 Ignac Ovari1†
Ignac Ovari1† Agnes Boglarka Brezniczky2†Attila Laczovics2Ervin Berényi2Tamas Erdei1
Agnes Boglarka Brezniczky2†Attila Laczovics2Ervin Berényi2Tamas Erdei1 Oluwatofunmi Ojo1Bence Hornok1
Oluwatofunmi Ojo1Bence Hornok1 Bela Juhasz1Zoltan Szilvassy1
Bela Juhasz1Zoltan Szilvassy1 Judit Zsuga3Gabor Viczjan1
Judit Zsuga3Gabor Viczjan1 Rudolf Gesztelyi1*
Rudolf Gesztelyi1*- 1Department of Pharmacology and Pharmacotherapy, Faculty of Medicine, University of Debrecen, Debrecen, Hungary
- 2Department of Radiology and Imaging Science, Faculty of Medicine, University of Debrecen, Debrecen, Hungary
- 3Department of Psychiatry, Faculty of Medicine, University of Debrecen, Debrecen, Hungary
Introduction: In this study, we investigated the influence of AB-680, a highly potent CD73 inhibitor, on the effect of NBTI, a nucleoside transport blocker, exerted on concentration-effect (E/c) curves generated with CPA, a relatively stable, selective A1 adenosine receptor full agonist, in isolated, paced guinea pig left atria.
Methods: Transformations of the CPA E/c curves, constructed in the absence and presence of AB-680 and NBTI (in all combinations), were used to assess the changes in the interstitial adenosine concentration. These changes were quantified with the receptorial responsiveness method (RRM), a unique procedure providing the CPA concentration (as cx), which is equieffective with the increase in the interstitial adenosine concentration caused by NBTI. AB-680 and NBTI were dissolved in DMSO (recommended for in vitro use) as well as in a buffer (recommended for in vivo use), and the results were compared.
Results and discussion: We found that AB-680, when added alone, did not affect the response to CPA. In turn, AB-680, when administered together with NBTI, was able to partially reverse the elevating effect of NBTI on the interstitial adenosine level. Nevertheless, the inhibitory action of AB-680 on the effect of NBTI appeared to be smaller than that of PSB-12379, another CD73 inhibitor investigated earlier in the same experimental model. We also found that DMSO interfered with our measurements to a lesser extent than the buffer recommended for in vivo studies. In addition, AB-680, when co-administered with NBTI (both dissolved in DMSO), reduced cx (i.e. probably also the surplus interstitial adenosine) by at least half.
1 Introduction
Adenosine, in addition to its central role as a metabolite (precursor and degradation product of ATP), is an important humoral regulator in almost all tissues of higher organisms (Fredholm et al., 2001; 2011; Burnstock et al., 2010; IJzerman et al., 2022). The regulatory effects of adenosine are predominantly mediated by cell-surface adenosine receptors (A1, A2A, A2B, and A3); thus, it is the interstitial adenosine level that plays a key role in the adenosinergic control. In the myocardium, A1 is the main adenosine receptor type (Fredholm et al., 2001; Szentmiklósi et al., 2011; Headrick et al., 2013; Qian et al., 2025).
In the heart, the complex regulatory actions of adenosine occur predominantly during cellular energy (ATP) deficiency, a pathological condition characterized by an increase in the interstitial adenosine concentration (Szentmiklósi et al., 2011; Headrick et al., 2013; Qian et al., 2025). In this situation, the main source of interstitial adenosine is the intracellularly formed adenosine that is released from the cells via equilibrative nucleoside transporters, including ENT1, which is NBTI-sensitive (Schreieck and Richardt, 1999; Deussen et al., 1999; 2006; Deussen, 2000a; 2000b; Szentmiklósi et al., 2011; Burnstock and Pelleg, 2015).
In turn, under physiological circumstances, interstitial adenosine is primarily formed from ATP, which is released from the cells and then dephosphorylated to adenosine by ectonucleotidases, mainly by CD39 (ecto-apyrase) and consecutively by CD73 (ecto-5′-nucleotidase) (Hesse et al., 2017; Zimmermann, 2000; Qian et al., 2025; for a brief synopsis of ectonucleotidases, see Table 1 in Viczjan et al., 2021). In this condition, contrary to the energy-depleted state, the equilibrative nucleoside carriers transport adenosine into the cells because the adenosine formation exceeds the adenosine elimination in the interstitium (Deussen et al., 1999; Deussen, 2000a; Deussen, 2000b).
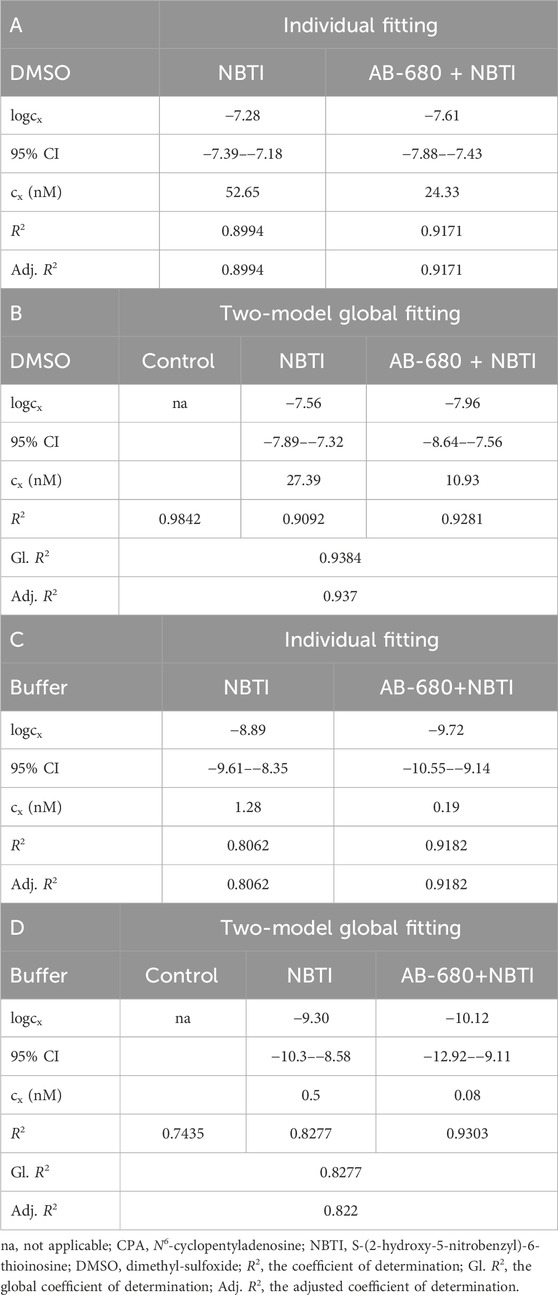
Table 1. Best-fit values (logcx) with their 95% confidence intervals (95% CI) and antilog values (cx) obtained with the receptorial responsiveness method (RRM). The cx values, converted to nM (nmol/L), are the estimates for the interstitial adenosine accumulation in response to NBTI (as the equieffective CPA concentrations). RRM was implemented in two alternative ways: individual fitting (panels A and C) and two-model global fitting (panels B and D). The CPA E/c curves (in the three experimental groups named control, NBTI, and AB-680 + NBTI) were constructed in the presence of DMSO (panels A and B) or a buffer recommended for in vivo use (panels C and D) as solvents.
Consistent with the above-mentioned well-established observations, we previously found that the inhibition of ENT1 by NBTI (S-(2-hydroxy-5-nitrobenzyl)-6-thioinosine) markedly elevated the interstitial adenosine level in the metabolically intact, isolated guinea pig left atrium (Karsai et al., 2006; 2007; Viczjan et al., 2021). On the conventionally constructed and plotted concentration–effect (E/c) curve of CPA (N6-cyclopentyladenosine), which is a relatively stable, selective A1 adenosine receptor full agonist (Fredholm et al., 2001; Pavan and IJzerman, 1998), this effect of NBTI manifested in a considerable decrease in the maximal effect (Emax) and potency (1/EC50). This phenomenon was explained by the fact that the surplus interstitial adenosine accumulated by NBTI consumed a part of the response capacity of the A1 adenosine receptors during the incubation period with NBTI (i.e., before the construction of the E/c curve with the given A1 adenosine receptor agonist) (Gesztelyi et al., 2003; Karsai et al., 2006; Karsai et al., 2007).
We also found that PSB-12379 (N6-benzyl-α,β-methyleneadenosine-5′-diphosphate), a potent CD73 inhibitor (Bhattarai et al., 2015; Schmies et al., 2020), significantly inhibited the aforementioned effect of NBTI on the CPA E/c curve; i.e., it increased the response to CPA in the presence of NBTI (compared to the state when only NBTI was present). This phenomenon was attributed to the fact that PSB-12379 decreased the interstitial adenosine formation and, thereby, the ability of NBTI to increase the interstitial level of endogenous adenosine (in an energetically physiological condition) (Viczjan et al., 2021).
AB-680 is an extremely potent but slow-onset competitive inhibitor of CD73 (Bowman et al., 2019). The significance of AB-680 stems from its promise as an antitumor agent that appears to restore the antineoplastic activity of the immune system, which has been impaired by the high adenosine level characteristic of the interior of solid tumors (Piovesan et al., 2022; Strickland et al., 2024; Gao et al., 2025). However, to the best of our knowledge, no investigation has yet dealt with the effect of AB-680 on the heart or cardiac-derived tissues. Given the pivotal impact of the interstitial adenosine level on the heart, it is important to examine any agent that may also affect it in this regard.
In the present study, we aimed to investigate the action of AB-680 on the effect of NBTI exerted on the A1 adenosine receptor-mediated response, which is detectable with conventionally constructed and plotted CPA E/c curves, in the isolated, paced guinea pig left atrium. Transformations of the CPA E/c curves generated in the absence and presence of AB-680 and NBTI (in all combinations) were used to assess the changes in the interstitial adenosine concentration. These changes were quantified with the receptorial responsiveness method (RRM). RRM is a unique procedure providing the concentration of the agonist used for the E/c curve, which is equieffective with the surplus interstitial adenosine concentration compared to a predefined baseline (Gesztelyi et al., 2004; Grenczer et al., 2010a; Grenczer et al., 2010b). RRM was implemented according to the recommendations of our recent study (Ovari et al., 2024), performing both individual fitting and two-model global fitting.
The evaluation of the influence of AB-680 on atrial contractility, both alone and in combination with a nucleoside transport inhibitor, can provide insights into the quantitative relationships of the adenosinergic mechanisms in the atrial myocardium. In addition, these results might inform us about the nature and extent of some potential side effects of this CD73-inhibitor drug candidate in the heart.
2 Materials and methods
2.1 Materials
The following chemicals were used: adenosine, N6-cyclopentyladenosine (CPA), S-(2-hydroxy-5-nitrobenzyl)-6-thioinosine (NBTI), and dimethyl-sulfoxide (DMSO), which were purchased from Sigma-Aldrich (St. Louis, MO, United States); furthermore, quemliclustat (AB-680), polyethylene glycol with an average molecular weight of 300 (PEG 300), and polysorbate 80 (Tween 80) were obtained from MedChemExpress (Monmouth Junction, NJ, United States).
Adenosine was always dissolved in 36 °C modified Krebs–Henseleit solution (Krebs solution) containing (in mmol/L): NaCl (118), KCl (4.7), CaCl2 (2.5), NaH2PO4 (1), MgCl2 (1.2), NaHCO3 (24.9), glucose (11.5), and ascorbic acid (0.1) dissolved in redistilled water. CPA was dissolved in an ethanol: water solution (1:4 v/v, respectively). Both stock solutions were adjusted to a concentration of 10 mmol/L, and then they were further diluted with Krebs solution (when appropriate).
In the first series of experiments, AB-680 and NBTI were dissolved in DMSO, as recommended by the manufacturer of AB-680 for in vitro use (MCE, 2025).
In turn, in the second series of experiments, AB-680 and NBTI were dissolved in a buffer composed of DMSO, PEG 300, Tween 80, and normal saline (0.9% w/v of NaCl) in a ratio of 10:40:5:45 (v/v/v/v, respectively), as recommended by the manufacturer of AB-680 for in vivo use (MCE, 2025).
In the organ baths, the concentration of DMSO, PEG 300, and Tween 80 did not exceed 0.1%, 0.04%, and 0.005%, respectively (v/v), at any time.
2.2 Animals and groups
The animal use protocols were approved by the Committee of Animal Research, University of Debrecen, Hungary (11/2021/DEMÁB). Naïve, male Hartley guinea pigs with approximately 500 g and 400 g body weight (used for the first and second series of experiments, respectively) were obtained from Animalab Hungary Ltd. (Vác, Hungary), the distributor of Charles River Laboratories International Inc. (Wilmington, MA, United States). After a 2-week accommodation period, the animals were guillotined, and then the left atria were quickly removed. The isolated left atria were mounted at 10 mN resting tension in 10-mL vertical organ chambers (TSZ-04; Experimetria Ltd., Budapest, Hungary), which were filled with Krebs solution and gassed with 95% O2 and 5% CO2 (36 °C; pH = 7.4). The atria were paced by platinum electrodes (3 Hz, 1 m, twice the threshold voltage) using a programmable stimulator (ST-02; Experimetria Ltd., Budapest, Hungary) and a power amplifier (PST-02; Experimetria Ltd., Budapest, Hungary). The atrial contractile force was characterized by the amplitude of the isometric twitches, which were measured by a transducer (SD-01; Experimetria Ltd., Hungary) connected to a WS-DA-02 workstation (MDE GmbH, Heidelberg, Germany) using SPEL Advanced ISOSYS software (SOFT-02; MDE GmbH, Heidelberg, Germany). From the contractile force values, effect values were computed as a percentage decrease in the initial contractile force of the atria.
In the first series of experiments (using DMSO as the solvent), the atria were randomized into four groups: control (n = 6), AB-680 (n = 4), NBTI (n = 12), and AB-680 + NBTI (n = 7).
In the second series of experiments (using the complex buffer as the solvent), the atria were randomly divided into three groups: control (n = 6), NBTI (n = 7), and AB-680 + NBTI (n = 5).
2.3 Protocols
All atria were first allowed to equilibrate in Krebs solution for 25 min; subsequently, they were exposed to 100 μM adenosine for 2 min (as a priming), followed by a washout with Krebs solution for 20 min. Afterward, a cumulative E/c curve was constructed with adenosine on all atria, followed by another 20-min washout period (using Krebs solution). From this point, the experiments could continue in two ways.
In the first series of experiments, the control group received 10 μL DMSO for 30 min. The AB-680 group was subjected to 3 μM AB-680 (administered in 10 μL DMSO) for 30 min. The NBTI group received 10 μL DMSO for 15 min, and then (after a washout) it received 10 μM NBTI (added in 10 μL DMSO) for an additional 15 min. The AB-680 + NBTI group received 3 μM AB-680 (added in 10 μL DMSO) for 15 min, and then (after a washout) it received 3 μM AB-680 and 10 μM NBTI (co-administered in 10 μL DMSO) for an additional 15 min. Finally, a cumulative E/c curve was constructed with CPA for all the groups.
In the second series of experiments, the control group received 20 μL buffer for 30 min. The NBTI group received 20 μL buffer for 15 min, and then (after a washout) it received 10 μM NBTI (administered in 20 μL buffer) for an additional 15 min. The AB-680 + NBTI group received 3 μM AB-680 (added in 20 μL buffer) for 15 min, and then (after a washout) it received 3 μM AB-680 and 10 μM NBTI (co-administered in 20 μL buffer) for an additional 15 min. Finally, a cumulative CPA E/c curve was generated for all the groups.
2.4 Characterization of the E/c curves
The effect values were plotted against the corresponding agonist concentrations that were administered. These E/c curves were fitted to the Hill equation to illustrate them and obtain the values of the three empirical Hill parameters to describe the control E/c curves. The classical form of the Hill equation is as follows (Hill, 1910; Gesztelyi et al., 2012):
where E is the effect, c is the concentration of the agonist in the bathing medium administered for the E/c curve [due to the short half-life of adenosine in the living tissues (Pavan and IJzerman, 1998; Fredholm et al., 2001), its concentrations in the organ bath and at the binding site of the adenosine receptors can significantly differ], Emax is the maximal effect (reachable by the agonist used), EC50 is the agonist concentration producing a half-maximal effect (reciprocal potency), and n is the Hill coefficient (slope factor).
For the regression with the Hill equation, the software’s built-in equation “log(agonist) vs. response-variable slope (four parameters)” was applied, with the “bottom” parameter constrained to 0 (GraphPad, 2025).
2.5 Quantification of the distortion of the CPA E/c curves caused by NBTI
The modification of the CPA E/c curves elicited by NBTI (alone or in the simultaneous presence of AB-680) was quantified using RRM. The core of RRM is described by the following relationship (Gesztelyi et al., 2004):
where (adapted to the conditions of the present study) E’ is the distorted effect (an effect that was determined when NBTI was present), c is the CPA concentration in the bathing medium (administered for the E/c curve and, due to the relative stability of CPA, assumed to be equal to the CPA concentration at the binding site of the A1 adenosine receptors), Emax, EC50, and n are the empirical Hill parameters defining the NBTI-naïve condition (upon individual fitting, these parameters were fixed at constant values obtained by fitting the Hill equation to the averaged CPA E/c curve of the appropriate control group, whereas upon two-model global fitting, these parameters were variable), and cx is the parameter indicating the CPA concentration that is equieffective with the surplus interstitial adenosine concentration elicited by NBTI (thought to be the sole cause of the E/c curve modification compared to the control condition).
RRM was performed by implementing both individual fitting and two-model global regression (using equations equivalent with Equations 1, 2), as described by Ovari et al. (2024).
2.6 Data analysis
Curve fitting and statistical analysis were performed using GraphPad Prism version 10.5.0 (GraphPad Software Inc., La Jolla, CA, United States), while other calculations were carried out using Microsoft Excel 2024 (Microsoft Co., Redmond, WA, United States).
The distribution of data and homogeneity of variances were tested using the Shapiro–Wilk test and the Brown–Forsythe test, respectively. To compare groups with homogeneous variances and data following a Gaussian distribution, ordinary one-way ANOVA with Tukey’s post-test was carried out. Groups without homogeneous variances but with Gaussian data were compared using Brown–Forsythe ANOVA, followed by Dunnett’s post-test. To compare groups without homogeneous variances and Gaussian distribution, the Kruskal–Wallis test with Dunn’s post-test was carried out. Differences in means or medians were considered significant at p < 0.05.
The precision of the regression was characterized by the width of the 95% confidence interval (CI) of logcx (the best-fit value that is most important for us). The precision of the curve fitting was indicated by the distance of the 95% confidence bands from the corresponding best-fit curve.
To check how well the experimental data define the model, the option “Identify “ambiguous” fits” was chosen (Graphpad, 2025) because, in general, more estimates could be obtained this way.
The goodness-of-fit was quantified by the coefficient of determination (R2) and its adjusted value.
The default option for computing 95% CIs was almost always “asymmetrical.” When not (i.e., during the definition of the models to fit), the “asymmetrical” option was chosen. For every setting not addressed above, the default option was used (GraphPad, 2025).
3 Results
3.1 Initial contractile forces
The initial contractile forces, which were measured before the construction of the adenosine E/c curve (in the absence of any solvents and inhibitors), did not differ significantly among the different groups in either the first (Figure 1A) or second series of experiments (Figure 1B).
Figure 1. Initial contractile force of atria in the four [first series of experiments; panel (A)] or three [second series of experiments; panel (B)] groups (mean ± SEM) measured for the adenosine E/c curve generated in the absence of any inhibitors and their solvents. The color code used for the groups is uniform for all figures in this work. E/c, concentration–effect; Krebs, Krebs–Henseleit solution; DMSO and buffer, solvents administered subsequently (but not here); NBTI, S-(2-hydroxy-5-nitrobenzyl)-6-thioinosine; ns, statistically non-significant.
3.2 Adenosine E/c curves
The concentration-dependent, direct negative inotropic effects of adenosine showed no statistically significant difference among the groups within the first (Figure 2A) and second (Figure 2B) series of experiments. Thus, the atria used in the same series of experiments proved to form a homogenous population in terms of their response to adenosine, the endogenous agonist of the A1 adenosine receptor.
![Graphs A and B depict the negative inotropic effect (%) versus log concentration of adenosine (log[Ado]) in Krebs solution. Graph A shows data for DMSO, while graph B is for buffer. Both graphs compare control, AB-680, NBTI, and AB-680+NBTI conditions. The curves for each condition represent changes in inotropic effects, with markers and error bars indicating mean values and variability.](https://www.frontiersin.org/files/Articles/1683950/fphar-16-1683950-HTML-r1/image_m/fphar-16-1683950-g002.jpg)
Figure 2. Direct negative inotropic effect of adenosine in the isolated guinea pig left atrium in the absence of any chemicals affecting the adenosinergic homeostasis (the names of the groups only refer to a subsequent exposure to the chemical(s) indicated). Panels (A,B) apply to the first and second series of experiments, respectively. The x-axis shows the common logarithm of the molar concentration of adenosine (used for the E/c curve), and the y-axis denotes the effect (as a percentage decrease in the initial contractile force). The symbols indicate the responses to adenosine averaged within the groups (±SEM). The continuous lines show the fitted Hill equation. The color code used for the groups is uniform for all figures in this work. E/c, concentration–effect; Ado, adenosine; Krebs, Krebs–Henseleit solution; DMSO and buffer, solvents administered subsequently (but not here); NBTI, S-(2-hydroxy-5-nitrobenzyl)-6-thioinosine.
3.3 Effect of AB-680 and NBTI, alone and together, on the CPA E/c curves
3.3.1 First series of experiments (carried out in the presence of DMSO, which is recommended for in vitro use as a solvent)
CPA also exerted concentration-dependent, direct negative inotropic effects in all groups. NBTI, as expected, significantly reduced the maximum of the CPA E/c curve. In contrast, AB-680, when administered alone, did not significantly affect the response to CPA. However, AB-680 co-administered with NBTI considerably reversed the suppressive action of NBTI on the CPA E/c curve (Figure 3A). In a direct comparison (AB-680 + NBTI vs. NBTI), this effect proved not to be statistically significant, but when using an indirect approach (comparing the levels of statistical significance for control vs. AB-680 + NBTI and control vs. NBTI), it appeared meaningful, although moderate (Figure 3A).
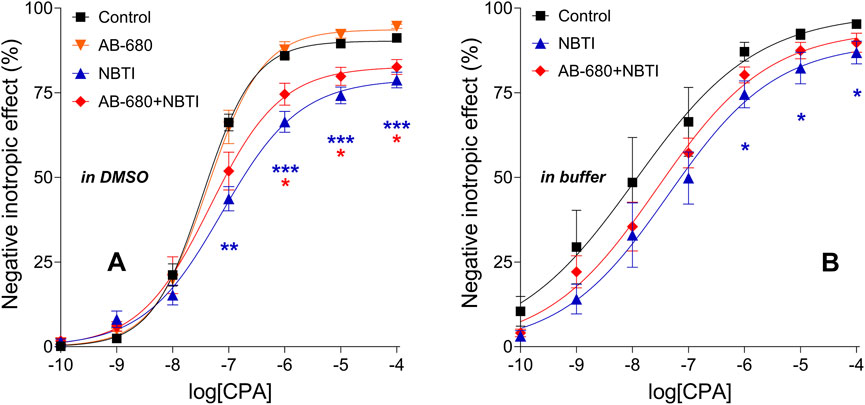
Figure 3. Direct negative inotropic effect of CPA in the isolated guinea pig left atrium in the absence and presence of 3 μM AB-680 and 10 μM NBTI (in all combinations). Panels (A,B) refer to the first (using DMSO as a solvent) and second (using a buffer as a solvent) series of experiments, respectively. The x-axis shows the common logarithm of the molar concentration of CPA (used for the E/c curve), while the y-axis denotes the effect (as a percentage decrease in the initial contractile force). The symbols indicate the responses to CPA averaged within the groups (±SEM). The continuous lines show the fitted Hill equation. The color code used for the groups is uniform for all figures in this work. E/c, concentration–effect; CPA, N6-cyclopentyladenosine; DMSO, dimethyl-sulfoxide (recommended to dissolve AB-680 when used for in vitro experiments); buffer, a mixture of chemicals presented in the 2.1 subsection (recommended to dissolve AB-680 when used for in vivo experiments); NBTI, S-(2-hydroxy-5-nitrobenzyl)-6-thioinosine; blue asterisks, comparison between control and NBTI groups; red asterisks, comparison between control and AB-680 + NBTI groups; one asterisk, p < 0.05; two asterisks, p < 0.01; three asterisks, p < 0.001.
The Emax values determined from the CPA E/c curves of the treated groups of the first series of experiments significantly differed from the control Emax. Although the maximal effect increased in the AB-680 group, it decreased in the NBTI and AB-680 + NBTI groups. Regarding the logEC50 values, the differences did not reach the level of statistical significance. Additionally, the Hill coefficient (n) in the NBTI group was significantly lower than the control value (Figures 4, A–C).
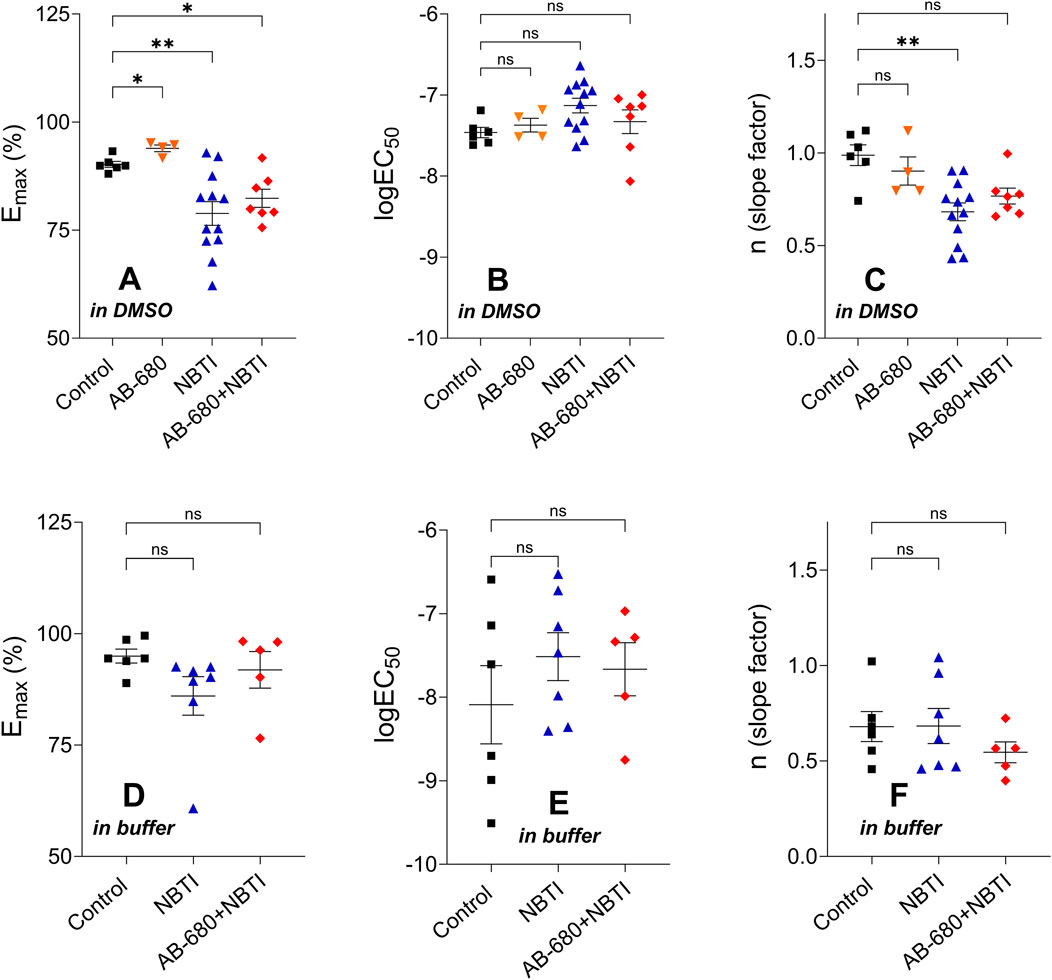
Figure 4. Hill parameters (Emax, logEC50, and n) of the CPA E/c curves in the four [first series of experiments; panels (A–C)] or three [second series of experiments; panels (D–F)] groups (mean ± SEM). The color code used for the groups is uniform for all figures in this work. In terms of the corresponding Hill parameters, the treated groups were compared to the appropriate control group. E/c, concentration–effect; CPA, N6-cyclopentyladenosine; DMSO, dimethyl-sulfoxide (recommended to dissolve AB-680 when used for in vitro experiments); buffer, a mixture of chemicals presented in the Subsection 2.1 (recommended to dissolve AB-680 when used for in vivo experiments); NBTI, S-(2-hydroxy-5-nitrobenzyl)-6-thioinosine; one asterisk, p < 0.05; two asterisks, p < 0.01; ns, statistically non-significant.
3.3.2 Second series of experiments (performed in the presence of a buffer, which is recommended for in vivo use as a solvent)
When the buffer was used to dissolve NBTI and AB-680, the effect of NBTI on the CPA E/c curve, both alone and in combination with AB-680, showed the same pattern as when DMSO was applied (Figure 3B). However, in the presence of the buffer, the scatter of the data was larger, the CPA E/c curves were closer to each other, and even the shape of the CPA E/c curves was somewhat different compared to that of the case when DMSO was present as a solvent (cf. Figures 3A, B). Statistically, only the control and NBTI groups differed significantly from each other (Figure 3B), but it was to a lesser extent than in the presence of DMSO (Figure 3A).
The corresponding Hill parameters of the CPA E/c curves did not show any significant differences among the groups of the second series of experiments (Figures 4, D–F).
3.4 Influence of AB-680 on the surplus interstitial adenosine induced by NBTI
In the first series of experiments (using DMSO), cx, the CPA concentration equieffective with the surplus endogenous adenosine concentration produced by NBTI, was approximately 2.2-fold and 2.5-fold higher in the NBTI group than in the AB-680 + NBTI group when assessed using RRM implemented with individual fitting and two-model global fitting, respectively (Table 1). In turn, the cx values determined with individual fitting were approximately 2-fold higher than the corresponding cx values obtained with two-model global fitting (Table 1) (we made sure that when comparing cx values, they only differed in one aspect).
In contrast, in the second set of experiments (using the buffer), cx was approximately 6.7-fold and 6.3-fold higher in the NBTI group than in the AB-680 + NBTI group when obtained with individual fitting and two-model global fitting, respectively (Table 1). Nevertheless, the cx values determined with individual fitting were approximately 2.5-fold higher than the corresponding cx values obtained with two-model global fitting, which was similar to the outcome observed in the first series of experiments (Table 1). However, when comparing the corresponding cx values between the two series of experiments, approximately 40–140-fold (!) higher cx values were obtained when DMSO was used as a solvent (Table 1).
Based on the 95% confidence bands indicating the precision of the curve fitting, the outcome of RRM, performed with both types of regression, appeared to be considerably more reliable when DMSO (and not the buffer) was used as a solvent. In the case of DMSO, the 95% confidence bands were clearly separated from each other, especially in the middle and terminal portions (individual fitting) or in the middle section (two-model global regression) of the CPA E/c curves (Figures 5A,B). Contrary to this, for the buffer, the 95% confidence bands largely overlapped (Figures 5C,D). Consequently, DMSO, a solvent widely used for in vitro investigations, interfered with the determination of cx to a lesser extent than the buffer recommended as a solvent for in vivo studies (MCE, 2025).
![Graphs A and B in DMSO and C and D in buffer show the negative inotropic effect versus log[CPA]. Each graph compares NBTI alone and with AB-680, using individual and two-model global fitting. Red diamonds, blue triangles, and black squares represent different conditions, with curves indicating model fits. Error bars are included for each data point.](https://www.frontiersin.org/files/Articles/1683950/fphar-16-1683950-HTML-r1/image_m/fphar-16-1683950-g005.jpg)
Figure 5. CPA E/c curves of the first [panels (A,B)] and second [panels (C,D)] series of experiments (except the AB-680 group) evaluated with RRM to obtain the CPA-equivalents of the surplus interstitial adenosine concentration caused by NBTI (i.e., cx values). The x-axis shows the common logarithm of the molar concentration of CPA, and the y-axis denotes the effect. The symbols indicate the responses to CPA averaged within the groups (±SEM). The continuous lines denote the model of RRM fitted individually [panels (A,C)] or globally using two equations [panels (B,D)]. The dotted lines indicate the 95% confidence bands. The color code used for the groups is uniform for all figures in this work. E/c, concentration–effect; CPA, N6-cyclopentyladenosine; DMSO, dimethyl-sulfoxide (recommended to dissolve AB-680 when used for in vitro experiments); buffer, a mixture of chemicals presented in the Subsection 2.1 (recommended to dissolve AB-680 when used for in vivo investigations); NBTI, S-(2-hydroxy-5-nitrobenzyl)-6-thioinosine.
In summary, in our ex vivo experimental setup, following the dissolution protocol of AB-680 recommended for in vitro use (MCE, 2025), AB-680 reduced cx, a measure of the interstitial adenosine accumulation caused by NBTI, by at least half. This outcome suggests that in response to AB-680, the concentration of the surplus interstitial adenosine accumulated by NBTI may also have been halved.
4 Discussion
In the present study, we found that AB-680, a highly potent CD73 inhibitor, did not affect the atrial direct negative inotropic response to CPA, a selective A1 adenosine receptor full agonist, when administered alone. In turn, when added together with NBTI, a nucleoside transport blocker, AB-680 was able to partially reverse the effect of NBTI on the response to CPA. Nevertheless, the inhibitory action of AB-680 on the effect of NBTI appeared to be less than that of PSB-12379, a less potent CD73 inhibitor, which was investigated earlier in the same experimental model (Viczjan et al., 2021). We also found that to dissolve AB-680 for our ex vivo investigation, the protocol recommended for in vitro use was preferable to that recommended for in vivo use (i.e., the simple DMSO should be preferred over the complex buffer). In addition, AB-680, co-administered with NBTI, reduced cx, the CPA-equivalent of the surplus interstitial adenosine concentration, by at least half (when DMSO was used as a solvent).
Adenosine exerts a variety of protective and regenerative effects in multicellular organisms (Fredholm et al., 2001; Fredholm et al., 2011; Burnstock et al., 2010; IJzerman et al., 2022). In the mammalian myocardium, all four adenosine receptor (sub)types are expressed and may influence cardiac function, including contractility (Chandrasekera et al., 2010). Nevertheless, most of the direct (proximate) effects of adenosine on the myocardium are mediated by A1 and (to a lesser extent) A3 adenosine receptors (Fredholm et al., 2001; Szentmiklósi et al., 2011; Headrick et al., 2013; Qian et al., 2025). The significance of the A2 subtypes has been highlighted in relation to atrial fibrillation (Maille et al., 2021).
The most striking acute cardiac actions of adenosine are the negative tropic effects, including negative inotropy (Headrick et al., 2013; Burnstock and Pelleg, 2015; Qian et al., 2025). Due to the extracellular location of the binding site of adenosine receptors, the interstitial level of adenosine plays a prominent role in the adenosinergic mechanisms in all organs, including the heart (Szentmiklósi et al., 2011; Headrick et al., 2013; Burnstock and Pelleg, 2015; Qian et al., 2025). Consequently, any agent influencing the interstitial adenosine levels should also be examined from this perspective.
The negative inotropic effect is particularly strong in the supraventricular myocardium (compared to the ventricular one), where, in most species, including the guinea pig and human, adenosine can significantly reduce the contractile force in a direct manner, thus without any previous positive inotropic stimulation (Belardinelli et al., 1995; Fredholm et al., 2001). The significance of atrial contractility stems from the fact that its decrease worsens the atrial booster pump function and, thereby, the ventricular filling (Rossi et al., 2000; Hoit, 2014). Moreover, the reduced atrial pumping capacity enhances the risk of atrial thrombus formation (Beigel et al., 2014; Yilmaz et al., 2020). Thus, it is important to consider atrial contractility (in addition to ventricular contractility) when investigating drugs or drug candidates that can (potentially or in a proven manner) influence the cardiac adenosinergic system.
In a previous study, we found that PSB-12379, a potent inhibitor of CD73, a key enzyme of the extracellular adenosine formation (Bhattarai et al., 2015; Schmies et al., 2020), significantly blunted the suppressing effect of NBTI on the response to CPA in an ex vivo guinea pig left atrium model (Viczjan et al., 2021). The transformation of the CPA E/c curve caused by NBTI (a decrease in the maximal effect and potency) was ascribed to the elevating effect of NBTI on the interstitial adenosine level (Gesztelyi et al., 2003; Karsai et al., 2006; 2007; Erdei et al., 2018; Viczjan et al., 2021). In the present work, we obtained a similar result using AB-680, an extremely potent CD73 inhibitor (Bowman et al., 2019), in the same experimental model (Figure 3). Nevertheless, there is an interesting difference between the previous and present results; i.e., the influence of PSB-12379 on the NBTI’s effect appears to be stronger than that of AB-680 (cf. Figure 6, left panel in Viczjan et al., 2021 with Figure 3A in this work). This difference suggests that, at least in our model, AB-680 affects the myocardial adenosinergic mechanisms to a lesser extent than PSB-12379, another CD73 inhibitor, does. Greater potency (cf. the picomolar affinity of AB-680 (Bowman et al., 2019) and the nanomolar affinity of PSB-12379 (Bhattarai et al., 2015) for CD73) does not necessarily mean greater efficacy (maximal effect) (Brunton and Knollmann, 2023). Regarding the potential cardiac side-effects of AB-680, this result is more favorable than undesirable.
It should also be addressed that AB-680, similarly to PSB-12379 (Viczjan et al., 2021), did not significantly affect the CPA E/c curve when administered alone (Figure 3A). This may be because under our experimental circumstances, the naïve interstitial adenosine level did not appear to be capable of activating the atrial A1 adenosine receptors (Pak et al., 2015). Hence, it is not surprising that a decrease in an already ineffective adenosine level could not influence the response to CPA (Figure 3A).
AB-680 was reported to have a slow onset regarding the inhibition of CD73. Increasing the pre-incubation time of human CD73 with AB-680 resulted in decreased IC50 values, with the plateau reached at approximately 30 min (Bowman et al., 2019). The 30-min incubation period used in the present investigation addressed this previous observation.
Regarding cx, the CPA concentration equieffective with an increase in the atrial interstitial adenosine concentration caused by NBTI, AB-680 reduced its value by at least half when added together with NBTI (and DMSO was used as a solvent) (Table 1). The cx values provided by RRM are unique in that they characterize the increase in the adenosine concentration in the microenvironment of the binding sites of myocardial A1 adenosine receptors, which is a tissue compartment that is otherwise difficult to assess in this regard (Karsai et al., 2006; 2007; Erdei et al., 2018). In light of our previous results, the current cx values (determined for NBTI alone, using DMSO), 52.65 nM and 27.39 nM (obtained with individual fitting and two-model global fitting, respectively; Table 1), can be considered average (Erdei et al., 2018).
In addition, DMSO, recommended as a solvent for AB-680 under in vitro conditions (MCE, 2025), interfered with our ex vivo measurements to a lesser extent than the buffer recommended to dissolve AB-680 in in vivo studies (MCE, 2025) (cf. Figure 3A with Figure 3B; Table 1). Therefore, for future ex vivo studies, we suggest using DMSO for dissolving AB-680 (or chemicals with a similar solubility profile).
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors, without undue reservation.
Ethics statement
The animal study was approved by Committee of Animal Research, University of Debrecen, Hungary (11/2021/DEMÁB). The study was conducted in accordance with the local legislation and institutional requirements.
Author contributions
IO: Writing – original draft, Investigation. ABB: Writing – original draft, Investigation. AL: Funding acquisition, Writing – review and editing. EB: Writing – review and editing, Funding acquisition. TE: Writing – review and editing, Resources, Data curation. OO: Data curation, Resources, Writing – review and editing. BH: Resources, Data curation, Writing – review and editing. BJ: Resources, Writing – review and editing, Data curation. ZS: Writing – review and editing, Funding acquisition. JZ: Data curation, Resources, Writing – review and editing. GV: Conceptualization, Writing – review and editing, Formal analysis. RG: Formal analysis, Conceptualization, Writing – review and editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. This research was supported by the European Union and the State of Hungary under the grant number TKP2021-EGA-19 (project no. TKP2021-EGA-19 has been implemented with the support provided by the Ministry of Culture and Innovation of Hungary from the National Research, Development, and Innovation Fund financed under the TKP2021-EGA funding scheme). This study was also supported by the University Research Scholarship Program of the Ministry for Culture and Innovation from the source of the National Research, Development, and Innovation fund/providing personal support for IO (EKÖP-24-4-I-DE-279). Furthermore, this research was supported by the University of Debrecen Program for Scientific Publication.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
Beigel, R., Wunderlich, N. C., Ho, S. Y., Arsanjani, R., and Siegel, R. J. (2014). The left atrial appendage: anatomy, function, and noninvasive evaluation. JACC Cardiovasc. Imaging 7, 1251–1265. doi:10.1016/j.jcmg.2014.08.009
Belardinelli, L., Shryock, J. C., Song, Y., Wang, D., and Srinivas, M. (1995). Ionic basis of the electrophysiological actions of adenosine on cardiomyocytes. FASEB J. 9, 359–365. doi:10.1096/fasebj.9.5.7896004
Bhattarai, S., Freundlieb, M., Pippel, J., Meyer, A., Abdelrahman, A., Fiene, A., et al. (2015). α,β-Methylene-ADP (AOPCP) derivatives and analogues: development of potent and selective ecto-5'-Nucleotidase (CD73) inhibitors. J. Med. Chem. 58, 6248–6263. doi:10.1021/acs.jmedchem.5b00802
Bowman, C. E., da Silva, R. G., Pham, A., and Young, S. W. (2019). An exceptionally potent inhibitor of human CD73. Biochemistry 58, 3331–3334. doi:10.1021/acs.biochem.9b00448
Brunton, L. L., and Knollmann, B. C. (2023). Goodman and Gilman’s the pharmacological basis of therapeutics. (McGraw Hill, LLC), 52–53.
Burnstock, G., and Pelleg, A. (2015). Cardiac purinergic signalling in health and disease. Purinergic Signal 11, 1–46. doi:10.1007/s11302-014-9436-1
Burnstock, G., Fredholm, B. B., North, R. A., and Verkhratsky, A. (2010). The birth and postnatal development of purinergic signalling. Acta Physiol. (Oxf.). 199, 93–147. doi:10.1111/j.1748-1716.2010.02114.x
Chandrasekera, P. C., McIntosh, V. J., Cao, F. X., and Lasley, R. D. (2010). Differential effects of adenosine A2a and A2b receptors on cardiac contractility. Am. J. Physiol. Heart Circ. Physiol. 299, H2082–H2089. doi:10.1152/ajpheart.00511.2010
Deussen, A. (2000a). Metabolic flux rates of adenosine in the heart. Naunyn Schmiedeb. Arch. Pharmacol. 362, 351–363. doi:10.1007/s002100000318
Deussen, A. (2000b). Quantitative integration of different sites of adenosine metabolism in the heart. Ann. Biomed. Eng. 28, 877–883. doi:10.1114/1.1308498
Deussen, A., Stappert, M., Schäfer, S., and Kelm, M. (1999). Quantification of extracellular and intracellular adenosine production: understanding the transmembranous concentration gradient. Circulation 99, 2041–2047. doi:10.1161/01.CIR.99.15.2041
Deussen, A., Weichsel, J., and Pexa, A. (2006). Features of adenosine metabolism of mouse heart. Purinergic Signal 2, 663–668. doi:10.1007/s11302-006-9013-3
Erdei, T., Szabo, A. M., Lampe, N., Szabo, K., Kiss, R., Zsuga, J., et al. (2018). FSCPX, a chemical widely used as an irreversible A1 adenosine receptor antagonist, modifies the effect of NBTI, a nucleoside transport inhibitor, by reducing the interstitial adenosine level in the Guinea pig atrium. Molecules 23, 2186. doi:10.3390/molecules23092186
Fredholm, B. B., Ijzerman, A. P., Jacobson, K. A., Klotz, K. N., and Linden, J. (2001). International union of pharmacology. XXV. Nomenclature and classification of adenosine receptors. Pharmacol. Rev. 53, 527–552. doi:10.1016/s0031-6997(24)01511-4
Fredholm, B. B., Ijzerman, A. P., Jacobson, K. A., Linden, J., and Müller, C. E. (2011). International union of Basic and Clinical Pharmacology. LXXXI. Nomenclature and classification of adenosine receptors–an update. Pharmacol. Rev. 63, 1–34. doi:10.1124/pr.110.003285
Gao, P. P., Jiang, H. F., Du, Y. W., Shu, Y., Wan, Y. W., Yin, E. Z., et al. (2025). CD73 inhibitor AB680 suppresses glioblastoma in mice by inhibiting purine metabolism and promoting P2RY12+ microglia transformation. Acta Pharmacol. Sin. doi:10.1038/s41401-025-01585-9
Gesztelyi, R., Zsuga, J., Hajdú, P., Szabó, J. Z., Cseppento, A., and Szentmiklósi, A. J. (2003). Positive inotropic effect of the inhibition of cyclic GMP-Stimulated 3',5'-cyclic nucleotide phosphodiesterase (PDE2) on Guinea pig left atria in eu- and hyperthyroidism. Gen. Physiol. Biophys. 22, 501–513.
Gesztelyi, R., Zsuga, J., Juhász, B., Der, P., Vecsernyes, M., and Szentmiklosi, A. J. (2004). Concentration estimation via curve fitting: quantification of negative inotropic agents by using a simple mathematical method in Guinea pig atria. Bull. Math. Biol. 66, 1439–1453. doi:10.1016/j.bulm.2004.03.001
Gesztelyi, R., Zsuga, J., Kemeny-Beke, A., Varga, B., Juhász, B., and Tosaki, A. (2012). The hill equation and the origin of quantitative pharmacology. Arch. Hist. Exact. Sci. 66, 427–438. doi:10.1007/s00407-012-0098-5
Graphpad (2025). Curve fitting guide. Available online at: https://www.graphpad.com/guides/prism/latest/curve-fitting/index.htm (Accessed January, 22, 2025).
Grenczer, M., Pinter, A., Zsuga, J., Kemeny-Beke, A., Juhasz, B., Szodoray, P., et al. (2010a). The influence of affinity, efficacy, and slope factor on the estimates obtained by the receptorial responsiveness method (RRM): a computer simulation study. Can. J. Physiol. Pharmacol. 88, 1061–1073. doi:10.1139/y10-078
Grenczer, M., Zsuga, J., Majoros, L., Pinter, A., Kemeny-Beke, A., Juhasz, B., et al. (2010b). Effect of asymmetry of concentration-response curves on the results obtained by the receptorial responsiveness method (RRM): an in silico study. Can. J. Physiol. Pharmacol. 88, 1074–1083. doi:10.1139/y10-089
Headrick, J. P., Ashton, K. J., Rose'meyer, R. B., and Peart, J. N. (2013). Cardiovascular adenosine receptors: expression, actions and interactions. Pharmacol. Ther. 140, 92–111. doi:10.1016/j.pharmthera.2013.06.002
Hesse, J., Leberling, S., Boden, E., Friebe, D., Schmidt, T., Ding, Z., et al. (2017). CD73-derived adenosine and tenascin-C control cytokine production by epicardium-derived cells formed after myocardial infarction. FASEB J. 31, 3040–3053. doi:10.1096/fj.201601307R
Hill, A. V. (1910). The heat produced in contracture and muscular tone. J. Physiol. Lond. 40, 389–403. doi:10.1113/jphysiol.1910.sp001377
Hoit, B. D. (2014). Left atrial size and function: role in prognosis. J. Am. Coll. Cardiol. 63, 493–505. doi:10.1016/j.jacc.2013.10.055
Ijzerman, A. P., Jacobson, K. A., Müller, C. E., Cronstein, B. N., and Cunha, R. A. (2022). International union of basic and clinical pharmacology. CXII: adenosine receptors: a further update. Pharmacol. Rev. 74, 340–372. doi:10.1124/pharmrev.121.000445
Karsai, D., Zsuga, J., Juhasz, B., Der, P., Szentmiklosi, A. J., Tosaki, A., et al. (2006). Effect of nucleoside transport blockade on the interstitial adenosine level characterized by a novel method in Guinea pig atria. J. Cardiovasc. Pharmacol. 47, 103–109. doi:10.1097/01.fjc.0000196239.51018.a0
Karsai, D., Gesztelyi, R., Zsuga, J., Jakab, A., Szendrei, L., Juhasz, B., et al. (2007). Influence of hyperthyroidism on the effect of adenosine transport blockade assessed by a novel method in Guinea pig atria. Cell. Biochem. Biophys. 47, 45–52. doi:10.1385/cbb:47:1:45
Maille, B., Fromonot, J., Guiol, C., Marlinge, M., Baptiste, F., Lim, S., et al. (2021). A2 adenosine receptor subtypes overproduction in atria of perioperative atrial fibrillation patients undergoing cardiac surgery: a pilot study. Front. Cardiovasc. Med. 8, 761164. doi:10.3389/fcvm.2021.761164
MCE (2025). Available online at: https://www.medchemexpress.com/AB-680.html (Accessed August 2, 2025).
Ovari, I., Viczjan, G., Erdei, T., Takacs, B., Tarjanyi, V., Zsuga, J., et al. (2024). The influence of the way of regression on the results obtained by the receptorial responsiveness method (RRM), a procedure to estimate a change in the concentration of a pharmacological agonist near the receptor. Front. Pharmacol. 15, 1375955. doi:10.3389/fphar.2024.1375955
Pak, K., Zsuga, J., Kepes, Z., Erdei, T., Varga, B., Juhasz, B., et al. (2015). The effect of adenosine deaminase inhibition on the A1 adenosinergic and M2 muscarinergic control of contractility in eu- and hyperthyroid Guinea pig atria. Naunyn Schmiedeb. Arch. Pharmacol. 388, 853–868. doi:10.1007/s00210-015-1121-6
Pavan, B., and Ijzerman, A. P. (1998). Processing of adenosine receptor agonists in rat and human whole blood. Biochem. Pharmacol. 56, 1625–1632. doi:10.1016/S0006-2952(98)00270-6
Piovesan, D., Tan, J. B. L., Becker, A., Banuelos, J., Narasappa, N., DiRenzo, D., et al. (2022). Targeting CD73 with AB680 (quemliclustat), a novel and potent small-molecule CD73 inhibitor, restores immune functionality and facilitates antitumor immunity. Mol. Cancer Ther. 21, 948–959. doi:10.1158/1535-7163.MCT-21-0802
Qian, Y., Zheng, Y., Leng, L., Liu, Q., Tian, X., Chen, S., et al. (2025). Adenosine and adenosine receptors: a double-edged sword in cardiovascular system. Front. Pharmacol. 16, 1538680. doi:10.3389/fphar.2025.1538680
Rossi, A., Zardini, P., and Marino, P. (2000). Modulation of left atrial function by ventricular filling impairment. Heart Fail. Rev. 5, 325–331. doi:10.1023/a:1026507128973
Schmies, C. C., Rolshoven, G., Idris, R. M., Losenkova, K., Renn, C., Schäkel, L., et al. (2020). Fluorescent probes for Ecto-5'-nucleotidase (CD73). ACS Med. Chem. Lett. 11, 2253–2260. doi:10.1021/acsmedchemlett.0c00391
Schreieck, J., and Richardt, G. (1999). Endogenous adenosine reduces the occurrence of ischemia-induced ventricular fibrillation in rat heart. J. Mol. Cell. Cardiol. 31, 123–134. doi:10.1006/jmcc.1998.0858
Strickland, L. N., Liu, W., Hussein, U., Mardik, N., Chen, X., Mills, T., et al. (2024). Preventive treatment with a CD73 small molecule inhibitor enhances immune surveillance in K-Ras mutant pancreatic intraepithelial neoplasia. Cancer Prev. Res. (Phila.) 17, 457–470. doi:10.1158/1940-6207.CAPR-24-0200
Szentmiklósi, A. J., Cseppento, A., Harmati, G., and Nánási, P. P. (2011). Novel trends in the treatment of cardiovascular disorders: Site- and event-selective adenosinergic drugs. Curr. Med. Chem. 18, 1164–1187. doi:10.2174/092986711795029753
Viczjan, G., Erdei, T., Ovari, I., Lampe, N., Szekeres, R., Bombicz, M., et al. (2021). A body of circumstantial evidence for the irreversible ectonucleotidase inhibitory action of FSCPX, an agent known as a selective irreversible A1 adenosine receptor antagonist so far. Int. J. Mol. Sci. 22, 9831. doi:10.3390/ijms22189831
Yilmaz, K. C., Akgun, A. N., Ciftci, O., Eroglu, S., Pirat, B., Sade, E., et al. (2020). Risk factors for left atrial appendage thrombus. Acta Cardiol. 75, 355–359. doi:10.1080/00015385.2020.1757852
Keywords: AB-680, CD73 inhibitor, nucleoside transport blocker, Guinea pig, atrium, A1 adenosine receptor, interstitial adenosine level
Citation: Ovari I, Brezniczky AB, Laczovics A, Berényi E, Erdei T, Ojo O, Hornok B, Juhasz B, Szilvassy Z, Zsuga J, Viczjan G and Gesztelyi R (2025) The interaction of AB-680, a CD73 inhibitor, with NBTI, a nucleoside transporter inhibitor, on the adenosinergic control of atrial contractility. Front. Pharmacol. 16:1683950. doi: 10.3389/fphar.2025.1683950
Received: 11 August 2025; Accepted: 30 September 2025;
Published: 21 October 2025.
Edited by:
Carlos Alan Dias-Junior, São Paulo State University, BrazilReviewed by:
Andrei Adrian Tica, University of Medicine and Pharmacy of Craiova, RomaniaDur E Shahwar, University of São Paulo, Brazil
Copyright © 2025 Ovari, Brezniczky, Laczovics, Berényi, Erdei, Ojo, Hornok, Juhasz, Szilvassy, Zsuga, Viczjan and Gesztelyi. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Rudolf Gesztelyi, Z2VzenRlbHlpLnJ1ZG9sZkBwaGFybS51bmlkZWIuaHU=
†These authors have contributed equally to this work