 Davide Cadeddu
Davide Cadeddu Anna Loftén
Anna Loftén Karin Ademar2
Karin Ademar2 Bo Söderpalm
Bo Söderpalm Louise Adermark
Louise Adermark Mia Ericson
Mia Ericson- 1Department of Pharmacology, Institute of Neuroscience and Physiology, The Sahlgrenska Academy at University of Gothenburg, Gothenburg, Sweden
- 2Addiction Biology Unit, Department of Psychiatry and Neurochemistry, Institute of Neuroscience and Physiology, The Sahlgrenska Academy at University of Gothenburg, Gothenburg, Sweden
- 3Beroendekliniken, Sahlgrenska University Hospital, Gothenburg, Sweden
Introduction: Alcohol use disorder (AUD) causes significant morbidity and mortality globally. Ethanol’s rewarding and reinforcing effects are attributed to activation of the mesolimbic dopamine system, increasing accumbal dopamine release. While activation of accumbal glycine receptors (GlyRs) is a prerequisite for ethanol-induced dopamine signaling, multiple transmitter systems may be involved; recent research implicates the GABA-rho receptor as a prominent target. Considering the structural and functional similarities between GlyRs and GABA-rho receptors, this study aimed to define the role of GlyRs and GABA-rho receptors in regulating baseline dopamine signalling and ethanol-induced elevation of extracellular dopamine and GlyR agonists, as well as to determine their involvement in the action of the ethanol relapse-preventing drug acamprosate.
Methods: To investigate this, in vivo microdialysis was conducted in male Wistar rats.
Results and discussion: Local perfusion with either the GABA-rho receptor antagonist TPMPA or the GlyR antagonist strychnine prior to ethanol administration significantly reduced the ethanol-induced increase in dopamine levels. These findings suggest that both GlyRs and GABA-rho receptors are involved in mediating the dopamine-elevating effect of ethanol. In addition, a significant attenuation of the ethanol-induced glycine and taurine elevation was observed following both pretreatment with TPMPA and strychnine, whilst only GlyR blockade inhibited the acamprosate-induced increase of dopamine. Unlike strychnine, TPMPA alone did not alter dopamine levels, suggesting that GABA-rho receptors display features that distinguish them from GlyR. In conclusion, GABA-rho receptors regulate ethanol-induced dopamine and glycine/taurine levels within the nAc without affecting basal dopamine neurotransmission, suggesting their potential as a pharmacological target for the treatment of AUD.
1 Introduction
With 3 million deaths per year (5.3% of global deaths), alcohol consumption is one of the main risk factors of premature death worldwide, and it accounts for 5.1% of the global burden of disease (Who, 2018). Despite the high prevalence, treatment options are few and defining new interventions is imperative. Ethanol exerts its effects on the brain by altering the functionality of brain areas involved in the rewarding and reinforcing properties of drugs of abuse. Especially, the rewarding property of ethanol has been attributed to the increase in dopamine release from dopaminergic neurons located in the ventral tegmental area (VTA) projecting to the nucleus accumbens (nAc) (Di Chiara, 1997; Imperato and Di Chiara, 1986). Acamprosate is one of few approved pharmacological treatments for alcohol use disorder (AUD). Our research group, and others, have shown that acamprosate increases dopamine in the mesolimbic dopamine system (Cano-Cebrian et al., 2003; Chau et al., 2010a; Olive et al., 2002), suggesting that acamprosate may act as a partial neurochemical substitution for ethanol (Ademar et al., 2022; Ademar et al., 2023; Chau et al., 2018).
While many neurotransmitter systems may be involved in regulating ethanol-induced dopamine release (Abrahao et al., 2017), inhibition of accumbal glycine receptors (GlyRs) both reduces basal dopamine levels in the nAc (Molander and Söderpalm, 2005a), and prevents ethanol-induced dopamine elevation (Molander and Söderpalm, 2005b). Furthermore, GlyRs may be involved in the alcohol relapse-preventing effect of acamprosate, as antagonism of GlyRs also blocks the acamprosate-induced elevation of dopamine and reverses the alcohol intake reducing effect of the drug in rats (Ademar et al., 2022; Chau et al., 2010a; Chau et al., 2010b). Recently, the atypical GABAergic receptor, GABA-rho (also known as GABA-ρ or GABAC or GABAAC), has received attention as a regulator of ethanol-induced dopamine release (Yorgason et al., 2022) and voluntary ethanol consumption (Blednov et al., 2014). Unlike GABAA and GABAB receptors, GABA-rho receptors are insensitive to bicuculline, baclofen, benzodiazepines, neurosteroids, and barbiturates (Naffaa et al., 2017; Woodward et al., 1993). However, they are selectively blocked by the competitive antagonist (1,2,5,6-tetrahydropyridin-4-yl) methylphosphinic acid (TPMPA) (Bormann, 2000; Mohammadi et al., 2017; Zhang et al., 2001) and are sensitive to the GlyR agonists glycine and taurine (Ochoa-de la Paz et al., 2008). This suggests that GABA-rho receptors in certain aspects show more similarities to GlyRs than to GABA-A receptors.
Given the role of GlyRs in regulating ethanol-induced responses and the similarity between GlyRs and the GABA-rho in receptor activation, we hypothesized that GlyR and GABA-rho would display comparable mechanisms of action. To test this hypothesis, we outlined the role of GABA-rho receptors and GlyRs in regulating basal dopamine levels and ethanol-induced dopamine elevation in vivo. We further assessed their involvement in ethanol-induced elevation of glycine receptor agonists (glycine and taurine), and in controlling acamprosate-induced dopamine elevation using in vivo microdialysis in Wistar rats.
2 Materials and methods
2.1 Animals
Male Wistar RccHan rats (Inotiv, the Netherlands) weighing 250–280 g (corresponding to an age of 9–10 weeks) at arrival to the facility were group-housed at a constant room temperature of 20–22 °C, humidity of 55–65% and with a regular light/dark cycle (light on at 7:00 a.m./light off at 7:00 p.m.). The rats had free access to regular rat chow and water and were allowed to acclimatize to the new environment for 1 week prior to the initiation of any experiments. All experiments were approved by the Ethics Committee for Animal Experiments, Gothenburg, Sweden (2401/19).
2.2 Drugs
(1,2,5,6-tetrahydropyridine-4-yl)methylphosphinic acid (TPMPA) (Tocris and Cayman Chemical Company) (10 μM, 100 μM or 500 μM), strychnine hydrochloride (Sigma-Aldrich, Stockholm, Sweden) (20 μM or 200 μM), calcium-bis(N-acetylhomotaurinate) (acamprosate) (Merck, Lyon, France) (1 mM), ethanol (95% Solveco, Sweden) (300 mM), taurine (50 µM) and glycine (50 µM) (Sigma-Aldrich, Stockholm, Sweden) were used in the study. All drugs were dissolved in Ringer’s solution (140 mM NaCl, 1.2 mM CaCl2, 3.0 mM KCl, and 1.0 mM MgCl2) and administered locally via reversed microdialysis in the nAc.
2.3 Surgery
Rats were deeply anesthetized with 4% isoflurane (Baxter Medical AB, Kista, Sweden), mounted onto a stereotaxic instrument (David Kopf Instruments, Lidingö, Sweden) and placed on a heating pad to prevent hypothermia. The skull was exposed, and three holes were drilled: one above the target area, the nAc, and two for attachment of anchoring screws. An I-shaped custom-made probe was implanted into the nAc core/shell borderline region (A/P: + 1.85, M/L: −1.4 relative to bregma, D/V: −7.8 mm relative to dura mater; Paxinos and Watson 7th compact ed. 2018) and fixed using Harvard cement (DAB Dental AB, Stockholm, Sweden). An active space of 2 mm and a molecular cut-off of 20 kDa was used for the dialysis probe. To prevent dehydration, rats received saline solution (2 mL, NaCl 0.9%) via subcutaneous (s.c.) injection at the end of the surgical procedure. Metacam vet (2 mg/mL, 1 mg/kg, s. c.; Apoteket AB, Sweden) was used as perioperative analgesia. Prior to the in vivo microdialysis experiments, rats were single-housed and allowed to recover for 48 h.
2.4 In vivo microdialysis
On the day of the experiment, rats were allowed to move freely in the cage. The probe was connected, via a swivel, to a microinjection pump (U-864 Syringe Pump, AgnTho’s, Lidingö Sweden), and perfused with Ringer’s solution (rate of 2 μL/min). A 2-h equilibrium phase was performed to stabilize fluid exchange, after which samples (40 μL) were collected every 20 min. A baseline period of 80 min was followed by the drug local administration of the drug when applicable. The microdialysate was analyzed using high-performance liquid chromatography (HPLC) with electrochemical detection to determine DA concentration, as previously described (Clarke et al., 2014). Additionally, HPLC with fluorescence detection was used to analyze levels of amino acids (glycine and taurine) as described previously (Ulenius et al., 2020). At the end of the experiment, rats were euthanized, brains gently removed from the skull and fixed in Accustain (Sigma diagnostics, United States) for 5–7 days. Brains were sectioned using a vibroslicer (Campden Instruments Ltd., Lafayette, IN, United States) to verify correct probe placement by gross examination (Figure 1A). Rats with incorrect probe placement or visual defects were excluded from the statistical analysis.
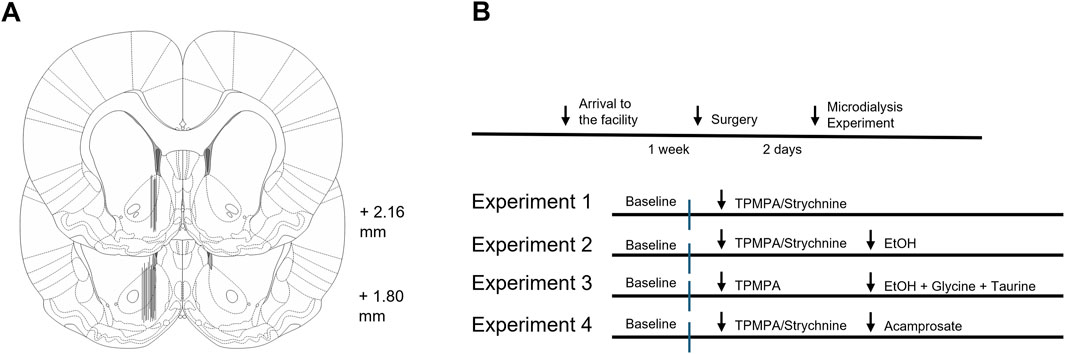
Figure 1. Histology and experimental design. (A) Schematic drawing of rat brain slices representing the nucleus accumbens. Black lines represent dialysis probe placements of approximately every fourth animal. Numbers indicate distance from bregma. (B) Schematic overview of the experimental design.
2.5 Experimental design
In a first experiment, TPMPA (10 μM, 100 μM, and 500 μM) or strychnine (20 μM and 200 μM) was administered in the nAc via the microdialysis probe to assess whether GABA-rho and GlyRs both are involved in maintaining basal dopamine levels within the nAc. In a second experiment, TPMPA (10 μM or 100 μM) or strychnine (20 μM) was administered in the nAc via the microdialysis probe prior to ethanol (300 mM, corresponding to approximately 67 mM in the tissue (Loftén et al., 2025)) locally in the nAc to assess if GABA-rho receptors and GlyRs both are involved in the ethanol-induced dopamine elevation. In the second experiment, we also analyzed the GlyR agonists glycine and taurine. In a third experiment we wanted to investigate if the addition of 50 μM glycine and 50 μM taurine to ethanol (300 mM) following TPMPA (100 μM) pretreatment would influence extracellular nAc dopamine. In the last study, we wanted to explore the influence of GABA-rho receptors on acamprosate-induced dopamine elevation. Here we perfused TPMPA (100 μM) or strychnine (20 μM) before the addition of acamprosate (1 mM). All drugs were applied via the dialysis probe. Experimental design is visualized in Figure 1B.
2.6 Statistical analysis
GraphPad Prism software version 10 (GraphPad Software, Inc., San Diego, CA, United States) was used for all statistical analyses. Two-way repeated-measures ANOVA followed by Tukey’s post hoc analysis was used for statistical evaluation of dopamine, glycine, and taurine levels for relevant timepoints. Only time points after the pharmacological intervention were included in the analysis. All data are presented as mean ± standard error of the mean (SEM), and p-value <0.05 was considered statistically significant.
3 Results
3.1 Glycine receptors, but not GABA-rho receptors, participate in maintaining the nucleus accumbens dopamine tone
We previously demonstrated that a high concentration of the GlyR antagonist strychnine (200 μM) decreases nAc dopamine levels in a manner that can be reversed by glycine, suggesting that GlyRs are involved in maintaining basal dopamine tone within the nAc. Since the GABA-rho receptor and GlyR share similar traits, we wanted to investigate whether the GABA-rho receptor also could be involved in regulating basal dopamine levels. As previously demonstrated, we found a dose-dependent decrease of nAc dopamine following strychnine administration (two-way ANOVA with repeated measures time points 20–180 min; F (2, 18) = 7.043; p = 0.0054, post hoc analysis, Ringer vs. strychnine 20, p = 0.883, Ringer vs. strychnine 200, p = 0.005, strychnine 20 vs. strychnine 200 p = 0.0189) (Figure 2A). Following perfusion of increasing concentrations of TPMPA, we did not find any influence on the extracellular levels of dopamine by the drug alone (two-way ANOVA with repeated measures time points 20–180 min; F (3, 28) = 2.0; p = 0.1263) (Figure 2B).
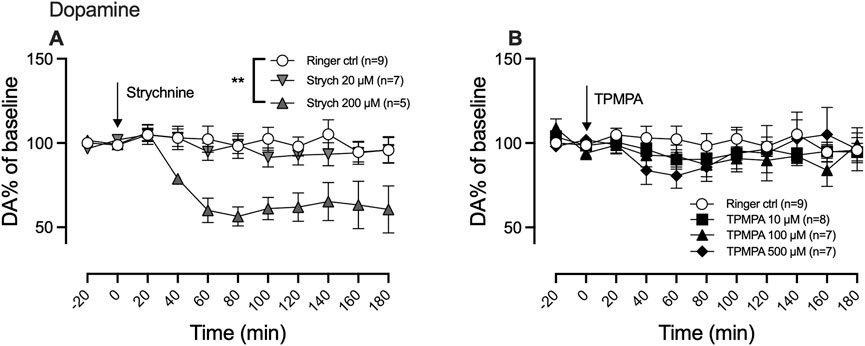
Figure 2. Glycine receptors, but not GABA-rho receptors, participate in maintaining the nucleus accumbens (nAc) dopamine tone. Strychnine (200 μM; Strych) significantly decreased nAc dopamine (DA) as compared to Ringer perfused rats (A), as measured by in vivo microdialysis in freely moving Wistar rats. Neither of the perfused doses of TPMPA produced a significant effect on nAc DA (B). Arrows indicate start of drug perfusion. Data are shown as means ± SEM. Statistical significance was determined by two-way repeated measures ANOVA followed by Tukey’s post hoc test, ** denotes p < 0.01.
3.2 Ethanol-induced dopamine elevation is blocked by the GABA-rho receptor antagonist TPMPA or the glycine receptor antagonist strychnine
To explore the involvement of GABA-rho receptors in ethanol-induced dopamine release, ethanol and the GABA-rho receptor antagonist TPMPA were administered locally in the nAc via reversed dialysis. Local administration of ethanol (300 mM) induced a significant elevation of extracellular dopamine as compared to Ringer control (two-way ANOVA with repeated measures time points 60–180 min; F (5, 44) = 7.8; p < 0.0001, Tukey’s post hoc analysis, Ringer vs. ethanol, p = 0.0005; Figure 3A). TPMPA did not influence dopamine on its own (Ringer vs. TPMPA 10 μM, p = 0.978; Ringer vs. TPMPA 100 μM, p = 0.941), but pre-treatment with either 10 μM or 100 μM TPMPA attenuated the ethanol-induced dopamine elevation (ethanol vs. TPMPA 10 μM + ethanol, p = 0.006 (Figure 3B); ethanol vs. TPMPA 100 μM + ethanol p = 0.004 (Figure 3C)), suggesting GABA-rho receptors to be involved in the ethanol-induced increase of dopamine. To verify the role of GlyRs in ethanol’s enhancement of the dopamine system, we locally administered the selective and competitive GlyR antagonist strychnine (20 μM) prior to local ethanol application. In line with our previous result (Molander and Söderpalm, 2005a), we were able to repeat that strychnine administration prevents the ethanol induced elevation of dopamine (two-way ANOVA with repeated measures time points 60–180 min; F (3, 30) = 7.8; p = 0.0005, post hoc analysis, Ringer vs. ethanol p = 0.0024, Ringer vs. strychnine p = 0.963, Ringer vs. strychnine + ethanol p = 0.976, strychnine + ethanol vs. ethanol p = 0.0072) (Figure 3D).
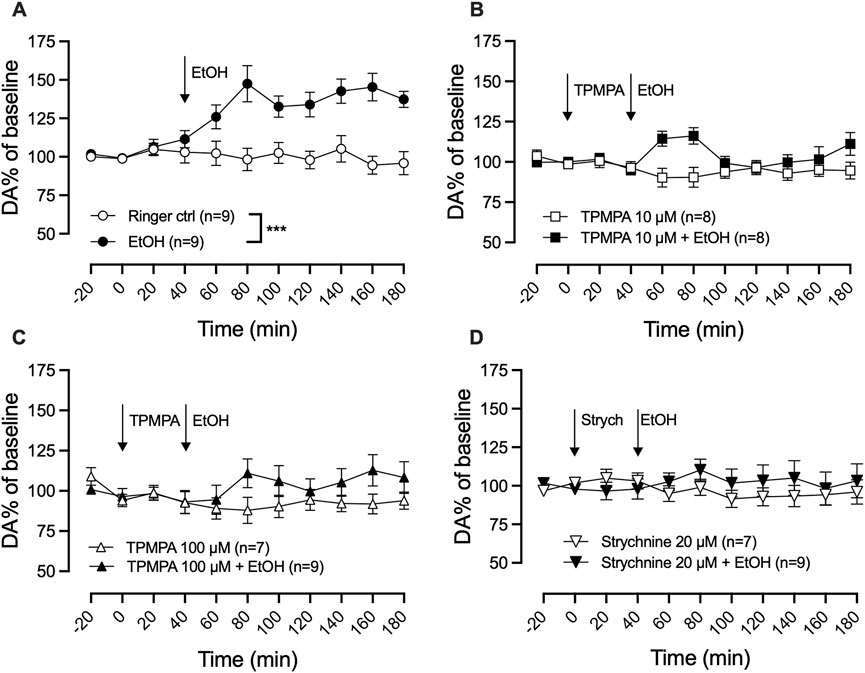
Figure 3. GABA-rho receptor or glycine receptor blockade attenuate ethanol-induced dopamine elevation in the nucleus accumbens. Time-course graph of dopamine (DA) levels as measured by in vivo microdialysis in freely moving Wistar rats, presented as DA percent of baseline following local perfusion of Ringer’s solution and ethanol (EtOH) 300 mM (A), TPMPA 10 μM alone and in combination with EtOH 300 mM (B), or TPMPA 100 μM alone and in combination with EtOH 300 mM (C), strychnine (Strych) 20 μM alone or in combination with EtOH 300 mM (D). Arrows indicate start of local drug administration. Data are shown as means ± SEM. Statistical significance was determined by two-way repeated measures ANOVA followed by Tukey’s post hoc test, *** denotes p < 0.001.
3.3 Both glycine receptor and GABA-rho receptor blockade modulate the ethanol-induced increase of glycine and taurine
Activation of GlyRs is a prerequisite for ethanol-induced dopamine release and local administration of ethanol (300 mM) significantly increased extracellular levels of both glycine (two-way ANOVA with repeated measures time points 60–180 min; F (5, 44) = 3.31; p = 0.013, Tukey’s post hoc analysis, Ringer vs. ethanol, p = 0.040) (Figure 4A) and taurine (two-way ANOVA with repeated measures time points 60–180 min; F (5, 44) = 17.62; p < 0.0001, post hoc analysis, Ringer vs. ethanol, p < 0.0001) (Figure 4) as compared to control.
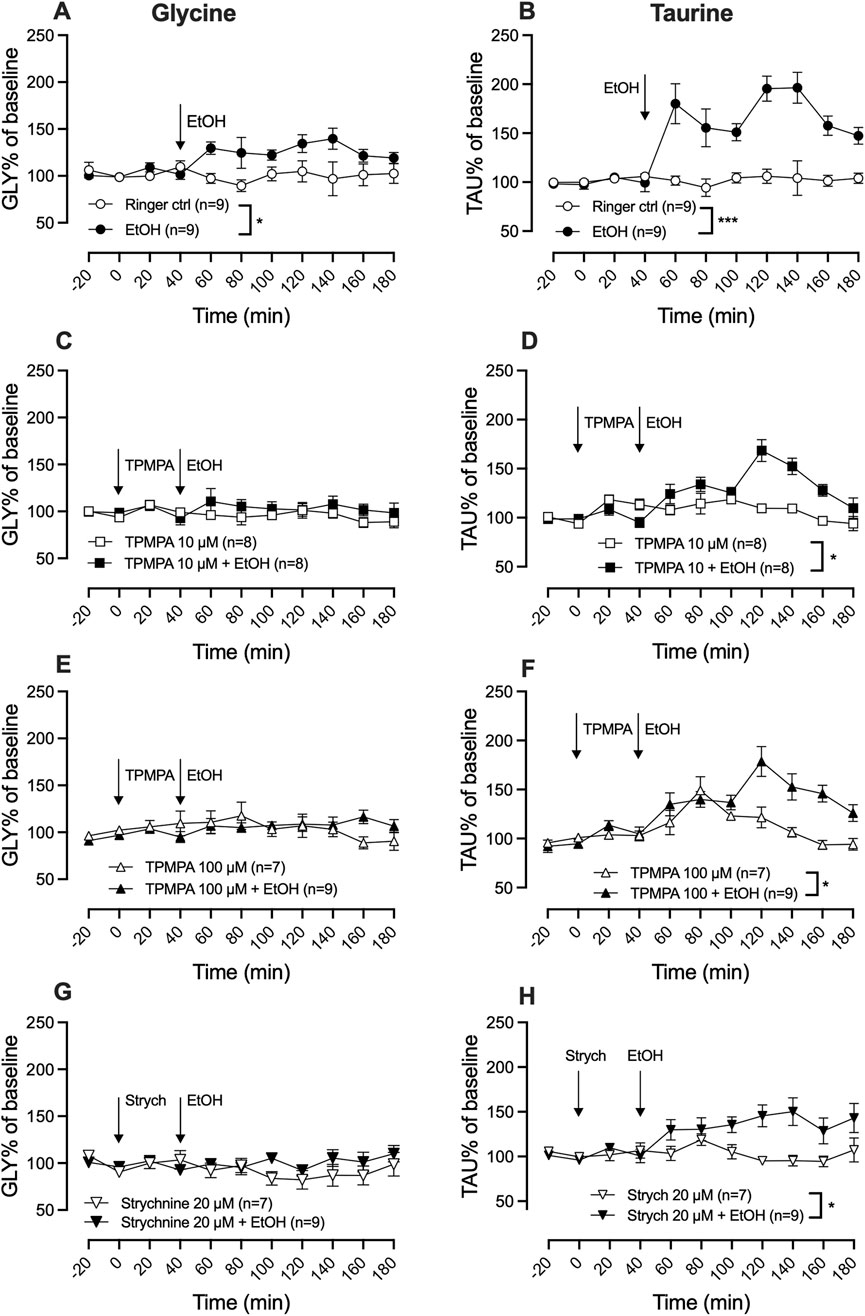
Figure 4. Blocking GABA-rho or glycine receptors attenuates ethanol-induced glycine and taurine elevation in the nucleus accumbens. Time-course graph of glycine (A,C,E,G) and taurine (B,D,F,H) levels as measured by in vivo microdialysis in freely moving Wistar rats, presented as percent of baseline following the local perfusion of Ringer’s solution and ethanol (EtOH) 300 mM (A,B), TPMPA 10 μM alone and in combination with EtOH 300 mM (C,D), TPMPA 100 μM alone and in combination with EtOH 300 mM (E,F), strychnine (strych) 20 μM alone or in combination with EtOH (G,H). Arrows indicate start of drug perfusion. Data are shown as means ± SEM. Statistical significance was determined by two-way repeated measures ANOVA followed by Tukey’s post hoc test, * denotes p < 0.05 and ***p < 0.001.
To assess whether ethanol-induced elevation of glycine and/or taurine could be modulated by GABA-rho receptors, TPMPA (10 μM or 100 μM) was administered prior to the start of local ethanol-perfusion (300 mM). Pre-treatment with TPMPA did not significantly alter extracellular glycine levels as compared to control (Tukey’s post hoc analysis; Ringer vs. TPMPA 10 μM, p = 0.995; Ringer vs. TPMPA 100 μM, p = 0.998) but inhibited ethanol’s ability to increase glycine (TPMPA 10 µM vs. TPMPA 10 µM + ethanol, p = 0.902; TPMPA 100 µM vs. TPMPA 100 µM + ethanol, p = 0.990; Figures 4C,E). Perfusion of the GABA-rho antagonist did not influence extracellular levels of taurine alone (Tukey’s post hoc analysis; Ringer vs. TPMPA 10 μM, p = 0.993; Ringer vs. TPMPA 100 μM, p = 0.732). However, post hoc analysis found TPMPA to partially inhibit the ethanol-induced elevation of taurine (TPMPA 10 µM vs. TPMPA 10 µM + ethanol, p = 0.043; TPMPA 100 µM vs. TPMPA 100 µM + ethanol, p = 0.021; ethanol vs. TPMPA 10 µM + ethanol, p = 0.004 and ethanol vs. TPMPA 100 µM + ethanol, p = 0.068; Figures 4D,F). Similar to TPMPA, strychnine administration also inhibited the ethanol induced elevation of glycine (two-way ANOVA with repeated measures time points 60–180 min; F (3, 30) = 5.2; p = 0.0050, post hoc analysis, Ringer vs. ethanol p = 0.031, Ringer vs. strychnine p = 0.797, Ringer vs. strychnine + ethanol p = 0.997, strychnine + ethanol vs. ethanol p = 0.049; Figure 4G) and partially the ethanol induced elevation of taurine (two-way ANOVA with repeated measures time points 60–180 min; F (3, 30) = 16.2; p < 0.0001, post hoc analysis, Ringer vs. ethanol p < 0.0001, Ringer vs. strychnine p = 0.999, Ringer vs. strychnine + ethanol p = 0.0152, strychnine vs. strychnine + ethanol p = 0.028; Figure 4H).
3.4 An artificial increase in glycine and taurine does not rescue ethanol-induced dopamine release
Considering the involvement of GlyRs in ethanol-induced dopamine release and the significant reduction of GlyR amino acid agonists following pretreatment with GABA-rho antagonist, we speculated that reduced GlyR signaling could underlie the effect of TPMPA. To test this hypothesis, we co-perfused a low level of glycine and taurine (50 µM of each amino acid via reversed microdialysis), which by themselves had no effect on basal dopamine. Co-perfusion of taurine has previously been demonstrated to restore ethanol-induced dopamine release during hypotonic conditions (Ericson et al., 2011). We administered TPMPA (100 µM) locally in the nAc 40 min before the addition of glycine (50 µM), taurine (50 µM), and ethanol (300 mM) in the perfusate. The addition of glycine and taurine did not restore the ethanol-induced dopamine elevation following GABA-rho blockade (two-way ANOVA with repeated measures time points 60–180 min; F (4, 37) = 5.9; p = 0.0009, post hoc analysis, Ringer vs. ethanol p = 0.0058, Ringer vs. TPMPA (100 µM)+ethanol p = 0.999, TPMPA (100 µM)+ethanol vs. TPMPA (100 µM)+ethanol + glycine + taurine) p = 0.999; Figure 5).
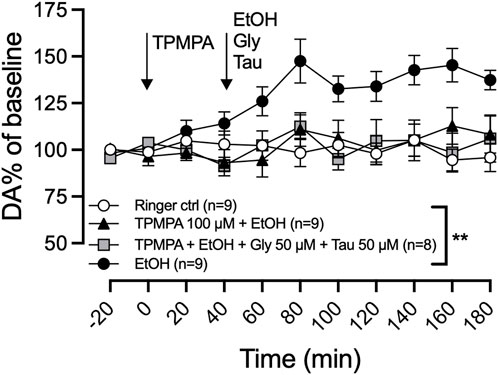
Figure 5. An artificial increase in glycine and taurine does not rescue ethanol-induced dopamine release. Time-course graph of extracellular levels of dopamine (DA) as measured by in vivo microdialysis in freely moving rats. Local administration, by means of reversed microdialysis in the nucleus accumbens, of Ringer’s solution, TPMPA 100 μM ethanol (EtOH) 300 mM, and in one group of rats, the addition of EtOH, glycine (Gly; 50 μM) and taurine (Tau; 50 μM) as indicated by the arrows. Data are shown as means ± SEM. Statistical significance was determined by two-way repeated measures ANOVA followed by Tukey’s post hoc test, ** denotes p < 0.01.
3.5 GABA-rho receptors are not involved in acamprosate-induced dopamine elevation
In a final set of experiments, we aimed to investigate the potential involvement of GABA-rho receptors in the acamprosate-induced increase in dopamine levels within the nAc. Local administration of acamprosate (1 mM) significantly increased dopamine levels in the nAc (two-way ANOVA with repeated measures time points 40–180 min; F (3, 30) = 10.15; p < 0.0001, post hoc analysis, Ringer vs. acamprosate p = 0.0004), an effect that was not affected by TPMPA 100 μM pre-treatment (Ringer vs. TPMPA p = 0.758, acamprosate vs. acamprosate + TPMPA p = 0.856, Ringer vs. acamprosate + TPMPA p = 0.023, Figure 6A), but fully blocked by strychnine (two-way ANOVA with repeated measures time points 40–180 min; F (3, 28) = 11.62; p < 0.0001, post hoc analysis, Ringer vs. acamprosate p = 0.0031, Ringer vs. strychnine p = 0.942, acamprosate vs. acamprosate + strychnine p < 0.001, Ringer vs. acamprosate + strychnine p = 0.999; Figure 6B).
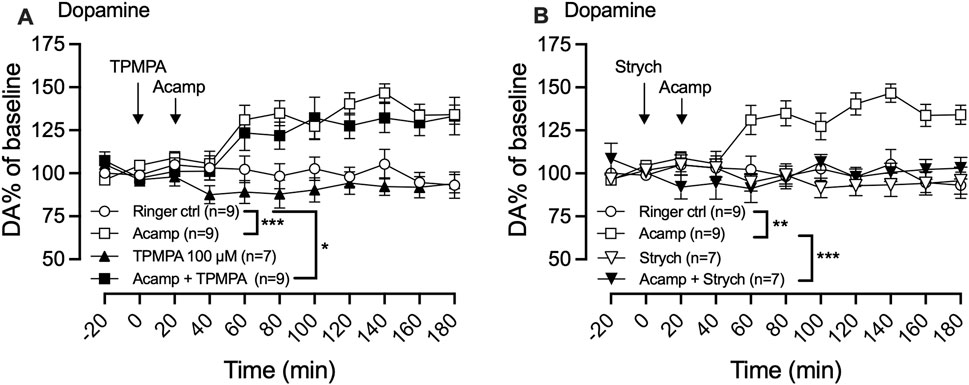
Figure 6. Acamprosate-induced dopamine elevation does not involve GABA-rho receptors. Time-course graph of extracellular levels of dopamine (DA) as measured by in vivo microdialysis in freely moving rats. Local administration, by means of reversed microdialysis in the nucleus accumbens, of Ringer’s solution, acamprosate (Acamp) 1 mM, TPMPA (100 μM), and the combination of acamprosate and TPMPA (A) and Ringer’s solution, acamprosate (Acamp) 1 mM, Strychnine (Strych; 20 μM), and the combination of acamprosate and strychnine (B). Arrows indicate start of local drug perfusion. Data are shown as mean ± SEM. Statistical significance was determined by two-way repeated measures ANOVA followed by Tukey’s post hoc test, * denotes p < 0.05, **p < 0.01 and ***p < 0.001.
4 Discussion
Recent studies have highlighted a role for GABA-rho receptors in ethanol-induced responses (Blednov et al., 2014; Yorgason et al., 2022). Considering the structural and functional similarity with GlyRs we hypothesized that the mechanisms associated with inhibition of GABA-rho receptors would be mimicked by GlyR antagonism. The data presented here demonstrate that GlyRs display a stronger influence over baseline dopamine levels and that a GlyR antagonist more robustly inhibits ethanol-induced dopamine release. In addition, while both TPMPA and strychnine reduce the ethanol-mediated release of GlyR agonists, only strychnine blocks dopamine release mediated by acamprosate. Thus, while GABA-rho receptors and GlyRs share some features, inhibition of GlyRs displays a more prominent effect on dopamine signaling and neurochemical responses to ethanol. Still, strychnine presents properties, e.g., deadly blockade of spinal GlyRs, that hinder its implementation as a possible intervention for AUD, while TPMPA appears to exert more ethanol-specific modulatory effects. Thus, GABA-rho receptors could be a promising target for future intervention strategies.
Local administration of the GlyR antagonist strychnine significantly reduced the microdialysate concentration of dopamine, indicating that nAc GlyRs are tonically active and play a key role in maintaining basal dopamine levels. This is in agreement with previous studies, demonstrating that strychnine produces a concentration-dependent decrease in accumbal dopamine concentrations which can be reversed by co-perfusion with glycine (Molander and Söderpalm, 2005a). TPMPA did not modulate extracellular dopamine levels, suggesting that GABA-rho receptors are not involved in maintaining basal dopamine levels in the nAc. This is further in line with an in vitro study, showing that TPMPA did not alter evoked dopamine release (Yorgason et al., 2022). The mechanism underlying GlyR-mediated regulation of dopamine levels has not been fully established, but it has been suggested that the GlyRs that are responsible for maintaining basal dopamine levels are not located on cholinergic interneurons (CINs), but rather involve other neurotransmitter systems (Loftén et al., 2023). Even though GABA-rho receptors may be expressed on a variety of neuronal cell types within the nAc, they appear to especially modulate CIN signaling (Yorgason et al., 2022). Thus, there appears to be a functional difference between glycine and GABA-rho receptors in the regulation of accumbal dopamine levels, which may be attributed to the distribution of expression. While GlyRs, putatively located on medium spiny neurons, appear to be critical for maintaining basal dopaminergic tone, GABA-rho receptors may play a more nuanced role, contributing to the dopaminergic response under conditions of ethanol exposure (Figure 7).
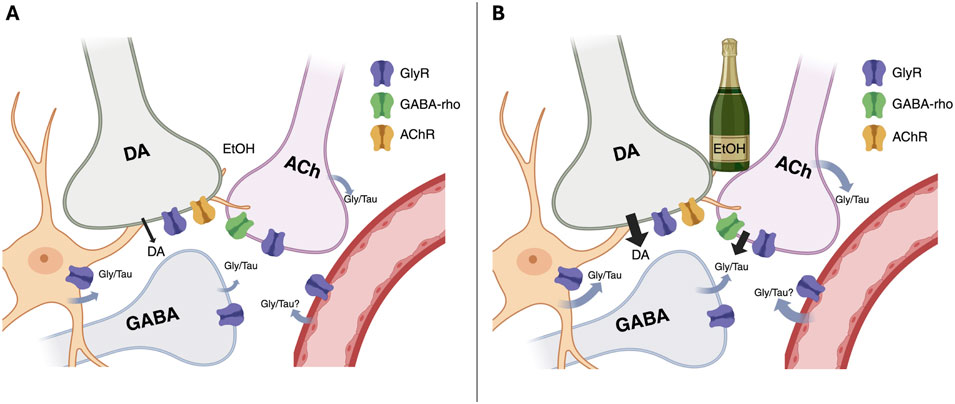
Figure 7. Simplified schematic illustrating the proposed roles of glycine receptors (GlyR) and GABA-rho receptors in regulating dopamine (DA) signaling under baseline conditions (A) and during ethanol exposure (B). At baseline, DA tone in the nucleus accumbens is sustained by tonic GlyR activity on non-cholinergic interneurons, with little contribution from GABA-rho receptors. Ethanol elevates extracellular glycine and taurine, which activate GlyRs on cholinergic interneurons (CINs), enhancing acetylcholine (ACh) release and subsequent acetylcholine receptor (AChR)-mediated DA output. Full expression of this ethanol-induced DA elevation additionally requires GABA-rho receptors, as blockade of either receptor type reduces glycine/taurine signaling and prevents DA enhancement. Together, these findings suggest that ethanol recruits a “dual-key” mechanism involving both GlyR and GABA-rho signaling. The image was created using BioRender.
Ethanol increased accumbal dopamine levels, which is in agreement with previous studies (Ericson et al., 1998; Ericson et al., 2020; Imperato and Di Chiara, 1986). The increase in dopamine was fully blocked by the GlyR antagonist strychnine and significantly reduced by the GABA-rho receptor antagonist TPMPA. Accumbal dopamine signaling is tightly regulated by various neurotransmitter systems, with cholinergic transmission identified as a key regulator of dopamine release (Abrahao et al., 2017; Söderpalm and Ericson, 2013; Zhou et al., 2001). Endogenous acetylcholine derives mainly from local CIN and selective activation of these cells is sufficient to induce a dopamine increase, an effect mediated via nicotinic acetylcholine receptors located on dopamine terminals (Brimblecombe et al., 2018; Cachope et al., 2012; Threlfell et al., 2012). Accumbal CINs have also been shown to be involved in ethanol-induced dopamine release (Loftén et al., 2021; Loftén et al., 2023; Wadsworth et al., 2023). CINs are tonically active (Aosaki et al., 1995), and are regulated by glutamatergic inputs from the cortex and GABAergic projection neurons originating in the VTA (Al-Hasani et al., 2021; Brown et al., 2012). Since CIN may express both GlyRs and GABA-rho receptors (Sergeeva and Haas, 2001; Yorgason et al., 2022), it is possible that the mechanisms underlying the inhibitory effect of strychnine and TPMPA on ethanol-induced dopamine elevation involve modulation of CIN signaling. This is further supported by previous findings, demonstrating that cholinergic antagonists block the ethanol-induced dopamine increase and that selective depletion of CIN attenuates ethanol-induced dopamine elevation (Loftén et al., 2021).
Glycine receptors are important regulators of ethanol-induced dopamine elevation in rats (Jonsson et al., 2014; Molander and Söderpalm, 2005b). Glycine and taurine are endogenous agonists for the GlyR (Pan and Slaughter, 1995), and the data presented here demonstrate a robust increase in both of these amino acids in response to ethanol challenge. Changes in endogenous GlyR agonists could thus play a key role in regulating the acute rewarding properties of alcohol. In fact, the ethanol-induced rise in taurine levels has previously been shown to be a prerequisite for ethanol-induced dopamine release (Ericson et al., 2011). Importantly, both strychnine and TPMPA blunted the ethanol-induced release of glycine and taurine. As both receptors may be expressed by CIN (Sergeeva and Haas, 2001; Yorgason et al., 2022), and as depletion of accumbal CINs has been shown to reduce ethanol-induced levels of glycine and taurine (Loftén et al., 2023), these findings could be mediated through changes in cholinergic signaling. The role of CINs in regulating amino acid release needs however to be further evaluated. Postulating that the release of glycine and taurine is required for ethanol-induced dopamine release, we speculated that co-perfusion of these agonists could restore dopamine signaling during conditions where the GABA-rho receptor was blocked. However, TPMPA blocked ethanol-induced dopamine release also during these circumstances. Since glycine and taurine are also endogenous agonists for the GABA-rho receptor (Calvo and Miledi, 1995; Ochoa-de la Paz et al., 2008), it is possible that the inhibition of the receptor prevents downstream signaling. Thus, GABA-rho receptors may be important not only for inducing the release of glycine and taurine, but also for conveying downstream signaling mediated by these amino acids.
Acamprosate is a pharmacological treatment for AUD approved by the United States Food and Drug Administration (FDA) and the European Medicines Agency (EMA). The mechanism of action is debated where some suggest modulation of N-Metyl-D-aspartate (NMDA) receptors, stabilizing excitatory and inhibitory neurotransmission, whilst others suggest interaction with accumbal GlyRs or that calcium and not homotaurine may underlie the effects of acamprosate (Ademar et al., 2022; Ademar et al., 2023; Al Qatari et al., 1998; Chau et al., 2010a; Chau et al., 2018; Spanagel et al., 2014). The interaction with GlyRs is suggested to mediate the acamprosate-induced dopamine increase, a pharmacological effect proposed to be important in the decreased ethanol consumption seen following treatment. Therefore, we wanted to investigate if part of acamprosate’s dopamine-releasing effect is mediated via GABA-rho receptors as this information could have important treatment implications. Supporting previous studies, we found that strychnine blocked dopamine release elicited by acamprosate. However, TPMPA did not attenuate acamprosate-induced dopamine increase, suggesting that GABA-rho receptors are not involved in acamprosate’s effect on dopamine. However, it should be noted that the present study was performed exclusively in male rats. Future studies should include both sexes to ensure broader applicability of the findings and to investigate potential sex differences.
In summary, our in vivo findings demonstrate that GABA-rho receptors in the nAc play an important role in mediating ethanol-induced elevation of dopamine levels and can influence ethanol-induced release of the GlyR agonists glycine and taurine. However, the GABA-rho receptor does not appear to contribute to the maintenance of basal dopamine levels in the nAc or to the dopamine-elevating effects of acamprosate. Although GlyR antagonism had a more prominent effect on dopamine signaling, the observation that GABA-rho receptors neither affect basal dopamine neurotransmission nor non-ethanol induced stimulation but appear to exert an ethanol-specific modulatory effect could make it a promising target for pharmacological interventions. Overall, these results highlight a selective involvement of GABA-rho receptors in ethanol-related neurochemical responses, providing new insights into the neurobiological mechanisms underlying ethanol’s effects on the reward system.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by Ethics Committee for Animal Experiments, Gothenburg, Sweden. The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
DC: Investigation, Formal Analysis, Writing – original draft. AL: Formal Analysis, Writing – review and editing, Investigation, Writing – original draft. KA: Formal Analysis, Writing – original draft, Investigation. BS: Conceptualization, Funding acquisition, Supervision, Writing – review and editing. LA: Validation, Funding acquisition, Supervision, Writing – review and editing. ME: Funding acquisition, Visualization, Validation, Project administration, Writing – original draft, Supervision, Writing – review and editing, Conceptualization.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This investigation was partially supported by grants from the Swedish Research Council, Grant/Award Numbers: 2020-02105, 2020-01346, 2020-00559, 2018-02814; LUA/ALF.
Acknowledgments
The authors would like to acknowledge Rosita Stomberg for expert technical and administrative support.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
The author(s) declared that they were an editorial board member of Frontiers, at the time of submission. This had no impact on the peer review process and the final decision.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Abrahao, K. P., Salinas, A. G., and Lovinger, D. M. (2017). Alcohol and the brain: neuronal molecular targets, synapses, and circuits. Neuron 96 (6), 1223–1238. doi:10.1016/j.neuron.2017.10.032
Ademar, K., Adermark, L., Söderpalm, B., and Ericson, M. (2022). Sodium acamprosate and calcium exert additive effects on nucleus accumbens dopamine in the rat. Addict. Biol. 27 (5), e13224. doi:10.1111/adb.13224
Ademar, K., Loftén, A., Nilsson, M., Domi, A., Adermark, L., Söderpalm, B., et al. (2023). Acamprosate reduces ethanol intake in the rat by a combined action of different drug components. Sci. Rep. 13 (1), 17863. doi:10.1038/s41598-023-45167-3
Al Qatari, M., Bouchenafa, O., and Littleton, J. (1998). Mechanism of action of acamprosate. Part II. Ethanol dependence modifies effects of acamprosate on NMDA receptor binding in membranes from rat cerebral cortex. Alcohol Clin. Exp. Res. 22 (4), 810–814. doi:10.1097/00000374-199806000-00006
Al-Hasani, R., Gowrishankar, R., Schmitz, G. P., Pedersen, C. E., Marcus, D. J., Shirley, S. E., et al. (2021). Ventral tegmental area GABAergic inhibition of cholinergic interneurons in the ventral nucleus accumbens shell promotes reward reinforcement. Nat. Neurosci. 24 (10), 1414–1428. doi:10.1038/s41593-021-00898-2
Aosaki, T., Kimura, M., and Graybiel, A. M. (1995). Temporal and spatial characteristics of tonically active neurons of the primate's striatum. J. Neurophysiol. 73 (3), 1234–1252. doi:10.1152/jn.1995.73.3.1234
Blednov, Y. A., Benavidez, J. M., Black, M., Leiter, C. R., Osterndorff-Kahanek, E., Johnson, D., et al. (2014). GABAA receptors containing ρ1 subunits contribute to in vivo effects of ethanol in mice. PLoS One 9 (1), e85525. doi:10.1371/journal.pone.0085525
Bormann, J. (2000). The 'ABC' of GABA receptors. Trends Pharmacol. Sci. 21 (1), 16–19. doi:10.1016/s0165-6147(99)01413-3
Brimblecombe, K. R., Threlfell, S., Dautan, D., Kosillo, P., Mena-Segovia, J., and Cragg, S. J. (2018). Targeted activation of cholinergic interneurons accounts for the modulation of dopamine by striatal nicotinic receptors. eNeuro 5 (5), ENEURO.0397–17.2018. doi:10.1523/ENEURO.0397-17.2018
Brown, M. T., Tan, K. R., O'Connor, E. C., Nikonenko, I., Muller, D., and Lüscher, C. (2012). Ventral tegmental area GABA projections pause accumbal cholinergic interneurons to enhance associative learning. Nature 492 (7429), 452–456. doi:10.1038/nature11657
Cachope, R., Mateo, Y., Mathur, B. N., Irving, J., Wang, H. L., Morales, M., et al. (2012). Selective activation of cholinergic interneurons enhances accumbal phasic dopamine release: setting the tone for reward processing. Cell Rep. 2 (1), 33–41. doi:10.1016/j.celrep.2012.05.011
Calvo, D. J., and Miledi, R. (1995). Activation of GABA rho 1 receptors by glycine and beta-alanine. Neuroreport 6 (8), 1118–1120. doi:10.1097/00001756-199505300-00011
Cano-Cebrian, M. J., Zornoza-Sabina, T., Guerri, C., Polache, A., and Granero, L. (2003). Local acamprosate modulates dopamine release in the rat nucleus accumbens through NMDA receptors: an in vivo microdialysis study. Naunyn Schmiedeb. Arch. Pharmacol. 367, 119–125. doi:10.1007/s00210-002-0674-3
Chau, P., Stomberg, R., Fagerberg, A., Söderpalm, B., and Ericson, M. (2010a). Glycine receptors involved in acamprosate's modulation of accumbal dopamine levels: an in vivo microdialysis study. Alcohol Clin. Exp. Res. 34 (1), 32–38. doi:10.1111/j.1530-0277.2009.01062.x
Chau, P., Höifödt-Lidö, H., Löf, E., Söderpalm, B., and Ericson, M. (2010b). Glycine receptors in the nucleus accumbens involved in the ethanol intake-reducing effect of acamprosate. Alcohol Clin. Exp. Res. 34 (1), 39–45. doi:10.1111/j.1530-0277.2009.01063.x
Chau, P., Lido, H. H., Soderpalm, B., and Ericson, M. (2018). Acamprosate's ethanol intake-reducing effect is associated with its ability to increase dopamine. Pharmacol. Biochem. Behav. 175, 101–107. doi:10.1016/j.pbb.2018.09.009
Clarke, R. B., Adermark, L., Chau, P., Soderpalm, B., and Ericson, M. (2014). Increase in nucleus accumbens dopamine levels following local ethanol administration is not mediated by acetaldehyde. Alcohol Alcohol 49 (5), 498–504. doi:10.1093/alcalc/agu047
Ericson, M., Blomqvist, O., Engel, J. A., and Söderpalm, B. (1998). Voluntary ethanol intake in the rat and the associated accumbal dopamine overflow are blocked by ventral tegmental mecamylamine. Eur. J. Pharmacol. 358 (3), 189–196. doi:10.1016/s0014-2999(98)00602-5
Ericson, M., Chau, P., Clarke, R. B., Adermark, L., and Soderpalm, B. (2011). Rising taurine and ethanol concentrations in nucleus accumbens interact to produce dopamine release after ethanol administration. Addict. Biol. 16 (3), 377–385. doi:10.1111/j.1369-1600.2010.00245.x
Ericson, M., Ulenius, L., Andrén, A., Jonsson, S., Adermark, L., and Söderpalm, B. (2020). Different dopamine tone in ethanol high- and low-consuming wistar rats. Addict. Biol. 25 (3), e12761. doi:10.1111/adb.12761
Imperato, A., and Di Chiara, G. (1986). Preferential stimulation of dopamine release in the nucleus accumbens of freely moving rats by ethanol. J. Pharmacol. Exp. Ther. 239 (1), 219–228. doi:10.1016/s0022-3565(25)23929-5
Jonsson, S., Adermark, L., Ericson, M., and Söderpalm, B. (2014). The involvement of accumbal glycine receptors in the dopamine-elevating effects of addictive drugs. Neuropharmacology 82, 69–75. doi:10.1016/j.neuropharm.2014.03.010
Loftén, A., Adermark, L., Ericson, M., and Söderpalm, B. (2021). An acetylcholine-dopamine interaction in the nucleus accumbens and its involvement in ethanol's dopamine-releasing effect. Addict. Biol. 26 (3), e12959. doi:10.1111/adb.12959
Loftén, A., Adermark, L., Ericson, M., and Söderpalm, B. (2023). Regulation of ethanol-mediated dopamine elevation by glycine receptors located on cholinergic interneurons in the nucleus accumbens. Addict. Biol. 28 (12), e13349. doi:10.1111/adb.13349
Loftén, A., Ademar, K., Danielsson, K., Söderpalm, B., Adermark, L., and Ericson, M. (2025). Alcohol-induced accumbal dopamine- and taurine release in female and male Wistar rats, an in vivo microdialysis study. J. Neural Transm. (Vienna) 132 (7), 1051–1062. doi:10.1007/s00702-025-02928-w
Mohammadi, E., Shamsizadeh, A., Salari, E., Fatemi, I., Allahtavakoli, M., and Roohbakhsh, A. (2017). Effect of TPMPA (GABA(C) receptor antagonist) on neuronal response properties in rat barrel cortex. Somatosens. Mot. Res. 34 (2), 108–115. doi:10.1080/08990220.2017.1317240
Molander, A., and Söderpalm, B. (2005a). Glycine receptors regulate dopamine release in the rat nucleus accumbens. Alcohol Clin. Exp. Res. 29 (1), 17–26. doi:10.1097/01.alc.0000150006.17168.f7
Molander, A., and Söderpalm, B. (2005b). Accumbal strychnine-sensitive glycine receptors: an access point for ethanol to the brain reward system. Alcohol Clin. Exp. Res. 29 (1), 27–37. doi:10.1097/01.alc.0000150012.09608.81
Naffaa, M. M., Hung, S., Chebib, M., Johnston, G. A. R., and Hanrahan, J. R. (2017). GABA-ρ receptors: distinctive functions and molecular pharmacology. Br. J. Pharmacol. 174 (13), 1881–1894. doi:10.1111/bph.13768
Ochoa-de la Paz, L. D., Martínez-Dávila, I. A., Miledi, R., and Martínez-Torres, A. (2008). Modulation of human GABArho1 receptors by taurine. Neurosci. Res. 61 (3), 302–308. doi:10.1016/j.neures.2008.03.009
Olive, M. F., Nannini, M. A., Ou, C. J., Koenig, H. N., and Hodge, C. W. (2002). Effects of acute acamprosate and homotaurine on ethanol intake and ethanolstimulated mesolimbic dopamine release. Eur. J. Pharmacol. 437, 55–61. doi:10.1016/s0014-2999(02)01272-4
Pan, Z. H., and Slaughter, M. M. (1995). Comparison of the actions of glycine and related amino acids on isolated third order neurons from the tiger salamander retina. Neuroscience 64 (1), 153–164. doi:10.1016/0306-4522(94)00399-p
Sergeeva, O. A., and Haas, H. L. (2001). Expression and function of glycine receptors in striatal cholinergic interneurons from rat and mouse. Neuroscience 104 (4), 1043–1055. doi:10.1016/s0306-4522(01)00130-0
Söderpalm, B., and Ericson, M. (2013). Neurocircuitry involved in the development of alcohol addiction: the dopamine system and its access points. Curr. Top. Behav. Neurosci. 13, 127–161. doi:10.1007/7854_2011_170
Spanagel, R., Vengeliene, V., Jandeleit, B., Fischer, W. N., Grindstaff, K., Zhang, X., et al. (2014). Acamprosate produces its anti-relapse effects via calcium. Neuropsychopharmacology 39 (4), 783–791. doi:10.1038/npp.2013.264
Threlfell, S., Lalic, T., Platt, N. J., Jennings, K. A., Deisseroth, K., and Cragg, S. J. (2012). Striatal dopamine release is triggered by synchronized activity in cholinergic interneurons. Neuron 75 (1), 58–64. doi:10.1016/j.neuron.2012.04.038
Ulenius, L., Andrén, A., Adermark, L., Söderpalm, B., and Ericson, M. (2020). Sub-chronic taurine administration induces behavioral sensitization but does not influence ethanol-induced dopamine release in the nucleus accumbens. Pharmacol. Biochem. Behav. 188, 172831. doi:10.1016/j.pbb.2019.172831
Wadsworth, H. A., Anderson, E. Q., Williams, B. M., Ronström, J. W., Moen, J. K., Lee, A. M., et al. (2023). Role of α6-Nicotinic receptors in alcohol-induced GABAergic synaptic transmission and plasticity to cholinergic interneurons in the nucleus accumbens. Mol. Neurobiol. 60 (6), 3113–3129. doi:10.1007/s12035-023-03263-5
Woodward, R. M., Polenzani, L., and Miledi, R. (1993). Characterization of bicuculline/baclofen-insensitive (rho-like) gamma-aminobutyric acid receptors expressed in Xenopus oocytes. II. Pharmacology of gamma-aminobutyric acidA and gamma-aminobutyric acidB receptor agonists and antagonists. Mol. Pharmacol. 43 (4), 609–625. doi:10.1016/s0026-895x(25)13444-5
Yorgason, J. T., Wadsworth, H. A., Anderson, E. J., Williams, B. M., Brundage, J. N., Hedges, D. M., et al. (2022). Modulation of dopamine release by ethanol is mediated by atypical GABA(A) receptors on cholinergic interneurons in the nucleus accumbens. Addict. Biol. 27 (1), e13108. doi:10.1111/adb.13108
Zhang, D., Pan, Z. H., Awobuluyi, M., and Lipton, S. A. (2001). Structure and function of GABA(C) receptors: a comparison of native versus recombinant receptors. Trends Pharmacol. Sci. 22 (3), 121–132. doi:10.1016/s0165-6147(00)01625-4
Keywords: dopamine, ethanol, taurine, GABA-rho, rat, nucleus accumbens, glycine
Citation: Cadeddu D, Loftén A, Ademar K, Söderpalm B, Adermark L and Ericson M (2025) The involvement of GABA-rho receptors in regulating ethanol-induced elevation of dopamine, glycine and taurine within the nucleus accumbens of Wistar rats. Front. Pharmacol. 16:1668669. doi: 10.3389/fphar.2025.1668669
Received: 18 July 2025; Accepted: 16 October 2025;
Published: 28 October 2025.
Edited by:
Priscila Vázquez-León, Universidad Autónoma de la Ciudad de México, MexicoReviewed by:
John J. Woodward, Medical University of South Carolina, United StatesMario Rivera-Meza, University of Chile, Chile
Copyright © 2025 Cadeddu, Loftén, Ademar, Söderpalm, Adermark and Ericson. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Mia Ericson, bWlhLmVyaWNzb25AbmV1cm8uZ3Uuc2U=