 Ying Jia
Ying Jia Yuanyuan Zhang2
Yuanyuan Zhang2 Tengyu Zhao
Tengyu Zhao Haichun Zhou
Haichun Zhou- 1The Second Clinical Medical College, Heilongjiang University of Chinese Medicine, Harbin, China
- 2The Fourth Affiliated Hospital of Heilongjiang University of Chinese Medicine, Harbin, China
- 3Medical Affairs Department, Heilongjiang Province Second Hospital, Harbin, China
- 4School of Basic Medicine, Heilongjiang University Of Chinese Medicine, Harbin, China
Parkinson’s disease (PD) is a progressive neurodegenerative disorder associated with α-synuclein deposition and dopaminergic neuron degeneration. Recent studies have revealed a close correlation between gut microbiota dysbiosis and the pathogenesis and progression of PD. Gut microbiota may influence the disease through multiple pathways, including promoting α-synuclein pathology, disrupting the gut-brain barrier, and triggering inflammation and neuronal damage. Currently, drug treatments for PD primarily focus on compensating for dopaminergic neurotransmission deficits, delaying neuronal degeneration, and clearing abnormal protein aggregates. However, these drugs can only slow disease progression and are associated with significant adverse effects. In contrast, natural products exhibit distinct advantages in modulating PD pathological features by targeting the “gut microbiota-metabolite axis,” owing to their multi-target synergistic regulation and favorable safety profiles, making them an ideal strategy for PD intervention. Based on this, we provide a comprehensive review of natural products that regulate the gut microbiota, analyze their specific mechanisms, and offer novel insights into this approach and provide a theoretical foundation for developing safe and effective PD therapeutics.
1 Introduction
Parkinson’s disease (PD) is the second most common progressive neurodegenerative disorder, with a prevalence of approximately 1%–2% among individuals aged 65 years or older, second only to Alzheimer’s disease. PD is more prevalent in males than females, and its global incidence continues to rise with the aging population, posing a pressing public health challenge (Ben-Shlomo et al., 2024). The clinical manifestations of PD primarily consist of characteristic motor symptoms. These motor symptoms include resting tremor, rigidity, bradykinesia, and postural instability. Additionally, PD is accompanied by various non-motor symptoms. A major manifestation of non-motor symptoms is gastrointestinal (GI) dysfunction, such as nausea, vomiting, gastroparesis, delayed gastric emptying, and constipation. Notably, a strong correlation exists between non-motor and motor symptoms. For instance, constipation affects up to 80% of PD patients and may precede motor symptoms by several years (Fasano et al., 2015). The pathophysiology of PD remains incompletely understood but is thought to involve multiple factors, including genetics, environmental influences, and lifestyle (Morris et al., 2024). Pathologically, PD is primarily characterized by the loss of dopaminergic neurons in the substantia nigra pars compacta (SNpc) and the abnormal aggregation of α-synuclein (α-syn). Recent studies suggest that pathological α-syn may originate in the GI tract, drawing attention to the potential link between gut microbiota and PD. This discovery has spurred the concept of the “gut-brain axis.” (Menozzi et al., 2025).
The gut microbiota (GM), the most complex microbial ecosystem in the human body, comprises trillions of microorganisms, including bacteria, fungi, viruses, and protozoa. It plays a pivotal role in health and disease and is often referred to as the “second brain.” Alterations in the diversity and abundance of GM have been implicated in the pathogenesis of various neurological disorders. Recent studies have demonstrated dysbiosis of GM in PD patients and animal models. Furthermore, fecal microbiota transplantation (FMT), probiotics, and prebiotics exhibit significant neuroprotective effects in PD (Tan et al., 2022). This indicates the ability of GM as a new target for PD intervention.
Natural products from natural sources exhibit properties such as water solubility, membrane permeability/transportability, biomolecular compatibility, and stability (Atanasov et al., 2021; Shenvi, 2024). They serve as a critical source for drug discovery, with 84% of clinically used central nervous system (CNS) drugs directly or indirectly originating from natural products. Remarkably, over 400 CNS drugs have been developed from just 20 natural products (Bharate et al., 2018). Bridging modern and traditional medicine, natural products have been extensively investigated for their therapeutic potential in PD. Numerous studies have demonstrated that natural products can modulate PD pathogenesis through multiple pathways, including anti-inflammatory and antioxidant effects. Their multi-target mechanisms and low toxicity profiles offer novel therapeutic avenues for PD management. Given these advantages, exploring natural products targeting the GM for PD treatment holds significant promise. This review summarizes several common classes of natural products with potential anti-PD effects, including polyphenols, flavonoids, polysaccharides, terpenoids and glycosides, and alkaloids. The aim is to provide a reference for the clinical application of GM-based natural product therapies in PD.
2 The relationship between gut microbiota and PD
2.1 Basic composition and function of gut microbiota
The microorganisms in the GI tract are predominantly bacteria, mainly strictly anaerobic bacteria (Zhernakova et al., 2016). The microbiota initially colonizes the human intestinal tract at birth, exhibiting maternal characteristics. The primitive microbiome (observed at approximately 1 year of age) is relatively simple but becomes increasingly complex during development due to environmental and dietary influences. The abundance and diversity of strict anaerobes have increased. These microbial communities persist throughout the host’s lifespan, maintaining relative stability despite being influenced by multiple factors (Milani et al., 2017). Notably, substantial interindividual variability exists in microbial composition among healthy subjects, particularly in infants, with gradual convergence toward similar phyla over time (Sorboni et al., 2022).
The GM comprises over 1,000 bacterial species (Hou et al., 2022). More than 90% of these are dominated by the phyla Firmicutes and Bacteroidetes, with lesser proportions of Actinobacteria, Fusobacteria, Proteobacteria, and Verrucomicrobia (Qin et al., 2010). The GM is characterized by high diversity, encompassing a vast array of microbial species and populations. This diversity maintains the equilibrium between microbes and the host, serving as a critical determinant of intestinal health (Li Z. et al., 2022). Existing in a symbiotic relationship with humans, the GM not only contributes to the regulation of GI and immune functions but also influences nutrient metabolism, metabolic regulation, neurofunctional modulation, as well as drug metabolism and absorption. It plays an essential role in maintaining host health and physiological functions, including those of the CNS.
2.2 Correlation between PD and gut microbiota
Disruption of GM homeostasis has been associated with various diseases, particularly PD. Alterations in microbial diversity and abundance have been observed in both animal models and human subjects. Through a systematic review and subgroup meta-analysis of 14 studies involving 1,045 PD patients and 821 healthy controls from different countries, Bai et al. demonstrated significant differences in GM abundance at the phylum, family, and genus levels between PD patients and healthy controls (Bai et al., 2024). Specifically, PD patients exhibited reduced abundance in four microbial families (Lachnospiraceae, Prevotellaceae, Erysipelotrichaceae, and Faecalibacterium) and increased abundance in six families (Lactobacillaceae, Verrucomicrobiaceae, Bifidobacteriaceae, Rikenellaceae, Christensenellaceae, and Ruminococcaceae). Heravi et al. conducted a systematic review of 26 PD studies and reported elevated abundance of Akkermansia, Verrucomicrobiaceae, Lachnospiraceae, and Ruminococcaceae in PD patients, whereas Blautia, Coprococcus, Prevotellaceae, and Roseburia were more abundant in controls (Heravi et al., 2023). Furthermore, GM composition in PD patients varies depending on geographical location and dietary habits (Shu et al., 2025). Microbial profiles also differ across PD stages: early-stage PD is characterized by dominant genera such as Akkermansia, Alistipes, Anaerotruncus, Bilophila, Rikenellaceae, Verrucomicrobia, and Verrucomicrobiae, whereas late-stage PD is associated with increased Actinobacteriota and Erysipelotrichaceae (Jiang et al., 2025). Similar microbiota alterations have been observed in non-human primate models of PD (Yan et al., 2021). These findings demonstrate that PD exhibits a distinct pattern of gut dysbiosis, which varies with disease progression and is influenced by geographical and dietary factors. This robustly supports the notion that GM dysregulation serves as a critical pathological feature and potential therapeutic target in PD.
The GM plays a pivotal role in the pathogenesis, progression, and severity of PD, exhibiting significant correlations with interindividual variability in clinical manifestations. It is associated not only with motor symptoms but also closely linked to non-motor impairments. Bifidobacteriaceae is strongly correlated with the severity of gait disturbances and the worsening of hallucinations (Zuo et al., 2022). The relative abundance of Enterobacteriaceae demonstrates a positive correlation with the severity of postural instability and gait difficulty (Scheperjans et al., 2015). In contrast, Faecalibacterium exhibits anti-inflammatory effects that confer protection to the intestinal epithelial barrier. A reduction in Faecalibacterium coupled with an increase in Enterobacteriaceae abundance may compromise the intestinal epithelial barrier, thereby rendering the enteric nervous system (ENS) more susceptible to luminal pathogens. Prevotellaceae is associated with the severity of PD. Its reduction correlates with decreased secretion of ghrelin (Silva-Reis et al., 2022). A negative correlation exists between the abundance of Lachnospiraceae and PD duration. Meanwhile, Lachnospiraceae can contribute to GI dysmotility in PD patients (Unger et al., 2016). Lactobacillaceae may exacerbate motor dysfunction (van der Maden et al., 2025). Barichella’s study (Barichella et al., 2019) demonstrated that alterations in the abundance of Lactobacillaceae (increased) and Lachnospiraceae (decreased) constitute the most profound factors associated with PD clinical features (cognitive impairment, gait disturbance, and postural instability). Notably, Lachnospiraceae was significantly more abundant in patients with the non-tremor-dominant phenotype. Mehanna et al. (2023) reported reduced Bifidobacterium levels in the GM of tremor-dominant PD patients, decreased Lactobacillus in non-tremor-dominant phenotypes, and a marked increase in Bacteroides among mixed phenotypes. A prospective study by Cilia et al. (2021) involving 39 early-stage PD patients revealed that the abundance of Ruminococcaceae and Actinobacteria correlated with accelerated decline in global cognitive function. The composition of the GM is significantly associated with the onset, progression, severity, and diverse clinical manifestations of PD. Changes in the abundance of specific bacterial families directly impact the pathological processes and individualized clinical presentations of PD.
2.3 Microbial metabolites in PD
Microbial metabolites act as pivotal mediators within the microbiota-gut-brain axis (MGBA) (Zheng et al., 2021). These metabolites are broadly classified into three categories: (1) Diet-derived metabolites produced directly by microbial digestion, such as short-chain fatty acids (SCFAs) and indole derivatives; (2) De novo synthesized metabolites originating from the microbiota; (3) Host- or diet-derived compounds modified by microbial activity.
2.3.1 SCFAs
The production of SCFAs is primarily mediated by the fermentation of carbohydrates and dietary proteins by Bifidobacterium, Lactobacillus, Faecalibacterium, Lachnospiraceae, and Ruminococcaceae. SCFAs are a group of organic fatty acids, predominantly comprising acetate, propionate, butyrate, and β-hydroxybutyrate. SCFAs play a profound role in maintaining intestinal homeostasis, modulating metabolism, regulating immune function, and influencing neurological activity (Dalile et al., 2019; Qu et al., 2025). Notably, the concentration of SCFAs is significantly reduced in patients with PD (Unger et al., 2016). Butyrate not only acts on the colonic mucosa, but also induces hepcidin expression to maintain iron homeostasis and mitigates neuronal ferroptosis (Mayneris-Perxachs et al., 2022). As a histone deacetylase (HDAC) inhibitor, sodium butyrate regulates gene expression, protects dopaminergic neurons, and prevents motor deficits in PD models (Wang F. et al., 2024; Zhang L. Y. et al., 2024). Propionate modulates energy metabolism in the gut and exerts neuroprotective effects via choline-neuronal signaling. Additionally, it suppresses neurodegeneration through nonautonomous mechanisms (Wang C. et al., 2024).
2.3.2 Bile acids
Primary BAs are predominantly synthesized in the liver from cholesterol and include free bile acids (CA and CDCA), as well as their glycine (Gly)- or taurine (Tau)-conjugated forms, which exhibit enhanced water solubility (GCA, TCA, GCDCA, and TUDCA). Approximately 95% of BAs released into the large intestine via the gallbladder are reabsorbed into the liver through the ileal apical sodium-dependent bile acid transporter (ASBT). A minor fraction undergoes deconjugation and dehydroxylation by GM (e.g., Lactobacillus, Bifidobacterium, Bacteroides, Clostridium) possessing bile salt hydrolase (BSH) activity, thereby converting them into secondary BAs such as LCA, DCA, and UDCA (Wang S. et al., 2023).
An increased abundance of BA-synthesizing bacteria (Burkholderiales and Clostridium) was observed in the appendix tissues of PD patients, accompanied by significantly elevated levels of LCA and DCA in the ileum (Li P. et al., 2021). In a prospective study investigating the relationship between PD and plasma microbial metabolites, Zhao et al. identified altered abundances of CDCA and GCA, which contrasts with previous reports of elevated secondary BAs levels in PD patients’ blood samples (Zhao et al., 2025).
2.3.3 Metabolic product of tryptophan
2.3.3.1 Indoles and derivatives
Tryptophan, the sole essential amino acid containing an indole structure, is primarily obtained from protein-rich foods. In the gut, it is metabolized through three major pathways—the indole pathway, kynurenine (Kyn) pathway, and serotonin pathway—with the GM playing a pivotal role in each. Particularly in the indole pathway, GM drive the catabolism of tryptophan into indole and its derivatives. The structural diversity of indole derivatives arises from variations in bacterial-encoded glycosidases. Over 85 bacterial species, including Escherichia coli, Bacteroides, and Clostridium, produce indole; Clostridium generates indole-3-propionic acid (IPA) and tryptamine; Lactobacillus synthesizes indole-3-lactic acid (ILA) and indole-3-aldehyde (IAld); Peptostreptococcus produces indole-3-acrylic acid (IA); and Ruminococcus and Clostridium yield tryptamine, among others. As a class of profoundly bioactive molecules, indole and its derivatives play a complex and critical role in the pathological mechanisms and therapeutic research of PD. Their functions span multiple dimensions, including GM metabolism, intestinal epithelial barrier integrity, mitochondrial function regulation, neuroinflammatory modulation, neuroprotection of dopaminergic neurons, and α-syn pathological aggregation (Zhou et al., 2023).
Shao’s study demonstrated that plasma ILA levels were reduced in PD patients (Shao et al., 2021), accompanied by decreased abundance of ILA-producing Clostridium saccharolyticum in fecal samples. Additionally, alterations were observed in several other Clostridium and Bacteroides species involved in tryptophan metabolic enzymes (Bedarf et al., 2017). Chen’s investigation of 56 PD patients and 43 age-, sex-, and diet-matched healthy controls (family members or friends) revealed unchanged plasma tryptophan levels but significantly elevated IPA concentrations in PD patients (Chen S. J. et al., 2022). IPA functions as a HDAC inhibitor and exhibits positive correlation with pathological protein aggregation.
2.3.3.2 5-HT and kynurenine
5-HT is synthesized from its precursor tryptophan and is widely distributed in both the central and peripheral nervous systems. The intestine serves as the primary site (accounting for ∼95%) of peripheral 5-HT synthesis. Most 5-HT is released from enterochromaffin cells (ECCs), which directly interact with enteric nerve terminals, playing a pivotal role in normal intestinal function. Through Mendelian randomization analysis of the GM-PD relationship, Li et al. demonstrated that intestinally derived 5-HT is associated with an earlier age of PD onset (Jiang et al., 2023). Postmortem studies of PD patients revealed elevated 5-HT1A receptor availability in the anterior cingulate cortex (Gonzaga et al., 2025). The Kyn pathway, representing the dominant route (≥95%) of tryptophan metabolism, exhibits broad pleiotropic bioactivities including neuroprotective/neurotoxic effects. Its directional shift profoundly impacts neuronal function and survival. Metabolomic analyses by Chang et al. identified a transition from neuroprotective kynurenic acid (KA) to neurotoxic quinolinic acid (QA) in PD patients (Chang et al., 2018). Serum KYN levels were significantly reduced in PD patients, accompanied by decreased KA/KYN conversion rates and substantial alterations in tryptophan metabolic pathways (Fan et al., 2024). The GM suppresses Kyn production and promotes 5-HT synthesis. Akkermansia modulates tryptophan metabolism, thereby shifting metabolic flux from Kyn to 5-HT and enhancing 5-HT activity (Wang J. et al., 2021; Zhang L. et al., 2023; Pan et al., 2025).
2.3.4 LPS
Lipopolysaccharide (LPS), a major component of the outer membrane of Gram-negative bacteria. The intestine serves as the primary source of LPS. Under physiological conditions, a healthy intestinal barrier (IB) confines the majority of LPS within the intestinal lumen. However, when the IB is compromised or dysbiosis occurs, LPS translocates into the bloodstream in large quantities, entering the portal circulation and triggering metabolic endotoxemia (Popovic et al., 2023; Gorecki et al., 2025). LPS and its associated inflammatory response compromise the blood-brain barrier (BBB), increasing permeability (Ma and Lieberman, 2024). This enables the translocation of LPS and inflammatory mediators into the CNS, subsequently activating microglia and astrocytes. Such activation propagates a neuroinflammatory cascade that promotes α-syn pathology, impairs dopaminergic neurons, and disrupts neuronal homeostasis (Huang et al., 2024). Additionally, LPS exerts direct neurotoxicity, inducing neuronal apoptosis or necrosis.
In patients with PD, the abundance of Gram-negative bacteria producing LPS is elevated, and this abundance positively correlates with the severity of motor symptoms. Studies have demonstrated increased levels of LPS-binding protein and soluble CD14 (proteins associated with the LPS/TLR4 signaling pathway) in PD patients, which correlate with disease progression. Administration of LPS into the SN or the GI tract of rodents successfully induces PD-like motor symptoms and pathological changes (Gorecki et al., 2019).
2.3.5 Gasotransmitters
Gasotransmitters, primarily produced by the GM, serve as pivotal signaling mediators and effector molecules in vivo. Sulfate-reducing bacteria generate substantial amounts of hydrogen sulfide (H2S), while other bacteria (e.g., certain Clostridium and Bacteroide) also produce H2S via enzymatic reactions. Lactobacillus, Bifidobacterium, and E.coli contribute to nitric oxide (NO) production. H2S exhibits a dual role. At physiological concentrations, it demonstrates neuroprotective properties, such as suppressing reactive oxygen species (ROS) generation, inhibiting microglia (MG) activation, and restoring mitochondrial function (Wang L. et al., 2023). Conversely, excessive H2S elevates ROS levels, disrupts barrier integrity, triggers inflammation, and promotes α-Syn aggregation (Murros, 2022). A meta-analysis revealed a significant increase in H2S-producing bacteria in PD patients (Marzouk et al., 2025). Similarly, NO concentration critically determines whether it induces or suppresses inducible nitric oxide synthase (iNOS) expression. iNOS participates in defense mechanisms involving immune cells and oxidative responses (Picón-Pagès et al., 2019).
2.4 Pathogenic mechanisms of gut microbiota in PD
Mounting evidence indicates the existence of a bidirectional gut-brain communication system between the CNS and the GM, termed the MGBA (Ohara and Hsiao, 2025). The microbial community can interact with the immune system, neuroendocrine pathways, and the sympathetic nervous systems, thereby modulating brain function, ultimately contributing to the pathogenesis and progression of PD (Zhang X. et al., 2023). Conversely, the CNS can induce alterations in intestinal motility, secretion, permeability, and inflammation through autonomic interactions. The specific mechanism diagram is shown in the figure (Figure1).
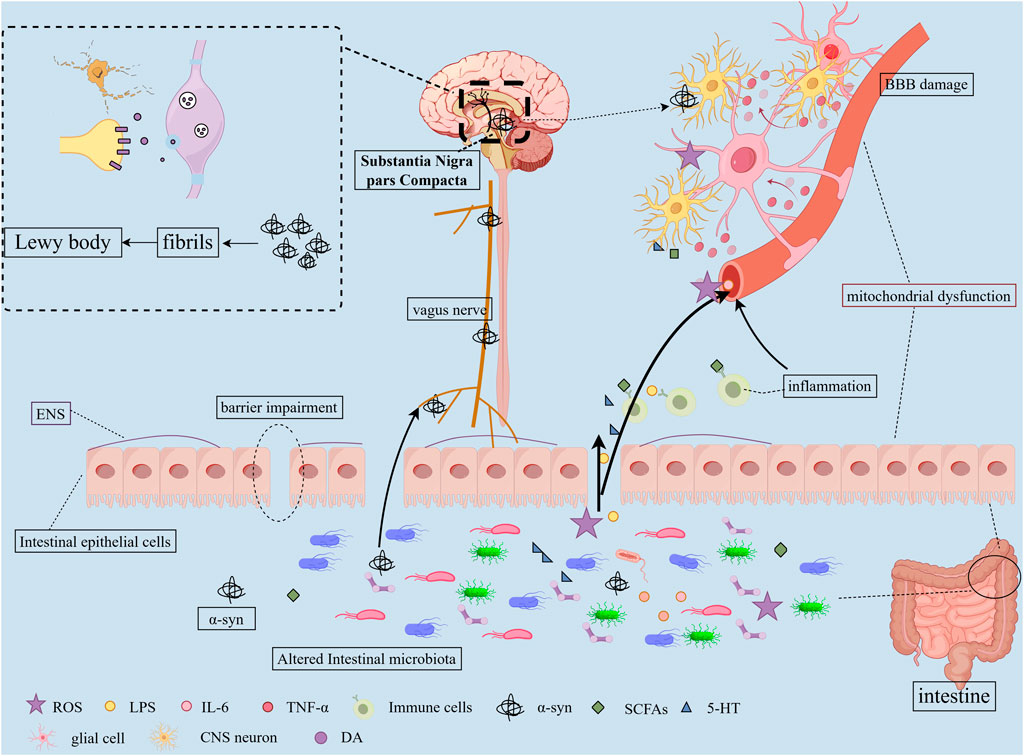
Figure 1. Pathogenic Mechanisms of Gut Microbiota in PD. 1) Dysbiosis of the GM alters microbial metabolites and compromises the intestinal barrier function, resulting in “leaky gut” syndrome; 2) These metabolites and harmful substances enter systemic circulation, damaging BBB structure and function while activating glial cells and triggering neuroinflammation; 3) Gut-derived α-syn may propagate to the CNS via vagal nerve pathways, ultimately forming fibrillary aggregates in the SNpc; 4) Homeostatic imbalance induces chronic low-grade inflammatory states in both CNS and peripheral systems; 5) Microbial-derived toxins contribute to mitochondrial dysfunction; 6) These cascades ultimately lead to dopaminergic neuron loss in the SN and α-syn aggregation.
2.4.1 Pathological α-synuclein production
α-Syn is primarily localized to presynaptic terminals in the CNS. Under physiological conditions, α-syn exists in a dynamic equilibrium between unfolded monomers and α-helix-folded tetramers, exhibiting low aggregation propensity and playing a crucial role in neurotransmission (Lashuel et al., 2013). Under pathological conditions, misfolded α-syn aggregates and further matures into fibrillar oligomers, disrupting normal cellular processes and contributing to neuronal death. Notably, phosphorylated α-syn levels were found to be significantly elevated in the GI up to 20 years before PD diagnosis (Hilton et al., 2014).
The GM is closely associated with pathological α-syn (Fang et al., 2024). In an eQTL analysis, it was found that the increased abundance of opportunistic bacteria in the intestines of PD patients was directly correlated with genetic variations in SNCA, the gene encoding α-syn (Wallen et al., 2021). α-syn-overexpressing (ASO) mice treated with broad-spectrum antibiotics did not develop motor dysfunction or exhibit α-syn aggregates in the brain. Furthermore, FMT from healthy donors ameliorated motor deficits in ASO mice, whereas colonization with microbiota from PD patients exacerbated the impairments (Sampson et al., 2016). Extracellular vesicles secreted by E.coli can deliver curli to colonic mucosal epithelial cells, upregulating DAPK1 expression and triggering α-syn aggregation (Liang et al., 2023). Enterobacteriaceae promote nitrite production. This mediates the oxidation of Fe2+ to Fe3+ and facilitates dopamine oxidation to o-quinone. Consequently, these processes induce misfolding and aggregation of α-syn in enteroendocrine cells (Ortiz de Ora et al., 2024). HpmA derived from Proteus mirabilis contributes to α-syn oligomerization and membrane pore formation (Huh et al., 2023a). Akkermansia muciniphila induces α-syn aggregation in enteroendocrine cells (EECs) (Amorim Neto et al., 2022).
Both LCA and DCA were found to markedly accelerate α-syn aggregation, exacerbating neurotoxicity while reducing the lag phase by 75% and 30%, respectively (Kaur et al., 2024). CDCA effectively reduces α-syn expression in PD model mice. Additionally, it alleviates motor deficits and anxiety-like behaviors in these models (Mehreen et al., 2025). Overexpression of α-syn leads to age-dependent dysregulation of the GM (Singh et al., 2023). Animal studies further demonstrate that α-syn drives ecological imbalance in the murine microbiome (Sampson et al., 2025).
2.4.2 Impairment of barrier function
The intestinal barrier (IB) and blood-brain barrier (BBB) are critical physiological barriers that maintain microenvironmental homeostasis (Aburto and Cryan, 2024). Dysfunctions of both barriers occur in PD patients and animal models. In PD patients, colonic ZO-1 expression is reduced (Liao et al., 2024). A PD monkey model shows significantly decreased occludin and ZO-1 expression in the duodenum, ileum, colon, and rectum, with concurrent intestinal mucosal damage (Zhang Y. et al., 2025). Postmortem PD studies reveal vascular changes and increased BBB permeability (Lau et al., 2024). Notably, Elabi et al. first observed impaired BBB integrity in an ASO mouse model. This impairment features increased extravascular fibrinogen, pericyte activation, and basilar membrane Col IV reduction (Elabi et al., 2021).
GM and metabolites modulate both IB and BBB functions. Germ-free mice exhibit enhanced BBB permeability and impaired tight junctions (Braniste et al., 2014). Lactobacillus rhamnosus (with strong adhesive properties) physically regulates goblet cells and mucus layers, restoring IB function via anti-inflammatory and cytoprotective effects (Martín et al., 2019). Propionate reverses antibiotic-induced BBB permeability and protects against BBB oxidative stress (Hoyles et al., 2018; Chenghan et al., 2025). It also upregulates tight junction proteins via AKT signaling, improving intestinal barrier function and motility (Huang et al., 2021). Additionally, propionate may exert neuroprotection through the FFAR3-GLP-1 axis in enteric neurons (Hou et al., 2021). Butyrate significantly increases occludin and claudin-5 expression in colonic tissue and the prefrontal cortex, repairing IB and BBB damage (Wang et al., 2025d). LPS induces pyroptosis in brain endothelial cells via the Casp11/Cd14-GSDMD pathway, causing BBB ultrastructural changes (Wei et al., 2024; Wei et al., 2025). Gut-derived α-syn impairs IB through caspase-1 inflammasome signaling and disrupts BBB via pericyte interactions (Dohgu et al., 2019; Pellegrini et al., 2022). IPA enhances IB by 1) reducing paracellular permeability by increasing transepithelial electrical resistance; 2) promoting mucin/goblet cell secretion; and 3) thickening mucus layers (Li J. et al., 2021). IAA activates AHR to promote goblet cell differentiation and mucus secretion (Cui W. et al., 2024). Indole supplementation repairs barrier damage in tryptophan-deficient mice. Tryptamine increases colonic ion flux and fluid secretion. This is supported by accelerated GI transit in germ-free mice colonized with tryptamine-producing engineered Bacteroides (Bhattarai et al., 2018). 5-HT regulates intestinal stem cells and mitigates IB damage via the Htr4-Kit-Wnt3 axis (Zhang D. et al., 2025).
2.4.3 Immunomodulation
The immune response drives PD pathophysiology. GM alterations trigger intestinal inflammation, damage the IB, and activate mucosal immune cells. These changes promote pro-inflammatory factor release alongside α-syn misfolding/aggregation. Such pathological products circulate to the brain, activating central immune cells and inducing neuroinflammation. Ultimately, this cascade causes dopaminergic neuron loss and PD progression. In PD patients, CD3+ T cell density in the SNpc negatively correlates with disease duration. Iba-1 expression is significantly elevated (Backman et al., 2025). Microglial and astrocyte activation is prominent (Kam et al., 2020). Peripheral T lymphocytes show increased PD-1 expression, which positively correlates with IFN-γ levels in CD4+ T cells (Zhang Z. L. et al., 2025). CD4+ T cell infiltration rises in cerebrospinal fluid and SNpc (observed in both patients and animal models) (Sun et al., 2024b). Clinical evidence further links gut immunity to PD. Chronic appendicitis-like lesions occur in 53% of PD patients (Chen et al., 2021). Appendectomy reduces PD risk (Nakahara et al., 2023). Inflammatory bowel disease (IBD) increases PD risk (Espinosa-Oliva et al., 2024; Kars et al., 2024). Anti-TNF-α therapy in IBD patients lowers PD risk by 78% (Peter et al., 2018).
Altered GM frequently coincides with elevated inflammatory cytokines. Specifically, Verrucomicrobia abundance correlates with plasma IFN-γ levels, whereas Bacteroides associates with TNF-α concentrations (Lin et al., 2019). Akkermansia muciniphila modulates SCFAs, ameliorating neuroinflammation and promoting hippocampal neurogenesis in PD models (Huang, 2024). Alistipes colonization regulates lipid metabolism and suppresses inflammatory mediators via outer membrane vesicles (Older et al., 2025). LPS crosses the BBB, activating MG through TLR4/MyD88/NF-κB signaling. FMT counteracts this by reducing LPS levels and inhibiting TLR4/MyD88/NF-κB in both gut and brain, attenuating inflammation (Zhao et al., 2021). Notably, LPS-stimulated macrophages secrete TNF-α, correlating with motor symptom severity. Concurrently, SCFAs associate with CD3+ T cells and TLR4+ cells while downregulating regulatory T cells (Perez-Pardo et al., 2019). Butyrate activates FFAR3, modulating sympathetic signaling and enteric neurogenesis. This consequently influences energy metabolism and inflammatory responses (Kalkan et al., 2025). BA mediate gut-brain communication via the FXR/TGR5-GLP-1 and FXR-FGF15/19 pathways, exerting neuroprotective effects (Kiriyama and Nochi, 2019). TUDCA activates TGR5, regulating AKT/NF-κB, AMPK/mTOR, and Pink1/Parkin pathways to suppress microglial activation and enhance autophagy (Ni et al., 2025). Indole derivatives act as core AhR ligands. AhR activation enhances intestinal barrier function, promotes IL-22 secretion, inhibits M1 macrophage polarization, and modulates gut immunity (Ma et al., 2020). Paradoxically, it may disrupt BBB integrity and exacerbate neuroinflammation (Salminen, 2023). IPA suppresses NF-κB via the PXR, reducing proinflammatory factors while exhibiting antioxidant activity (Zhao et al., 2022; Niu et al., 2025). Furthermore, IPA ameliorates dopaminergic dysfunction by inhibiting the IL-13Rα/JAK1-STAT6 signaling and reducing enteric glial cell gliosis (Shang et al., 2025).
2.4.4 Vagus nerve
The VN serves as the primary neural pathway connecting the gut and the brain, representing the fastest and most direct route. Although the VN is widely distributed throughout the gastrointestinal tract, it does not directly interact with the GM. Instead, communication occurs through specialized structures called neuropods on enteroendocrine cells (Peterson, 2020).
Epidemiological and histopathological findings suggest that pathological α-Syn may propagate from the gut to the brain via the VN, thereby increasing the risk of PD (Braak et al., 2003; Kim et al., 2019; Borghammer et al., 2022; Yang Z. X. et al., 2024). In mice inoculated with α-Syn PFFs, Lewy body (LB)-like aggregates formed in the dorsal motor nucleus of the VN and the myenteric plexus at 45 days and 12 months post-inoculation. However, these aggregates were completely absent when the VN was severed prior to inoculation (Uemura et al., 2019). Regarding the mechanism of α-Syn transport to the VN, Rashmi Chandra et al. provided evidence that intestinal mucosal cells facilitate this translocation (Chandra et al., 2023). High-resolution ultrasonography has revealed VN atrophy in PD patients (Dong S. et al., 2024). VN stimulation, as an adjunctive therapy, has been shown to ameliorate PD-related pathology and motor dysfunction (Evancho et al., 2024; Zhang H. et al., 2025).
Enterococcus faecalis can influence brain function in PD mice via the vagal pathway, mitigating BBB damage and neuroinflammation. However, this protective effect is attenuated upon subdiaphragmatic vagotomy (Shao et al., 2025). Lactobacillus rhamnosus modulates the expression of GABA receptors in the mouse brain and affects neuro-emotional behaviors, yet these changes are blocked by vagotomy (Bravo et al., 2011). LPS can activate vagal afferents through TLR4 on the nodose ganglion (Hosoi et al., 2005). SCFAs directly stimulate vagal afferent activity (Pang et al., 2026).
2.4.5 Mitochondrial dysfunction and oxidative stress
Mitochondrial dysfunction is a central mechanism in PD pathogenesis. As critical energy suppliers, mitochondria sustain dopaminergic neuron function. Impairment of the mitochondrial electron transport chain (ETC.) elevates ROS production. Excessive ROS damages mitochondrial components—including membrane lipids, proteins, and DNA (mtDNA)—compromising mitochondrial integrity. Concurrently, ROS attacks nuclear DNA, causing widespread cellular damage (Islam et al., 2018). Elevated oxidative stress markers confirm this pathology in PD patients (Korczowska-Łącka et al., 2023). Mitochondria also regulate Ca2+ uptake to maintain neuronal calcium homeostasis. Mitochondrial quality control (MQC) mechanisms sustain functional mitochondria through mitophagy, fusion/fission dynamics, and biogenesis. Dysregulation of these mechanisms increases ROS and reduces ATP. Notably, mutations in PINK1 and Parkin—key autophagy genes—are linked to early-onset PD (Samaranch et al., 2010). Reduced nigral PGC-1α expression in PD contrasts with its neuroprotective effects when activated (Mohammed et al., 2024; Ma et al., 2025). Both sporadic and familial PD exhibit mitochondrial dysfunction (Banerjee et al., 2009). Aberrant expression of mitochondrial-associated genes elevates PD risk (Wang et al., 2025e). Therapeutic targeting of oxidative stress and mitochondrial defects shows neuroprotective efficacy in PD models (Li X. et al., 2024). Clinically, blood mitochondrial DNA copy number (mtDNA-CN) may serve as a diagnostic and prognostic biomarker (Jo et al., 2025).
GM couples with mitochondrial dynamics (Zhu et al., 2022). SCFAs traverse the BBB, modulating mitochondrial metabolism (Chen and Vitetta, 2020). Butyrate curbs ROS/RNS in immune cells (Kalyanaraman et al., 2024); propionate reinforces mitochondrial integrity in Tregs (Duscha et al., 2020). LPS drives mitochondrial ROS accumulation, fueling inflammation. B. animalis ssp. Activates nigral PGC-1α to suppress neuroinflammation (Dong Y. et al., 2024). Microbial toxins target mitochondria: Clostridium’s TcdB inhibits mitoK+ channels (apoptosis induction) (Matarrese et al., 2007); H. pylori VacA triggers mitochondrial fission (Foo et al., 2010). Pathogenic α-syn disrupts mitochondria by:1) Inner membrane binding that inhibits MC I, driving dysfunction and mitophagy in A53T models; 3) disrupting morphology through autophagic flux in SH-SY5Y cells (Plotegher et al., 2014; Du et al., 2020). α-Syn PFFs induce dopaminergic oxidative stress via p-NMDAR2B/Nur77 (Lai et al., 2025). 5-HT elevates antioxidant defenses, reduces ROS, and enhances mitobiogenesis via SIRT1-PGC-1α (Fanibunda et al., 2019).
3 The related natural products regulating the PD gut microbiota
This review first delineates the most recent insights into the role of GM dysbiosis and the gut-brain axis in the pathogenesis of PD. Subsequently, we summarize and analyze experimental studies, and clinical evidence concerning the application of natural products for PD treatment via GM modulation over the past decade (Table 1). We synthesize the efficacy and underlying mechanisms by which these natural products ameliorate PD symptoms through restoring gut microbial balance and influencing gut-brain communication pathways (Figure 2). Representative chemical structures of key bioactive natural compounds are illustrated in Figure 3.
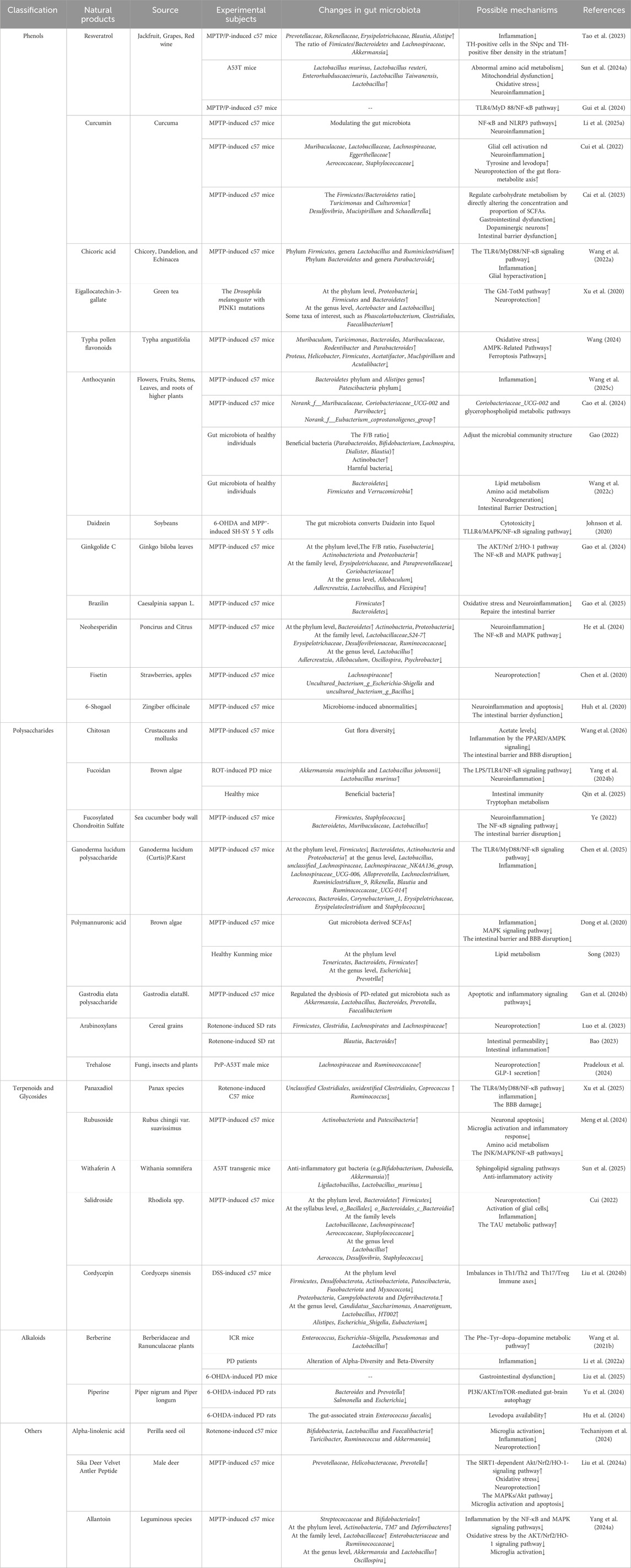
Table 1. Summary of recent studies on the Related Natural Products Regulating the Gut Microbiota in PD.
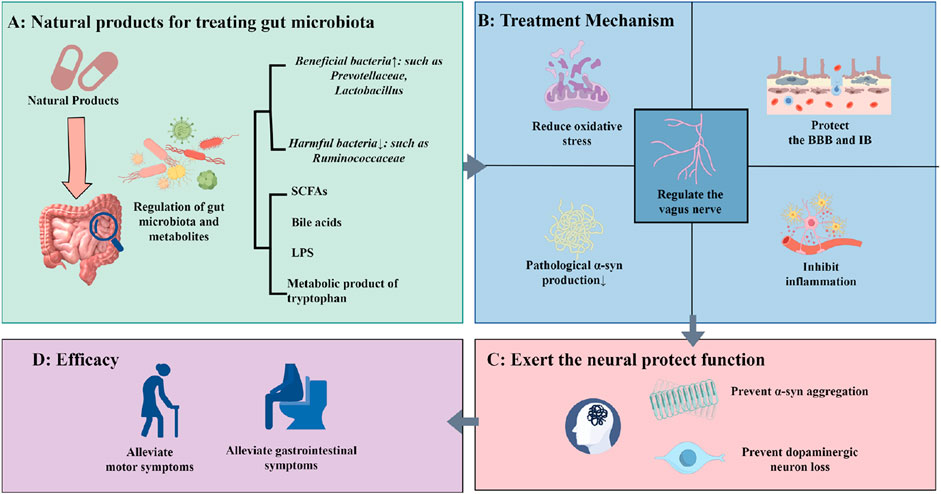
Figure 2. Natural Products Delay the Progression of PD via Gut Microbiota. (A) Regulation of gut microbiota and microbial metabolites by natural products; (B) Therapeutic mechanisms of natural products in PD treatment through gut microbiota modulation; (C) Neuroprotective effects mediated by attenuation of pathological changes in the PD brain; (D) Improvement of clinical PD symptoms by natural products.

Figure 3. Chemical structure of these natural products.
3.1 Phenols
3.1.1 Polyphenols
3.1.1.1 Resveratrol
Resveratrol (RSV), a naturally occurring polyphenolic compound, is predominantly found in jackfruit, grapes, red wine, and other sources. RSV supplementation has been demonstrated to elevate Akkermansia, Bacteroides, and Blautia abundance while increasing microbial metabolite 4-HPA and activating SIRT1 signaling (Wang P. et al., 2025). SIRT1 modulates oxidative stress and mitochondrial function to counteract neurodegenerative processes, thereby protecting dopaminergic neurons from α-syn-induced toxicity and apoptosis (Thapa et al., 2024). Notably, SIRT1 reduction occurs in both PD patients and models (Singh et al., 2017).
RSV significantly restores the levels of beneficial gut bacteria/metabolites, ameliorating PD-related pathological progression and symptoms. Preclinical studies demonstrate RSV’s dual regulatory actions: it modulates dopamine receptor transcription and calcium homeostasis-related genes while downregulating ZO-1/occludin expression. This molecular remodeling reconstructs the intestinal barrier. Consequently, colonic α-syn accumulation diminishes with concomitant reduction of low-grade inflammation (Sun et al., 2024a). Furthermore, FMT from RSV-treated mice markedly alleviates PD phenotypes in recipient mice and increases TH-positive cells in the SN and striatal TH-positive fiber density (Tao et al., 2023). Additionally, RSV reduces LPS, inhibits the TLR4/MyD88/NF-κB signaling pathway-mediated inflammation, repairs the IB, and modulates the gut-brain axis to exert neuroprotective effects (Gui et al., 2024). Through these multifaceted mechanisms targeting the microbiota-gut-brain axis, RSV demonstrates significant therapeutic potential for PD treatment.
3.1.1.2 Curcumin
Curcumin (Cur) is a bioactive low-molecular-weight polyphenolic compound primarily extracted from the rhizomes of Curcuma longa. It exhibits broad pharmacological effects, including anti-inflammatory, antioxidant, and neuroprotective properties (Wang G. et al., 2025). Clinical studies confirm its neuroprotective efficacy (Chang, 2025). Crucially, Cui et al. identify GM and metabolites as primary mediators of these effects (Cui et al., 2022). Cur alleviates gastrointestinal dysmotility and motor deficits in PD by modulating GM dysbiosis. This modulation suppresses inflammatory responses, leading to three key outcomes: 1) reduced α-syn aggregation; 2) enhanced IB integrity; 3) protection of dopaminergic neurons in SN. At the molecular level, preclinical studies demonstrate Cur’s activation of the SIRT1/NRF2 pathway. This activation inhibits AIM2-mediated pyroptosis and caspase-1. Simultaneously, Cur suppresses NF-κB and NLRP3 pathways. Consequently, proinflammatory cytokine production decreases while TH levels elevate in the SN and striatum (Zhong et al., 2022; Li W. et al., 2025). Furthermore, Cur enriches SCFA-producing genera like Turicimonas and Culturomica (Cai et al., 2023). This enrichment enhances carbohydrate metabolism and gastrointestinal motility. PD progression is thereby delayed. Additionally, Cur scavenges free radicals effectively. It concurrently suppresses neuroinflammation and oxidative stress while restoring mitochondrial function. These findings validate Cur’s multi-target therapeutic action against PD via the GMBA.
3.1.1.3 Chicoric acid
Chicoric acid (CA) is a core polyphenolic active component found in medicinal plants such as chicory, dandelion, and Echinacea. It exhibits anti-inflammatory and neuroprotective pharmacological effects and has a long history of clinical application (Yang et al., 2022). Recent studies have demonstrated that CA can modulate neuroinflammation and neurodegenerative pathology in PD through multi-target regulation of the gut-brain axis (Wang et al., 2022b). Specifically, Wang et al. confirmed that oral administration of CA reshapes GM balance and elevates SCFAs levels. Furthermore, CA restores intestinal epithelial integrity and suppresses the TLR4/MyD88/NF-κB inflammatory signaling pathway (Wang et al., 2022a). This cascade of actions blocks the gut-brain axis inflammatory cascade, thereby significantly improving motor dysfunction in PD.
3.1.1.4 Eigallocatechin-3-gallate
Eigallocatechin-3-gallate (EGCG), primarily derived from green tea, demonstrates neuroprotective properties. Epidemiological evidence indicates that consuming over 600 mL of green tea daily correlates with reduced PD risk (Hu et al., 2007). Preclinical PD studies have established EGCG’s efficacy in mitigating neurodegeneration in the SN by targeting pathogenic protein misfolding and aggregation (Gonçalves et al., 2021; Sergi, 2022). Notably, EGCG’s protective mechanisms involve modulation of the GM. As demonstrated by Xu et al., EGCG restructures GM composition and ameliorates neuronal/mitochondrial morphology in Drosophila PD models via the TotM pathway—a mechanism associated with mitochondrial function (Xu et al., 2020). Additionally, EGCG repairs compromised IB integrity and attenuates inflammatory responses (Li S. et al., 2024). Crucially, GM metabolizes EGCG into bioactive metabolites with enhanced BBB permeability (Pervin et al., 2019). Collectively, these findings indicate that GM serves as a pivotal hub mediating EGCG’s neuroprotective effects through metabolic conversion.
3.1.2 Flavonoids
3.1.2.1 Typha pollen flavonoids
Typhae Pollen Flavonoids (TPF), natural compounds extracted from the pollen of the traditional medicine Typha angustifolia, represent the core bioactive constituents responsible for its pharmacological effects in promoting blood circulation, resolving stasis, and hemostasis. In MPTP-induced PD models, TPF significantly restructures GM composition and ameliorate dysbiosis, evidenced by reduced pathogenic bacterial abundance and increased beneficial taxa. The regulatory mechanisms involve three primary pathways: 1) IB restoration: upregulation of tight junction protein occludin expression; 2) Microbial metabolite modulation: Enhanced SCFAs production activates the AMPK signaling pathway, thereby suppressing neuronal ferroptosis; 3) Amino acid metabolic reprogramming. Collectively, these effects attenuate oxidative stress in the SN and inhibit pathological α-syn aggregation (Wang, 2024). Our findings demonstrate that TAF protects dopaminergic neurons via the MGBA, ultimately improving motor dysfunction in PD mice.
3.1.2.2 Anthocyanin
Anthocyanins, a class of water-soluble flavonoid pigments ubiquitously present in vacuoles of flowers, fruits, stems, leaves, and roots of higher plants, are renowned for their antioxidant properties. Preclinical studies indicate that anthocyanins confer neuroprotective effects against PD (Zaa et al., 2023). In MPTP-induced PD models, anthocyanins paradoxically reduce GM α-diversity while modulating the Firmicutes/Bacteroidetes ratio, ameliorating microbial dysbiosis (Gao, 2022; Wang W. et al., 2022; Cao et al., 2024). This restructuring attenuates dopaminergic neuron damage and motor deficits via the gut-brain axis. Mechanistically, anthocyanins: downregulate pro-inflammatory cytokines (TNF-α, IL-1β, IL-6) and increase TH-positive neurons in the SN (Wang Y. et al., 2025). In addition, anthocyanins activate the Nrf2/GPX7 signaling pathway to mitigate mitochondrial oxidative stress, thereby restoring mitochondrial function and reducing neurodegeneration (Li X. N. et al., 2025).
3.1.2.3 Daidzein
Daidzein (DAI), dietary phytochemicals naturally occurring in soybeans and their derivatives, demonstrate ameliorative effects in neurodegenerative disorders. Following GM-mediated conversion to equol (EQL)—a metabolite with superior BBB permeability—this active compound accumulates in the CNS (Shimazu et al., 2021). Studies confirm that EQL significantly attenuates neurotoxicity and inflammation in both cellular and animal models of neurotoxin-induced PD (Johnson et al., 2020). Mechanistically, EQL suppresses microglial hyperactivation via inhibiting the TLR4/MAPK/NF-κB inflammatory cascade. Meanwhile, EQL enhances production of neurotrophic factors (e.g., BDNF, GDNF) in astrocytes. These coordinated actions collectively reduce neuronal apoptosis and confer resistance against inflammatory damage (Subedi et al., 2017). Our findings establish that the neuroprotection afforded by DAI critically depends on the anti-inflammatory mechanisms of its GM-derived metabolite EQL.
3.1.2.4 Ginkgolide C
Ginkgolide C (GC), a terpene lactone isolated from the traditional herbal medicine Ginkgo biloba leaves, exhibits anti-inflammatory, antioxidant, and free radical-scavenging properties with demonstrated BBB permeability for neuroprotection. Gao et al. revealed that oral GC administration significantly ameliorates MPTP-induced neurodegeneration in PD models, mechanistically linked to GM modulation—particularly through normalization of the Firmicutes/Bacteroidetes ratio (Gao et al., 2024). Further evidence indicates GC preserves IB integrity via activating the AKT/Nrf2/HO-1 antioxidant axis and suppresses NF-κB/MAPK inflammatory signaling cascades. Collectively, these actions attenuate gut-brain axis dysregulation, establishing GC as a dual-target therapeutic agent that concurrently mitigates oxidative stress and neuroinflammation to delay PD progression (Xu D. et al., 2022).
3.1.2.5 Brazilin
Brazilin is a characteristic homoisoflavonoid compound derived from the heartwood of Caesalpinia sappan L. (Leguminosae), mediates this plant’s traditional medicinal effects in promoting blood circulation, resolving stasis, and alleviating pain/swelling. Recent studies in PD have demonstrated that brazilin inhibits α-syn aggregation and ameliorates motor dysfunction. These findings position brazilin as a novel therapeutic candidate for PD (Cui Z. et al., 2024). Gao et al. reported that brazilin modulates GM to enhance butyrate production. This microbial metabolite upregulates the expression of ZO-1 and occludin to mitigate IB damage. Simultaneously, butyrate attenuates systemic inflammatorion and dopaminergic neurodegeneration (Gao et al., 2025). Butyrate has been shown to inhibit M1-like polarization of colonic macrophages and the production of pro-inflammatory cytokines, including TNF-α and IL-1β, via the PPARα-CYP4X1 axis (Chen Y. et al., 2022).
3.1.2.6 Neohesperidin
Neohesperidin (Neo), a natural dihydroflavonoid glycoside primarily derived from Poncirus and Citrus genera (Rutaceae family), exhibits notable anti-inflammatory activity. In an MPTP-induced PD mouse model, He et al. demonstrated that oral Neo administration upregulates the abundance of Prevotella and Bacteroides (He et al., 2024). Prevotella promotes dietary fiber fermentation to produce SCFAs, whereas Bacteroides enhances mucus layer integrity—mechanisms that collectively maintain IB homeostasis (Yu et al., 2023). Molecular analyses further confirmed SCFAs upregulate ZO-1 and occludin expression. These findings indicate Neo restores IB integrity via GM modulation. Additionally, their study revealed Neo suppresses NF-κB and MAPK signaling pathways, thereby reducing pro-inflammatory cytokines (TNF-α, IL-1β) in both intestinal and cerebral tissues. This dual inhibition protects dopaminergic neurons in the SN and mitigates neurodegeneration. Collectively, Neo exerts neuroprotective effects through synergistic modulation of GM and suppression of inflammatory cascades.
3.1.2.7 Fisetin
Fisetin, a naturally occurring flavonoid abundant in fruits (e.g., strawberries, apples), exhibits multi-targeted neuroprotective properties encompassing antioxidant, anti-inflammatory, and immunomodulatory activities (Calabrese et al., 2025). Recent studies have revealed an association between fisetin’s neuroprotective properties and the GM. Chen et al. demonstrated that fisetin intervention in MPTP-induced mice resulted in a significant increase in the abundance of Lachnospiraceae, accompanied by marked reductions in Escherichia-Shigella and Bacillus abundance (Chen et al., 2020). Lachnospiraceae promotes butyrate production, which exerts anti-inflammatory effects (Guo et al., 2024). These findings suggest that fisetin alleviates neurodegeneration and delays PD progression by remodeling the GM.
3.1.3 Alkylphenols
3.1.3.1 6-shogaol
6-Shogaol (6S), primarily derived from the dried rhizomes of the perennial herb Zingiber officinale, exhibits anti-inflammatory, antioxidant, barrier-repairing, and neuroprotective activities. Numerous studies have demonstrated the significant potential of 6S in the treatment of neurodegenerative diseases. It has been shown to modulate MPTP-induced intestinal inflammatory responses, intestinal barrier integrity, and enteric neuronal dysfunction (Huh et al., 2020). Eugene Huh et al. reported that 6S inhibited Proteus mirabilis, ameliorating intestinal barrier disruption, motor dysfunction, and neuronal death (Huh et al., 2023b).
3.2 Polysaccharides
3.2.1 Chitosan
Chitosan is primarily derived from the exoskeletons of crustaceans, mollusks, and fungal cell walls. Chitosan exerts neuroprotective effects through mechanisms including free radical scavenging, antioxidation, inhibition of apoptosis, and anti-inflammatory actions (Zhu et al., 2024). Preclinical studies demonstrate that chitosan enhances, ETC enzyme activity, restores dopamine levels, and ameliorates motor dysfunction and neurotoxicity (Pramod Kumar and Harish Prashanth, 2020). These actions establish its neuroprotective efficacy via mitochondrial functional restoration. Furthermore, chitosan crosses the BBB to suppress neuroinflammation through TSPO/c-Fos pathway-mediated downregulation of TNF-α and iNOS. This mechanism protects dopaminergic neurons from inflammatory damage (Chen, 2024). Moreover, emerging evidence indicates chitosan modulates gut-brain crosstalk by reducing acetate levels and repairing intestinal and BBB impairments (Wang et al., 2026). These dual-barrier interventions suggest microbial metabolite regulation contributes to PD pathology mitigation. Notably, the specific effects on GM composition and microbial metabolites remain unverified—a key limitation requiring further investigation.
3.2.1.1 Fucoidan
Fucoidan, a sulfated polysaccharide abundantly present in the mucilage of brown algae (e.g., Laminaria, Undaria pinnatifida, Fucus, and Nemacystus decipiens), exhibits potent immunomodulatory properties with emerging neuroprotective applications in PD via the MGBA. Preclinical evidence demonstrates that fucoidan remodels GM composition and suppresses LPS/TLR4/NF-κB signaling to modulate microglial polarization in SN (M1-type markers TNF-α and iNOS reduction, with M2-type markers Arg-1 and CD206 elevation) (Yang X. et al., 2024). Mechanistically, fucoidan enhances macrophage autophagy activity, amplifying anti-inflammatory responses (Hou et al., 2025). Fucoidan also regulates tryptophan metabolism via microbial modulation (kynurenine pathway) and strengthens IB integrity (Qin et al., 2025; Xue et al., 2025). Collectively, fucoidan coordinates gut-immune-brain crosstalk to mitigate PD pathogenesis through synergistic microbiota modulation and immunoregulation.
3.2.1.2 Fucosylated chondroitin sulfate
Fucosylated Chondroitin Sulfate (FuCS), a biologically active glycosaminoglycan (GAG) extracted from sea cucumber body wall. Ye’s study demonstrated that FuCS modulated the GM by reducing the abundance of the conditionally pathogenic bacterium Staphylococcus while increasing probiotic Muribaculaceae. Concomitantly, FuCS decreased the expression of Soyasapogenol E—a metabolite positively correlated with IL-1β—and lowered α-syn levels. These findings indicate that FuCS alleviates nigral neurodegeneration and motor dysfunction in PD mice through GM-mediated anti-inflammatory effects (Ye, 2022). Furthermore, FuCS enhanced IB function by upregulating colonic expression of tight junction proteins ZO-1, occludin, and claudin-1.
3.2.1.3 Ganoderma lucidum polysaccharide
Ganoderma lucidum Polysaccharide (GLP) is a naturally occurring polysaccharide primarily derived from the fruiting bodies, mycelia, or spores of the medicinal fungus Ganoderma lucidum (Curtis) P. Karst. Pharmacological studies have demonstrated that GLP exhibits anti-inflammatory, antioxidative, and anti-apoptotic effects (Liu et al., 2023). Recent studies demonstrate that GLP reduces α-syn expression in PD models while modulating GM abundance, diversity, and evenness. GLP concurrently elevates SCFA levels and suppresses the intestinal TLR4/MyD88/NF-κB pathway, thereby downregulating pro-inflammatory cytokines including IL-6 and IL-1β (Chen et al., 2025). These findings indicate that GLP exerts neuroprotective effects by mitigating inflammation through GM regulation and microbial metabolites-SCFAs.
3.2.1.4 Polymannuronic acid
Polymannuronic acid (PM), an oligosaccharide derived from brown algal alginate, is a structural polysaccharide exhibiting diverse biological activities—including antioxidant, immunomodulatory, antihypertensive, hypoglycemic, neuroprotective, and antimicrobial effects. Song’s research revealed that low-concentration PM significantly reduces the abundance of the opportunistic pathogen Escherichia while increasing the prevalence of the beneficial genus Prevotella in the GM (Song, 2023). Further studies indicate that PM modulates GM in PD, elevating SCFA levels to mediate neuroprotective effects via the gut-brain axis (Dong et al., 2020). Simultaneously, PM suppresses the MAPK pathway, mitigating systemic inflammation and enhancing the integrity of both IB and BBB.
3.2.1.5 Gastrodia elata polysaccharide
Gastrodia elata polysaccharide (GEP), derived from dried tubers of the orchidaceous plant Gastrodia elata Bl, is believed in traditional medicine to possess therapeutic effects such as calming endogenous wind, relieving convulsions, and suppressing hyperactive liver yang. Mechanistically, GEP modulates gut-brain axis activity to support neurological health (Gan Q. et al., 2024). As demonstrated by Gan et al., GEP ameliorates GM dysbiosis (notably enhancing Akkermansia and Lactobacillus abundance) while elevating SCFA levels (Gan Q. X. et al., 2024). Concomitantly, it upregulates occludin expression and reduces pro-inflammatory cytokines (TNF-α, IL-1β, IL-6), collectively inhibiting α-syn accumulation and dopaminergic neuron loss. These findings indicate GEP exerts neuroprotective effects in PD mice by restoring the IB and mitigating inflammation via GM.
3.2.1.6 Arabinoxylans
Arabinoxylan (AX), a non-starch polysaccharide primarily found in the cell walls of cereal grains and other plants, exhibits immunomodulatory activity. Preclinical evidence in PD indicates that AX increases the relative abundance of Firmicutes and Clostridia, elevates TH levels in the SN, and reduces α-syn accumulation (Luo et al., 2023). These findings suggest that AX exerts neuroprotective effects via modulation of the GM. Further studies by Bao revealed that AX promotes the growth of microbiota negatively associated with PD, enhances SCFA production, and increases GLP-1 secretion by 29% (Bao, 2023). This indicates that the neuroprotective effects of AX in PD are primarily mediated indirectly through the GM and GLP-1.
3.2.1.7 Trehalose
Trehalose, a non-reducing disaccharide composed of two glucose molecules, is primarily found in fungi, insects, plants, and certain animals. It exhibits biological activities, including antioxidant and neuroprotective effects. In a study by Pradeloux et al. using PrP-A53T transgenic mice as a model, the effects of trehalose on GM were investigated. The results demonstrated that trehalose increased Lachnospiraceae abundance, elevated GLP-1 expression, and reduced THR levels in both brain and gut tissues, thereby exerting neuroprotective effects (Pradeloux et al., 2024).
3.3 Terpenoids and glycosides
3.3.1 Panaxadiol
Panaxadiol, a naturally derived bioactive compound extracted from Panax species of the Araliaceae family, belongs to the dammarane-type triterpenoid sapogenins and exhibits multi-target regulatory capabilities, particularly in neuroprotection, cardiovascular protection, anti-inflammatory effects, and antioxidant activity. The highest concentration of panaxadiol is observed in the gastrointestinal tract of rats, where it exerts anti-inflammatory effects on macrophages by modulating the MAPK pathway, specifically through suppressing the phosphorylation (activation) of p38 and ERK (Xu L. et al., 2022). In a study by Xu et al., the impact of panaxadiol on PD was investigated at both cellular and animal levels (Xu et al., 2025). It was demonstrated for the first time that panaxadiol significantly restores GM composition, suppresses downstream pro-inflammatory cytokine production via the TLR4/MyD88/NF-κB pathway, reduces activation of SN glial cells, and attenuates peripheral and central inflammation. Additionally, it mitigates BBB damage by upregulating tight junction proteins (ZO-1, occludin, and claudin-5), repairs neuronal loss, and improves motor and gastrointestinal dysfunction.
3.3.2 Rubusoside
Rubusoside (Ru), a diterpene glycoside from the leaves of Rubus chingii var. suavissimus, is a high-intensity, low-calorie natural sweetener with potent anti-inflammatory properties. Preclinical evidence indicates that Ru alleviates LPS-induced low-grade chronic inflammation (Zhang et al., 2020). Ru has been demonstrated to modify the composition of gut microbiota and metabolite profiles (particularly amino acid metabolism) in PD model mice, inhibit microglial activation to reduce the release of proinflammatory mediators, regulate the JNK/p38 MAPK/NF-κB signaling pathway, mitigate oxidative stress, prevent neuronal apoptosis, and protect dopaminergic neurons while improving motor dysfunction (Meng et al., 2024). These findings suggest the potential of Ru in delaying PD progression through modulation of GM.
3.3.3 Withaferin A
Withaferin A (WFA) is a naturally occurring steroidal lactone compound with diverse biological activities, including anti-inflammatory, antiviral, and neuroprotective effects. It is primarily derived from the leaves of Withania somnifera. Sun et al. administered WFA to A53T transgenic mice (Sun et al., 2025), observing an increase in the abundance of anti-inflammatory GM and treversed alterations in 55 metabolites associated with sphingolipid metabolism, dopaminergic synapses, and neuroactive ligand-receptor interactions. These findings demonstrate that WFA modulates sphingolipid signaling pathways within the MGBA, mitigates neuroinflammation, and promotes neuronal repair.
3.3.4 Salidroside
Salidroside (SAL), a phenylethanoid glycoside from Rhodiola spp., exhibits multifaceted pharmacological activities, including GM modulation, antioxidative, anti-inflammatory, and neuroprotective effects. Cui et al. demonstrated that SAL intervention in PD models increased Lactobacillus abundance while reducing Aerococcus, Desulfovibrio, and Staphylococcus; modulated TAU-associated metabolic pathways; enhanced intestinal length and structural integrity; downregulated IL-6, IL-1β, and TNF-α expression; inhibited α-syn deposition and glial activation; and ameliorated motor dysfunction and neuronal loss (Cui, 2022).
3.3.5 Cordycepin
Cordycepin, a nucleoside compound derived from the medicinal fungus Cordyceps sinensis, demonstrates significant therapeutic potential against neuroinflammation in PD by targeting multiple pathways, including NLRP3 inflammasome and TLR/NF-κB signaling (Cheng and Zhu, 2019; Zhang et al., 2021; Sharma et al., 2023). Research by Liu et al. revealed cordycepin’s dual action: improving GM composition while modulating the Th1/Th2 and Th17/Treg immune axis balance (Liu Z. et al., 2024). Crucially, cordycepin suppresses PI3K/AKT/mTOR and ERK/JNK pathway activation, alleviates neuroinflammation, and enhances autophagy-related protein expression in the striatum and SN. This cascade ultimately reduces neuronal apoptosis in MPTP-induced PD mice (Wang L. et al., 2024). Furthermore, cordycepin regulates adenosine A2A receptors and ameliorates cognitive dysfunction in PD models, offering a novel therapeutic strategy for PD-associated dementia (Huang et al., 2023).
3.4 Alkaloids
3.4.1 Berberine
Berberine (BBR), an isoquinoline alkaloid primarily biosynthesized by Berberidaceae (e.g., Berberis spp.) and Ranunculaceae (e.g., Coptis and Thalictrum spp.) plants, exerts neuroprotective effects by crossing the BBB (Begh et al., 2025). Clinical evidence indicates that oral BBR administration activates the gut-brain axis, promoting levodopa production by Enterococcus to increase cerebral dopamine levels (Wang Y. et al., 2021). Li et al. demonstrated BBR’s efficacy in PD patients by improving GM dysbiosis and suppressing inflammatory cytokines (Li J. et al., 2022). In PD rat models, BBR ameliorated colonic dysfunction, restored mucosal permeability, modulated intestinal neurotransmitters and EGCs, and alleviated depressive-like behaviors (Liu et al., 2025). Collectively, these findings establish GM as a pivotal mediator of BBR’s neuroprotective effects.
3.4.2 Piperine
Piperine (PIP) is a pungent cinnamoyl amide alkaloid extracted from the fruits of Piper nigrum and Piper longum. As a medicinal and edible compound, it exhibits anti-inflammatory, antioxidant, cognitive-enhancing, gastrointestinal-protective, and antidepressant properties. Clinical evidence indicates that PIP modulates GM by upregulating Bacteroides and Prevotella while downregulating Salmonella and Escherichia. It elevates SCFA levels, restores IB integrity, and alleviates neuronal loss and α-Syn aggregation. These effects occur through gut-brain axis autophagy modulation via the PI3K/AKT/mTOR pathway, ultimately improving motor deficits and gastrointestinal dysfunction in PD rat models (Yu et al., 2024). Additionally, Hu et al. demonstrated that PIP enhances L-dopa bioavailability and increases cerebral dopamine by suppressing Enterococcus faecalis-mediated L-dopa metabolism in the gut, thereby ameliorating motor impairments in PD rats (Hu et al., 2024).
3.5 Others
3.5.1 Alpha-linolenic acid
Alpha-linolenic acid (ALA), an essential omega-3 polyunsaturated fatty acid, is abundant in perilla seed oil and constitutes 51%–64% of its composition. With potent antioxidant and anti-inflammatory properties, ALA exerts neuroprotective effects through gut-brain axis modulation (Mahnoor et al., 2025). Peerapa et al. demonstrated that ALA enhances GM diversity and boosts SCFA production. Concurrently, it reduces α-syn accumulation in both colonic myenteric plexus and hippocampal regions, suppresses microglial activation, attenuates inflammatory cascades, and increases colon tissue length (Techaniyom et al., 2024). Furthermore, ALA upregulates TH expression in SN and striatal regions while ameliorating motor and non-motor symptoms. These findings establish ALA as a multi-mechanism modulator of GM-mediated neuroprotection.
3.5.2 Sika deer velvet antler peptide
Sika Deer Velvet Antler Peptide (VAP) is derived from the non-ossified antlers of male deer, exhibiting tonic properties and immunomodulatory effects. Peptidomic analysis has identified 189 peptides with notable metal-binding characteristics. Studies have demonstrated that VAP increases the abundance of Prevotellaceae, Helicobacteraceae, and Prevotella while activating the SIRT1-mediated Akt/Nrf2/HO-1 pathway to mitigate MPP-induced apoptosis and oxidative stress (Liu Y. et al., 2024). Furthermore, it suppresses microglial activation and reduces the phosphorylation levels of the MAPK/Akt inflammatory pathway, subsequently lowering the expression of inflammatory mediators and preventing neuroinflammation. These findings suggest that VAP possesses neuroprotective potential in PD.
3.5.3 Allantoin
Allantoin, a naturally occurring compound predominantly found in plants (notably leguminous species), exhibits anti-inflammatory, antioxidant, and soothing properties. As demonstrated by Shuo Yang et al., allantoin modulates the GM, exerts antioxidative effects via the AKT/Nrf2/HO-1 signaling pathway, and suppresses inflammatory responses through inhibition of the NF-κB and MAPK signaling pathways (Yang S. et al., 2024). Additionally, it contributes to intestinal barrier restoration and alleviates neurodegeneration.
4 Limitations in the clinical translation of natural products
Current research on natural product-mediated GM modulation for PD treatment remains predominantly preclinical, lacking large-scale clinical evidence. Existing clinical data demonstrate that FMT significantly improves autonomic symptoms in PD patients, while probiotic therapy markedly reduces disease severity, anxiety, and gastrointestinal disturbances (Cheng et al., 2023; Andreozzi et al., 2024; Bruggeman et al., 2024; Zali et al., 2024). These findings suggest the potential of GM modulation in PD prevention and treatment. These findings highlight GM’s therapeutic potential for PD. Traditional medicine studies confirm that Chaihu Jia Longgu Muli decoction regulates GM and decreases oxidative stress in PD patients (Zhang S. et al., 2024).
However, clinical translation of natural products faces key limitations. The inherent complexity of natural products poses challenges in standardization. Poor solubility, structural instability, rapid metabolism, and low bioavailability hinder their clinical application. Additionally, pharmacokinetic challenges prevent these compounds from reaching effective concentrations in target tissues. The toxic side effects associated with natural products remain insufficiently studied, necessitating further toxicological evaluations. Furthermore, clinical trial design presents difficulties. Identifying sensitive and specific clinical endpoints and biomarkers presents a major challenge. However, these markers must reflect actual disease modification or microbiota modulation—not merely symptomatic relief.
5 Discussion and conclusion
As the second most prevalent neurodegenerative disorder, PD has been extensively linked to the GM, with mounting evidence suggesting a profound association between microbial communities and the interindividual variability in PD severity and clinical manifestations. Research on PD pathophysiology and therapeutic strategies has shifted from the traditional brain-centric paradigm to a novel perspective centered on the MGBA. Natural products, owing to their multi-target effects and low toxicity, exhibit unique advantages in modulating the MGBA. Although some natural products suffer from limitations such as poor bioavailability and limited BBB permeability, existing studies have demonstrated that their interactions with GM constitute a sophisticated “biotransformation-signal amplification” system, exerting direct or indirect therapeutic effects against PD.
In recent years, significant breakthroughs have been made in elucidating the regulatory role of natural products in PD treatment via GM modulation. This review systematically summarizes natural products that mediate GM-based PD therapy and their underlying mechanisms. Polyphenols, flavonoids, polysaccharides, alkaloids, terpenoids, and glycosides exert anti-PD effects by modulating the diversity and abundance of PD-associated GM, altering metabolite levels, inhibiting α-Syn pathological aggregation, improving barrier function, and mitigating neuroinflammation, mitochondrial dysfunction, and oxidative stress, thereby influencing immune, endocrine, and nervous system pathways. In multiple aforementioned preclinical studies (e.g., EGCG and Neo), the addition of antibiotics was found to attenuate the neuroprotective effects against PD, suggesting to some extent that alterations in the GM precede therapeutic efficacy. Notably, the vast diversity of GM enables the “personalized processing” of natural products, allowing these metabolites to serve as key players across multiple mechanistic pathways. Notably, although Akkermansia is enriched in the gut of PD patients, accumulating evidence indicates its neuroprotective potential. This apparent paradox highlights the bacterium’s environmentally modulated pleiotropy within the gut–brain axis. During homeostasis or compensatory states, it confers neuroprotection via immunomodulation and bioactive metabolite secretion. Conversely, under PD-associated chronic inflammation with IB disruption, its mucolytic activity may drive pathogenesis.
With deepening insights into the GM-PD relationship, microbiota-based therapeutic strategies hold broad clinical potential. Modulating GM may enhance patient responsiveness to conventional treatments, thereby improving therapeutic outcomes. Interdisciplinary research integrating nutriology and microbiology will provide novel perspectives for personalized PD therapy. Building on existing findings, future research faces several challenges: 1) elucidating mechanistic details and identifying therapeutic targets; 2) developing targeted delivery systems, such as nanocarriers or engineered bacterial shells, to achieve colon-specific release of natural products; 3) exploring combination therapies with natural products; 4) constructing multi-level intervention strategies; and 5) investigating pharmacokinetics, optimal dosing, and metabolic duration. Addressing these challenges remains a complex and demanding task.
In summary, natural products represent a promising avenue for PD intervention through GM modulation. With the advancement of the NIH-sponsored gut-brain axis research initiative and the maturation of personalized microbiota-based therapies, early prevention and treatment strategies targeting the gut may redefine PD clinical management, offering a means to delay the progression of this neurodegenerative disorder at its root.
Author contributions
YJ: Writing – original draft, Writing – review and editing. YZ: Software, Writing – original draft. XT: Investigation, Writing – original draft. TZ: Writing – review and editing. HnZ: Writing – review and editing. HiZ: Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This work was supported by the National Natural Science Foundation of China (82174232), the Scientific Development Project (Incubation Program) of the Chinese Association of Rehabilitation Medicine (KFKT-2023-037), the Key Project of Heilongjiang Provincial Educational Science Planning (GJB1424172), and the Fifth Cohort of National Training Program for Outstanding Clinical Talents in Traditional Chinese Medicine.
Acknowledgments
Figure 1 and Figure 2 were created with FigDraw.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Aburto, M. R., and Cryan, J. F. (2024). Gastrointestinal and brain barriers: unlocking gates of communication across the microbiota-gut-brain axis. Nat. Rev. Gastroenterol. Hepatol. 21 (4), 222–247. doi:10.1038/s41575-023-00890-0
Amorim Neto, D. P., Bosque, B. P., Pereira de Godoy, J. V., Rodrigues, P. V., Meneses, D. D., Tostes, K., et al. (2022). Akkermansia muciniphila induces mitochondrial calcium overload and α -synuclein aggregation in an enteroendocrine cell line. iScience 25 (3), 103908. doi:10.1016/j.isci.2022.103908
Andreozzi, V., Cuoco, S., Balestrieri, M., Fierro, F., Ferrara, N., Erro, R., et al. (2024). Synbiotic supplementation may globally improve non-motor symptoms in patients with stable Parkinson's disease: results from an open label single-arm study. Sci. Rep. 14 (1), 23095. doi:10.1038/s41598-024-74400-w
Atanasov, A. G., Zotchev, S. B., Dirsch, V. M., and Supuran, C. T. (2021). Natural products in drug discovery: advances and opportunities. Nat. Rev. Drug Discov. 20 (3), 200–216. doi:10.1038/s41573-020-00114-z
Backman, E. A., Gardberg, M., Luntamo, L., Peurla, M., Vahlberg, T., Borghammer, P., et al. (2025). Nigral neuroinflammation and dopaminergic neurons in Parkinson's disease and atypical parkinsonisms. Ann. Neurol. 97 (6), 1096–1109. doi:10.1002/ana.27202
Bai, F., You, L., Lei, H., and Li, X. (2024). Association between increased and decreased gut microbiota abundance and Parkinson's disease: a systematic review and subgroup meta-analysis. Exp. Gerontol. 191, 112444. doi:10.1016/j.exger.2024.112444
Banerjee, R., Starkov, A. A., Beal, M. F., and Thomas, B. (2009). Mitochondrial dysfunction in the limelight of Parkinson's disease pathogenesis. Biochim. Biophys. Acta 1792 (7), 651–663. doi:10.1016/j.bbadis.2008.11.007
Bao, C. (2023). The effects of madopar combined with Arabinoxylan on motor function and intestinal flora, intestinal permeability and inflammation in PD rats. [dissertation/master's thesis]. China: Youjiang Medical University For Nationalities. doi:10.27908/d.cnki.gymzy.2023.000305
Barichella, M., Severgnini, M., Cilia, R., Cassani, E., Bolliri, C., Caronni, S., et al. (2019). Unraveling gut microbiota in Parkinson's disease and atypical parkinsonism. Mov. Disord. 34 (3), 396–405. doi:10.1002/mds.27581
Bedarf, J. R., Hildebrand, F., Coelho, L. P., Sunagawa, S., Bahram, M., Goeser, F., et al. (2017). Functional implications of microbial and viral gut metagenome changes in early stage L-DOPA-naïve Parkinson's disease patients. Genome Med. 9 (1), 39. doi:10.1186/s13073-017-0428-y
Begh, M. Z. A., Amin, M. A., Shatu, M. M., Sweilam, S. H., Puri, S., Ramesh, R. B., et al. (2025). Unraveling berberine's molecular mechanisms in neuroprotection against neurodegeneration. Chem. Biodivers. 22 (8), e202500170. doi:10.1002/cbdv.202500170
Ben-Shlomo, Y., Darweesh, S., Llibre-Guerra, J., Marras, C., San Luciano, M., and Tanner, C. (2024). The epidemiology of Parkinson's disease. Lancet 403 (10423), 283–292. doi:10.1016/s0140-6736(23)01419-8
Bharate, S. S., Mignani, S., and Vishwakarma, R. A. (2018). Why are the majority of active compounds in the CNS domain natural products? A critical analysis. J. Med. Chem. 61 (23), 10345–10374. doi:10.1021/acs.jmedchem.7b01922
Bhattarai, Y., Williams, B. B., Battaglioli, E. J., Whitaker, W. R., Till, L., Grover, M., et al. (2018). Gut microbiota-produced tryptamine activates an epithelial G-protein-coupled receptor to increase colonic secretion. Cell Host Microbe 23 (6), 775–785. doi:10.1016/j.chom.2018.05.004
Borghammer, P., Just, M. K., Horsager, J., Skjærbæk, C., Raunio, A., Kok, E. H., et al. (2022). A postmortem study suggests a revision of the dual-hit hypothesis of Parkinson's disease. NPJ Park. Dis. 8 (1), 166. doi:10.1038/s41531-022-00436-2
Braak, H., Rüb, U., Gai, W. P., and Del Tredici, K. (2003). Idiopathic Parkinson's disease: possible routes by which vulnerable neuronal types may be subject to neuroinvasion by an unknown pathogen. J. Neural Transm. (Vienna) 110 (5), 517–536. doi:10.1007/s00702-002-0808-2
Braniste, V., Al-Asmakh, M., Kowal, C., Anuar, F., Abbaspour, A., Tóth, M., et al. (2014). The gut microbiota influences blood-brain barrier permeability in mice. Sci. Transl. Med. 6 (263), 263ra158. doi:10.1126/scitranslmed.3009759
Bravo, J. A., Forsythe, P., Chew, M. V., Escaravage, E., Savignac, H. M., Dinan, T. G., et al. (2011). Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc. Natl. Acad. Sci. U. S. A. 108 (38), 16050–16055. doi:10.1073/pnas.1102999108
Bruggeman, A., Vandendriessche, C., Hamerlinck, H., De Looze, D., Tate, D. J., Vuylsteke, M., et al. (2024). Safety and efficacy of faecal microbiota transplantation in patients with mild to moderate Parkinson's disease (GUT-PARFECT): a double-blind, placebo-controlled, randomised, phase 2 trial. EClinicalMedicine 71, 102563. doi:10.1016/j.eclinm.2024.102563
Cai, B., Zhong, L., Wang, Q., Xu, W., Li, X., and Chen, T. (2023). Curcumin alleviates 1-methyl- 4-phenyl- 1,2,3,6-tetrahydropyridine- induced Parkinson's disease in mice via modulating gut microbiota and short-chain fatty acids. Front. Pharmacol. 14, 1198335. doi:10.3389/fphar.2023.1198335
Calabrese, E. J., Pressman, P., Hayes, A. W., Agathokleous, E., Dhawan, G., Kapoor, R., et al. (2025). Fisetin: hormesis accounts for many of its chemoprotective effects. Biogerontology 26 (2), 90. doi:10.1007/s10522-025-10230-1
Cao, H., Tian, Q., Chu, L., Wu, L., Gao, H., and Gao, Q. (2024). Lycium ruthenicum Murray anthocyanin-driven neuroprotection modulates the gut microbiome and metabolome of MPTP-treated mice. Food Funct. 15 (24), 12210–12227. doi:10.1039/d4fo01878h
Chandra, R., Sokratian, A., Chavez, K. R., King, S., Swain, S. M., Snyder, J. C., et al. (2023). Gut mucosal cells transfer α-synuclein to the vagus nerve. JCI Insight 8 (23), e172192. doi:10.1172/jci.insight.172192
Chang, Y. H. (2025). Curcumin as a potential therapeutic agent for Parkinson's disease: a systematic review. Front. Pharmacol. 16, 1593191. doi:10.3389/fphar.2025.1593191
Chang, K. H., Cheng, M. L., Tang, H. Y., Huang, C. Y., Wu, Y. R., and Chen, C. M. (2018). Alternations of metabolic profile and kynurenine metabolism in the plasma of Parkinson's disease. Mol. Neurobiol. 55 (8), 6319–6328. doi:10.1007/s12035-017-0845-3
Chen, R. (2024). “TSPO/c-Fos pathway mediates the mechanism of Chitosan oligosaccharides in alleviating Parkinson's disease,” [dissertation/master's thesis]. China: Kunming Medical University doi:10.27202/d.cnki.gkmyc.2024.000726
Chen, J., and Vitetta, L. (2020). Mitochondria could be a potential key mediator linking the intestinal microbiota to depression. J. Cell Biochem. 121 (1), 17–24. doi:10.1002/jcb.29311
Chen, T. J., Feng, Y., Liu, T., Wu, T. T., Chen, Y. J., Li, X., et al. (2020). Fisetin regulates gut microbiota and exerts neuroprotective effect on mouse model of Parkinson's disease. Front. Neurosci. 14, 549037. doi:10.3389/fnins.2020.549037
Chen, Y., Wu, W., Zhao, S., Lv, X., Hu, J., Han, C., et al. (2021). Increased accumulation of α-synuclein in inflamed appendices of Parkinson's disease patients. Mov. Disord. 36 (8), 1911–1918. doi:10.1002/mds.28553
Chen, S. J., Chen, C. C., Liao, H. Y., Wu, Y. W., Liou, J. M., Wu, M. S., et al. (2022a). Alteration of gut microbial metabolites in the systemic circulation of patients with Parkinson's disease. J. Park. Dis. 12 (4), 1219–1230. doi:10.3233/jpd-223179
Chen, Y., Liu, Y., Wang, Y., Chen, X., Wang, C., Chen, X., et al. (2022b). Prevotellaceae produces butyrate to alleviate PD-1/PD-L1 inhibitor-related cardiotoxicity via PPARα-CYP4X1 axis in colonic macrophages. J. Exp. Clin. Cancer Res. 41 (1), 1. doi:10.1186/s13046-021-02201-4
Chen, L., Ling, Y., Sun, J., Zhou, S., Xiao, Y., Zou, X., et al. (2025). A glucan from Ganoderma lucidum: structural characterization and the anti-inflammatory effect on Parkinson's disease via regulating dysfunctions of intestinal microecology and inhibiting TLR4/MyD88/NF-κB signaling pathway. J. Ethnopharmacol. 343, 119446. doi:10.1016/j.jep.2025.119446
Cheng, C., and Zhu, X. (2019). Cordycepin mitigates MPTP-induced Parkinson's disease through inhibiting TLR/NF-κB signaling pathway. Life Sci. 223, 120–127. doi:10.1016/j.lfs.2019.02.037
Cheng, Y., Tan, G., Zhu, Q., Wang, C., Ruan, G., Ying, S., et al. (2023). Efficacy of fecal microbiota transplantation in patients with Parkinson's disease: clinical trial results from a randomized, placebo-controlled design. Gut Microbes 15 (2), 2284247. doi:10.1080/19490976.2023.2284247
Chenghan, M., Wanxin, L., Bangcheng, Z., Yao, H., Qinxi, L., Ting, Z., et al. (2025). Short-chain fatty acids mediate gut microbiota-brain communication and protect the blood-brain barrier integrity. Ann. N. Y. Acad. Sci. 1545 (1), 116–131. doi:10.1111/nyas.15299
Cilia, R., Piatti, M., Cereda, E., Bolliri, C., Caronni, S., Ferri, V., et al. (2021). Does gut microbiota influence the course of Parkinson's disease? A 3-year prospective exploratory study in de novo patients. J. Park. Dis. 11 (1), 159–170. doi:10.3233/jpd-202297
Cui, C. (2022). Study on the role and mechanism of salidrosideand curcumin in regulating gut microbiome-metabolome axis during Parkinson's disease. China: Tongji University. doi:10.27372/d.cnki.gtjsu.2022.000204
Cui, C., Han, Y., Li, H., Yu, H., Zhang, B., and Li, G. (2022). Curcumin-driven reprogramming of the gut microbiota and metabolome ameliorates motor deficits and neuroinflammation in a mouse model of Parkinson's disease. Front. Cell Infect. Microbiol. 12, 887407. doi:10.3389/fcimb.2022.887407
Cui, W., Guo, M., Liu, D., Xiao, P., Yang, C., Huang, H., et al. (2024a). Gut microbial metabolite facilitates colorectal cancer development via ferroptosis inhibition. Nat. Cell Biol. 26 (1), 124–137. doi:10.1038/s41556-023-01314-6
Cui, Z., Guo, F. Y., Li, L., Lu, F., Jin, C. H., Wang, X., et al. (2024b). Brazilin-7-acetate, a novel potential drug of Parkinson's disease, hinders the formation of α-synuclein fibril, mitigates cytotoxicity, and decreases oxidative stress. Eur. J. Med. Chem. 264, 115965. doi:10.1016/j.ejmech.2023.115965
Dalile, B., Van Oudenhove, L., Vervliet, B., and Verbeke, K. (2019). The role of short-chain fatty acids in microbiota-gut-brain communication. Nat. Rev. Gastroenterol. Hepatol. 16 (8), 461–478. doi:10.1038/s41575-019-0157-3
Dohgu, S., Takata, F., Matsumoto, J., Kimura, I., Yamauchi, A., and Kataoka, Y. (2019). Monomeric α-synuclein induces blood-brain barrier dysfunction through activated brain pericytes releasing inflammatory mediators in vitro. Microvasc. Res. 124, 61–66. doi:10.1016/j.mvr.2019.03.005
Dong, X. L., Wang, X., Liu, F., Liu, X., Du, Z. R., Li, R. W., et al. (2020). Polymannuronic acid prevents dopaminergic neuronal loss via brain-gut-microbiota axis in Parkinson's disease model. Int. J. Biol. Macromol. 164, 994–1005. doi:10.1016/j.ijbiomac.2020.07.180
Dong, S., Shen, B., Jiang, X., Zhu, J., Zhang, H., Zhao, Y., et al. (2024a). Comparison of vagus nerve cross-sectional area between brain-first and body-first Parkinson's disease. NPJ Park. Dis. 10 (1), 231. doi:10.1038/s41531-024-00844-6
Dong, Y., Qi, Y., Chen, J., Han, S., Su, W., Ma, X., et al. (2024b). Neuroprotective effects of Bifidobacterium animalis subsp. lactis NJ241 in a mouse model of Parkinson's disease: implications for gut microbiota and PGC-1α. Mol. Neurobiol. 61 (10), 7534–7548. doi:10.1007/s12035-024-04038-2
Du, X. Y., Xie, X. X., and Liu, R. T. (2020). The role of α-synuclein oligomers in Parkinson's disease. Int. J. Mol. Sci. 21 (22), 8645. doi:10.3390/ijms21228645
Duscha, A., Gisevius, B., Hirschberg, S., Yissachar, N., Stangl, G. I., Dawin, E., et al. (2020). Propionic acid shapes the multiple sclerosis disease course by an immunomodulatory mechanism. Cell 180 (6), 1067–1080. doi:10.1016/j.cell.2020.02.035
Elabi, O., Gaceb, A., Carlsson, R., Padel, T., Soylu-Kucharz, R., Cortijo, I., et al. (2021). Human α-synuclein overexpression in a mouse model of Parkinson's disease leads to vascular pathology, blood brain barrier leakage and pericyte activation. Sci. Rep. 11 (1), 1120. doi:10.1038/s41598-020-80889-8
Espinosa-Oliva, A. M., Ruiz, R., Soto, M. S., Boza-Serrano, A., Rodriguez-Perez, A. I., Roca-Ceballos, M. A., et al. (2024). Inflammatory bowel disease induces pathological α-synuclein aggregation in the human gut and brain. Neuropathol. Appl. Neurobiol. 50 (1), e12962. doi:10.1111/nan.12962
Evancho, A., Do, M., Fortenberry, D., Billings, R., Sartayev, A., and Tyler, W. J. (2024). Vagus nerve stimulation in Parkinson's disease: a scoping review of animal studies and human subjects research. NPJ Park. Dis. 10 (1), 199. doi:10.1038/s41531-024-00803-1
Fan, Y., Yang, W., Wu, W., Wang, X., Lin, Y., Wu, L., et al. (2024). Serum neurotransmitter analysis of motor and non-motor symptoms in Parkinson's patients. Front. Aging Neurosci. 16, 1423120. doi:10.3389/fnagi.2024.1423120
Fang, X., Liu, S., Muhammad, B., Zheng, M., Ge, X., Xu, Y., et al. (2024). Gut microbiota dysbiosis contributes to α-synuclein-related pathology associated with C/EBPβ/AEP signaling activation in a mouse model of Parkinson's disease. Neural Regen. Res. 19 (9), 2081–2088. doi:10.4103/1673-5374.391191
Fanibunda, S. E., Deb, S., Maniyadath, B., Tiwari, P., Ghai, U., Gupta, S., et al. (2019). Serotonin regulates mitochondrial biogenesis and function in rodent cortical neurons via the 5-HT(2A) receptor and SIRT1-PGC-1α axis. Proc. Natl. Acad. Sci. U. S. A. 116 (22), 11028–11037. doi:10.1073/pnas.1821332116
Fasano, A., Visanji, N. P., Liu, L. W., Lang, A. E., and Pfeiffer, R. F. (2015). Gastrointestinal dysfunction in Parkinson's disease. Lancet Neurol. 14 (6), 625–639. doi:10.1016/s1474-4422(15)00007-1
Foo, J. H., Culvenor, J. G., Ferrero, R. L., Kwok, T., Lithgow, T., and Gabriel, K. (2010). Both the p33 and p55 subunits of the Helicobacter pylori VacA toxin are targeted to mammalian mitochondria. J. Mol. Biol. 401 (5), 792–798. doi:10.1016/j.jmb.2010.06.065
Gan, Q., Chen, L., Xian, J., An, G., Wei, H., and Ma, Y. (2024a). Digestive characteristics of Gastrodia elata Blume polysaccharide and related impacts on human gut microbiota in vitro. J. Ethnopharmacol. 328, 118064. doi:10.1016/j.jep.2024.118064
Gan, Q. X., Peng, M. Y., Wei, H. B., Chen, L. L., Chen, X. Y., Li, Z. H., et al. (2024b). Gastrodia elata polysaccharide alleviates Parkinson's disease via inhibiting apoptotic and inflammatory signaling pathways and modulating the gut microbiota. Food Funct. 15 (6), 2920–2938. doi:10.1039/d3fo05169b
Gao, B. (2022). “Study on the effect and mechanism of cyanidin-3-O-glucoside on gut microbiota,”. [dissertation/master's thesis]. China: Shenyang Agricultural University. doi:10.27327/d.cnki.gshnu.2022.000115
Gao, X., Fu, S., Wen, J., Yan, A., Yang, S., Zhang, Y., et al. (2024). Orally administered ginkgolide C alleviates MPTP-induced neurodegeneration by suppressing neuroinflammation and oxidative stress through microbiota-gut-brain Axis in mice. J. Agric. Food Chem. 72 (40), 22115–22131. doi:10.1021/acs.jafc.4c03783
Gao, W., Wu, X., Wang, Y., Lu, F., and Liu, F. (2025). Brazilin-rich extract from Caesalpinia sappan L. Attenuated the motor deficits and neurodegeneration in MPTP/p-Induced Parkinson's disease mice by regulating gut microbiota and inhibiting inflammatory responses. ACS Chem. Neurosci. 16 (2), 181–194. doi:10.1021/acschemneuro.4c00679
Gonçalves, P. B., Sodero, A. C. R., and Cordeiro, Y. (2021). Green tea epigallocatechin-3-gallate (EGCG) targeting protein misfolding in drug discovery for neurodegenerative diseases. Biomolecules 11 (5), 767. doi:10.3390/biom11050767
Gonzaga, N. L., Karim, F., Liang, C., and Mukherjee, J. (2025). [(18)F]Mefway: imaging serotonin 5HT(1A) receptors in human postmortem Alzheimer's and Parkinson's disease anterior cingulate. Potential applications to human positron emission tomography studies. Biomolecules 15 (4), 592. doi:10.3390/biom15040592
Gorecki, A. M., Preskey, L., Bakeberg, M. C., Kenna, J. E., Gildenhuys, C., MacDougall, G., et al. (2019). Altered gut microbiome in Parkinson's disease and the influence of lipopolysaccharide in a human α-synuclein over-expressing mouse model. Front. Neurosci. 13, 839. doi:10.3389/fnins.2019.00839
Gorecki, A. M., Anyaegbu, C. C., Fitzgerald, M., Fuller, K. A., and Anderton, R. S. (2025). Imaging flow cytometry reveals LPS-induced changes to intracellular intensity and distribution of α-synuclein in a TLR4-dependent manner in STC-1 cells. Methods 234, 93–111. doi:10.1016/j.ymeth.2024.10.009
Gui, J., Sun, X., Wen, S., Liu, X., Qin, B., and Sang, M. (2024). Resveratrol protects dopaminergic neurons in a mouse model of Parkinson's disease by regulating the gut-brain axis via inhibiting the TLR4 signaling pathway. Nan Fang. Yi Ke Da Xue Xue Bao 44 (2), 270–279. doi:10.12122/j.issn.1673-4254.2024.02.09
Guo, M., He, S., Song, W., Mai, J., Yuan, X., Huang, Y., et al. (2024). The Lachnospiraceae-butyric acid axis and its role in glucocorticoid-associated osteonecrosis. J. Transl. Med. 22 (1), 1015. doi:10.1186/s12967-024-05813-4
He, D., Gao, X., Wen, J., Zhang, Y., Yang, S., Sun, X., et al. (2024). Orally administered neohesperidin attenuates MPTP-induced neurodegeneration by inhibiting inflammatory responses and regulating intestinal flora in mice. Food Funct. 15 (3), 1460–1475. doi:10.1039/d3fo04714h
Heravi, F. S., Naseri, K., and Hu, H. (2023). Gut microbiota composition in patients with neurodegenerative disorders (Parkinson's and Alzheimer's) and healthy controls: a systematic review. Nutrients 15 (20), 4365. doi:10.3390/nu15204365
Hilton, D., Stephens, M., Kirk, L., Edwards, P., Potter, R., Zajicek, J., et al. (2014). Accumulation of α-synuclein in the bowel of patients in the pre-clinical phase of Parkinson's disease. Acta Neuropathol. 127 (2), 235–241. doi:10.1007/s00401-013-1214-6
Hosoi, T., Okuma, Y., Matsuda, T., and Nomura, Y. (2005). Novel pathway for LPS-induced afferent vagus nerve activation: possible role of nodose ganglion. Auton. Neurosci. 120 (1-2), 104–107. doi:10.1016/j.autneu.2004.11.012
Hou, Y. F., Shan, C., Zhuang, S. Y., Zhuang, Q. Q., Ghosh, A., Zhu, K. C., et al. (2021). Gut microbiota-derived propionate mediates the neuroprotective effect of osteocalcin in a mouse model of Parkinson's disease. Microbiome 9 (1), 34. doi:10.1186/s40168-020-00988-6
Hou, K., Wu, Z. X., Chen, X. Y., Wang, J. Q., Zhang, D., Xiao, C., et al. (2022). Microbiota in health and diseases. Signal Transduct. Target Ther. 7 (1), 135. doi:10.1038/s41392-022-00974-4
Hou, T., Liu, X., Zhang, S., Tang, K., Liu, S., Liu, J., et al. (2025). Anti-inflammatory effects of the fucoidan from sea cucumber apostichopus japonicus. Mar. Biotechnol. (NY) 27 (1), 32. doi:10.1007/s10126-025-10410-7
Hoyles, L., Snelling, T., Umlai, U. K., Nicholson, J. K., Carding, S. R., Glen, R. C., et al. (2018). Microbiome-host systems interactions: protective effects of propionate upon the blood-brain barrier. Microbiome 6 (1), 55. doi:10.1186/s40168-018-0439-y
Hu, G., Bidel, S., Jousilahti, P., Antikainen, R., and Tuomilehto, J. (2007). Coffee and tea consumption and the risk of Parkinson's disease. Mov. Disord. 22 (15), 2242–2248. doi:10.1002/mds.21706
Hu, X., Yu, L., Li, Y., Li, X., Zhao, Y., Xiong, L., et al. (2024). Piperine improves levodopa availability in the 6-OHDA-lesioned rat model of Parkinson's disease by suppressing gut bacterial tyrosine decarboxylase. CNS Neurosci. Ther. 30 (2), e14383. doi:10.1111/cns.14383
Huang, W. (2024). The study of Akkermansia muciniphila regulating SCFAs to alleviate neuroinflammation and promote neurogenesisin Parkinson’s Disease mice. China: Jiangnan University. doi:10.27169/d.cnki.gwqgu.2024.000297
Huang, T., Shi, H., Xu, Y., and Ji, L. (2021). The gut microbiota metabolite propionate ameliorates intestinal epithelial barrier dysfunction-mediated Parkinson's disease via the AKT signaling pathway. Neuroreport 32 (3), 244–251. doi:10.1097/wnr.0000000000001585
Huang, S. Y., Su, Z. Y., Han, Y. Y., Liu, L., Shang, Y. J., Mai, Z. F., et al. (2023). Cordycepin improved the cognitive function through regulating adenosine A(2A) receptors in MPTP induced Parkinson's disease mice model. Phytomedicine 110, 154649. doi:10.1016/j.phymed.2023.154649
Huang, X., Wei, P., Fang, C., Yu, M., Yang, S., Qiu, L., et al. (2024). Compromised endothelial Wnt/β-catenin signaling mediates the blood-brain barrier disruption and leads to neuroinflammation in endotoxemia. J. Neuroinflammation 21 (1), 265. doi:10.1186/s12974-024-03261-x
Huh, E., Choi, J. G., Noh, D., Yoo, H. S., Ryu, J., Kim, N. J., et al. (2020). Ginger and 6-shogaol protect intestinal tight junction and enteric dopaminergic neurons against 1-methyl-4-phenyl 1,2,3,6-tetrahydropyridine in mice. Nutr. Neurosci. 23 (6), 455–464. doi:10.1080/1028415x.2018.1520477
Huh, E., Choi, J. G., Choi, Y., Ju, I. G., Kim, B., Shin, Y. J., et al. (2023a). P. mirabilis-derived pore-forming haemolysin, HpmA drives intestinal alpha-synuclein aggregation in a mouse model of neurodegeneration. EBioMedicine 98, 104887. doi:10.1016/j.ebiom.2023.104887
Huh, E., Choi, J. G., Choi, Y., Ju, I. G., Noh, D., Shin, D. Y., et al. (2023b). 6-Shogaol, an active ingredient of ginger, improves intestinal and brain abnormalities in Proteus mirabilis-induced Parkinson's disease mouse model. Biomol. Ther. Seoul. 31 (4), 417–424. doi:10.4062/biomolther.2023.098
Islam, S., Mir, A. R., Arfat, M. Y., Khan, F., Zaman, M., Ali, A., et al. (2018). Structural and immunological characterization of hydroxyl radical modified human IgG: clinical correlation in rheumatoid arthritis. Spectrochim. Acta A Mol. Biomol. Spectrosc. 194, 194–201. doi:10.1016/j.saa.2018.01.026
Jiang, L., Li, J. C., Tang, B. S., and Guo, J. F. (2023). Associations between gut microbiota and Parkinson disease: a bidirectional Mendelian randomization analysis. Eur. J. Neurol. 30 (11), 3471–3477. doi:10.1111/ene.15848
Jiang, T., Wang, Y., Fan, W., Lu, Y., Zhang, G., Li, J., et al. (2025). Intestinal microbiota distribution and changes in different stages of Parkinson's disease: a meta-analysis, bioinformatics analysis and in vivo simulation. Biosci. Trends 19 (1), 87–101. doi:10.5582/bst.2024.01352
Jo, S., Oh, J. H., Lee, E. J., Choi, M., Lee, J., Lee, S., et al. (2025). Mitochondrial DNA copy number as a potential biomarker for the severity of motor symptoms and prognosis in Parkinson's disease. Mov. Disord. 40 (3), 502–510. doi:10.1002/mds.30098
Johnson, S. L., Park, H. Y., Vattem, D. A., Grammas, P., Ma, H., and Seeram, N. P. (2020). Equol, a blood-brain barrier permeable gut microbial metabolite of dietary isoflavone Daidzein, exhibits neuroprotective effects against neurotoxins induced toxicity in human neuroblastoma SH-SY5Y cells and Caenorhabditis elegans. Plant Foods Hum. Nutr. 75 (4), 512–517. doi:10.1007/s11130-020-00840-0
Kalkan, A. E., BinMowyna, M. N., Raposo, A., Ahmad, M. F., Ahmed, F., Otayf, A. Y., et al. (2025). Beyond the gut: unveiling butyrate's global health impact through gut health and dysbiosis-related conditions: a narrative review. Nutrients 17 (8), 1305. doi:10.3390/nu17081305
Kalyanaraman, B., Cheng, G., and Hardy, M. (2024). Gut microbiome, short-chain fatty acids, alpha-synuclein, neuroinflammation, and ROS/RNS: relevance to Parkinson's disease and therapeutic implications. Redox Biol. 71, 103092. doi:10.1016/j.redox.2024.103092
Kam, T. I., Hinkle, J. T., Dawson, T. M., and Dawson, V. L. (2020). Microglia and astrocyte dysfunction in Parkinson's disease. Neurobiol. Dis. 144, 105028. doi:10.1016/j.nbd.2020.105028
Kars, M. E., Wu, Y., Stenson, P. D., Cooper, D. N., Burisch, J., Peter, I., et al. (2024). The landscape of rare genetic variation associated with inflammatory bowel disease and Parkinson's disease comorbidity. Genome Med. 16 (1), 66. doi:10.1186/s13073-024-01335-2
Kaur, H., Swadia, D., and Sinha, S. (2024). Bile acids as modulators of α-synuclein aggregation: implications for Parkinson's therapy. ACS Chem. Neurosci. 15 (21), 4055–4065. doi:10.1021/acschemneuro.4c00459
Kim, S., Kwon, S. H., Kam, T. I., Panicker, N., Karuppagounder, S. S., Lee, S., et al. (2019). Transneuronal propagation of pathologic α-synuclein from the gut to the brain models Parkinson's disease. Neuron 103 (4), 627–641. doi:10.1016/j.neuron.2019.05.035
Kiriyama, Y., and Nochi, H. (2019). The biosynthesis, signaling, and neurological functions of bile acids. Biomolecules 9 (6), 232. doi:10.3390/biom9060232
Korczowska-Łącka, I., Hurła, M., Banaszek, N., Kobylarek, D., Szymanowicz, O., Kozubski, W., et al. (2023). Selected biomarkers of oxidative stress and energy metabolism disorders in neurological diseases. Mol. Neurobiol. 60 (7), 4132–4149. doi:10.1007/s12035-023-03329-4
Lai, X., Wu, J., Kou, X., Zhang, Y., Shen, M., Yu, M., et al. (2025). Exogenous α-synuclein induces oxidative damage to dopaminergic neurons through p-NMDAR2B/Nur77. Mol. Neurobiol. 62 (5), 5664–5678. doi:10.1007/s12035-024-04625-3
Lashuel, H. A., Overk, C. R., Oueslati, A., and Masliah, E. (2013). The many faces of α-synuclein: from structure and toxicity to therapeutic target. Nat. Rev. Neurosci. 14 (1), 38–48. doi:10.1038/nrn3406
Lau, K., Kotzur, R., and Richter, F. (2024). Blood-brain barrier alterations and their impact on Parkinson's disease pathogenesis and therapy. Transl. Neurodegener. 13 (1), 37. doi:10.1186/s40035-024-00430-z
Li, J., Zhang, L., Wu, T., Li, Y., Zhou, X., and Ruan, Z. (2021a). Indole-3-propionic acid improved the intestinal barrier by enhancing epithelial barrier and mucus barrier. J. Agric. Food Chem. 69 (5), 1487–1495. doi:10.1021/acs.jafc.0c05205
Li, P., Killinger, B. A., Ensink, E., Beddows, I., Yilmaz, A., Lubben, N., et al. (2021b). Gut microbiota dysbiosis is associated with elevated bile acids in Parkinson's disease. Metabolites 11 (1), 29. doi:10.3390/metabo11010029
Li, J., Meng, P., Zhang, J., and He, M. (2022a). Effect of berberine hydrochloride on the diversity of intestinal flora in Parkinson's disease patients. Contrast Media Mol. Imaging 2022, 8381870. doi:10.1155/2022/8381870
Li, Z., Zhou, J., Liang, H., Ye, L., Lan, L., Lu, F., et al. (2022b). Differences in alpha diversity of gut microbiota in neurological diseases. Front. Neurosci. 16, 879318. doi:10.3389/fnins.2022.879318
Li, S., Wang, Z., Liu, G., and Chen, M. (2024a). Neurodegenerative diseases and catechins: (-)-epigallocatechin-3-gallate is a modulator of chronic neuroinflammation and oxidative stress. Front. Nutr. 11, 1425839. doi:10.3389/fnut.2024.1425839
Li, X., Hu, M., Zhou, X., Yu, L., Qin, D., Wu, J., et al. (2024b). Hederagenin inhibits mitochondrial damage in Parkinson's disease via mitophagy induction. Free Radic. Biol. Med. 224, 740–756. doi:10.1016/j.freeradbiomed.2024.09.030
Li, W., Zhao, Z., Wang, L., He, J., Liu, Y., and Han, Q. (2025a). Curcumin improving behavioral deficits in Parkinson’s disease mice via modulation of gut microbiota. Acta Anat. Sin. 56 (02), 143–149. doi:10.16098/j.issn.0529-1356.2025.02.003
Li, X. N., Fan, L. L., Zhu, Q., Liu, J. H., Xie, Z. Y., Cao, J. L., et al. (2025b). Blue honeysuckle (Lonicera caerulea L.)-anthocyanins and cyanidin-3-O-glucoside protect dopaminergic neurons against ferroptosis by activating the Nrf2-GPX7 axis. Free Radic. Biol. Med. 239, 242–256. doi:10.1016/j.freeradbiomed.2025.07.038
Liang, D., Liu, H., Jin, R., Feng, R., Wang, J., Qin, C., et al. (2023). Escherichia coli triggers α-synuclein pathology in the LRRK2 transgenic mouse model of PD. Gut Microbes 15 (2), 2276296. doi:10.1080/19490976.2023.2276296
Liao, P. H., Tung, H. Y., Lim, W. S., Jang, J. R., Li, H., Shun, C. T., et al. (2024). Impaired gut barrier integrity and reduced colonic expression of free fatty acid receptors in patients with Parkinson's disease. Neurol. Sci. 45 (11), 5297–5307. doi:10.1007/s10072-024-07641-2
Lin, C. H., Chen, C. C., Chiang, H. L., Liou, J. M., Chang, C. M., Lu, T. P., et al. (2019). Altered gut microbiota and inflammatory cytokine responses in patients with Parkinson's disease. J. Neuroinflammation 16 (1), 129. doi:10.1186/s12974-019-1528-y
Liu, X., Yang, L., Li, G., Jiang, Y., Zhang, G., and Ling, J. (2023). A novel promising neuroprotective agent: Ganoderma lucidum polysaccharide. Int. J. Biol. Macromol. 229, 168–180. doi:10.1016/j.ijbiomac.2022.12.276
Liu, Y., Li, H., Yang, M., Guo, J., Sun, Z., Wang, S., et al. (2024a). Sika deer Velvet antler peptide exerts neuroprotective effect in a Parkinson's disease model via regulating oxidative damage and gut microbiota. Pharm. (Basel) 17 (7), 972. doi:10.3390/ph17070972
Liu, Z., Wu, S., Zhang, W., Cui, H., Zhang, J., Yin, X., et al. (2024b). Cordycepin mitigates dextran sulfate sodium-induced colitis through improving gut microbiota composition and modulating Th1/Th2 and Th17/Treg balance. Biomed. Pharmacother. 180, 117394. doi:10.1016/j.biopha.2024.117394
Liu, Z. M., Zhang, X. L., Sun, Y. L., Zheng, L. F., Zhang, C. F., Yu, X., et al. (2025). Berberine improves depression-like behaviors and gastrointestinal dysfunction in 6-hydroxydopamine-induced Parkinson's disease rats. Neurogastroenterol. Motil. 37 (7), e70033. doi:10.1111/nmo.70033
Luo, X., Wang, G., Wang, J., Feng, Y., Huang, X., Bao, C., et al. (2023). Effect of arabinoxylan on behavior and intestinal flora in rats with Parkinson’s disease. J. Youjiang Med. Univ. Natl. 45 (02), 190–194+207. doi:10.3969/j.issn.1001-5817.2023.02.004
Ma, X., and Lieberman, J. (2024). Poking holes in the blood-brain barrier. Immunity 57 (6), 1192–1194. doi:10.1016/j.immuni.2024.05.012
Ma, N., He, T., Johnston, L. J., and Ma, X. (2020). Host-microbiome interactions: the aryl hydrocarbon receptor as a critical node in tryptophan metabolites to brain signaling. Gut Microbes 11 (5), 1203–1219. doi:10.1080/19490976.2020.1758008
Ma, J., Yang, Y., Zang, C., Chen, Q., Jiang, Y., Dong, Y., et al. (2025). Inhibiting mitochondrial excessive fission alleviates the neuronal damage in Parkinson's disease via regulating PGC-1α mediated mitochondrial biogenesis. Exp. Neurol. 391, 115288. doi:10.1016/j.expneurol.2025.115288
Mahnoor, , Jahan, S., Elahi, L., Zakria, M., and Ikram, M. (2025). Alpha-linolenic acid for mitigating neuroinflammation and dopaminergic neuronal loss in Parkinson's disease: insights from in vivo and in silico studies. Clin. Exp. Pharmacol. Physiol. 52 (6), e70043. doi:10.1111/1440-1681.70043
Martín, R., Chamignon, C., Mhedbi-Hajri, N., Chain, F., Derrien, M., Escribano-Vázquez, U., et al. (2019). The potential probiotic Lactobacillus rhamnosus CNCM I-3690 strain protects the intestinal barrier by stimulating both mucus production and cytoprotective response. Sci. Rep. 9 (1), 5398. doi:10.1038/s41598-019-41738-5
Marzouk, N. H., Rashwan, H. H., El-Hadidi, M., Ramadan, R., and Mysara, M. (2025). Proinflammatory and GABA eating bacteria in Parkinson's disease gut microbiome from a meta-analysis prospective. NPJ Park. Dis. 11 (1), 145. doi:10.1038/s41531-025-00950-z
Matarrese, P., Falzano, L., Fabbri, A., Gambardella, L., Frank, C., Geny, B., et al. (2007). Clostridium difficile toxin B causes apoptosis in epithelial cells by thrilling mitochondria. Involvement of ATP-sensitive mitochondrial potassium channels. J. Biol. Chem. 282 (12), 9029–9041. doi:10.1074/jbc.M607614200
Mayneris-Perxachs, J., Moreno-Navarrete, J. M., and Fernández-Real, J. M. (2022). The role of iron in host-microbiota crosstalk and its effects on systemic glucose metabolism. Nat. Rev. Endocrinol. 18 (11), 683–698. doi:10.1038/s41574-022-00721-3
Mehanna, M., AbuRaya, S., Ahmed, S. M., Ashmawy, G., Ibrahim, A., and AbdelKhaliq, E. (2023). Study of the gut microbiome in Egyptian patients with Parkinson's Disease. BMC Microbiol. 23 (1), 196. doi:10.1186/s12866-023-02933-7
Mehreen, M., Ali, M., Tariq, H., Noor, A., Mumtaz, S., and Zafar, S. (2025). Chenodeoxycholic Acid-Mediated neuroprotection via α-synuclein and BDNF Modulation in MPTP-Induced mouse model of Parkinson's disease. Neuroscience 573, 442–450. doi:10.1016/j.neuroscience.2025.03.050
Meng, T., Zhang, Y., Huang, J., Pandey, V., Fu, S., and Ma, S. (2024). Rubusoside mitigates neuroinflammation and cellular apoptosis in Parkinson's disease, and alters gut microbiota and metabolite composition. Phytomedicine 124, 155309. doi:10.1016/j.phymed.2023.155309
Menozzi, E., Schapira, A. H. V., and Borghammer, P. (2025). The gut-brain Axis in Parkinson disease: emerging concepts and therapeutic implications. Mov. Disord. Clin. Pract. 12 (7), 904–916. doi:10.1002/mdc3.70029
Milani, C., Duranti, S., Bottacini, F., Casey, E., Turroni, F., Mahony, J., et al. (2017). The first microbial colonizers of the human gut: composition, activities, and health implications of the infant gut microbiota. Microbiol. Mol. Biol. Rev. 81 (4), e00036-17. doi:10.1128/mmbr.00036-17
Mohammed, N. N., Tadros, M. G., and George, M. Y. (2024). Empagliflozin repurposing in Parkinson's disease; modulation of oxidative stress, neuroinflammation, AMPK/SIRT-1/PGC-1α, and wnt/β-catenin pathways. Inflammopharmacology 32 (1), 777–794. doi:10.1007/s10787-023-01384-w
Morris, H. R., Spillantini, M. G., Sue, C. M., and Williams-Gray, C. H. (2024). The pathogenesis of Parkinson's disease. Lancet 403 (10423), 293–304. doi:10.1016/s0140-6736(23)01478-2
Murros, K. E. (2022). Hydrogen sulfide produced by gut bacteria may induce Parkinson's disease. Cells 11 (6), 978. doi:10.3390/cells11060978
Nakahara, K., Nakane, S., Ishii, K., Ikeda, T., and Ando, Y. (2023). Gut microbiota of Parkinson's disease in an appendectomy cohort: a preliminary study. Sci. Rep. 13 (1), 2210. doi:10.1038/s41598-023-29219-2
Ni, C., Wang, L., Bai, Y., Huang, F., Shi, H., Wu, H., et al. (2025). Taurochenodeoxycholic acid activates autophagy and suppresses inflammatory responses in microglia of MPTP-induced Parkinson's disease mice via AMPK/mTOR, AKT/NFκB and Pink1/Parkin signaling pathways mediated by Takeda G protein-coupled receptor 5. Free Radic. Biol. Med. 235, 347–363. doi:10.1016/j.freeradbiomed.2025.04.053
Niu, B., Pan, T., Xiao, Y., Wang, H., Zhu, J., Tian, F., et al. (2025). The therapeutic potential of dietary intervention: based on the mechanism of a tryptophan derivative-indole propionic acid on metabolic disorders. Crit. Rev. Food Sci. Nutr. 65 (9), 1729–1748. doi:10.1080/10408398.2023.2299744
Ohara, T. E., and Hsiao, E. Y. (2025). Microbiota-neuroepithelial signalling across the gut-brain axis. Nat. Rev. Microbiol. 23 (6), 371–384. doi:10.1038/s41579-024-01136-9
Older, E. A., Mitchell, M. K., Campbell, A., Lian, X., Madden, M., Wang, Y., et al. (2025). Human gut commensal Alistipes timonensis modulates the host lipidome and delivers anti-inflammatory outer membrane vesicles to suppress colitis in an Il10-deficient mouse model. Gut Microbes 17 (1), 2517380. doi:10.1080/19490976.2025.2517380
Ortiz de Ora, L., Balsamo, J. M., Uyeda, K. S., and Bess, E. N. (2024). Discovery of a gut bacterial metabolic pathway that drives α-synuclein aggregation. ACS Chem. Biol. 19 (4), 1011–1021. doi:10.1021/acschembio.4c00095
Pan, M., Qian, C., Huo, S., Wu, Y., Zhao, X., Ying, Y., et al. (2025). Gut-derived lactic acid enhances tryptophan to 5-hydroxytryptamine in regulation of anxiety via Akkermansia muciniphila. Gut Microbes 17 (1), 2447834. doi:10.1080/19490976.2024.2447834
Pang, S., Ren, Z., Ding, H., and Chan, P. (2026). Short-chain fatty acids mediate enteric and central nervous system homeostasis in Parkinson's disease: innovative therapies and their translation. Neural Regen. Res. 21 (3), 938–956. doi:10.4103/nrr.Nrr-d-24-01265
Pellegrini, C., D'Antongiovanni, V., Miraglia, F., Rota, L., Benvenuti, L., Di Salvo, C., et al. (2022). Enteric α-synuclein impairs intestinal epithelial barrier through caspase-1-inflammasome signaling in Parkinson's disease before brain pathology. NPJ Park. Dis. 8 (1), 9. doi:10.1038/s41531-021-00263-x
Perez-Pardo, P., Dodiya, H. B., Engen, P. A., Forsyth, C. B., Huschens, A. M., Shaikh, M., et al. (2019). Role of TLR4 in the gut-brain axis in Parkinson's disease: a translational study from men to mice. Gut 68 (5), 829–843. doi:10.1136/gutjnl-2018-316844
Pervin, M., Unno, K., Takagaki, A., Isemura, M., and Nakamura, Y. (2019). Function of green tea catechins in the brain: epigallocatechin gallate and its metabolites. Int. J. Mol. Sci. 20 (15), 3630. doi:10.3390/ijms20153630
Peter, I., Dubinsky, M., Bressman, S., Park, A., Lu, C., Chen, N., et al. (2018). Anti-tumor necrosis factor therapy and incidence of Parkinson disease among patients with inflammatory bowel disease. JAMA Neurol. 75 (8), 939–946. doi:10.1001/jamaneurol.2018.0605
Peterson, C. T. (2020). Dysfunction of the microbiota-gut-brain Axis in neurodegenerative disease: the Promise of therapeutic modulation with prebiotics, medicinal herbs, probiotics, and synbiotics. J. Evid. Based Integr. Med. 25, 2515690x20957225. doi:10.1177/2515690x20957225
Picón-Pagès, P., Garcia-Buendia, J., and Muñoz, F. J. (2019). Functions and dysfunctions of nitric oxide in brain. Biochim. Biophys. Acta Mol. Basis Dis. 1865 (8), 1949–1967. doi:10.1016/j.bbadis.2018.11.007
Plotegher, N., Gratton, E., and Bubacco, L. (2014). Number and Brightness analysis of alpha-synuclein oligomerization and the associated mitochondrial morphology alterations in live cells. Biochim. Biophys. Acta 1840 (6), 2014–2024. doi:10.1016/j.bbagen.2014.02.013
Popovic, R., Mukherjee, A., Leal, N. S., Morris, L., Yu, Y., Loh, S. H. Y., et al. (2023). Blocking dPerk in the intestine suppresses neurodegeneration in a Drosophila model of Parkinson's disease. Cell Death Dis. 14 (3), 206. doi:10.1038/s41419-023-05729-9
Pradeloux, S., Coulombe, K., Ouamba, A. J. K., Isenbrandt, A., Calon, F., Roy, D., et al. (2024). Oral trehalose intake modulates the microbiota-gut-brain Axis and is neuroprotective in a synucleinopathy mouse model. Nutrients 16 (19), 3309. doi:10.3390/nu16193309
Pramod Kumar, P., and Harish Prashanth, K. V. (2020). Diet with low molecular weight chitosan exerts neuromodulation in rotenone induced Drosophila model of Parkinson's disease. Food Chem. Toxicol. 146, 111860. doi:10.1016/j.fct.2020.111860
Qin, J., Li, R., Raes, J., Arumugam, M., Burgdorf, K. S., Manichanh, C., et al. (2010). A human gut microbial gene catalogue established by metagenomic sequencing. Nature 464 (7285), 59–65. doi:10.1038/nature08821
Qin, W., Wei, B., Ren, P., Chang, Y., Xue, C., and Tang, Q. (2025). Fucoidan from Apostichopus japonicus enhances intestinal barrier function and promotes intestinal immunity via regulating the gut microbiota and tryptophan metabolism. Int. J. Biol. Macromol. 301, 139929. doi:10.1016/j.ijbiomac.2025.139929
Qu, Y., An, K., Wang, D., Yu, H., Li, J., Min, Z., et al. (2025). Short-chain fatty acid aggregates alpha-synuclein accumulation and neuroinflammation via GPR43-NLRP3 signaling pathway in a model Parkinson's disease. Mol. Neurobiol. 62 (5), 6612–6625. doi:10.1007/s12035-025-04726-7
Salminen, A. (2023). Activation of aryl hydrocarbon receptor (AhR) in Alzheimer's disease: role of tryptophan metabolites generated by gut host-microbiota. J. Mol. Med. Berl. 101 (3), 201–222. doi:10.1007/s00109-023-02289-5
Samaranch, L., Lorenzo-Betancor, O., Arbelo, J. M., Ferrer, I., Lorenzo, E., Irigoyen, J., et al. (2010). PINK1-linked parkinsonism is associated with Lewy body pathology. Brain 133 (Pt 4), 1128–1142. doi:10.1093/brain/awq051
Sampson, T. R., Debelius, J. W., Thron, T., Janssen, S., Shastri, G. G., Ilhan, Z. E., et al. (2016). Gut microbiota regulate motor deficits and neuroinflammation in a model of Parkinson's disease. Cell 167 (6), 1469–1480. doi:10.1016/j.cell.2016.11.018
Sampson, T. R., Wallen, Z. D., Won, W. J., Standaert, D. G., Payami, H., and Harms, A. S. (2025). Alpha synuclein overexpression can drive microbiome dysbiosis in mice. Sci. Rep. 15 (1), 4014. doi:10.1038/s41598-024-82139-7
Scheperjans, F., Aho, V., Pereira, P. A., Koskinen, K., Paulin, L., Pekkonen, E., et al. (2015). Gut microbiota are related to Parkinson's disease and clinical phenotype. Mov. Disord. 30 (3), 350–358. doi:10.1002/mds.26069
Sergi, C. M. (2022). Epigallocatechin gallate for Parkinson's disease. Clin. Exp. Pharmacol. Physiol. 49 (10), 1029–1041. doi:10.1111/1440-1681.13691
Shang, M., Ning, J., Zang, C., Ma, J., Yang, Y., Wan, Z., et al. (2025). Microbial metabolite 3-indolepropionic acid alleviated PD pathologies by decreasing enteric glia cell gliosis via suppressing IL-13Rα1 related signaling pathways. Acta Pharm. Sin. B 15 (4), 2024–2038. doi:10.1016/j.apsb.2025.02.029
Shao, Y., Li, T., Liu, Z., Wang, X., Xu, X., Li, S., et al. (2021). Comprehensive metabolic profiling of Parkinson's disease by liquid chromatography-mass spectrometry. Mol. Neurodegener. 16 (1), 4. doi:10.1186/s13024-021-00425-8
Shao, X., Wu, T., Li, M., Zheng, M., Lin, H., and Qi, X. (2025). Enterococcus faecalis exerts neuroprotective effects via the vagus nerve in a mouse model of Parkinson's disease. Mol. Neurobiol. 62 (6), 7875–7891. doi:10.1007/s12035-025-04741-8
Sharma, H., Sharma, N., and An, S. S. A. (2023). Unique bioactives from zombie fungus (Cordyceps) as promising multitargeted neuroprotective agents. Nutrients 16 (1), 102. doi:10.3390/nu16010102
Shenvi, R. A. (2024). Natural product synthesis in the 21st century: beyond the mountain top. ACS Cent. Sci. 10 (3), 519–528. doi:10.1021/acscentsci.3c01518
Shimazu, R., Anada, M., Miyaguchi, A., Nomi, Y., and Matsumoto, H. (2021). Evaluation of blood-brain barrier permeability of polyphenols, anthocyanins, and their metabolites. J. Agric. Food Chem. 69 (39), 11676–11686. doi:10.1021/acs.jafc.1c02898
Shu, Y., Hong, W., Liu, J., and Zhu, X. (2025). Exploring the Association of dietary index for gut microbiota with Parkinson's disease and depression: insights from NHANES. J. Affect Disord. 386, 119461. doi:10.1016/j.jad.2025.119461
Silva-Reis, R., Castro-Ribeiro, C., Gonçalves, M., Ferreira, T., Pires, M. J., Iglesias-Aguirre, C. E., et al. (2022). An integrative approach to characterize the early phases of dimethylhydrazine-induced colorectal carcinogenesis in the rat. Biomedicines 10 (2), 409. doi:10.3390/biomedicines10020409
Singh, P., Hanson, P. S., and Morris, C. M. (2017). SIRT1 ameliorates oxidative stress induced neural cell death and is down-regulated in Parkinson's disease. BMC Neurosci. 18 (1), 46. doi:10.1186/s12868-017-0364-1
Singh, Y., Trautwein, C., Romani, J., Salker, M. S., Neckel, P. H., Fraccaroli, I., et al. (2023). Overexpression of human alpha-Synuclein leads to dysregulated microbiome/metabolites with ageing in a rat model of Parkinson disease. Mol. Neurodegener. 18 (1), 44. doi:10.1186/s13024-023-00628-1
Song, S. (2023). Tissue distribution of Polymannuronic Acid in mice and its effects on gut microbiota. China: Shandong University. doi:10.27272/d.cnki.gshdu.2023.000116
Sorboni, S. G., Moghaddam, H. S., Jafarzadeh-Esfehani, R., and Soleimanpour, S. (2022). A comprehensive review on the role of the gut microbiome in human neurological disorders. Clin. Microbiol. Rev. 35 (1), e0033820. doi:10.1128/cmr.00338-20
Subedi, L., Ji, E., Shin, D., Jin, J., Yeo, J. H., and Kim, S. Y. (2017). Equol, a dietary daidzein gut metabolite attenuates microglial activation and potentiates neuroprotection in vitro. Nutrients 9 (3), 207. doi:10.3390/nu9030207
Sun, X., Feng, S., Qin, B., Ye, J., Xie, L., Gui, J., et al. (2024a). Integrated multi-omics profiling highlights the benefits of resveratrol hydroxypropyl-β-cyclodextrin inclusion complex for A53T transgenic mice through the microbiota-gut-brain axis. Food Funct. 15 (3), 1489–1512. doi:10.1039/d3fo03667g
Sun, X., Gu, R., and Bai, J. (2024b). Differentiation and regulation of CD4(+) T cell subsets in Parkinson's disease. Cell Mol. Life Sci. 81 (1), 352. doi:10.1007/s00018-024-05402-0
Sun, X., Qin, B., Guo, A., Gui, J., Weng, J., Ye, J., et al. (2025). Withaferin A maintained microbiome and metabolome features in A53T transgenic mice via multi-omics integrated analysis. Phytomedicine 141, 156725. doi:10.1016/j.phymed.2025.156725
Tan, A. H., Lim, S. Y., and Lang, A. E. (2022). The microbiome-gut-brain axis in Parkinson disease - from basic research to the clinic. Nat. Rev. Neurol. 18 (8), 476–495. doi:10.1038/s41582-022-00681-2
Tao, J., An, Y., Xu, L., Wang, Y., Wang, C., Li, P., et al. (2023). The protective role of microbiota in the prevention of MPTP/P-induced Parkinson's disease by resveratrol. Food Funct. 14 (10), 4647–4661. doi:10.1039/d2fo03379h
Techaniyom, P., Korsirikoon, C., Rungruang, T., Pakaprot, N., Prombutara, P., Mukda, S., et al. (2024). Cold-pressed perilla seed oil: investigating its protective influence on the gut-brain axis in mice with rotenone-induced Parkinson's disease. Food Sci. Nutr. 12 (9), 6259–6283. doi:10.1002/fsn3.4265
Thapa, R., Moglad, E., Afzal, M., Gupta, G., Bhat, A. A., Hassan Almalki, W., et al. (2024). The role of sirtuin 1 in ageing and neurodegenerative disease: a molecular perspective. Ageing Res. Rev. 102, 102545. doi:10.1016/j.arr.2024.102545
Uemura, N., Yagi, H., Uemura, M. T., Hatanaka, Y., Yamakado, H., and Takahashi, R. (2019). Correction to: inoculation of α-synuclein preformed fibrils into the mouse gastrointestinal tract induces Lewy body-like aggregates in the brainstem via the vagus nerve. Mol. Neurodegener. 14 (1), 31. doi:10.1186/s13024-019-0331-7
Unger, M. M., Spiegel, J., Dillmann, K. U., Grundmann, D., Philippeit, H., Bürmann, J., et al. (2016). Short chain fatty acids and gut microbiota differ between patients with Parkinson's disease and age-matched controls. Park. Relat. Disord. 32, 66–72. doi:10.1016/j.parkreldis.2016.08.019
van der Maden, M. M., Verbeek, M. M., and Beckers, M. (2025). Lactobacillaceae and Parkinson's disease: an apparent paradox. J. Park. Dis. 15 (2), 269–281. doi:10.1177/1877718x241312401
Wallen, Z. D., Stone, W. J., Factor, S. A., Molho, E., Zabetian, C. P., Standaert, D. G., et al. (2021). Exploring human-genome gut-microbiome interaction in Parkinson's disease. NPJ Park. Dis. 7 (1), 74. doi:10.1038/s41531-021-00218-2
Wang, H. (2024). “Investigation of the biological activity of pollen typhae flavonoids and its neuroprotective effects on Parkinson's disease in mice,” [dissertation/master's thesis]. China: Lanzhou University. doi:10.27204/d.cnki.glzhu.2024.001075
Wang, J., Xu, W., Wang, R., Cheng, R., Tang, Z., and Zhang, M. (2021a). The outer membrane protein Amuc_1100 of Akkermansia muciniphila promotes intestinal 5-HT biosynthesis and extracellular availability through TLR2 signalling. Food Funct. 12 (8), 3597–3610. doi:10.1039/d1fo00115a
Wang, Y., Tong, Q., Ma, S. R., Zhao, Z. X., Pan, L. B., Cong, L., et al. (2021b). Oral berberine improves brain dopa/dopamine levels to ameliorate Parkinson's disease by regulating gut microbiota. Signal Transduct. Target Ther. 6 (1), 77. doi:10.1038/s41392-020-00456-5
Wang, N., Feng, B. N., Hu, B., Cheng, Y. L., Guo, Y. H., and Qian, H. (2022a). Neuroprotection of chicoric acid in a mouse model of Parkinson's disease involves gut microbiota and TLR4 signaling pathway. Food Funct. 13 (4), 2019–2032. doi:10.1039/d1fo02216d
Wang, N., Li, R., Feng, B., Cheng, Y., Guo, Y., and Qian, H. (2022b). Chicoric acid prevents neuroinflammation and neurodegeneration in a mouse Parkinson's disease model: immune response and transcriptome profile of the spleen and colon. Int. J. Mol. Sci. 23 (4), 2031. doi:10.3390/ijms23042031
Wang, W., Zhu, G., Wang, Y., Li, W., Yi, S., Wang, K., et al. (2022c). Multi-Omics integration in mice with Parkinson's disease and the intervention effect of cyanidin-3-O-glucoside. Front. Aging Neurosci. 14, 877078. doi:10.3389/fnagi.2022.877078
Wang, L., Dan, Q., Xu, B., Chen, Y., and Zheng, T. (2023a). Research progress on gas signal molecular therapy for Parkinson's disease. Open Life Sci. 18 (1), 20220658. doi:10.1515/biol-2022-0658
Wang, S., Xu, C., Liu, H., Wei, W., Zhou, X., Qian, H., et al. (2023b). Connecting the gut microbiota and neurodegenerative diseases: the role of bile acids. Mol. Neurobiol. 60 (8), 4618–4640. doi:10.1007/s12035-023-03340-9
Wang, C., Yang, M., Liu, D., and Zheng, C. (2024a). Metabolic rescue of α-synuclein-induced neurodegeneration through propionate supplementation and intestine-neuron signaling in C. elegans. Cell Rep. 43 (3), 113865. doi:10.1016/j.celrep.2024.113865
Wang, F., Zhang, Z., Sun, Y., Yang, L., Guo, T., Pan, Y., et al. (2024b). Bmal1 mediates the neuroprotective effect of sodium butyrate in a mouse model of Parkinson's disease. Nan Fang. Yi Ke Da Xue Xue Bao 44 (5), 876–884. doi:10.12122/j.issn.1673-4254.2024.05.09
Wang, L., Tian, S., Ruan, S., Wei, J., Wei, S., Chen, W., et al. (2024c). Neuroprotective effects of cordycepin on MPTP-induced Parkinson's disease mice via suppressing PI3K/AKT/mTOR and MAPK-mediated neuroinflammation. Free Radic. Biol. Med. 216, 60–77. doi:10.1016/j.freeradbiomed.2024.02.023
Wang, G., Zhou, X., Pang, X., Ma, K., Li, L., Song, Y., et al. (2025a). Pharmacological effects, molecular mechanisms and strategies to improve bioavailability of curcumin in the treatment of neurodegenerative diseases. Front. Pharmacol. 16, 1625821. doi:10.3389/fphar.2025.1625821
Wang, P., Wang, R., Zhao, W., Zhao, Y., Wang, D., Zhao, S., et al. (2025b). Gut microbiota-derived 4-hydroxyphenylacetic acid from resveratrol supplementation prevents obesity through SIRT1 signaling activation. Gut Microbes 17 (1), 2446391. doi:10.1080/19490976.2024.2446391
Wang, Y., Li, Y., Gao, Q., Xu, J., Zhao, Z., Zhang, R., et al. (2025c). Effects of bilberry anthocyanins on inflammation and intestinal flora in Parkinson's disease mice. Chin. J. Microecology 37 (02), 147–154. doi:10.13381/j.cnki.cjm.202502004
Wang, Z., Ma, X., Shi, W., Zhu, W., Feng, X., Xin, H., et al. (2025d). The gut microbiota metabolite butyrate modulates acute stress-induced ferroptosis in the prefrontal cortex via the gut-brain Axis. Int. J. Mol. Sci. 26 (4), 1698. doi:10.3390/ijms26041698
Wang, Z., Sun, Y., Bai, Z., Li, M., Kong, D., and Wu, G. (2025e). Mitochondria-related genome-wide mendelian randomization identifies putatively causal genes for neurodegenerative diseases. Mov. Disord. 40 (4), 693–703. doi:10.1002/mds.30123
Wang, Y., Chen, R., Shi, G., Huang, X., Li, K., Wang, R., et al. (2026). Chitosan alleviates symptoms of Parkinson's disease by reducing acetate levels, which decreases inflammation and promotes repair of the intestinal barrier and blood-brain barrier. Neural Regen. Res. 21 (1), 377–391. doi:10.4103/nrr.Nrr-d-23-01511
Wei, C., Jiang, W., Wang, R., Zhong, H., He, H., Gao, X., et al. (2024). Brain endothelial GSDMD activation mediates inflammatory BBB breakdown. Nature 629 (8013), 893–900. doi:10.1038/s41586-024-07314-2
Wei, C., Jiang, W., Luo, M., and Shao, F. (2025). BBB breakdown caused by plasma membrane pore formation. Trends Cell Biol. doi:10.1016/j.tcb.2025.02.012
Xu, Y., Xie, M., Xue, J., Xiang, L., Li, Y., Xiao, J., et al. (2020). EGCG ameliorates neuronal and behavioral defects by remodeling gut microbiota and TotM expression in Drosophila models of Parkinson's disease. Faseb J. 34 (4), 5931–5950. doi:10.1096/fj.201903125RR
Xu, D., Zhuang, L., Gao, S., Ma, H., Cheng, J., Liu, J., et al. (2022a). Orally administered ginkgolide C attenuates DSS-induced colitis by maintaining gut barrier integrity, inhibiting inflammatory responses, and regulating intestinal flora. J. Agric. Food Chem. 70 (46), 14718–14731. doi:10.1021/acs.jafc.2c06177
Xu, L., Xiao, S., Lee, J. J., Li, X., and Zhao, Y. (2022b). Gender-related differences in tissue distribution, excretion, and metabolism studies of panaxadiol in rats and anti-inflammatory study. J. Agric. Food Chem. 70 (28), 8672–8679. doi:10.1021/acs.jafc.2c02618
Xu, L., Zhang, Z., Feng, Z., Niu, S., Yang, L., Xiao, B., et al. (2025). Study on the anti-Parkinson's disease activity mechanism and preparation of panaxadiol. Phytomedicine 136, 156296. doi:10.1016/j.phymed.2024.156296
Xue, L., Huan, Y., Chang, Y., Wang, Y., and Tang, Q. (2025). Investigating the alleviating effect of fucoidan from apostichopus japonicus on ulcerative colitis by mice experiments and in vitro simulation of human fecal fermentation. Foods 14 (4), 574. doi:10.3390/foods14040574
Yan, Y., Ren, S., Duan, Y., Lu, C., Niu, Y., Wang, Z., et al. (2021). Gut microbiota and metabolites of α-synuclein transgenic monkey models with early stage of Parkinson's disease. NPJ Biofilms Microbiomes 7 (1), 69. doi:10.1038/s41522-021-00242-3
Yang, M., Wu, C., Zhang, T., Shi, L., Li, J., Liang, H., et al. (2022). Chicoric acid: natural occurrence, chemical synthesis, biosynthesis, and their bioactive effects. Front. Chem. 10, 888673. doi:10.3389/fchem.2022.888673
Yang, S., Sun, X., Liu, D., Zhang, Y., Gao, X., He, J., et al. (2024a). Allantoin ameliorates dopaminergic neuronal damage in MPTP-induced Parkinson's disease mice via regulating oxidative damage, inflammation, and gut microbiota disorder. Food Funct. 15 (18), 9390–9408. doi:10.1039/d4fo02167c
Yang, X., Zhang, X., Ma, Y., Li, S., Wang, Q., Hong, J. S., et al. (2024b). Fucoidan ameliorates rotenone-induced Parkinsonism in mice by regulating the microbiota-gut-brain axis. Int. J. Biol. Macromol. 283 (Pt 2), 137373. doi:10.1016/j.ijbiomac.2024.137373
Yang, Z. X., Zhang, Y., Wang, Q., Zhang, L., Liu, Y. F., Zhang, Y., et al. (2024c). Addition of α-synuclein aggregates to the intestinal environment recapitulates Parkinsonian symptoms in model systems. Acta Pharmacol. Sin. 45 (1), 36–51. doi:10.1038/s41401-023-01150-2
Ye, Q. (2022). “Potential preventative effect of sea cucumber fucosylated chondroitin sulfate in Parkinson's disease mice model,” [dissertation/master's thesis]. China: Jinan University. doi:10.27167/d.cnki.gjinu.2022.002591
Yu, B., Wang, M., Teng, B., Veeraperumal, S., Cheung, P. C., Zhong, S., et al. (2023). Partially acid-hydrolyzed porphyran improved dextran sulfate sodium-induced acute colitis by modulation of gut microbiota and enhancing the mucosal barrier. J. Agric. Food Chem. 71 (19), 7299–7311. doi:10.1021/acs.jafc.2c08564
Yu, L., Hu, X., Xu, R., Zhao, Y., Xiong, L., Ai, J., et al. (2024). Piperine promotes PI3K/AKT/mTOR-mediated gut-brain autophagy to degrade α-Synuclein in Parkinson's disease rats. J. Ethnopharmacol. 322, 117628. doi:10.1016/j.jep.2023.117628
Zaa, C. A., Marcelo Á, J., An, Z., Medina-Franco, J. L., and Velasco-Velázquez, M. A. (2023). Anthocyanins: molecular aspects on their neuroprotective activity. Biomolecules 13 (11), 1598. doi:10.3390/biom13111598
Zali, A., Hajyani, S., Salari, M., Tajabadi-Ebrahimi, M., Mortazavian, A. M., and Pakpour, B. (2024). Co-administration of probiotics and vitamin D reduced disease severity and complications in patients with Parkinson's disease: a randomized controlled clinical trial. Psychopharmacol. Berl. 241 (9), 1905–1914. doi:10.1007/s00213-024-06606-9
Zhang, H., Qi, R., Zeng, Y., Tsao, R., and Mine, Y. (2020). Chinese sweet leaf tea (Rubus suavissimus) mitigates LPS-induced low-grade chronic inflammation and reduces the risk of metabolic disorders in a C57bl/6J mouse model. J. Agric. Food Chem. 68 (1), 138–146. doi:10.1021/acs.jafc.9b05975
Zhang, X. L., Huang, W. M., Tang, P. C., Sun, Y., Zhang, X., Qiu, L., et al. (2021). Anti-inflammatory and neuroprotective effects of natural cordycepin in rotenone-induced PD models through inhibiting Drp1-mediated mitochondrial fission. Neurotoxicology 84, 1–13. doi:10.1016/j.neuro.2021.02.002
Zhang, L., Ji, Q., Chen, Q., Wei, Z., Liu, S., Zhang, L., et al. (2023a). Akkermansia muciniphila inhibits tryptophan metabolism via the AhR/β-catenin signaling pathway to counter the progression of colorectal cancer. Int. J. Biol. Sci. 19 (14), 4393–4410. doi:10.7150/ijbs.85712
Zhang, X., Tang, B., and Guo, J. (2023b). Parkinson's disease and gut microbiota: from clinical to mechanistic and therapeutic studies. Transl. Neurodegener. 12 (1), 59. doi:10.1186/s40035-023-00392-8
Zhang, L. Y., Zhang, S. Y., Wen, R., Zhang, T. N., and Yang, N. (2024a). Role of histone deacetylases and their inhibitors in neurological diseases. Pharmacol. Res. 208, 107410. doi:10.1016/j.phrs.2024.107410
Zhang, S., You, J., Wei, L., Bai, Y., Liu, X., and Wang, S. (2024b). Effects of Chaihu plus Longgu Muli decoction combined with levodopa and benserazide hydrazine tablets on oxidative stress, intestinal flora and neurotrophic factors in patients with Parkinson's disease. Prog. Mod. Biomed. 24 (24), 4778–4780. doi:10.13241/j.cnki.pmb.2024.24.044
Zhang, D., Chen, K., Yu, Y., Feng, R., Cui, S. W., Zhou, X., et al. (2025a). Polysaccharide from Aloe vera gel improves intestinal stem cells dysfunction to alleviate intestinal barrier damage via 5-HT. Food Res. Int. 214, 116675. doi:10.1016/j.foodres.2025.116675
Zhang, H., Shan, A. D., Huang, Y. Y., Gao, M. X., Wan, C. H., Ye, S. Y., et al. (2025b). Transcutaneous auricular vagus nerve stimulation improves cortical functional topological properties and intracortical facilitation in patients with Parkinson's disease. NPJ Park. Dis. 11 (1), 38. doi:10.1038/s41531-025-00889-1
Zhang, Y., Liang, C., Weng, M., Zhang, Z., Zhang, L., Jiang, X., et al. (2025c). Intestinal alterations of mucosal barrier integrity, motility and enteric nerve in cynomolgus monkey model of Parkinson's disease. Exp. Neurol. 389, 115256. doi:10.1016/j.expneurol.2025.115256
Zhang, Z. L., Ye, P. H., Kong, W. X., Li, J., Zhu, S., Lin, Y. W., et al. (2025d). Upregulation of PD-1 on peripheral T cells subsets is associated with Parkinson's disease. Mol. Neurobiol. doi:10.1007/s12035-025-05153-4
Zhao, Z., Ning, J., Bao, X. Q., Shang, M., Ma, J., Li, G., et al. (2021). Fecal microbiota transplantation protects rotenone-induced Parkinson's disease mice via suppressing inflammation mediated by the lipopolysaccharide-TLR4 signaling pathway through the microbiota-gut-brain axis. Microbiome 9 (1), 226. doi:10.1186/s40168-021-01107-9
Zhao, Q., Chen, T., Ni, C., Hu, Y., Nan, Y., Lin, W., et al. (2022). Indole-3-propionic acid attenuates HI-related blood-brain barrier injury in neonatal rats by modulating the PXR signaling pathway. ACS Chem. Neurosci. 13 (19), 2897–2912. doi:10.1021/acschemneuro.2c00418
Zhao, Y., Lai, Y., Darweesh, S. K. L., Bloem, B. R., Forsgren, L., Hansen, J., et al. (2025). Gut microbial metabolites and future risk of Parkinson's disease: a metabolome-wide association study. Mov. Disord. 40 (3), 556–560. doi:10.1002/mds.30054
Zheng, S. Y., Li, H. X., Xu, R. C., Miao, W. T., Dai, M. Y., Ding, S. T., et al. (2021). Potential roles of gut microbiota and microbial metabolites in Parkinson's disease. Ageing Res. Rev. 69, 101347. doi:10.1016/j.arr.2021.101347
Zhernakova, A., Kurilshikov, A., Bonder, M. J., Tigchelaar, E. F., Schirmer, M., Vatanen, T., et al. (2016). Population-based metagenomics analysis reveals markers for gut microbiome composition and diversity. Science 352 (6285), 565–569. doi:10.1126/science.aad3369
Zhong, L., Cai, B., Wang, Q., Li, X., Xu, W., and Chen, T. (2022). Exploring the neuroprotective mechanism of curcumin inhibition of intestinal inflammation against Parkinson's disease based on the gut-brain Axis. Pharm. (Basel) 16 (1), 39. doi:10.3390/ph16010039
Zhou, Y., Chen, Y., He, H., Peng, M., Zeng, M., and Sun, H. (2023). The role of the indoles in microbiota-gut-brain axis and potential therapeutic targets: a focus on human neurological and neuropsychiatric diseases. Neuropharmacology 239, 109690. doi:10.1016/j.neuropharm.2023.109690
Zhu, Y., Li, Y., Zhang, Q., Song, Y., Wang, L., and Zhu, Z. (2022). Interactions between intestinal microbiota and neural mitochondria: a new perspective on communicating pathway from gut to brain. Front. Microbiol. 13, 798917. doi:10.3389/fmicb.2022.798917
Zhu, L., Ren, Y., Zhang, S., Guo, Y., Zong, J., and Liu, Y. (2024). Marine-derived polysaccharides: the potential agents against neurodegenerative diseases. Front. Pharmacol. 15, 1506789. doi:10.3389/fphar.2024.1506789
Keywords: Parkinson’s disease, natural product, gut microbiota, mechanisms, research progress
Citation: Jia Y, Zhang Y, Tai X, Zhao T, Zhang H and Zhou H (2025) Research progress on natural products in regulating the gut microbiota in Parkinson’s disease. Front. Pharmacol. 16:1667694. doi: 10.3389/fphar.2025.1667694
Received: 17 July 2025; Accepted: 21 August 2025;
Published: 08 September 2025.
Edited by:
Jacob Raber, Oregon Health and Science University, United StatesReviewed by:
Liu Han Deng, Chongqing Medical University, ChinaSidra Islam, Case Western Reserve University, United States
Copyright © 2025 Jia, Zhang, Tai, Zhao, Zhang and Zhou. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Haichun Zhou, ejEzOTM2NDYyNjY1QDEyNi5jb20=