 Rafael Batista João1
Rafael Batista João1 João Paulo Rocha Pereira Toiansk de Azevedo1Dácio Almeida Pereira1Paulo César Ragazzo1Paulo Maurício de Oliveira1,2,3*
João Paulo Rocha Pereira Toiansk de Azevedo1Dácio Almeida Pereira1Paulo César Ragazzo1Paulo Maurício de Oliveira1,2,3*- 1Neurophysiology Department, Goiânia Neurological Institute, Goiânia, Goiás, Brazil
- 2Psychiatry Department, Goiânia Neurological Institute, Goiânia, Goiás, Brazil
- 3Psychiatry Department, Federal University of Goiás, Goiânia, Goiás, Brazil
Background: Electroconvulsive therapy (ECT) remains an effective intervention for severe and treatment-resistant mood disorders, particularly major depressive disorder (MDD) and bipolar disorder (BD). While traditionally linked to neurotransmitter modulation, recent research suggests that ECT exerts broader biological effects. Currently, there is a necessity for identifying factors that could support a more accurate selection of individuals, predict their therapeutic response, and help investigate evidence of possible neuroplastic effects of this technique. In this setting, many studies have been published in the last few years, aiming to identify potential biomarkers by understanding immune-inflammatory, structural, and cellular mechanisms and their correlations with clinical outcomes post-ECT.
Methods: We searched PubMed, Embase, Scopus, Web of Science, PsycINFO, and Cochrane Library for studies published between 2020 and 2025. Studies were selected based on their relevance to inflammatory, immune, structural, and cellular mechanisms associated with ECT.
Results: Twenty-six studies were included. The main results reported post-ECT reductions in inflammatory markers, including C-reactive protein, interleukin-6, and tumor necrosis factor-alpha, or suggested a biphasic trajectory, with an initial transient immune activation preceding inflammatory partial resolution. Noteworthy differences were related to age, as younger patients showed more favorable immune adaptability in comparison with older individuals, who demonstrated elongated inflammatory activity. Neuroplastic changes following ECT were observed, including increased hippocampal neurogenesis, enhanced brain-derived neurotrophic factor expression, and structural changes in neuroimaging studies. Novel exploratory research on post-mortem analyses further confirmed the upregulation of neuroplasticity markers without evidence of sustained neuroinflammation. In addition, epigenetic mechanisms, particularly microRNA modulation following ECT, may induce long-term cellular reprogramming, potentially influencing treatment response. Moreover, one recent study suggested that elevated baseline levels of miR-223-3p may be a predictor of ECT response among treatment-resistant depression patients. Finally, a recent study exploring mitochondrial adaptations found that the interactions between mitochondrial DNA copy number, oxidative stress, and ECT remain inconclusive.
Conclusion: Recent studies have expanded the understanding of ECT’s neuroinflammation effects and beyond, adding data on its interactions with immune, neuroplastic, and genetic mechanisms in human samples. Although many gaps still exist, these findings pave the way for further research that may improve outcomes of treatment-resistant mood disorders.
Introduction
Electroconvulsive therapy (ECT) remains one of the most effective interventions for severe and treatment-resistant mood disorders, such as major depressive disorder (MDD) and bipolar disorder (BD) (1, 2). Meta-analytic data show that ECT achieves response and remission rates of 74.2% and 52.3% in MDD and 77.1% and 52.3% BD, respectively (3). While its clinical efficacy has been widely documented, the precise neurobiological mechanisms underlying ECT’s therapeutic effects are still under investigation (3, 4). ECT is one of the least understood biological treatments in psychiatry. Historically, ECT was found to act primarily through the modulation of neurotransmitter systems, particularly serotonin, dopamine, and gamma-aminobutyric acid (GABA) (4, 5). However, evidence suggests that its effects extend beyond synaptic neurotransmission, involving immune-inflammatory pathways, neuroplasticity, and genetic/epigenetic modifications (4, 6–8).
Neuroinflammation has been increasingly implicated in mood disorders, with elevated pro-inflammatory cytokines such as interleukin-6 (IL-6), tumor necrosis factor-alpha (TNF-α), and C-reactive protein (CRP) contributing to neuronal dysfunction and treatment resistance (9–11). ECT appears to modulate immune responses. However, research findings on this topic have been inconsistent, and many studies continue to explore how ECT influences inflammatory markers and clinical outcomes (12). Concomitantly, growing evidence suggests that ECT may support brain plasticity by increasing hippocampal volume, reshaping neural connections, and enhancing brain-derived neurotrophic factor (BDNF) expression (13). Recently, genetic and epigenetic changes, including DNA methylation shifts and altered microRNA expression, have been implicated in neural adaptation due to ECT modulation effects (8). Mitochondrial function has also been investigated in this setting, with studies linking ECT response to oxidative stress markers and mitochondrial DNA copy number (mtDNAcn) (12).
Understanding how ECT modulates immune-inflammatory pathways, neuroplasticity, and epigenetic processes is paramount to identifying biomarkers of response, improve patient selection, and guide the development of optimized neuromodulatory interventions. Despite recent advances, critical knowledge gaps persist. Thus, we performed this overview of recent findings of studies integrating ECT’s inflammatory, immune, neuroplastic, and genetic effects among individuals with mood disorders.
Methods
We searched PubMed, Embase, Scopus, Web of Science, PsycINFO, and Cochrane Library databases to identify original studies published between January 2020 and April 2025 that investigated the effects of ECT on inflammatory, immune, genetic, and neuroplastic processes in mood disorders. The search strategy included terms and keywords related to “electroconvulsive therapy,” “inflammation,” “immune response,” “neuroplasticity,” “genetics,” “epigenetics,” “mood disorders”, “depression”, “depressive”, and “bipolar disorder”. In addition, we manually reviewed the references of all included articles and previous relevant reviews for additional studies.
Studies with human samples and correlating inflammatory parameters, molecular profiling, neuroimaging, post-mortem histopathology, or genetic markers were included. Eligible studies included participants diagnosed with major depressive disorder (MDD) or bipolar disorder (BD), as defined by standardized diagnostic criteria (e.g., DSM-IV, DSM-5, or ICD-10/11) (14–16). Biological measures encompassed inflammatory cytokines (e.g., IL-6, TNF-α, IL-1β, IL-10, CRP), hematological inflammatory indexes (e.g., NLR, PLR, SII), neurotrophic factors (e.g., BDNF), DNA methylation changes, microRNA expression, mtDNAcn, and post-mortem markers of neuroplasticity (e.g., DCX, STMN1) and glial activation (e.g., GFAP, Iba1), as well as structural neuroimaging biomarkers.
No restrictions regarding study design were applied; both observational and interventional studies were eligible. Preprint publications were excluded. Data extraction focused on sample characteristics, study design, primary findings, and clinical correlations. Key details of ECT protocols — including electrode placement, dosing strategies, pulse width, frequency, and total number of sessions — are summarized for each study in Tables 1–3.
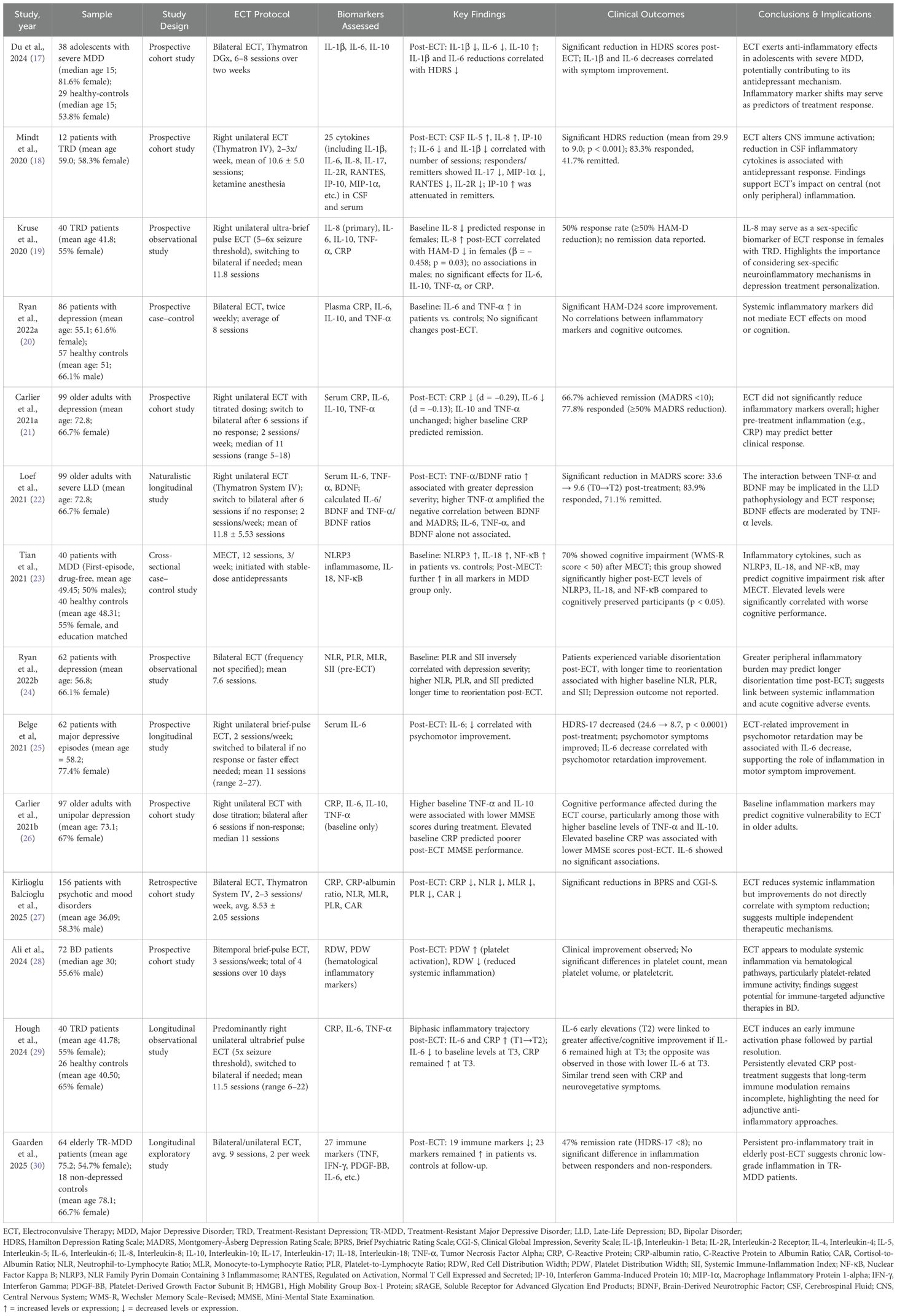
Table 1. Main features of studies evaluating peripheral inflammatory and hematological biomarkers in the context of ECT response, cognitive adverse events, and symptom clusters.

Table 2. Main features of studies evaluating neuroplasticity, neuroimmune processes, and brain changes related to ECT in mood disorders.
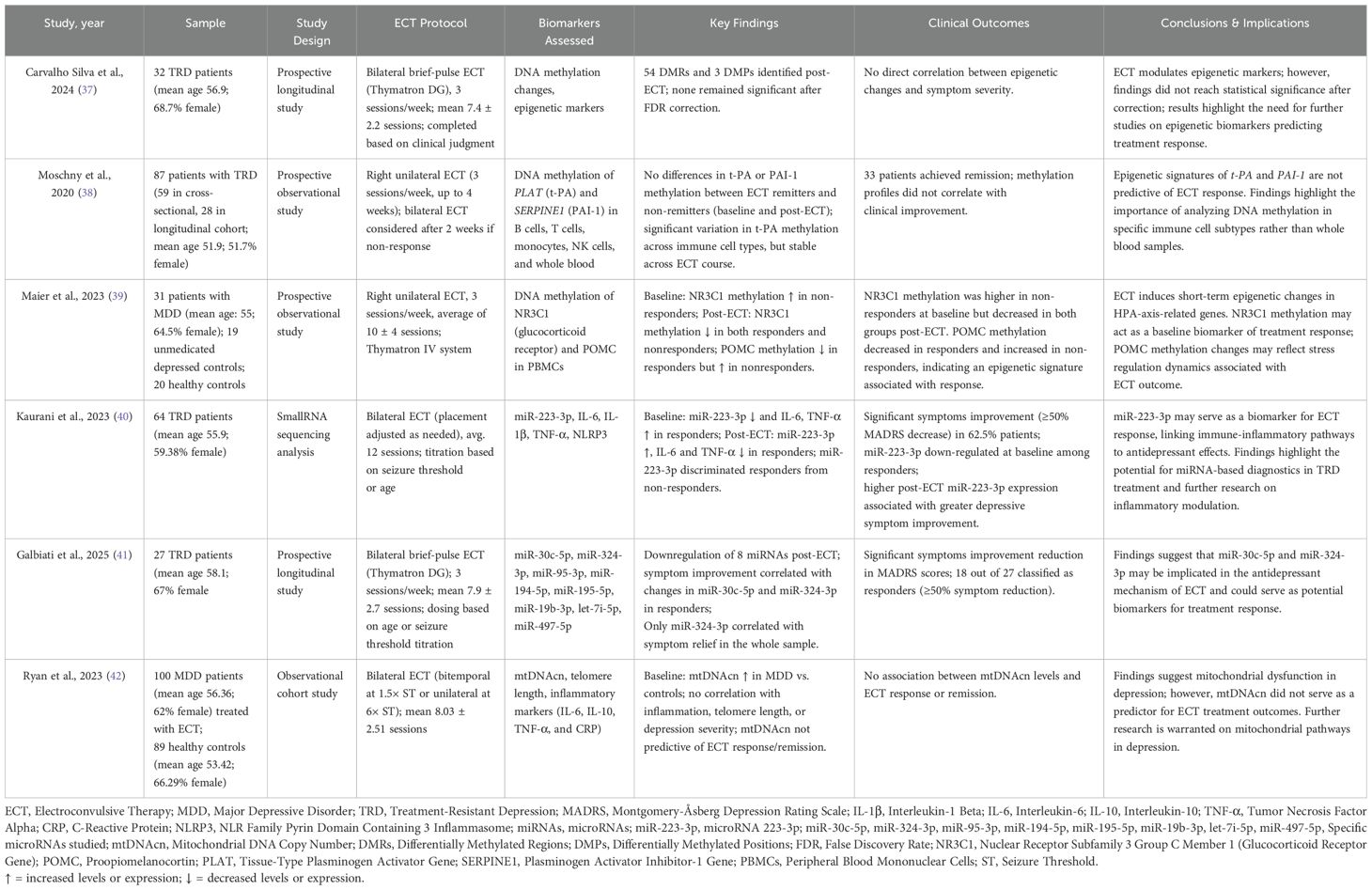
Table 3. Main features of studies on genetic, epigenetic, microRNA, and mitochondrial biomarkers after ECT in mood disorders.
Results
A total of 26 studies published between 2020 and 2025 were included, investigating the effects of ECT on immune, inflammatory, and cellular processes in patients with mood disorders (e.g., MDD and BD). Sample sizes varied widely, ranging from small pilot studies to large cohorts. The methodologies included longitudinal biomarker investigations, neuroimaging, molecular analyses, post-mortem histopathology, DNA/small RNA sequencing, and mtDNAcn assessments.
Inflammatory markers and antidepressant response
Studies focused on the impact of ECT on peripheral inflammatory markers, reporting reductions in systemic inflammation following treatment. In a prospective cohort study, Du et al. (2024) analyzed inflammatory biomarkers and depressive symptoms in 38 adolescents with severe MDD (median age [interquartile range]: 15 [14–16]; 81.6% female) compared to 29 healthy-controls (median age [interquartile range]: 15 [14.5–16.5]; 53.8% female). Baseline blood samples (e.g., interleukin-1 beta [IL-1β], interleukin-6 [IL-6], and interleukin-10 [IL-10]) were collected before treatment and after completing 6–8 ECT sessions spanning two weeks. The outcomes showed significant post-ECT decreases in IL-1β and IL-6, and an increase in IL-10. Interestingly, improvements in depressive symptoms (as assessed by the 17-item Hamilton Depression Rating Scale scores [HDRS]) were correlated with reductions in IL-1β (r = -0.343, p = 0.035) and in IL-6 (r = -0.403, p = 0.012), suggesting a link between anti-inflammatory and antidepressant effects of ECT in this population (17).
Mindt et al. (2020) examined CSF and plasma levels of 25 cytokines in 12 TRD patients (mean ± SD age: 59 ± 21.9; 58.3% female) undergoing ECT, focusing on central immune modulation. CSF levels of IL-6 and IL-1β showed negative correlations with the number of ECT sessions, suggesting progressive reductions over the treatment course. Moreover, patients who achieved remission exhibited reductions in CSF IL-17, MIP-1α, RANTES, and IL-2R, while IP-10 levels increased to a lesser extent compared to non-remitters. Such findings indicate that ECT may modulate central immune processes beyond peripheral inflammation (18). In addition, Kruse et al. (2020) analyzed serum levels of IL-6 and IL-8 in 40 patients with major depression (mean age ± SD: 41.8 ± 13.7; 55% female) treated with ECT. Their findings revealed sex-specific differences in inflammatory dynamics, with increases in IL-8 associated with symptom improvement in women but not in men. These results suggest that IL-8 may serve as a sex-dependent biomarker for ECT response and support further investigations of gender-specific immune signatures (19). In contrast, Ryan et al. (2022a) found no significant post-treatment changes in a panel of classic inflammatory cytokines. The authors examined plasma concentrations of CRP, IL-6, IL-10, TNF-α, and IL-1β in 86 patients with MDD (mean age ± SD: 55.1 ± 14.65; 61.63% female) and 57 healthy controls (mean age ± SD: 51 ± 12.7; 66.1% male). Although IL-6 and TNF-α were elevated at baseline in depressed individuals, these levels remained unchanged after ECT; no associations were found between cytokine concentrations and clinical or cognitive outcomes. IL-1β was undetectable in most samples. Taken together with other studies, these results illustrate the heterogeneity in ECT’s effects on systemic inflammatory markers and depressive symptoms (20).
Inflammation and treatment response in late-life depression
Studies have specifically investigated the immunoinflammatory response to ECT in older adults, aiming to clarify its relationship with treatment efficacy in late-life depression (LLD). Carlier et al. (2021) conducted a prospective cohort study evaluating the inflammatory response to ECT in 99 older adults with depression (mean age ± SD: 72.8 ± 8.3; 66.7% female). Patients received right unilateral ECT with dose titration; non-responders were switched to bilateral ECT after six sessions. Across a median of 11 sessions, small reductions in CRP and IL-6 were observed, although these changes did not reach statistical significance after Bonferroni correction. IL-10 and TNF-α levels remained unchanged. Notably, higher baseline CRP were significantly higher among remitters. Clinically, 77.8% of patients responded (e.g., ≥50% MADRS score reduction), and 66.7% achieved full remission (MADRS < 10) (21).
In a complementary analysis of the same cohort, Loef et al. (2021) investigated how the balance between inflammatory activity and neuroplastic support might influence ECT outcomes. In addition to assessing serum IL-6, TNF-α, and BDNF levels, the authors calculated the IL-6/BDNF and TNF-α/BDNF ratios to represent the relative dominance of immune versus neuroplastic signaling. While no direct associations were found between individual biomarker levels and MADRS scores at any time point, a significant interaction effect was observed: the relationship between BDNF and depressive severity became more negative in the presence of elevated TNF-α levels (p = 0.020). Moreover, a higher TNF-α/BDNF ratio was significantly associated with greater symptomatic burden (p = 0.007), suggesting that imbalance skewed toward proinflammatory signaling may hinder neuroplastic recovery and ECT effectiveness. Thus, these combined results suggest that ECT outcomes in LLD may be influenced by post-treatment biomarker changes, pre-existing inflammatory burden, and interactions with neuroplastic features (22).
Inflammatory dynamics and cognitive adverse events
Other studies investigated the potential relationship between inflammatory responses and cognitive adverse events of ECT in depression treatment. Tian et al., 2021 conducted a prospective controlled study including 40 patients with first-episode, drug-naive MDD (mean age ± SD: 49.5 ± 7.32; 50% female) and 40 healthy controls matched for age, sex, and education (mean age ± SD: 48.3 ± 6.76; 55% female). Individuals underwent 12 sessions of modified ECT (three per week), initiated alongside a stable antidepressant regimen. Blood samples were collected before and after treatment to assess peripheral levels of NLRP3 inflammasome, interleukin-18 (IL-18), and nuclear factor kappa B (NF-κB). Results showed significant post-ECT increases in all three markers (NLRP3: p < 0.01; IL-18: p < 0.05; NF-κB: p < 0.01). Notably, 70% of the patients experienced cognitive impairment, defined as a score below 50 on the Wechsler Memory Scale–Revised. The elevation of inflammatory markers was significantly associated with lower cognitive scores, suggesting that inflammatory activation, particularly via NLRP3-related pathways, may contribute to cognitive adverse events observed in a subset of persons undergoing ECT (23).
Ryan et al. (2022b) further assessed 62 patients with depression (mean age ± SD: 56.84 ± 15.3; 66.1% female) undergoing bilateral ECT and analyzed neutrophil-to-lymphocyte ratio (NLR), platelet-to-lymphocyte ratio (PLR), monocyte-to-lymphocyte ratio (MLR), and systemic immune-inflammation index (SII). While elevated PLR and SII correlated with lower HAMD-24 scores at baseline, higher NLR, PLR, and SII values were significantly associated with prolonged time to reorientation following ECT (24). Focusing on psychomotor aspects, Belge et al. (2021) investigated the relationship between IL-6 and depressive symptoms in a cohort of 62 participants undergoing ECT (mean age ± SD: 58.2 ± 14.8; 77.4% female). Following 8 to 12 bilateral sessions, serum IL-6 levels showed a modest yet statistically significant reduction (median from 0.74 to 0.65 pg/mL; p = 0.02). Notably, IL-6 changes correlated specifically with improvements in psychomotor retardation, as measured by a validated scale. Depressive symptoms also improved substantially, with HDRS-17 scores decreasing from 24.6 to 8.7 (p < 0.0001) (25).
Additionally, Carlier et al. (2021) examined whether baseline inflammatory markers were associated with cognitive outcomes in a cohort of 97 older adults with unipolar depression undergoing ECT (mean age ± SD: 73.1 ± 8.1; 67% female). Serum CRP, IL-6, IL-10, and TNF-α levels were measured prior to treatment, and Mini-Mental State Examination (MMSE) scores were assessed before, weekly during, and immediately after ECT. Higher baseline TNF-α and IL-10 were significantly associated with lower MMSE scores during treatment, while elevated baseline CRP predicted poorer cognitive performance immediately after ECT. These findings suggest that pre-existing inflammation may predispose older individuals to greater cognitive vulnerability during ECT (26).
Collectively, these studies indicate that peripheral inflammation may correlate with cognitive adverse events of ECT, particularly in older adults. While some markers reflect symptom improvement, others—such as TNF-α, IL-10, and inflammatory ratios—may signal greater cognitive vulnerability.
Hematological and systemic immune indices
In 2025, Kirlioglu Balcioglu et al. published a retrospective cohort study investigating the effects of ECT on inflammatory markers in a sample of 156 patients (mean ± SD age: 36.09 ± 13.99; 58.3% male) diagnosed with psychotic and mood disorders. Their results showed significant post-ECT decreases in CRP, CRP–albumin ratio, NLR, MLR, PLR, and cortisol-to-albumin ratio (CAR). Symptom improvement was also observed, with a reduction in the Brief Psychiatric Rating Scale (BPRS) score from 59.93 ± 10.95 pre-ECT to 21.98 ± 5.71 post-ECT (p < 0.01) and a Clinical Global Impression – Severity (CGI-S) score decrease from 6.04 ± 0.68 to 1.23 ± 0.52 (p < 0.01) (27).
Similarly, Ali et al. (2024) studied 72 BD patients (median age [range]: 30 [17-57]; 55.6% male) and found post-ECT shifts in complete blood count-derived inflammatory markers, such as platelet distribution width (PDW), which increased from 11.7 ± 2.2 pre-ECT to 12.7 ± 2.6 post-ECT (p = 0.004), and red cell distribution width (RDW), which decreased from 14.1 ± 1.1 to 13.6 ± 1.0 (p < 0.001). No significant differences were found in platelet count, mean platelet volume, and plateletcrit, showing that ECT did not impair platelet homeostasis despite the other observed hematological changes (28). Ryan et al. (2022b), as previously discussed, also explored whether pre-treatment hematological markers—specifically blood cell-derived ratios such as NLR, PLR, MLR, and the SII—could predict cognitive and mood outcomes following ECT. While no associations with mood improvement were observed, higher baseline values of NLR, PLR, and SII were significantly associated with delayed recovery of orientation (24).
Biphasic inflammatory responses and long-term immune activity
Some studies showed a biphasic trajectory of systemic inflammation following ECT. Hough et al. (2024) examined the levels of peripheral inflammatory markers in 40 patients with treatment-resistant depression (TRD) (mean ± SD age: 41.78 ± 13.73; 55% female) undergoing ECT, compared to 26 healthy controls (mean ± SD age: 40.50 ± 13.42; 65% female). Plasma inflammatory markers were evaluated at three time points: (T1) pre-treatment, (T2) post-second ECT session, and (T3) post-treatment. The study found a transient increase in CRP and IL-6 levels from T1 to T2, followed by a decline from T2 to T3. While IL-6 returned to baseline levels post-treatment, CRP remained elevated compared to pre-treatment values. Notably, no direct correlation was identified between general inflammatory changes and symptom improvement; however, a dynamic interaction between the early/acute inflammatory response and post-treatment inflammation compared to baseline correlated with clinical outcomes. While there was no direct correlation between inflammatory changes and overall symptom reduction, larger early elevations in IL-6 at T2 were associated with more remarkable improvement in affective and cognitive symptoms among individuals with higher levels of IL-6 post-treatment (T3), while a trend to the opposite was observed in those with lower IL-6 levels after the intervention. A similar trend was observed between CRP levels trajectory and neurovegetative symptoms (29).
In 2025, Gaarden et al. conducted a longitudinal study including 64 elderly patients with treatment-resistant unipolar depression (mean ± SD age: 75.2 ± 6.3; 54.7% female) treated with ECT and 18 non-depressed controls (mean ± SD age: 78.1 ± 4.8; 66.7% female). Blood samples for analysis of 27 immune markers were collected from patients pre, mid, and post-treatment and at 12 weeks of follow-up; for controls, the markers were assessed through identical methods at baseline and at 8 weeks of follow-up. The analysis showed that 47% achieved clinical remission (as classified by a score < 8 on the 17-item HDRS). Although a significant decrease of 19 immune markers was observed from pre- to post-treatment in the patients’ intra-group comparisons, higher concentrations of 23 immune markers were observed among patients compared to controls at the follow-up. Moreover, there were no differences in immune markers concentrations between responder and non-responder patients. These results raised questions about whether prolonged inflammatory activity in older populations might influence long-term treatment efficacy or relapse risk (30).
The main characteristics of the studies evaluating peripheral inflammatory and hematological biomarkers in relation to ECT outcomes are shown in Table 1.
Neuroplasticity, neuroimmune processes, and post-mortem findings
Beyond the ECT’s impact on inflammation, Han et al., in 2023, also added evidence of neuroplasticity in a prospective cohort study including severe MDD patients divided into 102 ECT-treated (mean ± SD age: 46.3 ± 11.2; 56.86% male) and 102 non-ECT treated (mean ± SD age: 45.8 ± 10.9; 58.82% male). The primary outcome was the change in HDRS scores from baseline to 12 weeks. Both groups showed significant symptom improvement, whereas the reduction in HDRS scores was significantly greater in the ECT group (-19.6 ± 6.4) compared to the non-ECT group (-14.2 ± 7.2, p < 0.001). Moreover, 68% of the ECT-treated patients achieved remission (HDRS ≤ 7) compared to 42% in the non-ECT group. Biomarker analysis revealed that BDNF levels decreased in both groups; however, the decline was significantly attenuated in the ECT group (-12.8 ± 4.6 vs. -18.6 ± 5.8, p < 0.001), suggesting a neuroprotective effect. Similarly, IL-6 increased in both groups, although with a less pronounced rise in the ECT group (+2.2 ± 0.9 vs. +3.4 ± 1.2, p < 0.001). No significant differences were found in cortisol levels between the groups. In addition, quality of life (as assessed by the WHO Quality of Life Brief Version) and cognitive function (evaluated by the Montreal Cognitive Assessment) improved more significantly in the ECT group. The findings of this study suggested that ECT led to significant symptomatic improvement and may influence neuroplasticity and inflammation in severe MDD (31).
Several studies explored neuroimmune interactions and cellular changes associated with ECT. Xu et al. (2023) investigated astrocytic markers and inflammatory cytokines in 40 TRD patients (mean ± SD age: 22.2 ± 4.5; 60% female) undergoing ECT and 35 healthy controls (mean ± SD age: 23.1 ± 3.9; 65.7% female). Pre-ECT, levels of glial fibrillary acidic protein (GFAP), S100 calcium-binding protein B (S100β), CD81, IL-1β, IL-4, IL-6, IL-10, IL-17, tumor necrosis factor-alpha (TNF-α), and inflammatory markers interferon γ (IFN-γ) were significantly higher in TRD patients than in controls. These elevations were positively correlated with cognitive dysfunction (r = 0.62, p < 0.001). Following ECT, there were significant decreases of GFAP, S100β, CD81, as well as IL-4 and IFN-γ in the TRD group compared to controls. Alongside these changes, patients showed significant improvement in all subscales of the Montgomery-Åsberg Depression Rating Scale (MADRS). Furthermore, the IL-4 decrease correlated with reductions in the MADRS vegetative subscale scores. This study added that ECT may also exert anti-inflammatory effects through astrocytic activity modulation in depressed individuals (32).
Abe et al. (2023) investigated the role of high mobility group box-1 protein (HMGB1) and soluble receptor for advanced glycation end-products (sRAGE) in the context of ECT, analyzing 25 MDD patients (median age [range]: 54.2 [48–63]; 56% female) and 25 healthy controls (median age [range]: 53.2 [47–61]; 56% female). HMGB1 is a damage-associated molecular pattern protein involved in neuroinflammation, blood-brain barrier dysfunction, and immune activation, while sRAGE functions as a decoy receptor that neutralizes pro-inflammatory HMGB1 signaling. Given prior evidence linking elevated HMGB1 to depression and stress-related neuroimmune dysregulation, it was hypothesized that ECT might exert therapeutic effects by downregulating HMGB1-mediated inflammatory pathways or modulating sRAGE expression. Surprisingly, however, no significant differences in HMGB1 or sRAGE levels at baseline were found between MDD patients and healthy controls. While ECT led to symptoms improvement (as assessed by the 21-item HDRS), it did not induce any significant post-treatment changes (HMGB1: p = 0.677; sRAGE: p = 0.922) (33).
A study by Brooks et al. (2024) assessed inflammation patterns, neuroimaging data, and its correlations with depressive symptoms in 20 TRD patients (mean ± SD age: 42.5 ± 13.7; 50% female) undergoing ECT. Magnetic resonance imaging (MRI) and inflammation markers (e.g., IL-6, IL-8, and TNF- α) from blood samples were obtained after an index ECT and six months after. MRI was evaluated with voxel-based morphometry. The results showed a correlation between IL-8 decrease and depression symptoms improvement, as evaluated by the 17-item HDRS (r = 0.65; p = 0.01). However, despite this correlation, inflammatory changes did not directly mediate neuroimaging structural changes and depressive symptoms. Four clusters of gray matter volume significant reductions were found; one cluster, including Brodmann’s area 22 and right insula, correlated with higher severity of symptoms in HAM-D over six months. The author discussed that such structural changes may not necessarily be a sign of neurotoxicity but part of a dynamic neuroadaptive process (34).
In another study exploring possible correlations between neuroinflammation and structural brain changes following neuromodulation, Andreou et al. (2022) investigated 42 TRD patients (mean age ± SD: 43.15 ± 13.82; 54.8% female) undergoing ECT (right unilateral ultra-brief pulse stimulus; 3 sessions/week; ~30% transitioned to bilateral). Pre- and post-treatment serum levels of IL-6, IL-8, TNF-α, and CRP were measured alongside diffusion-weighted MRI to assess free-water corrected fractional anisotropy (FAt) in 17 white matter tracts. Among responders (≥50% MADRS reduction), changes in IL-8 significantly correlated with FAt alterations in the right cingulum and superior longitudinal fasciculus (r = 0.65–0.70), suggesting a role for IL-8 in glia-mediated white matter remodeling. These neuroimmune associations were absent in non-responders, reinforcing the hypothesis that inflammatory signaling may support therapeutic plasticity induced by ECT (35).
Furthermore, post-mortem histopathological analyses contributed to elucidating the neurobiological effects of ECT. In 2023, Loef et al. published a first explorative study assessing direct histopathological evidence of ECT-induced neuroplasticity in human post-mortem brain tissue, examining hippocampal samples from 12 patients with bipolar or unipolar depression (mean ± SD age: 54.25 ± 21.85; 58.3% male) who had received ECT within the five years preceding death, compared to 10 depressed donor patients who were not treated with ECT (mean ± SD age: 66.9 ± 13.47; 60% female), and 15 healthy control donors (mean ± SD age: 68.87 ± 16.02; 53.3% female). In subjects previously treated with ECT, the analyses showed a significantly higher proportion of cells positive for doublecortin (DCX) in the hippocampal CA4 area (compared to healthy control donors) and subgranular zone (compared to both non-ECT depressed patients and healthy controls donors). DCX is a marker for the presence of young neurons and cellular plasticity. In addition, a higher percentage of positive stathmin 1 cell (STMN1), another marker for neuroplasticity, was found in the hippocampal subgranular zone of ECT-treated subjects compared with non-ECT depressed patients and healthy control donors. Notably, there were no evident differences in inflammatory markers reflecting microglial or astrocytic activity, as well as in structural changes (e.g., major hippocampal cell loss, overt cytoarchitectural changes) among the three groups. Based on these results, the authors discussed that ECT may enhance hippocampal neurogenesis without promoting chronic inflammatory responses or gliosis (36).
The main characteristics of studies investigating neuroplastic and structural brain changes in relation to ECT in mood disorders are shown in Table 2.
Molecular, epigenetic, and mitochondrial function modifications
Adding novelty to the field, molecular and epigenetic analyses investigated ECT-induced changes. In 2024, Carvalho Silva et al. published an epigenome-wide longitudinal study investigating DNA methylation in 32 TRD patients (mean ± SD age: 56.9 ± 14.3; 68.7% female) undergoing ECT. Clinical severity (evaluated with the MADRS) and ECT outcomes were assessed at baseline (T0) and 1 month after intervention (T1); genome-wide methylation was analyzed at T0 and T1. Analyses showed three differentially methylated probes (DMPs), with two annotated in genes CYB5B and PVRL4; nonetheless, these probes were not significant after false discovery rate (FDR) correction. In a covariate analysis including changes in clinical symptoms (as assessed by the MADRS), four DMPs were found annotated in the genes FAM20C, EPB41, OTUB1, and ADARBI1. Considering response status as a covariate to the model, three DMPs were detected, with two annotated in the genes IQCE and FAM20C. However, none of these probes remained significant after the FDR correction. Similarly, fifty-four differentially methylated regions (DMRs) were found, alongside the identification of 21 DMRs for symptoms variations and 26 DMRs for response status. Again, none of these regions remained significant after the FDR correction (37). In contrast to genome-wide approach, Moschny et al. (2020) analyzed DNA methylation patterns of tissue-type plasminogen activator (t-PA; gene: PLAT) and its inhibitor PAI-1 (SERPINE1) in sorted immune cells (B cells, T cells, monocytes, and NK cells), as well as in whole blood, from 87 TRD participants undergoing ECT (mean ± SD age: 51.9 ± 16.6; 51.7% female). Although significant variability in methylation across immune cell types was observed, no consistent differences in PLAT or SERPINE1 methylation were found between remitters and non-remitters. Clinical response (≥50% MADRS reduction) was achieved in 33 patients; however, methylation profiles were not associated with symptom improvement (38).
Maier et al. (2023) conducted a prospective observational study to examine the effects of ECT on DNA methylation in genes related to hypothalamic–pituitary–adrenal (HPA) axis regulation. The study included 31 patients with MDD (mean ± SD age: 55 ± 16; 64.5% female), alongside 19 unmedicated depressed controls and 20 healthy controls. All patients underwent right unilateral ECT (3 sessions/week; average of 10 ± 4 sessions) using the Thymatron IV system. Peripheral blood mononuclear cells (PBMCs) were collected to assess methylation patterns of NR3C1 (encoding the glucocorticoid receptor) and POMC. Results showed that NR3C1 methylation was significantly higher in ECT non-responders compared to responders at baseline; however, it decreased in both groups after treatment. In contrast, POMC methylation decreased in responders but increased in non-responders by the end of the ECT course. These findings suggest that ECT may induce short-term epigenetic modifications in stress-related genes and that NR3C1 methylation, in particular, could serve as a candidate biomarker for treatment response prediction (39).
Beyond methylation, new studies on small RNA sequencing have provided further insights into the molecular underpinnings of ECT response. In 2023, Kaurani et al. analyzed whole-blood miRNA profiles obtained from 64 patients with treatment-resistant MDD (mean ± SD age: 55.9 ± 16; 59.38% female) at three different times (e.g., before, after the first, and after the last ECT). Clinically, patients were assessed by the MADRS. The main outcome showed that miR-223-3p was down-regulated at baseline among responder subjects (e.g., 62.5% individuals with a reduction of ≥50% in MADRS total score) in comparison to non-responders. A ROC analysis showed that miR-223-3p distinguished these two groups with an area under curve of 0.76 (95% CI = 0.6–0.91; p = 0.0031), supporting its potential as a predictive biomarker. In addition, a negative correlation was observed between miR-223-3p expression and percentage changes in MADRS (r = -0.32; p = 0.04). This finding leads to an understanding that low miR-223-3p expression at baseline correlates with higher severity of depressive symptoms and that its higher post-treatment expression is associated with symptoms improvement. The results of this study also showed higher expressions of proinflammatory markers (NLRP3, IL-6, IL-1B, and TNF-α) at baseline in responders compared to non-responders, again underscoring the potential of ECT for inflammatory modulation (40). Similarly, Galbiati et al. (2025) performed miRNA analyses in 27 TRD individuals (mean ± SD age: 58.1 ± 11.2; 67% female) treated with ECT. Blood samples were obtained at baseline (T0) and 1 month after the last ECT session (T1); depression symptoms were assessed by the MADRS. Their outcomes showed that eight microRNAs were downregulated after ECT. Symptoms improvement among responders (18 out of 27 subjects) directly correlated with changes in miR-30c-5p (r = 0.616; p = 0.006) and miR-324-3p (r = 0.473; p = 0.048) levels between T0 and T1. In the whole sample (e.g., responders and non-responders), only miR-324-3p correlated with depressive symptoms relief (r = 0.460, p = 0.016) (41).
Moreover, mitochondrial function was investigated by Ryan et al. (2023) in a study including 100 individuals with depression (mean ± SD age: 56.36 ± 14.28; 62% female) treated with ECT and 89 healthy controls (mean age 53.42 ± 10.39; 66.29% female). Blood samples were obtained to measure mtDNAcn levels. Assessed outcomes were the depression severity, ECT response, telomere length, and inflammatory markers. The analyses showed significantly elevated mtDNAcn in depressed individuals compared to controls, even after adjustments for potential confounding factors. However, higher baseline mtDNAcn levels were insufficient as a biomarker for ECT outcome prediction, as no significant differences were observed between remitters and non-remitters or responders and non-responders. Alongside this finding, mtDNAcn was not associated with inflammatory markers levels, intensity of depression symptoms, or telomere length (42).
The main characteristics of studies investigating genetic, epigenetic, microRNA, and mitochondrial biomarkers in the context of ECT in mood disorders are shown in Table 3.
Discussion
Our review assessed recent evidence on ECT and mood disorders. The analyzed studies highlight key pathways through which ECT exerts its effects, spanning systemic immune modulation, neuroplasticity, genetic regulation, and mitochondrial function. Despite heterogeneity in studies designs, the collective evidence suggests, in accordance with previous studies of the field, that ECT does not merely act at the neurotransmitter level but induces broader biological adaptations that may underpin its therapeutic efficacy (43). Importantly, these processes are not isolated, as neurotransmitter alterations may act as upstream modulators influencing these multimodal effects.
Regarding systemic inflammation following ECT, some studies reported marked reductions in pro-inflammatory markers such as CRP, IL-6, and TNF-α post-ECT (17, 27, 28), while others revealed a biphasic response, characterized by initial immune activation followed by partial resolution (29, 30). Whether these distinct patterns can reflect consequences of influence by other factors (such as sample profile and assessment methods) or epiphenomenon is still a knowledge gap. While most biomarker studies have emphasized peripheral inflammation, emerging evidence also points to ECT’s capacity to modulate central immune activity. Findings from cerebrospinal fluid suggest that neuroimmune shifts may accompany clinical improvement, supporting a broader view of ECT as influencing both systemic and central pathways (18).
Interestingly, sex-specific immune responses to ECT—such as IL-8 increases associated with symptom improvement only in women—highlight the potential role of sex hormones and sex-linked immune regulation in treatment outcomes (19). Age-related differences have also been noted: adolescents with MDD exhibited a prolonged and sustained reduction in IL-1β and IL-6 post-ECT, alongside an increase in IL-10, an anti-inflammatory cytokine. These findings showed that younger individuals may exhibit a favorable profile in terms of ECT immune-inflammatory modulation, potentially due to greater immune plasticity or a lower baseline inflammatory burden (17). In contrast, among elderly TRD patients, inflammatory markers remained elevated in a subset of individuals, suggesting that, in this population, immune dysregulation may persist despite clinical symptom improvement; it raises concerns about whether residual inflammation contributes to long-term relapse risk (22). The differences between these age groups indicate the potential immune aging (inflammaging) on ECT response (22, 44). Future research should investigate whether baseline inflammatory profiles can predict differential responses to ECT across age groups and explore personalized approaches to optimize treatment efficacy.
Furthermore, specific symptom clusters may also be influenced by immune modulation. Psychomotor symptoms—often disabling and treatment-resistant—have been shown to associate with IL-6 shifts following ECT, suggesting that targeting immune pathways may have functional relevance beyond core mood symptoms (25). In parallel, cognitive vulnerability following ECT may be related to individual inflammatory profiles in patients with MDD. Inflammatory mediators such as NLRP3 inflammasome, IL-18, and NF-κB were significantly elevated after ECT in MDD patients and negatively correlated with memory scores. Similarly, higher pre-treatment TNF-α, IL-10, and CRP predicted poorer cognitive performance during or after ECT in depressed individuals. These findings suggest that pre-ECT immune phenotyping might aid in predicting short-term cognitive risk in this population (23).
Pharmacological strategies targeting neuroinflammation, such as the COX-2 inhibitor celecoxib, have been explored as potential adjuncts in mood disorders treatment (45–48). Indeed, Kargar et al. (2014) demonstrated that celecoxib, compared with placebo, effectively reduced TNF-α levels in patients with BD undergoing ECT. However, no reductions in other inflammatory markers (e.g., IL-6, IL-1, and high-sensitivity CRP) or direct link between TNF-α decrease and greater symptomatic improvement were observed. Specifically on age, a randomized controlled trial including a large sample of cognitively normal individuals with late-life depression showed that either celecoxib or naproxen was insufficient to improve depressive symptoms compared to placebo (47). Additionally, Kargar et al. (2015) explored the effects of celecoxib in manic patients receiving ECT, assessing its potential role in modulating BDNF levels, a critical mediator of neuroplasticity. While there was a trend toward increased BDNF in patients receiving celecoxib, it did not reach statistical significance, and no correlation was observed between BDNF changes and clinical response (48). These findings suggest that while anti-inflammatory strategies may modulate peripheral and central immune-inflammatory responses, their impact on neuroplasticity and symptomatic improvement remains uncertain (45–48).
The neuroplastic effects of ECT have been supported by studies demonstrating increased expression of hippocampal neurogenesis markers (36). These recent findings align with preclinical research showing that electroconvulsive seizures (ECS) in rodents stimulate neural progenitor cell proliferation and dendritic remodeling (49). The absence of significant microglial activation in post-mortem human hippocampal tissue from ECT-treated patients further suggests that ECT may promote structural brain recovery without triggering sustained neuroinflammation (36). However, contrasting results from animal models, such as ECS failing to prevent lipopolysaccharide-induced microglial activation and behavioral effects, reinforce that neuroimmune response to ECT may be context-dependent, varying with baseline inflammatory status and underlying pathology (50). Despite new data, it is still unclear whether neuroplastic changes correlate with long-term functional outcomes. Longitudinal studies assessing whether these biomarkers relate to relapse risk or cognitive effects could refine clinical decision-making; strategies for augmentation of ECT’s neuroplastic effect may further enhance the precision of neuromodulatory interventions. Notably, emerging neuroimaging studies in humans suggest that glial activity may mediate ECT-induced white matter reorganization. Rather than reflecting nonspecific volumetric effects, structural adaptations appear selectively associated with inflammatory shifts in responders, reinforcing the view that glia-mediated immune signaling may support functional plasticity in TRD (35).
Identifying trends in DNA methylation following ECT in the study by Carvalho Silva et al., albeit lacking statistical significance after multiple corrections, suggests possible effects on cellular reprogramming, which may contribute to neuroplasticity and clinical outcomes in MDD (37). This new analysis contributed to pre-existing investigations; however, newer investigations with larger populations are necessary for robust conclusions (7). Beyond global methylation patterns, recent evidence suggests that ECT may epigenetically modulate components of the stress-response system. Changes in the methylation of NR3C1 and POMC—two key HPA-axis genes—were associated with treatment response, indicating that epigenetic regulation of stress-related pathways may represent a mechanistic link between biological vulnerability and ECT efficacy, particularly in stress-linked mood phenotypes (39).
Still within genetics, the valuable findings of two included studies in this review on microRNA among TRD patients treated with ECT (40, 41) fit within the general trajectory of omics research on treatment-resistant depression (51). The findings indicate that biomarker-driven stratification might enable precision-medicine techniques to select appropriate candidates for ECT. In this setting, translational studies showed that antidepressant treatments have been associated with changes in miRNA expression profiles (such as miR-1202) (52), and current research reinforces that interventions targeting miRNAs may regulate immune functions and improve mood disorders treatment outcomes (53–55). However, gaps within the field involving ECT still remain. The stability of DNA methylation and miRNA changes over time, their sensitivity to external factors such as medication use (53, 56), and their specificity to ECT rather than other antidepressant interventions require deep investigation. Longitudinal studies with repeated sampling, larger cohorts, and controlled confounders are essential to determine whether these molecular alterations can be effectively integrated into clinical decision-making.
Mitochondrial function has also emerged as a relevant domain, with one cited study showing elevated mtDNA copy number in MDD patients, though without clear differentiation between ECT responders and non-responders (42). A plausible explanation for the lack of association between mtDNAcn levels and ECT response is that mitochondrial adaptations may occur at a functional rather than a purely quantitative level. While mtDNAcn is an indirect measure of mitochondrial activity, it does not fully capture key parameters such as mitochondrial membrane potential, ATP production, or reactive oxygen species generation (57, 58). The observed variability in mitochondrial adaptations following ECT requires further studies on comprehensive metabolic profiling, particularly in the context of oxidative stress and neuroenergetics (58). Given that mitochondrial dysfunction is implicated in multiple psychiatric disorders, including MDD, BD, and schizophrenia, further exploration of ECT’s role in mitochondrial resilience could refine understanding of its therapeutic potential (59–61). Additionally, interventions that enhance mitochondrial function, such as metabolic modulators or antioxidant therapies, may represent promising adjuncts to ECT, particularly for patients with high oxidative stress burden (58, 60, 61). Despite recent advances, interactions between mitochondrial function, inflammation, and neuroplasticity remain underexplored in mental disorders. Therefore, integrative research assessing these pathways could contribute to new mechanistic concepts.
Beyond ECT, other neuromodulation techniques have been investigated for their effects on immune, inflammatory, and cellular mechanisms in mood disorders. Repetitive transcranial magnetic stimulation (rTMS), transcranial direct current stimulation (tDCS), and vagus nerve stimulation (VNS) have all demonstrated varying degrees of immunomodulatory and neuroplastic effects. A systematic review encompassing both animal and human studies examined the impact of rTMS on inflammatory markers in psychiatric disorders. The review found that while animal models showed positive changes in microglial activity and anti-inflammatory effects associated with behavioral improvement, these findings were not consistently replicated in human studies focusing on TRD. Specifically, several human studies reported rTMS-induced alterations in peripheral inflammatory markers, but only a minority demonstrated an association with clinical treatment response. The review also noted that most studies exhibited poor or moderate quality in bias assessment, indicating a need for more rigorous research in this area (62). A pilot study published in 2024 by Lespérance et al. assessed the immunomodulatory effects of VNS in TRD patients over an extended period of up to four years. Plasma levels of 40 soluble inflammatory molecules, including cytokines, were measured before VNS implantation and throughout treatment. The results showed significant modifications in cytokine levels that were associated with clinical response in depressive symptoms, suggesting a potential relation between inflammatory modulation and response to treatment. Nonetheless, the small sample size and an extended follow-up of this study limits its generalizability (63). A study by Goerigk et al. (2020) examined the role of peripheral biomarkers associated with neuroplasticity and immune-inflammatory processes in patients with BD undergoing tDCS treatment. The findings suggested that tDCS modulates these biomarkers, particularly IL-8, which significantly reduced following active tDCS. Additionally, higher baseline IL-6 plasma levels were associated with symptomatic improvement post-tDCS (64), aligning with ECT and suggesting that pre-existing inflammatory states may influence treatment response (65, 66). Head-to-head comparisons between ECT and other modulatory interventions are needed to delineate their respective neurobiological mechanisms and optimize patient selection among different modalities.
The newer research in this review contributed to the field with noteworthy insights, yet numerous limitations still exist. First, different patients’ profile across samples (e.g., age, sex, clinical history, body mass index, years of education, ethnicity), heterogeneous outcome variables, distinct ECT protocols, serum collections, and time of follow-up may halt the overall interpretation of the included studies. Secondly, the multimodal responses to ECT, even in studies with similar methods, demonstrate the difficulty in identifying standard modulatory mechanisms. Thirdly, while novel results (such as neuroplasticity and genetic/epigenetic findings) TRD mechanistic insights and phenomenological correlations, their clinical translation remains preliminary, necessitating validation in larger cohorts/randomized controlled trials.
In summary, our review support the view that ECT may modulate immune-inflammatory, neuroplastic, and epigenetic mechanisms, reinforcing its function as a systemic neuromodulatory intervention rather than a purely neurotransmitter-based therapy. While these domains have often been studied in isolation, recent findings suggest a complex interplay between immune regulation, synaptic remodeling, and gene expression in underpinning ECT’s therapeutic effects (65). Neuroinflammatory changes may facilitate neuroplasticity by mitigating cytokine-induced synaptic dysfunction and neurodegeneration. This aligns with post-mortem and imaging studies demonstrating hippocampal neurogenesis and structural remodeling in ECT-treated patients (36). Moreover, epigenetic modifications may be molecular switches linking inflammatory modulation to synaptic plasticity (40, 41). Mitochondrial function represents another critical intersection in this neuroimmune interface. Since mitochondria are key oxidative stress and ATP production regulators, their dysfunction could exacerbate neuroinflammation and impair synaptic remodeling. However, current studies suggest that ECT-induced mitochondrial adaptations may occur at a functional rather than a quantitative level, requiring advanced metabolic profiling to elucidate their clinical significance (42, 58).
Conclusion
Novel research has added important insights into immune-inflammatory, neuroplastic, and genetic mechanisms related to ECT in mood disorders. Despite remaining gaps, these findings pave the way for precision-based therapeutic strategies.
Author contributions
RJ: Conceptualization, Data curation, Formal analysis, Investigation, Methodology, Project administration, Validation, Visualization, Writing – original draft, Writing – review & editing. JA: Conceptualization, Data curation, Investigation, Writing – original draft, Writing – review & editing. DP: Data curation, Investigation, Writing – original draft, Writing – review & editing. PR: Conceptualization, Data curation, Investigation, Methodology, Project administration, Supervision, Writing – original draft, Writing – review & editing. PO: Conceptualization, Data curation, Investigation, Methodology, Project administration, Supervision, Writing – original draft, Writing – review & editing.
Funding
The author(s) declare that no financial support was received for the research, and/or publication of this article.
Acknowledgments
We acknowledge all collaborators of Goiânia Neurological Institute and the Federal University of Goiás for supporting our academic staff activities.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Glossary
ADARB1: Adenosine Deaminase RNA Specific B1
BD: Bipolar Disorder
BDNF: Brain-Derived Neurotrophic Factor
BPRS: Brief Psychiatric Rating Scale
CAR: Cortisol-to-Albumin Ratio
CGI-S: Clinical Global Impression – Severity Scale
COX-2: Cyclooxygenase-2
CRP: C-Reactive Protein
CRP-albumin ratio: C-Reactive Protein to Albumin Ratio
CYB5B: Cytochrome B5 Type B
DBS: Deep Brain Stimulation
DCX: Doublecortin
DMP: Differentially Methylated Positions
DMR: Differentially Methylated Regions
DNA: Deoxyribonucleic Acid
ECT: Electroconvulsive Therapy
ECS: Electroconvulsive Seizures
EPB41: Erythrocyte Membrane Protein Band 4.1
FAM20C: Family with Sequence Similarity 20 Member C
FDR: False Discovery Rate
GABA: Gamma-Aminobutyric Acid
GFAP: Glial Fibrillary Acidic Protein
HAMD-24: Hamilton Depression Rating Scale 24-item version
HDRS: Hamilton Depression Rating Scale
HMGB1: High Mobility Group Box-1 Protein
Iba1: Ionized Calcium-Binding Adapter Molecule 1
IFN-γ: Interferon Gamma
IL-1β: Interleukin-1 Beta
IL-6: Interleukin-6
IL-8: Interleukin-8
IL-10: Interleukin-10
IL-18: Interleukin-18
IQCE: IQ Motif Containing E
MADRS: Montgomery-Åsberg Depression Rating Scale
MDD: Major Depressive Disorder
MEG: Magnetoencephalography
miRNA: MicroRNA
MLR: Monocyte-to-Lymphocyte Ratio
MMSE: Mini-Mental State Examination
MoCA: Montreal Cognitive Assessment
mtDNAcn: Mitochondrial DNA Copy Number
NF-κB: Nuclear Factor Kappa B
NLR: Neutrophil-to-Lymphocyte Ratio
NLRP3: NLR Family Pyrin Domain Containing 3 Inflammasome
NR3C1: Nuclear receptor subfamily 3 group C member 1 (glucocorticoid receptor)
OTUB1: OTU Deubiquitinase, Ubiquitin Aldehyde Binding 1
PDGF-BB: Platelet-Derived Growth Factor Subunit B
PDW: Platelet Distribution Width
PLR: Platelet-to-Lymphocyte Ratio
POMC: Proopiomelanocortin
PVRL4: Poliovirus Receptor-Related 4
QoL: Quality of Life
RDW: Red Cell Distribution Width
rTMS: Repetitive Transcranial Magnetic Stimulation
sRAGE: Soluble Receptor for Advanced Glycation End Products
SD: Standard Deviation
SII: Systemic Immune-Inflammation Index
STMN1: Stathmin 1
tDCS: Transcranial Direct Current Stimulation
TNF-α: Tumor Necrosis Factor Alpha
TRD: Treatment-Resistant Depression
TR-MDD: Treatment-Resistant Major Depressive Disorder
VNS: Vagus Nerve Stimulation
WMS-R: Wechsler Memory Scale–Revised
WHOQOL-BREF: World Health Organization Quality of Life – Brief Version
References
1. Jelovac A, Kolshus E, and McLoughlin DM. Relapse following successful electroconvulsive therapy for major depression: a meta-analysis. Neuropsychopharmacology. (2013) 38:2467–74. doi: 10.1038/npp.2013.149
2. Perugi G, Medda P, Toni C, Mariani MG, Socci C, and Mauri M. The role of electroconvulsive therapy (ECT) in bipolar disorder: effectiveness in 522 patients with bipolar depression, mixed-state, mania and catatonic features. Curr Neuropharmacol. (2017) 15:359–71. doi: 10.2174/1570159X14666161017233642
3. Bahji A, Hawken ER, Sepehry AA, Cabrera CA, and Vazquez G. ECT beyond unipolar major depression: systematic review and meta-analysis of electroconvulsive therapy in bipolar depression. Acta Psychiatr Scand. (2019) 139:214–26. doi: 10.1111/acps.12994
4. Young JR, Evans MK, Hwang J, Kritzer MD, Kellner CH, and Weiner RD. Electroconvulsive therapy changes immunological markers in patients with major depressive disorder: A scoping review. J ECT. (2024) 40:232–9. doi: 10.1097/YCT.0000000000001021
5. Cojocaru AM, Vasile AI, and Trifu SC. Neurobiological mechanisms and therapeutic impact of electroconvulsive therapy (ECT). Rom J Morphol Embryol. (2024) 65:13–7. doi: 10.47162/RJME.65.1.02
6. Baldinger P, Lotan A, Frey R, Kasper S, Lerer B, and Lanzenberger R. Neurotransmitters and electroconvulsive therapy. J ECT. (2014) 30:116–21. doi: 10.1097/YCT.0000000000000138
7. Yrondi A, Sporer M, Péran P, Schmitt L, Arbus C, and Sauvaget A. Electroconvulsive therapy, depression, the immune system and inflammation: A systematic review. Brain Stimul. (2018) 11:29–51. doi: 10.1016/j.brs.2017.10.013
8. Castro SCC, Bicca C, Bicca B, Araujo S, and Viola TW. A systematic mini-review of epigenetic mechanisms associated with electroconvulsive therapy in humans. Front Hum Neurosci. (2023) 17:1143332. doi: 10.3389/fnhum.2023.1143332
9. Jiang X and Xie Q. Efficacy and safety of modified electroconvulsive therapy for the refractory depression in older patients. Asia-Pac Psychiatry. (2020) 12:e12411. doi: 10.1111/appy.12411
10. Beurel E, Toups M, and Nemeroff CB. The bidirectional relationship of depression and inflammation: double trouble. Neuron. (2020) 107:234–56. doi: 10.1016/j.neuron.2020.06.002
11. Kouba BR, de Araujo Borba L, Borges de Souza P, Gil-Mohapel J, and Rodrigues ALS. Role of inflammatory mechanisms in major depressive disorder: from etiology to potential pharmacological targets. Cells. (2024) 13:423. doi: 10.3390/cells13050423
12. Maffioletti E, Carvalho Silva R, Bortolomasi M, Baune BT, Gennarelli M, and Minelli A. Molecular biomarkers of electroconvulsive therapy effects and clinical response: understanding the present to shape the future. Brain Sci. (2021) 11:1120. doi: 10.3390/brainsci11091120
13. Wilkinson ST, Sanacora G, and Bloch MH. Hippocampal volume changes following electroconvulsive therapy: a systematic review and meta-analysis. Biol Psychiatry Cognit Neurosci Neuroimaging. (2017) 2:327–35. doi: 10.1016/j.bpsc.2017.01.011
14. American Psychiatric Association. Diagnostic and statistical manual of mental disorders, 5th ed. (2013) (Arlington, VA: American Psychiatric Publishing). doi: 10.1176/appi.books.9780890425596.
15. World Health Organization. The ICD-10 classification of mental and behavioural disorders: Clinical descriptions and diagnostic guidelines. Geneva: World Health Organization (1992).
16. World Health Organization. International classification of diseases for mortality and morbidity statistics (11th Revision) (2019). Available online at: https://icd.who.int/en (Accessed January 15, 2025).
17. Du N, Wang Y, Geng D, Chen H, Chen F, Kuang L, et al. Effects of electroconvulsive therapy on inflammatory markers and depressive symptoms in adolescents with major depressive disorder. Front Psychiatry. (2024) 15:1447839. doi: 10.3389/fpsyt.2024.1447839
18. Mindt S, Neumaier M, Hoyer C, Sartorius A, and Kranaster L. Cytokine-mediated cellular immune activation in electroconvulsive therapy: A CSF study in patients with treatment-resistant depression. World J Biol Psychiatry. (2020) 21:139–47. doi: 10.1080/15622975.2019.1618494
19. Kruse JL, Olmstead R, Hellemann G, Wade B, Jiang J, Vasavada MM, et al. Inflammation and depression treatment response to electroconvulsive therapy: Sex-specific role of interleukin-8. Brain Behav Immun. (2020) 89:59–66. doi: 10.1016/j.bbi.2020.05.069
20. Ryan KM and McLoughlin DM. Peripheral blood inflammatory markers in depression: Response to electroconvulsive therapy and relationship with cognitive performance. Psychiatry Res. (2022) 315:114725. doi: 10.1016/j.psychres.2022.114725
21. Carlier A, Rhebergen D, Schilder F, Bouckaert F, Sienaert P, Veerhuis R, et al. The pattern of inflammatory markers during electroconvulsive therapy in older depressed patients. World J Biol Psychiatry. (2021) 22:770–7. doi: 10.1080/15622975.2021.1907718
22. Loef D, Vansteelandt K, Oudega ML, van Eijndhoven P, Carlier A, van Exel E, et al. The ratio and interaction between neurotrophin and immune signaling during electroconvulsive therapy in late-life depression. Brain Behav Immun Health. (2021) 18:100389. doi: 10.1016/j.bbih.2021.100389
23. Tian H, Li G, Xu G, Liu J, Wan X, Zhang J, et al. Inflammatory cytokines derived from peripheral blood contribute to the modified electroconvulsive therapy-induced cognitive deficits in major depressive disorder. Eur Arch Psychiatry Clin Neurosci. (2021) 271:475–85. doi: 10.1007/s00406-020-01128-9
24. Ryan KM, Lynch M, and McLoughlin DM. Blood cell ratios in mood and cognitive outcomes following electroconvulsive therapy. J Psychiatr Res. (2022) 156:729–36. doi: 10.1016/j.jpsychires.2022.11.016
25. Belge JB, Van Diermen L, Sabbe B, Moens J, Morrens M, Coppens V, et al. Improvement of psychomotor retardation after electroconvulsive therapy is related to decreased IL-6 levels. Prog Neuropsychopharmacol Biol Psychiatry. (2021) 105:110146. doi: 10.1016/j.pnpbp.2020.110146
26. Carlier A, Rhebergen D, Veerhuis R, Schouws S, Oudega ML, Eikelenboom P, et al. Inflammation and cognitive functioning in depressed older adults treated with electroconvulsive therapy: A prospective cohort study. J Clin Psychiatry. (2021) 82:20m13631. doi: 10.4088/JCP.20m13631
27. Kirlioglu Balcioglu SS, Kilictutan A, Ozer D, Guclu O, and Namli MN. Impact of electroconvulsive therapy on inflammatory markers in patients with severe mental disorders. J Psychiatr Res. (2025) 182:297–303. doi: 10.1016/j.jpsychires.2025.01.036
28. Ali E, Embaby A, Arafa SM, Elbana AK, Ghazala M, and Ibrahim D. Electroconvulsive therapy improves hematological inflammatory markers in bipolar disorder. Psychopharmacol (Berl). (2024) 241:351–7. doi: 10.1007/s00213-023-06491-8
29. Hough CM, Kruse JL, Espinoza RT, Brooks JO 3rd, Congdon EJ, Norris V, et al. Trajectory of peripheral inflammation during index ECT in association with clinical outcomes in treatment-resistant depression. Brain Behav Immun Health. (2024) 43:100925. doi: 10.1016/j.bbih.2024.100925
30. Gaarden TL, Engedal K, Benth JŠ, Larsen M, Lorentzen B, Mollnes TE, et al. Persistent pro-inflammatory trait in elderly patients following treatment-resistant major depressive disorder: a longitudinal exploratory study. Nord J Psychiatry. (2025) 79:42–51. doi: 10.1080/08039488.2024.2432981
31. Han KY, Wang CM, Du CB, Qiao J, Wang YL, and Lv LZ. Treatment outcomes and cognitive function following electroconvulsive therapy in patients with severe depression. World J Psychiatry. (2023) 13:949–57. doi: 10.5498/wjp.v13.i11.949
32. Xu SX, Xie XH, Yao L, Wang W, Zhang H, Chen MM, et al. Human in vivo evidence of reduced astrocyte activation and neuroinflammation in patients with treatment-resistant depression following electroconvulsive therapy. Psychiatry Clin Neurosci. (2023) 77:653–64. doi: 10.1111/pcn.13596
33. Abe H, Okada-Tsuchioka M, Kajitani N, Omori W, Itagaki K, Shibasaki C, et al. Serum levels of high mobility group box-1 protein (HMGB1) and soluble receptors of advanced glycation end-products (RAGE) in depressed patients treated with electroconvulsive therapy. Neuropsychopharmacol Rep. (2023) 43:359–64. doi: 10.1002/npr2.12358
34. Brooks JO 3rd, Kruse JL, Kubicki A, Hellemann G, Espinoza RT, Irwin MR, et al. Structural brain plasticity and inflammation are independently related to changes in depressive symptoms six months after an index ECT course. Psychol Med. (2024) 54:108–16. doi: 10.1017/S0033291722003555
35. Andreou B, Reid B, Lyall AE, Cetin-Karayumak S, Kubicki A, Espinoza R, et al. Longitudinal trajectory of response to electroconvulsive therapy associated with transient immune response & white matter alteration post-stimulation. Transl Psychiatry. (2022) 12:191. doi: 10.1038/s41398-022-01960-8
36. Loef D, Tendolkar I, van Eijndhoven PFP, Hoozemans JJM, Oudega ML, Rozemuller AJM, et al. Electroconvulsive therapy is associated with increased immunoreactivity of neuroplasticity markers in the hippocampus of depressed patients. Transl Psychiatry. (2023) 13:355. doi: 10.1038/s41398-023-02658-1
37. Carvalho Silva R, Martini P, Hohoff C, Mattevi S, Bortolomasi M, Abate M, et al. Unraveling epigenomic signatures and effectiveness of electroconvulsive therapy in treatment-resistant depression patients: a prospective longitudinal study. Clin Epigenetics. (2024) 16:93. doi: 10.1186/s13148-024-01704-z
38. Moschny N, Jahn K, Bajbouj M, Maier HB, Ballmaier M, Khan AQ, et al. DNA methylation of the t-PA gene differs between various immune cell subtypes isolated from depressed patients receiving electroconvulsive therapy. Front Psychiatry. (2020) 11:571. doi: 10.3389/fpsyt.2020.00571
39. Maier HB, Moschny N, Eberle F, Jahn K, Folsche T, Schülke R, et al. DNA methylation of POMC and NR3C1-1F and its implication in major depressive disorder and electroconvulsive therapy. Pharmacopsychiatry. (2023) 56:64–72. doi: 10.1055/a-2034-6536
40. Kaurani L, Besse M, Methfessel I, Methi A, Zhou J, Pradhan R, et al. Baseline levels of miR-223-3p correlate with the effectiveness of electroconvulsive therapy in patients with major depression. Transl Psychiatry. (2023) 13:294. doi: 10.1038/s41398-023-02582-4
41. Galbiati C, Dattilo V, Bortolomasi M, Vitali E, Abate M, Menesello V, et al. Plasma microRNA levels after electroconvulsive therapy in treatment-resistant depressed patients. J ECT. (2025) 41:7. doi: 10.1097/YCT.0000000000001100
42. Ryan KM, Doody E, and McLoughlin DM. Whole blood mitochondrial DNA copy number in depression and response to electroconvulsive therapy. Prog Neuropsychopharmacol Biol Psychiatry. (2023) 121:110656. doi: 10.1016/j.pnpbp.2022.110656
43. Singh A and Kar SK. How electroconvulsive therapy works?: Understanding the neurobiological mechanisms. Clin Psychopharmacol Neurosci. (2017) 15:210–21. doi: 10.9758/cpn.2017.15.3.210
44. Li X, Li C, Zhang W, Wang Y, Qian P, and Huang H. Inflammation and aging: signaling pathways and intervention therapies. Signal Transduct Target Ther. (2023) 8:239. doi: 10.1038/s41392-023-01502-8
45. Fields C, Drye L, Vaidya V, and Lyketsos C. Celecoxib or naproxen treatment does not benefit depressive symptoms in persons age 70 and older: findings from a randomized controlled trial. Am J Geriatr Psychiatry. (2012) 20:505. doi: 10.1097/JGP.0b013e318227f4da
46. Abbasi SH, Hosseini F, Modabbernia A, Ashrafi M, and Akhondzadeh S. Effect of celecoxib add-on treatment on symptoms and serum IL-6 concentrations in patients with major depressive disorder: randomized double-blind placebo-controlled study. J Affect Disord. (2012) 141:308–14. doi: 10.1016/j.jad.2012.03.033
47. Kargar M, Yousefi A, Mojtahedzadeh M, Akhondzadeh S, Artounian V, Abdollahi A, et al. Effects of celecoxib on inflammatory markers in bipolar patients undergoing electroconvulsive therapy: a placebo-controlled, double-blind, randomised study. Swiss Med Wkly. (2014) 144:w13880. doi: 10.4414/smw.2014.13880
48. Kargar M, Yoosefi A, Akhondzadeh S, Artonian V, Ashouri A, and Ghaeli P. Effect of adjunctive celecoxib on BDNF in manic patients undergoing electroconvulsive therapy: a randomized double-blind controlled trial. Pharmacopsychiatry. (2015) 48:268–73. doi: 10.1055/s-0035-1559667
49. Giacobbe J, Pariante CM, and Borsini A. The innate immune system and neurogenesis as modulating mechanisms of electroconvulsive therapy in pre-clinical studies. J Psychopharmacol. (2020) 34:1086–97. doi: 10.1177/0269881120936538
50. van Buel EM, Bosker FJ, van Drunen J, Strijker J, Douwenga W, Klein HC, et al. Electroconvulsive seizures (ECS) do not prevent LPS-induced behavioral alterations and microglial activation. J Neuroinflammation. (2015) 12:232. doi: 10.1186/s12974-015-0454-x
51. Amasi-Hartoonian N, Pariante CM, Cattaneo A, and Sforzini L. Understanding treatment-resistant depression using "omics" techniques: a systematic review. J Affect Disord. (2022) 318:423–55. doi: 10.1016/j.jad.2022.09.011
52. Lopez JP, Lim R, Cruceanu C, Crapper L, Fasano C, Labonte B, et al. miR-1202 is a primate-specific and brain-enriched microRNA involved in major depression and antidepressant treatment. Nat Med. (2014) 20:764–8. doi: 10.1038/nm.3582
53. Funatsuki T, Ogata H, Tahara H, Shimamoto A, Takekita Y, Koshikawa Y, et al. Changes in multiple microRNA levels with antidepressant treatment are associated with remission and interact with key pathways: a comprehensive microRNA analysis. Int J Mol Sci. (2023) 24:12199. doi: 10.3390/ijms241512199
54. Roy B and Dwivedi Y. An insight into the sprawling microverse of microRNAs in depression pathophysiology and treatment response. Neurosci Biobehav Rev. (2023) 146:105040. doi: 10.1016/j.neubiorev.2023
55. Roy B, Dunbar M, Shelton RC, and Dwivedi Y. Identification of microRNA-124-3p as a putative epigenetic signature of major depressive disorder. Neuropsychopharmacology. (2017) 42:864–75. doi: 10.1038/npp.2016.175
56. Davyson E, Shen X, Huider F, Adams M, Borges K, McCartney D, et al. Antidepressant exposure and DNA methylation: insights from a methylome-wide association study. medRxiv. (2024). doi: 10.1101/2024.05.01.24306640
57. Liao S, Chen L, Song Z, and He H. The fate of damaged mitochondrial DNA in the cell. Biochim Biophys Acta Mol Cell Res. (2022) 1869:119233. doi: 10.1016/j.bbamcr.2022.119233
58. Madireddy S and Madireddy S. Therapeutic interventions to mitigate mitochondrial dysfunction and oxidative stress-induced damage in patients with bipolar disorder. Int J Mol Sci. (2022) 23:1844. doi: 10.3390/ijms23031844
59. Fernström J, Mellon SH, McGill MA, Picard M, Reus VI, Hough CM, et al. Blood-based mitochondrial respiratory chain function in major depression. Transl Psychiatry. (2021) 11:593. doi: 10.1038/s41398-021-01723-x
60. Lam XJ, Xu B, Yeo PL, Cheah PS, and Ling KH. Mitochondria dysfunction and bipolar disorder: from pathology to therapy. IBRO Neurosci Rep. (2023) 14:407–18. doi: 10.1016/j.ibneur.2023.04.002
61. Rajasekaran A, Venkatasubramanian G, Berk M, and Debnath M. Mitochondrial dysfunction in schizophrenia: pathways, mechanisms and implications. Neurosci Biobehav Rev. (2015) 48:10–21. doi: 10.1016/j.neubiorev.2014.11.005
62. Pedraz-Petrozzi B, Insan S, Spangemacher M, Reinwald J, Lamadé EK, Gilles M, et al. Association between rTMS-induced changes in inflammatory markers and improvement in psychiatric diseases: a systematic review. Ann Gen Psychiatry. (2024) 23:31. doi: 10.1186/s12991-024-00514-0
63. Lespérance P, Desbeaumes Jodoin V, Drouin D, Racicot F, Miron JP, Longpré-Poirier C, et al. Vagus nerve stimulation modulates inflammation in treatment-resistant depression patients: a pilot study. Int J Mol Sci. (2024) 25:2679. doi: 10.3390/ijms25052679
64. Goerigk S, Cretaz E, Sampaio-Junior B, Vieira ÉLM, Gattaz W, Klein I, et al. Effects of tDCS on neuroplasticity and inflammatory biomarkers in bipolar depression: results from a sham-controlled study. Prog Neuropsychopharmacol Biol Psychiatry. (2021) 105:110119. doi: 10.1016/j.pnpbp.2020.110119
65. Belge JB, van Diermen L, Sabbe B, Parizel P, Morrens M, Coppens V, et al. Inflammation, hippocampal volume, and therapeutic outcome following electroconvulsive therapy in depressive patients: a pilot study. Neuropsychobiology. (2020) 79:222–32. doi: 10.1159/000506133
Keywords: electroconvulsive therapy, ECT, neuroinflammation, inflammation, neuroplasticity, genetics, epigenetics
Citation: João RB, Toiansk de Azevedo JPRP, Pereira DA, Ragazzo PC and de Oliveira PM (2025) Immune-inflammatory, neuroplastic, and epigenetic effects of electroconvulsive therapy in mood disorders: an overview of recent studies. Front. Psychiatry 16:1577530. doi: 10.3389/fpsyt.2025.1577530
Received: 16 February 2025; Accepted: 16 June 2025;
Published: 03 September 2025.
Edited by:
Joao Luciano De Quevedo, University of Texas Health Science Center at Houston, United StatesReviewed by:
Weronika Dębowska, Medical University of Warsaw, PolandSwetha K. Godavarthi, University of Pittsburgh, United States
Copyright © 2025 João, Toiansk de Azevedo, Pereira, Ragazzo and de Oliveira. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Paulo Maurício de Oliveira, cC5tLm9saXZlaXJhQGdtYWlsLmNvbQ==