 Antonia Toepffer1
Antonia Toepffer1 Marlene Früh1Tonia Rocktäschel1Johanna Ballez1Marie Troll1
Marlene Früh1Tonia Rocktäschel1Johanna Ballez1Marie Troll1 Daniel Güllmar2
Daniel Güllmar2 Kathrin Finke3,4,5
Kathrin Finke3,4,5 Philipp A. Reuken6
Philipp A. Reuken6 Andreas Stallmach6Sabine Vonderlind6
Andreas Stallmach6Sabine Vonderlind6 Ildiko Rita Dunay4,5,7,8
Ildiko Rita Dunay4,5,7,8 Christian Gaser1,3,4,5
Christian Gaser1,3,4,5 Martin Walter1,4,5
Martin Walter1,4,5 Bianca Besteher1,4,5*
Bianca Besteher1,4,5*- 1Department of Psychiatry and Psychotherapy, Jena University Hospital, Jena, Germany
- 2Medical Physics Group, Institute for Diagnostic and Interventional Radiology, Jena University Hospital, Jena, Germany
- 3Department of Neurology, Jena University Hospital, Jena, Germany
- 4German Center for Mental Health (DZPG), Jena, Germany
- 5Center for Intervention and Research on Adaptive and Maladaptive Brain Circuits Underlying Mental Health (C-I-R-C), site Halle-Jena-Magdeburg, Jena, Germany
- 6Department of Internal Medicine IV, Gastroenterology, Hepatology and Infectious Diseases, Jena University Hospital, Jena, Germany
- 7Center for Behavioral Brain Sciences (CBBS), Magdeburg, Germany
- 8Institute of Inflammation and Neurodegeneration, Otto von Guericke University Magdeburg, Magdeburg, Germany
Introduction: The long-term effects of the coronavirus disease 2019 (COVID-19) are a major concern in today’s society, with cognitive impairment being an important manifestation. Notably, men and women exhibit differences in disease progression and the prevalence of long-COVID. This study aims to investigate sex differences in cognitively impaired long-COVID individuals and their potential association with alterations in gray matter volume (GMV).
Methods: We conducted MRI at 3 Tesla to investigate brain structural correlates of cognitive impairment in long-COVID patients using voxel-based morphometry (VBM) and compared these patients to a healthy control (HC) group (n=30, female=13, male=17). Long-COVID patients underwent scanning and neuropsychiatric assessment on average 9.9 months after their acute and mostly mild COVID-19 infection. Based on Montreal Cognitive Assessment (MoCA) scores, they were classified into two groups: the PCn group, showing preserved cognitive function with MoCA scores of 26 or higher (n=36, female=23, male=13), and the PCcog group, characterized by cognitive impairment with MoCA scores below 26 (n=28, female=15, male=13). Subsequent analyses were performed separately for males and females to investigate sex-specific brain structural correlates of cognitive impairment.
Results: Our analysis revealed significant GMV alterations in long-COVID patients across various brain regions, encompassing both shared and sex-specific regional changes. In females, these alterations were more restricted, affecting anterior frontal, limbic, and diencephalic regions. In males, GMV alterations were more widespread, involving neocortical regions such as the parietal, occipital, and motor cortices, and were characterized by a greater number of affected clusters.
Discussion: Our findings demonstrate GMV alterations in both men and women with cognitive impairment, exhibiting sex-specific differences in affected regions. These differences suggest potentially distinct underlying mechanisms, highlighting the need for further research into their functional implications and relevance for personalized treatment strategies.
Introduction
The coronavirus disease 2019 (COVID-19) pandemic, caused by severe acute respiratory syndrome coronavirus 2 (SARS−CoV−2) has raised awareness of its long-term effects on human health. While many individuals recover fully, some continue to experience symptoms well beyond the initial infection. This condition is referred to by various terms, including long-COVID, post-COVID-19 syndrome, post-COVID condition (PCC), or post-acute COVID-19 syndrome (PACS). In this study, the term long-COVID is used.
Long-COVID encompasses both ongoing symptomatic COVID-19 (4–12 weeks) and post-COVID-19 condition (12 weeks or more) (1). It can occur in both hospitalized and non-hospitalized individuals and is known to affect multiple organ systems (2). During the early stages of the pandemic cognitive impairment has been reported in 13,5% to 28.85% of individuals with prior SARS-CoV2 infection (3, 4). A large English study conducted between 2020 and 2022 with 112,964 participants found that objectively measurable cognitive deficits persisted for a year or more following SARS-CoV-2 infection, particularly in those with severe illness, prolonged symptoms, or infections during the early phase of the pandemic (5). However, cognitive impairment has also been observed regardless of disease severity (6, 7).
From the onset of the pandemic, sex differences in infection rates and disease progression became evident. Men exhibited higher mortality rates and more severe disease courses (8, 9). A Swedish study on ICU patients conducted between 2020 and 2022 found that critically ill men faced a greater risk of poor long-term outcomes (10), a disparity linked to comorbidities, behavioral and lifestyle factors (9, 11), aging, and biological sex differences (8). While men were more prone to severe acute illness, women appeared to be at higher risk for persistent symptoms (12, 13). A comprehensive review involving 1.32 million patients revealed that women were significantly more likely than men to experience long-term effects of COVID-19 across multiple categories (14, 15). This may be attributed to greater symptom self-awareness in women compared to men (16) as well as a more persistent immune response (17). Beyond these biological and perceptual factors, sex-specific differences in pandemic-related psychosocial stressors and in coping strategies may also contribute to the observed disparities (18).
While previous research has investigated structural brain alterations following SARS-CoV-2 infection using various imaging techniques, findings remain inconsistent. Some studies suggest that COVID-19 can lead to changes in brain structure, including negative associations between gray matter volume (GMV) and neuropsychiatric symptoms, indicative of atrophy and loss of connectivity (19, 20). Others report positive correlations of GMV in specific brain regions and memory loss, a key neuropsychiatric symptom (21), or no alteration at all (22). These positive correlations, likely reflecting ongoing low-grade inflammation in the hippocampus, basal ganglia, thalamus, and insula, were also observed by our group (23). We hypothesized that structural alterations in these regions, which are partly components of the limbic system and the secondary olfactory network, might contribute to the neuropsychiatric symptoms observed in long-COVID (23).
Additionally, we found a negative correlation between functional connectivity in the caudate and the left precentral gyrus and Montreal Cognitive Assessment (MoCA) scores (24).
Given the inconsistent findings regarding long-COVID related brain structural changes and the lack of sex-disaggregated data, further investigation is essential to clarify the potential impact of COVID-19 on brain structure and its relationship to cognitive impairment. Reliable and standardized techniques are necessary to investigate structural brain changes in people with long-COVID, focusing on well characterized subgroups defined by age, sex, clinical features, and recovery time. Voxel-based morphometry (VBM) is a well-established neuroimaging technique for assessing GMV changes in specific brain regions (25) and has been widely applied in the study of structural alterations across various neurological and psychiatric conditions (26).
This study aimed to determine whether cognitive deficits in long-COVID individuals, assessed by the MoCA, are associated with GMV changes compared to a long-COVID group without cognitive deficits and a healthy control (HC) group. Given the previously described sex-related clinical disparities, we also hypothesized sex-specific patterns of GMV alterations.
2 Methods
2.1 Participants and assessments
A total of 94 participants were included in this cross-sectional case-control study and assigned to one of three groups.
All long-COVID patients were recruited from the long-COVID outpatient clinics of the Department of Internal Medicine IV (Infectiology) and the Department of Neurology at the University Hospital of Jena. Participants were included based solely on a confirmed long-COVID diagnosis to ensure a broad symptom spectrum; without requiring specific symptom profiles. A positive polymerase chain reaction (PCR) test was used to verify SARS-CoV-2 infection at both clinics. The diagnosis of long-COVID was based on the 2021 NICE and AWMF guidelines, which defined long-COVID as symptoms newly occurring after SARS-CoV-2 infection, not explained by other medical conditions, and persisting for > 4 weeks after infection onset (27, 28). The presence and duration of long-COVID symptoms were systematically recorded in a descriptive way via self-report, the symptom spectrum was as multi-facetted as in representative population-based studies with pronounced fatigue and cognitive impairment (29). Accordingly, 61.1% of the PCn cohort and 73.1% of the PCcog cohort reported cognitive impairment, while 75% of the PCn group and 96.2% of the PCcog group reported fatigue. Additional symptoms are listed in Table 1 (see also Supplementary Table 2). On average, the PCn group experienced 5.14 long-COVID symptoms, whereas the PCcog group reported 6.9 (see Table 1). Medical history, including information on the timing of previous SARS-CoV-2 infection(s), COVID-vaccination status and the severity of the acute COVID-19 infection as defined by the World Health Organization (WHO), was collected by a certified physician. Severity of COVID-19 was categorized into five levels based on the clinical manifestations and disease progression: uninfected (score 0), ambulatory mild disease (scores 1-3), hospitalized moderate disease (scores 4-5), hospitalized severe disease (scores 6-9), and deceased (score 10) (30). The mean WHO severity among long-COVID participants (both groups) was 2.27 (range 1 to 5, SD 0.89). Patients were enrolled and assessed in the study on average 9.9 months after infection (range 1 to 24.5 months, SD 4).
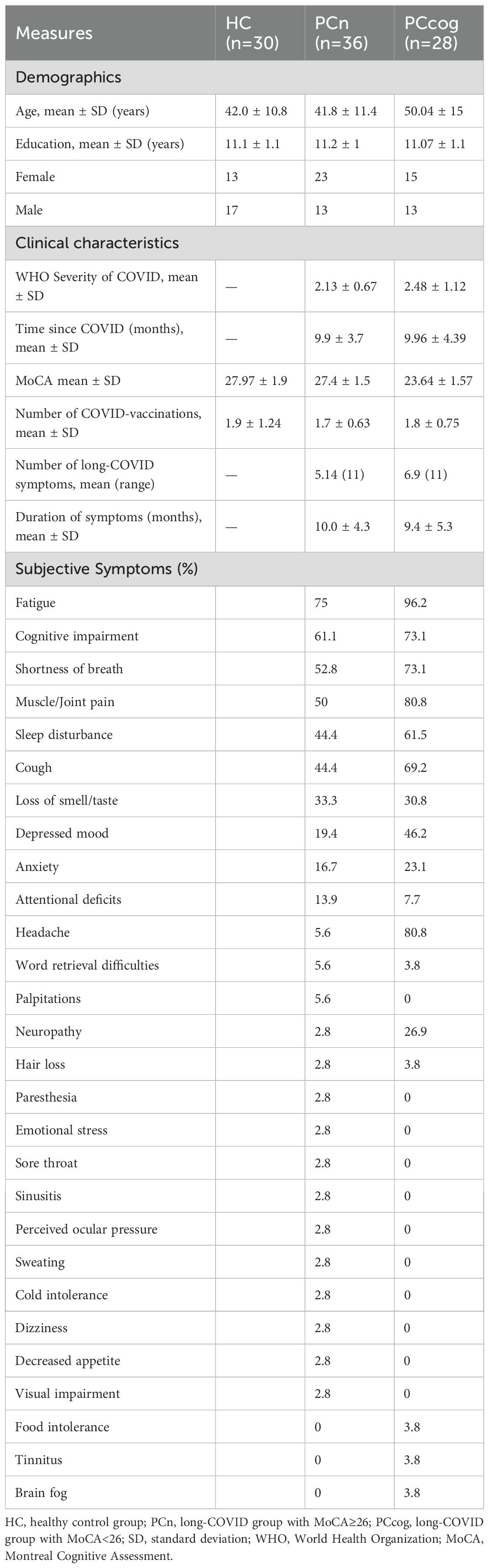
Table 1. Demographic data of the HC, PCn and PCcog groups.
Long-COVID individuals were stratified based on their cognitive performance assessed using the MoCA score (31): those with a MoCA score ≥ 26 were assigned to the cognitively unimpaired long-COVID group (PCn; n=36; mean age 41.8; SD 11.4), while participants with a MoCA score < 26 were assigned to the cognitively impaired long-COVID group (PCcog; n= 28; mean age 50.04; SD 15). A healthy control group (HC n= 30; mean age yrs 42.0; SD 10.8) was included for comparison. All participants self-identified as either male or female, and subsequent analyses were stratified accordingly by sex. To enable sex-specific analysis, each group was further subdivided by sex. Female and male participants were labeled f-PCn/m-PCn, f-PCcog/m-PCcog and f-HC/m-HC, respectively. Group distributions were as follows: the cognitively unimpaired long-COVID group (PCn) included 23 female (f-PCn) and 13 male participants (m-PCn); the cognitively impaired long-COVID group (PCcog) comprised 15 female (f-PCcog) and 13 male participants (m-PCcog); and the healthy control group (HC) consisted of 13 female (f-HC) and 17 male participants (m-HC)
The HC group was recruited via announcements of the study in the local newspaper and on social media accounts of the clinic. To ensure that participants in the HC group had not previously been infected with SARS-CoV-2 at the time of assessment, the absence of SARS-CoV-2–specific antibodies was confirmed by serological testing. Serology results were further validated using a Western blot to distinguish between antibodies arising from natural infection and those induced by vaccination.
All participants were screened via telephone to exclude those with past or current psychiatric disorders and current addiction. Additional exclusion criteria included contraindications for magnetic resonance imaging (MRI), diseases of the nervous system, a history of traumatic brain injury or loss of consciousness, unmedicated internal medical conditions and severe cognitive impairment (IQ < 80). To exclude the latter, IQ was estimated using the German Multiple Choice Vocabulary Test B (MWT-B) (32).
The study protocol was approved by the local ethics committee of the Jena University Hospital. All participants gave written informed consent. Tables 1, 2 summarize the demographic and psychometric data.
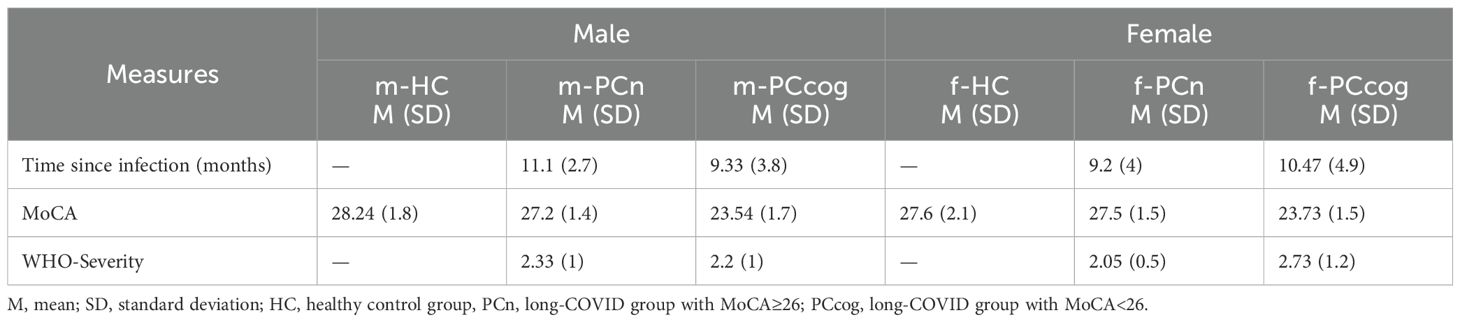
Table 2. Sex-stratified clinical and cognitive measures across cohorts.
2.2 Magnetic resonance imaging
Each participant underwent high-resolution T1-weighted MRI scans using a standard quadrature head coil and an axial 3-dimensional magnetization-prepared rapid gradient echo (MP-RAGE) sequence (TR 2400 ms, TE 2.22 ms, α 8°, 208 contiguous sagittal slices, FoV 256 mm, voxel resolution 0.8 x 0.8 x 0.8 mm, acquisition time 6:38 min) on a 3 Tesla Siemens Prisma fit (Siemens, Erlangen, Germany). All scans were checked for imaging artefacts.
2.3 Voxel-based morphometry
VBM analysis was performed using the CAT12 (Computational Anatomy Toolbox 12) developed by the Structural Brain Mapping group at University Hospital Jena, Germany, and implemented in SPM12 (Statistical Parametric Mapping, Institute of Neurology, London, UK). The T1-weighted images underwent bias-field correction to account for field homogeneity, followed by spatial normalization using the DARTEL algorithm (33). The images were segmented into white matter, gray matter and cerebrospinal fluid (34). To improve the accuracy of the segmentation process, it was extended to account for partial volume effects (35). Adaptive maximum a posteriori estimation was applied, and a hidden Markov random field model was used. To exclude artefacts at the grey-white matter boundary, an internal grey matter threshold of 0.1 was applied. After pre-processing, all scans were subjected to an automated quality control protocol. As CAT12 does not apply a fixed motion-exclusion threshold; instead, we used its image quality ratings and visual inspection to identify and exclude scans affected by motion artifacts. 2 participants from the long- COVID patient group had to be excluded from further analysis due to poor image quality at that point. The remaining images were smoothed with an 8 mm FWHM Gaussian kernel, which represents a widely used compromise between sensitivity and anatomical specificity, satisfies assumptions of Gaussian random field theory for voxel-based inference, and facilitates comparability with prior morphometry studies (36, 37).
2.4 Statistics
The statistical analysis was performed using the general linear model approach, implemented in SPM12. Groups were compared using two-sample t-tests. To account for associated variance, we included total intracranial volume (TIV), age and sex as confounding variables in the VBM analysis. As a non-parametric statistic, we applied threshold-free cluster enhancement (TFCE) with 5000 permutations (38) to all analyses and corrected for multiple comparisons via the family wise error method (FWE) at p<0.05. For atlas labelling of significant clusters we used the Neuromorphometrics Atlas (http://www.neuromorphometrics.com). We first performed this analysis for the overall groups of healthy controls (HC), long-COVID patients without cognitive symptoms according to MoCA (PCn) and long-COVID patients with cognitive impairment (PCcog). In a second step we performed these analyses separately for female and male participants, adding TIV and age as confounding variables. Complementary statistical analysis of clinical and demographic data was conducted in IBM SPSS Statistics (version 29 0.2.0.). A significance level of p≤ 0.05 was applied. The chi-square test assessed sex distribution among groups. Kruskal-Wallis tests were used to compare age, years of education and TIV. Mann-Whitney U tests examined differences in WHO severity of acute COVID-19 infection and time since infection between PCn, f-PCn, m-PCn and PCcog. f-PCcog, m-PCcog. ANCOVA was performed to assess the influence of covariates on MoCA scores.
3 Results
3.1 Clinical and demographic results
3.1.1 Overall analysis
No significant difference in sex distribution was observed between the HC, PCn and PCcog groups (χ2 (2)=2.793, p=0.247; Chi-square test). Similarly, there were no significant differences in years of education (H (2)=0.153, p=0.926) or TIV (n=92, H (2)=0,266, p=0.875; Kruskal-Wallis test). A significant group difference in age was found (H (2)=7.061, p=0.029; Kruskal-Wallis test), with PCcog participants being significantly older than those in the HC (U = 280.0, Z=-2.180, p=0.029) and PCn groups (U = 323.0, Z=-2.451, p=0.014; Mann-Whitney U test). No significant differences between the PCn and PCcog groups in WHO severity scores (U = 276.0, Z=-1.401, p=0.161) and time since infection (U = 455,5, Z=-0.242, p=0.809; Mann-Whitney U test) were found.
As expected, due to group definitions based on cognitive status, MoCA scores differed significantly between the PCcog group and the HC (U = 41.500, Z=-5.963, p=<0.001), and the PCn groups (U = 0.000, Z=-6.900, p=<0.001), while the HC and PCn groups did not differ significantly (U = 406.000, Z=-1.759, p= 0.079; Mann-Whitney U test). ANCOVA results showed that TIV, age, years of education, time since infection, and WHO severity were not significantly associated with MoCA performance. Only group affiliation, used to define cognitive status, had a significant effect on MoCA score (see Table 3).
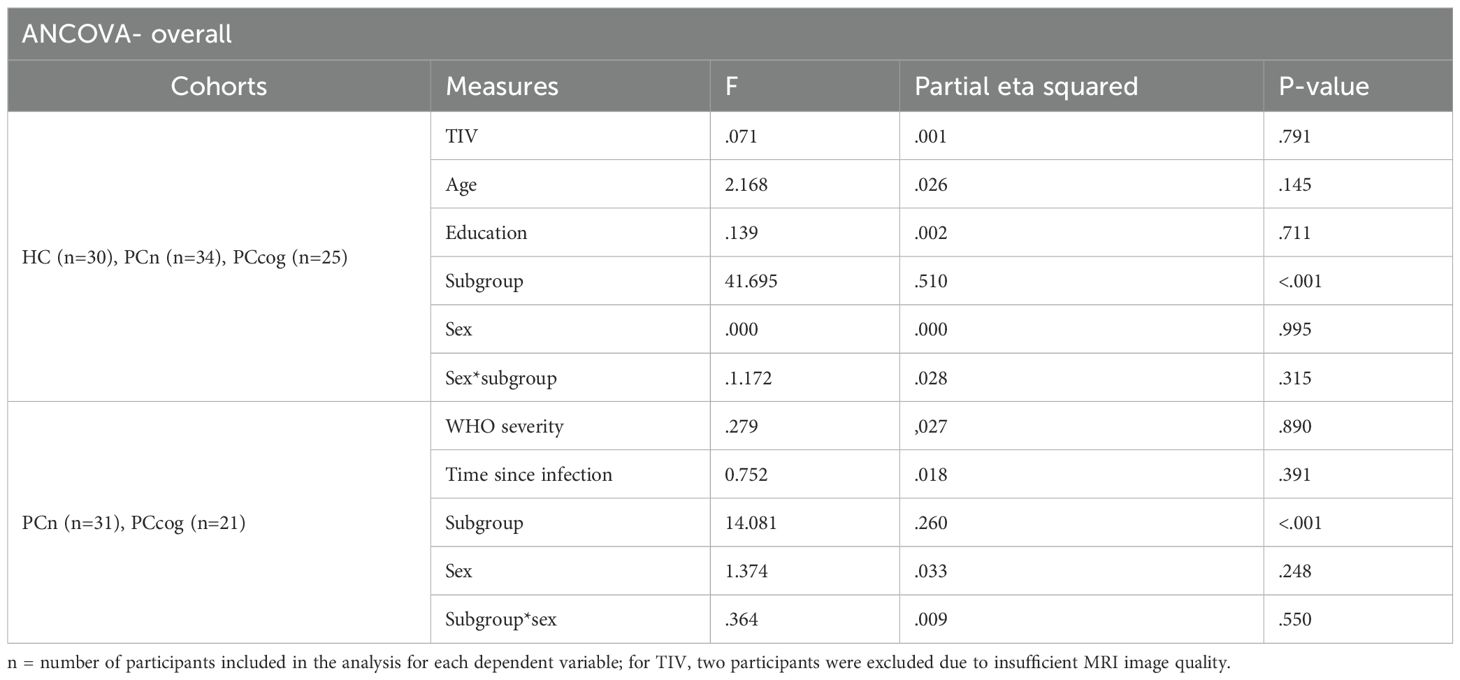
Table 3. overall ANCOVA.
3.1.2 Sex-stratified analysis
In both female and male subgroups, no significant differences were observed between f-PCn and f-PCcog and m-PCn and m-PCcog in terms of WHO severity (females: U = 79.0, Z=-2.385, p=0.114, males: U = 40.5, Z=-0.579, p=0.720) or time since infection (females: U = 158.5, Z=-0.419, p=0.680, males: U = 58.5 Z=-0.781, p=0.443; Mann-Whitney U test).
Among women, no significant group differences in years of education (H (2)=0.353, p=0,838), TIV (n=92, H (2)=0,459, p=0,795) or age (H (2)=2,004, p=0,367; Kruskal-Wallis test) were found. Similarly, no significant differences were found among men for education (H (2)=0,192, p=0,908), TIV (n=92, H (2)=0,610, p=0,737) or age (H (2)=5,421, p=0,066; Kruskal-Wallis test). Separate ANCOVAs for women and men confirmed that none of the potential confounding variables significantly influenced MoCA performance (see Table 4). As expected, group classification significantly affected MoCA scores in both subgroups, consistent with the criteria used for defining cognitive status.
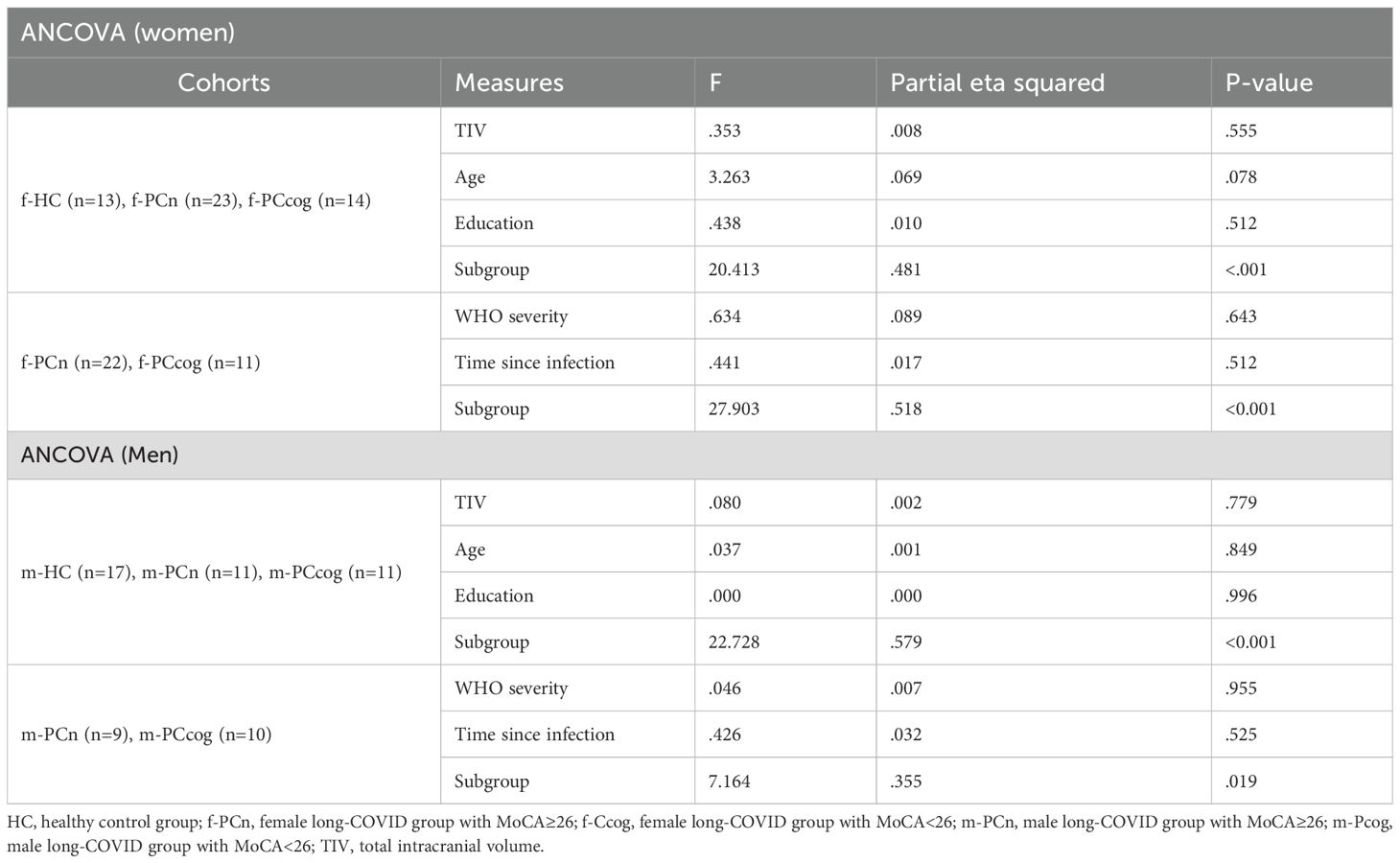
Table 4. Sex-stratified ANCOVA.
3.2 Imaging results
Overall analyses and sex-separated analyses of the HC, PCn and PCcog groups revealed several significant clusters (p<0.05, FWE-corrected) showing GMV alterations between the HC and both PC groups.
3.2.1 Overall comparison
Significant clusters exceeding a size threshold of kE>100 were identified for each comparison and are displayed in Supplementary Table 1 and Supplementary Figure 1.
3.2.2 Sex-stratified analysis
In women, no significant clusters of significant GMV differences were found for the comparisons f-PCn<f-PCcog and f-PCn>f-PCcog, whereas the remaining comparisons revealed significant clusters. In men, no significant clusters emerged for the contrasts m-HC<m-PCcog, m-HC>m-PCn, m-PCn<m-PCcog; all other comparisons yielded significant results.
An overview of all significant clusters (kE>100) is provided in Table 5, with their spatial distribution illustrated in Figure 1 (women) and Figure 2 (men).
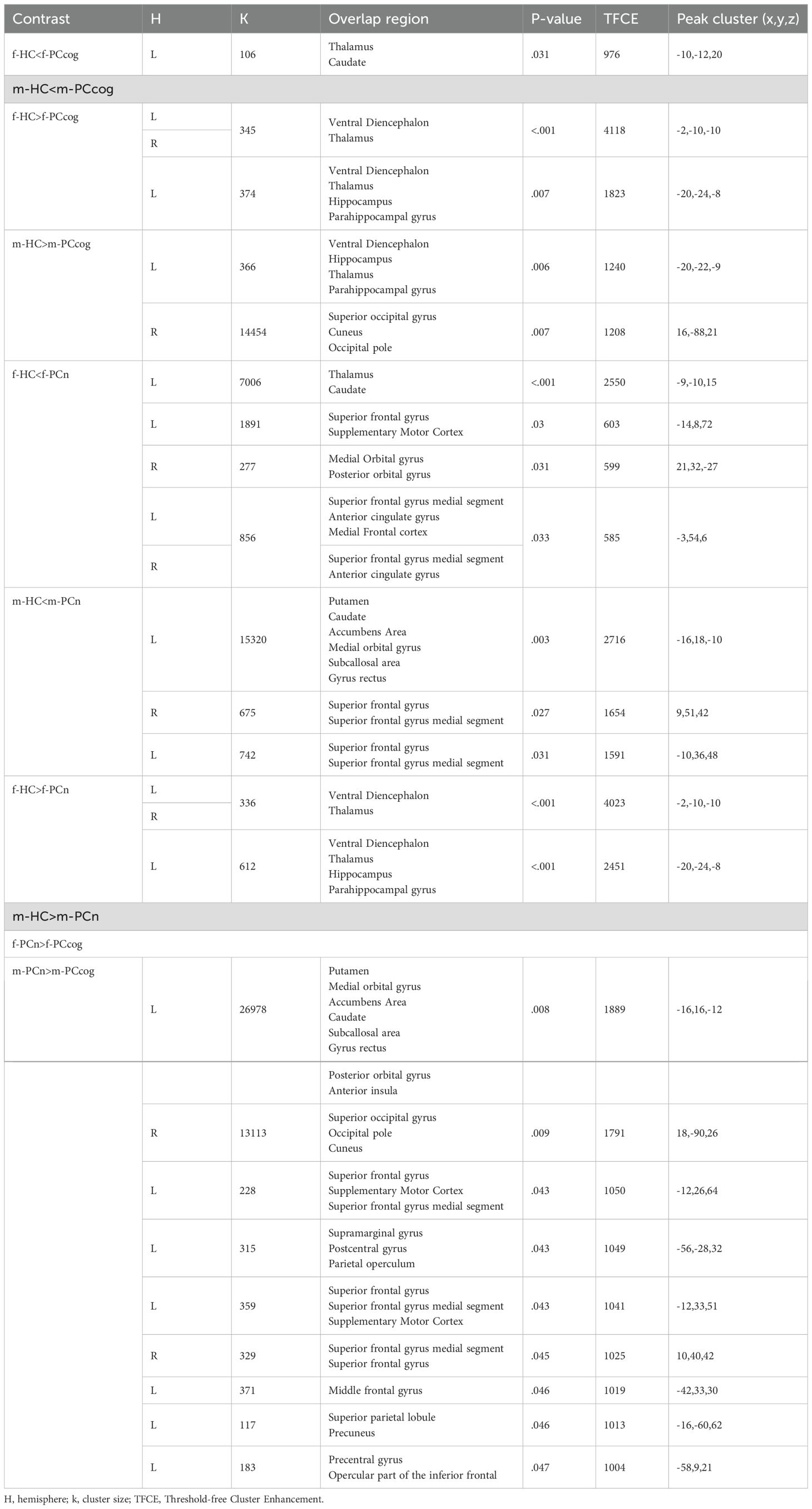
Table 5. Sex-stratified GMV differences between long-COVID cohorts and controls.
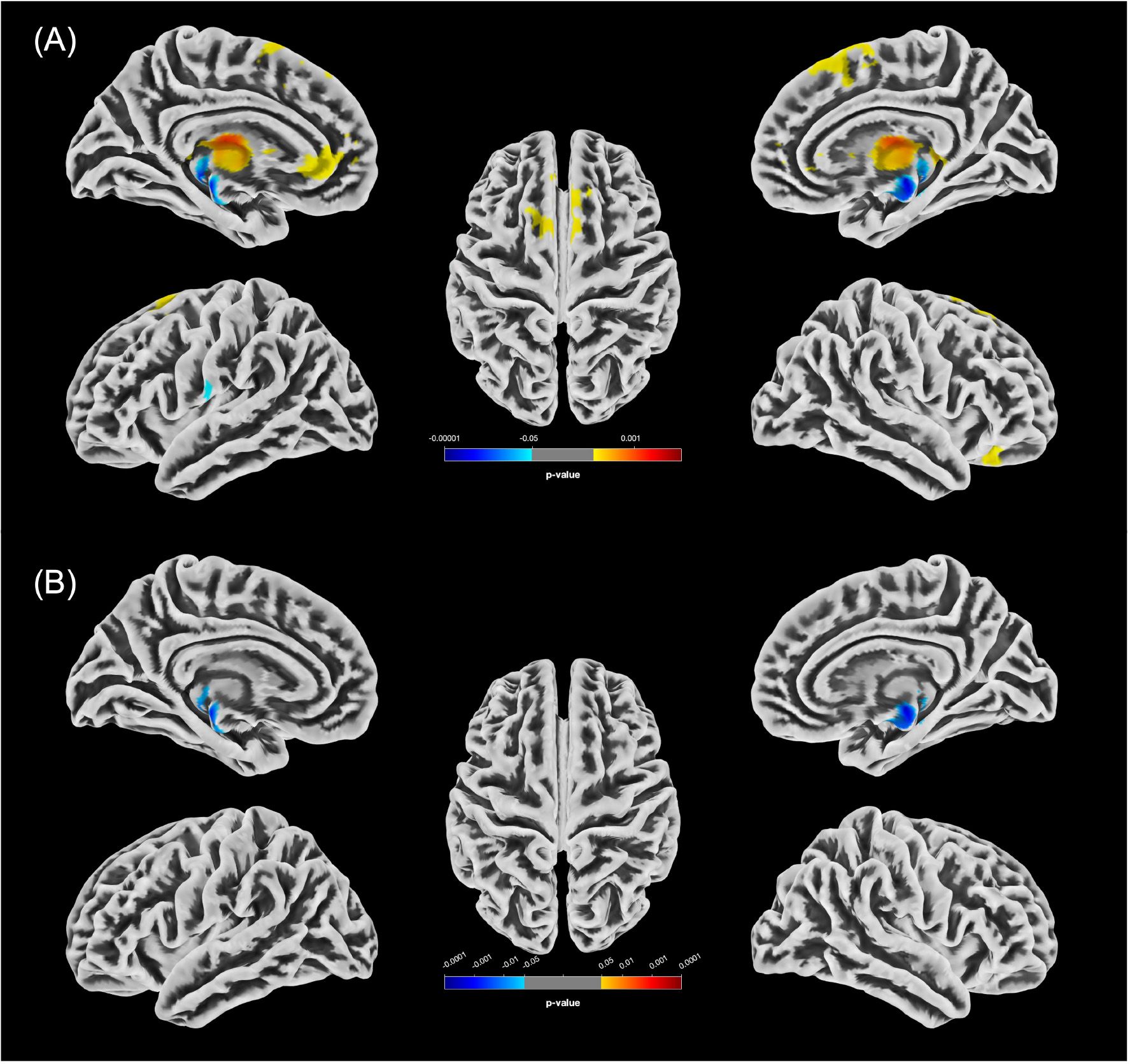
Figure 1. Statistically significant GMV differences between the female groups (p<0.05, FWE-corrected). (A) Increases in GMV in f-PCn relative to f-HC (f-HC<f-PCn) are shown in yellow/red; decreases (f-HC>f-PCn) in blue. (B) Significant clusters of reduced GMV in f-PCcog compared to f-HC are displayed in blue.
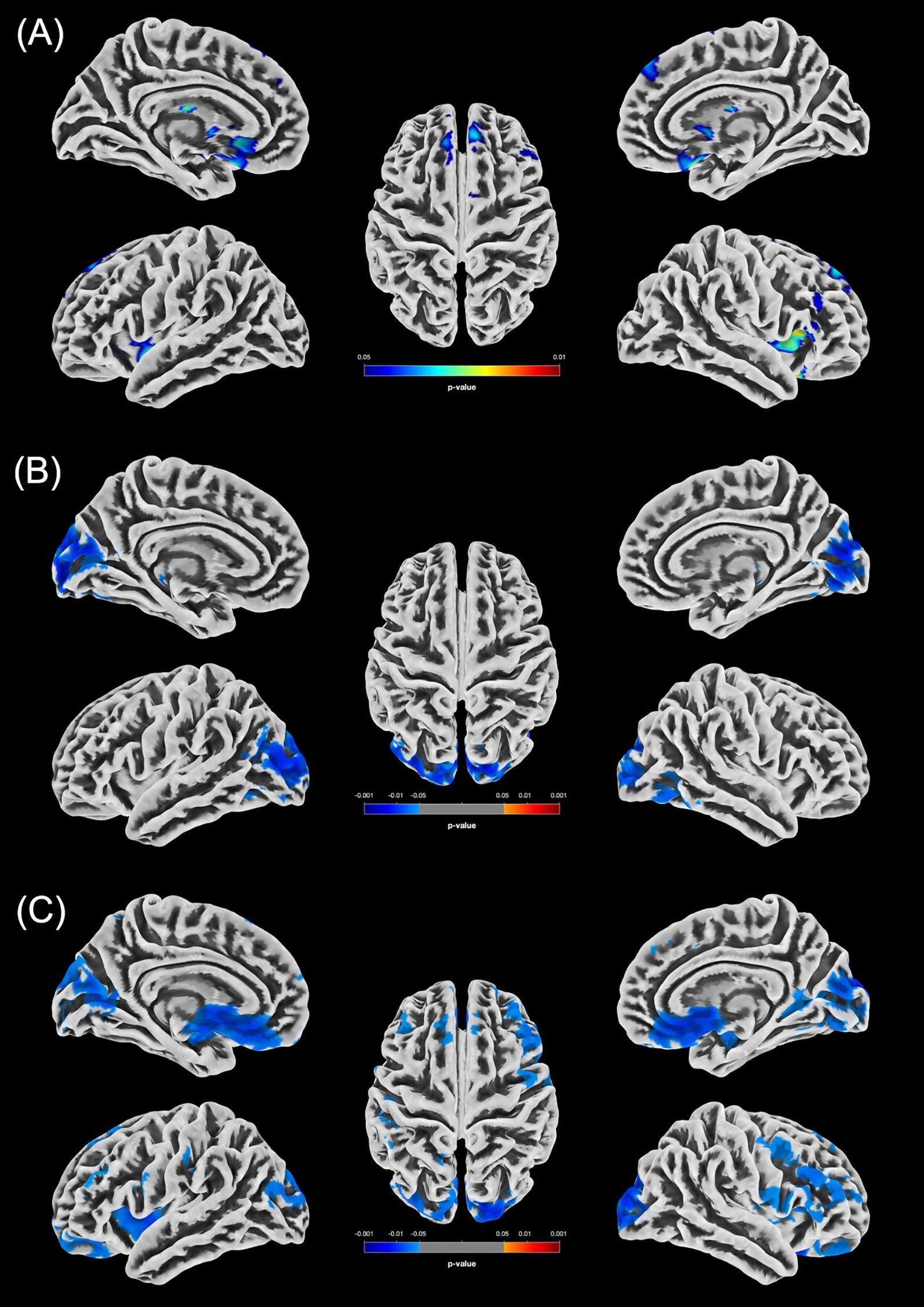
Figure 2. Statistically significant GMV differences between the male groups (p < 0.05, FWE-corrected). (A) Increased GMV in the m-PCn group compared to m-HC (m-HC < m-PCn) is displayed in blue/green. (B) Reduced GMV in m-PCcog compared to m-HC is displayed in blue. (C) Increased GMV in the m-PCn group compared to the m-PCcog group (m-PCn > m-PCcog) is displayed in blue.
4 Discussion
Our analyses revealed significant GMV alterations across multiple brain regions, comprising both shared and sex-specific patterns.
4.1 Common GMV alterations in male and female participants
In the HC>PCcog comparison, several brain regions showed overlapping GMV reductions in both sexes, indicating shared structural alterations. Both m-PCcog and f-PCcog participants exhibited GMV reductions in the left ventral diencephalon, thalamus, hippocampus, and parahippocampal gyrus. Among these, the thalamus and hippocampus are particularly critical for cognitive function (39, 40). Men additionally showed alterations in right occipital areas. These findings are consistent with previous findings on GMV alterations associated with long-COVID. Diez-Cirarda et al. reported GMV reductions in limbic areas, among others, associated with cognitive dysfunction (41). Similarly, a UK Biobank study investigating brain changes in 401 participants over a long-term follow-up period found reductions in gray matter thickness and tissue contrast in the parahippocampal gyrus and orbitofrontal cortex, along with an overall greater progression of cognitive decline in COVID-19 patients (20).
The hippocampus has likewise emerged as a region of concern in individuals recovering from SARS-CoV-2 infection. As a key structure for cognition and particularly episodic memory (42), it is critically implicated in the pathophysiology of neurodegenerative disorders, such as Alzheimer`s disease, and psychiatric conditions including major depressive disorder (43–45). In the context of long-COVID, hippocampal structural and functional alterations, potentially affecting adult neurogenesis, have been linked to memory loss and an accelerated progression of neurodegenerative processes (46, 47).
In the present study, long-COVID patients exhibited consistently reduced hippocampal GMV compared to HC, in line with findings from Capelli et al., Kamasak et al. and Invernizzi et al. (48–50). However, contrasting results have been reported in two large-scale studies that found increased hippocampal GMV (51), with Lu et al. additionally describing a positive correlation with memory impairment (Lu et al., 2020). These divergent findings underscore the complexity of structural brain alterations associated with long-COVID.
The coexistence of both increases and decreases in GMV of different brain regions may reflect a dynamic interplay between neurodegenerative processes and compensatory mechanisms, such as neuroplasticity (52, 53). Neuroinflammation, a key focus of long-COVID research, is thought to contribute to structural brain changes through cytokine-mediated disruption of the blood-brain barrier, neurovascular damage, and impaired neurogenesis (54–62). Evidence from previous coronavirus outbreaks (SARS, MERS) supports this mechanism (63, 64). These pro-inflammatory responses resemble those implicated in cancer therapy-related cognitive impairment, suggesting shared pathophysiological pathways (65, 66). Under certain conditions, however, microglial activation may promote neurogenesis, depending on cytokine profiles and concentrations (67). This dual role could help explain the heterogeneous pattern of GMV alterations observed in long-COVID.
Beyond hippocampal and cortical regions, the amygdala has been implicated in COVID-19 related neurocognitive changes (50, 68). Invernizzi et al. reported structural and functional alterations not only in the hippocampus but also the left amygdala, where reduced connectivity was shown to specifically mediate spatial working memory deficits (50). A cross-sectional study involving 75 individuals, including COVID-19 survivors with and without brain fog and healthy controls, found that both COVID-19 groups showed reduced gray matter concentrations in the left inferior temporal gyrus, left fusiform gyrus, and right orbital gyri compared to healthy controls. In addition, participants with brain fog exhibited further reductions in the bilateral caudate nuclei, right putamen/pallidum, and amygdala (68).
This highlights the role of limbic circuitry in cognitive sequalae following SARS-CoV-2 infection. Complementary longitudinal work in healthy individuals without prior SARS-CoV-2 infection linked transient volumetric increases in the amygdalae to stress- and anxiety-related processes following the COVID-19 outbreak and lockdown, with GMV gradually decreasing over time after lockdown relief (69). In our study, however, the amygdala did not emerge as a region of interest. Nevertheless, other limbic structures showed relevant alterations.
Although the overall analysis was secondary to the sex-stratified results, it revealed several regions of interest relevant to cognitive functioning, including the hippocampus, entorhinal area, posterior cingulate gyrus, angular gyrus, and planum temporale (see Supplementary Table 1 and Supplementary Figure 1).
4.2 Sex-specific GMV alterations
4.2.1 GMV alterations observed in female participants
The distribution of clusters with altered GMV in female participants compared to males in our analysis was more restricted, predominantly involving anterior frontal areas as well as limbic and diencephalic regions, including the ventral diencephalon, hippocampus and thalamus. No significant differences were found between the f-PCn and f-PCcog groups. Notably, the left thalamus consistently demonstrated GMV alterations across all statistically significant female subgroup comparisons, with both increases and decreases observed. Given its heterogenous structure and its central role in cognitive processes (70), thalamic involvement may be particularly relevant to neuropsychiatric manifestations of long-COVID. Supporting this, VBM in patients with mild cognitive impairment, unrelated to COVID-19, similarly revealed volumetric reductions in the left thalamus, along with alterations in the hippocampus and amygdala (39). This convergence underscores the thalamus as a central node whose vulnerability may extend across different conditions associated with cognitive decline.
GMV alterations specific to female participants were also detected in the anterior cingulate gyrus and medial frontal cortex.
4.2.2 GMV alterations observed in male participants
In men, the distribution of statistically significant GMV clusters was broader than in women, extending into parietal, occipital and motor areas. Additionally, the number of clusters was greater compared to women. A consistent pattern of reduced GMV emerged in the m-PCcog group, with the occipital pole, cuneus, and superior occipital gyrus repeatedly showing GMV reductions. While the occipital pole and superior occipital gyrus are not primarily associated with cognitive functions, the cuneus plays a role in working memory, which is crucial for performance in complex cognitive tasks (71, 72) and is often implicated in early stages of neurodegenerative and psychiatric conditions (73, 74). In addition to the previously mentioned regions, the putamen exhibited notable GMV increases in the m-PCn group compared to both the m-HC and the m-PCcog groups, reinforcing the notion of sex-specific structural alterations in long-COVID. The putamen, a key component of the basal ganglia along with the caudate nucleus and pallidum (75), plays a central role in motor control, learning, behavior regulation, and emotional processing (76) and has increasingly been implicated in the context of long-COVID. A systematic review highlighted the frontal, temporal, and parietal lobes, as well as the cerebellum, hippocampus, amygdala, and basal ganglia as key regions affected in post-COVID conditions (77). In line with this, Vakani et al. found that persistent COVID-19 symptoms were significantly associated with smaller putamen volume, impaired cognitive performance and poorer mental health and sleep quality (78). Heine et al. reported shape deformations and decreased GMV in the left thalamus, putamen and pallidum in post-COVID fatigue patients (79). These findings converge with our results and underscore the relevance of basal ganglia alterations in long-COVID. Moreover, recent work from our group linked changes in corticostriatal connectivity to cognitive impairment in long-COVID patients (24), potentially mediated by ACE2 receptor expression in the basal ganglia, which facilitates SARS-CoV-2 entry (24).
The broader distribution of GMV alterations observed in cognitively impaired men compared to women may be linked to sex-specific immune response patterns. Men are more prone to excessive inflammatory responses, including cytokine storms, which are associated with poor COVID-19 outcomes and may contribute to neural damage (8). However, since our cohort primarily included individuals with mild disease courses, this mechanism alone is unlikely to fully account for the observed sex-differences, particularly considering the absence of significant GMV alterations between m-HC and m-PCn participants. Instead, the findings likely reflect underlying biological factors such as hormonal influences and immune regulatory differences. Females generally exhibit stronger innate immune responses, greater resistance to viral infections, and lower levels of inflammatory mediators (8, 16, 80, 81), potentially mitigating neuroinflammation and limiting GMV changes in long-COVID. Importantly, the differing spatial distribution of GMV alterations between sexes was not accompanied by measurable differences in cognitive performance, as indicated by comparable MoCA scores across male and female participants.
Direct comparisons with prior work are limited, as few studies have examined sex-specific structural brain changes in long-COVID. One VBM study in men reported right hippocampal volume reductions shortly after Omicron infection, but was limited by the absence of a control group, small sample size, and the fact that cognitive impairment was not addressed in the study (82).
In summary, our findings demonstrate sex-specific GMV alterations in individuals with long-COVID, with men showing a broader distribution of affected regions despite the higher reported prevalence of long-COVID in women. Both sexes exhibited changes in brain areas relevant to cognition, with notable overlap between groups. However, given the cross-sectional design and limited sample size, the generalizability and temporal stability of these findings remain uncertain. Furthermore, the MoCA may have limited sensitivity in younger participants, potentially affecting the accuracy of cognitive assessment (83). Longitudinal, sex-stratified studies are needed to clarify the long-term neuropsychiatric effects of SARS-CoV-2. Also, our future work will focus on extending our analyses to larger and more heterogeneous samples through national and international collaborations, thereby improving the generalizability and robustness of our findings.
Data availability statement
The datasets presented in this article are not readily available because informed consent and ethics approval for anonymous public use were not obtained and cannot be secured retrospectively. Pseudonymized data may be provided upon reasonable request. Requests to access the datasets should be directed to Bianca Besteher (bianca.besteher@med.uni-jena.de). All image and statistical analyses were conducted using the openly available CAT12 toolbox and IBM SPSS Statistics; no custom code beyond these packages was developed.
Ethics statement
The studies involving humans were approved by Ethik-Kommission der Friedrich-Schiller-Universität Jena Bachstraße 18/Gebäude 1 07740 Jena. The studies were conducted in accordance with the local legislation and institutional requirements. The participants provided their written informed consent to participate in this study.
Author contributions
AT: Investigation, Formal Analysis, Writing – original draft, Data curation. MF: Data curation, Writing – review & editing, Investigation. TR: Writing – review & editing, Data curation. JB: Investigation, Writing – review & editing, Data curation. MT: Writing – review & editing, Data curation. DG: Writing – review & editing, Formal Analysis, Data curation. KF: Writing – review & editing, Investigation. PR: Data curation, Writing – review & editing. AS: Writing – review & editing, Investigation. SV: Writing – review & editing, Investigation. ID: Writing – review & editing. CG: Writing – review & editing, Data curation, Formal Analysis. MW: Methodology, Writing – review & editing, Conceptualization. BB: Project administration, Writing – review & editing, Conceptualization, Methodology, Supervision.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. The project was in part funded by the German Center for Mental Health (BB, MW) and the Interdisciplinary Center for Clinical Research of JUH (Advanced Clinician Scientist Program, ACS001, BB).
Acknowledgments
We are grateful to Ines Krumbein for overseeing MRI measurements and to Lara Krickow, Eva-Maria Dommaschk, Maximilian Vollmer and Marlene Müller for the collection of MRI data. ChatGPT (version GTP-4) was used exclusively for language editing. The content, reasoning, and analysis are entirely the authors’ own.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that Generative AI was used in the creation of this manuscript. ChatGPT (version GTP-4) was used exclusively for language editing. The content, reasoning, and analysis are entirely the authors’ own.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary Material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fpsyt.2025.1653295/full#supplementary-material
References
1. COVID-19 rapid guideline: managing the long-term effects of COVID-19 (London: National Institute for Health and Care Excellence (NICE)) (2024). (NICE Guideline, No. 188.).
2. Davis HE, McCorkell L, Vogel JM, and Topol EJ. Long COVID: major findings, mechanisms and recommendations. Nat Rev Microbiol. (2023) 21:133–46. doi: 10.1038/s41579-022-00846-2
3. Di Gennaro F, Belati A, Tulone O, Diella L, Fiore Bavaro D, Bonica R, et al. Incidence of long COVID-19 in people with previous SARS-Cov2 infection: a systematic review and meta-analysis of 120,970 patients. Intern Emerg Med. (2023) 18:1573–81. doi: 10.1007/s11739-022-03164-w
4. Natarajan A, Shetty A, Delanerolle G, Zeng Y, Zhang Y, Raymont V, et al. A systematic review and meta-analysis of long COVID symptoms. Syst Rev. (2023) 12:88. doi: 10.1186/s13643-023-02250-0
5. Hampshire A, Azor A, Atchison C, Trender W, Hellyer PJ, Giunchiglia V, et al. Cognition and memory after covid-19 in a large community sample. N Engl J Med. (2024) 390:806–18. doi: 10.1056/NEJMoa2311330
6. Evans RA, McAuley H, Harrison EM, Shikotra A, Singapuri A, Sereno M, et al. Physical, cognitive, and mental health impacts of COVID-19 after hospitalization (PHOSP-COVID): a UK multicenter, prospective cohort study. Lancet Respir Med. (2021) 9:1275–87. doi: 10.1016/S2213-2600(21)00383-0
7. Sykes DL, Holdsworth L, Jawad N, Gunasekera P, Morice AH, and Crooks MG. Post-COVID-19 symptom burden: what is long-COVID and how should we manage it? Lung. (2021) 199:113–9. doi: 10.1183/13993003.congress-2021.OA4189
8. Haitao T, Vermunt JV, Abeykoon J, Ghamrawi R, Gunaratne M, Jayachandran M, et al. COVID-19 and sex differences: mechanisms and biomarkers. Mayo Clin Proc. (2020) 95:2189–203. doi: 10.1016/j.mayocp.2020.07.024
9. Williamson EJ, Walker AJ, Bhaskaran K, Bacon S, Bates C, Morton CE, et al. Factors associated with COVID-19-related death using OpenSAFELY. Nature. (2020) 584:430–6. doi: 10.1038/s41586-020-2521-4
10. Zettersten E, Engerström L, Bell M, Jäderling G, Mårtensson J, Block L, et al. Long-term outcome after intensive care for COVID-19: differences between men and women-a nationwide cohort study. Crit Care. (2021) 25:86. doi: 10.1186/s13054-021-03511-x
11. The L. The gendered dimensions of COVID-19. Lancet. (2020) 395:1168. doi: 10.1016/S0140-6736(20)30823-0
12. Munblit D, Bobkova P, Spiridonova E, Shikhaleva A, Gamirova A, Blyuss O, et al. Incidence and risk factors for persistent symptoms in adults previously hospitalized for COVID-19. Clin Exp Allergy. (2021) 51:1107–20. doi: 10.1111/cea.13997
13. Ceban F, Ling S, Lui LMW, Lee Y, Gill H, Teopiz KM, et al. Fatigue and cognitive impairment in Post-COVID-19 Syndrome: A systematic review and meta-analysis. Brain Behav Immun. (2022) 101:93–135. doi: 10.1016/j.bbi.2021.12.020
14. Sylvester SV, Rusu R, Chan B, Bellows M, O'Keefe C, and Nicholson S. Sex differences in sequelae from COVID-19 infection and in long COVID syndrome: a review. Curr Med Res Opin. (2022) 38:1391–9. doi: 10.1080/03007995.2022.2081454
15. Bai F, Tomasoni D, Falcinella C, Barbanotti D, Castoldi R, Mulè G, et al. Female gender is associated with long COVID syndrome: a prospective cohort study. Clin Microbiol Infect. (2022) 28:611.e9–.e16. doi: 10.1016/j.cmi.2021.11.002
16. Ortona E and Malorni W. Long COVID: to investigate immunological mechanisms and sex/gender related aspects as fundamental steps for tailored therapy. Eur Respir J. (2022) 59(2):2102245. doi: 10.1183/13993003.02245-2021
17. Kitselman AK, Bédard-Matteau J, Rousseau S, Tabrizchi R, and Daneshtalab N. Sex differences in vascular endothelial function related to acute and long COVID-19. Vascul Pharmacol. (2024) 154:107250. doi: 10.1016/j.vph.2023.107250
18. Patrono A, Invernizzi A, Placidi D, Cagna G, Calza S, Oppini M, et al. Impact of COVID-19-related social isolation on behavioral outcomes in young adults residing in northern Italy. Int J Environ Res Public Health. (2022) 19(24):16496. doi: 10.3390/ijerph192416496
19. Benedetti F, Palladini M, Paolini M, Melloni E, Vai B, De Lorenzo R, et al. Brain correlates of depression, post-traumatic distress, and inflammatory biomarkers in COVID-19 survivors: A multimodal magnetic resonance imaging study. Brain Behav Immun Health. (2021) 18:100387. doi: 10.1016/j.bbih.2021.100387
20. Douaud G, Lee S, Alfaro-Almagro F, Arthofer C, Wang C, McCarthy P, et al. SARS-CoV-2 is associated with changes in brain structure in UK Biobank. Nature. (2022) 604:697–707. doi: 10.1038/s41586-022-04569-5
21. Lu Y, Li X, Geng D, Mei N, Wu PY, Huang CC, et al. Cerebral micro-structural changes in COVID-19 patients - an MRI-based 3-month follow-up study. EClinicalMedicine. (2020) 25:100484. doi: 10.1016/j.eclinm.2020.100484
22. Pelizzari L, Cazzoli M, Lipari S, Lagana MM, Cabinio M, Isernia S, et al. Mid-term MRI evaluation reveals microstructural white matter alterations in COVID-19 fully recovered subjects with anosmia presentation. Ther Adv Neurol Disord. (2022) 15:17562864221111995. doi: 10.1177/17562864221111995
23. Besteher B, Machnik M, Troll M, Toepffer A, Zerekidze A, Rocktaschel T, et al. Larger gray matter volumes in neuropsychiatric long-COVID syndrome. Psychiatry Res. (2022) 317:114836. doi: 10.1016/j.psychres.2022.114836
24. Troll M, Li M, Chand T, Machnik M, Rocktäschel T, Toepffer A, et al. Altered corticostriatal connectivity in long-COVID patients is associated with cognitive impairment. Psychol Med. (2025) 55:e49. doi: 10.1017/S0033291725000054
25. Whitwell JL. Voxel-based morphometry: an automated technique for assessing structural changes in the brain. J Neurosci. (2009) 29:9661–4. doi: 10.1523/JNEUROSCI.2160-09.2009
26. Goto M, Abe O, Hagiwara A, Fujita S, Kamagata K, Hori M, et al. Advantages of using both voxel- and surface-based morphometry in cortical morphology analysis: A review of various applications. Magn Reson Med Sci. (2022) 21:41–57. doi: 10.2463/mrms.rev.2021-0096
27. Venkatesan P. NICE guideline on long COVID. Lancet Respir Med. (2021) 9:129. doi: 10.1016/S2213-2600(21)00031-X
28. AWMF. S1-leitlinie: post-COVID/long-COVID2021 (2025). Available online at: https://register.awmf.org/de/leitlinien/detail/020-027 (Accessed September 15, 2025).
29. Peter RS, Nieters A, Kräusslich HG, Brockmann SO, Göpel S, Kindle G, et al. Post-acute sequelae of covid-19 six to 12 months after infection: population based study. Bmj. (2022) 379:e071050. doi: 10.1136/bmj-2022-071050
30. Marshall JC, Murthy S, Diaz J, Adhikari NK, Angus DC, Arabi YM, et al. Characterization WHOWGotC, Management of C-i. A minimal common outcome measure set for COVID-19 clinical research. Lancet Infect Dis. (2020) 20:e192–e7. doi: 10.1016/S1473-3099(20)30483-7
31. Nasreddine ZS, Phillips NA, Bédirian V, Charbonneau S, Whitehead V, Collin I, et al. The Montreal Cognitive Assessment, MoCA: a brief screening tool for mild cognitive impairment. J Am Geriatr Soc. (2005) 53:695–9. doi: 10.1111/j.1532-5415.2005.53221.x
33. Ashburner J. A fast diffeomorphic image registration algorithm. Neuroimage. (2007) 38:95–113. doi: 10.1016/j.neuroimage.2007.07.007
34. Ashburner J and Friston KJ. Unified segmentation. Neuroimage. (2005) 26:839–51. doi: 10.1016/j.neuroimage.2005.02.018
35. Tohka J, Zijdenbos A, and Evans A. Fast and robust parameter estimation for statistical partial volume models in brain MRI. Neuroimage. (2004) 23:84–97. doi: 10.1016/j.neuroimage.2004.05.007
36. Shen S and Sterr A. Is DARTEL-based voxel-based morphometry affected by width of smoothing kernel and group size? A study using simulated atrophy. J Magn Reson Imaging. (2013) 37:1468–75. doi: 10.1002/jmri.23927
37. Michael AM, Evans E, and Moore GJ. Influence of group on individual subject maps in SPM voxel based morphometry. Front Neurosci. (2016) 10:522. doi: 10.3389/fnins.2016.00522
38. Spisak T, Spisak Z, Zunhammer M, Bingel U, Smith S, Nichols T, et al. Probabilistic TFCE: A generalized combination of cluster size and voxel intensity to increase statistical power. Neuroimage. (2019) 185:12–26. doi: 10.1016/j.neuroimage.2018.09.078
39. Wang J, Liang X, Lu J, Zhang W, Chen Q, Li X, et al. Cortical and subcortical gray matter abnormalities in mild cognitive impairment. Neuroscience. (2024) 557:81–8. doi: 10.1016/j.neuroscience.2024.07.036
40. Zhang J, Liu Y, Lan K, Huang X, He Y, Yang F, et al. Gray matter atrophy in amnestic mild cognitive impairment: A voxel-based meta-analysis. Front Aging Neurosci. (2021) 13:627919. doi: 10.3389/fnagi.2021.627919
41. Diez-Cirarda M, Yus M, Gomez-Ruiz N, Polidura C, Gil-Martinez L, Delgado-Alonso C, et al. Multimodal neuroimaging in post-COVID syndrome and correlation with cognition. Brain. (2022) 146(5):2142–52. doi: 10.1093/brain/awac384
42. Lisman J, Buzsáki G, Eichenbaum H, Nadel L, Ranganath C, and Redish AD. Viewpoints: how the hippocampus contributes to memory, navigation and cognition. Nat Neurosci. (2017) 20:1434–47. doi: 10.1038/nn.4661
43. Sheline YI, Wang PW, Gado MH, Csernansky JG, and Vannier MW. Hippocampal atrophy in recurrent major depression. Proc Natl Acad Sci U S A. (1996) 93:3908–13. doi: 10.1073/pnas.93.9.3908
44. Minkova L, Habich A, Peter J, Kaller CP, Eickhoff SB, and Klöppel S. Gray matter asymmetries in aging and neurodegeneration: A review and meta-analysis. Hum Brain Mapp. (2017) 38:5890–904. doi: 10.1002/hbm.23772
45. Zackova L, Jani M, Brazdil M, Nikolova YS, and Mareckova K. Cognitive impairment and depression: Meta-analysis of structural magnetic resonance imaging studies. NeuroImage Clin. (2021) 32:102830. doi: 10.1016/j.nicl.2021.102830
46. Nouraeinejad A. The functional and structural changes in the hippocampus of COVID-19 patients. Acta Neurol Belg. (2023) 123:1247–56. doi: 10.1007/s13760-023-02291-1
47. Saikarthik J, Saraswathi I, Padhi BK, Shamim MA, Alzerwi N, Alarifi A, et al. Structural and functional neuroimaging of hippocampus to study adult neurogenesis in long COVID-19 patients with neuropsychiatric symptoms: a scoping review. PeerJ. (2025) 13:e19575. doi: 10.7717/peerj.19575
48. Capelli S, Arrigoni A, Napolitano A, Pezzetti G, Remuzzi A, Zangari R, et al. MRI evidence of gray matter loss in COVID-19 patients with cognitive and olfactory disorders. Ann Clin Transl Neurol. (2024) 11:2457–72. doi: 10.1002/acn3.52164
49. Kamasak B, Ulcay T, Nisari M, Gorgulu O, Akca V, Alpaslan M, et al. Effects of COVID-19 on brain and cerebellum: a voxel based morphometrical analysis. Bratisl Lek Listy. (2023) 124(6):442–8. doi: 10.4149/BLL_2023_068
50. Invernizzi A, Renzetti S, van Thriel C, Rechtman E, Patrono A, Ambrosi C, et al. COVID-19 related cognitive, structural and functional brain changes among Italian adolescents and young adults: a multimodal longitudinal case-control study. Transl Psychiatry. (2024) 14:402. doi: 10.1038/s41398-024-03108-2
51. Guo Z, Sun S, Xiao S, Chen G, Chen P, Yang Z, et al. COVID-19 is associated with changes in brain function and structure: A multimodal meta-analysis of neuroimaging studies. Neurosci Biobehav Rev. (2024) 164:105792. doi: 10.1016/j.neubiorev.2024.105792
52. Gulyaeva NV. Molecular mechanisms of neuroplasticity: an expanding universe. Biochem (Mosc). (2017) 82:237–42. doi: 10.1134/S0006297917030014
53. Konorski J. Conditioned reflexes and neuron organization. (New York: Hafner Publishing Company) (1948). 277 p.
54. Lyra ESNM, Barros-Aragão FGQ, De Felice FG, and Ferreira ST. Inflammation at the crossroads of COVID-19, cognitive deficits and depression. Neuropharmacology. (2022) 209:109023. doi: 10.1016/j.neuropharm.2022.109023
55. Schou TM, Joca S, Wegener G, and Bay-Richter C. Psychiatric and neuropsychiatric sequelae of COVID-19 - A systematic review. Brain Behav Immun. (2021) 97:328–48. doi: 10.1016/j.bbi.2021.07.018
56. Mazza MG, Palladini M, De Lorenzo R, Magnaghi C, Poletti S, Furlan R, et al. Persistent psychopathology and neurocognitive impairment in COVID-19 survivors: Effect of inflammatory biomarkers at three-month follow-up. Brain Behav Immun. (2021) 94:138–47. doi: 10.1016/j.bbi.2021.02.021
57. Jarczak D and Nierhaus A. Cytokine storm-definition, causes, and implications. Int J Mol Sci. (2022) 23(19):11740. doi: 10.3390/ijms231911740
58. Zanza C, Romenskaya T, Manetti AC, Franceschi F, La Russa R, Bertozzi G, et al. Cytokine storm in COVID-19: immunopathogenesis and therapy. Med (Kaunas). (2022) 58(2):144. doi: 10.3390/medicina58020144
59. Yang RC, Huang K, Zhang HP, Li L, Zhang YF, Tan C, et al. SARS-CoV-2 productively infects human brain microvascular endothelial cells. J Neuroinflammation. (2022) 19:149. doi: 10.1186/s12974-022-02514-x
60. Vanderheiden A and Klein RS. Neuroinflammation and COVID-19. Curr Opin Neurobiol. (2022) 76:102608. doi: 10.1016/j.conb.2022.102608
61. Chesnokova V, Pechnick RN, and Wawrowsky K. Chronic peripheral inflammation, hippocampal neurogenesis, and behavior. Brain Behav Immun. (2016) 58:1–8. doi: 10.1016/j.bbi.2016.01.017
62. Borsini A, Zunszain PA, Thuret S, and Pariante CM. The role of inflammatory cytokines as key modulators of neurogenesis. Trends Neurosci. (2015) 38:145–57. doi: 10.1016/j.tins.2014.12.006
63. Sheng B, Cheng SK, Lau KK, Li HL, and Chan EL. The effects of disease severity, use of corticosteroids and social factors on neuropsychiatric complaints in severe acute respiratory syndrome (SARS) patients at acute and convalescent phases. Eur Psychiatry. (2005) 20:236–42. doi: 10.1016/j.eurpsy.2004.06.023
64. Rogers JP, Chesney E, Oliver D, Pollak TA, McGuire P, Fusar-Poli P, et al. Psychiatric and neuropsychiatric presentations associated with severe coronavirus infections: a systematic review and meta-analysis with comparison to the COVID-19 pandemic. Lancet Psychiatry. (2020) 7:611–27. doi: 10.1016/S2215-0366(20)30203-0
65. Fernández-Castañeda A, Lu P, Geraghty AC, Song E, Lee MH, Wood J, et al. Mild respiratory COVID can cause multi-lineage neural cell and myelin dysregulation. Cell. (2022) 185:2452–68.e16. doi: 10.1016/j.cell.2022.06.008
66. Theoharides TC, Cholevas C, Polyzoidis K, and Politis A. Long-COVID syndrome-associated brain fog and chemofog: Luteolin to the rescue. Biofactors. (2021) 47:232–41. doi: 10.1002/biof.1726
67. Ekdahl CT, Kokaia Z, and Lindvall O. Brain inflammation and adult neurogenesis: the dual role of microglia. Neuroscience. (2009) 158:1021–9. doi: 10.1016/j.neuroscience.2008.06.052
68. Samanci B, Ay U, Gezegen H, Yörük SS, Medetalibeyoğlu A, Kurt E, et al. Persistent neurocognitive deficits in long COVID: Evidence of structural changes and network abnormalities following mild infection. Cortex. (2025) 187:98–110. doi: 10.1016/j.cortex.2025.04.004
69. Salomon T, Cohen A, Barazany D, Ben-Zvi G, Botvinik-Nezer R, Gera R, et al. Brain volumetric changes in the general population following the COVID-19 outbreak and lockdown. Neuroimage. (2021) 239:118311. doi: 10.1016/j.neuroimage.2021.118311
70. Shine JM, Lewis LD, Garrett DD, and Hwang K. The impact of the human thalamus on brain-wide information processing. Nat Rev Neurosci. (2023) 24:416–30. doi: 10.1038/s41583-023-00701-0
71. Owens MM, Duda B, Sweet LH, and MacKillop J. Distinct functional and structural neural underpinnings of working memory. Neuroimage. (2018) 174:463–71. doi: 10.1016/j.neuroimage.2018.03.022
73. Kirova AM, Bays RB, and Lagalwar S. Working memory and executive function decline across normal aging, mild cognitive impairment, and Alzheimer's disease. BioMed Res Int. (2015) 2015:748212. doi: 10.1155/2015/748212
74. Nikolin S, Tan YY, Schwaab A, Moffa A, Loo CK, and Martin D. An investigation of working memory deficits in depression using the n-back task: A systematic review and meta-analysis. J Affect Disord. (2021) 284:1–8. doi: 10.1016/j.jad.2021.01.084
75. Fazl A and Fleisher J. Anatomy, physiology, and clinical syndromes of the basal ganglia: A brief review. Semin Pediatr Neurol. (2018) 25:2–9. doi: 10.1016/j.spen.2017.12.005
76. Florio TM, Scarnati E, Rosa I, Di Censo D, Ranieri B, Cimini A, et al. The Basal Ganglia: More than just a switching device. CNS Neurosci Ther. (2018) 24:677–84. doi: 10.1111/cns.12987
77. Mohammadi S and Ghaderi S. Post-COVID-19 conditions: a systematic review on advanced magnetic resonance neuroimaging findings. Neurol Sci. (2024) 45:1815–33. doi: 10.1007/s10072-024-07427-6
78. Vakani K, Norbury R, Vanova M, Ratto M, Parton A, Antonova E, et al. Cognitive function and brain structure in COVID-19 survivors: The role of persistent symptoms. Behav Brain Res. (2025) 476:115283. doi: 10.1016/j.bbr.2024.115283
79. Heine J, Schwichtenberg K, Hartung TJ, Rekers S, Chien C, Boesl F, et al. Structural brain changes in patients with post-COVID fatigue: a prospective observational study. EClinicalMedicine. (2023) 58:101874. doi: 10.1016/j.eclinm.2023.101874
80. Ferretti VV, Klersy C, Bruno R, Cutti S, and Nappi RE. Men with COVID-19 die. Women survive. Maturitas. (2022) 158:34–6. doi: 10.1016/j.maturitas.2021.11.014
81. Ciarambino T, Para O, and Giordano M. Immune system and COVID-19 by sex differences and age. Womens Health (Lond). (2021) 17:17455065211022262. doi: 10.1177/17455065211022262
82. Du Y, Zhao W, Huang S, Huang C, Li C, Chen Y, et al. Gray matter thickness and subcortical nuclear volume in men after SARS-coV-2 omicron infection. JAMA Netw Open. (2023) 6:e2345626. doi: 10.1001/jamanetworkopen.2023.45626
Keywords: long-covid, post-COVID, COVID-19, GMV, VBM, MOCA, sex-difference, cognitive impairment
Citation: Toepffer A, Früh M, Rocktäschel T, Ballez J, Troll M, Güllmar D, Finke K, Reuken PA, Stallmach A, Vonderlind S, Dunay IR, Gaser C, Walter M and Besteher B (2025) Cognition-associated gray matter volume alterations in long-COVID show sex-specific patterns. Front. Psychiatry 16:1653295. doi: 10.3389/fpsyt.2025.1653295
Received: 24 June 2025; Accepted: 08 September 2025;
Published: 06 October 2025.
Edited by:
Jack Jiaqi Zhang, Hong Kong Polytechnic University, Hong Kong SAR, ChinaReviewed by:
Azzurra Invernizzi, Icahn School of Medicine at Mount Sinai, United StatesEzekiel Gonzalez-Fernandez, University of Mississippi Medical Center, United States
Copyright © 2025 Toepffer, Früh, Rocktäschel, Ballez, Troll, Güllmar, Finke, Reuken, Stallmach, Vonderlind, Dunay, Gaser, Walter and Besteher. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Bianca Besteher, YmlhbmNhLmJlc3RlaGVyQG1lZC51bmktamVuYS5kZQ==