 Yi Hou
Yi Hou Hong Li
Hong Li Luochen Zhu
Luochen Zhu Tian Quan
Tian Quan Xianhu Feng4
Xianhu Feng4 Yuan Bian
Yuan Bian Yuxun Wei
Yuxun Wei- 1Pharmacy Department, The Center of Clinical Trials, The People’s Hospital of Zhongjiang, Deyang, China
- 2West China School of Medicine, Sichuan University, Chengdu, China
- 3Department of Pharmacy, Nanjing University of Chinese Medicine, Nanjing, China
- 4Department of Pharmacy, Nanchong Central Hospital, North Sichuan Medical College, Nanchong, Sichuan, China
- 5Molecular Urooncology Department of Urology Klinikum rechts der Isar Technical University of Munich, München, Germany
- 6Department of Oncology, Xichang People’s Hospital, Xichang, China
Paeoniflorin (PF) is widely present in species of the Paeonia genus. In recent years, numerous preclinical studies have shown that PF has preventive and therapeutic effects on various neurological diseases, particularly in the prevention and treatment of depression. Additionally, some classic traditional Chinese medicine formulas containing PF, such as Xiaoyao San, Chaihu Shugan San, and Sini San, have been proven to significantly improve depressive symptoms. However, the antidepressant mechanisms of PF and its containing classic traditional Chinese medicine formulas are not yet fully understood. PF, as a natural glycoside metabolite with a wide margin of safety and good tolerance, exhibits certain toxicity at high concentrations. The differences in standardized methods between the traditional formulations, such as extraction processes, dosages, and inherent metabolite variability in formulations, may affect the interpretation of results and clinical applications. Therefore, this article reviews the antidepressant mechanisms of PF from the perspectives of inhibiting the hypothalamic-pituitary-adrenal axis, increasing the levels of monoamine neurotransmitters, suppressing oxidative stress and apoptosis, regulating calcium homeostasis, inhibiting neuroinflammation, modulating mitochondrial function, regulating cellular autophagy, and increasing the levels of brain-derived neurotrophic factor, and elucidates the antidepressant effects and mechanisms of traditional Chinese medicine formulations containing PF. Additionally, we describe the physicochemical properties, toxinology, pharmacokinetic characteristics, and the transformation of PF in vivo. This review may contribute to the application of PF and its formulations in depression.
1 Introduction
Depression, the most prevalent mental disorder, imposes a huge burden on individual health and wellbeing as well as social and economic development, which has become one of the major global mental health challenges. According to Global Burden of Diseases (GBD) data survey statistics, depression is one of the leading causes of disability-adjusted life years (DALYs) for people aged 10–49 years (10–24 years: fourth, 25–49 years: Sixth) (Diseases and Injuries, 2020). When only mental disorders were considered, depression ranked highest of DALYs in all age groups except those aged 0–14 years, as behavioral disorders were the main cause of burden (Collaborators, 2022; Whiteford et al., 2013). According to the World Health Organization (WHO), depression will become the leading cause of death worldwide by 2030 (Tian et al., 2013). Depression, as defined by the American Psychiatric Association in 2013, is a common and profoundly heterogeneous disorder (Johnston et al., 2019), usually accompanied by physical, behavioral, and psychological symptoms such as changes in appetite, headaches, sleep disturbances, persistent low mood, lack of pleasure, sexual problems, and suicidal ideation (Marwaha et al., 2023; Disner et al., 2011). These physical and emotional disorders seriously affect the work and daily life of individuals, and significantly reduce their quality of life and happiness. Therefore, the search for effective treatment should be an important research topic.
Brain stimulation, including repetitive transcranial magnetic stimulation, transcranial direct current stimulation, and deep brain stimulation are common treatment for depression nowadays (Marwaha et al., 2023). However, regrettably, these seemingly attractive treatment methods can still trigger a series of side effects, including headaches, scalp discomfort, fatigue, pain, dizziness, insomnia, eye and nose issues, as well as gastrointestinal problems (Miuli et al., 2021; Hett and Marwaha, 2020; Chen L. et al., 2020; Moffa et al., 2020; Berlow et al., 2019; Zhou et al., 2018; Dougherty et al., 2015). Due to the time and economic loss caused by the treatment, the benefit to the audience is limited. Medication still seems to play an irreplaceable and important role in severe cases of depression. In clinical settings, antidepressants such as TCAs, MAOIs, selective 5-HT reuptake inhibitors, and 5-HT-NE reuptake inhibitors are widely used for the treatment of depression. However, these traditional antidepressants still have inevitable side effects and drug-drug interactions, such as dry mouth, blurred vision, inability to drive, sexual dysfunction, decreased libido, headache, gastrointestinal symptoms, anxiety, agitation, and other common adverse events (Ray et al., 1987; Roose et al., 1994). More disappointingly, less than half of the patients treated with these medications show a beneficial therapeutic response and induce drug tolerance if used for a prolonged period of time (Cipriani et al., 2018; Rush et al., 2009). Therefore, there is an urgent need to develop antidepressant medications that act more rapidly, have better tolerability, offer superior therapeutic efficacy, and are associated with fewer side effects.
Numerous studies have shown that Complementary and Alternative Medicine (CAM) has extensive application value in the field of mental health treatment, particularly in the treatment of depression (Kessler et al., 2002; Unutzer et al., 2000; Knaudt et al., 1999). Natural drugs as alternative treatment methods may be a promising attempt, which have significant therapeutic effects, relatively small side effects, and low prices. Certain extracts or single units of natural medicines have shown great potential to treat psychiatric disorders, such as PF (Peng et al., 2022; Hong et al., 2022; Guo et al., 2022).
Paeoniflorin (PF; C23H28O11) is a water-soluble monoterpene bicyclic glycoside extracted from Paeonia lactiflora Pall. [Paeoniaceae, Paeonia lactiflora root] is a plant known for its medicinal and edible properties. First isolated in 1963, its chemical structure was determined in 1972 (Wang Z. et al., 2022) (Figure 1). As an additional nutritional metabolite in plant-based foods, PF offers health benefits that extend beyond basic nutritional value, with increasing evidence supporting its positive impact on human health. In recent years, most studies have found that PF has a wide range of pharmacological effects in vitro and in vivo, including anti-inflammatory (Tu et al., 2019; Xin et al., 2019; Wang C. et al., 2013), anti-oxidation (Chen et al., 2011), anti-thrombosis (Ye et al., 2016), anti-convulsion (Hino et al., 2012), analgesia (Zhang et al., 2009), cardioprotection (Chen H. et al., 2018; Wang et al., 2020), neuroprotection (Mao et al., 2012a; Kong et al., 2020), liver protection (Jiang et al., 2014), antidepressant (Qiu et al., 2013; Cheng et al., 2021), anti-tumor (Lu et al., 2014), immunomodulation and so on (Wang Z. et al., 2022; Zhang and Wei, 2020). PF can be used as a potential therapeutic agent for many diseases, such as psoriasis (Bai et al., 2020), atherosclerosis (Yu et al., 2022; Duan et al., 2021), and depression (Hong et al., 2022; Guo et al., 2022), due to the inherent advantages of low toxicity, high efficiency, and safety. The intrinsic pharmacological mechanism of PF has received great attention from researchers and clinicians. Currently, more and more evidence suggests that PF has significant pharmacological activity against various neurological diseases, for instance, cerebral ischemia (Wu et al., 2020; Guo et al., 2012; Liu et al., 2005), vascular dementia (Luo et al., 2018; Zhang et al., 2016), Parkinson’s disease (Zheng et al., 2017; Liu H. Q. et al., 2006), and depression (Li et al., 2020; Chen et al., 2019). Especially for depression, PF may exert surprising antidepressant effects through oxidative stress, apoptosis, neuroinflammation, and other mechanisms (Guo et al., 2022). In traditional Chinese medicine, some prescriptions containing PF, such as Xiaoyaosan (XYS) (Chen C. et al., 2020), Chaihu-Shugan-San (CSS) (Kim et al., 2005), and Sinisan (SNS), have been proven to have significant improvement effects on depression and are used as an alternative treatment for depression. This study reviews the pharmacokinetic properties of PF in vivo and pharmacological mechanisms in the treatment of depression, further determines the clinical application of PF in depression, and provides literature support for its drug formation research.
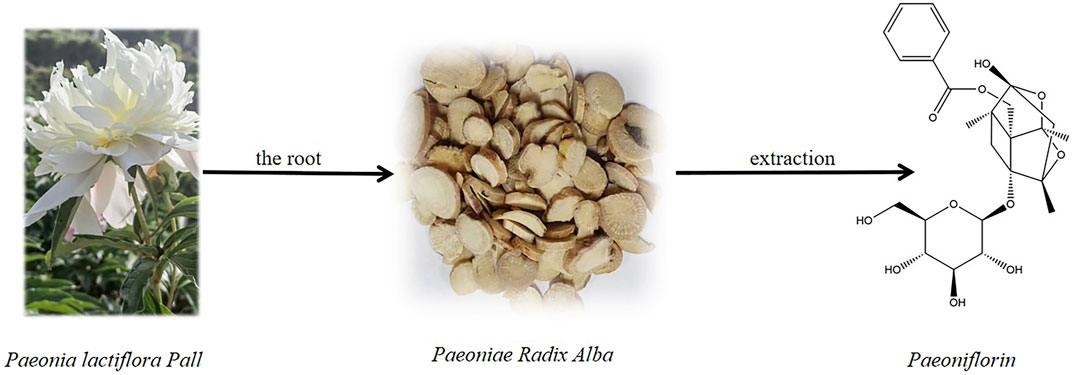
Figure 1. Paeoniflorin and the source from the roots of Paeonia lactiflora Pall.
2 Methods
A comprehensive search was conducted across popular and widely used databases, including PubMed, Web of Science, Scopus, Science Direct, Google Scholar, and CNKI, utilizing various search strings. The search terms included, but were not limited to, “paeoniflorin, physicochemical properties, pharmacokinetics, antidepressant mechanisms, hypothalamic-pituitary-adrenal (HPA) axis, monoamine neurotransmitters, oxidative stress, apoptosis, calcium homeostasis, neuroinflammation, mitochondrial function, autophagy, and brain-derived neurotrophic factor (BDNF),” as well as the application of paeoniflorin in traditional Chinese medicine formulations (e.g., XiaoyaoSan, ChaihuShuganSan, SiniSan, etc.). The search was conducted in both English and Chinese, relying solely on online databases without incorporating other physical sources. Inclusion criteria encompassed studies related to the aforementioned aspects of paeoniflorin, including basic pharmacological research, pharmaceutical research, clinical research, and other formally published literature. Exclusion criteria consisted of studies that were irrelevant or minimally relevant to the specified aspects of paeoniflorin, duplicate publications, low-quality literature, and studies for which full texts were unavailable.
3 Physicochemical properties and metabolism
Paeoniflorin (molecular weight 480.5) is a monoterpene glucoside, a strong hydrophilic plant metabolite (logP: 2.88). The pharmacokinetics study found that the low bioavailability of PF was about 3%–4%, related to the low permeability caused by the high hydrophilicity of PF. The absorption site of PF is mainly the intestine, and the absorption rate of the aglycone is 48 times that of PF (Takeda et al., 1995; Takeda et al., 1997; Yu et al., 2019) (Figure 2). Liu ZQ et al. demonstrated that the effect of P-GP efflux protein on PF and the metabolism of intestinal microorganisms is another important reason for the low bioavailability of PF (Liu Z. Q. et al., 2006). PF is widely distributed in various tissues after entering the systemic circulation, mainly in the stomach, intestine, and heart (Minmin Zhao et al., 2014). Significantly, PF has a smaller ability to penetrate the blood-brain barrier (permeability coefficient: 0.587 × 10–6 to 0.705 × 10–6 cm/s) (Hu et al., 2016). The metabolism of PF is divided into two pathways: intestinal flora metabolism and enzyme metabolism in vivo. After oral administration, the part of PF was metabolized into two chiral counterparts (7R or 7S paeonimetabolin Ⅰ, 7R or 7S paeonimetabolin Ⅱ) by the intestinal flora in the human being (Hu et al., 2016; Shu et al., 1987), and about 42% of the remaining PF was converted to paeoniflorgenin in vivo by β-glucosidase LDH enzyme (Hsiu et al., 2003). Interestingly, PF decomposes into paeoniflorgenin, similar to human fecal bacteria. Further studies have found that some bacteria can achieve mutual conversion of PF and aglycones. In addition, both pathways can metabolize PF to benzoic acid and can cross the blood-brain barrier to act on the central nervous system (CNS), which proves to some extent that PF has a certain neuroprotective effect (Yu et al., 2019). PF is mainly excreted in urine as benzoic acid after oral administration, and its cumulative excretion is 50%. On the contrary, less is excreted with the prototype drug, in urine, bile, and feces (Takeda et al., 1995; Yu et al., 2019), of which the property is inseparable from the low bioavailability of PF and the conversion of PF to benzoic acid by intestinal flora (Liu Z. Q. et al., 2006). Meanwhile, PF entering the systemic circulation is still mainly excreted in the form of urine.
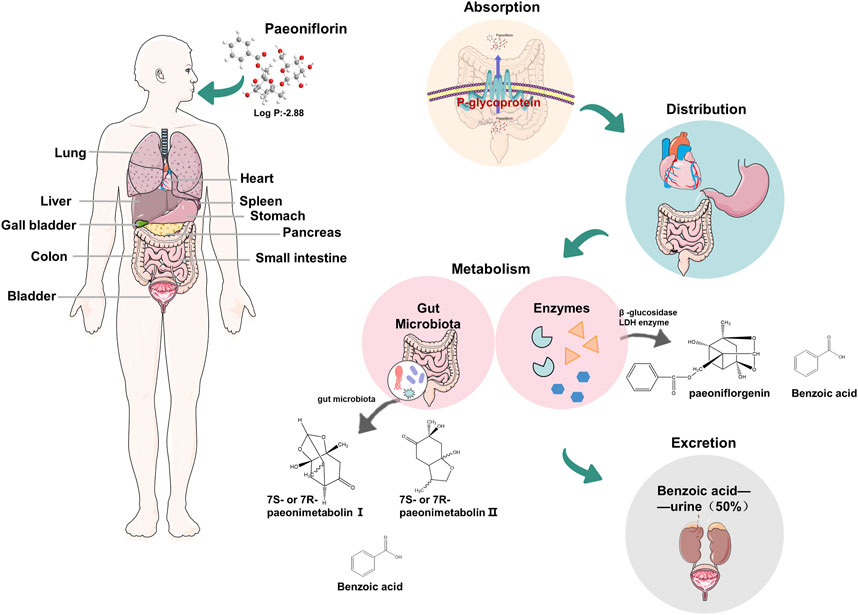
Figure 2. Physicochemical properties of paeoniflorin and the pharmacokinetics in humans.
4 Molecular mechanism of PF in depression
4.1 Hypothalamic-pituitary-adrenal axis
The hypothalamic-pituitary-adrenal (HPA) axis is one of the major biological stress response systems in humans, of which activation is essential to provide an appropriate biological response to stress (Guerry and Hastings, 2011). Activation of the HPA axis in stressful situations is a normal homeostatic mechanism that can contribute to the maintenance of stability and health by enabling adaptive changes in the body. For a healthy population, normal levels return when this stress response is no longer biologically significant. However, studies have found that abnormal activity of the HPA axis, such as excessive activity or persistent disorder, is closely related to the development of depression (Nelson and Davis, 1997; Curtis and Mendels, 1976; Bardeleben and Holsboer, 1989). Previous research has shown that the activity of the HPA axis is governed by vasopressin (AVP) and corticotropin-releasing factor (CRF) which is secreted by the hypothalamus and undergoes a cascade of events. Specifically, when the body is stimulated by stress, the adrenal glands secrete a surge of adrenaline, followed by the activation of the HPA axis. The hypothalamus secretes CRF for release into the portal circulation, and this in turn activates the pituitary to secrete the adrenocorticotropic hormone (ACTH), which ultimately stimulates the adrenal cortex to secrete glucocorticoids and cortisol (Pariante and Lightman, 2008). In addition, cortisol induces inhibitory feedback of the HPA axis by interacting with glucocorticoid (GR) and mineralocorticoid (MR), such as the secretion of ACTH, CRF, and AVP (Pariante and Lightman, 2008; Juruena et al., 2018; De Kloet et al., 1998). The normal physiological activity of the HPA described above maintains the relative homeostasis and health of the organism under certain stressful circumstances. However, excessive or prolonged psychological stress may alter the normal state of homeostasis in the body, which leads to depression, a serious mental illness that affects people’s mental and physical health. Studies have found that the most common biological abnormality in depression is HPA axis overactivity, which is characterized by increased cortisol, adrenal hyperplasia, and negative feedback abnormalities (Dinan and Scott, 2005; Dwyer et al., 2020). Fortunately, previous studies have shown that monoamine-based antidepressants can reverse stress-induced hyperactivity of the HPA axis (Seki et al., 2018). GR receptor antagonists, press-in receptor antagonists, and adrenocorticotropin-releasing hormone receptor antagonists can also exert antidepressant effects by blocking receptor activity to terminate hormone secretion resulting from HPA axis overactivity (Menke, 2019), suggesting that components of the HPA axis may be potential targets for the treatment of mood disorders (Thakore et al., 1997).
Most researchers suggest that PF has promising antidepressant activity by modulating HPA axis dysfunction. Qiu et al. found that intraperitoneal injection of PF (30 or 60 mg/kg) for 4 weeks significantly increased sucrose consumption, and decreased serum corticosterone (CORT) and ACTH levels in the chronic unpredictable stress (CUS) model group, which suggests that PF may exert antidepressant effects in CUS rats by regulating the HPA axis (Qiu et al., 2013). This preclinical study suggests and explores the antidepressant effects of PF and its underlying mechanisms; however, several significant limitations persist. Firstly, the generalizability of the findings is restricted due to the exclusive use of male SD rats as experimental subjects. Additionally, the CUS paradigm employed may not fully replicate the complex stressors encountered by humans. Furthermore, the study utilized only intraperitoneal (i.p.) injection as the method of administration, which may differ from clinical drug delivery methods. Increasing the number of studies on multiple routes of administration, such as oral and transdermal patches, and comparing the antidepressant effects and pharmacokinetic profiles of PF under different modes of administration may provide some reference for the clinical application of the drug. Similarly, the depression model of rats established that the forced swimming trial (FST) was treated with PF (10 mg/kg) or fluoxetine (20 mg/kg) by gavage three times at 24 h, 5 h, and 1 h prior to the behavioral experiments, including the forced swimming and the open field trials. Both fluoxetine and PF could significantly shorten the immobilization time of the 5-min forced swim. In addition, the distance traveled in the open field did not significantly change in both groups compared to the normal control in the Open-Field Test (OFT) (Mu et al., 2020). Further studies revealed that PF was able to increase tolerance to stressors in rats by modulating the hyperactive HPA axis, exerting antidepressant effects similar to those of fluoxetine, as evidenced by the significant reduction of corticotropin-releasing hormone (CRH), ACTH, and CORT in plasma and hippocampus (Yan-xia et al., 2014). Meanwhile, the total glucosides of paeoniflorin (TGP) are active metabolites extracted from Paeonia lactiflora Pall. [Paeoniaceae, Paeonia lactiflora root], including PF, hydroxyl-paeoniflorin, paeonin, albiflorin, benzoylpaeoniflorin. Whereas, PF is the main active metabolite of TGP, which accounts for more than 40% of TGP (Zhang and Wei, 2020). Mao et al. found that TGP significantly inhibited the behavioral and biochemical changes in the chronic unpredictable mild stress (CUMS) mice, and could dose-dependently reduce the serum CORT levels in CUMS-exposed mice. The TGP may also exert an antidepressant-like effect by regulating the function of HPA (Mao et al., 2009a).
Numerous studies have found that prenatal stress (PS) increases the risk of neurological, endocrine, and metabolic disorders, which induces depression in offspring. To stressed offspring, PF (15, 30, and 60 mg/kg/day) administered for 28 consecutive days significantly increased sucrose intake and reduced immobility time as well as the total number of crossings, center crossings, rearing, and grooming in model rats. However, PF could restore the levels of ACTH, CRH, and COR in PS offspring (Li et al., 2020). For the ovariectomized and CUS-induced menopause depression model rats, PF could downregulate the serum levels of CRH, CORT, and ACTH, correct the hyperfunctioning of the HPA axis, which resulted in a significant improvement in the abnormal behaviors of the model rats (Huang et al., 2015). In summary, PF results suggest that PF could regulate mood disorders and exert antidepressant-like effects by modulating the disordered HPA axis function.
4.2 Monoaminergic nervous system
Most trial and clinical evidence have shown that monoaminergic neurotransmitters such as serotonin (5-HT), norepinephrine (NE), and dopamine (DA) have a wide range of biological activities and are important regulators of a series of physiological activities, such as mental activity, behavioral state, and emotion in the CNS (Nemeroff, 2002). Thus, physiological changes in abnormal monoaminergic neurotransmitters, including 5-HT, NE, and DA signal transduction could change receptor regulation and function, or impaired intracellular signal processing (Liu Y. et al., 2018), which trigger various emotional changes (Hamon and Blier, 2013). 5-HT, a biochemical messenger and modulator synthesized by decarboxylation of L-tryptophan, produces a variety of “state-dependent” behavioral responses to different stimuli, which have been shown to be useful in the treatment of anxiety and obsessive-compulsive disorder. The defects in the 5-HT system lead to disorders such as depression, phobias, obsessive-compulsive disorder, generalized anxiety disorder, and post-traumatic stress disorder (De-Miguel and Trueta, 2005). The most common of the pathophysiologic hypotheses in depression is the monoamine hypothesis, which posits that alterations in monoamine neurotransmitters are responsible for the pathogenesis of depression, such as decreased concentrations, abnormal function, and defective transmission across the synaptic gaps in the brain (Hirschfeld, 2000). Evidence for this hypothesis comes mainly from clinical observations and animal experiments. Reserpine, the antihypertensive drug, has been shown to deplete central stores of monoamines, which can trigger depressive-like manifestations such as bradykinesia and sedation (Delgado, 2000). In contrast, isoniazid could increase the concentration of NE and 5-HT in the brain by inhibiting monoamine oxidase, making the subject feel euphoric and active (Buda et al., 2022). The monoamine oxidase inhibitors (MAOIs) iproniazid also have antidepressant effects when used in patients with tuberculosis (Delgado, 2000). Therefore, increasing the monoaminergic neurotransmitter levels and enhancing the function of monoaminergic neurotransmitter systems are effective options for the treatment of depression. According to this hypothesis, various antidepressant drugs have been discovered, such as tricyclic antidepressants (TCAs) that block the reuptake of monoamines in presynaptic neurons, and MAOIs that prevent the breakdown of monoamines after reuptake and enhance neurotransmitters to exert antidepressant effects (Kessing et al., 2024).
The study found that the treatment of PF significantly attenuated the decrease of NA, DA, 5-HT, and metabolite 5-hydroxyindoleacetic acid (5-HIAA) in chronic CUS model mice, as well as the increase in the ratio of 5-HIAA/5-HT in the model (Qiu et al., 2013; Wang Jing-xia et al., 2010). Similarly, Intragastric administration of PF (10 mg/kg) could increase the levels of 5-HT and NE in plasma and hippocampus of the forced swimming test (FST) depression model rats (Mu et al., 2020). Reserpine is a vesicle reuptake inhibitor that irreversibly inhibits the vesicular uptake of monoamine neurotransmitters, including NA, DA, and 5-HT, which are metabolized and depleted by monoamine oxidase (MAO) inducing behavioral and physiological changes in animals (Ye et al., 2024). Based on the above principles, the rifampicin-antagonistic experimental model was the first established animal model of depression (Bourin et al., 1983). Intragastric administration of PF or TGP dose-dependently antagonized hypothermia, akinesia, and blepharoptosis in reserpine-induced mice, and reversed reserpine-induced decreases in monoamine transmitters such as NE, DA, and 5-HT in the brain, which showed an obvious anti-resensitization effect (Mao et al., 2008; Guang-zhi, 2012; Jin Shumei and Cui, 2013). These studies suggested that the antidepressant-like effect of PF or TGP may be realized by protecting monoamine neurotransmitters. In addition, PF or TGP could protect monoamine neurotransmitters by dose-dependently inhibiting the activities of MAO-A and MAO-B in the mouse brain monoamine oxidase (Mao et al., 2009b).
5-HT1A receptors, the largest class of 5-HT receptor subtypes, are mainly distributed in the frontal cortex, hippocampal area, lateral septal nucleus, and dorsal nucleus of the middle suture, which are closely associated with anxiety and depression (Sargent et al., 2010). 5-HT2A receptors are densely distributed in the hippocampal area, amygdala, prefrontal cortex, and olfactory cortex executive area, closely associated with suicide, depression, and schizophrenia (Papp et al., 1994). The mRNA and protein expression levels of the 5-HT1A receptor in the hypothalamus of menopausal depression rats by CUMS model were significantly lower than those of normal rats, while the mRNA and protein expression levels of the 5-HT2A receptor were significantly higher than those of normal rats. After gavage of PF (10 mg/kg) for 2 weeks, the mRNA and protein expression levels of the 5-HT1A receptor in the hypothalamus of the model rats were increased, while the expression level of the 5-HT2A receptor was decreased. These results indicate that PF can treat climacteric depression by adjusting the different 5-HT receptor subtypes in the hypothalamic region of rats (Huang et al., 2015). The model established by ovariectomy combined with long-term CUS in this study, while effectively simulating the physiological changes and stressors associated with menopause, still fails to capture the complex etiology and manifestations of human menopausal depression, and cannot fully replicate the realities of the human condition. In addition, although serum CRH, ACTH, CORT, and prefrontal cortex monoaminergic neurotransmitter levels, mRNA, and protein expression were measured, which confirmed the antidepressant effect of PF at the molecular level. On this basis, we suggest an in-depth exploration of the specific molecular targets where PF acts and the related signalling pathways. In addition, PF could activate the release of monoamines in the rodent brain, inhibit the reuptake of NA and 5-HT, and increase the content of DOPAC and 5-HIAA in the brain, which are the metabolites of 5-HT and DA (Lin et al., 2019; Qingwei, 2018).
4.3 Oxidative stress
Oxidative stress is caused by the overproduction of reactive oxygen species (ROS) or defective antioxidant systems, strongly associated with diseases such as cardiovascular disease, cancer, and diabetes (Poprac et al., 2017). In addition, ROS is widely recognized as a main cause of brain damage. Specifically, the brain effectively regulates oxygen consumption and redox-generating capacity by neutralizing the deleterious effects of ROS production through the antioxidant system, under normal physiological conditions. When this regulatory system is dysregulated, excessive ROS will cause oxidative damage to a series of biomolecules such as DNA, proteins, and lipids, and even lead to functional decline (Patel, 2016). Therefore, oxidative stress may be associated with the development of a range of neurodegenerative diseases or mental disorders, such as Alzheimer’s disease, Parkinson’s disease, cerebrovascular disease, attention deficit hyperactivity disorder, schizophrenia, and autism spectrum disorders (Vavakova et al., 2015). Most studies have shown that oxidative stress is closely related to depression. The depressed patients have elevated levels of ROS, reactive nitrogen species (RNS) (Suzuki and Colasanti, 2001; Dhir and Kulkarni, 2011; Maes et al., 2011), and reduced activity of the antioxidant glutathione (GSH) in postmortem brain samples (Gawryluk et al., 2011). In addition, the expression of the enzymes involved in ROS production (xanthine oxidase and monoamine oxidase) was increased in depressed patients. For example, xanthine oxidase (XO), which catalyzes the oxidation of xanthine to produce superoxide and hydrogen peroxide, has been observed to elevate levels in the serum of depressed patients and in the thalamic region of post-mortem depressed patients (Michel et al., 2010). MAO levels, of which the by-products, such as hydrogen peroxide, leading to excessive production of ROS, are higher in depressed and postpartum depressed patients than in nondepressed subjects (Sacher et al., 2015), resulting in neuronal apoptosis and mitochondrial dysfunction. Therefore, anti-oxidative stress may be an effective strategy for the treatment of depression.
Numerous studies have demonstrated that PF could exert neuroprotective effects by inhibiting oxidative stress (Wang X. et al., 2021) (Figure 3). Superoxide dismutase (SOD) is an important antioxidant enzyme that catalyzes the breakdown of superoxide into oxygen and hydrogen peroxide, which is degraded by catalase under physiological conditions (Sies, 2015). In addition, the increase of ROS could lead to lipid peroxidation of the cell membrane to produce a large amount of malondialdehyde (MDA), a product of oxidative stress. Clinical studies have reported higher serum levels of MDA in patients with major depression compared with controls (Sies, 2015). After multiple intragastric administrations of PF, the plasma SOD level of the depression rats in the FST model was increased, and the plasma MDA level was decreased (Mu et al., 2020). PF significantly shortened the 5-min swimming immobility time of the rats in the FST model and increased the 5-min moving distance in the open-field trials, showing an antidepressant effect similar to fluoxetine. These studies, however, still exhibit certain deficiencies in their design. Firstly, the use of fluoxetine exclusively as a positive control in the antidepressant group restricts a comprehensive evaluation of the advantages and disadvantages of PF in comparison to other similar medications. Additionally, the absence of a detailed randomization methodology for the experimental subgroups may compromise the results. Furthermore, the experimental design featured a limited range of behavioral tests, encompassing only the forced swimming test and the open field test, and was conducted at only a few specific time points, which hindered the ability to observe the effects of PF across different time periods. In CORT- or glutamate-treated PC12 cells, PF increased the cell viability, the levels of GSH, the SOD and catalase activities, meanwhile, decreased intracellular reactive ROS and MDA levels in a dose-dependent manner (Mao et al., 2012a; Mao et al., 2011a; Mao et al., 2010). In addition, long-term PF (15 mg/kg and 30 mg/kg, i.p.) treatment of Aβ (1–42) (1 μg/μL) in rats restored the decreased activities of SOD and catalase, increased the level of MDA and the content of reduced GSH, which suggests that the PF can exert neuroprotective effects by alleviating oxidative stress (Lan et al., 2013; Zhong et al., 2009). Furthermore, Mao QQ et al. found that TGP (80 and 160 mg/kg) treatment in CUS mice dose-dependently reduced GSH depletion and MDA production, which further suggests that the antidepressant-like activity of TGP may be mediated by attenuating oxidative stress in the mouse brain (Mao et al., 2009b).
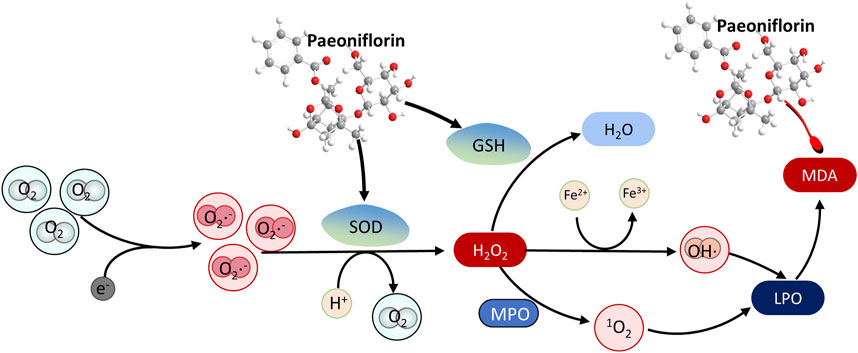
Figure 3. The antioxidant mechanism of paeoniflorin in depression by promoting the degradation of peroxides. The red arrow represents the inhibition of the protein, and the black arrow represents the promotion of the protein.
4.4 Apoptosis
Apoptosis, a form of programmed cell death, plays a critical role in tissue homeostasis, immune cell renewal, and neuronal development, which prevents the damage of surrounding tissues by timely eliminating senescent or damaged cells in the body (McKernan et al., 2009). Apoptosis is a tightly regulated and energy-dependent process, characterized by cytoplasmic shrinkage, chromatin condensation, nuclear pyknosis, ordered DNA fragmentation, cell rounding, and membrane blistering. Cells eventually form the membrane-bound vesicles called “apoptotic vesicles”, which could be phagocytosed by neighboring cells. Most studies have shown that neuronal apoptosis plays a crucial role in most CNS diseases. For instance, excessive apoptosis of a series of neurons can lead to neurodegeneration such as Alzheimer’s disease, Parkinson’s disease, and Huntington’s disease (Su et al., 1994; Jenner and Olanow, 1998; Kim et al., 1999). In addition, apoptosis may also be caused by stress (Lucassen et al., 2006). The study showed numbers of apoptotic and necrotic cells were observed in depressed mice with chronic mild stress (CMS) (Tian et al., 2019). Kosten et al. found that exposure to unpredictable stress decreased Bcl-2 mRNA in limbic structures of the brain and frontal cortex, and Bcl-xL mRNA in the hippocampus (Kosten et al., 2008). Meanwhile, the Administration of alanylcyclopropionate, reboxetine, and fluoxetine upregulated Bcl-2 mRNA levels and also increased Bcl-xL mRNA expression. Moreover, Bcl-2 expression in the hippocampus was increased after 14 days of administration of citalopram, promethazine, and amitriptyline (Kosten et al., 2008), which suggests that potential antidepressants could be developed through an anti-apoptotic mechanism.
Studies have shown that PF, the natural plant metabolite from the treasure trove of nature, has surprising neuroprotective effects, inhibiting excessive neuronal apoptosis and exhibiting good therapeutic effects on neurodegenerative diseases (AD, Parkinson’s disease, etc.) and mental disorders (anxiety, depression, etc.) (Zhong et al., 2009; Zhai et al., 2019; Li et al., 2014; Zhang et al., 2017; Cong et al., 2019; Zhang et al., 2015; Wang et al., 2013b; Wu et al., 2013; Sun et al., 2012; Zheng et al., 2016). The accumulation of glutamate (Glu) in the synaptic cleft could produce excitatory neurotoxicity that may contribute to depression, anxiety, post-traumatic stress disorder, schizophrenia, cognitive impairment, and other psychiatric disorders (Mehta et al., 2013). Wang X et al. suggest that PF significantly improves the Glu-induced decrease in SH-SY5Y cell viability in human neuroblastoma cells by affecting the expression of Bax/Bcl2, cleaved caspase-3, and cleaved caspase-9, suggesting that PF significantly reduces cell apoptosis and exerts neuroprotective effects through the Bax/Bcl2 pathway (Wang X. et al., 2021) (Figure 4). Moreover, PF exhibited a suppressive effect on apoptosis by attenuating mitochondrial membrane potential, promoting cytochrome c release, and counteracting the upregulation of caspase-3 and caspase-9 in the case of Aβ 25–35 induced PC12 cell damage (Li et al., 2014). Treatment with 200 μM CORT resulted in apoptosis of PC12 cells for 48 h. Interestingly, the treatment of TGP protected PC12 cells against CORT-induced toxicity in a dose-dependent manner, which was associated with the inhibition of caspase-3 activity involved in the mitochondrial pathway and the upregulation of the bcl-2/bax mRNA ratio. Meanwhile, TGP in mice also produced similar antidepressant effects (Mao et al., 2009c). Similarly, XiaoyaoSan, a characteristic Chinese traditional formula, reduced chronic stress-induced anxiety and depression behaviors in mice. PF, one of the main active plant metabolites in XYS, could protect primary neurons from CORT-induced neurotoxicity and reverse neuronal apoptosis caused by miR-200a-3p and miR-200b-3p overexpression (Yuan et al., 2022). It is important to note that XiaoyaoSan, as a Chinese medicine formula, has a complex composition that includes various plant metabolites beyond PF. This study focused solely on miR-200a/b-3p and PF, neglecting the potential roles and interrelationships of other miRNAs and metabolites within the formula. Furthermore, the use of a stereotactic microinjection method to investigate the role of miR-200a/b-3p in stress behavior may pose a risk of damaging rat brain tissue, which could subsequently interfere with the experimental results. The above studies suggest that PF may exert antidepressant effects through an anti-apoptotic molecular mechanism.
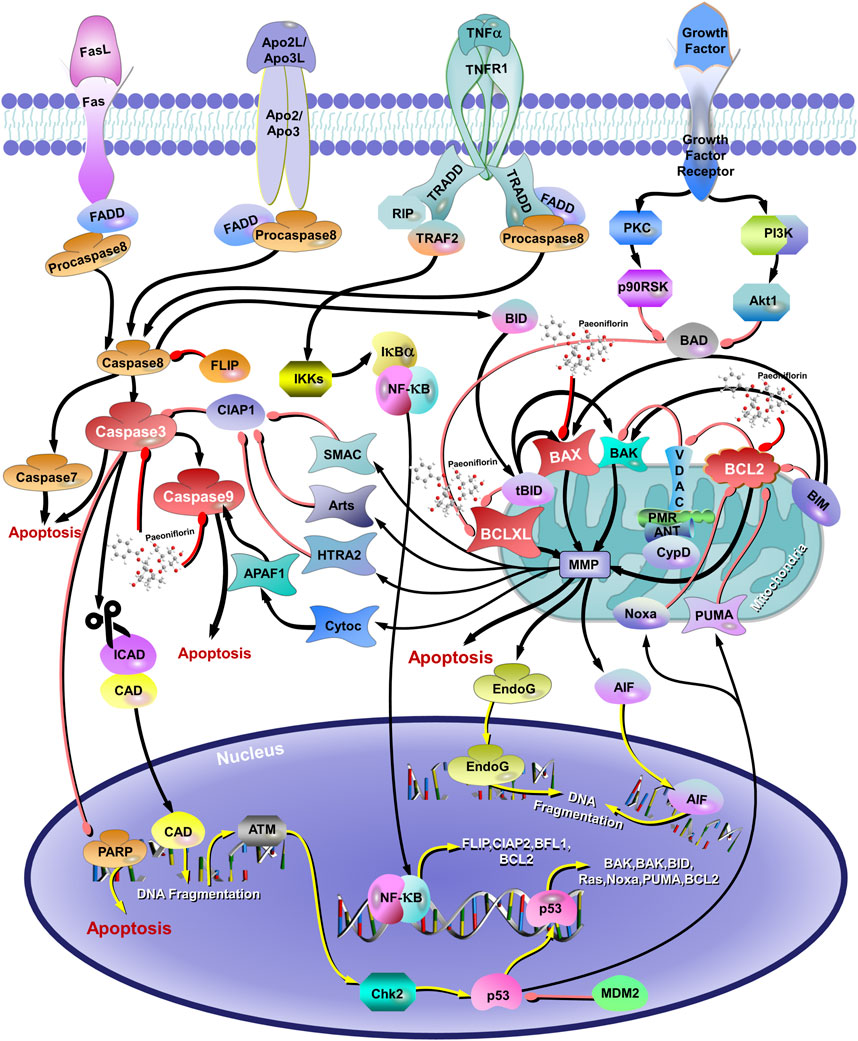
Figure 4. The Antiapoptotic mechanisms of paeoniflorin in depression. The red arrows represent the inhibition of proteins and the black arrows represent the promotion of proteins.
4.5 Calcium homeostasis and calcium signaling pathways
Calcium signaling regulates a range of neuronal activities by controlling the release of multiple neurotransmitters (Bezprozvanny and Mattson, 2008; Bergantin et al., 2013). Dysregulation of calcium signaling, such as excessive Ca2+ influx, may be associated with aging, Alzheimer’s disease, and major depression (Bergantin and Caricati-Neto, 2016). The abnormality of serum calcium level may be related to cognitive impairment in patients with depression (Grutzner et al., 2018). Normally, the intracellular Ca2+ concentration is maintained at a stable low level. Excessive Ca2+ influx may over-activate the Ca2+/CaM/CaMKII signaling pathway, impair neuronal activity, and induce depression when the homeostasis is changed. In addition, previous studies have found that the development of depression leads to neuronal death in the hippocampus and limbic brain. And the dysregulation of intracellular Ca2+ homeostasis seems to be closely related to this abnormal neuronal death (Duman, 2004; Bergantin, 2020). Therefore, maintaining normal Ca2+ homeostasis and safeguarding the proper regulation of calcium signaling pathways is one of the mechanisms to protect the nervous system.
Calbindin-D28K, one of the major calcium-binding proteins in the brain, could maintain intracellular Ca2+ homeostasis by binding excess Ca2+ to protect normal neuronal activation and function and inhibit neuronal apoptosis caused by intracellular Ca2+ overload (Carter et al., 2008; Meng et al., 2007). Experiments showed that PF could reverse the decrease of Calbindin-D28K mRNA level and the increase of Ca2+ concentration in PC12 cells induced by Glu, suggesting that the neuroprotective effect of PF is related to the inhibition of intracellular Ca2+ overload (Mao et al., 2010; Mao et al., 2011b). This result may be one of the pathways of antidepressant activity of PF in vivo. Meanwhile, PF treatment widely observed in other in vitro and animal trials could significantly block intracellular calcium influx caused by adverse stimuli such as methyl-4 phenylpyridine ion (MPP+) (Wang et al., 2013b), Aβ(1–42) oligomer (Zhong et al., 2009), and Glu (Wang X. et al., 2021; Wang et al., 2013c), which improve intracellular calcium homeostasis to play a neuroprotective role. In addition, previous studies have shown that the Ca2+/CaMKI/CREB signaling pathway plays an important role in intracellular signaling pathways involved in cell proliferation, cell survival, inflammation, and metabolism (Meng et al., 2007; Valera et al., 2008; Zhang et al., 2012). Calcium/calmodulin-dependent pathways may be overactivated to irreversible cellular damagewhen calcium is overloaded in neurons. PF reversed the significant reduction of p-CaMKII and p-CREB and regulated the expression of downstream proteins in the Middle Cerebral Artery Occlusion(MCAO) model and N-methyl-D-aspartic acid receptor(NMDA) induced excitatory toxicity model of primary hippocampal neurons, including Bax, Bcl2, Bad, and Caspase3, along the Ca2+/CaMKI/CREB signaling pathway (Zhang et al., 2017). Song et al. found that the effect of PF on the Ca2+/CaMKI/CREB signaling pathway may be achieved by regulating the current density of voltage-gated Ca2+ channel Cav1.2 (Song et al., 2017). Another study found that PF could significantly inhibit Glu-induced CaMKII over-expression and prevent intracellular calcium overload in PC12 cells (Wang et al., 2013c), thus playing an important role in the treatment and remission of affective disorders.
4.6 Neuroinflammation
Recently, clinical and preclinical evidence suggestthat neuroinflammation is an important factor in major depressive disorder. Studies using positron emission computed tomography (PET) imaging and 18 kDa translocator protein (TSPO) as a microglia biomarker have demonstrated neuroinflammation in several brain regions in depressed patients (Li et al., 2018; Setiawan et al., 2018). The research in animal models has also revealed the release of proinflammatory factors and activation of microglia in the animal brain, showing signs of anxiety and depression (Munshi et al., 2020; Wang Y. L. et al., 2018). The persistent sympathetic and parasympathetic under-activity in chronic stress situations and major depressive disorder (MDD) increase catecholamine levels and decreases acetylcholine levels, which increases levels of pro-inflammatory cytokines (TNF-α, IL-1β, IL-6, and IL-18, among others), which explains that depression-like symptoms can be induced directly by proinflammatory cytokines (Won et al., 2021). Moreover, proinflammatory cytokines could activate the kynurenine pathway, leading to the increase of neurotoxic metabolites, including 3-hydroxykynurenine, 3-hydroxy-anthranilic acid, and quinolinic acid, to cause brain damage (Kim and Won, 2017). Meanwhile, researchers have found that the activation of microglia by stress stimulation releases a large number of proinflammatory cytokines, which could destroy the neuroprotective mechanisms in the brain, impair neuroplasticity, and inhibit adult hippocampal neurogenesis, leading to the occurrence of depression-related symptoms (McKim et al., 2016).
The researchers have recently found that PF significantly inhibits the neuroinflammatory response by decreasing the over-activation of astrocytes (AST) and microglia, as well as the expression of pro-inflammatory cytokines such as IL-1β, IL-6 and TNF-α (Zhou et al., 2019; Liu H. et al., 2015; Nam et al., 2013) (Figure 5). PF prevented the upregulation of pro-inflammatory mediators (TNF-α, IL-1b, iNOS, COX2, and 5-LOX) in plasma and brain to the sustained activation of hippocampal AST and microglia caused by chronic cerebral insufficiency of cerebral perfusion or cerebral ischemia, suggesting that PF exerts delayed protective effects in ischemia-injured rats by inhibiting the peripheral and cerebral tissue inflammatory responses mediated in MAPKs/NF-kB (Guo et al., 2012; Liu J. et al., 2006). TLR4/NF-κB/NLRP3 signaling has been shown to regulate the inflammatory response of microglia, of which the activation promotes the over-expression and over-release of proinflammatory cytokines, leading to neuronal damage (Zhong et al., 2019). Cheng et al. found that PF can reduce the release of pro-inflammatory cytokines by regulating TLR4/NF-κB/NLRP3 signaling, thus reducing the damage of cytokines to neurons and reversing LPS-induced depression-like behavior in mice (Cheng et al., 2021). The study found obvious changes in inflammation in the amygdala for 4 weeks after subcutaneous injection of Interferon-α 15 × 106 IU/kg, as modeled by depression-inducing mice. Interestingly, the trial after 4 weeks of pretreatment with PF (20 mg/kg or 40 mg/kg) reversed the depression behavior of mice, and the abnormal level of inflammatory cytokines in serum, medial prefrontal cortex(mPFC), ventral hippocampus (vHi) and the amygdala, including IL - 6, IL - 1β, TNF-α, IL - 9, IL - 10, IL - 12 and monocyte chemotactic protein 1 (Li et al., 2017). However, the experiment only established two PF dose groups, specifically 20 mg/kg and 40 mg/kg, which limits the ability to ascertain the optimal effective dose of PF and its dose-effect relationship. Furthermore, while PF was observed to reverse the increase in microglia density and reduce the levels of inflammatory factors, the specific molecular pathways through which PF influences microglia in the context of neuroinflammation and depressive-like behaviors have not been thoroughly investigated. Furthermore, recent studies have found that PF reduces neuroinflammation by inhibiting Casp-11-dependent pyroptosis signaling induced by the overactivation of hippocampal microglia in reserpine-treated mice, representing a novel mechanism by which PF attenuates depression (Tian et al., 2021).
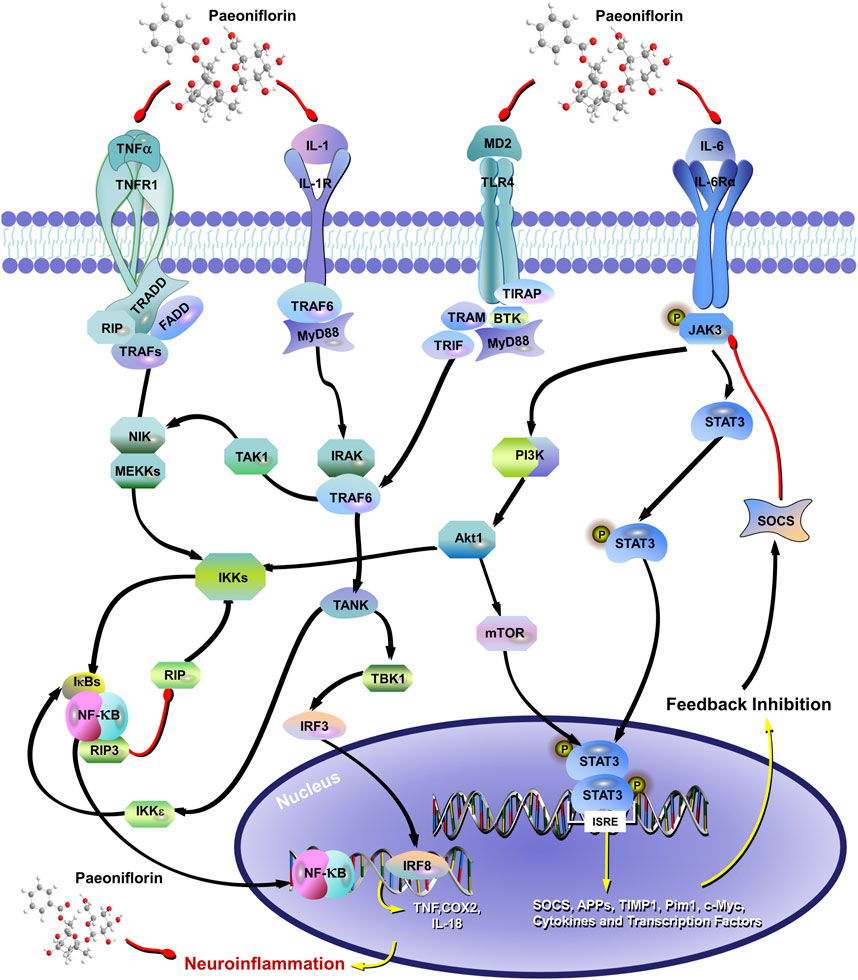
Figure 5. The mechanism of paeoniflorin inhibiting neuronal inflammation involves the inhibition of IL-1, IL-6, and TNF-α Expression, and affects the TLR4/NF-κB/NLRP3 signaling pathway. The red arrow represents the inhibition of the protein, and the black arrow represents the promotion of the protein.
4.7 Mitochondrial dysfunction
Mitochondria, the energy factory of eukaryotic cells, produce energy through the metabolism of lipids, steroids, and proteins, which play a key role in maintaining cellular stability by regulating Ca2+ levels, maintaining ROS levels, and regulating apoptosis (Nunnari and Suomalainen, 2012). Mitochondrial biosynthesis occurs more rapidly during neuronal development because neuronal differentiation requires an increase in the mitochondrial genome and mitochondrial proteins (Calingasan et al., 2008; Cuperfain et al., 2018; Kasahara and Kato, 2018). Therefore, mitochondrial dysfunction prevents cells from meeting their energy requirements and involves impairment of neuronal communication, cellular resilience, and hippocampal neurogenesis, which leads to mood and psychiatric disorders (Perkins et al., 1997; Nagashima et al., 2020). Most studies have shown that mitochondrial dysfunction in brain regions may be closely related to the development of depression. For example, brain mitochondrial dysfunction and ultrastructural damage have been reported in mouse models of depression (Gong et al., 2011). 54% of patients were also found to exhibit depressive symptoms in a study of the prevalence of psychiatric comorbidities in subjects with mitochondrial cytopathies (Feng et al., 2020), suggesting that targeting mechanisms of mitochondrial dysfunction may be a possibility for developing new treatments for depression.
Studies have shown that PF could exert neuroprotective effects by preventing mitochondrial dysfunction. PF could significantly improve the viability of SH-SY5Y cells and human neuroblastoma cells damaged by Glu or Ab25-35, and significantly inhibit the increase of mitochondrial membrane potential and calcium concentration to protect SH-SY5Y cells from Glu-induced excitatory neurotoxicity (Wang X. et al., 2021; Li et al., 2014). Daily administration of PF (10 mg/kg once a day) for 21 days was able to significantly ameliorate the cognitive dysfunction for streptozotocin (STZ)-induced in mice model, This research has demonstrated that PF may be related to the significant attenuation of the STZ-induced mitochondrial dysfunction, including a significant increase in cytochrome C oxidase activity and ATP synthesis to significantly restore the function in hippocampal area and cerebral cortex (Wang D. et al., 2018). Moreover, PF can regulate mitochondrial membrane potential and maintain mitochondrial membrane integrity to attenuate or restore PC12 cell injury induced by unfavorable stimuli such as Aβ25-35, MPP+, Glu, and CORT (Li et al., 2014; Wang et al., 2013b; Sun et al., 2012; Zheng et al., 2016; Mao et al., 2009c). Further studies revealed that this preventive-protective effect by PF might be achieved by inhibiting the MKK4-JNK signaling pathway (Cong et al., 2019).
4.8 Autophagy
Autophagy, a process of cellular waste removal and regeneration, transports cytosolic components such as proteins and organelles to lysosomes for degradation and recycling to provide a constitutive mechanism for the turnover and recruitment of cellular components (Xiao et al., 2018; Miranda et al., 2018). In the CNS, autophagy is involved in cell surveillance, neuroinflammation, and neuroplasticity. The investigators found that neurons are vulnerable to autophagy defects and depend heavily on the level of autophagy composition for survival (Hara et al., 2006; Nishiyama et al., 2007). Several preclinical and clinical studies have shown that dysfunction of cellular autophagy may be an important factor in the occurrence and progression of psychiatric disorders such as depression (Komatsu et al., 2006; Nouri et al., 2020; Zhao et al., 2017). For example, autophagy biomarkers, such as the LC3II/LC3I ratio and BECN1, are significantly reduced in animal models of depression. Notably, PF can exert neuronal protection by modulating the autophagy pathway (Jiang P. et al., 2017). PF can have a significant protective effect against acidosis or MPP + -induced injury in PC12 cells; further research suggested that PF improved acidosis-induced activation of acid-sensing ion channels (ASIC).
Meanwhile, PF weakened the autophagy inhibition induced by MPP+ and significantly upregulated autophagy and the ubiquitin-proteasome pathway, which prevents α-synuclein accumulation(α-SYN) of synaptic nucleoproteins to reduce cell damage (Sun et al., 2011; Cai et al., 2015; Cao et al., 2010). α-SYN is a characteristic marker of all types of PD, which can be degraded by the ubiquitin-proteasome system (UPS) and autophagy-lysosomal pathway (ALP) (Kaushik and Cuervo, 2012). In another study, systemic administration of PF in a 6-hydroxydopamine (6-OHDA) -induced PD rat model showed that PF enhanced the autophagic degradation of α-SYN to protect DA neurons from 6-OHDA-induced neurotoxicity. Further Western blot results showed that PF could significantly reduce the level of ASIC1a, suggesting that PF may play a neuroprotective role by inhibiting the activation of ASIC, especially ASIC1a (Gu et al., 2016).
4.9 Brain-derived neurotrophic factor
Brain-derived neurotrophic factor (BDNF), a key mediator of activity-dependent neuronal plasticity in the brain (Park and Poo, 2013), has a major impact on neuronal morphology, survival, and differentiation, increasing synapse sprouting and synaptic stability, and facilitating long-duration strengthening (Zagrebelsky and Korte, 2014). BDNF synthesis occurs predominantly in regions involved in emotional and cognitive functions, such as the hippocampus and frontal regions (Phillips, 2017). BDNF could bind and activate tropomyosin receptor kinase B (TrkB) to regulate the different cellular processes for the development and maintenance of normal brain function. In addition, several lines of evidence suggest that BDNF/TrkB signaling is involved with adult neurogenesis in the hippocampus and has differential effects on the dentate gyrus (DG) and subventricular zone (SVZ) (Colucci-D'Amato et al., 2020). Numerous studies have confirmed that BDNF is one of the important biomarkers of depression. The low levels of BDNF are associated with reduced synaptic plasticity and neuronal atrophy, which is consistent with the neurogenic hypothesis of depression. Several autopsy studies have demonstrated decreased BDNF expression in the hippocampus and prefrontal cortex of depressed patients (Dwivedi et al., 2003). In addition, significantly elevated BDNF levels were detected in the serum of patients treated with antidepressants. This suggests that BDNF is not only a marker of disease but also a potential predictor of antidepressant efficacy. Taking BDNF signaling as a breakthrough point would be a potential direction for the development of antidepressants.
Fortunately, a large number of studies have found that the natural plant metabolite PF could upregulate the expression of plasma BDNF and BDNF mRNA in the hippocampus and frontal cortex (Mu et al., 2020; Mao et al., 2009a; Qingwei, 2018; Mao et al., 2012b), as well as postsynaptic density protein 95 (PSD 95) in model animals with mood disorders (Liu S. C. et al., 2019), thereby preventing CUMS-induced synaptic plasticity defects and providing neuroprotection in animal models of mood disorders. Previous studies suggested that BDNF combined with p-CREB is the main regulator of neurogenesis and emotion regulation, which may be closely related to neural plasticity (Sasaki and Yoshizaki, 2015; Orlovsky et al., 2014). Hu et al. found that PF therapy plays a positive role in neural and emotional regulation by reversing the decreased expression of BDNF and p-CREB in the hippocampus caused by the MCAO and CUMS model (Hu et al., 2019). In addition, PF also significantly increased the levels of other neurotrophic factors, such as NGF protein and mRNA, in the frontal cortex of CUMS rats. Further study found that the treatment of PF not only significantly enhanced the protein expression and gene transcription of BDNF in CUMS rats, but also activated the expression of TrkB, a high-affinity receptor for BDNF, which promoted the proliferation of neural stem cells, differentiation into AST, and neurogenesis in the DG of the hippocampus in stressed rats. This result suggests that PF may play the role of an antidepressant through the BDNF/TrkB signaling pathway (Chen et al., 2019). Meanwhile, Chen et al. used PF to treat the withdrawal hormone simulated pregnancy (HSP) in a Postpartum depression (PPD) rat model and found that PF improved PPD symptoms by promoting the activation of the transporter TspO and BDNF/mTOR pathways in PPD rats, proving that PF may be an effective anti-PPD and anti-depression drug (Chen J. et al., 2022).
5 Safety overview
Currently, toxicological studies on PF are still very limited. Most of the studies have investigated the toxicity of PF at the cellular level. In general, PF is a low-toxicity natural plant metabolite. The safety results of PF still vary in different cell types. For mouse thymocytes, PF did not show cytotoxicity at concentrations of 0–1000 μg/mL (Li et al., 2007). Two studies reported that PF showed a favorable safety profile against U937 cells, the human myelomonocytic cell line, which did not exhibit cytotoxicity even at 640 μg/mL (Salunga et al., 2007; Jin et al., 2011). However, our study found that PF at 400 μg/mL exhibited some growth inhibition against BEAS-2B cells, the human normal lung epithelial cells (Hou et al., 2024). Meanwhile, PF also exhibited some toxicity with HaCat cells in a concentration-dependent manner (Wang D. et al., 2022). The safety studies of PF with animal models are still scarce in vivo. PF was found to inhibit the hatching rate of zebrafish at high concentrations (100 and 200 μg/mL) (Rao et al., 2024). Some clinical trials of formulas containing PF have reported the safety evaluation. The pharmacokinetic trial involving Chinese volunteers showed that no adverse events (AEs) and serious adverse events (SAEs) were observed in the investigators who received a single intravenous infusion of Huoxue-Tongluo lyophilized powder for injection (HTLPI) containing PF (Li et al., 2016). Interestingly, one male subject experienced elevated transaminase levels with multiple dose infusions that recovered after 2 weeks (Li et al., 2016). Another clinical trial conducted on Chinese healthy volunteers reported that injections containing PF can cause AEs such as dizziness and diarrhea, but no SAEs occurred (Chen et al., 2013). Indeed, both clinical trials used formula preparations containing PF, and the subjects were all from China, which may interfere with the trial results. These clinical results can only provide a reference for the safety evaluation of PF.
In conclusion, although the above studies reported some safety results of PF, most of them were not systematically investigated. Meanwhile, the long-term evaluative safety assessment of PF should be emphasized by researchers. Particularly, how to select reference reagents and placebos is also a challenge for the clinical design of natural plant metabolites, which is necessary for the pharmacological research of PF.
6 Traditional Chinese medicine formula
In the application of traditional Chinese medicine, certain formulas containing PF have demonstrated notable efficacy in treating depression, such as Xiaoyao San, Chaihu Shugan San, and Sini San. These formulas, which include PF as one of their active ingredients, exert their effects through multiple targets and pathways, involving the regulation of neural pathways, neurotransmitters, synaptic plasticity, the neuroendocrine system, and the immune system. Although PF may not be the decisive factor in the prescription, TCM formulas containing PF still hold significant value in the treatment of depression. TCM formulas are composed of multiple botanical drugs that are synergistically combined, with each botanical drug containing various plant metabolites that interact to produce therapeutic effects. The following section will summarize the applications and molecular mechanisms of several classic TCM formulas containing PF in the treatment of depression.
Xiaoyaosan (XYS) is a classic TCM formula originating from the “Taiping Hui Min He Ji Ju Fang”, which is composed of Bupleurum chinense DC. [Apiaceae, Bupleurum chinense root], Angelica sinensis (Oliv.) Diels [Apiaceae, A. sinensis radix et rhizoma], Paeonia lactiflora Pall. [Paeoniaceae, Paeonia lactiflora root], Atractylodes macrocephala Koidz. [Asteraceae, Atractylodes macrocephala rhizoma et root], Wolfiporia cocos (F.A. Wolf) Ryvarden & Gilb. [Polyporaceae, W. cocos sclerotium], Glycyrrhiza uralensis Fisch. ex DC. [Fabaceae, Glycyrrhiza uralensis radix et rhizoma], Mentha canadensis L. [Lamiaceae, M. canadensis leaves], and Zingiber officinale Roscoe [Zingiberaceae, Z. officinale rhizoma et root], with a recommended ratio of 6:6:6:6:6:3:2:2. Its active metabolites include PF, bupleurum saponins A/C/D, ferulic acid, ligustilide, atractylodes lactone I/II/III, paeonia lactiflora glycoside, glycyrrhiza glycoside, glycyrrhetic acid, and poria acid (Lu et al., 2018). This formula has a history of over 2,000 years in China and is widely employed to treat various types of depression due to its liver-soothing, depression-relieving, spleen-nourishing, and blood-nourishing effects (Feng et al., 2016; Li et al., 2022). Recent studies have demonstrated that XYS exerts its antidepressant effects through multi-target regulation, involving the nervous, endocrine, and immune systems (Ma et al., 2019) (Tables 1, 2). For example, XYS can reduce the expression of miR-200a/b-3p in the prefrontal cortex induced by chronic stress, regulating the miR-200a/b-3p/NR3c1 signaling pathway. Further studies have found that PF, its main metabolite, inhibits miR-200a/b-3p-mediated neuronal apoptosis (Yuan et al., 2022). Additionally, XYS can reverse CORT elevation in the HPA axis of CUMS model rats, upregulate glial fibrillary acidic protein (GFAP) expression in the hippocampus, influence astrocyte (AST) activity, downregulate the NMDA receptor NR2B subunit level in the hippocampus, and improve depressive-like behavior (Liu T. et al., 2020). Studies on APJ receptors indicate that XYS can upregulate hypothalamic apelin levels and downregulate APJ levels, resulting in antidepressant behavioral improvements comparable to those of fluoxetine (Yan et al., 2018). Additionally, XYS can also increase microtubule-associated protein 2 (MAP2) expression in the CA1 region of the hippocampus in CUMS rats, enhance NR2B and PI3K expression to regulate the NR2B and PI3K/Akt signaling pathways, and alleviate Glu-induced neuronal damage (Zhou et al., 2021). Interestingly, XYS may exert its antidepressant effects by regulating gut microbiota composition and restoring abnormal levels of cecal metabolites (Lv et al., 2021).
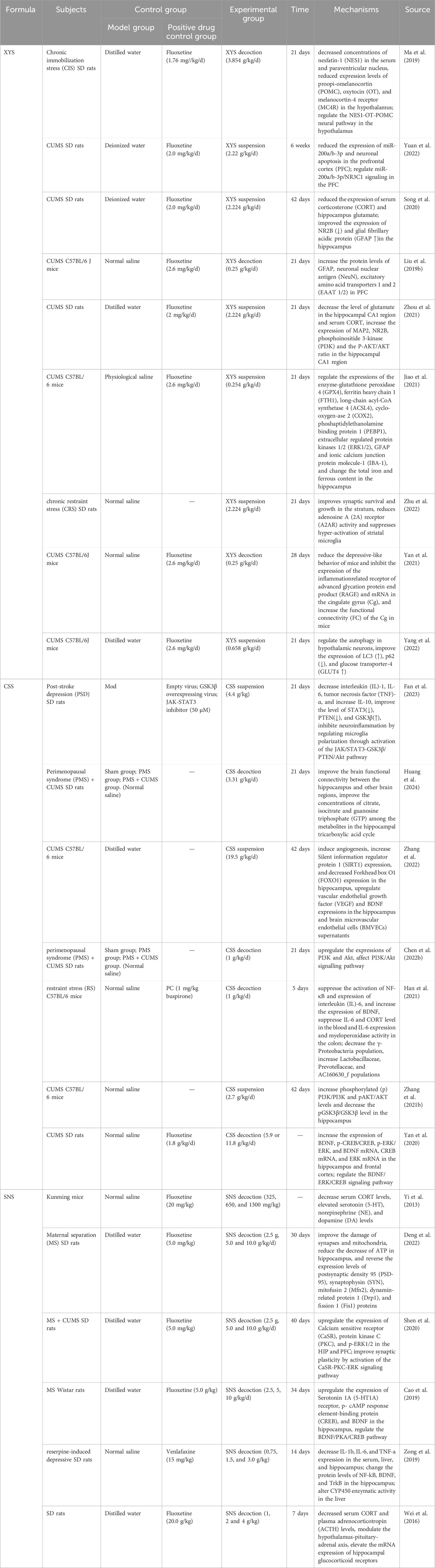
Table 1. Antidepressant Mechanisms of Formulas in vivo. (XYS: Xiaoyaosan; CSS: Chaihu-Shugan-San; SNS: Si-Ni-San).
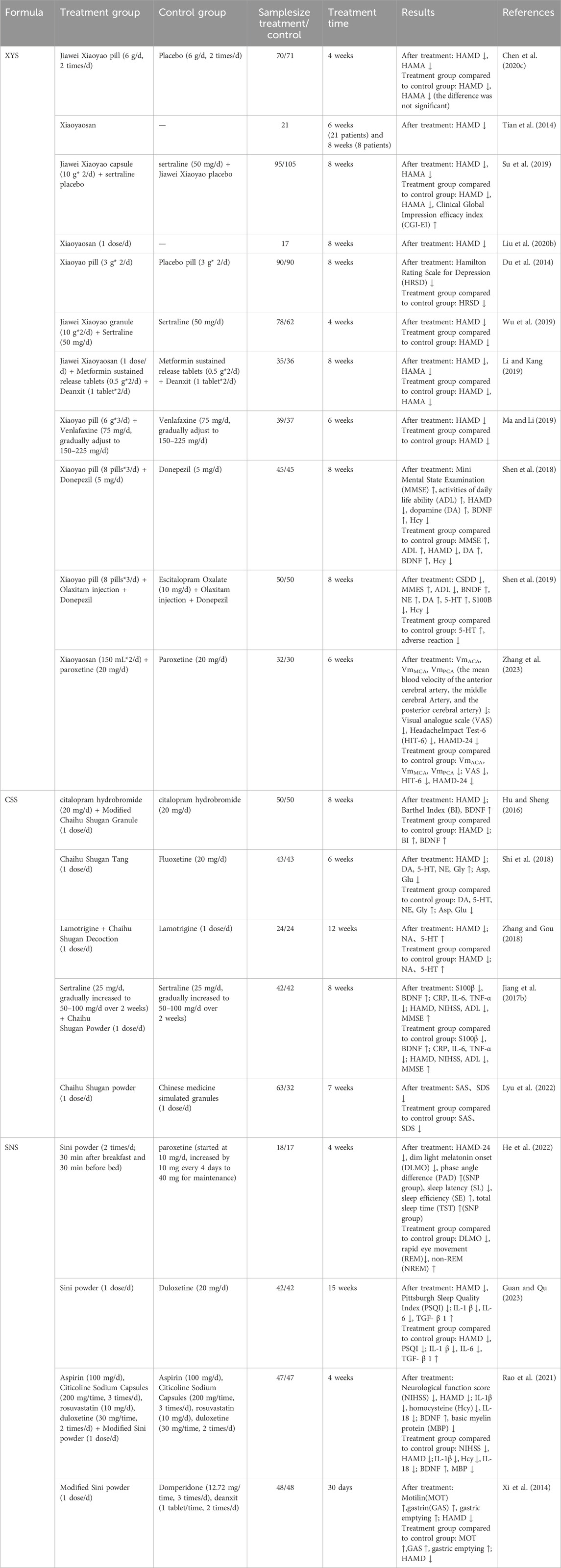
Table 2. Clinical researches of formulas. (XYS: Xiaoyaosan; CSS: Chaihu-Shugan-San; SNS: Si-Ni-San).
Chaihu Shugan San (CSS), first documented in the Ming Dynasty’s Jingyue Quanshu, is a classical formula in traditional Chinese medicine. As per traditional Chinese medicine theory, it is utilized for the treatment of liver qi stagnation syndrome. This formula synergistically combines Paeonia lactiflora Pall. [Paeoniaceae, Paeonia lactiflora root] with Bupleurum chinense DC. [Apiaceae, Bupleurum chinense root], complemented by Cyperus rotundus L. [Cyperaceae, C. rotundus rhizomaa et root], Conioselinum anthriscoides ‘Chuanxiong’ [Apiaceae, C. anthriscoides ‘Chuanxiong’ rhizoma et root], Citrus reticulata Blanco [Rutaceae, C. reticulata pericarp], Citrus × aurantium L. [Rutaceae, Citrus aurantium fruits], and Glycyrrhiza uralensis Fisch. ex DC. [Fabaceae, Glycyrrhiza uralensis radix et rhizoma] (Gao et al., 2022). Modern research has substantiated its significant efficacy in alleviating depression (Wang et al., 2012; Qiu et al., 2014a; Qiu et al., 2014b) (Table 2), although the underlying mechanisms remain elusive. In recent years, extensive studies have explored the antidepressant mechanisms of CSS (Table 1). Bioinformatics analyses indicate that CSS addresses depression through multiple targets and pathways, including the regulation of 110 differentially expressed proteins (DEPs) in the hippocampus and the modulation of various neurotransmitter transport and circulation (Zhu et al., 2021). Furthermore, CSS has been shown to reverse hyperactivity of the HPA axis, enhance cerebral blood flow perfusion, and mitigate depressive symptoms. Research indicates that it significantly increases regional cerebral blood flow (rCBF) in patients suffering from major depressive disorder (Vangu et al., 2003), and single-photon emission computed tomography (SPECT) has validated its efficacy in improving cerebral perfusion deficits, correlating with clinical symptom relief (Qiu et al., 2014a). Numerous studies have demonstrated that CSS enhances monoamine neurotransmitter levels, regulates BDNF, and modulates the BDNF-TrkB-ERK/Akt signaling pathway, thereby exerting antidepressant effects (Qiu et al., 2014b; Li et al., 2009; Chen X. Q. et al., 2018; Fan et al., 2023). Additionally, CSS shows promise in treating post-stroke depression by downregulating pro-inflammatory factors, including interleukin-1 (IL-1), interleukin-6 (IL-6), and tumor necrosis factor-α (TNF-α), as well as associated proteins such as STAT3 and PTEN, while upregulating glycogen synthase kinase 3β (GSK3β). It also regulates microglial polarization and mitigates neuroinflammation through the activation of the JAK/STAT3-GSK3β/PTEN/Akt pathway (Fan et al., 2023). Recent research indicates that CSS downregulates microRNA-124 (miR-124), upregulates target genes such as mitogen-activated protein kinase 14 (MAPK14) and glutamate ionotropic receptor AMPA subunit 3 (Gria3), promotes synaptic reconstruction in the hippocampus of CUMS rats, and ameliorates depressive behavior (Liu Q. et al., 2018). In conclusion, CSS, with Paeonia lactiflora Pall. [Paeoniaceae, Paeonia lactiflora root] as its primary metabolite, exerts antidepressant effects through multiple targets and pathways.
SiniSan(SNS), originating from the Shanghan Lun of the Han Dynasty, consists of equal parts of white Paeonia lactiflora Pall. [Paeoniaceae, Paeonia lactiflora root], Bupleurum chinense DC. [Apiaceae, Bupleurum chinense root], Citrus × aurantium L. [Rutaceae, C. aurantium fruits], and Glycyrrhiza uralensis Fisch. ex DC. [Fabaceae, Glycyrrhiza uralensis radix et rhizoma], making it a standard formula for treating depression caused by liver qi stagnation (Shen et al., 2023). Recent clinical trials have confirmed its significant efficacy in addressing various types of depression (Zhang Qi et al., 2022) (Table 2). Animal studies indicate that SNS significantly elevates the levels of serotonin (5-HT), norepinephrine (NE), and dopamine (DA) in the brains of depressed mice, while concurrently reducing serum cortisol (CORT) levels (Yi et al., 2013) (Table 1). Prior research has established a link between mitochondrial diseases, depression, and diminished hippocampal synaptic plasticity and neuronal atrophy, all of which are associated with abnormal mitochondrial function and reduced ATP levels in the hippocampus and prefrontal cortex under stress conditions (Aleksandrova et al., 2019; Malberg et al., 2021; Tartt et al., 2022). The SNS has been shown to enhance synaptic plasticity and alleviate mitochondrial damage in maternally separated (MS) female rats under stress, prevent ATP depletion in the hippocampus, increase the postsynaptic density (PSD) at glutamatergic neurotransmitter transmission sites—specifically at asymmetric synapses—and inhibit the overexpression of key proteins involved in mitochondrial fission and fusion (Deng et al., 2022). SNS can activate the CaSR-PKC-ERK signaling pathway, upregulate the expression of serotonin 1α receptors (5-HT1α), phosphorylated cAMP response element-binding protein (p-CREB), and BDNF in the hippocampus, thereby alleviating depressive and anxious behaviors in MS rats (Shen et al., 2020; Cao et al., 2019). Furthermore, SNS enhances the activity of cytochrome P450-related enzymes (CYP1A2, CYP2D1, CYP2E1, CYP3A2), decreases the expression of inflammatory cytokines (IL-1β, IL-6, TNF-α) in reserpine-induced depressive rats, and increases the protein levels of nuclear factor-κB (NF-κB), BDNF, and tyrosine kinase B (TrkB) in the hippocampus. Its mechanism of action in treating depression may be associated with the modulation of CYP450 enzyme activity in the liver (Zong et al., 2019). In models of CORT-induced neuronal damage, SNS regulates the expression of autophagy-related proteins, activates the phosphoinositide 3-kinase/protein kinase B/mammalian target of rapamycin (PI3K/AKT/mTOR) pathway, and prevents excessive autophagy (Zhang M. et al., 2021). Additionally, its extract can modulate the HPA axis, diminish acute stress-induced elevations in serum CORT and plasma corticotropin-releasing hormone (CRH), and reverse the reduction in hippocampal glucocorticoid receptor (GR) mRNA levels (Wei et al., 2016).
Existing evidence indicates that PF, as the primary active metabolite, exerts certain antidepressant effects in formulas such as XYS, CSS, and SNS; however, it accounts for only a portion of the overall effect. The antidepressant efficacy of these traditional Chinese medicine formulas arises from the integrated synergistic effects of multiple plant metabolites, pathways, and targets. While PF could serve as one of the quality markers, it should not be simplistically equated with the evaluation of the entire formulation. Current studies, however, are unable to distinguish between the isolated effects of PF and the overall effects of the formula. We propose that future research adopt a progressive strategy of “formula - combination - metabolite”. Moreover, the research could utilize an integrated approach of network pharmacology, metabolomics, and microbiomics. According to PF as a reference, researchers can systematically compare the differences in targets and molecular mechanisms among single drugs, the combination of PF and core protein, and the entire formula. This approach may clarify the precise target and contribution weight of PF within the formula and further explore the mechanisms of other key active metabolites and their synergistic effects within the formula.
7 Research limitations and future prospects
In recent years, greater emphasis has been placed on mental health and wellbeing. However, depression has become more common, and the incidence has gradually increased. Depression exhibits characteristics that are not confined to a specific age group and may be in the elderly, youth, or even children. Depression, characterized by symptoms such as negative mood, persistent low mood, self-depreciation, and loss of interest in life, seriously affects the life and health of individuals. Although a variety of antidepressant drugs have been developed, including TCAs, MAOIs, and selective 5-HT reuptake inhibitors, they still have defects such as constipation, urinary retention, cardiovascular risk, gastrointestinal symptoms, emotional agitation, or tolerance (Marwaha et al., 2023; Peng et al., 2015). Fortunately, some time-honored formulas documented in traditional Chinese medicine, such as the XYS, have demonstrated promising antidepressant efficacy for millennia (Fathinezhad et al., 2019; Liu L. et al., 2015). Modern studies have found that PF, the main plant metabolite contained in these formulas, may be one of the important factors for good antidepressant and neuroprotective effects (Wu et al., 2024). PF is a bioactive metabolite with the characteristic of being both medicinal and edible, widely present in plant-based foods. It not only possesses nutritional functions but also exerts positive effects on human health through various bioactive mechanisms. In animal experiments, PF has shown good preventive and therapeutic effects on the menopausal depression model, CUMS model, FST, and PPD model. Moreover, a wide range of researchers have focused on exploring the intrinsic molecular mechanisms by which PF exerts antidepressant effects. PF may exert antidepressant activity through a variety of mechanisms, such as preventing the overactivity of the HPA axis, regulating the monoaminergic nervous system, maintaining normal calcium homeostasis and calcium signaling pathway, inhibiting oxidative stress and apoptosis, and regulating the expression of neurotrophic factors in the brain (Figure 6). However, some urgent problems of PF are still to be solved (Guo et al., 2022). Although PF, a water-soluble plant metabolite, could be conveniently administered to patients in the clinic. These qualities, such as the chemical instability, the necessity of storing at low temperatures, avoiding alkaline environments, and the low oral bioavailability, have led to some limitations in application (Yu et al., 2019). The explorations study has found that PF may help patients recover physical and mental health through multi-pathway and multi-target ways. Unfortunately, the mechanism has been extensively studied but not deeply, leading to more potential pharmacological effects and targets that may not be found yet (Hong et al., 2022; Wang X. L. et al., 2021). Particularly, the differences in analytical methods and composition still have a certain impact on the results of most studies with the medicine formula containing PF, which suggests that researchers should consider the importance of developing the standardization of research methods. Besides, the evaluation of side effects and toxicity of PF on target organs is rarely reported. Therefore, long-term systematic drug safety trials are encouraged to achieve the optimal level of safety. Most importantly, clinical trials and data on PF for depression are still lacking. Future studies may require well-designed and adequate clinical trials to explore the deeper intrinsic mechanisms of PF on the one hand and to look forward to designing more specific treatment regimens to achieve optimal efficacy on the other. Indeed, clinical trials with a single natural plant metabolite should also consider the selection of reference reagents and the development of validated indicator judgments. We suggest that in antidepressant clinical trials of PF, study designers should also consider the setting of the Run-in period while focusing on blinding and randomization, which could avoid confounding factors that may interfere with the results of the trial and thus improve the reliability of the study.
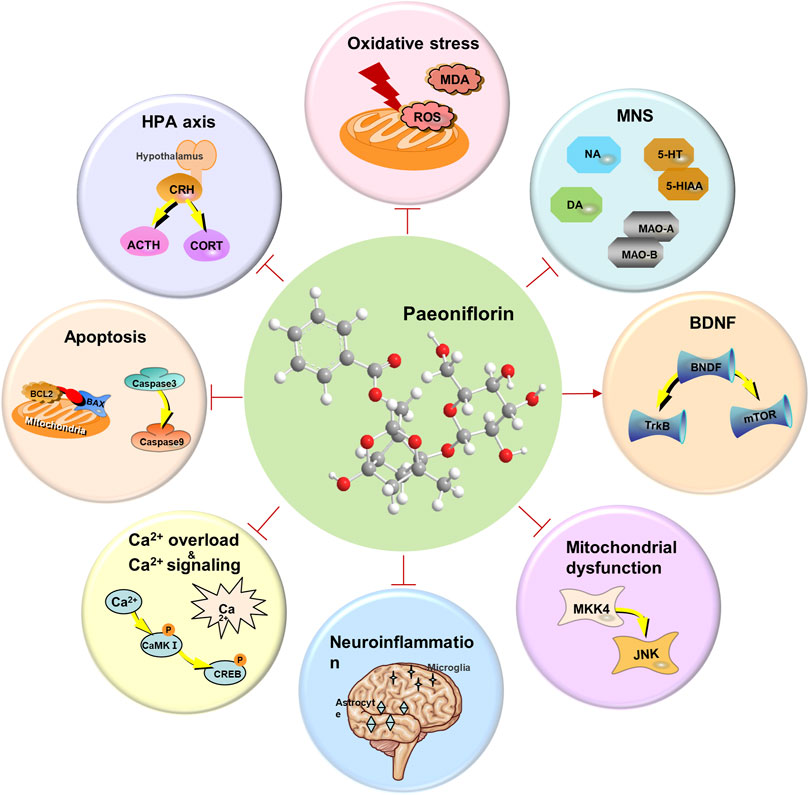
Figure 6. The mechanism of paeoniflorin exerting antidepressant activity.
8 Conclusion
Overall, PF, as a bioactive plant metabolite with dual purposes in medicine and food, not only provides new insights for the treatment of depression but also offers strong support for the health benefits of bioactive metabolites in plant-based foods. In this paper, we present the first narrative review of the mechanisms of PF in antidepressant therapy and the antidepressant applications of traditional compounding. Compared to a systematic review, this study is a better reference for researchers who could contribute to the study of the molecular pathways of PF in depression, as well as the formulation. In particular, these constructive comments on the methodology of PF in antidepressant clinical trials provide a basis for the systematic evaluation of its safety and efficacy in the clinic.
Author contributions
YH: Writing – original draft, Visualization, Conceptualization, Writing – review and editing. HL: Writing – review and editing, Writing – original draft. LZ: Funding acquisition, Writing – review and editing, Writing – original draft. TQ: Writing – original draft, Software, Writing – review and editing, Validation. XF: Validation, Writing – original draft, Writing – review and editing, Software. YL: Writing – original draft, Writing – review and editing, Funding acquisition. YB: Writing – review and editing, Writing – original draft, Validation, Investigation. YW: Conceptualization, Writing – review and editing, Writing – original draft, Visualization.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This study was supported by the Science and Technology Fund of Deyang (No. 2022SZ043, 2024SZY035, 2024SZY034); Special Project for Science and Technology Strategic Cooperation between City and School in Nanchong City, Sichuan Province (22SXQT0166).
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aleksandrova, L. R., Wang, Y. T., and Phillips, A. G. (2019). Evaluation of the wistar-Kyoto rat model of depression and the role of synaptic plasticity in depression and antidepressant response. Neurosci. Biobehav Rev. 105, 1–23. doi:10.1016/j.neubiorev.2019.07.007
Bai, X., Yu, C., Yang, L., Luo, Y., Zhi, D., Wang, G., et al. (2020). Anti-psoriatic properties of paeoniflorin: suppression of the NF-kappaB pathway and Keratin 17. Eur. J. Dermatol 30 (3), 243–250. doi:10.1684/ejd.2020.3770
Bardeleben, U., and Holsboer, F. (1989). Cortisol response to a combined dexamethasone-human corticotrophin-releasing hormone challenge in patients with depression. J. Neuroendocrinol. 1 (6), 485–488. doi:10.1111/j.1365-2826.1989.tb00150.x
Bergantin, L. B. (2020). A Hypothesis for the relationship between depression and cancer: role of Ca2+/cAMP signalling. Anticancer Agents Med. Chem. 20 (7), 777–782. doi:10.2174/1871520620666200220113817
Bergantin, L. B., and Caricati-Neto, A. (2016). Challenges for the pharmacological treatment of neurological and psychiatric disorders: implications of the Ca(2+)/cAMP intracellular signalling interaction. Eur. J. Pharmacol. 788, 255–260. doi:10.1016/j.ejphar.2016.06.034
Bergantin, L. B., Souza, C. F., Ferreira, R. M., Smaili, S. S., Jurkiewicz, N. H., Caricati-Neto, A., et al. (2013). Novel model for “calcium paradox” in sympathetic transmission of smooth muscles: role of cyclic AMP pathway. Cell Calcium 54 (3), 202–212. doi:10.1016/j.ceca.2013.06.004
Berlow, Y. A., Zandvakili, A., Carpenter, L. L., and Philip, N. S. (2019). Transcranial direct current stimulation for unipolar depression and risk of treatment emergent mania: an updated meta-analysis. Brain Stimul. 12 (4), 1066–1068. doi:10.1016/j.brs.2019.03.025
Bezprozvanny, I., and Mattson, M. P. (2008). Neuronal calcium mishandling and the pathogenesis of Alzheimer's disease. Trends Neurosci. 31 (9), 454–463. doi:10.1016/j.tins.2008.06.005
Bourin, M., Poncelet, M., Chermat, R., and Simon, P. (1983). The value of the reserpine test in psychopharmacology. Arzneimittelforschung 33 (8), 1173–1176.
Buda, O., Hostiuc, S., Popa-Velea, O., and Boroghina, S. (2022). Altered states, alkaloids, and catatonia: monoaminoxidase inhibitors and their role in the history of psychopharmacology. Front. Pharmacol. 13, 1053534. doi:10.3389/fphar.2022.1053534
Cai, Z., Zhang, X., Zhang, Y., Li, X., Xu, J., and Li, X. (2015). The impact of paeoniflorin on alpha-Synuclein degradation pathway. Evid. Based Complement. Altern. Med. 2015, 182495. doi:10.1155/2015/182495
Calingasan, N. Y., Ho, D. J., Wille, E. J., Campagna, M. V., Ruan, J., Dumont, M., et al. (2008). Influence of mitochondrial enzyme deficiency on adult neurogenesis in mouse models of neurodegenerative diseases. Neuroscience 153 (4), 986–996. doi:10.1016/j.neuroscience.2008.02.071
Cao, B. Y., Yang, Y. P., Luo, W. F., Mao, C. J., Han, R., Sun, X., et al. (2010). Paeoniflorin, a potent natural compound, protects PC12 cells from MPP+ and acidic damage via autophagic pathway. J. Ethnopharmacol. 131 (1), 122–129. doi:10.1016/j.jep.2010.06.009
Cao, K., Shen, C., Yuan, Y., Bai, S., Yang, L., Guo, L., et al. (2019). SiNiSan ameliorates the depression-like behavior of rats that experienced maternal separation through 5-HT1A Receptor/CREB/BDNF pathway. Front. Psychiatry 10, 160. doi:10.3389/fpsyt.2019.00160
Carter, D. S., Harrison, A. J., Falenski, K. W., Blair, R. E., and DeLorenzo, R. J. (2008). Long-term decrease in calbindin-D28K expression in the hippocampus of epileptic rats following pilocarpine-induced status epilepticus. Epilepsy Res. 79 (2-3), 213–223. doi:10.1016/j.eplepsyres.2008.02.006
Chen, T., Guo, Z. P., Jiao, X. Y., Zhang, Y. H., Li, J. Y., and Liu, H. J. (2011). Protective effects of peoniflorin against hydrogen peroxide-induced oxidative stress in human umbilical vein endothelial cells. Can. J. Physiol. Pharmacol. 89 (6), 445–453. doi:10.1139/y11-034
Chen, L., Qi, H., Jiang, D., Wang, R., Chen, A., Yan, Z., et al. (2013). The new use of an ancient remedy: a double-blinded randomized study on the treatment of rheumatoid arthritis. Am. J. Chin. Med. 41 (2), 263–280. doi:10.1142/s0192415x13500195
Chen, H., Dong, Y., He, X., Li, J., and Wang, J. (2018a). Paeoniflorin improves cardiac function and decreases adverse postinfarction left ventricular remodeling in a rat model of acute myocardial infarction. Drug Des. Devel Ther. 12, 823–836. doi:10.2147/DDDT.S163405
Chen, X. Q., Li, C. F., Chen, S. J., Liang, W. N., Wang, M., Wang, S. S., et al. (2018b). The antidepressant-like effects of Chaihu Shugan San: dependent on the hippocampal BDNF-TrkB-ERK/Akt signaling activation in perimenopausal depression-like rats. Biomed. Pharmacother. 105, 45–52. doi:10.1016/j.biopha.2018.04.035
Chen, L. B., Qiu, F. M., Zhong, X. M., Hong, C., and Huang, Z. (2019). Promoting neurogenesis in hippocampal dentate gyrus of chronic unpredictable stress-induced depressive-like rats with paeoniflorin. J. Integr. Neurosci. 18 (1), 43–49. doi:10.31083/j.jin.2019.01.116
Chen, L., Hudaib, A. R., Hoy, K. E., and Fitzgerald, P. B. (2020a). Efficacy, efficiency and safety of high-frequency repetitive transcranial magnetic stimulation applied more than once a day in depression: a systematic review. J. Affect Disord. 277, 986–996. doi:10.1016/j.jad.2020.09.035
Chen, C., Yin, Q., Tian, J., Gao, X., Qin, X., Du, G., et al. (2020b). Studies on the potential link between antidepressant effect of Xiaoyao san and its pharmacological activity of hepatoprotection based on multi-platform metabolomics. J. Ethnopharmacol. 249, 112432. doi:10.1016/j.jep.2019.112432
Chen, G., Feng, P., Wang, S., Ding, X., Xiong, J., Wu, J., et al. (2020c). An herbal formulation of jiawei xiaoyao for the treatment of functional dyspepsia: a multicenter, randomized, placebo-Controlled, clinical trial. Clin. Transl. Gastroenterol. 11 (10), e00241. doi:10.14309/ctg.0000000000000241
Chen, J., Zhu, W., Zeng, X., Yang, K., Peng, H., and Hu, L. (2022a). Paeoniflorin exhibits antidepressant activity in rats with postpartum depression via the TSPO and BDNF-mTOR pathways. Acta Neurobiol. Exp. (Wars) 82 (3), 347–357. doi:10.55782/ane-2022-033
Chen, Q., Li, C., Tao, E., Asakawa, T., and Zhang, Y. (2022b). Exploration of a brain-liver-communication-related mechanism involved in the experimental perimenopausal depression rat model using Chaihu-Shugan-San. Neurochem. Res. 47 (5), 1354–1368. doi:10.1007/s11064-022-03534-y
Cheng, J., Chen, M., Wan, H. Q., Chen, X. Q., Li, C. F., Zhu, J. X., et al. (2021). Paeoniflorin exerts antidepressant-like effects through enhancing neuronal FGF-2 by microglial inactivation. J. Ethnopharmacol. 274, 114046. doi:10.1016/j.jep.2021.114046
Cipriani, A., Furukawa, T. A., Salanti, G., Chaimani, A., Atkinson, L. Z., Ogawa, Y., et al. (2018). Comparative efficacy and acceptability of 21 antidepressant drugs for the acute treatment of adults with major depressive disorder: a systematic review and network meta-analysis. Focus Am Psychiatr. Publ. 16 (4), 420–429. doi:10.1176/appi.focus.16407
Collaborators, G. M. D. (2022). Global, regional, and national burden of 12 mental disorders in 204 countries and territories, 1990-2019: a systematic analysis for the global burden of disease study 2019. Lancet Psychiatry 9 (2), 137–150. doi:10.1016/S2215-0366(21)00395-3
Colucci-D'Amato, L., Speranza, L., and Volpicelli, F. (2020). Neurotrophic Factor BDNF, physiological functions and therapeutic potential in depression, neurodegeneration and brain cancer. Int. J. Mol. Sci. 21 (20), 7777. doi:10.3390/ijms21207777
Cong, C., Kluwe, L., Li, S., Liu, X., Liu, Y., Liu, H., et al. (2019). Paeoniflorin inhibits tributyltin chloride-induced apoptosis in hypothalamic neurons via inhibition of MKK4-JNK signaling pathway. J. Ethnopharmacol. 237, 1–8. doi:10.1016/j.jep.2019.03.030
Cuperfain, A. B., Zhang, Z. L., Kennedy, J. L., and Goncalves, V. F. (2018). The complex interaction of mitochondrial genetics and mitochondrial pathways in psychiatric disease. Mol. Neuropsychiatry 4 (1), 52–69. doi:10.1159/000488031
Curtis, G. C., and Mendels, J. (1976). Neuroendocrine regulation in depression. I. Limbic system-adrenocortical dysfunction. Arch. Gen. Psychiatry 33 (9), 1039–1044. doi:10.1001/archpsyc.1976.01770090029002
De Kloet, E. R., Vreugdenhil, E., Oitzl, M. S., and Joels, M. (1998). Brain corticosteroid receptor balance in health and disease. Endocr. Rev. 19 (3), 269–301. doi:10.1210/edrv.19.3.0331
De-Miguel, F. F., and Trueta, C. (2005). Synaptic and extrasynaptic secretion of serotonin. Cell Mol. Neurobiol. 25 (2), 297–312. doi:10.1007/s10571-005-3061-z
Delgado, P. L. (2000). Depression: the case for a monoamine deficiency. J. Clin. Psychiatry 61 (Suppl. 6), 7–11.
Deng, D., Cui, Y., Gan, S., Xie, Z., Cui, S., Cao, K., et al. (2022). Sinisan alleviates depression-like behaviors by regulating mitochondrial function and synaptic plasticity in maternal separation rats. Phytomedicine 106, 154395. doi:10.1016/j.phymed.2022.154395
Dhir, A., and Kulkarni, S. K. (2011). Nitric oxide and major depression. Nitric Oxide 24 (3), 125–131. doi:10.1016/j.niox.2011.02.002
Dinan, T. G., and Scott, L. V. (2005). Anatomy of melancholia: focus on hypothalamic-pituitary-adrenal axis overactivity and the role of vasopressin. J. Anat. 207 (3), 259–264. doi:10.1111/j.1469-7580.2005.00443.x
Diseases, G. B. D., and Injuries, C. (2020). Global burden of 369 diseases and injuries in 204 countries and territories, 1990-2019: a systematic analysis for the global burden of Disease study 2019. Lancet 396 (10258), 1204–1222. doi:10.1016/S0140-6736(20)30925-9
Disner, S. G., Beevers, C. G., Haigh, E. A., and Beck, A. T. (2011). Neural mechanisms of the cognitive model of depression. Nat. Rev. Neurosci. 12 (8), 467–477. doi:10.1038/nrn3027
Dougherty, D. D., Rezai, A. R., Carpenter, L. L., Howland, R. H., Bhati, M. T., O'Reardon, J. P., et al. (2015). A randomized sham-controlled trial of deep brain stimulation of the ventral capsule/ventral striatum for chronic treatment-resistant depression. Biol. Psychiatry 78 (4), 240–248. doi:10.1016/j.biopsych.2014.11.023
Du, H. G., Ming, L., Chen, S. J., and Li, C. D. (2014). Xiaoyao pill for treatment of functional dyspepsia in perimenopausal women with depression. World J. Gastroenterol. 20 (44), 16739–16744. doi:10.3748/wjg.v20.i44.16739
Duan, H., Zhang, Q., Liu, J., Li, R., Wang, D., Peng, W., et al. (2021). Suppression of apoptosis in vascular endothelial cell, the promising way for natural medicines to treat atherosclerosis. Pharmacol. Res. 168, 105599. doi:10.1016/j.phrs.2021.105599
Duman, R. S. (2004). Depression: a case of neuronal life and death? Biol. Psychiatry 56 (3), 140–145. doi:10.1016/j.biopsych.2004.02.033
Dwivedi, Y., Rizavi, H. S., Conley, R. R., Roberts, R. C., Tamminga, C. A., and Pandey, G. N. (2003). Altered gene expression of brain-derived neurotrophic factor and receptor tyrosine kinase B in postmortem brain of suicide subjects. Arch. Gen. Psychiatry 60 (8), 804–815. doi:10.1001/archpsyc.60.8.804
Dwyer, J. B., Aftab, A., Radhakrishnan, R., Widge, A., Rodriguez, C. I., Carpenter, L. L., et al. (2020). Hormonal treatments for major depressive disorder: state of the art. Am. J. Psychiatry 177 (8), 686–705. doi:10.1176/appi.ajp.2020.19080848
Fan, Q., Liu, Y., Sheng, L., Lv, S., Yang, L., Zhang, Z., et al. (2023). Chaihu-shugan-san inhibits neuroinflammation in the treatment of post-stroke depression through the JAK/STAT3-GSK3β/PTEN/Akt pathway. Biomed. Pharmacother. 160, 114385. doi:10.1016/j.biopha.2023.114385
Fathinezhad, Z., Sewell, R. D. E., Lorigooini, Z., and Rafieian-Kopaei, M. (2019). Depression and treatment with effective herbs. Curr. Pharm. Des. 25 (6), 738–745. doi:10.2174/1381612825666190402105803
Feng, D. D., Tang, T., Lin, X. P., Yang, Z. Y., Yang, S., Xia, Z. A., et al. (2016). Nine traditional Chinese herbal formulas for the treatment of depression: an ethnopharmacology, phytochemistry, and pharmacology review. Neuropsychiatr. Dis. Treat. 12, 2387–2402. doi:10.2147/NDT.S114560
Feng, T., Tripathi, A., and Pillai, A. (2020). Inflammatory pathways in psychiatric disorders: the case of schizophrenia and depression. Curr. Behav. Neurosci. Rep. 7 (3), 128–138. doi:10.1007/s40473-020-00207-4
Gao, Z., Wang, Y., and Yu, H. (2022). A Chinese classical prescription chaihu shugan powder in treatment of post-stroke depression: an overview. Med. Kaunas. 59 (1), 55. doi:10.3390/medicina59010055
Gawryluk, J. W., Wang, J. F., Andreazza, A. C., Shao, L., and Young, L. T. (2011). Decreased levels of glutathione, the major brain antioxidant, in post-mortem prefrontal cortex from patients with psychiatric disorders. Int. J. Neuropsychopharmacol. 14 (1), 123–130. doi:10.1017/S1461145710000805
Gong, Y., Chai, Y., Ding, J. H., Sun, X. L., and Hu, G. (2011). Chronic mild stress damages mitochondrial ultrastructure and function in mouse brain. Neurosci. Lett. 488 (1), 76–80. doi:10.1016/j.neulet.2010.11.006
Grutzner, T. M., Listunova, L., Fabian, G. A., Kramer, B. A., Flach, D., Weisbrod, M., et al. (2018). Serum calcium levels and neuropsychological performance in depression and matched healthy controls: reversal of correlation a marker of the aging cognitive clock? Psychoneuroendocrinology 91, 198–205. doi:10.1016/j.psyneuen.2018.03.012
Gu, X. S., Wang, F., Zhang, C. Y., Mao, C. J., Yang, J., Yang, Y. P., et al. (2016). Neuroprotective effects of paeoniflorin on 6-OHDA-Lesioned rat model of Parkinson's disease. Neurochem. Res. 41 (11), 2923–2936. doi:10.1007/s11064-016-2011-0
Guan, R., and Qu, F. (2023). Observation on the therapeutic effect of Sini powder on Adolescent depression with stagnation of liver-Qi and its effect on the levels of serum IL-1 β, TGF- β 1 and IL-6. Lishizhen Med. Materia Medica Res. 34 (05), 1138–1140.
Guang-zhi, C. (2012). Effect of paeoniflorin on reserpine-induced depression model in mice. Chin. J. Exp. Traditional Med. Formulae 18 (22), 272–274. doi:10.13422/j.cnki.syfjx.2012.22.079
Guerry, J. D., and Hastings, P. D. (2011). In search of HPA axis dysregulation in child and adolescent depression. Clin. Child. Fam. Psychol. Rev. 14 (2), 135–160. doi:10.1007/s10567-011-0084-5
Guo, R. B., Wang, G. F., Zhao, A. P., Gu, J., Sun, X. L., and Hu, G. (2012). Paeoniflorin protects against ischemia-induced brain damages in rats via inhibiting MAPKs/NF-κB-mediated inflammatory responses. PLoS One 7 (11), e49701. doi:10.1371/journal.pone.0049701
Guo, W., Yao, X., Cui, R., Yang, W., and Wang, L. (2022). Mechanisms of Paeoniaceae action as an antidepressant. Front. Pharmacol. 13, 934199. doi:10.3389/fphar.2022.934199
Hamon, M., and Blier, P. (2013). Monoamine neurocircuitry in depression and strategies for new treatments. Prog. Neuropsychopharmacol. Biol. Psychiatry 45, 54–63. doi:10.1016/j.pnpbp.2013.04.009
Han, S. K., Kim, J. K., Park, H. S., Shin, Y. J., and Kim, D. H. (2021). Chaihu-shugan-san (shihosogansan) alleviates restraint stress-generated anxiety and depression in mice by regulating NF-κB-mediated BDNF expression through the modulation of gut microbiota. Chin. Med. 16 (1), 77. doi:10.1186/s13020-021-00492-5
Hara, T., Nakamura, K., Matsui, M., Yamamoto, A., Nakahara, Y., Suzuki-Migishima, R., et al. (2006). Suppression of basal autophagy in neural cells causes neurodegenerative disease in mice. Nature 441 (7095), 885–889. doi:10.1038/nature04724
He, X., Zhang, R., Li, Z., Yao, Z., Xie, X., Bai, R., et al. (2022). Sini powder with paroxetine ameliorates major depressive disorder by modulating circadian rhythm: a randomized, double-blind, placebo-controlled trial. J. Pineal Res. 73 (4), e12832. doi:10.1111/jpi.12832
Hett, D., and Marwaha, S. (2020). Repetitive transcranial magnetic stimulation in the treatment of bipolar disorder. Ther. Adv. Psychopharmacol. 10, 2045125320973790. doi:10.1177/2045125320973790
Hino, H., Takahashi, H., Suzuki, Y., Tanaka, J., Ishii, E., and Fukuda, M. (2012). Anticonvulsive effect of paeoniflorin on experimental febrile seizures in immature rats: possible application for febrile seizures in children. PLoS One 7 (8), e42920. doi:10.1371/journal.pone.0042920
Hirschfeld, R. M. (2000). History and evolution of the monoamine hypothesis of depression. J. Clin. Psychiatry 61 (Suppl. 6), 4–6.
Hong, H., Lu, X., Wu, C., Chen, J., Chen, C., Zhang, J., et al. (2022). A review for the pharmacological effects of paeoniflorin in the nervous system. Front. Pharmacol. 13, 898955. doi:10.3389/fphar.2022.898955
Hou, Y., Feng, X. H., Leng, Y. E., Wei, Y. X., Zeng, Y., Shi, L., et al. (2024). The antitumor activity of paeoniflrin in non-small cell lung cancer A549 cells via EGFR/rhoA pathway. J. NORTH SICHUAN Med. Coll. 39 (09), 1161–1166. doi:10.3969/j.issn.1005.3697.2024.09.002
Hsiu, S. L., Lin, Y. T., Wen, K. C., Hou, Y. C., and Chao, P. D. (2003). A deglucosylated metabolite of paeoniflorin of the root of Paeonia lactiflora and its pharmacokinetics in rats. Planta Med. 69 (12), 1113–1118. doi:10.1055/s-2003-45192
Hu, D., and Sheng, L. (2016). Clinical observation on efficacy of modified Chaihu shugan granule for treating post ischemic stroke depression. Chin. Traditional Herb. Drugs 47 (21), 3866–3870.
Hu, P. Y., Liu, D., Zheng, Q., Wu, Q., Tang, Y., and Yang, M. (2016). Elucidation of transport mechanism of paeoniflorin and the influence of ligustilide, senkyunolide I and senkyunolide A on paeoniflorin transport through Mdck-Mdr1 cells as blood-brain barrier in vitro model. Molecules 21 (3), 300. doi:10.3390/molecules21030300
Hu, M. Z., Wang, A. R., Zhao, Z. Y., Chen, X. Y., Li, Y. B., and Liu, B. (2019). Antidepressant-like effects of paeoniflorin on post-stroke depression in a rat model. Neurol. Res. 41 (5), 446–455. doi:10.1080/01616412.2019.1576361
Huang, H., Zhao, J., Jiang, L., Xie, Y., Xia, Y., Lv, R., et al. (2015). Paeoniflorin improves menopause depression in ovariectomized rats under chronic unpredictable mild stress. Int. J. Clin. Exp. Med. 8 (4), 5103–5111.
Huang, R., Gong, M., Tan, X., Shen, J., Wu, Y., Cai, X., et al. (2024). Effects of chaihu shugan san on brain functional network connectivity in the hippocampus of a perimenopausal depression rat model. Mol. Neurobiol. 61 (3), 1655–1672. doi:10.1007/s12035-023-03615-1
Jenner, P., and Olanow, C. W. (1998). Understanding cell death in parkinson's disease. Ann. Neurol. 44 (3 Suppl. 1), S72–S84. doi:10.1002/ana.410440712
Jiang, Z., Chen, W., Yan, X., Bi, L., Guo, S., and Zhan, Z. (2014). Paeoniflorin protects cells from GalN/TNF-alpha-induced apoptosis via ER stress and mitochondria-dependent pathways in human L02 hepatocytes. Acta Biochim. Biophys. Sin. (Shanghai) 46 (5), 357–367. doi:10.1093/abbs/gmu010
Jiang, P., Guo, Y., Dang, R., Yang, M., Liao, D., Li, H., et al. (2017a). Salvianolic acid B protects against lipopolysaccharide-induced behavioral deficits and neuroinflammatory response: involvement of autophagy and NLRP3 inflammasome. J. Neuroinflammation 14 (1), 239. doi:10.1186/s12974-017-1013-4
Jiang, R., Guo, Z. W., Mao, Y. X., Sun, J., and Jin, S. M. (2017b). Effect of chaihu shugan powder on serum cell factors of patients with post cerebral infarction depression(PCID). Liaoning J. Traditional Chin. Med. 44 (05), 985–988. doi:10.13192/j.issn.1000-1719.2017.05.029
Jiao, H., Yang, H., Yan, Z., Chen, J., Xu, M., Jiang, Y., et al. (2021). Traditional Chinese formula xiaoyaosan alleviates depressive-like behavior in CUMS mice by regulating PEBP1-GPX4-Mediated ferroptosis in the hippocampus. Neuropsychiatr. Dis. Treat. 17, 1001–1019. doi:10.2147/NDT.S302443
Jin, L., Zhang, L. M., Xie, K. Q., Ye, Y., and Feng, L. (2011). Paeoniflorin suppresses the expression of intercellular adhesion molecule-1 (ICAM-1) in endotoxin-treated human monocytic cells. Br. J. Pharmacol. 164 (2b), 694–703. doi:10.1111/j.1476-5381.2011.01464.x
Jin Shumei, S. W., and Cui, G. (2013). Observations on the antidepressant effects of paeoniflorin in a model of drug-induced depression. Shandong Med. J. 53 (19), 28–29.
Johnston, K. J. A., Adams, M. J., Nicholl, B. I., Ward, J., Strawbridge, R. J., McIntosh, A. M., et al. (2019). Identification of novel common variants associated with chronic pain using conditional false discovery rate analysis with major depressive disorder and assessment of pleiotropic effects of LRFN5. Transl. Psychiatry 9 (1), 310. doi:10.1038/s41398-019-0613-4
Juruena, M. F., Bocharova, M., Agustini, B., and Young, A. H. (2018). Atypical depression and non-atypical depression: is HPA axis function a biomarker? A systematic review. J. Affect Disord. 233, 45–67. doi:10.1016/j.jad.2017.09.052
Kasahara, T., and Kato, T. (2018). What can mitochondrial DNA analysis tell us about mood disorders? Biol. Psychiatry 83 (9), 731–738. doi:10.1016/j.biopsych.2017.09.010
Kaushik, S., and Cuervo, A. M. (2012). Chaperone-mediated autophagy: a unique way to enter the lysosome world. Trends Cell Biol. 22 (8), 407–417. doi:10.1016/j.tcb.2012.05.006
Kessing, L. V., Ziersen, S. C., Andersen, F. M., Gerds, T., and Budtz-Jorgensen, E. (2024). Comparative responses to 17 different antidepressants in major depressive disorder: results from a 2-year long-term nation-wide population-based study emulating a randomized trial. Acta Psychiatr. Scand. 149, 378–388. doi:10.1111/acps.13673
Kessler, D., Bennewith, O., Lewis, G., and Sharp, D. (2002). Detection of depression and anxiety in primary care: follow up study. BMJ 325 (7371), 1016–1017. doi:10.1136/bmj.325.7371.1016
Kim, Y. K., and Won, E. (2017). The influence of stress on neuroinflammation and alterations in brain structure and function in major depressive disorder. Behav. Brain Res. 329, 6–11. doi:10.1016/j.bbr.2017.04.020
Kim, M., Lee, H. S., LaForet, G., McIntyre, C., Martin, E. J., Chang, P., et al. (1999). Mutant huntingtin expression in clonal striatal cells: dissociation of inclusion formation and neuronal survival by caspase inhibition. J. Neurosci. 19 (3), 964–973. doi:10.1523/JNEUROSCI.19-03-00964.1999
Kim, S. H., Han, J., Seog, D. H., Chung, J. Y., Kim, N., Hong Park, Y., et al. (2005). Antidepressant effect of Chaihu-Shugan-San extract and its constituents in rat models of depression. Life Sci. 76 (11), 1297–1306. doi:10.1016/j.lfs.2004.10.022
Knaudt, P. R., Connor, K. M., Weisler, R. H., Churchill, L. E., and Davidson, J. R. (1999). Alternative therapy use by psychiatric outpatients. J. Nerv. Ment. Dis. 187 (11), 692–695. doi:10.1097/00005053-199911000-00007
Komatsu, M., Waguri, S., Chiba, T., Murata, S., Iwata, J., Tanida, I., et al. (2006). Loss of autophagy in the central nervous system causes neurodegeneration in mice. Nature 441 (7095), 880–884. doi:10.1038/nature04723
Kong, Y., Peng, Q., Lv, N., Yuan, J., Deng, Z., Liang, X., et al. (2020). Paeoniflorin exerts neuroprotective effects in a transgenic mouse model of alzheimer's disease via activation of adenosine A(1) receptor. Neurosci. Lett. 730, 135016. doi:10.1016/j.neulet.2020.135016
Kosten, T. A., Galloway, M. P., Duman, R. S., Russell, D. S., and D'Sa, C. (2008). Repeated unpredictable stress and antidepressants differentially regulate expression of the bcl-2 family of apoptotic genes in rat cortical, hippocampal, and limbic brain structures. Neuropsychopharmacology 33 (7), 1545–1558. doi:10.1038/sj.npp.1301527
Lan, Z., Chen, L., Fu, Q., Ji, W., Wang, S., Liang, Z., et al. (2013). Paeoniflorin attenuates amyloid-beta peptide-induced neurotoxicity by ameliorating oxidative stress and regulating the NGF-Mediated signaling in rats. Brain Res. 1498, 9–19. doi:10.1016/j.brainres.2012.12.040
Li, F. F., and Kang, X. D. (2019). Clinical observation of jiawei xiaoyaosan in treating liver depression and spleen deficiency type 2 diabetes with emotional disorder. Chin. J. Exp. Traditional Med. Formulae 25 (02), 134–138. doi:10.13422/j.cnki.syfjx.20190116
Li, C. R., Zhou, Z., Zhu, D., Sun, Y. N., Dai, J. M., and Wang, S. Q. (2007). Protective effect of paeoniflorin on irradiation-induced cell damage involved in modulation of reactive oxygen species and the mitogen-activated protein kinases. Int. J. Biochem. Cell Biol. 39 (2), 426–438. doi:10.1016/j.biocel.2006.09.011
Li, Y. H., Zhang, C. H., Wang, S. E., Qiu, J., Hu, S. Y., and Xiao, G. L. (2009). Effects of chaihu shugan san on behavior and plasma levels of corticotropin releasing hormone and adrenocorticotropic hormone of rats with chronic mild unpredicted stress depression. Zhong Xi Yi Jie He Xue Bao 7 (11), 1073–1077. doi:10.3736/jcim20091110
Li, J., Ji, X., Zhang, J., Shi, G., Zhu, X., and Wang, K. (2014). Paeoniflorin attenuates Aβ25-35-induced neurotoxicity in PC12 cells by preventing mitochondrial dysfunction. Folia Neuropathol. 52 (3), 285–290.
Li, X., Shi, F., Zhang, R., Sun, C., Gong, C., Jian, L., et al. (2016). Pharmacokinetics, safety, and tolerability of amygdalin and paeoniflorin after single and multiple intravenous infusions of Huoxue-Tongluo lyophilized powder for injection in healthy Chinese volunteers. Clin. Ther. 38 (2), 327–337. doi:10.1016/j.clinthera.2015.12.005
Li, J., Huang, S., Huang, W., Wang, W., Wen, G., Gao, L., et al. (2017). Paeoniflorin ameliorates interferon-alpha-induced neuroinflammation and depressive-like behaviors in mice. Oncotarget 8 (5), 8264–8282. doi:10.18632/oncotarget.14160
Li, H., Sagar, A. P., and Keri, S. (2018). Translocator protein (18kDa TSPO) binding, a marker of microglia, is reduced in major depression during cognitive-behavioral therapy. Prog. Neuropsychopharmacol. Biol. Psychiatry 83, 1–7. doi:10.1016/j.pnpbp.2017.12.011
Li, Y. C., Zheng, X. X., Xia, S. Z., Li, Y., Deng, H. H., Wang, X., et al. (2020). Paeoniflorin ameliorates depressive-like behavior in prenatally stressed offspring by restoring the HPA axis- and glucocorticoid receptor-associated dysfunction. J. Affect Disord. 274, 471–481. doi:10.1016/j.jad.2020.05.078
Li, Y., Li, L., Guo, R., Yu, H., Wang, X., Wang, B., et al. (2022). Clinical efficacy of shugan granule in the treatment of mixed anxiety-depressive disorder: a multicenter, randomized, double-blind, placebo-controlled trial. J. Ethnopharmacol. 290, 115032. doi:10.1016/j.jep.2022.115032
Lin, Y. T., Huang, W. S., Tsai, H. Y., Lee, M. M., and Chen, Y. F. (2019). In vivo microdialysis and in vitro HPLC analysis of the impact of paeoniflorin on the monoamine levels and their metabolites in the rodent brain. Biomed. (Taipei) 9 (2), 11. doi:10.1051/bmdcn/2019090211
Liu, D. Z., Xie, K. Q., Ji, X. Q., Ye, Y., Jiang, C. L., and Zhu, X. Z. (2005). Neuroprotective effect of paeoniflorin on cerebral ischemic rat by activating adenosine A1 receptor in a manner different from its classical agonists. Br. J. Pharmacol. 146 (4), 604–611. doi:10.1038/sj.bjp.0706335
Liu, H. Q., Zhang, W. Y., Luo, X. T., Ye, Y., and Zhu, X. Z. (2006a). Paeoniflorin attenuates neuroinflammation and dopaminergic neurodegeneration in the MPTP model of parkinson's disease by activation of adenosine A1 receptor. Br. J. Pharmacol. 148 (3), 314–325. doi:10.1038/sj.bjp.0706732
Liu, Z. Q., Jiang, Z. H., Liu, L., and Hu, M. (2006b). Mechanisms responsible for poor oral bioavailability of paeoniflorin: role of intestinal disposition and interactions with sinomenine. Pharm. Res. 23 (12), 2768–2780. doi:10.1007/s11095-006-9100-8
Liu, J., Jin, D. Z., Xiao, L., and Zhu, X. Z. (2006c). Paeoniflorin attenuates chronic cerebral hypoperfusion-induced learning dysfunction and brain damage in rats. Brain Res. 1089 (1), 162–170. doi:10.1016/j.brainres.2006.02.115
Liu, H., Wang, J., Wang, J., Wang, P., and Xue, Y. (2015a). Paeoniflorin attenuates Aβ1-42-induced inflammation and chemotaxis of microglia in vitro and inhibits NF-κB- and VEGF/Flt-1 signaling pathways. Brain Res. 1618, 149–158. doi:10.1016/j.brainres.2015.05.035
Liu, L., Liu, C., Wang, Y., Wang, P., Li, Y., and Li, B. (2015b). Herbal medicine for anxiety, depression and insomnia. Curr. Neuropharmacol. 13 (4), 481–493. doi:10.2174/1570159x1304150831122734
Liu, Y., Zhao, J., and Guo, W. (2018a). Emotional roles of mono-aminergic neurotransmitters in major depressive disorder and anxiety disorders. Front. Psychol. 9, 2201. doi:10.3389/fpsyg.2018.02201
Liu, Q., Sun, N. N., Wu, Z. Z., Fan, D. H., and Cao, M. Q. (2018b). Chaihu-shugan-san exerts an antidepressive effect by downregulating miR-124 and releasing inhibition of the MAPK14 and Gria3 signaling pathways. Neural Regen. Res. 13 (5), 837–845. doi:10.4103/1673-5374.232478
Liu, S. C., Hu, W. Y., Zhang, W. Y., Yang, L., Li, Y., Xiao, Z. C., et al. (2019a). Paeoniflorin attenuates impairment of spatial learning and hippocampal long-term potentiation in mice subjected to chronic unpredictable mild stress. Psychopharmacol. Berl. 236 (9), 2823–2834. doi:10.1007/s00213-019-05257-5
Liu, Y., Ding, X. F., Wang, X. X., Zou, X. J., Li, X. J., Liu, Y. Y., et al. (2019b). Xiaoyaosan exerts antidepressant-like effects by regulating the functions of astrocytes and EAATs in the prefrontal cortex of mice. BMC Complement. Altern. Med. 19 (1), 215. doi:10.1186/s12906-019-2613-6
Liu, T., Kong, W., Liu, Y., and Song, D. (2020a). Efficacy and prognostic factors of concurrent chemoradiotherapy in patients with stage Ib3 and IIa2 cervical cancer. Ginekol. Pol. 91 (2), 57–61. doi:10.5603/gp.2020.0017
Liu, X., Liu, C., Tian, J., Gao, X., Li, K., Du, G., et al. (2020b). Plasma metabolomics of depressed patients and treatment with xiaoyaosan based on mass spectrometry technique. J. Ethnopharmacol. 246, 112219. doi:10.1016/j.jep.2019.112219
Lu, J. T., He, W., Song, S. S., and Wei, W. (2014). Paeoniflorin inhibited the tumor invasion and metastasis in human hepatocellular carcinoma cells. Bratisl. Lek. Listy 115 (7), 427–433. doi:10.4149/bll_2014_084
Lu, J., Fu, L., Qin, G., Shi, P., and Fu, W. (2018). The regulatory effect of xiaoyao san on glucocorticoid receptors under the condition of chronic stress. Cell Mol. Biol. (Noisy-le-grand) 64 (6), 103–109. doi:10.14715/cmb/2018.64.6.17
Lucassen, P. J., Heine, V. M., Muller, M. B., van der Beek, E. M., Wiegant, V. M., De Kloet, E. R., et al. (2006). Stress, depression and hippocampal apoptosis. CNS Neurol. Disord. Drug Targets 5 (5), 531–546. doi:10.2174/187152706778559273
Luo, X. Q., Li, A., Yang, X., Xiao, X., Hu, R., Wang, T. W., et al. (2018). Paeoniflorin exerts neuroprotective effects by modulating the M1/M2 subset polarization of microglia/macrophages in the hippocampal CA1 region of vascular dementia rats via cannabinoid receptor 2. Chin. Med. 13, 14. doi:10.1186/s13020-018-0173-1
Lv, M., Wang, Y., Qu, P., Li, S., Yu, Z., Qin, X., et al. (2021). A combination of cecum microbiome and metabolome in CUMS depressed rats reveals the antidepressant mechanism of traditional Chinese medicines: a case study of xiaoyaosan. J. Ethnopharmacol. 276, 114167. doi:10.1016/j.jep.2021.114167
Lyu, R., Wang, W., Liu, L. J., Li, N. J., Xu, W. H., and Yao, S. K. (2022). Effects of chaihu shugan powder on symptoms, anxiety, depression and radionuclide gastric emptying of functional dyspepsia of liver stomach qi stagnation type. China J. Traditional Chin. Med. Pharm. 37 (03), 1815–1818.
Ma, X. L., and Li, L. Y. (2019). Clinical effect of xiaoyao pill combined with venlafaxine on postpartum depression patients with deficiency of Qi and blood. Chin. Tradit. Pat. Med. 41 (09), 2261–2263.
Ma, Q., Li, X., Yan, Z., Jiao, H., Wang, T., Hou, Y., et al. (2019). Xiaoyaosan ameliorates chronic immobilization stress-induced depression-like behaviors and anorexia in rats: the role of the Nesfatin-1-Oxytocin-Proopiomelanocortin neural pathway in the hypothalamus. Front. Psychiatry 10, 910. doi:10.3389/fpsyt.2019.00910
Maes, M., Galecki, P., Chang, Y. S., and Berk, M. (2011). A review on the oxidative and nitrosative stress (O&NS) pathways in major depression and their possible contribution to the (neuro)degenerative processes in that illness. Prog. Neuropsychopharmacol. Biol. Psychiatry 35 (3), 676–692. doi:10.1016/j.pnpbp.2010.05.004
Malberg, J. E., Hen, R., and Madsen, T. M. (2021). Adult neurogenesis and antidepressant treatment: the Surprise finding by ron duman and the field 20 years later. Biol. Psychiatry 90 (2), 96–101. doi:10.1016/j.biopsych.2021.01.010
Mao, Q. Q., Ip, S. P., Tsai, S. H., and Che, C. T. (2008). Antidepressant-like effect of peony glycosides in mice. J. Ethnopharmacol. 119 (2), 272–275. doi:10.1016/j.jep.2008.07.008
Mao, Q. Q., Ip, S. P., Ko, K. M., Tsai, S. H., and Che, C. T. (2009a). Peony glycosides produce antidepressant-like action in mice exposed to chronic unpredictable mild stress: effects on hypothalamic-pituitary-adrenal function and brain-derived neurotrophic factor. Prog. Neuropsychopharmacol. Biol. Psychiatry 33 (7), 1211–1216. doi:10.1016/j.pnpbp.2009.07.002
Mao, Q. Q., Ip, S. P., Ko, K. M., Tsai, S. H., Xian, Y. F., and Che, C. T. (2009b). Effects of peony glycosides on mice exposed to chronic unpredictable stress: further evidence for antidepressant-like activity. J. Ethnopharmacol. 124 (2), 316–320. doi:10.1016/j.jep.2009.04.019
Mao, Q. Q., Ip, S. P., Ko, K. M., Tsai, S. H., Zhao, M., and Che, C. T. (2009c). Peony glycosides protect against corticosterone-induced neurotoxicity in PC12 cells. Cell Mol. Neurobiol. 29 (5), 643–647. doi:10.1007/s10571-009-9357-7
Mao, Q. Q., Zhong, X. M., Feng, C. R., Pan, A. J., Li, Z. Y., and Huang, Z. (2010). Protective effects of paeoniflorin against glutamate-induced neurotoxicity in PC12 cells via antioxidant mechanisms and Ca(2+) antagonism. Cell Mol. Neurobiol. 30 (7), 1059–1066. doi:10.1007/s10571-010-9537-5
Mao, Q. Q., Xian, Y. F., Ip, S. P., Tsai, S. H., and Che, C. T. (2011a). Protective effects of peony glycosides against corticosterone-induced cell death in PC12 cells through antioxidant action. J. Ethnopharmacol. 133 (3), 1121–1125. doi:10.1016/j.jep.2010.11.043
Mao, Q. Q., Zhong, X. M., Li, Z. Y., and Huang, Z. (2011b). Paeoniflorin protects against NMDA-Induced neurotoxicity in PC12 cells via Ca2+ antagonism. Phytother. Res. 25 (5), 681–685. doi:10.1002/ptr.3321
Mao, Q. Q., Zhong, X. M., Qiu, F. M., Li, Z. Y., and Huang, Z. (2012a). Protective effects of paeoniflorin against corticosterone-induced neurotoxicity in PC12 cells. Phytother. Res. 26 (7), 969–973. doi:10.1002/ptr.3673
Mao, Q. Q., Huang, Z., Ip, S. P., Xian, Y. F., and Che, C. T. (2012b). Peony glycosides reverse the effects of corticosterone on behavior and brain BDNF expression in rats. Behav. Brain Res. 227 (1), 305–309. doi:10.1016/j.bbr.2011.11.016
Marwaha, S., Palmer, E., Suppes, T., Cons, E., Young, A. H., and Upthegrove, R. (2023). Novel and emerging treatments for major depression. Lancet 401 (10371), 141–153. doi:10.1016/S0140-6736(22)02080-3
McKernan, D. P., Dinan, T. G., and Cryan, J. F. (2009). Killing the blues: a role for cellular suicide (apoptosis) in depression and the antidepressant response? Prog. Neurobiol. 88 (4), 246–263. doi:10.1016/j.pneurobio.2009.04.006
McKim, D. B., Niraula, A., Tarr, A. J., Wohleb, E. S., Sheridan, J. F., and Godbout, J. P. (2016). Neuroinflammatory dynamics underlie memory impairments after repeated social defeat. J. Neurosci. 36 (9), 2590–2604. doi:10.1523/JNEUROSCI.2394-15.2016
Mehta, A., Prabhakar, M., Kumar, P., Deshmukh, R., and Sharma, P. L. (2013). Excitotoxicity: bridge to various triggers in neurodegenerative disorders. Eur. J. Pharmacol. 698 (1-3), 6–18. doi:10.1016/j.ejphar.2012.10.032
Meng, H., Li, C., Feng, L., Cheng, B., Wu, F., Wang, X., et al. (2007). Effects of ginkgolide B on 6-OHDA-induced apoptosis and calcium over load in cultured PC12. Int. J. Dev. Neurosci. 25 (8), 509–514. doi:10.1016/j.ijdevneu.2007.09.010
Menke, A. (2019). Is the HPA axis as target for depression outdated, or is there a new hope? Front. Psychiatry 10, 101. doi:10.3389/fpsyt.2019.00101
Michel, T. M., Camara, S., Tatschner, T., Frangou, S., Sheldrick, A. J., Riederer, P., et al. (2010). Increased xanthine oxidase in the thalamus and putamen in depression. World J. Biol. Psychiatry 11 (2 Pt 2), 314–320. doi:10.3109/15622970802123695
Minmin Zhao, M. G., Tian, Y., Du, Y., Wang, C., Xu, H., Zhang, L., et al. (2014). Pharmacokinetic and tissue distribution studies of paeoniflorin and albiflorin in rats after oral administration of total glycosides of paeony by HPLC-MS/MS. J. Chin. Pharm. Sci. 23 (06), 403–411. doi:10.5246/jcps.2014.06.054
Miranda, A. M., Lasiecka, Z. M., Xu, Y., Neufeld, J., Shahriar, S., Simoes, S., et al. (2018). Neuronal lysosomal dysfunction releases exosomes harboring APP C-terminal fragments and unique lipid signatures. Nat. Commun. 9 (1), 291. doi:10.1038/s41467-017-02533-w
Miuli, A., Sepede, G., Stigliano, G., Mosca, A., Di Carlo, F., d'Andrea, G., et al. (2021). Hypomanic/Manic switch after transcranial magnetic stimulation in mood disorders: a systematic review and meta-analysis. World J. Psychiatry 11 (8), 477–490. doi:10.5498/wjp.v11.i8.477
Moffa, A. H., Martin, D., Alonzo, A., Bennabi, D., Blumberger, D. M., Bensenor, I. M., et al. (2020). Efficacy and acceptability of transcranial direct current stimulation (tDCS) for major depressive disorder: an individual patient data meta-analysis. Prog. Neuropsychopharmacol. Biol. Psychiatry 99, 109836. doi:10.1016/j.pnpbp.2019.109836
Mu, D. Z., Xue, M., Xu, J. J., Hu, Y., Chen, Y., Ren, P., et al. (2020). Antidepression and prokinetic effects of paeoniflorin on rats in the forced swimming test via polypharmacology. Evid. Based Complement. Altern. Med. 2020, 2153571. doi:10.1155/2020/2153571
Munshi, S., Loh, M. K., Ferrara, N., DeJoseph, M. R., Ritger, A., Padival, M., et al. (2020). Repeated stress induces a pro-inflammatory state, increases amygdala neuronal and microglial activation, and causes anxiety in adult Male rats. Brain Behav. Immun. 84, 180–199. doi:10.1016/j.bbi.2019.11.023
Nagashima, S., Tabara, L. C., Tilokani, L., Paupe, V., Anand, H., Pogson, J. H., et al. (2020). Golgi-derived PI(4)P-containing vesicles drive late steps of mitochondrial division. Science 367 (6484), 1366–1371. doi:10.1126/science.aax6089
Nam, K. N., Yae, C. G., Hong, J. W., Cho, D. H., Lee, J. H., and Lee, E. H. (2013). Paeoniflorin, a monoterpene glycoside, attenuates lipopolysaccharide-induced neuronal injury and brain microglial inflammatory response. Biotechnol. Lett. 35 (8), 1183–1189. doi:10.1007/s10529-013-1192-8
Nelson, J. C., and Davis, J. M. (1997). DST studies in psychotic depression: a meta-analysis. Am. J. Psychiatry 154 (11), 1497–1503. doi:10.1176/ajp.154.11.1497
Nemeroff, C. B. (2002). Recent advances in the neurobiology of depression. Psychopharmacol. Bull. 36 (Suppl. 2), 6–23.
Nishiyama, J., Miura, E., Mizushima, N., Watanabe, M., and Yuzaki, M. (2007). Aberrant membranes and double-membrane structures accumulate in the axons of Atg5-null purkinje cells before neuronal death. Autophagy 3 (6), 591–596. doi:10.4161/auto.4964
Nouri, A., Hashemzadeh, F., Soltani, A., Saghaei, E., and Amini-Khoei, H. (2020). Progesterone exerts antidepressant-like effect in a mouse model of maternal separation stress through mitigation of neuroinflammatory response and oxidative stress. Pharm. Biol. 58 (1), 64–71. doi:10.1080/13880209.2019.1702704
Nunnari, J., and Suomalainen, A. (2012). Mitochondria: in sickness and in health. Cell 148 (6), 1145–1159. doi:10.1016/j.cell.2012.02.035
Orlovsky, M. A., Dosenko, V. E., Spiga, F., Skibo, G. G., and Lightman, S. L. (2014). Hippocampus remodeling by chronic stress accompanied by GR, proteasome and caspase-3 overexpression. Brain Res. 1593, 83–94. doi:10.1016/j.brainres.2014.09.059
Papp, M., Klimek, V., and Willner, P. (1994). Effects of imipramine on serotonergic and beta-adrenergic receptor binding in a realistic animal model of depression. Psychopharmacol. Berl. 114 (2), 309–314. doi:10.1007/BF02244853
Pariante, C. M., and Lightman, S. L. (2008). The HPA axis in major depression: classical theories and new developments. Trends Neurosci. 31 (9), 464–468. doi:10.1016/j.tins.2008.06.006
Park, H., and Poo, M. M. (2013). Neurotrophin regulation of neural circuit development and function. Nat. Rev. Neurosci. 14 (1), 7–23. doi:10.1038/nrn3379
Patel, M. (2016). Targeting oxidative stress in central nervous system disorders. Trends Pharmacol. Sci. 37 (9), 768–778. doi:10.1016/j.tips.2016.06.007
Peng, G. J., Tian, J. S., Gao, X. X., Zhou, Y. Z., and Qin, X. M. (2015). Research on the pathological mechanism and drug treatment mechanism of depression. Curr. Neuropharmacol. 13 (4), 514–523. doi:10.2174/1570159x1304150831120428
Peng, W., Chen, Y., Tumilty, S., Liu, L., Luo, L., Yin, H., et al. (2022). Paeoniflorin is a promising natural monomer for neurodegenerative diseases via modulation of Ca(2+) and ROS homeostasis. Curr. Opin. Pharmacol. 62, 97–102. doi:10.1016/j.coph.2021.11.009
Perkins, G., Renken, C., Martone, M. E., Young, S. J., Ellisman, M., and Frey, T. (1997). Electron tomography of neuronal mitochondria: three-dimensional structure and organization of cristae and membrane contacts. J. Struct. Biol. 119 (3), 260–272. doi:10.1006/jsbi.1997.3885
Phillips, C. (2017). Brain-derived neurotrophic factor, depression, and physical activity: making the neuroplastic connection. Neural Plast. 2017, 7260130. doi:10.1155/2017/7260130
Poprac, P., Jomova, K., Simunkova, M., Kollar, V., Rhodes, C. J., and Valko, M. (2017). Targeting free radicals in oxidative stress-related human diseases. Trends Pharmacol. Sci. 38 (7), 592–607. doi:10.1016/j.tips.2017.04.005
Qingwei, C. Z. L. Y. C. (2018). Study on the effect of albiflorin and paeoniflorin on rat models with blood deficiency and the liver depression syndrome and the mechanism. J. New Chin. Med. 50 (10), 14–17. doi:10.13457/j.cnki.jncm.2018.10.004
Qiu, F. M., Zhong, X. M., Mao, Q. Q., and Huang, Z. (2013). Antidepressant-like effects of paeoniflorin on the behavioural, biochemical, and neurochemical patterns of rats exposed to chronic unpredictable stress. Neurosci. Lett. 541, 209–213. doi:10.1016/j.neulet.2013.02.029
Qiu, J., Hu, S. Y., Shi, G. Q., and Wang, S. E. (2014a). Changes in regional cerebral blood flow with Chaihu-Shugan-San in the treatment of major depression. Pharmacogn. Mag. 10 (40), 503–508. doi:10.4103/0973-1296.141775
Qiu, J., Hu, S. Y., Zhang, C. H., Shi, G. Q., Wang, S. E., and Xiang, T. (2014b). The effect of Chaihu-Shugan-San and its components on the expression of ERK5 in the hippocampus of depressed rats. J. Ethnopharmacol. 152 (2), 320–326. doi:10.1016/j.jep.2014.01.004
Rao, K. H., Xiao, J. F., Zhao, L. Q., Huang, C. H., and Li, D. H. (2021). Effect of jiawei sini powder on plasma IL-1 β, IL-18, hcy and neurokines in patients with depression after acute cerebral infarction. Lishizhen Med. Materia Medica Res. 32 (05), 1097–1100.
Rao, C., Hu, R., Hu, Y., Jiang, Y., Zou, X., Tang, H., et al. (2024). Theoretical exploring of potential mechanisms of antithrombotic ingredients in danshen-chishao herb-pair by network pharmacological study, molecular docking and zebrafish models. Chin. Med. 19 (1), 100. doi:10.1186/s13020-024-00970-6
Ray, W. A., Griffin, M. R., Schaffner, W., Baugh, D. K., and Melton, L. J. (1987). Psychotropic drug use and the risk of hip fracture. N. Engl. J. Med. 316 (7), 363–369. doi:10.1056/NEJM198702123160702
Roose, S. P., Glassman, A. H., Attia, E., and Woodring, S. (1994). Comparative efficacy of selective serotonin reuptake inhibitors and tricyclics in the treatment of melancholia. Am. J. Psychiatry 151 (12), 1735–1739. doi:10.1176/ajp.151.12.1735
Rush, A. J., Warden, D., Wisniewski, S. R., Fava, M., Trivedi, M. H., Gaynes, B. N., et al. (2009). STAR*D: revising conventional wisdom. CNS Drugs 23 (8), 627–647. doi:10.2165/00023210-200923080-00001
Sacher, J., Rekkas, P. V., Wilson, A. A., Houle, S., Romano, L., Hamidi, J., et al. (2015). Relationship of monoamine oxidase-A distribution volume to postpartum depression and postpartum crying. Neuropsychopharmacology 40 (2), 429–435. doi:10.1038/npp.2014.190
Salunga, T. L., Tabuchi, Y., Takasaki, I., Feril, L. B., Zhao, Q. L., Ohtsuka, K., et al. (2007). Identification of genes responsive to paeoniflorin, a heat shock protein-inducing compound, in human leukemia U937 cells. Int. J. Hyperth. 23 (6), 529–537. doi:10.1080/02656730701639499
Sargent, P. A., Rabiner, E. A., Bhagwagar, Z., Clark, L., Cowen, P., Goodwin, G. M., et al. (2010). 5-HT(1A) receptor binding in euthymic bipolar patients using positron emission tomography with [carbonyl-(11)C]WAY-100635. J. Affect Disord. 123 (1-3), 77–80. doi:10.1016/j.jad.2009.07.015
Sasaki, K., and Yoshizaki, F. (2015). Investigation into hippocampal nerve cell damage through the mineralocorticoid receptor in mice. Mol. Med. Rep. 12 (5), 7211–7220. doi:10.3892/mmr.2015.4406
Seki, K., Yoshida, S., and Jaiswal, M. K. (2018). Molecular mechanism of noradrenaline during the stress-induced major depressive disorder. Neural Regen. Res. 13 (7), 1159–1169. doi:10.4103/1673-5374.235019
Setiawan, E., Attwells, S., Wilson, A. A., Mizrahi, R., Rusjan, P. M., Miler, L., et al. (2018). Association of translocator protein total distribution volume with duration of untreated major depressive disorder: a cross-sectional study. Lancet Psychiatry 5 (4), 339–347. doi:10.1016/S2215-0366(18)30048-8
Shen, Y., Yu, G., and Zhang, H. S. (2018). Clinical observation of xiaoyao pills combined with donepezil hydrochloride in treatment of vascular dementia complicated with depression. Chin. Archives Traditional Chin. Med. 36 (07), 1724–1726. doi:10.13193/j.issn.1673-7717.2018.07.051
Shen, Y., Chen, S., Yu, G., and Yang, H. (2019). Comparative analysis of clinical efficacy of xiaoyao pills and escitalopram in treatment of vascular dementia patients with depression. Chin. Archives Traditional Chin. Med. 37 (02), 396–399. doi:10.13193/j.issn.1673-7717.2019.02.034
Shen, C., Cao, K., Cui, S., Cui, Y., Mo, H., Wen, W., et al. (2020). SiNiSan ameliorates depression-like behavior in rats by enhancing synaptic plasticity via the CaSR-PKC-ERK signaling pathway. Biomed. Pharmacother. 124, 109787. doi:10.1016/j.biopha.2019.109787
Shen, Z., Yu, M., and Dong, Z. (2023). Research progress on the pharmacodynamic mechanisms of sini powder against depression from the perspective of the central nervous system. Med. Kaunas. 59 (4), 741. doi:10.3390/medicina59040741
Shi, J. J., Yuan, Z., Shen, L., Li, H. N., and Zhang, J. R. (2018). Effect of modified chaihu shugan tang in treatment of Liver-Qi stagnation type of post-stroke depression and on serum nerve transmitters and nerve function ecovery. Chin. J. Exp. Traditional Med. Formulae 24 (21), 188–192. doi:10.13422/j.cnki.syfjx.20182029
Shu, Y. Z., Hattori, M., Akao, T., Kobashi, K., Kagei, K., Fukuyama, K., et al. (1987). Metabolism of paeoniflorin and related compounds by human intestinal bacteria. II. Structures of 7S- and 7R-paeonimetabolines I and II formed by Bacteroides fragilis and Lactobacillus brevis. Chem. Pharm. Bull. (Tokyo) 35 (9), 3726–3733. doi:10.1248/cpb.35.3726
Sies, H. (2015). Oxidative stress: a concept in redox biology and medicine. Redox Biol. 4, 180–183. doi:10.1016/j.redox.2015.01.002
Song, C., Wang, J., Gao, D., Yu, Y., Li, F., Wei, S., et al. (2017). Paeoniflorin, the main active ingredient of shuyu capsule, inhibits Cav1.2 and regulates Calmodulin/calmodulin-dependent protein kinase II signalling. Biomed. Res. Int. 2017, 8459287. doi:10.1155/2017/8459287
Song, M., Zhang, J., Li, X., Liu, Y., Wang, T., Yan, Z., et al. (2020). Effects of xiaoyaosan on depressive-like behaviors in rats with chronic unpredictable mild stress through HPA axis induced astrocytic activities. Front. Psychiatry 11, 545823. doi:10.3389/fpsyt.2020.545823
Su, J. H., Anderson, A. J., Cummings, B. J., and Cotman, C. W. (1994). Immunohistochemical evidence for apoptosis in alzheimer's disease. Neuroreport 5 (18), 2529–2533. doi:10.1097/00001756-199412000-00031
Su, R., Fan, J., Li, T., Cao, X., Zhou, J., Han, Z., et al. (2019). Jiawei xiaoyao capsule treatment for mild to moderate major depression with anxiety symptoms: a randomized, double-blind, double-dummy, controlled, multicenter, parallel-treatment trial. J. Tradit. Chin. Med. 39 (3), 410–417.
Sun, X., Cao, Y. B., Hu, L. F., Yang, Y. P., Li, J., Wang, F., et al. (2011). ASICs mediate the modulatory effect by paeoniflorin on alpha-synuclein autophagic degradation. Brain Res. 1396, 77–87. doi:10.1016/j.brainres.2011.04.011
Sun, R., Wang, K., Wu, D., Li, X., and Ou, Y. (2012). Protective effect of paeoniflorin against glutamate-induced neurotoxicity in PC12 cells via Bcl-2/Bax signal pathway. Folia Neuropathol. 50 (3), 270–276. doi:10.5114/fn.2012.30527
Suzuki, H., and Colasanti, M. (2001). NO: a molecule with two masks of 'NO' theatre. Biofactors 15 (2-4), 123–125. doi:10.1002/biof.5520150216
Takeda, S., Isono, T., Wakui, Y., Matsuzaki, Y., Sasaki, H., Amagaya, S., et al. (1995). Absorption and excretion of paeoniflorin in rats. J. Pharm. Pharmacol. 47 (12A), 1036–1040. doi:10.1111/j.2042-7158.1995.tb03293.x
Takeda, S., Isono, T., Wakui, Y., Mizuhara, Y., Amagaya, S., Maruno, M., et al. (1997). In-vivo assessment of extrahepatic metabolism of paeoniflorin in rats: relevance to intestinal floral metabolism. J. Pharm. Pharmacol. 49 (1), 35–39. doi:10.1111/j.2042-7158.1997.tb06748.x
Tartt, A. N., Mariani, M. B., Hen, R., Mann, J. J., and Boldrini, M. (2022). Dysregulation of adult hippocampal neuroplasticity in major depression: pathogenesis and therapeutic implications. Mol. Psychiatry 27 (6), 2689–2699. doi:10.1038/s41380-022-01520-y
Thakore, J. H., Barnes, C., Joyce, J., Medbak, S., and Dinan, T. G. (1997). Effects of antidepressant treatment on corticotropin-induced cortisol responses in patients with melancholic depression. Psychiatry Res. 73 (1-2), 27–32. doi:10.1016/s0165-1781(97)00106-6
Tian, J. S., Shi, B. Y., Xiang, H., Gao, S., Qin, X. M., and Du, G. H. (2013). 1H-NMR-based metabonomic studies on the anti-depressant effect of genipin in the chronic unpredictable mild stress rat model. PLoS One 8 (9), e75721. doi:10.1371/journal.pone.0075721
Tian, J. S., Peng, G. J., Gao, X. X., Zhou, Y. Z., Xing, J., Qin, X. M., et al. (2014). Dynamic analysis of the endogenous metabolites in depressed patients treated with TCM formula xiaoyaosan using urinary (1)H NMR-Based metabolomics. J. Ethnopharmacol. 158 (Pt A), 1–10. doi:10.1016/j.jep.2014.10.005
Tian, R. H., Bai, Y., Li, J. Y., and Guo, K. M. (2019). Reducing PRLR expression and JAK2 activity results in an increase in BDNF expression and inhibits the apoptosis of CA3 hippocampal neurons in a chronic mild stress model of depression. Brain Res. 1725, 146472. doi:10.1016/j.brainres.2019.146472
Tian, D. D., Wang, M., Liu, A., Gao, M. R., Qiu, C., Yu, W., et al. (2021). Antidepressant effect of paeoniflorin is through inhibiting pyroptosis CASP-11/GSDMD pathway. Mol. Neurobiol. 58 (2), 761–776. doi:10.1007/s12035-020-02144-5
Tu, J., Guo, Y., Hong, W., Fang, Y., Han, D., Zhang, P., et al. (2019). The regulatory effects of paeoniflorin and its derivative Paeoniflorin-6'-O-Benzene sulfonate CP-25 on inflammation and immune diseases. Front. Pharmacol. 10, 57. doi:10.3389/fphar.2019.00057
Unutzer, J., Klap, R., Sturm, R., Young, A. S., Marmon, T., Shatkin, J., et al. (2000). Mental disorders and the use of alternative medicine: results from a national survey. Am. J. Psychiatry 157 (11), 1851–1857. doi:10.1176/appi.ajp.157.11.1851
Valera, E., Sanchez-Martin, F. J., Ferrer-Montiel, A. V., Messeguer, A., and Merino, J. M. (2008). NMDA-Induced neuroprotection in hippocampal neurons is mediated through the protein kinase A and CREB (cAMP-response element-binding protein) pathway. Neurochem. Int. 53 (5), 148–154. doi:10.1016/j.neuint.2008.07.007
Vangu, M. D., Esser, J. D., Boyd, I. H., and Berk, M. (2003). Effects of electroconvulsive therapy on regional cerebral blood flow measured by 99mtechnetium HMPAO SPECT. Prog. Neuropsychopharmacol. Biol. Psychiatry 27 (1), 15–19. doi:10.1016/s0278-5846(02)00309-3
Vavakova, M., Durackova, Z., and Trebaticka, J. (2015). Markers of oxidative stress and neuroprogression in depression disorder. Oxid. Med. Cell Longev. 2015, 898393. doi:10.1155/2015/898393
Wang, Y., Fan, R., and Huang, X. (2012). Meta-analysis of the clinical effectiveness of traditional Chinese medicine formula Chaihu-Shugan-San in depression. J. Ethnopharmacol. 141 (2), 571–577. doi:10.1016/j.jep.2011.08.079
Wang, C., Yuan, J., Wu, H. X., Chang, Y., Wang, Q. T., Wu, Y. J., et al. (2013a). Paeoniflorin inhibits inflammatory responses in mice with allergic contact dermatitis by regulating the balance between inflammatory and anti-inflammatory cytokines. Inflamm. Res. 62 (12), 1035–1044. doi:10.1007/s00011-013-0662-8
Wang, D., Wong, H. K., Feng, Y. B., and Zhang, Z. J. (2013b). Paeoniflorin, a natural neuroprotective agent, modulates multiple anti-apoptotic and pro-apoptotic pathways in differentiated PC12 cells. Cell Mol. Neurobiol. 33 (4), 521–529. doi:10.1007/s10571-013-9914-y
Wang, D., Tan, Q. R., and Zhang, Z. J. (2013c). Neuroprotective effects of paeoniflorin, but not the isomer albiflorin, are associated with the suppression of intracellular calcium and calcium/calmodulin protein kinase II in PC12 cells. J. Mol. Neurosci. 51 (2), 581–590. doi:10.1007/s12031-013-0031-7
Wang, Y. L., Han, Q. Q., Gong, W. Q., Pan, D. H., Wang, L. Z., Hu, W., et al. (2018a). Microglial activation mediates chronic mild stress-induced depressive- and anxiety-like behavior in adult rats. J. Neuroinflammation 15 (1), 21. doi:10.1186/s12974-018-1054-3
Wang, D., Liu, L., Li, S., and Wang, C. (2018b). Effects of paeoniflorin on neurobehavior, oxidative stress, brain insulin signaling, and synaptic alterations in intracerebroventricular streptozotocin-induced cognitive impairment in mice. Physiol. Behav. 191, 12–20. doi:10.1016/j.physbeh.2018.03.016
Wang, T., Xu, L., Gao, L., Zhao, L., Liu, X. H., Chang, Y. Y., et al. (2020). Paeoniflorin attenuates early brain injury through reducing oxidative stress and neuronal apoptosis after subarachnoid hemorrhage in rats. Metab. Brain Dis. 35 (6), 959–970. doi:10.1007/s11011-020-00571-w
Wang, X., Hao, J. C., Shang, B., Yang, K. L., He, X. Z., Wang, Z. L., et al. (2021a). Paeoniflorin ameliorates oxidase stress in Glutamate-stimulated SY5Y and prenatally stressed female offspring through Nrf2/HO-1 signaling pathway. J. Affect Disord. 294, 189–199. doi:10.1016/j.jad.2021.07.054
Wang, X. L., Feng, S. T., Wang, Y. T., Chen, N. H., Wang, Z. Z., and Zhang, Y. (2021b). Paeoniflorin: a neuroprotective monoterpenoid glycoside with promising anti-depressive properties. Phytomedicine 90, 153669. doi:10.1016/j.phymed.2021.153669
Wang, Z., Ren, X., Liu, Z., Li, Y., and Wang, T. (2022a). Multimodality treatment for multiple recurrences of cervical cancer after radiotherapy: a case report. Transl. Cancer Res. 11 (4), 943–951. doi:10.21037/tcr-21-2250
Wang, D., Cheng, S., Zou, G., and Ding, X. (2022b). Paeoniflorin inhibits proliferation and migration of psoriatic keratinocytes via the lncRNA NEAT1/miR-3194-5p/Galectin-7 axis. Anticancer Drugs 33 (1), e423–e433. doi:10.1097/cad.0000000000001225
Wang Jing-xia, Z. J.-j., Wei, Li, Chun-ping, M., Yan, L., Lin, Q., and Chen, Z.-zhen (2010). Effects of extract of white peony root on the behavior and cerebral cortex monoamine neurotransmitters in depressive model of chronic unpredictable mild stress rats. China J. Traditional Chin. Med. Pharm. 25 (11), 1895–1897.
Wei, S. S., Yang, H. J., Huang, J. W., Lu, X. P., Peng, L. F., and Wang, Q. G. (2016). Traditional herbal formula sini powder extract produces antidepressant-like effects through stress-related mechanisms in rats. Chin. J. Nat. Med. 14 (8), 590–598. doi:10.1016/S1875-5364(16)30069-3
Whiteford, H. A., Degenhardt, L., Rehm, J., Baxter, A. J., Ferrari, A. J., Erskine, H. E., et al. (2013). Global burden of disease attributable to mental and substance use disorders: findings from the global burden of disease study 2010. Lancet 382 (9904), 1575–1586. doi:10.1016/S0140-6736(13)61611-6
Won, E., Na, K. S., and Kim, Y. K. (2021). Associations between melatonin, neuroinflammation, and brain alterations in depression. Int. J. Mol. Sci. 23 (1), 305. doi:10.3390/ijms23010305
Wu, Y. M., Jin, R., Yang, L., Zhang, J., Yang, Q., Guo, Y. Y., et al. (2013). Phosphatidylinositol 3 kinase/protein kinase B is responsible for the protection of paeoniflorin upon H₂O₂-Induced neural progenitor cell injury. Neuroscience 240, 54–62. doi:10.1016/j.neuroscience.2013.02.037
Wu, Y. F., Yang, M. Z., and Yu, L. (2019). Microbial regulation mechanism of jiawei xiaoyao granule in the treatment of depression with liver stagnation and spleen deficiency syndrome. Lishizhen Med. Materia Medica Res. 30 (03), 633–635.
Wu, C., Li, B., Zhang, Y., Chen, T., Chen, C., Jiang, W., et al. (2020). Intranasal delivery of paeoniflorin nanocrystals for brain targeting. Asian J. Pharm. Sci. 15 (3), 326–335. doi:10.1016/j.ajps.2019.11.002
Wu, Y., Liu, L., Zhao, Y., Li, X., Hu, J., Li, H., et al. (2024). Xiaoyaosan promotes neurotransmitter transmission and alleviates CUMS-Induced depression by regulating the expression of Oct1 and Oct3 in astrocytes of the prefrontal cortex. J. Ethnopharmacol. 326, 117923. doi:10.1016/j.jep.2024.117923
Xi, Y. H., Dang, Z. Q., Zhang, L. L., Niu, X. E., Wang, Y. L., Xu, X. Q., et al. (2014). Clinical study on intervening effects on functional dyspepsia plus depression treated with jiawei sinisan. Chin. J. Exp. Traditional Med. Formulae 20 (03), 202–204.
Xiao, F. H., Chen, X. Q., Yu, Q., Ye, Y., Liu, Y. W., Yan, D., et al. (2018). Transcriptome evidence reveals enhanced autophagy-lysosomal function in centenarians. Genome Res. 28 (11), 1601–1610. doi:10.1101/gr.220780.117
Xin, Q., Yuan, R., Shi, W., Zhu, Z., Wang, Y., and Cong, W. (2019). A review for the anti-inflammatory effects of paeoniflorin in inflammatory disorders. Life Sci. 237, 116925. doi:10.1016/j.lfs.2019.116925
Yan, Z., Jiao, H., Ding, X., Ma, Q., Li, X., Pan, Q., et al. (2018). Xiaoyaosan improves depressive-like behaviors in mice through regulating Apelin-APJ system in hypothalamus. Molecules 23 (5), 1073. doi:10.3390/molecules23051073
Yan, L., Xu, X., He, Z., Wang, S., Zhao, L., Qiu, J., et al. (2020). Antidepressant-like effects and cognitive enhancement of coadministration of chaihu shugan san and fluoxetine: dependent on the BDNF-ERK-CREB signaling pathway in the hippocampus and frontal cortex. Biomed. Res. Int. 2020, 2794263. doi:10.1155/2020/2794263
Yan, W., Dong, Z., Zhao, D., Li, J., Zeng, T., Mo, C., et al. (2021). Xiaoyaosan exerts antidepressant effect by downregulating RAGE expression in cingulate gyrus of depressive-like mice. Front. Pharmacol. 12, 703965. doi:10.3389/fphar.2021.703965
Yan-xia, L. I., Jing-xia, W., Chun, W., Yang, Ge, Yin-feng, H., and Xu, W. (2014). Effects of paeonifl orin on HPA axis and monoamine neurotransmitters in the rats of syndrome of stagnation of liver qi and blood defi ciency. China J. Traditional Chin. Med. Pharm. 29 (08), 2591–2595.
Yang, F. R., Zhu, X. X., Kong, M. W., Zou, X. J., Ma, Q. Y., Li, X. J., et al. (2022). Xiaoyaosan exerts antidepressant-like effect by regulating autophagy involves the expression of GLUT4 in the mice hypothalamic neurons. Front. Pharmacol. 13, 873646. doi:10.3389/fphar.2022.873646
Ye, S., Mao, B., Yang, L., Fu, W., and Hou, J. (2016). Thrombosis recanalization by paeoniflorin through the upregulation of urokinase-type plasminogen activator via the MAPK signaling pathway. Mol. Med. Rep. 13 (6), 4593–4598. doi:10.3892/mmr.2016.5146
Ye, J., Chen, H., Wang, K., Wang, Y., Ammerman, A., Awasthi, S., et al. (2024). Structural insights into vesicular monoamine storage and drug interactions. Nature 629, 235–243. doi:10.1038/s41586-024-07290-7
Yi, L. T., Li, J., Liu, B. B., and Li, C. F. (2013). Screening of the antidepressant-like effect of the traditional Chinese medicinal formula si-ni-san and their possible mechanism of action in mice. Pharmacogn. Res. 5 (1), 36–42. doi:10.4103/0974-8490.105647
Yu, J. B., Zhao, Z. X., Peng, R., Pan, L. B., Fu, J., Ma, S. R., et al. (2019). Gut microbiota-based pharmacokinetics and the antidepressant mechanism of paeoniflorin. Front. Pharmacol. 10, 268. doi:10.3389/fphar.2019.00268
Yu, W., Ilyas, I., Hu, X., Xu, S., and Yu, H. (2022). Therapeutic potential of paeoniflorin in atherosclerosis: a cellular action and mechanism-based perspective. Front. Immunol. 13, 1072007. doi:10.3389/fimmu.2022.1072007
Yuan, N., Li, X., Tang, K., Gan, H., Da, X., Hao, W., et al. (2022). Xiaoyaosan inhibits neuronal apoptosis by regulating the miR-200/NR3C1 signaling in the prefrontal cortex of chronically stressed rats. Phytomedicine 103, 154239. doi:10.1016/j.phymed.2022.154239
Zagrebelsky, M., and Korte, M. (2014). Form follows function: BDNF and its involvement in sculpting the function and structure of synapses. Neuropharmacology 76 Pt C, 628–638. doi:10.1016/j.neuropharm.2013.05.029
Zhai, A., Zhang, Z., and Kong, X. (2019). Paeoniflorin alleviates H(2)O(2)-Induced oxidative injury through down-regulation of MicroRNA-135a in HT-22 cells. Neurochem. Res. 44 (12), 2821–2831. doi:10.1007/s11064-019-02904-3
Zhang, X. M., and Gou, Y. L. (2018). Clinical observation of the effect of chaihu shugan decoction in adjuvant treatment of simple partial seizure combine depression. China J. Traditional Chin. Med. Pharm. 33 (07), 3183–3185.
Zhang, L., and Wei, W. (2020). Anti-inflammatory and immunoregulatory effects of paeoniflorin and total glucosides of paeony. Pharmacol. Ther. 207, 107452. doi:10.1016/j.pharmthera.2019.107452
Zhang, X. J., Chen, H. L., Li, Z., Zhang, H. Q., Xu, H. X., Sung, J. J., et al. (2009). Analgesic effect of paeoniflorin in rats with neonatal maternal separation-induced visceral hyperalgesia is mediated through adenosine A(1) receptor by inhibiting the extracellular signal-regulated protein kinase (ERK) pathway. Pharmacol. Biochem. Behav. 94 (1), 88–97. doi:10.1016/j.pbb.2009.07.013
Zhang, C., Du, F., Shi, M., Ye, R., Cheng, H., Han, J., et al. (2012). Ginsenoside Rd protects neurons against glutamate-induced excitotoxicity by inhibiting ca(2+) influx. Cell Mol. Neurobiol. 32 (1), 121–128. doi:10.1007/s10571-011-9742-x
Zhang, Y., Li, H., Huang, M., Huang, M., Chu, K., Xu, W., et al. (2015). Paeoniflorin, a monoterpene glycoside, protects the brain from cerebral ischemic injury via inhibition of apoptosis. Am. J. Chin. Med. 43 (3), 543–557. doi:10.1142/S0192415X15500342
Zhang, Y., Wang, L. L., Wu, Y., Wang, N., Wang, S. M., Zhang, B., et al. (2016). Paeoniflorin attenuates hippocampal damage in a rat model of vascular dementia. Exp. Ther. Med. 12 (6), 3729–3734. doi:10.3892/etm.2016.3849
Zhang, Y., Qiao, L., Xu, W., Wang, X., Li, H., Xu, W., et al. (2017). Paeoniflorin attenuates cerebral ischemia-induced injury by regulating Ca(2+)/CaMKII/CREB signaling pathway. Molecules 22 (3), 359. doi:10.3390/molecules22030359
Zhang, M., Zhang, Y., Sun, H., Ni, H., Sun, J., Yang, X., et al. (2021a). Sinisan protects primary hippocampal neurons against corticosterone by inhibiting autophagy via the PI3K/Akt/mTOR pathway. Front. Psychiatry 12, 627056. doi:10.3389/fpsyt.2021.627056
Zhang, S., Lu, Y., Chen, W., Shi, W., Zhao, Q., Zhao, J., et al. (2021b). Network pharmacology and experimental evidence: PI3K/AKT signaling pathway is involved in the antidepressive roles of chaihu shugan san. Drug Des. Devel Ther. 15, 3425–3441. doi:10.2147/DDDT.S315060
Zhang, S., Lu, Y., Shi, W., Ren, Y., Xiao, K., Chen, W., et al. (2022). SIRT1/FOXO1 axis-mediated hippocampal angiogenesis is involved in the antidepressant effect of chaihu shugan san. Drug Des. Devel Ther. 16, 2783–2801. doi:10.2147/DDDT.S370825
Zhang, M., Wang, M. X., Zhao, Q., Zhang, Y., Chen, Y. M., and Zhang, Y. Y. (2023). Clinical efficacy of xiaoyaosan combined with paroxetine tablets in treatment of migraine with liver depression and blood deficiency syndrome and depression based on transcranial doppler. Chin. J. Exp. Traditional Med. Formulae 29 (02), 113–118. doi:10.13422/j.cnki.syfjx.20230195
Zhang Qi, D. S.-T., Ji, B., and Feng, L. (2022). Clinical observation on sini powder combined with gan mai dazao decoction in the treatment of type 2 diabetes mellitus complicated with depression and anxiety. J. Guangzhou Univ. Traditional Chin. Med. 39 (04), 763–769. doi:10.13359/j.cnki.gzxbtcm.2022.04.006
Zhao, Z., Zhang, L., Guo, X. D., Cao, L. L., Xue, T. F., Zhao, X. J., et al. (2017). Rosiglitazone exerts an anti-depressive effect in unpredictable chronic mild-stress-induced depressive mice by maintaining essential neuron autophagy and inhibiting excessive astrocytic apoptosis. Front. Mol. Neurosci. 10, 293. doi:10.3389/fnmol.2017.00293
Zheng, M., Liu, C., Fan, Y., Shi, D., and Zhang, Y. (2016). Protective effects of paeoniflorin against MPP(+)-Induced neurotoxicity in PC12 cells. Neurochem. Res. 41 (6), 1323–1334. doi:10.1007/s11064-016-1834-z
Zheng, M., Liu, C., Fan, Y., Yan, P., Shi, D., and Zhang, Y. (2017). Neuroprotection by paeoniflorin in the MPTP mouse model of parkinson's disease. Neuropharmacology 116, 412–420. doi:10.1016/j.neuropharm.2017.01.009
Zhong, S. Z., Ge, Q. H., Li, Q., Qu, R., and Ma, S. P. (2009). Peoniflorin attentuates Abeta((1-42))-mediated neurotoxicity by regulating calcium homeostasis and ameliorating oxidative stress in hippocampus of rats. J. Neurol. Sci. 280 (1-2), 71–78. doi:10.1016/j.jns.2009.01.027
Zhong, X., Liu, M., Yao, W., Du, K., He, M., Jin, X., et al. (2019). Epigallocatechin-3-Gallate attenuates Microglial Infmammation andiNeurotoxicity by nuppressing thesActivation of aanonical andcNoncanonical Infnammasome viaiTLRviaF-κB Pathway. Mol. Nutr. Food Res. 63 (21), e1801230. doi:10.1002/mnfr.201801230
Zhou, C., Zhang, H., Qin, Y., Tian, T., Xu, B., Chen, J., et al. (2018). A systematic review and meta-analysis of deep brain stimulation in treatment-resistant depression. Prog. Neuropsychopharmacol. Biol. Psychiatry 82, 224–232. doi:10.1016/j.pnpbp.2017.11.012
Zhou, D., Zhang, S., Hu, L., Gu, Y. F., Cai, Y., Wu, D., et al. (2019). Inhibition of apoptosis signal-regulating kinase by paeoniflorin attenuates neuroinflammation and ameliorates neuropathic pain. J. Neuroinflammation 16 (1), 83. doi:10.1186/s12974-019-1476-6
Zhou, X. M., Liu, C. Y., Liu, Y. Y., Ma, Q. Y., Zhao, X., Jiang, Y. M., et al. (2021). Xiaoyaosan Alleviates Hippocampal Glutamate-Induced Toxicity in the CUMS Rats via NR2B and PI3K/Akt Signaling Pathway. Front. Pharmacol. 12, 586788. doi:10.3389/fphar.2021.586788
Zhu, X., Li, T., Hu, E., Duan, L., Zhang, C., Wang, Y., et al. (2021). Proteomics Study Reveals the Anti-Depressive Mechanisms and the Compatibility Advantage of Chaihu-Shugan-San in a Rat Model of Chronic Unpredictable Mild Stress. Front. Pharmacol. 12, 791097. doi:10.3389/fphar.2021.791097
Zhu, X., Ma, Q., Yang, F., Li, X., Liu, Y., Chen, J., et al. (2022). Xiaoyaosan Ameliorates Chronic Restraint Stress-Induced Depression-Like Phenotype by Suppressing A2AR Signaling in the Rat Striatum. Front. Pharmacol. 13, 897436. doi:10.3389/fphar.2022.897436
Keywords: paeoniflorin, medicine formula, depression, pharmacological mechanism, pharmacokinetics
Citation: Hou Y, Li H, Zhu L, Quan T, Feng X, Li Y, Bian Y and Wei Y (2025) Advances of paeoniflorin in depression: the molecular mechanism and formula application. Front. Pharmacol. 16:1614429. doi: 10.3389/fphar.2025.1614429
Received: 18 April 2025; Accepted: 20 August 2025;
Published: 03 September 2025.
Edited by:
Fabien Schultz, Bernhard Nocht Institute for Tropical Medicine (BNITM), GermanyReviewed by:
Juan Francisco Rodríguez-Landa, Universidad Veracruzana, MexicoDong Xu, Xijing Hospital, Air Force Medical University, China
Copyright © 2025 Hou, Li, Zhu, Quan, Feng, Li, Bian and Wei. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Yuan Bian, Ymlhbnl1YW54Y0AxNjMuY29t; Yuxun Wei, d2VpeXV4dW4zNTgzQDE2My5jb20=