 Marion Brollo1
Marion Brollo1 Stanislas Grassin-Delyle2,3,4
Stanislas Grassin-Delyle2,3,4 Camille Roquencourt1,3,4Elisabeth Longchampt5Isabelle Miguet-Besson5Matthieu Glorion1,2,6Hélène Salvator1,2,7
Camille Roquencourt1,3,4Elisabeth Longchampt5Isabelle Miguet-Besson5Matthieu Glorion1,2,6Hélène Salvator1,2,7 Philippe Devillier1,2,4*
Philippe Devillier1,2,4*- 1Laboratory of Research in Respiratory Pharmacology, VIM Suresnes, UMR-0892, Foch Hospital, Suresnes, France
- 2Faculty of Health Sciences Simone Veil, University of Versailles Saint Quentin en Yvelines, Paris Saclay University, Montigny-leBretonneux, France
- 3INSERM UMR1173 & Mass Spectrometry Platform, University of Versailles Saint Quentin en Yvelines, University Paris Saclay, Montigny-leBretonneux, France
- 4Exhalomics®, Exhaled Breath Analysis Platform, Department of Pulmonology, Foch Hospital, Suresnes, France
- 5Pathology Department, Foch Hospital, Suresnes, France
- 6Department of Thoracic Surgery, Foch Hospital, Suresnes, France
- 7Department of Pulmonology, Foch Hospital, Suresnes, France
Aim: The role of somatostatin (SST) in the modulation of cholinergic neurotransmission has not been explored previously in human bronchi. We investigated the effects of SST, selective agonists of the five SST receptors SSTR, and octreotide (a SSTR2,3,5 agonist) on the cholinergic contraction induced in vitro either by acetylcholine or by electrical field stimulation (EFS) in human bronchial rings.
Methods: Human bronchial rings (n = 326) were obtained from 32 patients undergoing surgery for lung carcinoma. 5 Hz EFS (biphasic pulse width: 1 ms; constant current: 320 mA for 10 s) induced contractions that reached about ∼30% of the maximum contraction caused by 40 Hz EFS. Bronchial rings were stimulated for 240 min in the presence or absence of various concentrations of SST, octreotide, and selective agonists of each of the five SSTR receptors. Furthermore, the tissue and cellular locations of each of the five types of SSTR was determined by immunohistochemistry.
Results: SST, octreotide, and the SSTR agonists did not change the resting tone or the contractions produced by the cumulative addition of acetylcholine (10−9 to 10−3 M). In contrast, octreotide and the SSTR3 and SSTR5 agonists significantly increased the EFS-induced contractions. Immunoreactivity for all SSTR subtypes was detected in the airway’s neural ganglia.
Conclusion: The present study provided new data on the location of SSTR in the human lung: notably, all types of receptor were found in the parasympathetic nerve ganglia of the bronchial wall. We suggest that the activation of prejunctional SSTR3 and SSTR5 receptors potentiates cholinergic-nerve-mediated contraction induced by EFS in human bronchi.
1 Background
The multiple physiological functions and actions of somatostatin (SST) are mediated through five receptors (SSTR1-SSTR5). These receptors belong to the G–protein–coupled receptor family and are involved in several distinct signal transduction pathways (Günther et al., 2018). The gene coding for SSTR2 (SSTR2) is subject to alternate mRNA splicing and thus produces two receptor isoforms (SSTR2A and SSTR2B), which differ only with regard to the number of amino acids and the amino acid composition of the cytoplasmic carboxy-terminal tail (Günther et al., 2018).
The SSTRs’ expression and (in particular) functions in the human lung have not been extensively characterized. With regard to protein expression, all the SSTRs other than SSTR2A have been found on cells in the bronchial glands (Taniyama et al., 2005). Furthermore, SSTR2A and SSTR4 have been found on bronchial, bronchiolar and alveolar epithelial cells (Gugger et al., 2004; Borie et al., 2008; Varecza et al., 2009), SSTR2B, SSTR4 and SSTR5 are present on alveolar macrophages, and SSTR4 has been identified on smooth muscle and vascular endothelial cells (Borie et al., 2008; Varecza et al., 2009; Günther et al., 2018). SST is known to inhibit lipopolysaccharide-induced cytokine production by human lung macrophages, although the receptors involved have not been identified (Balibrea et al., 1994). Furthermore, SSTR2A receptors might be involved in the inhibition of pulmonary fibrosis (Borie et al., 2008). All the other data on SST’s effects in the lung come from animal studies. In rodents, SST and its synthetic analogues inhibited endotoxin- and ovalbumin-induced airway inflammation and the resulting bronchial hyperreactivity (Helyes et al., 2006; Helyes et al., 2007; Helyes et al., 2009; Elekes et al., 2008). These effects involved (at least in part) the activation of SSTR4 on bronchial smooth muscles and the prejunctional SSTR4 that mediates the inhibition of neuropeptide release from afferent sensory nerves (Helyes et al., 2006; Elekes et al., 2008). Although the vast majority of SST’s effects are inhibitory (particularly for neurogenically mediated contractile responses (Pintér et al., 2006; Günther et al., 2018), direct contractile effects have been described with several isolated preparations: guinea pig isolated ileum and vas deferens (Feniuk et al., 1993), human saphenous veins (Dimech et al., 1995) and colonic smooth muscle cells (Corleto et al., 2006). Furthermore, SST has been shown to potentiate cholinergic neurotransmission in isolated ferret trachea (Sekizawa et al., 1989). On the other hand, adverse effects such as dyspnea or bronchospasm have been reported in clinical trials of somatostatin and octreotide, a stable SSTR2, SSTR3 and SSTR5 agonist (see the summary of product characteristics).
The objectives of the present study were to (i) describe the expression of the various SSTRs in the human lung more fully, and (ii) analyze the direct and indirect effects of SST, octreotide, and other SSTR-specific agonists on isolated human bronchial preparations.
2 Methods
Our hospital’s pathology department supplied lung tissue from macroscopically healthy parts of the lungs of 32 patients (26 males and 6 females; mean ± standard deviation (SD) age: 63.8 ± 8.7 years; 11 current smokers, 18 former smokers, and 3 never-smokers; mean ± SD pack-years: 37.5 ± 19.8; mean ± SD FEV1 = 78.8 ± 15.1%; mean ± SD FEV1/FVC ratio: 0.81 ± 0.15. Four of the patients had non-severe COPD. All the patients had undergone surgical resection for lung carcinoma and had not received chemotherapy or radiotherapy before surgery. The use of human lung tissue for in vitro experiments was approved by the local independent ethics committee (Comité de Protection des Personnes Ile de France VIII, Boulogne-Billancourt, France; reference: CPP8 - DC 11 10 05). Each patient gave their informed consent to the use of surgical samples for research purposes.
2.1 Immunohistochemical detection of the SST receptors
Lung tissue samples were obtained from five patients who had all stopped smoking for at least 3 years. The samples were fixed in 4% buffered formaldehyde within 1 hour after lobectomy, embedded in paraffin, and cut into 4 µm serial sections. The tissue sections were then dewaxed in xylene and rehydrated with ethanol. Before the immunohistochemical procedure, the antigens were retrieved by incubating the tissue sections in a boiling water canner for 20 min. Non-specific binding of the secondary antibody was prevented by incubation with normal goat serum for 1 h. The slides were stained with an automated immunohistochemistry system (Ventana NexeS IHC, Roche Diagnostics, Meylan, France), incubated with human SSTR subtype-specific rabbit polyclonal antibodies (Gramsch Laboratories/Biotrend, Köln, Germany) (Taniyama et al., 2005) at an optimal dilution/concentration ratio and optimal incubation time (SS-840 (anti-SSTR1): 1/200, 22 min; SS-800 (anti-SSTR2A) 1/200, 22 min; SS-860 (anti-SSTR2B): 1/200 22 min; SS-850 (anti-SSTR3): 1/200, 32 min; SS-880 (anti-SSTR4): 2 μg/mL, 3 h; and SS-890 (anti-SSTR5): 1/200, 32 min)) and then processed with a rabbit-specific horseradish peroxidase/3,3′-diaminobenzidine kit (RUO, Roche Diagnostics, Meylan, France). The specificity of the immunohistochemical reaction was confirmed by the absence of a positive signal after the secondary antibody was used without the primary antibody. The positive control experiments involved normal human pancreatic parenchyma and/or pulmonary carcinoid tumour tissue.
2.2 Experiments on rings of isolated human bronchus
The bronchi (mean ± SD inner diameter: 1.9 ± 0.5 mm) were dissected free from adhering lung parenchyma and connective tissue and then cut into rings of 4–7 mm in length. Eight to 24 rings were obtained from a given patient’s sample and then used as paired preparations. Before use, the rings were stored overnight at +4 °C in a Krebs-Henseleit solution (KHS, in mM: NaCl 119, KCl 5.4, CaCl2 2.5, KH2PO4 1.2, MgSO4 1.2, NaHCO3 25, glucose 11.7) equilibrated with O2/CO2 (95:5). On the following day, human bronchial segments were placed in an isolated organ bath filled with 5 mL of KHS, oxygenated with O2/CO2 (95:5) and thermostated at +37 °C (pH 7.4). No peptidase inhibitors were used. Tension was measured isometrically with a strain gauge (UF1; Piodem, Canterbury, Kent, United Kingdom) connected to an amplifier (EMKA Technologies, Paris, France). Data were acquired, processed and analyzed with a computerized system running IOX v1.56.8 and Datanalyst v1.58 softwares (EMKA Technologies). In all experiments, the bronchial rings were suspended with an initial load of 2 g and equilibrated for 60–90 min. The KHS in the bath was changed every 15–20 min. At the end of the equilibration period, the resting load was stable at ∼1–3 g. As described previously, bronchi were first contracted maximally with acetylcholine (ACh, 3 mM) and then washed and equilibrated again for 60 min before the experimental procedures were initiated (Grassin-Delyle et al., 2014; Heusler et al., 2015; Naline et al., 2018). A total of 326 rings were used in the experiments on isolated rings of human bronchus. The ranges of agonist concentrations employed here were chosen to maximize the discrimination between the various SSTRs, based on the literature data (Rohrer et al., 1998; Patel, 1999; Rohrer and Schaeffer, 2000; Günther et al., 2018).
2.2.1 Effects on basal tone and the contractile responses to exogenous ACh
Each agonist’s effect on a ring’s basal tone was assessed by cumulative addition (0.1 nM–1 μM, in logarithmic increments) at 15 min intervals. To investigate the agonist’s respective effects on ACh- or KCl-induced contractions, a first cumulative concentration-response curve (CRC) was obtained for ACh (10 nM–1 mM) or KCl (1 mM–1 M). After extensive washing and equilibration for 1 h, the rings were incubated with the agonist for 10 min prior to the measurement of a second cumulative CRC for ACh or KCl (Advenier et al., 1986; Naline et al., 2018). The CRC-derived data on efficacy and potency were expressed as Emax and–log EC50 (pD2), respectively.
2.2.2 Electrical field stimulation–induced contraction of human bronchus
In order to trigger the neural release of ACh, electrical field stimulation (EFS) experiments were performed as described previously (Grassin-Delyle et al., 2014; Naline et al., 2018). EFS was performed in organ baths fitted with two platinum plate electrodes placed alongside the tissue (10 mm apart) and connected to a stimulator (EMKA Technologies). A stimulation train consists in biphasic square-wave pulses with a constant current of 320 mA and a pulse duration of 1 ms delivered for 10 s at 5 Hz. The EFS-induced contractions at frequencies ranging from 5 to 40 Hz in paired human bronchi samples with inner diameters ranging from 1 to 3 mm were similar (Naline et al., 2018). These contractions were fully blocked by atropine (1 µM) and tetrodotoxin (1 µM); this demonstrated the involvement of ACh release from cholinergic nerves, as shown previously in our laboratory (Grassin-Delyle et al., 2014; Naline et al., 2018) and in others (Watson et al., 1998). For the EFS experiments, the cyclo-oxygenase inhibitor indomethacin (1 µM), and the cysteinyl leukotriene antagonist MK476 (1 µM) were added to the KHS at the beginning of the experiments; this respectively avoided the influence of leukotrienes and prostaglandins on the neuronal responses (Watson et al., 1998; Naline et al., 2007; Naline et al., 2018; Grassin-Delyle et al., 2014). At the end of the equilibration period, the resting load was stable at ∼1 g. Eight to 16 bronchial rings were simultaneously tested, along with at least one time-control preparation per series of 8 rings (to assess the response for the duration of the experiments). To assess each preparation’s baseline response, a first train of EFS was applied twice at 10 min intervals. This stimulation (at 5 Hz, as used by Sekizawa et al. (1989)) caused a contraction corresponding to 32% ± 6% of the maximum response to 40 Hz stimulation (n = 8), in order to leave room for an increase in EFS-induced contraction (Sekizawa et al., 1989; Fernandes et al., 1999; Grassin-Delyle et al., 2014; Naline et al., 2018). SST, the SSTR agonists or vehicle was added to the bath 10 min before the beginning of a second train of stimulations (delivered every 10 min for 1 h and then every 20 min for 3 h). A given ring was used to study only one concentration of one compound (Grassin-Delyle et al., 2014; Naline et al., 2018).
2.3 Drugs
SST (UCB, Nanterre, France) and octreotide (Sandostatin™, Novartis Pharma, Rueil-Malmaison, France) were obtained as solutions for injection. The SSTR1 agonist CH-275 (Des-AA1,2,5-[DTrp8,IAmp9]-SRIF) was purchased from Neo MPS (Strasbourg, France) (Rivier et al., 2001). The other selective SSTR agonists (SSTR2: L-779,976; SSTR3: L-796,778; SSTR4: L-803,087; SSTR5: L-817,818) were kindly provided by Dr Susan P. Rohrer (Merck Research Laboratories, Rahway, NJ, United States). L-796,778 is a partial agonist for SSTR3 (Rohrer et al., 1998; Rohrer and Schaeffer, 2000). The SSTR agonists’ selectivity is described in Supplementary Table S1. All the agonists were dissolved to a concentration of 10−2 M in DMSO, and subsequent dilutions were prepared in assay buffer. ACh and atropine were obtained from Sigma (St Louis, MI, United States).
2.4 Data analysis
The experimental data are quoted as the mean ± standard error of the mean (SEM); the n represents the number of donors. For each bronchial ring, the values were expressed as the percentage of the contraction obtained with either the maximum concentration of ACh or during the initial EFS train.
A one- or two-way repeated-measures analysis of variance (ANOVA) and then Dunnett’s post-test for multiple comparisons were used to assess the EFS data. The effects of agonist concentration and stimulation time were evaluated in a two-way ANOVA. The EFS data were also evaluated in a one-way ANOVA after calculation of the areas under the concentration-time curves (AUC), using the linear trapezoidal method. The agonists’ effects on the CRC (Emax and pD2) for ACh or KCl were analyzed in a one-way ANOVA. The threshold for statistical significance was set to p < 0.05. All analyses were performed using GraphPad Prism software (version 8.4.2, GraphPad Software Inc., San Diego, CA, United States).
3 Results
3.1 Immunohistochemical detection of the SST receptors on human bronchus and lung parenchyma
In the bronchus, all the SSTRs were expressed on the parasympathetic ganglia (rank order: SST4 > other SSTRs) and the submucosal glands (rank order: SSTR1,2A,2B,3,5 > SSTR4). On smooth muscle, SSTR4 and (to a lesser extent) SSTR2A, SSTR3 and SSTR5 were expressed but SSTR1 and SSTR2B were not (Table 1; Figure 1). In the parenchyma, all the SSTRs other than SSTR4 were expressed by bronchiolar epithelial cells and some pneumocytes. Lastly, all the SSTRs other than SST2A and SST4 were expressed by alveolar macrophages (Table 1).
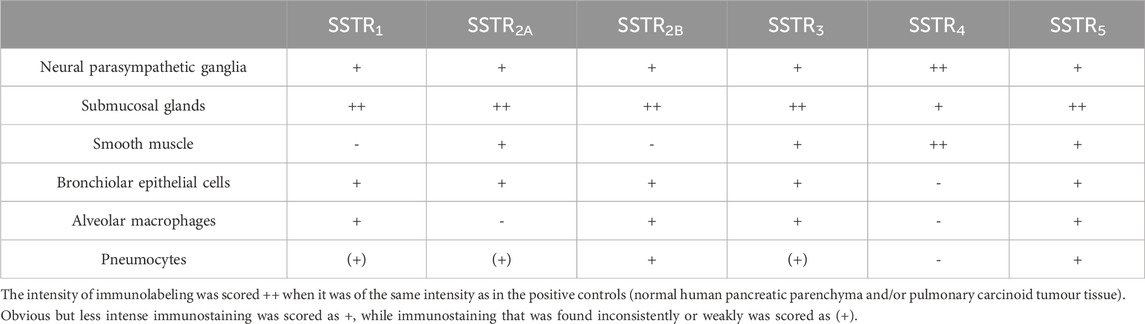
Table 1. The relative abundance of SST receptor immunoreactivity in human bronchus and lung parenchyma.
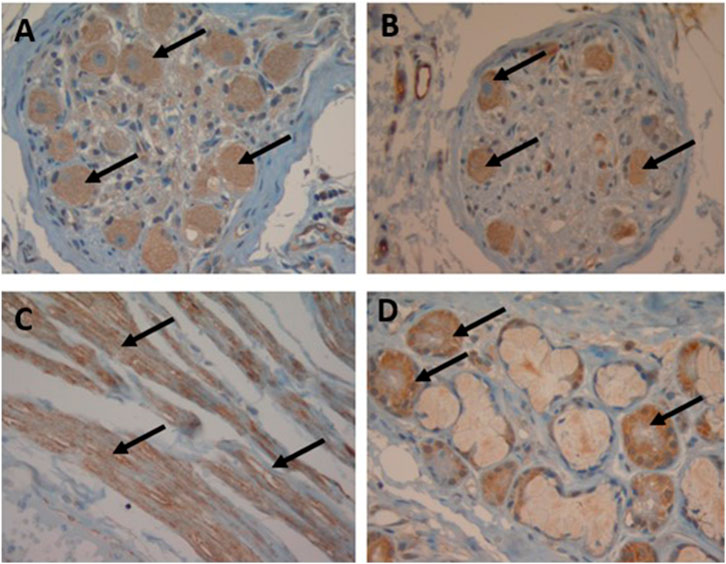
Figure 1. Immunohistochemical localization of SST receptors in the human bronchus. Arrows indicate SST receptor positivity (magnification: 40x): neural ganglia staining with SSTR3 (A) and SSTR5 (B), smooth muscle staining with SSTR4 (C) and submucosal gland staining with SSTR1 (D).
3.2 Effect of SST and SSTR agonists on the EFS-induced contraction of human bronchus
SST at a concentration of 10−7 M induced a small (∼10%, p = 0.06) increase in EFS-induced contraction. The increase in EFS-induced contraction caused by octreotide was concentration-dependent. At the optimal concentration of 10−7 M, octreotide caused a significant increase in EFS-induced contraction versus the control; the mean maximum increase was 22% ± 8% (p < 0.05) at the 50th minute (Figure 2). The mean increase in the AUC was also significant with octreotide 10−7 M (33% ± 5%*min, P < 0.001).
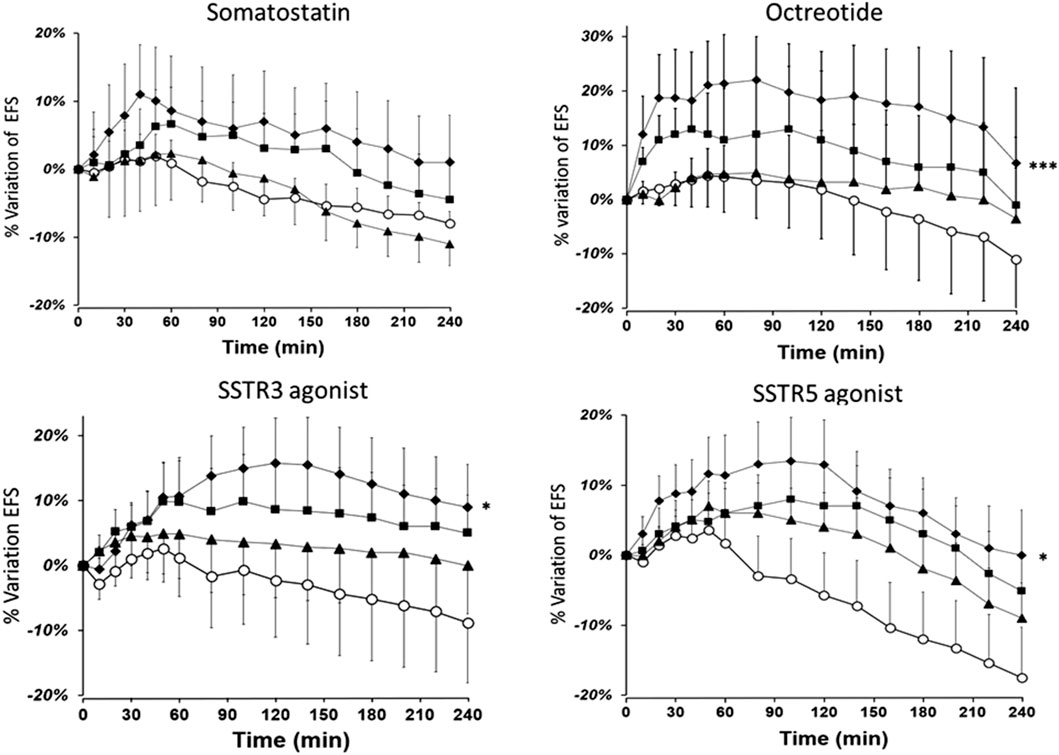
Figure 2. The time course of the enhancing effect of octreotide and the SSTR3 and SSTR5 agonists on the EFS-induced contraction of human bronchial rings. The data are quoted as the mean ± SEM percentage of EFS-induced contraction of paired bronchial rings from 9 to 10 patients. (O: controls; ▲10−9 M, ■: 10−8 M, and ◆10−7 M for octreotide and the SSTR5 agonist; ▲10−8 M, ■: 10−7 M, and ◆ 3 × 10−7 M for the SSTR3 agonist), *: P < 0.05, ***: P < 0.001).
Furthermore, the optimal concentrations of the SSTR3 (3 × 10-7 M) and SSTR5 (10−7 M) agonists caused a significant increase in EFS-induced contraction. The mean maximum increases induced by the SSTR3 and SSTR5 agonists were 16% ± 4% and 19% ± 5% after 90 and 120 min of stimulation, respectively. At the optimal concentrations, the mean increases in the AUC were also significant (19% ± 6%*min, p = 0.012 for SSTR3, and 22% ± 7%*min, p = 0.013 for SSTR5). In contrast, the agonists of the other SSTR receptors (SSTR1, SSTR2, and SSTR4) did not alter EFS-induced contraction (n = 6–8) (Supplementary Figure S1).
3.3 Effect of SST and SSTR agonists on the resting tone and the acetylcholine-induced contraction of human bronchus
Neither SST, octreotide, nor any of the SSTR agonists was associated with a change in the resting tone. The contractile responses to the cumulative addition of increasing concentrations of ACh were not modified by pre-incubation of the rings with SST, octreotide, or the SSTR agonists, since there was no shift in the CRC and no significant difference in the maximum response (Table 2; Supplementary Figure S2). Since SSTRs can inhibit voltage-operated calcium channels, we also tested the effects of octreotide and SSTR3 and SSTR4 agonists on KCl-induced contraction (Advenier et al., 1984; Advenier et al., 1986). None of these three SSTR agonists altered KCl-induced contraction (n = 5-6, data not shown).
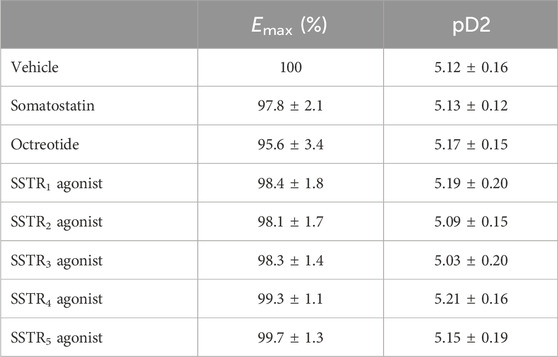
Table 2. The effects of the SSTR agonists on the contraction induced by exogenous ACh (n = 5–6). The concentrations of SSTR agonists used here were either the optimal concentration with respect to the effect on the EFS-induced contraction (for octreotide (0.1 µM), the SSTR3 agonist (0.3 µM) and the SSTR5 agonist (0.1 µM)) or the maximum concentration tested (for somatostatin and the other SSTR agonists). The data are quoted as the mean ± SEM.
4 Discussion
Our study generated new data on the locations of SST receptors in the human lung. We notably found that in the bronchial wall, all types of receptor were expressed in the parasympathetic nerve ganglia. In bronchial smooth muscle, only SSTR1 and SSTR2B receptors were not expressed. We suggest that in human bronchi, (i) the activation of prejunctional SSTR3 and SSTR5 receptors potentiates the cholinergic-nerve-mediated contraction induced by EFS, and (ii) neither SSTR has a direct effect on muscle tone or acetylcholine-induced contraction.
Our present results confirmed the report by Taniyama et al. (2005) and showed that all types of SSTR are indeed expressed by bronchial submucosal glands. It has been reported that human tissues contain only the SSTR2A variant, whereas both SSTR2A and SSTR2B have been identified in rodent tissues (Günther et al., 2018). However, expression of SSTR2A and SSTR2B has been reported in human tissues, including thyrotropin-secreting pituitary adenomas, thyroid tumours (Pisarek et al., 2015; Thodou and Kontogeorgos, 2020), and the lung (alveolar macrophages (SSTR2B), pneumocytes, submucosal glands) (Taniyama et al., 2005; Borie et al., 2008). In the present study, the two SSTR2 forms were found on the neural ganglia, the submucosal glands, the bronchiolar epithelial cells and the pneumocytes, while SSTR2A was expressed by smooth muscle and SSTR2B was expressed by alveolar macrophages. SSTR2B receptor expression is low in normal lung parenchyma but is reported elevated in the fibrotic lung (particularly on epithelial cells) (Borie et al., 2008). Our results suggest that alveolar macrophages express SSTR1 and SSTR3 in addition to SSTR2B (Borie et al., 2008) and SSTR5 (Günther et al., 2018).
We did not detect SSTR4 on bronchiolar epithelial cells (which nevertheless expressed all the other SSTRs) or macrophages (in contrast to the report by Varecza et al. (2009)). However, Varecza et al. mainly observed SSTR4 labelling of macrophages in samples from inflamed lungs (pneumonia and bronchiectasis). To the best of our knowledge, only SSTR4 expression has been documented in human bronchial smooth muscle (Varecza et al., 2009). We also found that SSTR2A, SSTR3 and SSTR5 were expressed by human bronchial smooth muscle.
SST is present in enteric neurons and SSTR1, SSTR2, and SSTR3 receptors are expressed on enteric ganglia and the corresponding nerve fibres and nerve terminals (Günther et al., 2018). Given that the airways’ nerves are derived embryologically from those of the gut, the presence of SSTR in the airway ganglia was not unexpected (Barnes, 1990). Indeed, we found that all the SSTR were expressed by the airway neural ganglia. In the present study, lung tissue was obtained from former smokers who had undergone surgery for cancer, which limits the extrapolation of our findings on SSTR receptor localization to healthy human lungs.
To the best of our knowledge, the present study is the first to have studied the effects of SST and SSTR agonists on the human bronchus. Neither SST, octreotide, nor any of the other SSTR agonists changed the resting tone or maximum response or shifted the CRC for ACh or KCl. The absence of direct effects is in line with a report on the ferret trachea, in which SST did not significantly change the resting muscle tone or the CRC for ACh (Sekizawa et al., 1989).
With regard to human smooth muscles other than those of bronchial origin, SSTR1, SSTR2, and SSTR4 are expressed in the aorta, the internal mammary artery, and the saphenous vein (Curtis et al., 2000), whereas SSTR2 is involved in the contraction of isolated saphenous vein (Dimech et al., 1995). In the gastrointestinal tract, the circular smooth muscle expresses SSTR2 only and the longitudinal smooth muscle expresses SSTR1, SSTR2, and SSTR3 (Corleto et al., 2006). SST exerted a relatively weak contractile effect (around a third of that of carbachol) on the circular and longitudinal human colonic smooth muscle cells; this effect was thought to be mediated by the activation of SSTR1 and SSTR2 (Corleto et al., 2006). However, SST’s main effect was SSTR2-mediated relaxation of carbachol-induced contraction (Corleto et al., 2006).
Most of the best-characterized actions of the SST receptors are inhibitory. Activation of Gi/Go proteins by SST caused a reduction in cAMP by inhibition of adenylyl cyclase, and inhibition of calcium influx through voltage-operated calcium channels (Günther et al., 2018). These inhibitory effects are additive in excitable cells, such as neurons (Günther et al., 2018). However, it has been shown that SST stimulated the release of ACh in the myenteric and submucous plexus of the guinea pig ileum (Yau et al., 1983; Lu et al., 1990). More specifically, SST potentiated EFS-induced contraction of the ferret trachea, although the researchers did not determine which SSTR were involved (Sekizawa et al., 1989). Our results suggest that SSTR agonists potentiate EFS-induced contraction of the human bronchus. SST’s weak modulatory effect on EFS-induced contraction in the present study is probably related to the molecule’s short half-life–a drawback that limits its therapeutic use (Günther et al., 2018). Accordingly, two stable SST analogues (octreotide and lanreotide) have been developed for clinical use (Chen et al., 2023). Octreotide is highly selective for SSTR2, SSTR3 and SSTR5 (Supplementary Table S1). We found that SSTR3 and SSTR5 agonists potentiated EFS-induced contraction, whereas an SSTR2 agonist had no effect on EFS-induced contraction. The apparently greater effect of octreotide on EFS-induced contraction is probably explained by the dual stimulation exerted by this agonist on the SSTR3 and SSTR5 receptors and by the partial agonist activity of the SSTR3 agonist. Given that SSTR are definitely present on bronchial parasympathetic nerve ganglia and probably present on post-ganglionic nerve fibres, our results suggest that stimulation of the SSTR3 and SSTR5 receptors potentiates the cholinergic-nerve-mediated contraction induced by EFS in human bronchi via presynaptic mechanisms. Our findings may help explain the occurrence of dyspnea or bronchospasm during treatment with somatostatin and octreotide.
Very few mediators have been shown to potentiate cholinergic neurotransmission in the human bronchus. Along with SST, endothelin-1 was found to potentiate EFS-mediated contraction in the human bronchus via activation of cholinergic nerve ETA and ETB receptors (Fernandes et al., 1999). The amplitude of the endothelin-potentiating effect ( ̴ 20%) was of the same order as that of octreotide and the SSTR3 and SSTR5 agonists (Fernandes et al., 1999; D’Agostino et al., 2001). Even though neurokinins can facilitate cholinergic neurotransmission in the rabbit airways (via NK1 and NK2 receptors) and guinea pig airways (via the NK1 receptor), they have no demonstrable effects on human airways (Belvisi et al., 1994).
5 Conclusion
The present study provided new data on the location of SST receptors in the human lung. Our results suggest that the activation of prejunctional SST3 and SST5 receptors potentiates the cholinergic-nerve-mediated contraction induced by EFS in human bronchi.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The studies involving humans were approved by Comité de Protection des Personnes Ile de France VIII, Boulogne-Billancourt, France; reference: CPP8 - DC 11 10 05. The studies were conducted in accordance with the local legislation and institutional requirements. The human samples used in this study were acquired from Our hospital’s pathology department supplied lung tissue from macroscopically healthy parts of the lungs patients The use of human lung tissue for in vitro experiments was approved by the local independent ethics committee (Comité de Protection des Personnes Ile de France VIII, Boulogne-Billancourt, France; reference: CPP8 - DC 11 10 05). Each patient gave their informed consent to the use of surgical samples for research purposes. Written informed consent for participation was not required from the participants or the participants’; legal guardians/next of kin in accordance with the national legislation and institutional requirements.
Author contributions
MB: Conceptualization, Formal Analysis, Investigation, Writing – original draft. SG-D: Data curation, Formal Analysis, Investigation, Writing – review and editing. CR: Data curation, Formal Analysis, Writing – review and editing. EL: Resources, Writing – review and editing. IM-B: Investigation, Resources, Writing – review and editing. MG: Resources, Writing – review and editing. HS: Writing – review and editing. PD: Conceptualization, Formal Analysis, Writing – original draft, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The publications costs were covered by Foch Hospital.
Acknowledgements
The authors thank Emmanuel Naline, PharmD, PhD for technical expertise and assistance with the EFS experiments and David Fraser, D. Phil. (Biotech Communication SARL, Ploudalmézeau, France) for copy-editing services.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fphar.2025.1677183/full#supplementary-material
References
Advenier, C., Cerrina, J., Duroux, P., Floch, A., and Renier, A. (1984). Effects of five different organic calcium antagonists on guinea-pig isolated trachea. Br. J Pharmacol. 82, 727–733. doi:10.1111/j.1476-5381.1984.tb10812.x
Advenier, C., Naline, E., and Renier, A. (1986). Effects of Bay K 8644 on contraction of the human isolated bronchus and guinea-pig isolated trachea. Br. J Pharmacol. 88, 33–39. doi:10.1111/j.1476-5381.1986.tb09468.x
Balibrea, J. L., Arias-Díaz, J., García, C., and Vara, E. (1994). Effect of pentoxifylline and somatostatin on tumour necrosis factor production by human pulmonary macrophages. Circ. Shock 43, 51–56.
Barnes, P. J. (1990). Muscarinic receptors in airways: recent developments. J. Appl. Physiology 68, 1777–1785. doi:10.1152/jappl.1990.68.5.1777
Belvisi, M. G., Patacchini, R., Barnes, P. J., and Maggi, C. A. (1994). Facilitatory effects of selective agonists for tachykinin receptors on cholinergic neurotransmission: evidence for species differences. Br. J. Pharmacol. 111, 103–110. doi:10.1111/j.1476-5381.1994.tb14030.x
Borie, R., Fabre, A., Prost, F., Marchal-Somme, J., Lebtahi, R., Marchand-Adam, S., et al. (2008). Activation of somatostatin receptors attenuates pulmonary fibrosis. Thorax 63, 251–258. doi:10.1136/thx.2007.078006
Chen, S., Teng, X., and Zheng, S. (2023). Molecular basis for the selective G protein signaling of somatostatin receptors. Nat. Chem. Biol. 19, 133–140. doi:10.1038/s41589-022-01130-3
Corleto, V. D., Severi, C., Romano, G., Tattoli, I., Weber, H. C., Stridsberg, M., et al. (2006). Somatostatin receptor subtypes mediate contractility on human colonic smooth muscle cells. Neurogastroenterol. Motil. 18, 217–225. doi:10.1111/j.1365-2982.2005.00752.x
Curtis, S. B., Hewitt, J., Yakubovitz, S., Anzarut, A., Hsiang, Y. N., and Buchan, A. M. J. (2000). Somatostatin receptor subtype expression and function in human vascular tissue. Am. J. Physiology-Heart Circulatory Physiology 278, H1815–H1822. doi:10.1152/ajpheart.2000.278.6.H1815
Dimech, J., Feniuk, W., Latimer, R. D., and Humphrey, P. P. A. (1995). Somatostatin-induced contraction of human isolated saphenous vein involves sst2 receptor-mediated activation of L-Type calcium channels. J. Cardiovasc. Pharmacol. 26, 721–728. doi:10.1097/00005344-199511000-00008
D’Agostino, B., Advenier, C., Falciani, M., Gallelli, L., Marrocco, G., Piegari, E., et al. (2001). Endothelin-1 increases cholinergic nerve-mediated contraction of human bronchi via tachykinin synthesis induction. Br. J Pharmacol. 134, 1447–1454. doi:10.1038/sj.bjp.0704395
Elekes, K., Helyes, Z., Kereskai, L., Sándor, K., Pintér, E., Pozsgai, G., et al. (2008). Inhibitory effects of synthetic somatostatin receptor subtype 4 agonists on acute and chronic airway inflammation and hyperreactivity in the mouse. Eur. J. Pharmacol. 578, 313–322. doi:10.1016/j.ejphar.2007.09.033
Feniuk, W., Dimech, J., and Humphrey, P. P. A. (1993). Characterization of somatostatin receptors in guinea-pig isolated ileum, vas deferens and right atrium. Br. J Pharmacol. 110, 1156–1164. doi:10.1111/j.1476-5381.1993.tb13935.x
Fernandes, L., Henry, P., and Goldie, R. (1999). Endothelin-1 potentiates cholinergic nerve-mediated contraction in human isolated bronchus. Eur. Respir. J. 14, 439–442. doi:10.1034/j.1399-3003.1999.14b33.x
Grassin-Delyle, S., Naline, E., Buenestado, A., Faisy, C., Alvarez, J.-C., Salvator, H., et al. (2014). Cannabinoids inhibit cholinergic contraction in human airways through prejunctional CB1 receptors. Br. J. Pharmacol. 171, 2767–2777. doi:10.1111/bph.12597
Gugger, M., Waser, B., Kappeler, A., Schonbrunn, A., and Reubi, J. (2004). Immunohistochemical localization of somatostatin receptor sst2A in human gut and lung tissue: possible implications for physiology and carcinogenesis. Ann. N. Y. Acad. Sci. 1014, 132–136. doi:10.1196/annals.1294.013
Günther, T., Tulipano, G., Dournaud, P., Bousquet, C., Csaba, Z., Kreienkamp, H.-J., et al. (2018). International union of basic and clinical pharmacology. CV. Somatostatin receptors: structure, function, ligands, and new nomenclature. Pharmacol. Rev. 70, 763–835. doi:10.1124/pr.117.015388
Helyes, Z., Pintér, E., Németh, J., Sándor, K., Elekes, K., Szabó, Á., et al. (2006). Effects of the somatostatin receptor subtype 4 selective agonist J-2156 on sensory neuropeptide release and inflammatory reactions in rodents. Br. J Pharmacol. 149, 405–415. doi:10.1038/sj.bjp.0706876
Helyes, Z., Elekes, K., Németh, J., Pozsgai, G., Sándor, K., Kereskai, L., et al. (2007). Role of transient receptor potential vanilloid 1 receptors in endotoxin-induced airway inflammation in the mouse. Am. J. Physiology-Lung Cell. Mol. Physiology 292, L1173–L1181. doi:10.1152/ajplung.00406.2006
Helyes, Z., Pintér, E., Sándor, K., Elekes, K., Bánvölgyi, Á., Keszthelyi, D., et al. (2009). Impaired defense mechanism against inflammation, hyperalgesia, and airway hyperreactivity in somatostatin 4 receptor gene-deleted mice. Proc. Natl. Acad. Sci. U.S.A. 106, 13088–13093. doi:10.1073/pnas.0900681106
Heusler, P., Cussac, D., Naline, E., Tardif, S., Clerc, T., and Devillier, P. (2015). Characterization of V0162, a new long-acting antagonist at human M3 muscarinic acetylcholine receptors. Pharmacol. Res. 100, 117–126. doi:10.1016/j.phrs.2015.07.033
Lu, Y., Wiley, J., and Owyang, C. (1990). Mechanisms regulating somatostatin release and somatostatin-induced acetylcholine release from the myenteric plexus. Metabolism 39, 131–133. doi:10.1016/0026-0495(90)90230-A
Naline, E., Trifilieff, A., Fairhurst, R. A., Advenier, C., and Molimard, M. (2007). Effect of indacaterol, a novel long-acting β2 -agonist, on isolated human bronchi. Eur. Respir. J. 29, 575–581. doi:10.1183/09031936.00032806
Naline, E., Grassin Delyle, S., Salvator, H., Brollo, M., Faisy, C., Victoni, T., et al. (2018). Comparison of the in vitro pharmacological profiles of long-acting muscarinic antagonists in human bronchus. Pulm. Pharmacol. and Ther. 49, 46–53. doi:10.1016/j.pupt.2018.01.003
Patel, Y. C. (1999). Somatostatin and its receptor family. Front. Neuroendocrinol. 20, 157–198. doi:10.1006/frne.1999.0183
Pintér, E., Helyes, Z., and Szolcsányi, J. (2006). Inhibitory effect of somatostatin on inflammation and nociception. Pharmacol. and Ther. 112, 440–456. doi:10.1016/j.pharmthera.2006.04.010
Pisarek, H., Pawlikowski, M., Marchlewska, M., Minias, R., and Winczyk, K. (2015). An immunohistochemical investigation of the expression of somatostatin receptor subtypes – should therapeutic trials be performed to determine the efficacy of somatostatin analogs in treating advanced thyroid malignances? Exp. Clin. Endocrinol. Diabetes 123, 342–346. doi:10.1055/s-0035-1548825
Rivier, J. E., Hoeger, C., Erchegyi, J., Gulyas, J., DeBoard, R., Craig, A. G., et al. (2001). Potent somatostatin undecapeptide agonists selective for somatostatin receptor 1 (sst1). J. Med. Chem. 44, 2238–2246. doi:10.1021/jm010037+
Rohrer, S. P., and Schaeffer, J. M. (2000). Identification and characterization of subtype selective somatostatin receptor agonists. J. Physiology-Paris 94, 211–215. doi:10.1016/S0928-4257(00)00215-1
Rohrer, S. P., Birzin, E. T., Mosley, R. T., Berk, S. C., Hutchins, S. M., Shen, D.-M., et al. (1998). Rapid identification of subtype-selective agonists of the somatostatin receptor through combinatorial chemistry. Science 282, 737–740. doi:10.1126/science.282.5389.737
Sekizawa, K., Graf, P. D., and Nadel, J. A. (1989). Somatostatin potentiates cholinergic neurotransmission in ferret trachea. J. Appl. Physiology 67, 2397–2400. doi:10.1152/jappl.1989.67.6.2397
Taniyama, Y., Suzuki, T., Mikami, Y., Moriya, T., Satomi, S., and Sasano, H. (2005). Systemic distribution of somatostatin receptor subtypes in human: an immunohistochemical study. Endocr. J. 52, 605–611. doi:10.1507/endocrj.52.605
Thodou, E., and Kontogeorgos, G. (2020). Somatostatin receptor profile in pituitary thyrotroph adenomas. Clin. Neurology Neurosurg. 195, 105865. doi:10.1016/j.clineuro.2020.105865
Varecza, Z., Elekes, K., László, T., Perkecz, A., Pintér, E., Sándor, Z., et al. (2009). Expression of the somatostatin receptor subtype 4 in intact and inflamed pulmonary tissues. J. Histochem Cytochem 57, 1127–1137. doi:10.1369/jhc.2009.953919
Watson, N., Magnussen, H., and Rabe, K. F. (1998). The relevance of resting tension to responsiveness and inherent tone of human bronchial smooth muscle. Br. J Pharmacol. 123, 694–700. doi:10.1038/sj.bjp.0701637
Keywords: somatostatin receptors, human, lung, bronchus, contraction, immunohistochemistry, cholinergic nerve
Citation: Brollo M, Grassin-Delyle S, Roquencourt C, Longchampt E, Miguet-Besson I, Glorion M, Salvator H and Devillier P (2025) Somatostatin receptors 3 and 5 potentiate cholinergic-nerve-mediated contraction in human bronchus. Front. Pharmacol. 16:1677183. doi: 10.3389/fphar.2025.1677183
Received: 31 July 2025; Accepted: 22 October 2025;
Published: 06 November 2025.
Edited by:
Venkata Ramireddy Narala, Yogi Vemana University, IndiaReviewed by:
Katarzyna Kaczynska, Polish Academy of Sciences, PolandAparna Renigunta, University of Marburg, Germany
Copyright © 2025 Brollo, Grassin-Delyle, Roquencourt, Longchampt, Miguet-Besson, Glorion, Salvator and Devillier. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Philippe Devillier, cC5kZXZpbGxpZXJAaG9waXRhbC1mb2NoLmNvbQ==