering, Germany) with DAD detection and an AmaZon SL ion trap mass spectrometer with an ESI interface (Bruker Daltonik GmbH, Bremen, Germany). Separation was performed on a Zorbax SB–C18 column (150 × 2.1 mm, 1.9 μm) (Agilent, Santa Clara, California, USA). The mobile phase consisted of water +0.1% formic acid (A) and acetonitrile +0.1% formic acid (B). A gradient was applied as follows: 0–60 min, 5%–40% B. All solvents used for analysis (methanol, formic acid acetonitrile; Avantor Performance Materials Poland S.A., Gliwice, Poland) were of gradient grade. The LC eluate was introduced into the ESI interface without splitting, and the analysis was performed in negative ion modes with the following parameters: nebuliser pressure at 40 psi; drying gas flow rate at 9 L/min; nitrogen gas temperature at 300°C; and a capillary voltage of 4.5 kV. The mass scan range was set from 100 to 2,200 m/z. UV–VIS detection covered the 190–600 nm range. All samples were analysed using the UHPLC–DAD–ESI–MS/MS method, and the predominant substances in each extract were identified by comparing retention times and spectra (UV, MS, MS/MS) with standard and literature data.
2.5 Preparation of tested extracts for bioassay
Lyophilisates were dissolved in DMSO (Sigma–Aldrich Chemie GmbH, Steinheim, Germany) and then diluted with (Mg2+, Ca2+)–free PBS buffer at pH 7.4, reaching a final 1 mg/mL concentration. Control with dexamethasone (Merck, Darmstadt, Germany) as well as the reference marker–rosavin, were initially dissolved in DMSO to obtain 10 mM stock solutions, which were then diluted with (Mg2+, Ca2+)–free PBS buffer. The infusions and ethanolic extracts were tested at a concentration of 50 μg/mL. Rosavin was evaluated across a concentration range of 0.1–5 μM, while the control samples containing dexamethasone were assessed at a concentration of 20 µM. The concentration of DMSO (<0.1% in the final added quantity) did not affect the outcomes of the assays. Appropriate dilutions and final concentrations of LPS were prepared using DPBS solution.
2.6 Determination of BV2 cell viability
The immortalised murine microglial cell line BV2 (passages 1–4) was purchased from DSMZ–German Collection of Microorganisms and Cell Cultures GmbH. BV–2 cells (passages 1–5) were cultured in RPMI medium 1,640 (Thermo Fisher Scientific, Waltham, MA, USA) supplemented with 10% FBS and antibiotics (1% penicillin–streptomycin and 0.5% amphotericin B; PAA, Laboratories GmbH, Pasching, Austria) at 37°C, 5% CO2. For all experiments, BV–2 cells were incubated in RPMI 1640 medium containing 2% FBS, supplemented with antibiotics.
Cells were seeded at 4 × 103 cells/ml in 96–well microtiter plates (Nunc™, Thermo Fisher Scientific) and cultivated for 24 h at standard treatment conditions. BV2 microglia were exposed to the tested extracts (or appropriate controls), with or without stimulation with LPS (100 ng/mL) from Escherichia coli 0111:B4 (Sigma–Aldrich Chemie GmbH, Steinheim, Germany) for 24 h. Following treatment, the cell culture medium was replaced with a culture medium containing NRU dye supplement (50 μg/mL) or MTT solution (0.5 mg/mL). Following 6 h incubation, intracellular NRU dye was extracted using a destaining solution (ethanol, water, and acetic acid, mixed in a 50:49:1 ratio). For the MTT assay, following a 4–hour incubation, the medium was removed, the cells were washed with DPBS, and the formazan crystals were dissolved in 200 μL of DMSO. Absorbance was subsequently measured at 540 nm for NRU and 580 nm for MTT assay, using a Synergy 4 BioTek microplate reader (Winooski, VT, USA).
2.7 TNF-α, IL-6 secretion by BV2 cells
BV2 cells were plated in 24–well plates at the density of 1 × 105 cells/mL and cultivated for 24 h, at standard treatment conditions. Subsequently, the cells were treated with tested extracts (50 μg/mL) or rosavin (20 µM), followed by the stimulation with LPS (100 ng/mL). After 24 h, collected supernatants were centrifuged, and the cytokines’ release was determined by ELISA assay kits (BD Biosciences, San Jose, CA, USA) following the manufacturer’s instructions using a microplate reader. The effect on cytokine production was determined through the percentage of released cytokines relative to the LPS–stimulated control. Dexamethasone (20 µM) was used as a positive control.
2.8 Statistics and data analysis
The results were expressed as mean ± SEM for three independent experiments performed at least in triplicate. One–way analysis of variance (ANOVA), HSD Tukey test, and Dunnett’s test were applied to evaluate the statistical significance of the mean values, with *p < 0.05, **p < 0,01, ***p < 0.001, regarded as statistically significant. GraphPad Prism 10 was used for all analyses and graphs.
3 Results
3.1 HPTLC analysis of ethanolic extracts prepared from Rhodiola rosea commercial samples
Qualitative evaluation of the composition of the tested samples, declaring the content of R. rosea roots/rhizomes or their extracts, was conducted using the HPTLC method. The analysis enabled the preliminary verification of the composition of 13 dietary supplements. The major bioactive compounds of R. rosea, e.g., rosavin, are often used for the quality evaluation of R. rosea preparations (Booker et al., 2016). Notably, the analysis confirmed the presence of rosavin in 5 of the 13 tested preparations. After the development of chromatograms, the band corresponding to rosavin was observed under UV light at a wavelength of 254 nm, with an Rf value of 0.22 (Figure 1A). Further verification through chemical derivatization using vanillin in a sulfuric acid solution confirmed the presence of rosavin as a violet band (details provided in the Supplementary Figures S1, S2), which was visible in samples Z1A, Z3A, K2A, K3A, and K5A (Figure 1B). Notably, HPTLC profiling revealed that the characteristic marker compound, rosavin, was absent in 8 products claiming to contain Rhodiola rosea.
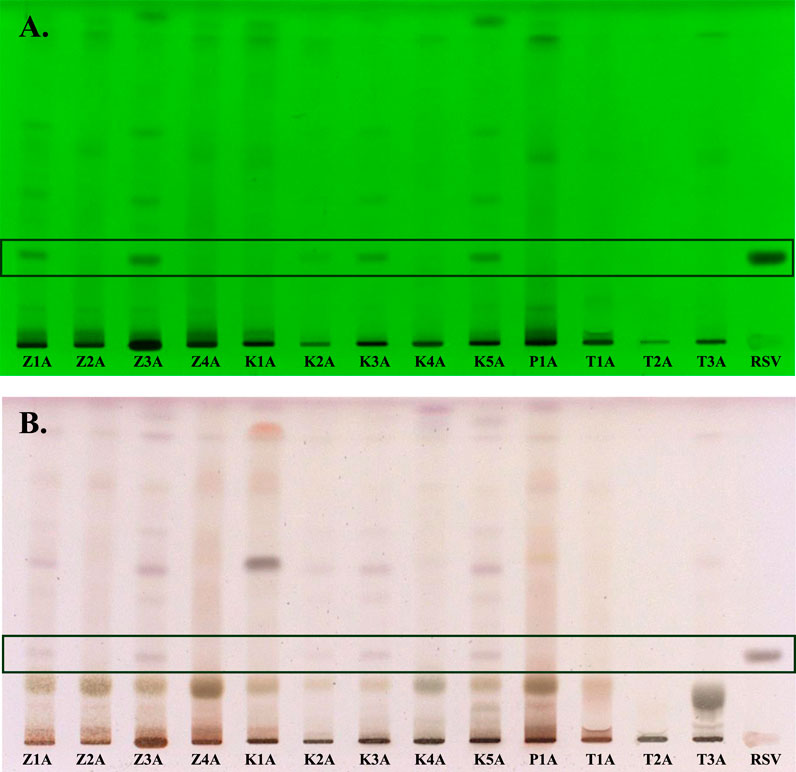
Figure 1. HPTLC chromatograms of the Rhodiola rosea dietary supplements, separated on HPTLC plates silica gel F254, using ethyl acetate–methanol–water–formic acid (77:13:10:2, v/v/v/v) mobile phase and detected at (A) 254 nm (before derivatization) and in (B) white light illumination (after derivatization). Z1A–Z4A–unprocessed raw materials of Rhodiola rosea roots/rhizomes; K1A–K4A–capsules; P1A–powder; T1A–T3A–tablets; RSV–rosavin.
The HPTLC analysis also revealed that the K1A, P1A, and T1A samples exhibited a fingerprint inconsistent with that of R. rosea (e.g., Z3A) and were likely adulterated with other Rhodiola species. Additionally, two of the tested samples (T1A and T2A) showed the presence of unspecified components unrelated to R. rosea.
3.2 LC–MS analysis of Rhodiola rosea roots aqueous and ethanolic extract
From the reference material–cut, unprocessed roots of Rhodiola rosea, an infusion and a 60% ethanolic extract were prepared. Phytochemical analysis of the prepared solutions was conducted using the HPLC–DAD–MS/MS method. The chromatogram and the analysis of the reference extracts (60% ethanolic–green and infusion–blue), recorded at a wavelength of 240 and 280 nm, are presented below (Figures 2A,B; Table 2).
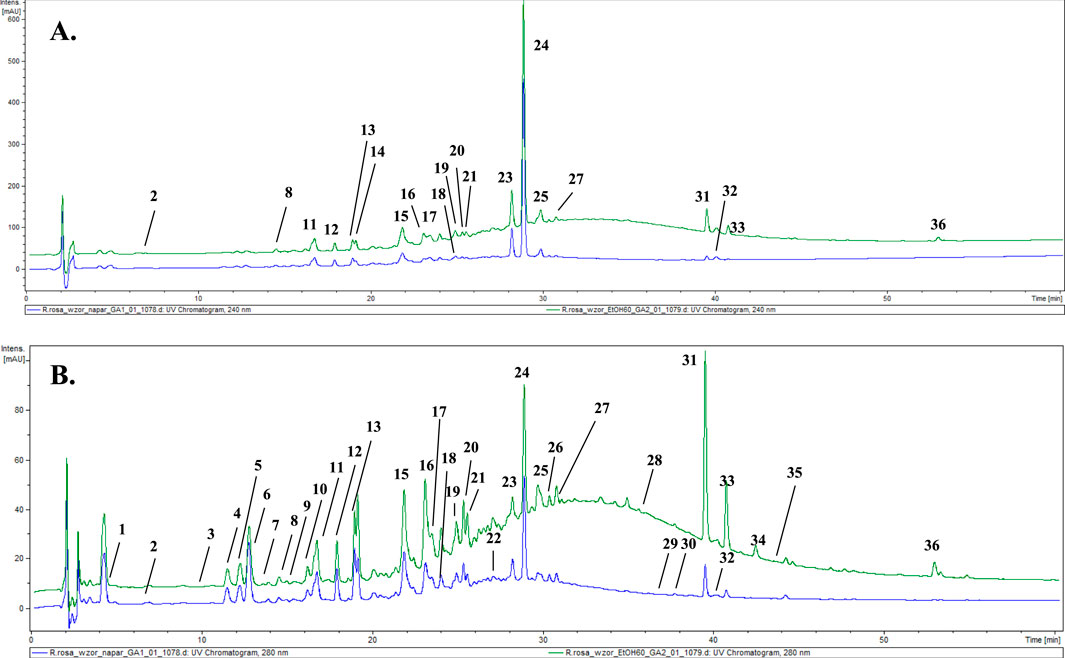
Figure 2. LC-DAD-MS/MS chromatograms of extracts (60% ethanolic extract–green and aqueous extract–blue) from reference plant material, recorded at (A) 240 nm and (B) 280 nm. The numbered peaks are denoted in Table 2.
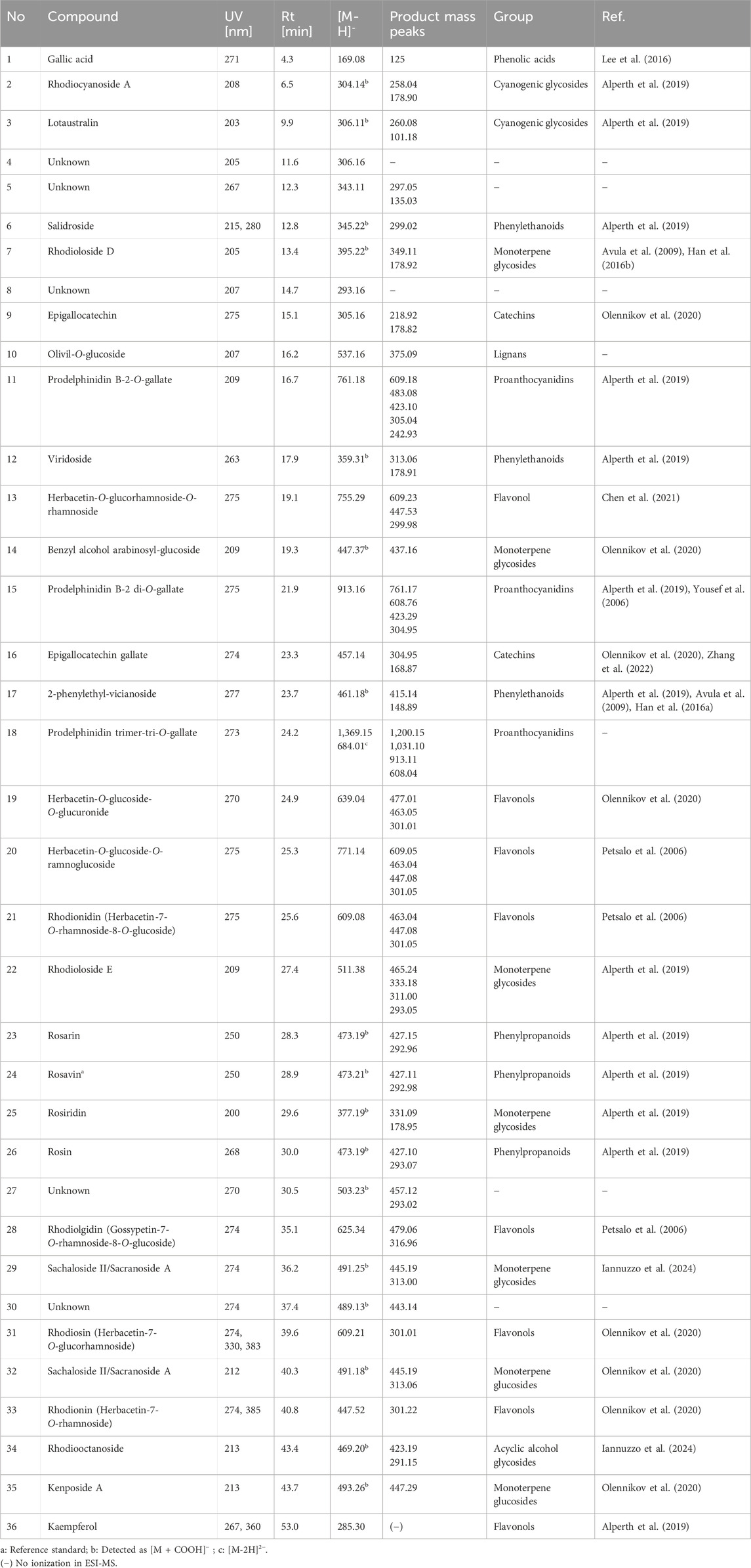
Table 2. Retention time, UV, and MS/MS data of the compounds present in the tested extract.
In the course of our analysis, we identified 36 metabolites, encompassing a diverse range of chemical groups, including phenolic acids, monoterpene alcohols glycosides, cyanogenic glycosides (hydroxynitrile glycosides), flavonoids, phenylethanoids, phenylpropanoids, proanthocyanidins, and others. Constituents were tentatively identified by comparison with literature data describing compounds previously found in this genus. Analysis was performed mainly considering negative ionisation data. Negative molecular ions are indicated by m/z values equal to [M−H]− or [M−2H]2−. In several peaks, formic acid adducts were detected as ([M−H]− + COOH), which fully dissociated into [M−H]− in MS2.
The representative of the phenolic acid group was gallic acid (1, tr = 4.3 min) with m/z 169.08 [M−H]−.
Compounds 2 and 3 were identified as hydroxynitrile glucosides structurally related to cyanogenic glycosides. Compound 2 (tr = 6.5 min) showing a pseudomolecular ion at m/z 304.14 [M + COOH]−, with fragments at m/z 258.04 and 178.90 was identified as rhodiocyanoside A, while compound 3 was found to be lotaustralin (tr = 9.9 min; m/z 260.08 [M−H]−).
Two phenyloethanoids were identified. Salidroside with m/z 299.02 [M−H]− (6) is the most frequently reported phenyloethanoid, representing the Rhodiola genus, found in the aerial parts as well as roots as rhizomes (Zakharenko et al., 2021). Moreover, viridoside (12), identified at tr = 17.9, with its pseudomolecular ion at m/z 359.31 [M + COOH]− and fragmentation pattern at m/z 313.06 and 178.91 was also identified (Lee et al., 2016).
From the group of oligomeric procyanidins, compounds 11 (tr = 16.7 min) and 15 (tr = 21.9 min) were identified as prodelphinidin dimers, as its O–gallate (761.18 → 609.16 → 423.14 → 305.07) and di–O–gallate (913.19 → 761.18 → 608.76 → 423.14 → 305.07), respectively. Similarly, compound 18 (tr = 24.2) was also classified into the proanthocyanidin group and identified as prodelphinidin trimer–tri–O–gallate (1,369.15 → 913.19 → 608.76).
Six glycosides of monoterpene alcohols were also identified (or partially identified) during the analysis. Compounds 7 and 22 were structurally elucidated and designated as rhodioloside D and rhodioloside E, respectively. High–resolution mass spectrometric analysis of compound 7 revealed a pseudomolecular ion at m/z 349.11 [M−H]-. Compound 22 was eluted at 22.4 min and exhibited a pseudomolecular ion at m/z 511.38 [M−H]-.
Rosiridin (rosiridol 1–O–glucoside), eluted at tr = 29.6 min with m/z 331.09 [M−H]− and fragmentation ion at m/z 178.95, corresponding to the loss of glucose moiety was identified as compound 25. Sacranoside A and sachalinoside II can be seen as a possible candidates for 29 (tr = 36.2) or/and 32 (tr = 40.3) with their pseudomolecular ion at m/z 445.19 [M−H]−. Based on pseudomolecular ion at m/z 447.29 [M−H]− compound 35 was identified as kenposide A.
A distinctive characteristic of R. rosea is the presence of cinnamic alcohol glucosides and a notably high concentration of phenylpropanoids, called rosavins, specified for Rhodiola rosea. Compounds 23 (tr = 28.3 min), 24 (tr = 28.9 min), and 26 (tr = 30.0 min) were identified as rosarin, rosavin and rosin respectively, while rosavin was identified in comparison with reference substance. The sugar moieties consist of arabinopyranosyl–glucopyranoside for (23), arabinofuranosyl–glucopyranoside for (24), and glucopyranoside for (26) respectively.
Another important group of compounds identified in R. rosea roots/rhizomes are flavonols and their derivatives. Compound 31 (tr = 39.6) with pseudomolecular ion m/z 609.21 [M−H]− and fragmentation pattern m/z 445.17 [M−H−164]− and 300.94 [M−H−146]− corresponding to loss of glucose and rhamnose moiety was identified as rhodiosin (herbacetin–7–O–glucorhamnoside). Compound 37 (tr = 40.8) was identified as rhodionin (herbacetin–7–O–rhamnoside), based on its fragmentation pattern with m/z 447.52 → 301.22 [M−H−146] −. Compound 13 (tr = 19.1), with pseudomolecular ion at m/z 755.29 [M−H]−, was identified as rhodiosine rhamnoside (herbacetin–3–O–glucorhamnoside–7–O–rhamnoside) based on its fragmentation pattern indicating the loss of 2 rhamnose and 1 glucose molecules. In turn, compound 28 (tr = 35.1) was identified as rhodiolgidin (gossypetin–7–O–rhamnoside–8–O–glucoside), based on fragment ions including 316.96 [M−H−146–162]−, indicating the presence of aglycone–gossypetin. Compound 36 showed [M−H]− ion at m/z 285.30 (tr = 53.0 min) and could be identified as kaempferol.
3.3 LC–MS analysis of Rhodiola rosea roots dietary supplements
Phytochemical analysis and identification of the primary compounds were conducted on ethanolic extracts and infusions prepared from 13 dietary supplements declared to contain R. roseae roots.
Based on the comparison of the phytochemical profiles of various species within the Rhodiola genus, salidroside’s presence can be considered a generic marker compound. In addition to R. rosea, its presence has been recorded in several other species, including R. quadrifida, R. algila (Ledeb.) Fisch, R. sachalinensis, R. kirilowii, R. crenulata, R. heterodonta, and R. semenovii (Zakharenko et al., 2021). As demonstrated in our analysis, salidroside was confirmed in 11 out of the 13 tested formulations containing both R. rosea and other related species. Based on the peak sizes, the highest content of this compound was identified in K1, T3, and P1 samples. Notably, salidroside was not detected in T1/T1A and T2/T2A, while T2 did not contain any compounds characteristic of the Rhodiola genus, as confirmed by HPTLC and LC–MS methods.
On the other hand, the occurrence of cinnamic alcohol and its glycosides–rosavin, rosarin, and rosin, is described as a characteristic marker specific to the R. rosea species (Bykov et al., 1999). The confirmation of these marker compounds’ presence is a crucial factor in the quality assessment of formulations containing R. rosea. Based on the results obtained from HPTLC fingerprinting, the presence of rosavin was confirmed in only five samples (Z1, Z3, K1, K3, K5). However, using the more sensitive liquid chromatography–mass spectrometry (LC–MS) method, rosavin was additionally detected in trace amounts in three further samples (Z2, K2, K4). This may indicate the poor quality of certain preparations, including the predominant presence of contaminants or a significant presence of other Rhodiola species in their composition.
In turn, in 5 of the tested samples, all three phenylpropanoids (rosarin, rosavin, rosin) were identified. In samples K4 and Z2, Z4 a small (trace) amount of rosavin was possibly detected using LC–MS, along with compounds characteristic of other Rhodiola species, such as R. crenulata, R. gelida, R. sacra, and others, e.g., crenulatin and creoside I.
The characteristics of the individual forms of administration of the tested dietary supplements declaring the content of R. rosea or their extracts are presented below. The detailed characteristics of the presence of compounds identified in the individual samples are summarised in Table 3, available below. The numbering of the compounds in the graphical representations corresponds to that in Table 3.
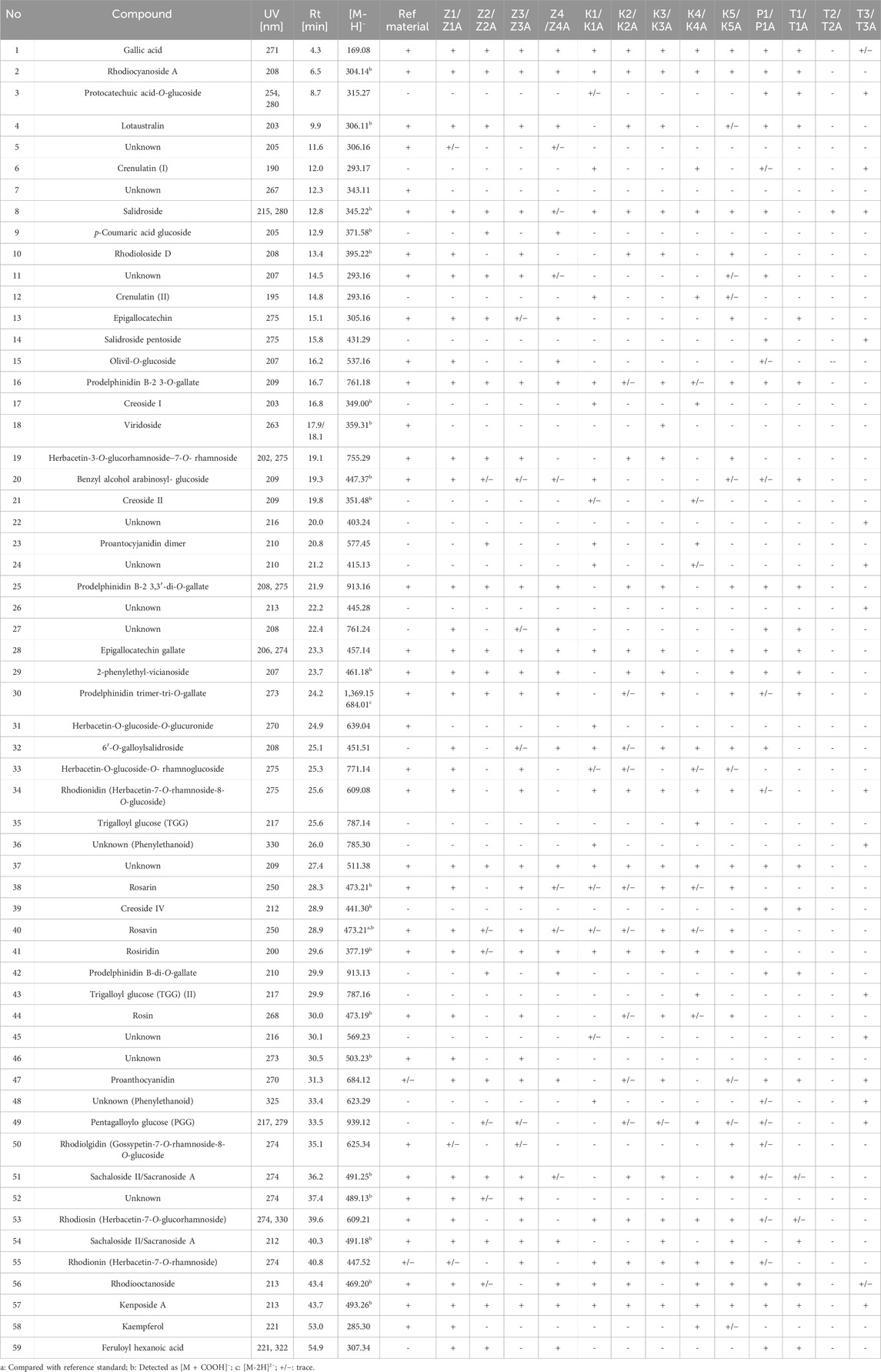
Table 3. The content of compounds in individual dietary supplements identified on the basis of HPLC-MS - retention times, UV, mass spectra fragmentation.
3.3.1 Cut raw material
The phytochemical analysis of the preparations in the form of unprocessed raw material confirmed the presence of rosavin (40, tr = 28.9) as a marker compound for R. rosea in four tested preparations (Z1–Z4) (Figure 3). However, in samples Z2 and Z4, the presence of rosavin was minimal. However, the presence of compounds of ambiguous species affiliation was observed. In both Z2 and Z4 p–coumaric acid glucoside (9) with m/z 371.25 [M + COOH]− (MS2 325.05 and 162.84 [M−H]−) and feruloyl hexanoic acid (59) with m/z 307.34 [M−H]− (MS2 193.00 [M−H]−) characteristic for R. wallichiana, were detected (Liu et al., 2019). Phytochemical profile of Z4 was matching with our reference extract of R. kirilowii (data nor shown), especially rich in proanthocyanidins such as: prodelphinidin B–2 3,3′–di–O–gallate (25), epigallocatechin gallate (28), prodelphinidin trimer–tri–O–gallate (30) and prodelphinidin B–di–O–gallate (42). This may suggest the presence of a mixture of species in the tested preparations (R. rosea, R. wallichiana and R. kirilowii), although we did not identify the presence of compounds typical of R. crenulata, e.g., crenulatin or creosides I–IV. Based on the HPTLC chromatogram and comparison of the LC–MS Z1 and Z3 fingerprints with the reference material profile, we can determine the presence of the R. rosea species as the leading (or only) component.
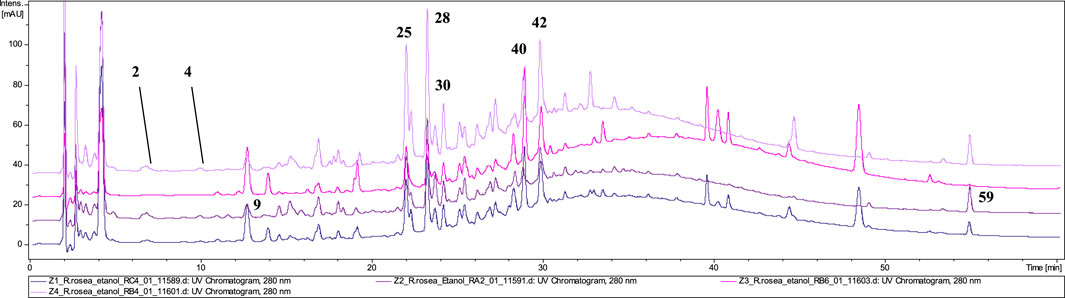
Figure 3. LC-DAD-MS/MS chromatograms of 60% ethanolic extracts prepared from tested cut raw materials, recorded at 280 nm.
3.3.2 Capsules
The analysis of the capsules’ composition confirmed the presence of rosavin (40, tr = 29.0) in all five tested extracts (K1–K5) (Figure 4). However, the content of this compound in formulations K1 and K4 was only in trace amount. Interestingly, despite the presence of this compound characteristic of R. rosea, the identity of this species could only be conclusively confirmed in 2 of the 5 tested preparations–K3, and K5. In capsules K1 and K4, only minimal amounts of rosavins were detected, accompanied by the presence of compounds characteristic of other Rhodiola species. Notably, creoside I (17, tr = 16.8) with m/z at 349.00 [M + COOH]- and crenulatin (6, tr = 12.0 and 12, tr = 14.8), which are also characteristic of R. crenulata, were identified (Han et al., 2016a; Han et al., 2016b).
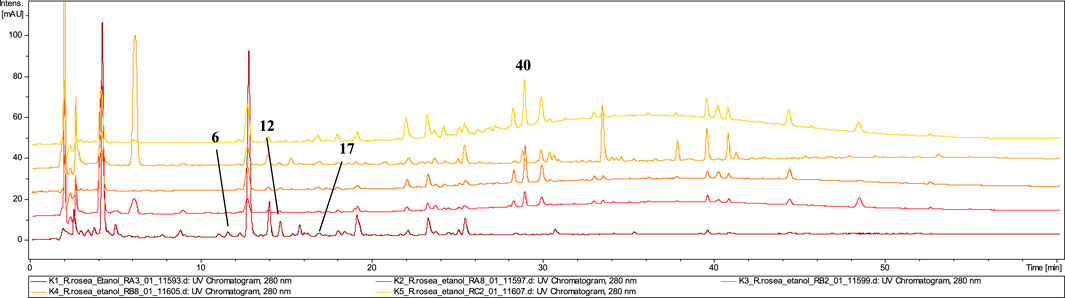
Figure 4. LC-DAD-MS/MS chromatograms of 60% ethanolic extracts prepared from tested capsules, recorded at 280 nm.
3.3.3 Powder
The dominating compounds of P1 powdered raw material were gallic acid (1, tr = 4.3), identified by its pseudomolecular ion at m/z 169.08 [M−H]−, salidroside (8), salidroside pentoside (14), prodelphinidin B–2 3–O–gallate (16), epigallocatechin gallate (28), and 2–phenylethyl–vicianoside (29). No rosavin was detected in the P1 extracts, confirming the absence of R. rosea (Figure 5). Additionally, the presence of traces of crenulatin (6) and creoside IV (39, tr = 28.9) in the tested extracts suggests potential adulteration of the preparation with R. crenulata with other undefined Rhodiola spp. excluding R. rosea (Han et al., 2016a; Han et al., 2016b).
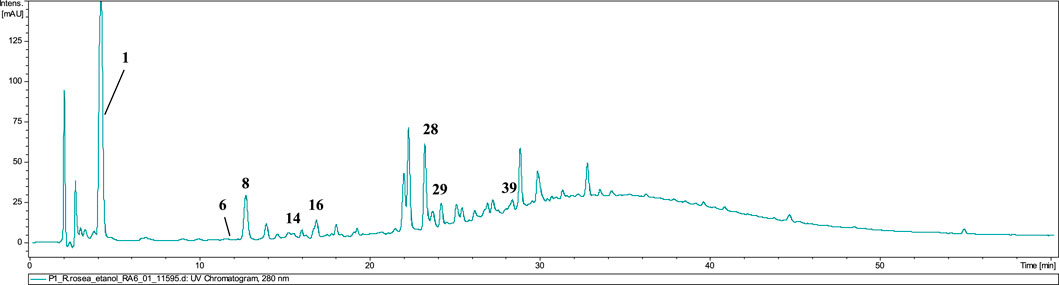
Figure 5. LC-DAD-MS/MS chromatograms of 60% ethanolic extract prepared from tested powder, recorded at 280 nm.
3.3.4 Tablets
The dietary supplements in tablet form (T1, T2, T3) exhibited the lowest quality among the tested preparations (Figure 6). LC–MS analysis revealed that none of the three tablet–formed preparations contained extracts from R. rosea roots/rhizomes. Neither rosavin nor other characteristic cinnamic alcohol derivatives were identified in T2 and T3 extracts.
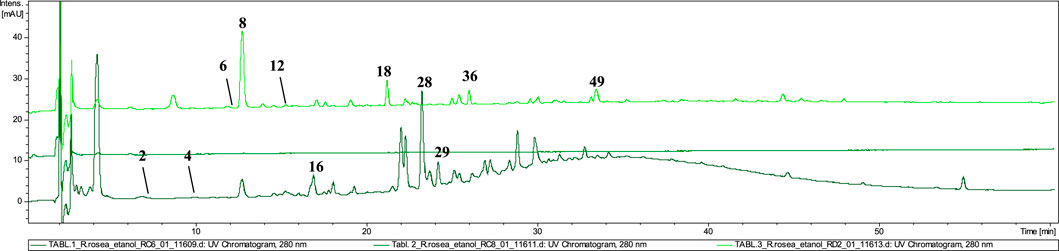
Figure 6. LC-DAD-MS/MS chromatograms of 60% ethanolic extracts prepared from tested tablets, recorded at 280 nm.
However, in extract T1, we identified compounds characteristic of other species within the Rhodiola genus, such as prodelphinidin B–2 3–O–gallate (16), epigallocatechin gallate (28), and 2–phenylethyl–vicianoside (29), which are also commonly found in R. gelida, R. recticaulis, R. sachalinensis, R. kirilowii and R. sacra (Tao et al., 2019). In tablet T3, compounds specific to R. crenulata, such as crenulatin (6, tr = 12.0 and 12, tr = 14.8) with m/z at 293.17 [M−H]− was detected together with compound not present in Rhodiola spp. (compounds 36 and 49) (Nakamura et al., 2008). Moreover, similarly to the P1 preparation, in the T3 tablet we observed the dominant presence of salidroside (8) and small amounts of its derivative–salidroside pentoside (14).
3.4 Anti–inflammatory activity of Rhodiola rosea root extracts
Microglial cells represent the primary immune defense within the central nervous system (CNS), playing a crucial role in responding to pathological stimuli and maintaining neural homeostasis. To assess the biological activity of the tested samples (towards TNF-α and IL-6 release), we employed the LPS-stimulated BV2 microglial cell line, a widely used in vitro model for investigating the effects of plant-derived compounds on neuroinflammatory processes (Marrazzo et al., 2023).
The most substantial inhibition of TNF-α secretion was observed following incubation with infusions derived from the Z3 raw material, which contained the highest concentration of rosavin among the unprocessed samples, as indicated in the Supplementary Material (Supplementary Figure S3). Furthermore, infusions prepared from formulations containing only trace amounts or entirely lacking the marker compound, including Z4, capsule K4, tablet T3, powder P1, and the T1 ethanolic extract, also demonstrated inhibitory effects. For example, ethanolic extracts from products Z3 and Z4 reduced TNF-α secretion to 62.0% ± 9.5% and 66.6% ± 13.7%, respectively. In contrast, incubation of cells with K4, T3 ethanolic extracts, and the T1 infusion induced TNF-α secretion at levels of 76.0% ± 6.5%, 74.3% ± 5.1%, and 77.8% ± 4.3%, respectively, relative to the LPS–stimulated control (100% secretion) (Figure 7).
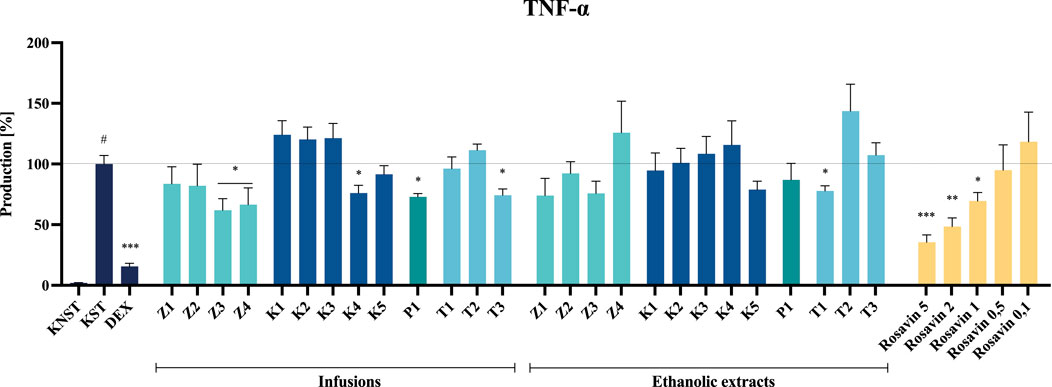
Figure 7. The influence of tested extracts (50 μg/mL) and rosavin (0.1–5 μM) on TNF-α secretion by LPS-stimulated microglia BV2. Data from three separate experiments assayed in duplicate are expressed as mean ± SEM. Dexamethasone (DEX, 20 μM) was used as a positive control. Absorbance values for all samples were expressed as percentages relative to the LPS-stimulated control (KST), which was set at 100%. Statistical significance: *p < 0.05, **p < 0.01, ***p < 0.001 vs. stimulated control (KST), # statistically significant (p < 0.001) vs. non-stimulated control (KNST).
Despite the noticeable inhibitory effect of certain tested preparations (e.g., ethanolic extracts from raw materials or the infusion from P1) on IL-6 secretion, none of the analyzed samples demonstrated a statistically significant effect at the tested concentration.
Rosavin, as the primary marker compound, exhibited a statistically significant, concentration–dependent inhibitory effect on the secretion of the tested inflammatory mediators. Notably, rosavin suppressed TNF-α secretion at concentrations of 5, 2 and 1 μM, reducing its levels to 35.5% ± 6.0%, 48.5% ± 6.9% and 69.5% ± 6.9% respectively, relative to the LPS-stimulated control (100% secretion) (Figure 7). The effect of rosavin on IL-6 secretion was equally pronounced. The analyzed compound significantly reduced IL-6 levels to 34.3% ± 3.3% at a concentration of 5 μM, 43.5% ± 4.9% at 2 μM, and 51.6% ± 6.2% at 1 µM (Figure 8).
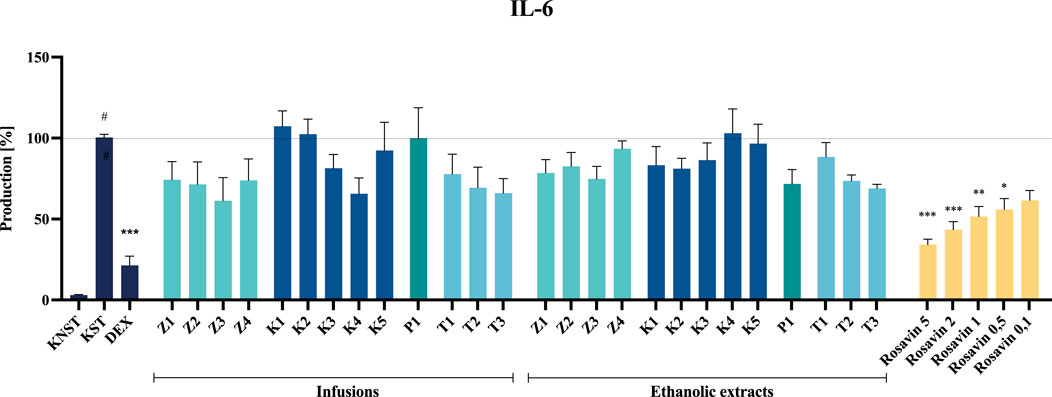
Figure 8. The influence of tested extracts (50 μg/mL) and rosavin (0.1–5 μM) on IL-6 secretion by LPS-stimulated microglia BV2. Data from three separate experiments assayed in duplicate are expressed as mean ± SEM. Dexamethasone (DEX, 20 μM) was used as a positive control. Absorbance values for all samples were expressed as percentages relative to the LPS-stimulated control (KST), which was set at 100%. Statistical significance: *p < 0.05, **p < 0.01, ***p < 0.001 vs. stimulated control (KST), # statistically significant (p < 0.001) vs. non-stimulated control (KNST).
Extracts prepared from dietary supplements of questionable quality or those containing significant admixtures of other species exhibited minimal or no effect on LPS-stimulated mediator production in the BV2 cell model at a concentration of 50 μg/mL (Figures 7, 8). None of the tested samples exhibited cytotoxic effects on BV2 microglial cells (see Supplementary Material, Supplementary Figures S4, S5).
4 Discussion
Neuroinflammation plays a crucial role in the pathogenesis of neuropsychiatric and neurodegenerative disorders. A growing body of evidence suggests that excessive microglial activation and the overproduction of pro–inflammatory mediators underlie the development of psychophysical exhaustion, depression, anxiety, and neurodegeneration. In addition to its role in synaptic plasticity, learning, and memory, microglia are essential for maintaining central nervous system (CNS) homeostasis. Due to their involvement in these processes, microglia may become an important target for therapeutic interventions aimed at modulating neuroinflammatory responses and mitigating disease progression (Gao et al., 2023; Li et al., 2022).
Rhodiola rosea L., a traditional medicinal plant widely used in Eastern Europe and Asia, is recognized for its neurostimulatory, antidepressant, and adaptogenic properties. Studies have demonstrated its efficacy in enhancing cognitive performance, reducing fatigue, and alleviating stress–related symptoms. In shift workers, short–term administration of R. rosea extract improved concentration, focus, and reaction speed under stress. Similarly, in a study on 80 healthy students, a 14–days regimen of 400 mg R rosea root extract significantly reduced anxiety, stress, anger, and confusion while improving mood (Cropley et al., 2015). Additionally, a phase II clinical trial comparing R. rosea with sertraline indicated a moderate antidepressant effect in mild to moderate depression, with fewer adverse effects than the conventional drug (Mao et al., 2015). The available literature provides partial insights into the pathomechanism underlying this effect. Available review reports systematize the available evidence indicating the inhibitory effects of the raw material on the secretion of inflammatory mediators, reactive oxygen species (ROS), and nitric oxide (NO) as well as its anti–apoptotic functions in in vitro models employing microglial and macrophage cells. Notably, the majority of studies focus on assessing the bioactivity of isolated single compounds, predominantly salidroside, a constituent found in multiple species within this genus and rosavins as leading anti–inflammatory components (Liu et al., 2024). Furthermore, there is a limited number of studies that establish a correlation between the biological activity of the extract and its chemical composition (Ma et al., 2018; Pu et al., 2020).
The growing global demand for Rhodiola rosea products, spurred by increasing scientific evidence supporting its efficacy, presents significant challenges to the conservation and sustainable use of this species. The expanding use of R. rosea in herbal medicines, dietary supplements, cosmetics, and food additives, combined with its limited cultivation areas, high agricultural costs, lengthy growing period, low dispersal ability, and low rate of germination and seedling survival is contributing to a gradual decline in the availability of this raw material worldwide (Galambosi, 2006). Depletion of natural populations and habitat destruction have led to the disappearance of Rhodiola species in many regions, as most of the raw material is collected from the wild, and the plant needs several years to regenerate (Howes et al., 2020).
The increasing demand for raw materials, coupled with their decreasing availability, has led to a significant rise in cases of intentional and accidental adulteration of R. rosea products. A major concern in this context is the admixture or substitution of R. rosea roots and rhizomes with those from other Rhodiola species, which are morphologically similar and difficult to distinguish, particularly in underground structures. Recent phylogenetic studies have highlighted significant evolutionary convergence among the 74 species of Rhodiola (Zhang et al., 2014). Although the color of the flowers can differentiate R. rosea from R. crenulata and other purple–flowering species, this convergence is particularly noticeable in the roots and rhizomes of Rhodiola, which lack distinct diagnostic features, making species identification based on these structures difficult (Cuerrier A and Rapinski, 2014; Zhu et al., 2017).
According to a 2016 market study, approximately 75% of the global Rhodiola extract production originates from Asia, with around 13% sourced from Europe, 5% from the United States, and 7% from other regions (Center, 2016). The high volume of imports from Asia, particularly from China, into the U.S. and European herbal supplement markets has led to frequent adulteration or substitution of R. rosea raw materials with other Asian Rhodiola species. This is compounded by the interchangeable use of various Rhodiola species names in different regions, with species like R. crenulata, R. heterodonta, and R. kirilowii being labeled under the common term “rhodiola” in China (Cunningham et al., 2020). Despite limited data, it is widely recognized that European sources of R. rosea are scarce. The commercial collection of R. rosea in Europe is of relatively low economic significance due to high labor costs and logistical challenges associated with harvesting in remote mountainous regions. Furthermore, many European populations of R. rosea are legally protected, including those in Bulgaria, the Czech Republic, Austria, and Germany (Fisheries, 2022).
The United States Herbal Medicines Compendium identifies several Rhodiola species as potentially confounding in the commercial supply of Rhodiola rosea, including R. crenulata, R. kirilowii, R. sacra, R. serrata, R. sachalinensis, and R. yunnanensis (United States Pharmacopeial Convention, 2014). Our analysis indicates the intentional or accidental substitution of R. rosea with R. crenulata in at least 5 of the tested products, as suggested by the presence of compounds characteristic of R. crenulata, such as creoside I, IV, and crenulatin. A typical example of a species involved in the adulteration of roseroot preparations is also R. sachalinensis, a species with a similar chemical composition to R. rosea but containing lower amounts of rosavins and salidroside. Some botanists even regard R. sachalinensis as the same species or a subspecies of R. rosea (Gontcharova et al., 2009). However, chemotaxonomic studies conducted by Booker et al. show differences between R. rosea and R. sachalinensis, which, based on general metabolomic methods and specific high–performance thin–layer chromatography (HPTLC) spectra and limited geographical range, allow treating them as separate species (Booker et al., 2016; Liu et al., 2013).
Although adulteration of preparations with other Rhodiola species seems to be a major challenge for the pharmaceutical market, meeting the requirements for good quality raw materials for the growing industrial demand is also a significant problem. The key aspect seems to be the requirements of the European Pharmacopoeia for R. rosea roots and rhizomes in terms of the content of marker compounds, i.e., salidroside and rosavins.
R. rosea is available on the market in various forms, including cut/dried, powdered rhizomes and roots, dry or liquid extracts, as well as dry extracts in solid dosage forms like capsules and tablets (Brinckmann et al., 2021). Market analysis indicates that the global demand for R. rosea products is constantly growing, and with it, the number of adulterated or poor–quality preparations (Bernard, 2016). The scale of the problem is illustrated by earlier studies assessing the quality of preparations declaring the content of R. rosea, available on the European market. Ma et al. found that approximately 33% of 18 commercial Rhodiola rosea rhizome powder extract samples did not display a consistent Rapid Resolution Liquid Chromatography (RRLC) profile and were missing the characteristic peaks of rosarin, rosavin, and rosin or did not meet the claimed contents standards for the marker compounds for authentic R. rosea rhizomes (Ma et al., 2011). Similarly, a study by Booker A. et al. (2016) assessing the composition of 40 commercial preparations available on the EU market, claiming to contain R. rosea, confirmed the absence of rosavin in approximately 20% of the cases. Furthermore, in the remaining 80%, based on NMR Spectroscopy and HPTLC methods, the rosavin content was lower than declared. Additionally, 25% of tested dietary supplements were found to be substituted with other Rhodiola species, and one product was adulterated with synthetic 5–hydroxytryptophan (5–HTP) (A. Booker et al., 2016). A study conducted by Ruhsam M et al., in 2018 found that only 5 of 13 roseroot supplements contained R. rosea, with no visible admixture of other species (Ruhsam and Hollingsworth, 2018). In a study presented by Marchev A. et al., of 30 commercial products purchased from various suppliers, only 23% had rosavin and salidroside levels consistent with the amounts specified on the label, as confirmed by HPLC analysis. Additionally, 37% of the products did not meet the typical standardisation (1% salidroside and 3% rosavins), which may suggest potential adulteration (Marchev et al., 2020). In the study by Cunningham A. et al., they also found the presence of additives of extracts from the roots of Astragalus and Lycium species in preparations, declaring the presence of species of the Rhodiola genus (Cunningham et al., 2020).
Despite a smaller research group, our study indicates that almost 60% of the tested preparations did not contain the declared content of a single raw material–R. rosea roots/rhizomes or their extracts. Moreover, we detected the presence of rosavin in only 9 out of 13 tested preparations (using the LC–MS method), with 4 of these showing only trace amounts of the marker compound. Notably, the misidentification or accidental use of other Rhodiola species was most frequently observed in the tablet/capsule group. This could be attributed to the ease of substituting powdered extracts and the challenges associated with their precise identification.
Numerous studies have evaluated the quality and composition of commercial R. rosea products, highlighting the potential substitution of R. rosea with other Rhodiola species in dietary supplements. Such substitutions are often driven by the greater availability and ease of large–scale cultivation of alternative species. Notably, while the substitution of R. rosea with R. crenulata is widely recognized as a primary authenticity issue in R. rosea products, R. crenulata roots and rhizomes are preferred in Traditional Chinese Medicine (TCM) due to their higher availability in Asia, consistent with previous reports. However, from a safety perspective, the consumption of R. rosea is endorsed by the European Medicines Agency, whereas other species within the Rhodiola genus lack similar regulatory support. Furthermore, prior research underscores the efficacy of R. rosea as an adaptogen, demonstrating its beneficial effects in managing stress–related conditions, enhancing both mental and physical performance, and supporting immune function. In addition, few reports indicate its potential effectiveness in the adjunctive therapy and prevention of microglia–generated neuroinflammation associated with the development of neurodegenerative diseases, e.g., Parkinson’s or Alzheimer’s disease (Morgan and Grundmann, 2017).
The interchangeable use of different species within the same genus may, therefore, create some variations in chemical composition, potentially affecting the quality, safety, and biological activity of the product. The assessment of the anti–inflammatory effect of the tested preparations in the model of LPS–stimulated microglia may allow for the indirect correlation of the quality of the tested extracts with their biological effect.
Among the tested samples, the highest anti–inflammatory activity was observed for Z3 and Z4 among the unmodified raw material samples, K4 among the capsule formulations, T1 and T3 among the tablet formulations, and P1 in the powdered form (Figures 7, 8). Notably, the most significant inhibition of both TNF–α and IL–6 secretion was recorded for the ethanolic extract of preparation Z3, which was confirmed to contain Rhodiola rosea. Specifically, the infusion of Z3 resulted in TNF–α secretion inhibition, reducing its level to 62.0% ± 9.5%. Furthermore, Z3 reduced IL–6 secretion to 61.4% ± 14.2% and 74.8% ± 7.7% for the ethanol extract and infusion, respectively, relative to the 100% release observed in the stimulated control (Figure 8). The anti–inflammatory effects of R. rosea crude extract and its primary constituents, as well as their concentration–dependent modulation of iNOS, TNF–α, IL–1β, IL–6, and other inflammatory mediators, have been previously documented in the studies of Lee et al. and Jiang et al., among others (Borgonetti et al., 2020; Jiang et al., 2024; Lee et al., 2013). These findings are particularly relevant in the context of neuroinflammation modulation, a mechanism proposed to underlie the anti–stress effects of adaptogenic formulations (Ahmed et al., 2016; Guo et al., 2021).
An analysis of the composition of biologically active preparations suggests that their anti–inflammatory effects may partially correlate with the concentration of the marker compound for Rhodiola rosea. A comparative assessment of rosavin peak intensities in the analyzed samples of unprocessed plant material (Supplementary Table S3A, Supplementary Material) revealed a significantly higher concentration of rosavin in the Z3 preparation compared to the other samples.
Previous research has emphasized the role of phenylpropanoid derivatives, such as rosavins, and phenylethanol derivatives in contributing to the adaptogenic properties of R. rosea, exerting pleiotropic pharmacological effects on the neuroendocrine and immune systems (Fan et al., 2022; Maadawi, 2017). However, most studies have primarily focused on the biological activity of salidroside, with limited investigations into the effects of other compounds, including rosavin, within this cellular model (Lee et al., 2013; Zhang et al., 2022).
Thus, the neuroinflammation model employed in the present study enabled the evaluation of rosavin activity, representing one of the first investigations of this compound in a microglial cell model. As demonstrated, rosavin significantly and dose–dependently inhibited IL–6 and TNF–α secretion in LPS–stimulated microglia. Specifically, at a concentration of 5 μM, rosavin reduced IL–6 secretion to 34.3% ± 3.3% compared to the stimulated control (100% secretion), while a 43.5% ± 4.9% inhibition was observed at a concentration of 2 µM (Figure 8). Although data on rosavin’s effects in microglial cell models remain scarce, the present findings align with previous reports on its anti–inflammatory properties in other models, such as murine macrophages (Liu et al., 2024).
Interestingly, some preparations that did not contain R. rosea or rosavin—specifically P1, T1, and T3—also exhibited notable anti-inflammatory activity. These findings indicate that, although the selected cellular model is effective in capturing the activity of rosavin and standardized extracts, it may be insufficient for evaluating the overall quality or authenticity of complex commercial formulations.
It is also important to note that, although the BV2 microglial cell line is an immortalized murine model that does not fully capture the complexity of human microglia, it remains a widely used and well-established system for the preliminary investigation of neuroinflammatory mechanisms. Owing to their reproducible and well-characterized response to pro-inflammatory stimuli such as LPS, BV2 cells offer a practical and informative platform for the initial evaluation of anti-inflammatory properties in natural products (Marrazzo et al., 2023). Nevertheless, limitations related to species origin and monoculture conditions highlight the need for future studies employing more physiologically relevant models, including human-derived systems or co-culture approaches, to more accurately assess the biological activity of such preparations.
Nevertheless, our study confirms the potential of Rhodiola rosea as an agent mitigating neuroinflammatory processes, with rosavin identified as a compound potentially responsible for the observed biological activity of the raw material. With regard to other species within the Rhodiola genus, a comprehensive analysis should be conducted to assess their biological activities and to identify the specific compounds responsible for the observed effects.
The growth of the dietary supplement market necessitates detailed analyses of product composition and quality control. The increasing number of companies producing dietary supplements, along with rising consumer demand increases the risk of intentional or accidental contamination or incorrect product composition. European regulations governing the content of medicinal products containing Rhodiola rosea roots, rhizomes or their extracts necessitate the production of high–quality preparations with confirmed efficacy. The questionable quality of available preparations, claiming to contain R. rosea roots or rhizomes, may stem from either intentional or unintentional substitution of raw materials, as well as improper storage, which can lead to the degradation of key marker compounds. The objectives of this study were achieved by using an interdisciplinary approach (primarily based on LC–MS analysis) to better understand the sources of potential adulteration of individual herbal samples and their impact on biological activity. The identification of R. rosea in only 40% of the preparations underlines that rigorous control and standardisation of herbal supplements are crucial to understanding their therapeutic activity and preventing adulteration.
Data availability statement
The original contributions presented in the study are included in the article/Supplementary Material, further inquiries can be directed to the corresponding author.
Ethics statement
Ethical approval was not required for the studies on animals in accordance with the local legislation and institutional requirements because only commercially available established cell lines were used.
Author contributions
MK-J: Methodology, Data curation, Writing – original draft, Conceptualization, Investigation, Visualization, Funding acquisition. KC: Investigation, Writing – review and editing. AP: Writing – review and editing, Investigation. AB: Writing – review and editing, Investigation, Funding acquisition. MO: Resources, Writing – review and editing. AK: Conceptualization, Methodology, Writing – review and editing, Supervision, Formal Analysis, Project administration, Data curation.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the Medical University of Warsaw, the Students’ Mini-grant (No. WF4/3/F/MG/N/23).
Acknowledgments
The authors wish to thank Monika Marciniak, Małgorzata Lipowska and Sebastian Doktór (all Medical University of Warsaw) for their technical assistance.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2025.1608767/full#supplementary-material
References
Ahmed, T., Raza, S. H., Maryam, A., Setzer, W. N., Braidy, N., Nabavi, S. F., et al. (2016). Ginsenoside Rb1 as a neuroprotective agent: a review. Brain Res. Bull. 125, 30–43. doi:10.1016/j.brainresbull.2016.04.002
Alperth, F., Turek, I., Weiss, S., Vogt, D., and Bucar, F. (2019). Qualitative and quantitative analysis of different Rhodiola rosea rhizome extracts by UHPLC-DAD-ESI-MSn. Sci. Pharm. 87 (2), 8. doi:10.3390/scipharm87020008
Avula, B., Wang, Y. H., Ali, Z., Smillie, T. J., Filion, V., Cuerrier, A., et al. (2009). RP-HPLC determination of phenylalkanoids and monoterpenoids in Rhodiola rosea and identification by LC-ESI-TOF. Biomed. Chromatogr. 23 (8), 865–872. doi:10.1002/bmc.1198
Blomkvist, J., Taube, A., and Larhammar, D. (2009). Perspective on roseroot (Rhodiola rosea) studies. Planta Med. 75 (11), 1187–1190. doi:10.1055/s-0029-1185720
Booker, A., Jalil, B., Frommenwiler, D., Reich, E., Zhai, L., Kulic, Z., et al. (2016). The authenticity and quality of Rhodiola rosea products. Phytomedicine 23 (7), 754–762. doi:10.1016/j.phymed.2015.10.006
Booker, A., Zhai, L., Gkouva, C., Li, S., and Heinrich, M. (2016). From traditional resource to global commodities: a comparison of rhodiola species using NMR spectroscopy-metabolomics and HPTLC [original research]. Front. Pharmacol. 7, 254. doi:10.3389/fphar.2016.00254
Borgonetti, V., Governa, P., Biagi, M., Dalia, P., and Corsi, L. (2020). Rhodiola rosea L. modulates inflammatory processes in a CRH-activated BV2 cell model. Phytomedicine, 68, 153143. doi:10.1016/j.phymed.2019.153143
Brinckmann, J. A., Cunningham, A. B., and Harter, D. E. V. (2021). Running out of time to smell the roseroots: reviewing threats and trade in wild Rhodiola rosea L. J. Ethnopharmacol. 269, 113710. doi:10.1016/j.jep.2020.113710
Bykov, V. A., Zapesochnaya, G. G., and Kurkin, V. A. (1999). Traditional and biotechnological aspects of obtaining medicinal preparations from Rhodiola Rosea L. (A review). Pharm. Chem. J. 33 (1), 29–40. doi:10.1007/BF02508414
Center, Q. C. M. R. (2016). Global Rhodiola rosea P.E. market 2016: industry trends, sales, supply, demand, analysis and forecast to 2021. Beijing, China.
Chen, K., Liu, J., Ma, Z., Duan, F., Guo, Z., and Xiao, H. (2021). Rapid identification of chemical constituents of Rhodiola crenulata using liquid chromatography-mass spectrometry pseudotargeted analysis. J. Sep. Sci. 44 (20), 3747–3776. doi:10.1002/jssc.202100342
Cropley, M., Banks, A. P., and Boyle, J. (2015). The effects of Rhodiola rosea L. Extract on anxiety, stress, cognition and other mood symptoms. Phytother. Res. 29 (12), 1934–1939. doi:10.1002/ptr.5486
Cuerrier A, T. Y., and Rapinski, M. (2014). “Ethnobotany and conservation of Rhodiola species,” in Rhodiola rosea. Editors A. Cuerrier, and N. K. Ampong (Boca Raton, FL: CRC Press).
Cunningham, A. B., Li, H. L., Luo, P., Zhao, W. J., Long, X. C., and Brinckmann, J. A. (2020). There “ain't no mountain high enough”? the drivers, diversity and sustainability of China's Rhodiola trade. J. Ethnopharmacol. 252, 112379. doi:10.1016/j.jep.2019.112379
European Food Safety Authority (2012). General function health claims under Article 13. Available online at: http://www.efsa.europa.eu/en/topics/topic/article13.htm.
European Commission (2020). Cosmetic ingredient database. Eur. Comm. Dir. general Intern. Mark. industry, Entrepreneursh. SMEs. Available online at: https://ec.europa.eu/growth/sectors/cosmetics/cosing_en.
European Food Safety Authority (EFSA) (2012). Scientific Opinion on the substantiation of a health claim related to Rhodiola rosea L. extract and reduction of mental fatigue pursuant toArticle 13(5) of Regulation (EC) No 1924/20061. Available online at: https://efsa.onlinelibrary.wiley.com/doi/epdf/10.2903/j.efsa.2012.2805.
European Medicine Agency (2012). Community herbal monograph on Rhodiola rosea L., rhizoma et radix. Available online at: https://www.ema.europa.eu/en/documents/herbal-monograph/final-community-herbal-monograph-rhodiola-rosea_en.pdf.
Ezra Bejar, R. U., and John, H. C. I. I. (2017). Adulteration of rhodiola (Rhodiola rosea) rhizome and root and extracts. Bot. Adulterants Bull.
Fan, F., Xu, N., Sun, Y., Li, X., Gao, X., Yi, X., et al. (2022). Uncovering the metabolic mechanism of salidroside alleviating microglial hypoxia inflammation based on microfluidic chip-mass spectrometry. J. Proteome Res. 21 (4), 921–929. doi:10.1021/acs.jproteome.1c00647
Fisheries, N. (2022). Convention on international trade in endangered species of wild fauna and flora.
Galambosi, B. (2006). Demand and availability of rhodiola rosea L. Raw material. Frontis 17, 223–236. doi:10.1007/1-4020-5449-1_16
Gao, C., Jiang, J., Tan, Y., and Chen, S. (2023). Microglia in neurodegenerative diseases: mechanism and potential therapeutic targets. Signal Transduct. Target. Ther. 8 (1), 359. doi:10.1038/s41392-023-01588-0
Gontcharova, S. B., Gontcharov, A. A., Yakubov, V. V., and Kondo, K. (2009). Seed surface morphology in some representatives of the genus rhodiola sect. Rhodiola (Crassulaceae) in the Russian far east. Flora - Morphol. Distribution, Funct. Ecol. Plants, 204(1), 17–24. doi:10.1016/j.flora.2008.01.009
Guo, Y., Xie, J., Zhang, L., Yang, L., Ma, J., Bai, Y., et al. (2021). Ginsenoside Rb1 exerts antidepressant-like effects via suppression inflammation and activation of AKT pathway. Neurosci. Lett. 744, 135561. doi:10.1016/j.neulet.2020.135561
Han, F., Li, Y., Ma, L., Liu, T., Wu, Y., Xu, R., et al. (2016a). A rapid and sensitive UHPLC-FT-ICR MS/MS method for identification of chemical constituents in Rhodiola crenulata extract, rat plasma and rat brain after oral administration. Talanta, 160, 183–193. doi:10.1016/j.talanta.2016.07.014
Han, F., Li, Y., Mao, X., Xu, R., and Yin, R. (2016b). Characterization of chemical constituents in Rhodiola Crenulate by high-performance liquid chromatography coupled with Fourier-transform ion cyclotron resonance mass spectrometer (HPLC-FT-ICR MS). J. Mass Spectrom. 51 (5), 363–368. doi:10.1002/jms.3764
Howes, M. J. R., Quave, C. L., Collemare, J., Tatsis, E. C., Twilley, D., Lulekal, E., et al. (2020). Molecules from nature: reconciling biodiversity conservation and global healthcare imperatives for sustainable use of medicinal plants and fungi. Plants, People, Planet 2 (5), 463–481. doi:10.1002/ppp3.10138
Hung, S. K., Perry, R., and Ernst, E. (2011). The effectiveness and efficacy of Rhodiola rosea L.: a systematic review of randomized clinical trials. Phytomedicine, 18(4), 235–244. doi:10.1016/j.phymed.2010.08.014
Iannuzzo, F., Schiano, E., Pastore, A., Guerra, F., Tenore, G. C., Novellino, E., et al. (2024). Controlled cultivation confers Rhodiola rosea synergistic activity on muscle cell homeostasis, metabolism and antioxidant defense in primary human myoblasts. Antioxidants 13 (8), 1000. doi:10.3390/antiox13081000
Jiang, L., Yang, D., Zhang, Z., Xu, L., Jiang, Q., Tong, Y., et al. (2024). Elucidating the role of Rhodiola rosea L. in sepsis-induced acute lung injury via network pharmacology: emphasis on inflammatory response, oxidative stress, and the PI3K-AKT pathway. Pharm. Biol. 62 (1), 272–284. doi:10.1080/13880209.2024.2319117
Lamadrid, P., Williams, D. K., Kilpatrick, M. W., Bickford, P. C., and Sanberg, C. (2019). The impact of dietary supplement NT-020 with Rhodiola rosea on energy, fatigue, and perceived exertion. Funct. Foods Health Dis. 9 (11), 706–718. doi:10.31989/ffhd.v9i11.656
Lee, T. H., Hsu, C. C., Hsiao, G., Fang, J. Y., Liu, W. M., and Lee, C. K. (2016). Anti-MMP-2 activity and skin-penetrating capability of the chemical constituents from Rhodiola rosea. Planta Med. 82 (8), 698–704. doi:10.1055/s-0042-101033
Lee, Y., Jung, J. C., Jang, S., Kim, J., Ali, Z., Khan, I. A., et al. (2013). Anti-inflammatory and neuroprotective effects of constituents isolated from rhodiola rosea. Evid. Based Complement. Altern. Med. 2013, 514049. doi:10.1155/2013/514049
Li, B., Yang, W., Ge, T., Wang, Y., and Cui, R. (2022). Stress induced microglial activation contributes to depression. Pharmacol. Res., 179, 106145. doi:10.1016/j.phrs.2022.106145
Liu, Q. H., Zhang, K., Feng, S. S., Zhang, L. J., Li, S. Y., Wang, H. Y., et al. (2024). Rosavin alleviates LPS-induced acute lung injure by modulating the TLR-4/NF-κB/MAPK singnaling pathways. Int. J. Mol. Sci. 25 (3), 1875. doi:10.3390/ijms25031875
Liu, Y., Chen, C., Qiu, J., Fang, Z., Wu, H., Zhang, X., et al. (2019). Characterization of the chemical constituents in Hongjingtian injection by liquid chromatography quadrupole time-of-flight mass spectrometry. Biomed. Chromatogr. 33 (3), e4446. doi:10.1002/bmc.4446
Liu, Z., Liu, Y., Liu, C., Song, Z., Li, Q., Zha, Q., et al. (2013). The chemotaxonomic classification of Rhodiola plants and its correlation with morphological characteristics and genetic taxonomy. Chem. Central J. 7 (1), 118. doi:10.1186/1752-153X-7-118
Lu, Y., Deng, B., Xu, L., Liu, H., Song, Y., and Lin, F. (2022). Effects of rhodiola rosea supplementation on exercise and sport: a systematic review. Front. Nutr. 9, 856287–2022. doi:10.3389/fnut.2022.856287
Ma, G. P., Zheng, Q., Xu, M. B., Zhou, X. L., Lu, L., Li, Z. X., et al. (2018). Rhodiola rosea L. Improves learning and memory function: preclinical evidence and possible mechanisms. Front. Pharmacol. 9, 1415. doi:10.3389/fphar.2018.01415
Ma, Y. C., Wang, X. Q., Hou, F. F., Ma, J., Luo, M., Lu, S., et al. (2011). Rapid resolution liquid chromatography (RRLC) analysis for quality control of Rhodiola rosea roots and commercial standardized products. Nat. Prod. Commun. 6 (5), 645–650. doi:10.1177/1934578x1100600515
Maadawi, Z. M. E. (2017). Conditioned medium derived from salidroside-pretreated mesenchymal stem cell culture ameliorates mouse lipopolysaccharide-induced cerebral neuroinflammation- histological and immunohistochemical study. Int. J. Stem Cells 10 (1), 60–68. doi:10.15283/ijsc16055
Mao, J. J., Xie, S. X., Zee, J., Soeller, I., Li, Q. S., Rockwell, K., et al. (2015). Rhodiola rosea versus sertraline for major depressive disorder: a randomized placebo-controlled trial. Phytomedicine 22 (3), 394–399. doi:10.1016/j.phymed.2015.01.010
Marchev, A. S., Koycheva, I. K., Aneva, I. Y., and Georgiev, M. I. (2020). Authenticity and quality evaluation of different Rhodiola species and commercial products based on NMR-spectroscopy and HPLC. Phytochem. Anal. 31 (6), 756–769. doi:10.1002/pca.2940
Marrazzo, P., Mandrone, M., Chiocchio, I., Zambonin, L., Barbalace, M. C., Zalambani, C., et al. (2023). By-product extracts from Castanea sativa counteract hallmarks of neuroinflammation in a microglial model. Antioxidants 12 (4), 808. doi:10.3390/antiox12040808
Morgan, L. A., and Grundmann, O. (2017). Preclinical and potential applications of common western herbal supplements as complementary treatment in Parkinson's disease. J. Diet. Suppl. 14 (4), 453–466. doi:10.1080/19390211.2016.1263710
Nakamura, S., Li, X., Matsuda, H., and Yoshikawa, M. (2008). Bioactive constituents from Chinese natural medicines. XXVIII. Chemical structures of acyclic alcohol glycosides from the roots of Rhodiola crenulata. Chem. Pharm. Bull. (Tokyo) 56 (4), 536–540. doi:10.1248/cpb.56.536
Nikolaichuk, H., Typek, R., Gnat, S., Studziński, M., and Choma, I. M. (2021). Effect-directed analysis as a method for quality and authenticity estimation of Rhodiola rosea L. preparations. J. Chromatogr. A, 1649, 462217. doi:10.1016/j.chroma.2021.462217
Olennikov, D. N., Chirikova, N. K., Vasilieva, A. G., and Fedorov, I. A. (2020). LC-MS profile, gastrointestinal and gut microbiota stability and antioxidant activity of Rhodiola rosea herb metabolites: a comparative study with subterranean organs. Antioxidants (Basel) 9 (6), 526. doi:10.3390/antiox9060526
Petsalo, A., Jalonen, J., and Tolonen, A. (2006). Identification of flavonoids of Rhodiola rosea by liquid chromatography-tandem mass spectrometry. J. Chromatogr. A, 1112(1), 224–231. doi:10.1016/j.chroma.2005.11.056
Pu, W.-l., Zhang, M.-y., Bai, R.-y., Sun, L.-k., Li, W.-h., Yu, Y.-l., et al. (2020). Anti-inflammatory effects of Rhodiola rosea L.: a review. Biomed. and Pharmacother., 121, 109552. doi:10.1016/j.biopha.2019.109552
Ruhsam, M., and Hollingsworth, P. M. (2018). Authentication of Eleutherococcus and Rhodiola herbal supplement products in the United Kingdom. J. Pharm. Biomed. Analysis, 149, 403–409. doi:10.1016/j.jpba.2017.11.025
Tao, H., Wu, X., Cao, J., Peng, Y., Wang, A., Pei, J., et al. (2019). Rhodiola species: a comprehensive review of traditional use, phytochemistry, pharmacology, toxicity, and clinical study. Med. Res. Rev. 39 (5), 1779–1850. doi:10.1002/med.21564
United States Pharmacopeial Convention (2014). “United States Pharmacopeial Convention Rhodiola rosea root and rhizome. Final authorized version 1.0,” in Herbal medicines Compendium.
Yousef, G. G., Grace, M. H., Cheng, D. M., Belolipov, I. V., Raskin, I., and Lila, M. A. (2006). Comparative phytochemical characterization of three Rhodiola species. Phytochemistry, 67(21), 2380–2391. doi:10.1016/j.phytochem.2006.07.026
Zakharenko, A. M., Razgonova, M. P., Pikula, K. S., and Golokhvast, K. S. (2021). Simultaneous determination of 78 compounds of Rhodiola rosea extract by supercritical CO(2)-extraction and HPLC-ESI-MS/MS spectrometry. Biochem. Res. Int. 2021, 9957490. doi:10.1155/2021/9957490
Zhang, J.-Q., Meng, S.-Y., Wen, J., and Rao, G.-Y. (2014). Phylogenetic relationships and character evolution of Rhodiola (Crassulaceae) based on nuclear ribosomal ITS and plastid trnL-F and psbA-trnH sequences. Syst. Bot. 39 (2), 441–451. doi:10.1600/036364414x680753
Zhang, S., Jiang, S., Deng, N., Zheng, B., Li, T., and Liu, R. H. (2022). Phytochemical profiles, antioxidant activity and antiproliferative mechanism of Rhodiola rosea L. Phenolic extract. Nutrients 14 (17), 3602. doi:10.3390/nu14173602
Keywords: Rhodiola L., golden root, rosavin, adaptogens, dietary supplement, microglia, neuroinflammation
Citation: Kołtun-Jasion M, Czerwiec K, Parzonko A, Bakiera A, Ożarowski M and Kiss AK (2025) Comprehensive profiling of Rhodiola rosea roots and corresponding products: phytochemical insights and modulation of neuroinflammation in BV2 microglial cell model. Front. Pharmacol. 16:1608767. doi: 10.3389/fphar.2025.1608767
Received: 09 April 2025; Accepted: 05 June 2025;
Published: 02 July 2025.
Edited by:
Agnieszka Szopa, Jagiellonian University Medical College, PolandReviewed by:
Pasquale Marrazzo, University of Urbino Carlo Bo, ItalyTimea Teglas, Hungarian University of Sports Science, Hungary
Copyright © 2025 Kołtun-Jasion, Czerwiec, Parzonko, Bakiera, Ożarowski and Kiss. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Małgorzata Kołtun-Jasion, bWFsZ29yemF0YS5rb2x0dW5Ad3VtLmVkdS5wbA==