 Jhommara Bautista
Jhommara Bautista Camila Hidalgo-TinocoMiranda Di Capua DelgadoJuliana Viteri-Recalde
Camila Hidalgo-TinocoMiranda Di Capua DelgadoJuliana Viteri-Recalde Antonio Guerra-Guerrero
Antonio Guerra-Guerrero Andrés López-Cortés*
Andrés López-Cortés*- Cancer Research Group (CRG), Faculty of Medicine, Universidad de Las Américas, Quito, Ecuador
Anxiety and depressive disorders rank among the most prevalent psychiatric conditions worldwide, yet remission rates remain unsatisfactory despite advances in pharmacological and psychotherapeutic interventions. The gut–brain axis has emerged as a transformative framework for understanding these disorders, emphasizing bidirectional communication between the central nervous system, the enteric nervous system, the endocrine and immune systems, and the gut microbiota. Preclinical studies demonstrate that germ-free or dysbiotic states exaggerate hypothalamic–pituitary–adrenal (HPA) reactivity, remodel synaptic plasticity, and induce anxiety- and depression-like behaviors, while fecal microbiota transplantation confirms the causal influence of microbial communities. Mechanistically, neural (e.g., vagal), endocrine (e.g., cortisol), immune (e.g., cytokine), and metabolic (e.g., short-chain fatty acids, tryptophan metabolites, bile acids) pathways converge to regulate mood and stress resilience. An underappreciated yet critical dimension of this model is circadian rhythmicity. Both host endocrine cycles and microbial communities exhibit diurnal oscillations that synchronize metabolism, immune activity, and neural signaling. Disruption of these rhythms, through factors such as sleep disturbance, irregular feeding, or shift work, alters microbial diversity, dampens metabolite oscillations, destabilizes HPA regulation, and enhances neuroinflammation, thereby amplifying vulnerability to psychiatric disorders. Collectively, evidence supports a model in which anxiety and depression are systemic conditions arising from integrated neural, immune, endocrine, metabolic, and circadian dysregulation, rather than isolated brain-based pathologies. This reconceptualization positions microbial taxa and metabolites as candidate biomarkers and therapeutic targets. Precision interventions, ranging from diet and psychobiotics to fecal microbiota transplantation, chrononutrition, and immune-modulatory strategies, offer promising avenues for personalized psychiatry.
Introduction
Anxiety and depressive disorders are among the most prevalent and debilitating psychiatric conditions, collectively affecting nearly 10% of the global population and ranking as leading causes of disability worldwide (1, 2). Despite advances in pharmacological and psychotherapeutic strategies, remission rates remain disappointingly low, underscoring the need for novel mechanistic frameworks to explain disease onset, persistence, and treatment resistance (3). Over the past two decades, the gut–brain axis has emerged as one such paradigm, emphasizing the bidirectional interplay between the central nervous system (CNS), the enteric nervous system (ENS), the endocrine and immune systems, and the intestinal microbiota (4, 5) (Figure 1).
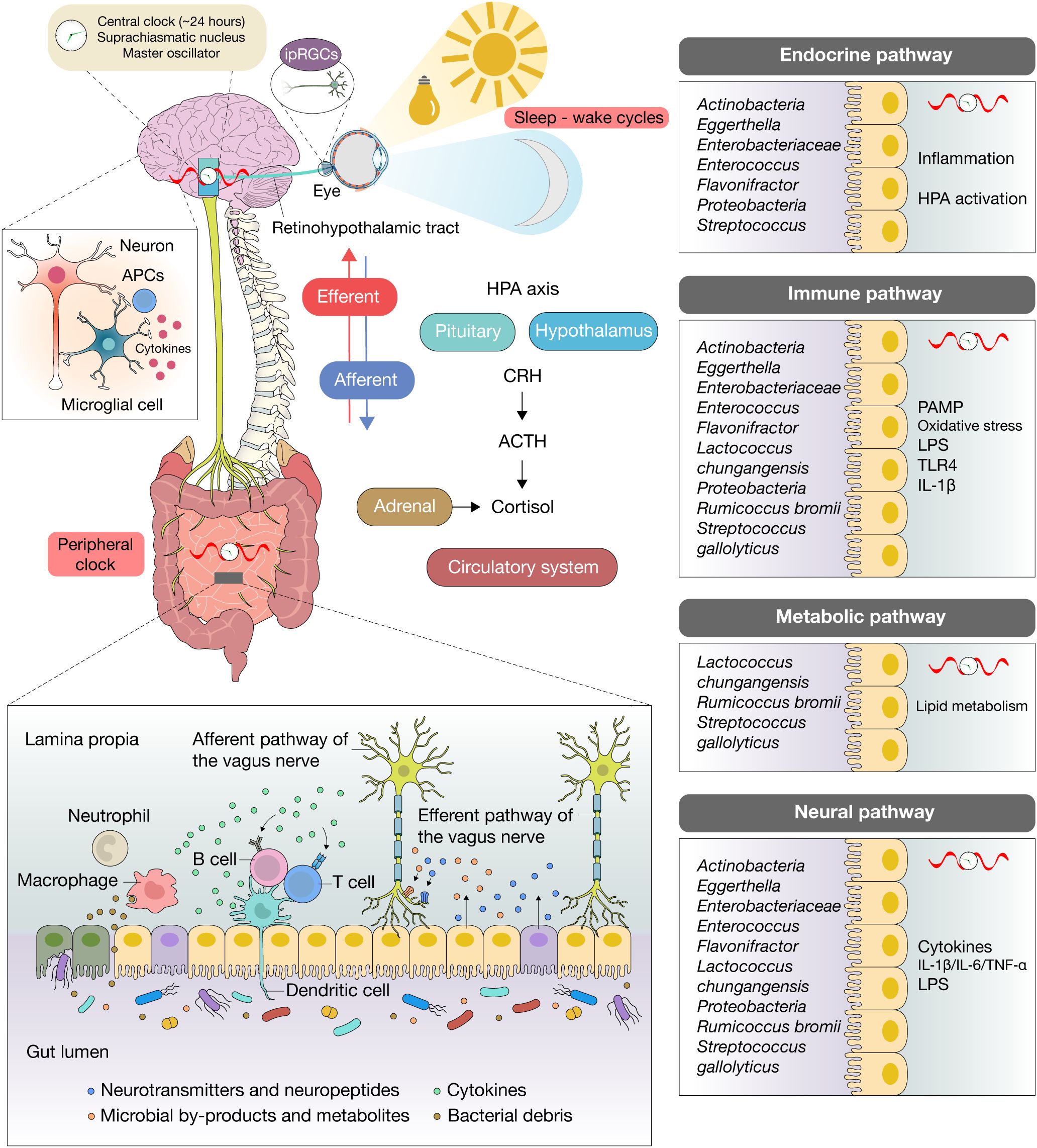
Figure 1. Gut–brain–circadian axis in anxiety and depression. Schematic focusing on risk-enriched microbial signals and their downstream effects across the neural, endocrine (HPA), immune, and metabolic arms of the gut–brain axis: barrier compromise facilitates translocation of PAMPs/LPS, activating TLR pathways and pro-inflammatory cytokines (IL-1β, IL-6, TNF-α) that drive microglial activation, synaptic remodeling, and dysfunction of mood-relevant circuits, contributing to anxiety- and depression-related phenotypes; in parallel, inflammatory signaling diverts tryptophan toward kynurenine via IDO/TDO, diminishing serotonergic tone and generating neuroactive metabolites; stress engages the HPA axis, and cortisol feedback further weakens barrier function and reshapes microbial ecology, creating a feed-forward loop; circadian rhythms and peripheral clocks gate the timing of microbial composition shifts and metabolite fluxes, as well as time-of-day windows for barrier permeability, cytokine responsiveness, vagal tone, and cortisol dynamics; circadian misalignment desynchronizes these rhythms, increases LPS leakage and inflammatory set points, and amplifies risk-associated signaling to the brain.
The human gastrointestinal tract harbors nearly 1014 microbes, outnumbering human cells and carrying a gene pool vastly exceeding the host genome (6–8). These commensals contribute to digestion, immune maturation, and host metabolism, but accumulating evidence reveals their equally critical role in shaping CNS development and behavior (4, 9). Germ-free mice display exaggerated hypothalamic–pituitary–adrenal (HPA) reactivity, anxiety-like phenotypes, and altered synaptic plasticity, which normalize upon colonization with commensals (9, 10). Converging lines of evidence, antibiotic-induced dysbiosis with behavioral rescue after recolonization, monoassociation with specific taxa (e.g., Bifidobacterium infantis) that normalizes HPA output, and metabolite supplementation that restores microglial maturation and synaptic function, support a causal role for microbiota–brain signaling without relying solely on fecal microbiota transplantation (FMT) models (11, 12).
Multiple biological pathways mediate this crosstalk. Neural signaling is exemplified by the vagus nerve, which conveys microbial cues to mood-related brain circuits; indeed, vagotomy abolishes the behavioral effects of probiotics and FMT (6, 13). Endocrine routes converge on the HPA axis, where dysbiosis disrupts glucocorticoid rhythms, elevates cortisol, and exacerbates stress sensitivity (3, 14). Immune signaling provides another conduit: lipopolysaccharide (LPS) and other pathogen-associated molecular patterns can breach a compromised gut barrier, triggering cytokine release such as IL-1β, IL-6, and TNF-α, which activate microglia and remodel synapses (9, 15, 16). Metabolic signaling represents a further layer of complexity, as microbiota-derived short-chain fatty acids (SCFAs) regulate blood–brain barrier (BBB) integrity, synaptic plasticity, and anti-inflammatory tone, while tryptophan metabolites influence serotonin biosynthesis and kynurenine pathway flux (11, 17, 18). Together, these pathways establish a framework in which microbial imbalance, or dysbiosis, contributes to psychiatric vulnerability.
Importantly, SCFAs and tryptophan metabolites exert convergent but mechanistically distinct influences on the CNS. SCFAs primarily act via G-protein–coupled receptors (FFAR2/FFAR3) and histone deacetylase inhibition, modulating microglial maturation, neurotransmission, and neuroinflammation (19). In parallel, tryptophan-derived metabolites access the brain through LAT1 transporters and act via serotonin receptors, kynurenine–N-methyl-D-aspartate (NMDA) signaling, and the aryl hydrocarbon receptor (AhR), ultimately producing overlapping outcomes such as modulation of mood, synaptic function, and stress reactivity (20–22). Alterations in bile acid pools further modulate dopaminergic and noradrenergic circuits, affecting limbic regions such as the prefrontal cortex and amygdala that are critical for mood regulation (23). Concurrently, these metabolic shifts enhance systemic inflammation by amplifying IL-6 and TNF-α release, priming microglia, and compromising the BBB integrity, mechanisms long implicated in depression and other affective disorders (24). Collectively, these findings establish a mechanistic link between microbial dysbiosis, aberrant neurotransmission, and neuroimmune dysfunction.
Historically, depression was viewed primarily through neurochemical models, but immunological perspectives now dominate (25). Peripheral immune activation induces “sickness behavior,” an adaptive, cytokine-driven syndrome characterized by anhedonia, fatigue, and social withdrawal, that overlaps strikingly with core features of depression (15, 26). Proinflammatory cytokines cross the BBB and modulate neurotransmitter systems including serotonin, dopamine, and glutamate, thereby precipitating behavioral and cognitive changes (3, 24). Evolutionary theories posit that depressive behaviors, while maladaptive in modern societies, once conferred survival benefits by conserving energy during infection or injury (3). Contemporary studies confirm that individuals with heightened inflammatory responses to psychosocial stressors are at greater risk of developing depression, reinforcing the gut–immune–brain axis as a central mechanism (9, 27).
Human studies now provide strong evidence for these links. Large-scale metagenomic analyses, such as those from the Flemish Gut Flora Project, revealed that butyrate-producing genera like Faecalibacterium and Coprococcus are associated with improved quality of life, whereas their depletion, along with reduced Dialister, correlates with depression (11). In contrast, enrichment of pro-inflammatory taxa such as Eggerthella and Enterobacteriaceae has been reported in MDD patients. Multi-omics studies consistently demonstrate that these microbial signatures are mirrored by metabolic shifts, reduced SCFAs, indoles, and serotonin precursors alongside elevated kynurenine pathway metabolites, that converge to impair neurotransmitter balance and promote chronic neuroinflammation (11, 18, 28, 29). These microbial signatures are paralleled by metabolic shifts, including reduced SCFAs and indoles and increased kynurenine pathway metabolites, all of which influence neurotransmitter signaling and immune activation (4, 11). Clinically, many patients with depression and anxiety also present with gastrointestinal comorbidities, highlighting the translational relevance of the microbiota–gut–brain axis (4).
An often-overlooked dimension in this model is the role of circadian rhythms. Both host endocrine cycles, such as cortisol and melatonin secretion, and microbial communities display diurnal oscillations that synchronize metabolic and neural processes (9, 30). Disruption of these rhythms through factors such as shift work, irregular feeding, or sleep disturbances alters microbial diversity, dampens SCFA oscillations, and destabilizes HPA regulation (4, 31). Experimental evidence now shows that circadian rhythm disruption (CRD) induces disturbed expression of the intestinal epithelial gene Per2, drives gut microbiota dysbiosis, and via FMT, transfers depression-like phenotypes to naïve recipients, thereby implicating microbiota circadian misalignment as a causal driver of mood vulnerability (9, 32, 33). Recent studies further reveal that circadian disruption not only exacerbates neurotransmitter dysregulation but also heightens systemic inflammation, thereby creating a temporal dimension to gut–brain–immune interactions (31, 34, 35). These findings integrate circadian biology into the gut–brain–immune framework and open the door to chronotherapeutic and microbiota-targeted interventions.
Together, this evidence converges on the conclusion that microbial dysbiosis contributes causally to the onset and persistence of anxiety and depression (6, 29). This reconceptualizes psychiatric disorders as systemic conditions arising from the intersection of neural, endocrine, immune, metabolic, and circadian networks, all shaped by the microbiota, rather than isolated brain-based pathologies. Beyond mechanistic insights, these findings have translational implications: microbial taxa and metabolites are emerging as biomarkers for disease risk and treatment response, while interventions such as psychobiotics, FMT, precision diets, chrononutrition, and immune-modulatory therapies hold promise for individualized psychiatric care (9, 36).
The gut–brain axis in the pathophysiology of anxiety and depression
The gut–brain axis has emerged as a central conceptual framework for understanding anxiety and depressive disorders, emphasizing the bidirectional communication between the CNS, the ENS, the endocrine and immune systems, and the gut microbiota (6, 37, 38). Communication across this axis occurs through neural, endocrine, immune, metabolic, and circadian pathways that collectively orchestrate stress responses, emotional regulation, and systemic homeostasis. Neural signaling is exemplified by the vagus nerve, which transmits microbial cues directly from the gut to the brain; indeed, vagotomy abolishes depression-like behaviors induced by FMT from depressed individuals into rodents, demonstrating a causal link between microbial signals and mood regulation (39, 40). Endocrine pathways converge on the HPA axis, where microbial alterations influence glucocorticoid rhythms, cortisol secretion, and stress reactivity (41–43). Immune mechanisms provide another conduit: LPS and other microbial components can cross a compromised gut barrier, triggering systemic cytokine release such as IL-1β, IL-6, and TNF-α, which activate microglia, remodel synaptic function, and promote neuroinflammation (19, 41). In parallel, microbial metabolites such as SCFAs enhance BBB integrity and synaptic plasticity, while tryptophan metabolites regulate serotonin and kynurenine pathways that influence mood and cognition (6, 23). Importantly, these neural, endocrine, immune, and metabolic pathways are not temporally static, they are entrained by circadian clocks, and disruption of daily rhythmicity amplifies gut dysbiosis, HPA hyperactivity, and neuroinflammatory tone, ultimately heightening vulnerability to anxiety and depression (31, 44–47) (Table 1).
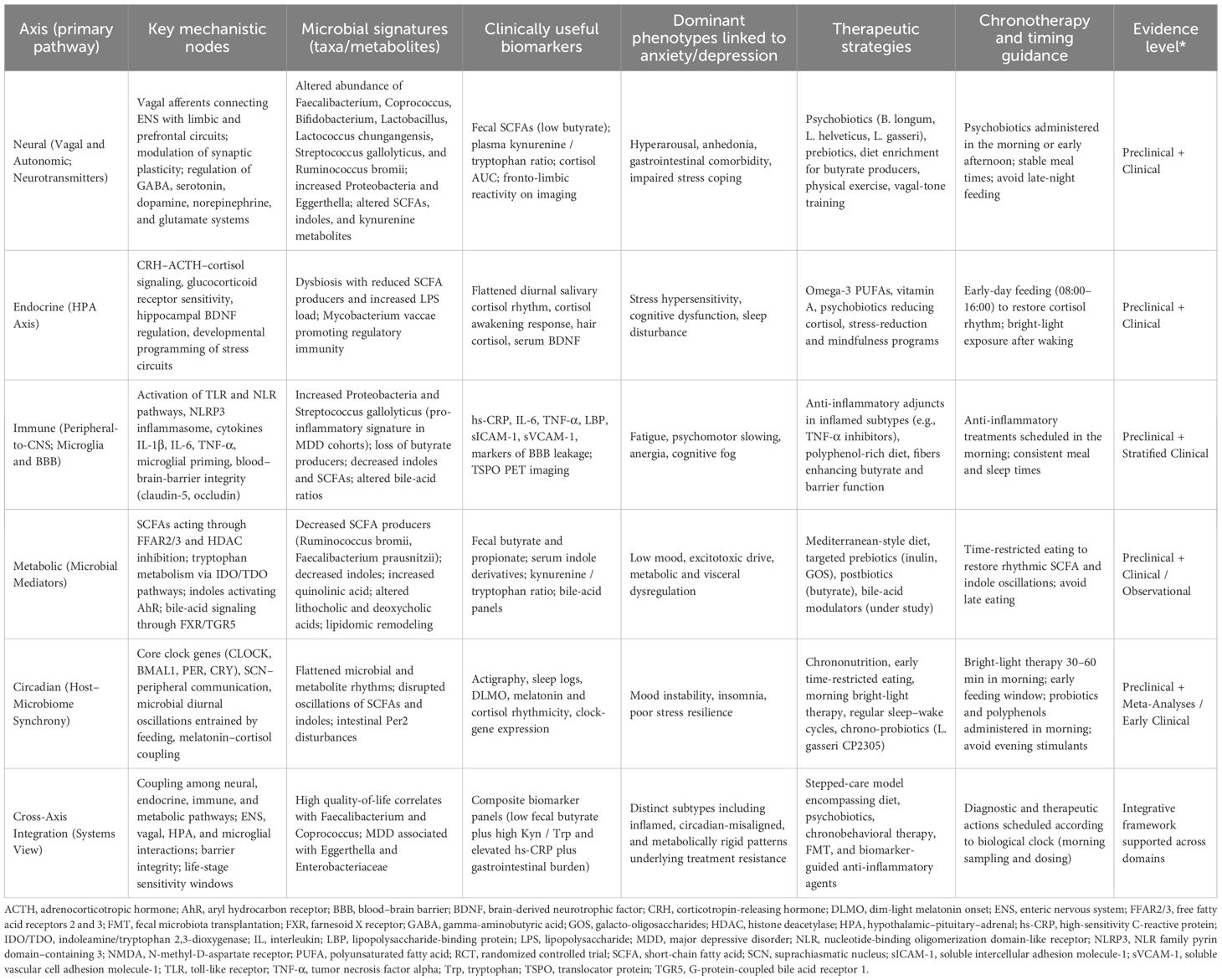
Table 1. Integrated gut–brain–circadian axis in anxiety and depression: mechanisms, microbial signatures, and therapeutic opportunities.
Preclinical studies have established that the microbiota is an active regulator of stress circuitry rather than a passive correlate. Germ-free mice exhibit exaggerated HPA responses and anxiety-like behaviors that normalize following colonization with commensal microbiota, demonstrating that microbial input is essential for appropriate stress regulation (6, 37, 48). Complementary strategies, perturbation, replacement, and rescue, further reinforce causality: antibiotic-induced dysbiosis elicits anxiety- and depression-like phenotypes reversible by recolonization; monoassociation with Bifidobacterium infantis or selected Lactobacillus species recalibrates HPA tone and reduces behavioral despair; supplementation with microbial metabolites such as butyrate or indole derivatives restores microglial maturation, BBB integrity, and synaptic plasticity; and vagal deafferentation abolishes probiotic-specific behavioral effects, confirming that neural pathways mediate microbial influence on mood (39, 49). These converging lines of evidence highlight that gut microbial signals actively shape affect-related neural circuitry through endocrine and immune cross-talk.
Beyond these mechanisms, emerging data reveal that the gut microbiota and the host circadian system are dynamically intertwined. The microbiota exhibits robust diurnal oscillations in composition, localization, and metabolic activity that align with host feeding–fasting cycles and hormonal rhythms. These microbial rhythms, in turn, entrain peripheral clocks within intestinal, hepatic, and immune tissues, coordinating metabolic and neuroendocrine homeostasis (31, 46). Microbial depletion through antibiotics disrupts rhythmic gene expression in the hippocampus, amygdala, and SCN, alters corticosterone rhythmicity, and produces time-of-day–dependent changes in stress reactivity, indicating that microbial cues synchronize neuroendocrine rhythms (50). Transcriptomic analyses further show enrichment of dysregulated genes related to BBB permeability, circadian timing, and stress adaptation, revealing a molecular bridge between microbial signals and host clock machinery (50). Likewise, gut microbes drive daily oscillations in tryptophan metabolism: microbial regulation is required for the diurnal production of serotonin and kynurenine metabolites that modulate mood-related neurotransmission (51). Conversely, chronic stress flattens microbial rhythmicity; in depression-model mice, restraint stress abolishes circadian oscillations of key microbial taxa and metabolic pathways (52). Loss of host clock genes such as Clock or Bmal1 produces parallel effects, erasing microbial oscillations and disrupting metabolic synchrony (31).
At the systems level, meal timing emerges as a dominant synchronizer of microbiota–clock interactions. Irregular feeding schedules blunt microbial oscillations and propagate circadian misalignment across the gut–brain–immune axis, leading to metabolic, inflammatory, and behavioral dysregulation (53). Together, these findings indicate that gut dysbiosis in mood disorders reflects not only compositional imbalance but also a loss of temporal order, flattened microbial rhythms, mistimed metabolite release, and desynchronized clocks across host tissues. This concept redefines dysbiosis as a dynamic, time-sensitive disruption of microbiota–host synchrony rather than a static state of imbalance.
Clinical studies reinforce these mechanistic insights. Large-scale metagenomic analyses in major depressive disorder (MDD) reveal depletion of butyrate-producing genera such as Faecalibacterium prausnitzii and Coprococcus, alongside enrichment of proinflammatory taxa including Enterobacteriaceae (41, 54, 55). These microbial shifts correlate with systemic inflammation, impaired intestinal barrier integrity, and altered connectivity in emotion-related brain regions (42, 56). Moreover, microbial and metabolite signatures correlate with treatment response, suggesting their potential as biomarkers to stratify patients and predict therapeutic outcomes (54, 56). When integrated with circadian evidence, these findings suggest that psychiatric vulnerability arises from temporal desynchrony across microbial, immune, and neural systems, and that restoring rhythmic harmony through interventions such as time-restricted feeding, chrononutrition, or circadian-timed psychobiotics may enhance therapeutic efficacy.
Collectively, evidence from animal models and human cohorts supports a model in which gut microbial dysbiosis contributes causally to the onset and persistence of anxiety and depression through integrated neural, endocrine, immune, metabolic, and circadian pathways. This time-sensitive systems framework redefines psychiatric disorders as dynamic, body-wide conditions shaped by microbiota–clock co-regulation. It also underscores the need for chronobiologically optimized interventions, ranging from diet and microbiome modulation to light–sleep alignment, to re-entrain disrupted microbial and host rhythms for improved mental health outcomes.
Neural circuits and neurotransmitter modulation in the gut–brain axis
The microbiota shapes anxiety- and depression–relevant emotional and cognitive functions through well-defined neuroanatomical circuits that connect the gut with the CNS (57). The vagus nerve represents the most prominent communication pathway, transmitting microbial and luminal signals from the gastrointestinal tract to brainstem nuclei implicated in mood regulation and projecting to limbic/prefrontal targets that govern fear, negative affect, and stress coping (58). Experimental studies show that probiotics alter vagal afferent activity, while vagotomy abolishes these effects, eliminating anxiolytic/antidepressant-like behavioral responses and confirming vagal dependency (6, 59). Complementing this route, spinal and sympathetic pathways convey microbial cues related to visceral nociception, stress responsiveness, and autonomic regulation which can amplify hyperarousal and stress reactivity typical of anxiety and depressive phenotypes (4, 60). Within the ENS, microbial metabolites modulate neuronal excitability and gut motility, reshaping interoceptive signaling that feeds forward to affective circuits and helps explain frequent GI comorbidity in anxiety and depression (61, 62). In what follows, we focus specifically on how the microbiota modulates core neurotransmitter systems that mechanistically link gut signals to anxiety and depressive symptom domains.
Beyond structural connectivity, the microbiota exerts profound effects on neurotransmitter systems. Several bacterial taxa directly synthesize or regulate bioactive amines. Lactobacillus and Bifidobacterium species produce γ-aminobutyric acid (GABA), a key inhibitory neurotransmitter strongly linked to anxiety phenotypes (60, 63, 64). Reduced abundance of GABA-producing taxa is associated with heightened amygdala reactivity and anxiety-like behavior, whereas probiotic supplementation that increases microbial GABA production correlates with decreased behavioral despair and reduced physiological stress markers in preclinical and clinical settings (60, 63, 64). Microbiota also influence serotonergic tone through interactions with enterochromaffin cells: spore-forming bacteria stimulate peripheral serotonin biosynthesis, which constitutes a major portion of circulating serotonin that regulates both gut motility and central serotonergic signaling (6, 9). In depressive phenotypes, dysbiosis diverts tryptophan away from 5-HT synthesis toward the kynurenine pathway, contributing to anhedonia and low mood; restoration of eubiotic communities or targeted psychobiotics increases 5-HT–related signaling and tracks with symptom improvement (6, 9, 65). Dopaminergic and noradrenergic systems are similarly modulated by microbiota-derived metabolites, including SCFAs and tryptophan catabolites, which alter catecholamine synthesis and receptor sensitivity in mood-related brain regions such as the prefrontal cortex and amygdala (7, 28, 61). These catecholaminergic effects map onto motivational deficits, psychomotor slowing, and heightened arousal commonly observed in depression and anxiety, linking microbial metabolite availability to reward processing and stress responsivity (66, 67).
Microbial regulation of neurotransmission extends beyond production to the modulation of synaptic plasticity and receptor signaling. Indole derivatives and bile acid metabolites cross the intestinal barrier and act on neuronal and glial targets, reshaping synaptic connectivity in cortical and limbic regions (60, 62). Indole–AhR signaling supports serotonergic and glutamatergic balance and dampens microglial activation, thereby constraining anxiety-like behavior and depressive affect; loss of indole producers removes this brake on excitatory tone and neuroinflammation (68, 69). SCFAs also play a pivotal role in restoring microglial maturation and dampening neuroinflammation in germ-free mice, suggesting that microbial metabolites not only influence neurotransmitter availability but also support cellular processes fundamental for CNS homeostasis (9, 17). Functionally, butyrate’s histone deacetylase (HDAC) inhibition enhances expression of GABAergic and neurotrophic genes, normalizes synaptic plasticity in stress-responsive circuits, and is repeatedly linked to reductions in anxiety- and depression-like behaviors (70).
Glutamatergic signaling provides a key convergence point for anxiety and depression. Microbiota-driven activation of indoleamine 2,3-dioxygenase (IDO) shifts tryptophan toward kynurenine metabolites; quinolinic acid (NMDA agonist) promotes excitotoxicity and anxiety/depressive behaviors, while kynurenic acid (NMDA antagonist) can be neuroprotective (71–74). Elevated Kyn/Trp ratios in dysbiosis thus index a neurotransmitter imbalance (low 5-HT, high Glu/NMDA) that correlates with symptom severity and treatment resistance. Interventions that restore SCFAs and indoles, or reduce peripheral inflammation (which drives IDO), rebalance this axis and improve affective outcomes (75, 76).
Neurotransmitter dynamics are further synchronized by circadian clocks, which regulate their synthesis and release (34). Microbial metabolites such as SCFAs and indoles exhibit daily oscillations that tune GABAergic, serotonergic, and dopaminergic signaling. Disruption of these rhythms compromises stress resilience and mood stability, providing an additional temporal dimension to gut–brain interactions (44, 77, 78). Accordingly, timing interventions (chrononutrition, circadian-aligned psychobiotics) to coincide with peak microbial metabolite release enhances GABAergic tone and stabilizes monoaminergic balance, offering a mechanistic rationale for time-of-day–specific treatment windows in anxiety and depression (35, 79–81).
Importantly, the relationship between neurotransmission and microbiota is reciprocal. Stress and altered neural activity reshape gut microbial composition, which in turn modifies neurotransmitter availability and signaling efficacy (4, 6). Chronic stress reduces microbial diversity and disrupts metabolic outputs such as SCFAs and indoles, thereby reinforcing maladaptive neurotransmission patterns underlying anxiety and depression (7, 82). This creates a feed-forward loop, stress hormones remodel the microbiota, microbial outputs skew neurotransmitters (low GABA, low 5-HT, high Kyn/Trp, high Glu/NMDA), and these shifts intensify anxiety and depressive symptoms, highlighting neurotransmitter modulation as the proximate pathway through which the microbiota shapes affect. Thus, microbial control of inhibitory (GABA), monoaminergic (5-HT/DA/NE), and glutamatergic (NMDA) systems provides a direct, mechanistic bridge from gut dysbiosis to the core symptom dimensions of anxiety and depression, and a tractable target for biomarker-guided interventions (60, 83–85).
Microbiota–endocrine crosstalk and hpa axis dysregulation
The HPA axis is the central coordinator of the stress response, and its dysregulation is strongly implicated in anxiety and depression (86). Emerging evidence indicates that the gut microbiota critically shapes HPA axis activity through endocrine and metabolic pathways, creating a reciprocal loop in which microbial communities influence stress hormones while glucocorticoids remodel the gut ecosystem (87, 88).
Preclinical studies provide direct behavioral evidence that microbiota–endocrine interactions modulate anxiety- and depression-like outcomes. In rodent models, adolescent stress not only induces hippocampal BDNF downregulation and cognitive impairments but also produces pronounced anxiety- and depression-like behaviors, such as increased immobility in the forced swim test (FST), reduced sucrose preference (SPT), and enhanced avoidance in the elevated plus maze (EPM), changes that parallel gut microbial dysbiosis (89, 90). Dietary interventions with omega-3 polyunsaturated fatty acids or vitamin A attenuate these affective phenotypes, restoring exploratory behavior and anhedonia scores, normalizing hippocampal BDNF expression, and partially reestablishing microbial composition, suggesting that microbial–endocrine signaling buffers against HPA hyperactivation (63, 90). Similarly, administration of Mycobacterium vaccae confers robust anxiolytic and antidepressant-like effects, reducing immobility in FST, improving open-field exploration, and normalizing corticosterone levels, through Treg-mediated immunomodulation and glucocorticoid dampening, thereby enhancing resilience to chronic stress and inflammation (91).
Clinical findings parallel these experimental observations. Patients with depression and anxiety frequently show altered gut microbial signatures associated with hypercortisolemia and disrupted diurnal cortisol rhythms (92, 93). Translational models confirm that microbiota perturbations heighten HPA axis reactivity and aggravate depressive phenotypes (6).
Importantly, probiotic supplementation with Bifidobacterium and Lactobacillus strains reduces cortisol levels and improves both mood and stress responsiveness (94, 95). Randomized controlled trials further demonstrate that specific probiotic formulations, such as Lactobacillus casei Shirota, Lactobacillus gasseri CP2305, and Bifidobacterium longum 1714, lower salivary cortisol and attenuate anxiety or stress-induced physiological responses in humans (96–98). Additionally, a multi-strain combination of L. helveticus R0052 and B. longum R0175 has been shown to reduce urinary cortisol and psychological distress in both preclinical and clinical models (99, 100).
The mechanistic basis of this crosstalk lies partly in microbial metabolites. SCFAs modulate glucocorticoid receptor sensitivity and neurotransmitter release, directly linking bacterial fermentation products to stress hormone regulation (101). Developmental studies add another layer of complexity: germ-free animals display exaggerated ACTH and corticosterone responses to stress, which normalize only when colonization with Bifidobacterium infantis occurs during early life, revealing a sensitive period for microbiota-driven endocrine programming (102). In humans, early-life adversity correlates with persistent microbial alterations and long-term cortisol dysregulation, suggesting that developmental disruptions in microbiota–endocrine communication may predispose individuals to psychiatric vulnerability (90). These developmental findings emphasize that timing of microbial exposure, particularly during critical windows such as infancy and adolescence, can have enduring consequences on HPA-axis regulation and stress resilience.
Notably, the HPA axis is tightly regulated by circadian clocks. Dysbiosis-driven cortisol abnormalities often coincide with flattened diurnal rhythms, reinforcing maladaptive stress responses and increasing susceptibility to anxiety and depression (103–105). Circadian misalignment not only alters cortisol rhythmicity but also disrupts gut microbial oscillations, amplifying systemic inflammation and stress sensitivity. Chronotherapeutic interventions that restore circadian–microbial synchrony, such as time-restricted feeding, chrononutrition, and light–sleep alignment, are now being investigated as strategies to recalibrate endocrine and microbial rhythms in mood disorders (31, 44, 106). Together, these findings establish the microbiota as an active regulator of HPA axis function. By integrating endocrine, immune, and circadian control, microbial communities determine the threshold and recovery of stress responses. Dysbiosis amplifies stress hormone release and psychiatric vulnerability, whereas targeted interventions, including psychobiotics, prebiotics, dietary supplementation, and timing-aligned (chronobiotic) interventions, offer promising avenues to restore endocrine balance and enhance resilience against anxiety- and depression-related disorders (107).
Neuroimmune mechanisms linking gut dysbiosis to anxiety and depression
Gut dysbiosis profoundly influences the immune system, triggering a cascade of neuroimmune alterations that extend to the CNS (108, 109). Perturbations in microbial composition promote chronic low-grade inflammation through the release of pathogen-associated molecular patterns (PAMPs) and danger-associated molecular patterns (DAMPs), which engage pattern recognition receptors such as Toll-like receptors and NOD-like receptors (110). These signals activate inflammasomes and upregulate proinflammatory cytokines, including IL-1β, IL-6, and TNF-α, which in turn modulate microglial activity and neural plasticity, fostering vulnerability to psychiatric disorders (110–112). Seminal frameworks now link sustained peripheral inflammation to “sickness behavior” and depression via immune-to-brain signaling, anchoring dysbiosis-driven inflammation within modern depression pathophysiology (15).
A study demonstrates that gut microbiota disturbances compromise the BBB, facilitating peripheral immune mediators’ access to the brain (113). Chronic social stress reduces tight-junction protein claudin-5, breaches the BBB, and enables peripheral IL-6 infiltration into limbic circuits, driving depression-like behaviors; convergent work confirms stress-induced BBB pathology across models (114). Beyond animal models, vascular activation markers, including soluble adhesion molecules, are altered in depression, and in women with MDD, circulating sE-selectin is significantly elevated, underscoring sex-linked endothelial contributions to neuroimmune dysregulation (115). Cytokines can also access or signal across the BBB via endothelial and perivascular routes, providing direct channels by which peripheral immune tone shapes CNS function (116).
At the cellular level, gut-derived metabolites further orchestrate neuroimmune interactions. SCFAs, normally protective, become depleted in dysbiosis, removing a key brake on histone deacetylases and G-protein-coupled receptor signaling (117, 118). This loss enhances microglial reactivity and reduces regulatory T cell function, driving maladaptive inflammatory signaling (17, 119). Additionally, kynurenine pathway metabolites, shaped by microbial metabolism of tryptophan, engage the AhR to promote Treg expansion and dampen cytotoxic T cell activity, thereby linking metabolic disturbances to immune escape and neurodegeneration (120–123). Microglial NLRP3 inflammasome activation has emerged as a central node that converts stress and metabolic danger signals into IL-1β-driven neuroinflammation and depressive-like behavior, highlighting druggable checkpoints within the gut–immune–brain axis (124, 125).
Sterile inflammation, independent of pathogens, can be initiated by psychological stress and gut dysbiosis, culminating in increased NLRP3 activation and IL-1β release in both the periphery and the CNS (125–127). These processes compromise neuronal survival and synaptic plasticity, fostering cognitive and mood disturbances. Furthermore, peripheral immune activation translates into CNS changes via vagal and humoral routes, with immune-to-brain signaling shaping behavioral responses to stress and contributing to depression and anxiety phenotypes (128). Importantly, human molecular-imaging studies show increased brain TSPO binding, a proxy for microglial activation, during major depressive episodes, providing in vivo evidence that heightened neuroinflammation accompanies symptom severity (129).
Microglia, the resident immune cells of the brain, are especially sensitive to gut microbial signals. Dysbiosis-induced priming of microglia amplifies proinflammatory cytokine release and impairs phagocytic clearance of pathological proteins. This dysfunctional state is closely linked to aberrant synaptic pruning and neuronal circuit remodeling (3). Crucially, systemic immune alterations dictate microglial phenotypes: chronic elevations of IL-6, TNF-α, and IL-1β “prime” microglia, lowering activation thresholds and exaggerating responses to subsequent stressors, whereas IL-10 and TGF-β bias microglia toward homeostatic states. This systemic-to-central coupling reframes microglia as sensors of peripheral immune tone, not merely local responders (3). Microbiota are indispensable for normal microglial maturation and function, and their depletion disrupts microglial transcriptional homeostasis, further connecting gut ecology to central immune set-points (130). Parallel evidence shows that immune-cell trafficking into the CNS, facilitated by BBB disruption and chemokine gradients, intensifies neuroinflammation and exacerbates behavioral deficits (114, 131, 132).
Clinical and translational evidence corroborates these mechanistic insights. Patients with depression exhibit systemic inflammation, BBB leakage, and altered immune signatures in brain tissue, providing convergent evidence that gut dysbiosis-driven neuroimmune dysregulation is not a secondary phenomenon but a mechanistic driver of psychiatric pathology (133, 134). Finally, circadian control powerfully shapes these processes: immune transcripts and cytokines oscillate across the day, and circadian misalignment amplifies pro-inflammatory output and microglial reactivity, linking disrupted temporal organization to persistent neuroinflammation in mood disorders (135–138).
Metabolic mediators of microbiota–brain communication
The gut microbiota generates a wide array of metabolites that directly and indirectly influence brain physiology, many of which have been implicated in the pathophysiology of anxiety and depression (139, 140). Among the most studied are SCFAs such as acetate, propionate, and butyrate. These molecules act as epigenetic regulators by inhibiting histone deacetylases, thereby modulating the transcription of genes involved in neuroplasticity and stress responses (130, 141, 142). In parallel, SCFAs interact with G protein–coupled receptors, including GPR41 and GPR43, shaping systemic immune activity and favoring the production of anti-inflammatory cytokines, which in turn influence mood regulation (11, 143). Evidence from animal models shows that depletion of SCFA-producing bacteria heightens anxiety-like behaviors, while butyrate supplementation restores synaptic integrity and normalizes HPA axis function (29, 144).
Tryptophan metabolism constitutes another crucial microbiota-controlled pathway linking the gut and brain (18). Beneficial microbes such as Bifidobacterium and Lactobacillus spp. enhance serotonin biosynthesis by stimulating enterochromaffin cells, influencing central serotonergic signaling through vagal and circulatory routes (18, 145). However, in the context of dysbiosis, tryptophan is preferentially metabolized via the kynurenine pathway through activation of IDO, leading to the accumulation of neuroactive metabolites such as quinolinic acid, which acts as an NMDA receptor agonist and exerts neurotoxic effects, and kynurenic acid, which functions as an NMDA antagonist with neuroprotective properties (11, 146). This imbalance has been linked to excitotoxicity, chronic neuroinflammation, and depressive symptomatology, and clinical studies consistently report altered kynurenine-to-tryptophan ratios in patients with MDD (146).
Beyond these classical metabolites, bile acids have recently emerged as key modulators of microbiota–brain communication. Primary bile acids synthesized in the liver undergo microbial transformation into secondary bile acids such as deoxycholic acid and lithocholic acid, which can cross the BBB and act on receptors including FXR and TGR5 (147, 148). These receptors influence systemic metabolism, immune signaling, and microglial activation, processes that collectively shape neural plasticity and emotional regulation. Dysregulated bile acid pools have been associated with impaired hippocampal function, altered energy balance, and enhanced anxiety-like behavior, positioning bile acid signaling as an additional therapeutic target in mood disorders (149, 150). Microbial catabolism of tryptophan also yields indole derivatives such as indole-3-acetic acid and indole-3-aldehyde, which activate the AhR in epithelial and immune cells (151). Activation of this receptor supports intestinal barrier integrity and restrains pro-inflammatory cascades that otherwise propagate systemic inflammation and brain dysfunction. Reduced levels of indole-producing microbes are frequently observed in anxiety and depression, suggesting that impaired indole–AhR signaling contributes to disease pathology and may be restored through microbiome-targeted interventions (151). Lactate, another microbial metabolite, is increasingly recognized not only as a byproduct of glycolysis but also as a key neuroenergetic shuttle. Microbiota-derived lactate fuels astrocyte–neuron metabolic coupling, thereby sustaining synaptic plasticity and modulating neuronal excitability (152). Animal studies indicate that higher microbial lactate production correlates with resilience to stress-induced behavioral alterations, underscoring its neuroprotective role (153). Many of these metabolites, including SCFAs, bile acids, and kynurenine pathway intermediates, exhibit circadian oscillations synchronized with host feeding–fasting cycles. Disruption of these rhythms contributes to metabolic misalignment, impaired neurotransmission, and mood dysregulation (154, 155).
Taken together, these metabolites constitute an interconnected signaling network that links gut microbial activity to brain function. Anxiety and depression are consistently associated with a metabolic signature characterized by reduced SCFAs and indoles, elevated kynurenine pathway metabolites, and disrupted bile acid profiles (142, 146, 150, 156). Therapeutic approaches aimed at restoring microbial balance, through prebiotics, probiotics, or dietary strategies, have demonstrated the capacity to reestablish metabolite production, normalize neurotransmission and inflammatory tone, and ultimately alleviate mood-related symptoms (5).
Circadian rhythm disruption and the gut–brain axis
Circadian rhythms, generated by transcription–translation feedback loops of core clock genes such as CLOCK, BMAL1, PER, and CRY, orchestrate nearly all aspects of physiology, including sleep–wake cycles, hormone secretion, metabolism, and immune responses (157, 158). While the suprachiasmatic nucleus (SCN) serves as the central pacemaker, peripheral clocks in the gut, liver, and immune system also display robust oscillations that synchronize host metabolism and stress responsivity to environmental cues (157). Importantly, the gut microbiota itself undergoes pronounced diurnal oscillations in composition, localization, and metabolite production, which are tightly entrained by host feeding–fasting cycles and circadian signaling (159, 160).
Seminal studies demonstrated that intestinal bacteria coordinate with the circadian system to shape host gene expression and metabolic rhythms. For instance, diurnal microbial relocation along the gut epithelium and rhythmic metabolite fluxes program oscillations in the host liver transcriptome and epigenetic landscape (160). Similarly, the microbiota programs HDAC3-dependent diurnal transcription in the intestine, controlling lipid uptake and energy balance (44). Perturbation of these rhythms, either by antibiotics, germ-free conditions, or arrhythmic feeding, disrupts circadian gene expression and increases susceptibility to obesity and inflammation (50, 161).
Circadian disruption also alters gut–immune interactions. Segmented filamentous bacteria (SFB) exhibit rhythmic attachment to epithelial cells, driving ILC3–STAT3–mediated (80) antimicrobial protein oscillations that enhance time-of-day–specific resistance to infection. These microbiota–immune interactions are synchronized with feeding rhythms, underscoring the integration of diet, circadian clocks, and microbial dynamics. In parallel, the microbiota regulates diurnal corticosterone secretion, modulating HPA axis activity and stress responsiveness (81). Microbial depletion abolishes rhythmicity of corticosterone and stress pathway gene expression in the hippocampus and amygdala, leading to time-of-day–specific impairments in stress resilience (81, 162).
Disruption of circadian rhythmicity, through shift work, jet lag, high-fat diets, or sleep disturbance, induces microbial dysbiosis, dampens metabolite oscillations, and desynchronizes host clocks (50, 158, 161). Human studies show that such misalignment exacerbates anxiety and depression, as circadian disruption not only destabilizes sleep but also amplifies neuroinflammatory signaling and HPA hyperactivity (35). This bidirectional dysfunction creates a feed-forward loop in which stress and microbial imbalance reinforce circadian misalignment, heightening psychiatric vulnerability (81, 160). Integrating circadian biology into the gut–brain research opens promising therapeutic avenues. Chrononutrition and time-restricted feeding restore microbial rhythmicity and improve host metabolic and mood outcomes (50, 159). Likewise, probiotics such as Lactobacillus reuteri have been shown to normalize glucocorticoid rhythms and stress responsivity (81). Emerging chronotherapeutic strategies suggest that aligning microbial interventions such as psychobiotics, FMT, or engineered microbiomes with circadian windows may maximize efficacy for anxiety and depression (35, 157, 160).
Microbial signatures and biomarkers
The gut microbiome has emerged as a central determinant of neuropsychiatric health, with specific microbial taxa and their metabolites increasingly recognized as candidate biomarkers for anxiety and depression (163). Multi-omics studies demonstrate that microbial dysbiosis directly correlates with immune activation, host metabolism, and brain structure. For example, clinical cohorts revealed altered abundance of Ruminococcus bromii, Lactococcus chungangensis, and Streptococcus gallolyticus in individuals with MDD, with these taxa increasing pro-inflammatory signaling via LPS- and peptidoglycan-mediated TLR/NOD pathways, elevating IL-1β and perturbing lipid oxidation, and reductions in grey matter volume in the inferior frontal gyrus (164). Notably, these microbial shifts were associated with functional brain imaging readouts, including altered fronto-limbic connectivity that tracks peripheral cytokine load, providing neuroanatomical correlates of dysbiosis. Extending this, large population-level data from the Flemish Gut Flora Project identified butyrate-producing Faecalibacterium and Coprococcus as positive correlates of quality of life and inverse correlates of depression scores, where butyrate inhibits HDACs, activates FFAR2/FFAR3, tightens epithelial junctions, and suppresses IL-6/TNF-α, establishing these genera as reproducible, population-scale candidates for biomarker panels (165).
Meta-analyses reinforce these findings by showing consistent depletion of butyrate-producing genera such as Butyricicoccus, Coprococcus, and Faecalibacterium, alongside enrichment of pro-inflammatory taxa including Eggerthella, Enterococcus, Flavonifractor, and Streptococcus in depressive patients (41, 166). Loss of SCFA producers reduces mucosal butyrate, weakens claudin/occludin expression, increases microbial translocation, and amplifies NLRP3-IL-1β signaling, both of which are critical drivers of HPA axis overactivation and mood disturbances (167). Such immune-related microbial signatures are increasingly proposed as stratification tools to identify patients likely to benefit from anti-inflammatory or microbiota-directed interventions. Recent systematic syntheses confirm these compositional trends across studies and tie them to quantitative biomarker thresholds (e.g., low fecal butyrate, high kynurenine/tryptophan ratio, elevated LBP/CRP), supporting their use as putative diagnostic and prognostic markers rather than mere correlates (168).
Specific taxa repeatedly appear as microbial signatures of psychiatric dysfunction. Bifidobacterium and Lactobacillus are frequently diminished in major depression, which reduces microbial GABA production and attenuates enterochromaffin 5-HT biosynthesis via TPH1 stimulation, thereby linking gut dysbiosis to neurotransmitter imbalance (11, 169). Conversely, expansion of Actinobacteria and Proteobacteria has been correlated with inflammatory cytokine release and oxidative stress pathways (170, 171), with Proteobacteria enriching endotoxin load that activates TLR4/NF-κB and elevates IL-6/TNF-α. Importantly, these compositional imbalances have been shown to normalize following probiotic supplementation or targeted diets, accompanied by restored fecal SCFAs, lower Kyn/Trp ratios, and reduced circulating sICAM-1/sVCAM-1, underscoring their translational potential. Causality is supported by FMT: transfer of “depression microbiota” increases hippocampal quinolinic acid, decreases butyrate, and induces microglial priming and depressive-like behavior, whereas healthy-donor microbiota does not, functionally validating these signatures (169).
Integrative omics has further advanced biomarker discovery. Metagenomic and metabolomic studies highlight disrupted tryptophan metabolism as a central pathway, with microbial activation of the kynurenine pathway via IDO/TDO diverting tryptophan from 5-HT, accumulating neuroactive kynurenines (quinolinic > kynurenic), and potentiating NMDA-dependent excitotoxicity (164, 165). At the same time, SCFA depletion and altered phosphoethanolamine levels suggest lipid and energy metabolism as crucial biomarker domains for depression and anxiety (165, 168). Across psychiatric cohorts, elevated kynurenine-to-tryptophan ratios and shifts in neuroactive kynurenines (e.g., quinolinic vs. kynurenic acid) are among the most consistent metabolic readouts, and are mechanistically tied to glutamatergic signaling and neuroinflammation (84). Additional axes include bile-acid remodeling, microbial deoxycholic/lithocholic acids that activate FXR/TGR5 and modulate microglial tone, and indole-AhR signaling, where reduced indole-3 derivatives diminish barrier and permit pro-inflammatory drift (172).
Collectively, these findings define an evidence-based biomarker panel that includes butyrate-producing genera such as Faecalibacterium and Coprococcus, consistently associated with improved quality of life and reduced depressive symptoms (11); depletion of Bifidobacterium and Lactobacillus, which are linked to diminished production of GABA and serotonin precursors and thus to neurotransmitter imbalance; and enrichment of pro-inflammatory taxa including Eggerthella and Enterobacteriaceae, which correlate with heightened cytokine release and neuroinflammation. Metabolically, reduced levels of SCFAs and indole derivatives together with elevated kynurenine-pathway intermediates, long implicated in glutamatergic dysregulation and neurotoxicity, represent reproducible molecular signatures across psychiatric cohorts. Importantly, these microbial and metabolic profiles correlate with clinical outcomes and can transfer depressive phenotypes in FMT models, providing causal validation. Beyond these canonical markers, recent studies highlight bile-acid remodeling and microbial-derived lactate as novel metabolic signatures influencing limbic activity and resilience to stress. Finally, because microbial taxa and metabolites exhibit diurnal oscillations entrained by host circadian rhythms, biomarker sampling and interpretation must account for temporal dynamics, as circadian misalignment can confound associations and obscure their diagnostic and prognostic value (35).
Taken together, these biomarkers not only delineate mechanistic pathways but also serve as direct entry points for therapeutic targeting, bridging discovery with intervention.
Therapeutic strategies targeting the gut–brain axis in anxiety and depression
The gut–brain axis is increasingly recognized as a modifiable therapeutic target in anxiety and depression, with interventions spanning diet, microbial modulation, targeted microbiota transfers, and host-directed anti-inflammatory strategies (173). Collectively, these approaches demonstrate that reshaping microbiota–brain communication can produce measurable effects on emotional, cognitive, and systemic outcomes. Importantly, such interventions align with the microbial, metabolic, and immune biomarkers described above, offering a foundation for precision psychiatry (Table 1).
Among the available strategies, dietary modification remains the most robustly supported intervention. In the landmark SMILES randomized controlled trial, adjunctive dietary support led to a marked reduction in clinician-rated depression (MADRS; Cohen’s d ≈ −1.16) and quadrupled remission compared with an active social-support control (32.3% vs 8.0%) (174). A meta-analysis of 16 randomized clinical trials (RCTs) further confirmed a small-to-moderate pooled benefit of dietary interventions for depressive symptoms (g≈0.28) but not for anxiety, underscoring phenotype-specific effects (175). Long-term prospective evidence from 180,446 UK Biobank participants demonstrated that greater adherence to the EAT-Lancet dietary pattern significantly reduced the risk of depression and anxiety (HR for depression 0.71–0.84) (176). Mechanistic studies reveal that such dietary patterns enrich butyrate-producing taxa, including Faecalibacterium and Coprococcus, thereby restoring SCFA-mediated signaling and countering the microbiome deficits observed in depressive disorders.
Probiotic and prebiotic supplementation, collectively referred to as psychobiotics, has emerged as a promising adjunctive approach rather than a stand-alone therapy. A comprehensive meta-analysis showed that probiotics significantly reduced depressive symptoms when co-administered with antidepressants (SMD = 0.83), whereas no significant effect was observed in isolation (177). Mechanistic RCTs provide convergent evidence: in MDD, a four-week course of multi-strain probiotics added to standard therapy reduced Hamilton Depression Rating (HAM-D) scores more than placebo, preserved microbial diversity, increased Lactobacillus abundance correlated with symptom improvement, and attenuated putaminal reactivity during emotion-processing tasks (178). Similarly, Bifidobacterium longum NCC3001 improved depressive symptoms in patients with irritable bowel syndrome and reduced amygdala and fronto-limbic reactivity to negative stimuli, linking peripheral microbial modulation to central neural changes (179). Collectively, these studies support the capacity of psychobiotics to recalibrate neuroimmune and neuroendocrine pathways that underpin mood regulation (180).
FMT is transitioning from proof-of-concept to early clinical application. Preclinical studies demonstrate that transplantation of “depression microbiota” from affected individuals to rodents induces depressive-like behavior, establishing causality between dysbiosis and mood pathology (181). The first double-blind pilot RCT in adults with moderate-to-severe MDD confirmed the feasibility and safety of FMT via enema, reporting improved gastrointestinal symptoms and quality of life with no major adverse events (181). Future studies are moving toward precision-matched donor–recipient approaches based on specific biomarker signatures, such as low SCFA or indole levels and elevated kynurenine pathway activity, thereby translating mechanistic findings into personalized interventions.
Host-targeted anti-inflammatory strategies are gaining attention, particularly for inflammation-driven subtypes of depression. In a pivotal trial in treatment-resistant MDD, the TNF-α antagonist infliximab produced no overall advantage over placebo. However, patients with elevated baseline hs-CRP (> 5 mg/L) exhibited significantly greater reductions in HAM-D scores and higher response rates, while those with low CRP worsened on treatment, highlighting the potential harm of non-stratified immunotherapy (182). These results emphasize the necessity of integrating inflammatory biomarkers into treatment algorithms and reserving biologic agents for well-defined, inflammation-positive subgroups.
A rapidly advancing frontier integrates circadian biology into gut–brain therapeutics, aligning microbial and host rhythmicity to optimize efficacy. Chrononutrition and time-restricted eating (TRE) restore microbial diurnal oscillations, re-establish SCFA rhythmicity, and improve metabolic and mood-related parameters. Although meta-analyses report variable outcomes, consistent early-day feeding schedules (eTRE; 8 a.m.–4 p.m.) have been shown to enhance circadian synchrony and lower stress reactivity (155, 183–185). In parallel, chrono-probiotics, specifically Lactobacillus gasseri CP2305, exhibit stress-buffering and sleep-regulating properties, reducing salivary cortisol and improving subjective sleep quality in chronically stressed adults (186, 187). These findings indicate that synchronizing probiotic administration with host circadian cycles may potentiate neuroendocrine resilience and mood stability.
Beyond microbial interventions, circadian behavioral therapies are now a core component of integrated treatment frameworks. Morning bright-light therapy (BLT) acts as a circadian phase-advancing intervention that accelerates antidepressant response and improves outcomes in nonseasonal depression, with meta-analyses confirming enhanced remission and response rates when combined with pharmacotherapy or cognitive behavioral therapy (Lieverse et al., 2022; Legenbauer et al., 2024; Perera et al., 2016). These interventions normalize melatonin and cortisol rhythmicity, thereby complementing microbiota- and metabolism-targeted strategies.
When these findings are considered together, a coherent therapeutic hierarchy emerges. Dietary modification represents a scalable, evidence-based foundation for depression prevention and treatment, while anxiety may require more targeted or multimodal approaches (174–176). Psychobiotics serve as effective adjuncts that modulate neurocircuitry and immune tone (177, 178). FMT remains investigational but increasingly feasible, emphasizing donor selection and mechanistic alignment (179, 181). Inflammation-stratified biologics may benefit well-defined inflammatory subgroups (182). Finally, circadian-aligned strategies, chrononutrition, chrono-probiotics, and BLT, extend these interventions into the temporal dimension, restoring synchronization across the gut–brain–immune axis. Across modalities, precision gut microbiota modulators, encompassing diet, probiotics, engineered consortia, bacteriophages, and targeted delivery systems, should be conceptualized as biomarker- and rhythm-guided tools rather than one-size-fits-all therapies (188).
Despite these advances, important limitations persist. Probiotic and FMT studies remain small, heterogeneous in strains, dosing, and duration, and often confounded by comorbidities and concomitant medications (178, 179). Dietary interventions consistently demonstrate antidepressant efficacy but less impact on anxiety (175), and anti-TNF strategies illustrate the risks of non-stratified application (182). These limitations reinforce a stepped, integrated model: optimize diet as the foundational layer; incorporate psychobiotics with biomarker and behavioral monitoring; introduce circadian-aligned interventions such as light therapy, regular feeding rhythms, and sleep regulation; and reserve immunomodulation for biomarker-defined inflammatory phenotypes. Altogether, these therapeutic layers anchor treatment to the molecular, microbial, and temporal signatures of psychiatric disease, defining a roadmap for biomarker- and chronobiology-informed precision psychiatry (189).
Conclusions and future perspectives
The collective evidence presented across current studies underscores that the gut–brain–circadian axis is a central regulatory hub for mental health, particularly in the context of anxiety and depression (190, 191). Mechanistic insights consistently demonstrate that microbial metabolites such as SCFAs, bile acids, and tryptophan derivatives influence neuronal signaling, immune activity, and endocrine homeostasis, ultimately shaping stress responses and emotional regulation (4, 6, 11). Dysbiosis-driven alterations in vagal, HPA-axis, and immune pathways provide a causal basis for mood-related disorders, while circadian disruption emerges as a critical modifier that amplifies these effects by desynchronizing host–microbiome rhythms (15, 36). Together, these findings highlight the necessity of considering circadian alignment as an integral component in gut–brain research and therapy design.
Therapeutically, the data indicate that multiple strategies hold promise in modulating the gut–brain axis. Diet remains the most clinically validated intervention, with trials demonstrating robust improvements in depression following structured dietary modifications (3). Beyond diet, probiotics, prebiotics, and psychobiotics target microbial communities with the potential to restore balanced signaling through neuroimmune and neuroendocrine pathways (4). More advanced modalities such as FMT, engineered microbial consortia, and bacteriophage-based interventions are gaining attention, though their long-term efficacy and safety require further validation (14). Anti-inflammatory and immunomodulatory strategies targeting microbiota-driven neuroinflammation also show preclinical efficacy, opening an avenue for adjunctive therapies in patients with treatment-resistant depression and anxiety (3).
A critical frontier lies in multi-omic integration and systems biology approaches. Current multi-omic platforms have begun to delineate disease-associated microbial modules that link species, pathways, and metabolites into coherent signatures predictive of psychiatric outcomes (9, 192). However, challenges persist in harmonizing complex datasets across cohorts, accounting for inter-individual variability, and defining causality rather than correlation. Artificial intelligence and machine learning pipelines, applied to large-scale, longitudinal microbiome and host-omic datasets, will be essential to establish predictive biomarkers and patient-tailored interventions (7, 193).
Importantly, developmental and aging trajectories define sensitive “age windows” during which microbiota–brain–circadian interactions exert disproportionate influence on psychiatric vulnerability. Early-life stages such as the perinatal period and adolescence represent critical windows for microbial and circadian programming of the HPA axis and neuroimmune circuits, with disruptions during these phases leading to long-term effects on stress responsivity and mood regulation (194, 195). In contrast, midlife transitions and aging are characterized by inflammaging, reduced microbial diversity, and dampened circadian amplitude, processes that compromise resilience and increase vulnerability to anxiety and depression (19, 196). These life-stage–specific vulnerabilities also open therapeutic opportunities: early interventions with breastfeeding, structured diet, and chrononutrition may prevent maladaptive trajectories, while targeted psychobiotics, dietary polyphenols, and circadian-aligned sleep–light hygiene may restore resilience in midlife and older age. Thus, “when” to intervene becomes as critical as “what” intervention is applied.
Looking forward, three research avenues appear most critical. First, precision psychiatry must integrate circadian biology, microbial ecology, and life-stage–specific vulnerabilities with host genetics to optimize interventions for individual patients (197). Second, longitudinal and life-course studies are required to capture the developmental windows during which microbiota–brain interactions exert the strongest influence, particularly in childhood, adolescence, and aging (6). Third, clinical translation should focus on designing controlled, reproducible interventions that bridge dietary, microbial, and pharmacological approaches, while monitoring adverse outcomes such as immune-related side effects or unintended microbial shifts (36). In conclusion, the gut–brain–circadian framework offers a transformative paradigm for understanding and managing anxiety and depression. By advancing mechanistic dissection, leveraging integrative omics, and prioritizing translational research, this field is poised to deliver personalized, biologically grounded interventions that could reshape psychiatric care in the coming decade.
Author contributions
JB: Visualization, Writing – original draft, Writing – review & editing, Conceptualization, Investigation, Resources, Validation, Project administration, Formal Analysis, Supervision, Methodology, Data curation. CH-T: Visualization, Validation, Resources, Formal Analysis, Methodology, Data curation, Writing – review & editing, Writing – original draft, Investigation. MD: Methodology, Visualization, Data curation, Investigation, Validation, Writing – review & editing, Writing – original draft, Resources, Formal Analysis. JV-R: Methodology, Writing – original draft, Formal Analysis, Visualization, Investigation, Resources, Writing – review & editing, Data curation, Validation. AG-G: Resources, Investigation, Writing – original draft, Formal Analysis, Visualization, Data curation, Validation, Writing – review & editing, Methodology. AL-C: Funding acquisition, Validation, Conceptualization, Project administration, Supervision, Writing – review & editing, Formal Analysis, Writing – original draft, Methodology, Data curation, Resources, Visualization, Investigation.
Funding
The author(s) declare financial support was received for the research and/or publication of this article. This work was supported by Universidad de Las Américas (UDLA) from Quito, Ecuador.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Generative AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
References
1. Kieling C, Buchweitz C, Caye A, Silvani J, Ameis SH, Brunoni AR, et al. Worldwide prevalence and disability from mental disorders across childhood and adolescence: evidence from the global burden of disease study. JAMA Psychiatry. (2024) 81:347–56. doi: 10.1001/jamapsychiatry.2023.5051
2. Fan Y, Fan A, Yang Z, and Fan D. Global burden of mental disorders in 204 countries and territories, 1990-2021: results from the global burden of disease study 2021. BMC Psychiatry. (2025) 25:486. doi: 10.1186/s12888-025-06932-y
3. Miller AH and Raison CL. The role of inflammation in depression: from evolutionary imperative to modern treatment target. Nat Rev Immunol. (2016) 16:22–34. doi: 10.1038/nri.2015.5
4. Rogers GB, Keating DJ, Young RL, Wong ML, Licinio J, and Wesselingh S. From gut dysbiosis to altered brain function and mental illness: mechanisms and pathways. Mol Psychiatry. (2016) 21:738–48. doi: 10.1038/mp.2016.50
5. Ashique S, Mohanto S, Ahmed MG, Mishra N, Garg A, Chellappan DK, et al. Gut-brain axis: A cutting-edge approach to target neurological disorders and potential synbiotic application. Heliyon. (2024) 10:e34092. doi: 10.1016/j.heliyon.2024.e34092
6. Foster JA and McVey Neufeld K-A. Gut-brain axis: how the microbiome influences anxiety and depression. Trends Neurosci. (2013) 36:305–12. doi: 10.1016/j.tins.2013.01.005
7. Jabbari Shiadeh SM, Chan WK, Rasmusson S, Hassan N, Joca S, Westberg L, et al. Bidirectional crosstalk between the gut microbiota and cellular compartments of brain: Implications for neurodevelopmental and neuropsychiatric disorders. Transl Psychiatry. (2025) 15:278. doi: 10.1038/s41398-025-03504-2
8. Sender R, Fuchs S, and Milo R. Revised estimates for the number of human and bacteria cells in the body. PloS Biol. (2016) 14:e1002533. doi: 10.1371/journal.pbio.1002533
9. Yano JM, Yu K, Donaldson GP, Shastri GG, Ann P, Ma L, et al. Indigenous bacteria from the gut microbiota regulate host serotonin biosynthesis. Cell. (2015) 161:264–76. doi: 10.1016/j.cell.2015.02.047
10. Farzi A, Fröhlich EE, and Holzer P. Gut microbiota and the neuroendocrine system. Neurotherapeutics. (2018) 15:5–22. doi: 10.1007/s13311-017-0600-5
11. Valles-Colomer M, Falony G, Darzi Y, Tigchelaar EF, Wang J, Tito RY, et al. The neuroactive potential of the human gut microbiota in quality of life and depression. Nat Microbiol. (2019) 4:623–32. doi: 10.1038/s41564-018-0337-x
12. Chang M, Chang KT, and Chang F. Just a gut feeling: Faecal microbiota transplant for treatment of depression - A mini-review. J Psychopharmacol (Oxford). (2024) 38:353–61. doi: 10.1177/02698811241240308
13. Bonaz B, Bazin T, and Pellissier S. The vagus nerve at the interface of the microbiota-gut-brain axis. Front Neurosci. (2018) 12:49. doi: 10.3389/fnins.2018.00049
14. Park JC, Chang L, Kwon H-K, and Im S-H. Beyond the gut: decoding the gut–immune–brain axis in health and disease. Cell Mol Immunol. (2025). doi: 10.1038/s41423-025-01333-3
15. Dantzer R, O’Connor JC, Freund GG, Johnson RW, and Kelley KW. From inflammation to sickness and depression: when the immune system subjugates the brain. Nat Rev Neurosci. (2008) 9:46–56. doi: 10.1038/nrn2297
16. Jiao W, Lin J, Deng Y, Ji Y, Liang C, Wei S, et al. The immunological perspective of major depressive disorder: unveiling the interactions between central and peripheral immune mechanisms. J Neuroinflamm. (2025) 22:10. doi: 10.1186/s12974-024-03312-3
17. Silva YP, Bernardi A, and Frozza RL. The role of short-chain fatty acids from gut microbiota in gut-brain communication. Front Endocrinol (Lausanne). (2020) 11:25. doi: 10.3389/fendo.2020.00025
18. Gao K, Mu C-L, Farzi A, and Zhu W-Y. Tryptophan metabolism: A link between the gut microbiota and brain. Adv Nutr. (2020) 11:709–23. doi: 10.1093/advances/nmz127
19. Cryan JF, O’Riordan KJ, Cowan CSM, Sandhu KV, Bastiaanssen TFS, Boehme M, et al. The microbiota-gut-brain axis. Physiol Rev. (2019) 99:1877–2013. doi: 10.1152/physrev.00018.2018
20. Stone TW and Darlington LG. Endogenous kynurenines as targets for drug discovery and development. Nat Rev Drug Discov. (2002) 1:609–20. doi: 10.1038/nrd870
21. Schwarcz R and Stone TW. The kynurenine pathway and the brain: Challenges, controversies and promises. Neuropharmacology. (2017) 112:237–47. doi: 10.1016/j.neuropharm.2016.08.003
22. Zaragozá R. Transport of amino acids across the blood-brain barrier. Front Physiol. (2020) 11:973. doi: 10.3389/fphys.2020.00973
23. Margolis KG, Cryan JF, and Mayer EA. The microbiota-gut-brain axis: from motility to mood. Gastroenterology. (2021) 160:1486–501. doi: 10.1053/j.gastro.2020.10.066
24. Felger JC and Lotrich FE. Inflammatory cytokines in depression: neurobiological mechanisms and therapeutic implications. Neuroscience. (2013) 246:199–229. doi: 10.1016/j.neuroscience.2013.04.060
25. Kaltenboeck A and Harmer C. The neuroscience of depressive disorders: A brief review of the past and some considerations about the future. Brain Neurosci Adv. (2018) 2:2398212818799269. doi: 10.1177/2398212818799269
26. Raison CL and Miller AH. The evolutionary significance of depression in Pathogen Host Defense (PATHOS-D). Mol Psychiatry. (2013) 18:15–37. doi: 10.1038/mp.2012.2
27. Hassamal S. Chronic stress, neuroinflammation, and depression: an overview of pathophysiological mechanisms and emerging anti-inflammatories. Front Psychiatry. (2023) 14:1130989. doi: 10.3389/fpsyt.2023.1130989
28. Caspani G, Kennedy S, Foster JA, and Swann J. Gut microbial metabolites in depression: understanding the biochemical mechanisms. Microb Cell. (2019) 6:454–81. doi: 10.15698/mic2019.10.693
29. Liu L, Wang H, Chen X, Zhang Y, Zhang H, and Xie P. Gut microbiota and its metabolites in depression: from pathogenesis to treatment. EBioMedicine. (2023) 90:104527. doi: 10.1016/j.ebiom.2023.104527
30. Wollmuth EM and Angert ER. Microbial circadian clocks: host-microbe interplay in diel cycles. BMC Microbiol. (2023) 23:124. doi: 10.1186/s12866-023-02839-4
31. Bishehsari F, Voigt RM, and Keshavarzian A. Circadian rhythms and the gut microbiota: from the metabolic syndrome to cancer. Nat Rev Endocrinol. (2020) 16:731–9. doi: 10.1038/s41574-020-00427-4
32. Cai T, Zheng S-P, Shi X, Yuan L-Z, Hu H, Zhou B, et al. Therapeutic effect of fecal microbiota transplantation on chronic unpredictable mild stress-induced depression. Front Cell Infect Microbiol. (2022) 12:900652. doi: 10.3389/fcimb.2022.900652
33. Zhang H, Qin X, Song H, Zhou J, Wei H, Zhang L, et al. A peripheral mechanism of depression: disturbed intestinal epithelial per2 gene expression causes depressive behaviors in mice with circadian rhythm disruption via gut barrier damage and microbiota dysbiosis. Adv Sci (Weinh). (2025):e01818. doi: 10.1002/advs.202501818
34. Kiehn J-T, Faltraco F, Palm D, Thome J, and Oster H. Circadian clocks in the regulation of neurotransmitter systems. Pharmacopsychiatry. (2023) 56:108–17. doi: 10.1055/a-1027-7055
35. Thaiss CA, Levy M, Korem T, Dohnalová L, Shapiro H, Jaitin DA, et al. Microbiota diurnal rhythmicity programs host transcriptome oscillations. Cell. (2016) 167:1495–1510.e12. doi: 10.1016/j.cell.2016.11.003
36. Hu J, Zhang Y, Liu C, Gkaravella A, and Yu J. Effects of microbiota-based interventions on depression and anxiety in children and adolescents-A systematic review. J Pediatr Gastroenterol Nutr. (2025) 81:404–16. doi: 10.1002/jpn3.70092
37. Ullah H, Arbab S, Tian Y, Liu C-Q, Chen Y, Qijie L, et al. The gut microbiota-brain axis in neurological disorder. Front Neurosci. (2023) 17:1225875. doi: 10.3389/fnins.2023.1225875
38. Appleton J. The gut-brain axis: influence of microbiota on mood and mental health. Integr Med (Encinitas). (2018) 17:28–32.
39. Chang L, Wei Y, and Hashimoto K. Brain-gut-microbiota axis in depression: A historical overview and future directions. Brain Res Bull. (2022) 182:44–56. doi: 10.1016/j.brainresbull.2022.02.004
40. Zhao Y, Yang L, Sahakian BJ, Langley C, Zhang W, Kuo K, et al. The brain structure, immunometabolic and genetic mechanisms underlying the association between lifestyle and depression. Nat Ment Health. (2023) 1:736–50. doi: 10.1038/s44220-023-00120-1
41. Simpson CA, Diaz-Arteche C, Eliby D, Schwartz OS, Simmons JG, and Cowan CSM. The gut microbiota in anxiety and depression - A systematic review. Clin Psychol Rev. (2021) 83:101943. doi: 10.1016/j.cpr.2020.101943
42. Cryan JF and Dinan TG. Mind-altering microorganisms: the impact of the gut microbiota on brain and behaviour. Nat Rev Neurosci. (2012) 13:701–12. doi: 10.1038/nrn3346
43. Herman JP, McKlveen JM, Ghosal S, Kopp B, Wulsin A, Makinson R, et al. Regulation of the hypothalamic-pituitary-adrenocortical stress response. Compr Physiol. (2016) 6:603–21. doi: 10.1002/cphy.c150015
44. Bautista J, Ojeda-Mosquera S, Ordóñez-Lozada D, and López-Cortés A. Peripheral clocks and systemic zeitgeber interactions: from molecular mechanisms to circadian precision medicine. Front Endocrinol (Lausanne). (2025) 16:1606242. doi: 10.3389/fendo.2025.1606242
45. Geng F, Zhao N, and Ren Q. Circadian rhythm, microglia-mediated neuroinflammation, and Alzheimer’s disease. Neurosci Biobehav Rev. (2025) 170:106044. doi: 10.1016/j.neubiorev.2025.106044
46. Bertollo AG, Santos CF, Bagatini MD, and Ignácio ZM. Hypothalamus-pituitary-adrenal and gut-brain axes in biological interaction pathway of the depression. Front Neurosci. (2025) 19:1541075. doi: 10.3389/fnins.2025.1541075
47. Warren A, Nyavor Y, Beguelin A, and Frame LA. Dangers of the chronic stress response in the context of the microbiota-gut-immune-brain axis and mental health: a narrative review. Front Immunol. (2024) 15:1365871. doi: 10.3389/fimmu.2024.1365871
48. Luczynski P, McVey Neufeld K-A, Oriach CS, Clarke G, Dinan TG, and Cryan JF. Growing up in a bubble: using germ-free animals to assess the influence of the gut microbiota on brain and behavior. Int J Neuropsychopharmacol. (2016) 19. doi: 10.1093/ijnp/pyw020
49. Rosell-Cardona C, Cryan JF, Clarke G, and Kittel-Schneider S. Host-microbiome relationship in depression: can human induced pluripotent stem cells play a role in unravelling mechanisms? NPJ Biofilms Microbiomes. (2025) 11:117. doi: 10.1038/s41522-025-00749-z
50. Tofani GSS, Leigh S-J, Gheorghe CE, Bastiaanssen TFS, Wilmes L, Sen P, et al. Gut microbiota regulates stress responsivity via the circadian system. Cell Metab. (2025) 37:138–153.e5. doi: 10.1016/j.cmet.2024.10.003
51. Gheorghe CE, Leigh S-J, Tofani GSS, Bastiaanssen TFS, Lyte JM, Gardellin E, et al. The microbiota drives diurnal rhythms in tryptophan metabolism in the stressed gut. Cell Rep. (2024) 43:114079. doi: 10.1016/j.celrep.2024.114079
52. Wang Y, Cui P, Cao M, Ai L, Zeng L, Li X, et al. Chronic restraint stress affects the diurnal rhythms of gut microbial composition and metabolism in a mouse model of depression. BMC Microbiol. (2025) 25:38. doi: 10.1186/s12866-025-03764-4
53. Lotti S, Dinu M, Colombini B, Amedei A, and Sofi F. Circadian rhythms, gut microbiota, and diet: Possible implications for health. Nutr Metab Cardiovasc Dis. (2023) 33:1490–500. doi: 10.1016/j.numecd.2023.05.009
54. Martin CR, Osadchiy V, Kalani A, and Mayer EA. The brain-gut-microbiome axis. Cell Mol Gastroenterol Hepatol. (2018) 6:133–48. doi: 10.1016/j.jcmgh.2018.04.003
55. Samuthpongtorn C, Chan AA, Ma W, Wang F, Nguyen LH, Wang DD, et al. prausnitzii potentially modulates the association between citrus intake and depression. Microbiome. (2024) 12:237. doi: 10.1186/s40168-024-01961-3
56. Chakrabarti A, Geurts L, Hoyles L, Iozzo P, Kraneveld AD, La Fata G, et al. The microbiota-gut-brain axis: pathways to better brain health. Perspectives on what we know, what we need to investigate and how to put knowledge into practice. Cell Mol Life Sci. (2022) 79:80. doi: 10.1007/s00018-021-04060-w
57. Ramadan YN, Alqifari SF, Alshehri K, Alhowiti A, Mirghani H, Alrasheed T, et al. Microbiome gut-brain-axis: impact on brain development and mental health. Mol Neurobiol. (2025) 62:10813–33. doi: 10.1007/s12035-025-04846-0
58. Breit S, Kupferberg A, Rogler G, and Hasler G. Vagus nerve as modulator of the brain-gut axis in psychiatric and inflammatory disorders. Front Psychiatry. (2018) 9:44. doi: 10.3389/fpsyt.2018.00044
59. Bravo JA, Forsythe P, Chew MV, Escaravage E, Savignac HM, Dinan TG, et al. Ingestion of Lactobacillus strain regulates emotional behavior and central GABA receptor expression in a mouse via the vagus nerve. Proc Natl Acad Sci USA. (2011) 108:16050–5. doi: 10.1073/pnas.1102999108
60. Strandwitz P, Kim KH, Terekhova D, Liu JK, Sharma A, Levering J, et al. GABA-modulating bacteria of the human gut microbiota. Nat Microbiol. (2019) 4:396–403. doi: 10.1038/s41564-018-0307-3
61. Carabotti M, Scirocco A, Maselli MA, and Severi C. The gut-brain axis: interactions between enteric microbiota, central and enteric nervous systems. Ann Gastroenterol. (2015) 28:203–9.
62. Bercik P, Denou E, Collins J, Jackson W, Lu J, Jury J, et al. The intestinal microbiota affect central levels of brain-derived neurotropic factor and behavior in mice. Gastroenterology. (2011) 141:599–609. doi: 10.1053/j.gastro.2011.04.052
63. Foster JA, Rinaman L, and Cryan JF. Stress & the gut-brain axis: Regulation by the microbiome. Neurobiol Stress. (2017) 7:124–36. doi: 10.1016/j.ynstr.2017.03.001
64. Braga JD, Thongngam M, and Kumrungsee T. Gamma-aminobutyric acid as a potential postbiotic mediator in the gut-brain axis. NPJ Sci Food. (2024) 8:16. doi: 10.1038/s41538-024-00253-2
65. Réus GZ, Jansen K, Titus S, Carvalho AF, Gabbay V, and Quevedo J. Kynurenine pathway dysfunction in the pathophysiology and treatment of depression: Evidences from animal and human studies. J Psychiatr Res. (2015) 68:316–28. doi: 10.1016/j.jpsychires.2015.05.007
66. Felger JC and Treadway MT. Inflammation effects on motivation and motor activity: role of dopamine. Neuropsychopharmacology. (2017) 42:216–41. doi: 10.1038/npp.2016.143
67. Felger JC. The role of dopamine in inflammation-associated depression: mechanisms and therapeutic implications. Curr Top Behav Neurosci. (2017) 31:199–219. doi: 10.1007/7854_2016_13
68. Salminen A. Activation of aryl hydrocarbon receptor (AhR) in Alzheimer’s disease: role of tryptophan metabolites generated by gut host-microbiota. J Mol Med. (2023) 101:201–22. doi: 10.1007/s00109-023-02289-5
69. Hu J, Ji W-J, Liu G-Y, Su X-H, Zhu J-M, Hong Y, et al. IDO1 modulates pain sensitivity and comorbid anxiety in chronic migraine through microglial activation and synaptic pruning. J Neuroinflamm. (2025) 22:42. doi: 10.1186/s12974-025-03367-w
70. Authement ME, Kodangattil JN, Gouty S, Rusnak M, Symes AJ, Cox BM, et al. Histone deacetylase inhibition rescues maternal deprivation-induced GABAergic metaplasticity through restoration of AKAP signaling. Neuron. (2015) 86:1240–52. doi: 10.1016/j.neuron.2015.05.024
71. Wichers MC and Maes M. The role of indoleamine 2,3-dioxygenase (IDO) in the pathophysiology of interferon-alpha-induced depression. J Psychiatry Neurosci. (2004) 29:11–7. doi: 10.1139/jpn.0402
72. Kim Y-K and Jeon SW. Neuroinflammation and the immune-kynurenine pathway in anxiety disorders. Curr Neuropharmacol. (2018) 16:574–82. doi: 10.2174/1570159X15666170913110426
73. Dolšak A, Gobec S, and Sova M. Indoleamine and tryptophan 2,3-dioxygenases as important future therapeutic targets. Pharmacol Ther. (2021) 221:107746. doi: 10.1016/j.pharmthera.2020.107746
74. Savitz J, Drevets WC, Smith CM, Victor TA, Wurfel BE, Bellgowan PSF, et al. Putative neuroprotective and neurotoxic kynurenine pathway metabolites are associated with hippocampal and amygdalar volumes in subjects with major depressive disorder. Neuropsychopharmacology. (2015) 40:463–71. doi: 10.1038/npp.2014.194
75. Lin P, Li D, Shi Y, Li Q, Guo X, Dong K, et al. Dysbiosis of the gut microbiota and kynurenine (Kyn) pathway activity as potential biomarkers in patients with major depressive disorder. Nutrients. (2023) 15:1752. doi: 10.3390/nu15071752
76. Brown J, Robusto B, and Morel L. Intestinal dysbiosis and tryptophan metabolism in autoimmunity. Front Immunol. (2020) 11:1741. doi: 10.3389/fimmu.2020.01741
77. Kim Y-I, Kim E, Lee Y, and Park J. Role of late-night eating in circadian disruption and depression: a review of emotional health impacts. Phys Act Nutr. (2025) 29:18–24. doi: 10.20463/pan.2025.0003
78. Dos Santos A and Vasylyshyn A. The modulatory role of short-chain fatty acids on peripheral circadian gene expression: a systematic review. Front Physiol. (2025) 16:1595057. doi: 10.3389/fphys.2025.1595057
79. Bajaj P and Sharma M. Chrononutrition and gut health: exploring the relationship between meal timing and the gut microbiome. Curr Nutr Rep. (2025) 14:79. doi: 10.1007/s13668-025-00670-z
80. Kuang Z, Wang Y, Li Y, Ye C, Ruhn KA, Behrendt CL, et al. The intestinal microbiota programs diurnal rhythms in host metabolism through histone deacetylase 3. Science. (2019) 365:1428–34. doi: 10.1126/science.aaw3134
81. Teichman EM, O’Riordan KJ, Gahan CGM, Dinan TG, and Cryan JF. When rhythms meet the blues: circadian interactions with the microbiota-gut-brain axis. Cell Metab. (2020) 31:448–71. doi: 10.1016/j.cmet.2020.02.008
82. Cheng J, Hu H, Ju Y, Liu J, Wang M, Liu B, et al. Gut microbiota-derived short-chain fatty acids and depression: deep insight into biological mechanisms and potential applications. Gen Psych. (2024) 37:e101374. doi: 10.1136/gpsych-2023-101374
83. Agus A, Planchais J, and Sokol H. Gut microbiota regulation of tryptophan metabolism in health and disease. Cell Host Microbe. (2018) 23:716–24. doi: 10.1016/j.chom.2018.05.003
84. Savitz J. The kynurenine pathway: a finger in every pie. Mol Psychiatry. (2020) 25:131–47. doi: 10.1038/s41380-019-0414-4
85. Bailey MT, Dowd SE, Galley JD, Hufnagle AR, Allen RG, and Lyte M. Exposure to a social stressor alters the structure of the intestinal microbiota: implications for stressor-induced immunomodulation. Brain Behav Immun. (2011) 25:397–407. doi: 10.1016/j.bbi.2010.10.023
86. Keller J, Gomez R, Williams G, Lembke A, Lazzeroni L, Murphy GM, et al. HPA axis in major depression: cortisol, clinical symptomatology and genetic variation predict cognition. Mol Psychiatry. (2017) 22:527–36. doi: 10.1038/mp.2016.120
87. Rusch JA, Layden BT, and Dugas LR. Signalling cognition: the gut microbiota and hypothalamic-pituitary-adrenal axis. Front Endocrinol (Lausanne). (2023) 14:1130689. doi: 10.3389/fendo.2023.1130689
88. Rosin S, Xia K, Azcarate-Peril MA, Carlson AL, Propper CB, Thompson AL, et al. A preliminary study of gut microbiome variation and HPA axis reactivity in healthy infants. Psychoneuroendocrinology. (2021) 124:105046. doi: 10.1016/j.psyneuen.2020.105046
89. Hueston CM, Cryan JF, and Nolan YM. Stress and adolescent hippocampal neurogenesis: diet and exercise as cognitive modulators. Transl Psychiatry. (2017) 7:e1081. doi: 10.1038/tp.2017.48
90. Provensi G, Schmidt SD, Boehme M, Bastiaanssen TFS, Rani B, Costa A, et al. Preventing adolescent stress-induced cognitive and microbiome changes by diet. Proc Natl Acad Sci USA. (2019) 116:9644–51. doi: 10.1073/pnas.1820832116
91. Langgartner D, Amoroso M, Kempter E, Kustermann M, Scheurer J, Lowry CA, et al. Mycobacterium vaccae protects against glucocorticoid resistance resulting from combined physical and psychosocial trauma in mice. Brain Behav Immun. (2023) 109:221–34. doi: 10.1016/j.bbi.2023.01.018
92. Dinan TG and Cryan JF. Regulation of the stress response by the gut microbiota: implications for psychoneuroendocrinology. Psychoneuroendocrinology. (2012) 37:1369–78. doi: 10.1016/j.psyneuen.2012.03.007
93. Codoñer-Franch P, Gombert M, Martínez-Raga J, and Cenit MC. Circadian disruption and mental health: the chronotherapeutic potential of microbiome-based and dietary strategies. Int J Mol Sci. (2023) 24:7579. doi: 10.3390/ijms24087579
94. Dacaya P, Sarapis K, and Moschonis G. The role and mechanisms of probiotic supplementation on depressive symptoms: A narrative review. Curr Nutr Rep. (2025) 14:53. doi: 10.1007/s13668-025-00644-1
95. Moloney RD, Johnson AC, O’Mahony SM, Dinan TG, Greenwood-Van Meerveld B, and Cryan JF. Stress and the microbiota-gut-brain axis in visceral pain: relevance to irritable bowel syndrome. CNS Neurosci Ther. (2016) 22:102–17. doi: 10.1111/cns.12490
96. Kato-Kataoka A, Nishida K, Takada M, Suda K, Kawai M, Shimizu K, et al. Fermented milk containing Lactobacillus casei strain Shirota prevents the onset of physical symptoms in medical students under academic examination stress. Benef Microbes. (2016) 7:153–6. doi: 10.3920/BM2015.0100
97. Sawada D, Kawai T, Nishida K, Kuwano Y, Fujiwara S, and Rokutan K. Daily intake of Lactobacillus gasseri CP2305 improves mental, physical, and sleep quality among Japanese medical students enrolled in a cadaver dissection course. J Funct Foods. (2017) 31:188–97. doi: 10.1016/j.jff.2017.01.042
98. Allen AP, Hutch W, Borre YE, Kennedy PJ, Temko A, Boylan G, et al. Bifidobacterium longum 1714 as a translational psychobiotic: modulation of stress, electrophysiology and neurocognition in healthy volunteers. Transl Psychiatry. (2016) 6:e939. doi: 10.1038/tp.2016.191
99. Walden KE, Moon JM, Hagele AM, Allen LE, Gaige CJ, Krieger JM, et al. A randomized controlled trial to examine the impact of a multi-strain probiotic on self-reported indicators of depression, anxiety, mood, and associated biomarkers. Front Nutr. (2023) 10:1219313. doi: 10.3389/fnut.2023.1219313
100. Messaoudi M, Lalonde R, Violle N, Javelot H, Desor D, Nejdi A, et al. Assessment of psychotropic-like properties of a probiotic formulation (Lactobacillus helveticus R0052 and Bifidobacterium longum R0175) in rats and human subjects. Br J Nutr. (2011) 105:755–64. doi: 10.1017/S0007114510004319
101. Cao Q, Shen M, Li R, Liu Y, Zeng Z, Zhou J, et al. Elucidating the specific mechanisms of the gut-brain axis: the short-chain fatty acids-microglia pathway. J Neuroinflamm. (2025) 22:133. doi: 10.1186/s12974-025-03454-y
102. Jian C, Carpén N, Helve O, de Vos WM, Korpela K, and Salonen A. Early-life gut microbiota and its connection to metabolic health in children: Perspective on ecological drivers and need for quantitative approach. EBioMedicine. (2021) 69:103475. doi: 10.1016/j.ebiom.2021.103475
103. Nader N, Chrousos GP, and Kino T. Interactions of the circadian CLOCK system and the HPA axis. Trends Endocrinol Metab. (2010) 21:277–86. doi: 10.1016/j.tem.2009.12.011
104. O’Byrne NA, Yuen F, Butt WZ, and Liu PY. Sleep and circadian regulation of cortisol: A short review. Curr Opin Endocrine Metab Res. (2021) 18:178–86. doi: 10.1016/j.coemr.2021.03.011
105. Liu PY. Rhythms in cortisol mediate sleep and circadian impacts on health. Sleep. (2024) 47:zsae151. doi: 10.1093/sleep/zsae151
106. Bautista J, Echeverría CE, Maldonado-Noboa I, Ojeda- Mosquera S, Hidalgo-Tinoco C, and López-Cortés A. The human microbiome in clinical translation: from bench to bedside. Front Microbiol. (2025). doi: 10.3389/fmicb.2025.1632435
107. Oroojzadeh P, Bostanabad SY, and Lotfi H. Psychobiotics: the influence of gut microbiota on the gut-brain axis in neurological disorders. J Mol Neurosci. (2022) 72:1952–64. doi: 10.1007/s12031-022-02053-3
108. Müller L and Di Benedetto S. Bridging the brain and gut: neuroimmune mechanisms of neuroinflammation and therapeutic insights. Front Cell Neurosci. (2025) 19:1590002. doi: 10.3389/fncel.2025.1590002
109. Shen Y, Fan N, Ma S, Cheng X, Yang X, and Wang G. Gut microbiota dysbiosis: pathogenesis, diseases, prevention, and therapy. MedComm. (2025) 6:e70168. doi: 10.1002/mco2.70168
110. Chen R, Zou J, Chen J, Zhong X, Kang R, and Tang D. Pattern recognition receptors: function, regulation and therapeutic potential. Signal Transduct Target Ther. (2025) 10:216. doi: 10.1038/s41392-025-02264-1
111. Zhao M, Chu J, Feng S, Guo C, Xue B, He K, et al. Immunological mechanisms of inflammatory diseases caused by gut microbiota dysbiosis: A review. BioMed Pharmacother. (2023) 164:114985. doi: 10.1016/j.biopha.2023.114985
112. Warren A, Nyavor Y, Zarabian N, Mahoney A, and Frame LA. The microbiota-gut-brain-immune interface in the pathogenesis of neuroinflammatory diseases: a narrative review of the emerging literature. Front Immunol. (2024) 15:1365673. doi: 10.3389/fimmu.2024.1365673
113. Tang W, Zhu H, Feng Y, Guo R, and Wan D. The impact of gut microbiota disorders on the blood-brain barrier. Infect Drug Resist. (2020) 13:3351–63. doi: 10.2147/IDR.S254403
114. Menard C, Pfau ML, Hodes GE, Kana V, Wang VX, Bouchard S, et al. Social stress induces neurovascular pathology promoting depression. Nat Neurosci. (2017) 20:1752–60. doi: 10.1038/s41593-017-0010-3
115. Dion-Albert L, Cadoret A, Doney E, Kaufmann FN, Dudek KA, Daigle B, et al. Vascular and blood-brain barrier-related changes underlie stress responses and resilience in female mice and depression in human tissue. Nat Commun. (2022) 13:164. doi: 10.1038/s41467-021-27604-x
116. Banks WA. Blood-brain barrier transport of cytokines: a mechanism for neuropathology. Curr Pharm Des. (2005) 11:973–84. doi: 10.2174/1381612053381684
117. He M, Wei W, Zhang Y, Xiang Z, Peng D, Kasimumali A, et al. Gut microbial metabolites SCFAs and chronic kidney disease. J Transl Med. (2024) 22:172. doi: 10.1186/s12967-024-04974-6
118. Sankarganesh P, Bhunia A, Ganesh Kumar A, Babu AS, Gopukumar ST, and Lokesh E. Short-chain fatty acids (SCFAs) in gut health: Implications for drug metabolism and therapeutics. Med Microecology. (2025) 25:100139. doi: 10.1016/j.medmic.2025.100139
119. Drevets WC, Wittenberg GM, Bullmore ET, and Manji HK. Immune targets for therapeutic development in depression: towards precision medicine. Nat Rev Drug Discov. (2022) 21:224–44. doi: 10.1038/s41573-021-00368-1
120. Li S. Modulation of immunity by tryptophan microbial metabolites. Front Nutr. (2023) 10:1209613. doi: 10.3389/fnut.2023.1209613
121. Bautista J, Coral-Riofrio EC, Suárez Urresta S, Palacios-Zavala D, Echeverría CE, Araujo-Abad S, et al. Neurodegeneration rewires the tumor microenvironment via the neuro–immune–cancer axis. iScience. (2025) 28:113550. doi: 10.1016/j.isci.2025.113550
122. Kyriakidis NC, Echeverría CE, Bautista J, Rivera-Orellana S, Ramos-Medina MJ, Salazar-Santoliva C, et al. Reprogramming cancer immunity with next-generation combination therapies. Front Cell Dev Biol. (2025) 13:1652047. doi: 10.3389/fcell.2025.1652047
123. Bautista J, Ávila-Coello D, Hidalgo-Tinoco C, Bueno-Miño J, and López-Cortés A. Unraveling the gut-brain-immune interplay in herpes simplex virus-associated neurodegeneration. J Med Virol. (2025) 97:e70504. doi: 10.1002/jmv.70504
124. Pan Y, Chen X-Y, Zhang Q-Y, and Kong L-D. Microglial NLRP3 inflammasome activation mediates IL-1β-related inflammation in prefrontal cortex of depressive rats. Brain Behav Immun. (2014) 41:90–100. doi: 10.1016/j.bbi.2014.04.007
125. Pinzón-Fernández MV, Saavedra-Torres JS, López Garzón NA, Pachon-Bueno JS, Tamayo-Giraldo FJ, Rojas Gomez MC, et al. NLRP3 and beyond: inflammasomes as central cellular hub and emerging therapeutic target in inflammation and disease. Front Immunol. (2025) 16:1624770. doi: 10.3389/fimmu.2025.1624770
126. Hodes GE, Pfau ML, Leboeuf M, Golden SA, Christoffel DJ, Bregman D, et al. Individual differences in the peripheral immune system promote resilience versus susceptibility to social stress. Proc Natl Acad Sci USA. (2014) 111:16136–41. doi: 10.1073/pnas.1415191111
127. Yang D, Wang Z, Chen Y, Guo Q, and Dong Y. Interactions between gut microbes and NLRP3 inflammasome in the gut-brain axis. Comput Struct Biotechnol J. (2023) 21:2215–27. doi: 10.1016/j.csbj.2023.03.017
128. Feng X, Jia M, Cai M, Zhu T, Hashimoto K, and Yang J-J. Central-peripheral neuroimmune dynamics in psychological stress and depression: insights from current research. Mol Psychiatry. (2025) 30:4881–98. doi: 10.1038/s41380-025-03085-y
129. Setiawan E, Wilson AA, Mizrahi R, Rusjan PM, Miler L, Rajkowska G, et al. Role of translocator protein density, a marker of neuroinflammation, in the brain during major depressive episodes. JAMA Psychiatry. (2015) 72:268–75. doi: 10.1001/jamapsychiatry.2014.2427
130. Erny D, Hrabě de Angelis AL, Jaitin D, Wieghofer P, Staszewski O, David E, et al. Host microbiota constantly control maturation and function of microglia in the CNS. Nat Neurosci. (2015) 18:965–77. doi: 10.1038/nn.4030
131. Chen T, Dai Y, Hu C, Lin Z, Wang S, Yang J, et al. Cellular and molecular mechanisms of the blood-brain barrier dysfunction in neurodegenerative diseases. Fluids Barriers CNS. (2024) 21:60. doi: 10.1186/s12987-024-00557-1
132. Mou Y, Du Y, Zhou L, Yue J, Hu X, Liu Y, et al. Gut microbiota interact with the brain through systemic chronic inflammation: implications on neuroinflammation, neurodegeneration, and aging. Front Immunol. (2022) 13:796288. doi: 10.3389/fimmu.2022.796288
133. Fleshner M, Frank M, and Maier SF. Danger signals and inflammasomes: stress-evoked sterile inflammation in mood disorders. Neuropsychopharmacology. (2017) 42:36–45. doi: 10.1038/npp.2016.125
134. Padhi P, Worth C, Zenitsky G, Jin H, Sambamurti K, Anantharam V, et al. Mechanistic insights into gut microbiome dysbiosis-mediated neuroimmune dysregulation and protein misfolding and clearance in the pathogenesis of chronic neurodegenerative disorders. Front Neurosci. (2022) 16:836605. doi: 10.3389/fnins.2022.836605
135. Jiao H, Kalsbeek A, and Yi C-X. Microglia, circadian rhythm and lifestyle factors. Neuropharmacology. (2024) 257:110029. doi: 10.1016/j.neuropharm.2024.110029
136. Guzmán-Ruiz MA, Guerrero-Vargas NN, Lagunes-Cruz A, González-González S, García-Aviles JE, Hurtado-Alvarado G, et al. Circadian modulation of microglial physiological processes and immune responses. Glia. (2023) 71:155–67. doi: 10.1002/glia.24261
137. Fonken LK, Frank MG, Kitt MM, Barrientos RM, Watkins LR, and Maier SF. Microglia inflammatory responses are controlled by an intrinsic circadian clock. Brain Behav Immun. (2015) 45:171–9. doi: 10.1016/j.bbi.2014.11.009
138. Zeng Y, Guo Z, Wu M, Chen F, and Chen L. Circadian rhythm regulates the function of immune cells and participates in the development of tumors. Cell Death Discov. (2024) 10:199. doi: 10.1038/s41420-024-01960-1
139. Dong TS and Mayer E. Advances in brain-gut-microbiome interactions: A comprehensive update on signaling mechanisms, disorders, and therapeutic implications. Cell Mol Gastroenterol Hepatol. (2024) 18:1–13. doi: 10.1016/j.jcmgh.2024.01.024
140. Lee S-H, Han C, and Shin C. IUPHAR review: Microbiota-gut-brain axis and its role in neuropsychiatric disorders. Pharmacol Res. (2025) 216:107749. doi: 10.1016/j.phrs.2025.107749
141. O’Riordan KJ, Collins MK, Moloney GM, Knox EG, Aburto MR, Fülling C, et al. Short chain fatty acids: Microbial metabolites for gut-brain axis signalling. Mol Cell Endocrinol. (2022) 546:111572. doi: 10.1016/j.mce.2022.111572
142. Dalile B, Van Oudenhove L, Vervliet B, and Verbeke K. The role of short-chain fatty acids in microbiota-gut-brain communication. Nat Rev Gastroenterol Hepatol. (2019) 16:461–78. doi: 10.1038/s41575-019-0157-3
143. Kim MH, Kang SG, Park JH, Yanagisawa M, and Kim CH. Short-chain fatty acids activate GPR41 and GPR43 on intestinal epithelial cells to promote inflammatory responses in mice. Gastroenterology. (2013) 145:396–406.e1. doi: 10.1053/j.gastro.2013.04.056
144. Wu J-T, Sun C-L, Lai T-T, Liou C-W, Lin Y-Y, Xue J-Y, et al. Oral short-chain fatty acids administration regulates innate anxiety in adult microbiome-depleted mice. Neuropharmacology. (2022) 214:109140. doi: 10.1016/j.neuropharm.2022.109140
145. Nunzi E, Pariano M, Costantini C, Garaci E, Puccetti P, and Romani L. Host-microbe serotonin metabolism. Trends Endocrinol Metab. (2025) 36:83–95. doi: 10.1016/j.tem.2024.07.014
146. Inam ME, Fernandes BS, Salagre E, Grande I, Vieta E, Quevedo J, et al. The kynurenine pathway in major depressive disorder, bipolar disorder, and schizophrenia: a systematic review and meta-analysis of cerebrospinal fluid studies. Rev Bras Psiquiatr. (2023) 45:343–55. doi: 10.47626/1516-4446-2022-2973
147. Darmanto AG, Yen T-L, Jan J-S, Linh TTD, Taliyan R, Yang C-H, et al. Beyond metabolic messengers: Bile acids and TGR5 as pharmacotherapeutic intervention for psychiatric disorders. Pharmacol Res. (2025) 211:107564. doi: 10.1016/j.phrs.2024.107564
148. Sabahat SE, Saqib M, Talib M, Shaikh TG, Khan T, and Kailash SJ. Bile acid modulation by gut microbiota: a bridge to understanding cognitive health. Ann Med Surg (Lond). (2024) 86:5410–5. doi: 10.1097/MS9.0000000000002433
149. Chen S, Shao Q, Chen J, Lv X, Ji J, Liu Y, et al. Bile acid signalling and its role in anxiety disorders. Front Endocrinol (Lausanne). (2023) 14:1268865. doi: 10.3389/fendo.2023.1268865
150. Chiang JYL and Ferrell JM. Bile acids as metabolic regulators and nutrient sensors. Annu Rev Nutr. (2019) 39:175–200. doi: 10.1146/annurev-nutr-082018-124344
151. Ye X, Li H, Anjum K, Zhong X, Miao S, Zheng G, et al. Dual role of indoles derived from intestinal microbiota on human health. Front Immunol. (2022) 13:903526. doi: 10.3389/fimmu.2022.903526
152. Mason S. Lactate shuttles in neuroenergetics-homeostasis, allostasis and beyond. Front Neurosci. (2017) 11:43. doi: 10.3389/fnins.2017.00043
153. Figley CR. Lactate transport and metabolism in the human brain: implications for the astrocyte-neuron lactate shuttle hypothesis. J Neurosci. (2011) 31:4768–70. doi: 10.1523/JNEUROSCI.6612-10.2011
154. Zhang M, Zhou C, Li X, Li H, Han Q, Chen Z, et al. Interactions between gut microbiota, host circadian rhythms, and metabolic diseases. Adv Nutr. (2025) 16:100416. doi: 10.1016/j.advnut.2025.100416
155. Zeb F, Osaili T, Obaid RS, Naja F, Radwan H, Cheikh Ismail L, et al. Gut microbiota and time-restricted feeding/eating: A targeted biomarker and approach in precision nutrition. Nutrients. (2023) 15:259. doi: 10.3390/nu15020259
156. Tang Y, Fu A, Wang L, and Ge Q. Microbiota-dependent metabolites - New engine for T cell warriors. Gut Microbes. (2025) 17:2523815. doi: 10.1080/19490976.2025.2523815
157. Brooks JF, Behrendt CL, Ruhn KA, Lee S, Raj P, Takahashi JS, et al. The microbiota coordinates diurnal rhythms in innate immunity with the circadian clock. Cell. (2021) 184:4154–4167.e12. doi: 10.1016/j.cell.2021.07.001
158. Pérez-Villa A, Echeverría-Garcés G, Ramos-Medina MJ, Prathap L, Martínez-López M, Ramírez-Sánchez D, et al. Integrated multi-omics analysis reveals the molecular interplay between circadian clocks and cancer pathogenesis. Sci Rep. (2023) 13:14198. doi: 10.1038/s41598-023-39401-1
159. Litichevskiy L and Thaiss CA. The oscillating gut microbiome and its effects on host circadian biology. Annu Rev Nutr. (2022) 42:145–64. doi: 10.1146/annurev-nutr-062320-111321
160. Thaiss CA, Zeevi D, Levy M, Zilberman-Schapira G, Suez J, Tengeler AC, et al. Transkingdom control of microbiota diurnal oscillations promotes metabolic homeostasis. Cell. (2014) 159:514–29. doi: 10.1016/j.cell.2014.09.048
161. Meyer N, Lok R, Schmidt C, Kyle SD, McClung CA, Cajochen C, et al. The sleep-circadian interface: A window into mental disorders. Proc Natl Acad Sci USA. (2024) 121:e2214756121. doi: 10.1073/pnas.2214756121
162. Kinlein SA and Karatsoreos IN. The hypothalamic-pituitary-adrenal axis as a substrate for stress resilience: Interactions with the circadian clock. Front Neuroendocrinol. (2020) 56:100819. doi: 10.1016/j.yfrne.2019.100819
163. Cao Y, Cheng Y, Pan W, Diao J, Sun L, and Meng M. Gut microbiota variations in depression and anxiety: a systematic review. BMC Psychiatry. (2025) 25:443. doi: 10.1186/s12888-025-06871-8
164. Jiang H, Ling Z, Zhang Y, Mao H, Ma Z, Yin Y, et al. Altered fecal microbiota composition in patients with major depressive disorder. Brain Behav Immun. (2015) 48:186–94. doi: 10.1016/j.bbi.2015.03.016
165. Zhao H, Jin K, Jiang C, Pan F, Wu J, Luan H, et al. A pilot exploration of multi-omics research of gut microbiome in major depressive disorders. Transl Psychiatry. (2022) 12:8. doi: 10.1038/s41398-021-01769-x
166. Sanada K, Nakajima S, Kurokawa S, Barceló-Soler A, Ikuse D, Hirata A, et al. Gut microbiota and major depressive disorder: A systematic review and meta-analysis. J Affect Disord. (2020) 266:1–13. doi: 10.1016/j.jad.2020.01.102
167. Kelly JR, Borre Y, O’ Brien C, Patterson E, El Aidy S, Deane J, et al. Transferring the blues: Depression-associated gut microbiota induces neurobehavioural changes in the rat. J Psychiatr Res. (2016) 82:109–18. doi: 10.1016/j.jpsychires.2016.07.019
168. Gao M, Wang J, Liu P, Tu H, Zhang R, Zhang Y, et al. Gut microbiota composition in depressive disorder: a systematic review, meta-analysis, and meta-regression. Transl Psychiatry. (2023) 13:379. doi: 10.1038/s41398-023-02670-5
169. Zheng P, Zeng B, Zhou C, Liu M, Fang Z, Xu X, et al. Gut microbiome remodeling induces depressive-like behaviors through a pathway mediated by the host’s metabolism. Mol Psychiatry. (2016) 21:786–96. doi: 10.1038/mp.2016.44
170. Cheung SG, Goldenthal AR, Uhlemann A-C, Mann JJ, Miller JM, and Sublette ME. Systematic review of gut microbiota and major depression. Front Psychiatry. (2019) 10:34. doi: 10.3389/fpsyt.2019.00034
171. Barandouzi ZA, Starkweather AR, Henderson WA, Gyamfi A, and Cong XS. Altered composition of gut microbiota in depression: A systematic review. Front Psychiatry. (2020) 11:541. doi: 10.3389/fpsyt.2020.00541
172. Han B, Lv X, Liu G, Li S, Fan J, Chen L, et al. Gut microbiota-related bile acid metabolism-FXR/TGR5 axis impacts the response to anti-α4β7-integrin therapy in humanized mice with colitis. Gut Microbes. (2023) 15:2232143. doi: 10.1080/19490976.2023.2232143
173. Berding K, Vlckova K, Marx W, Schellekens H, Stanton C, Clarke G, et al. Diet and the microbiota-gut-brain axis: sowing the seeds of good mental health. Adv Nutr. (2021) 12:1239–85. doi: 10.1093/advances/nmaa181
174. Jacka FN, O’Neil A, Opie R, Itsiopoulos C, Cotton S, Mohebbi M, et al. A randomised controlled trial of dietary improvement for adults with major depression (the “SMILES” trial). BMC Med. (2017) 15:23. doi: 10.1186/s12916-017-0791-y
175. Firth J, Marx W, Dash S, Carney R, Teasdale SB, Solmi M, et al. The effects of dietary improvement on symptoms of depression and anxiety: A meta-analysis of randomized controlled trials. Psychosom Med. (2019) 81:265–80. doi: 10.1097/PSY.0000000000000673
176. Lu X, Wu L, Shao L, Fan Y, Pei Y, Lu X, et al. Adherence to the EAT-Lancet diet and incident depression and anxiety. Nat Commun. (2024) 15:5599. doi: 10.1038/s41467-024-49653-8
177. Nikolova VL, Cleare AJ, Young AH, and Stone JM. Updated review and meta-analysis of probiotics for the treatment of clinical depression: adjunctive vs. Stand-alone treatment. J Clin Med. (2021) 10:647. doi: 10.3390/jcm10040647
178. Schaub A-C, Schneider E, Vazquez-Castellanos JF, Schweinfurth N, Kettelhack C, Doll JPK, et al. Clinical, gut microbial and neural effects of a probiotic add-on therapy in depressed patients: a randomized controlled trial. Transl Psychiatry. (2022) 12:227. doi: 10.1038/s41398-022-01977-z
179. Pinto-Sanchez MI, Hall GB, Ghajar K, Nardelli A, Bolino C, Lau JT, et al. Probiotic bifidobacterium longum NCC3001 reduces depression scores and alters brain activity: A pilot study in patients with irritable bowel syndrome. Gastroenterology. (2017) 153:448–459.e8. doi: 10.1053/j.gastro.2017.05.003
180. Sarkar A, Lehto SM, Harty S, Dinan TG, Cryan JF, and Burnet PWJ. Psychobiotics and the manipulation of bacteria-gut-brain signals. Trends Neurosci. (2016) 39:763–81. doi: 10.1016/j.tins.2016.09.002
181. Green JE, Berk M, Mohebbi M, Loughman A, McGuinness AJ, Castle D, et al. Feasibility, acceptability, and safety of faecal microbiota transplantation in the treatment of major depressive disorder: A pilot randomized controlled trial. Can J Psychiatry. (2023) 68:315–26. doi: 10.1177/07067437221150508
182. Raison CL, Rutherford RE, Woolwine BJ, Shuo C, Schettler P, Drake DF, et al. A randomized controlled trial of the tumor necrosis factor antagonist infliximab for treatment-resistant depression: the role of baseline inflammatory biomarkers. JAMA Psychiatry. (2013) 70:31–41. doi: 10.1001/2013.jamapsychiatry.4
183. Collado MC, Engen PA, Bandín C, Cabrera-Rubio R, Voigt RM, Green SJ, et al. Timing of food intake impacts daily rhythms of human salivary microbiota: a randomized, crossover study. FASEB J. (2018) 32:2060–72. doi: 10.1096/fj.201700697RR
184. Kreuzer K, Birkl-Toeglhofer AM, Haybaeck J, Reiter A, Dalkner N, Fellendorf FT, et al. PROVIT-CLOCK: A potential influence of probiotics and vitamin B7 add-on treatment and metabolites on clock gene expression in major depression. Neuropsychobiology. (2024) 83:135–51. doi: 10.1159/000538781
185. Lowe DA, Wu N, Rohdin-Bibby L, Moore AH, Kelly N, Liu YE, et al. Effects of time-restricted eating on weight loss and other metabolic parameters in women and men with overweight and obesity: the TREAT randomized clinical trial. JAMA Intern Med. (2020) 180:1491–9. doi: 10.1001/jamainternmed.2020.4153
186. Chu A, Samman S, Galland B, and Foster M. Daily consumption of Lactobacillus gasseri CP2305 improves quality of sleep in adults - A systematic literature review and meta-analysis. Clin Nutr. (2023) 42:1314–21. doi: 10.1016/j.clnu.2023.06.019
187. Nishida K, Sawada D, Kuwano Y, Tanaka H, and Rokutan K. Health benefits of lactobacillus gasseri CP2305 tablets in young adults exposed to chronic stress: A randomized, double-blind, placebo-controlled study. Nutrients. (2019) 11:1859. doi: 10.3390/nu11081859
188. Wadan A-HS, El-Aziz MKA, and Ellakwa DE-S. The microbiota-gut-brain-axis theory: role of gut microbiota modulators (GMMs) in gastrointestinal, neurological, and mental health disorders. Naunyn Schmiedebergs Arch Pharmacol. (2025) 398:13397–426. doi: 10.1007/s00210-025-04155-2
189. Rees NP, Shaheen W, Quince C, Tselepis C, Horniblow RD, Sharma N, et al. Systematic review of donor and recipient predictive biomarkers of response to faecal microbiota transplantation in patients with ulcerative colitis. EBioMedicine. (2022) 81:104088. doi: 10.1016/j.ebiom.2022.104088
190. Clapp M, Aurora N, Herrera L, Bhatia M, Wilen E, and Wakefield S. Gut microbiota’s effect on mental health: The gut-brain axis. Clin Pract. (2017) 7:987. doi: 10.4081/cp.2017.987
191. Doenyas C, Clarke G, and Cserjési R. Gut–brain axis and neuropsychiatric health: recent advances. Sci Rep. (2025) 15:3415. doi: 10.1038/s41598-025-86858-3
192. Muller E, Shiryan I, and Borenstein E. Multi-omic integration of microbiome data for identifying disease-associated modules. Nat Commun. (2024) 15:2621. doi: 10.1038/s41467-024-46888-3
193. Probul N, Huang Z, Saak CC, Baumbach J, and List M. AI in microbiome-related healthcare. Microb Biotechnol. (2024) 17:e70027. doi: 10.1111/1751-7915.70027
194. Borre YE, O’Keeffe GW, Clarke G, Stanton C, Dinan TG, and Cryan JF. Microbiota and neurodevelopmental windows: implications for brain disorders. Trends Mol Med. (2014) 20:509–18. doi: 10.1016/j.molmed.2014.05.002
195. Cowan CSM, Dinan TG, and Cryan JF. Annual Research Review: Critical windows - the microbiota-gut-brain axis in neurocognitive development. J Child Psychol Psychiatry. (2020) 61:353–71. doi: 10.1111/jcpp.13156
196. Simbirtseva KY and O’Toole PW. Healthy and unhealthy aging and the human microbiome. Annu Rev Med. (2025) 76:115–27. doi: 10.1146/annurev-med-042423-042542
Keywords: anxiety, depression, gut microbiota, neural pathway, immune pathway, endocrine pathway, metabolic pathway, psychobiotics
Citation: Bautista J, Hidalgo-Tinoco C, Di Capua Delgado M, Viteri-Recalde J, Guerra-Guerrero A and López-Cortés A (2025) The gut–brain–circadian axis in anxiety and depression: a critical review. Front. Psychiatry 16:1697200. doi: 10.3389/fpsyt.2025.1697200
Received: 03 September 2025; Accepted: 17 October 2025;
Published: 30 October 2025.
Edited by:
Elvira Anna Carbone, University Magna Graecia of Catanzaro, ItalyReviewed by:
Panida Sittipo, Burapha University, ThailandZahra A. Barandouzi, Nell Hodgson Woodruff School of Nursing, Emory University, United States
Copyright © 2025 Bautista, Hidalgo-Tinoco, Di Capua Delgado, Viteri-Recalde, Guerra-Guerrero and López-Cortés. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Andrés López-Cortés, YWFsYzg0QGdtYWlsLmNvbQ==