 Ádám Kerek1,2*
Ádám Kerek1,2* Bence Török1Ábel Szabó1
Bence Török1Ábel Szabó1 Levente Laczkó3,4,5
Levente Laczkó3,4,5 Gábor Kardos2,3,6,7Krisztián Bányai1,2,8,9
Gábor Kardos2,3,6,7Krisztián Bányai1,2,8,9 Eszter Kaszab2,3,5,10
Eszter Kaszab2,3,5,10 Krisztina Bali2,10
Krisztina Bali2,10 Ákos Jerzsele1,2
Ákos Jerzsele1,2- 1Department of Pharmacology and Toxicology, University of Veterinary Medicine Budapest, Budapest, Hungary
- 2National Laboratory of Infectious Animal Diseases, Antimicrobial Resistance, Veterinary Public Health and Food Chain Safety, University of Veterinary Medicine Budapest, Budapest, Hungary
- 3One Health Institute, University of Debrecen, Debrecen, Hungary
- 4HUN-REN–UD Conservation Biology Research Group, Debrecen, Hungary
- 5Department of Bioinformatics, University of Debrecen, Debrecen, Hungary
- 6National Public Health Center, Budapest, Hungary
- 7Department of Gerontology, Faculty of Health Sciences, University of Debrecen, Nyíregyháza, Hungary
- 8Department of Medical Biology, Medical School, University of Pécs, Pécs, Hungary
- 9HUN-REN Veterinary Medical Research Institute, Budapest, Hungary
- 10Department of Microbiology and Infectious Diseases, University of Veterinary Medicine, Budapest, Hungary
Introduction: Antimicrobial resistance (AMR) poses an escalating global health threat. Potentiated sulphonamides are widely used in both veterinary and human medicine. This study aimed to investigate the in vitro adaptation of Escherichia coli strains to increasing concentrations of potentiated sulphonamides, focusing on co-selection and the genetic mechanisms of resistance.
Methods: The MEGA-plate evolutionary model was used to expose E. coli ATCC 25922 to increased concentrations (0 × to 1000×) of a potentiated sulphonamide. Clones isolated from different concentration zones were analyzed for phenotypic resistance via minimum inhibitory concentration (MIC) testing and genotypically through next-generation sequencing.
Results: In strains adapted to 1000 × potentiated sulphonamide, MIC values significantly increased for most tested antibiotics. Mutations were identified in key folate pathway genes (folP, folA), as well as in efflux pump regulator genes (emrR, marR, acrR, mdtM). These genetic changes indicated activation of multiple multidrug efflux systems, including acrAB-tolC, emrAB-tolC, and mdtEF-tolC. Mutations were also detected in genes associated with SOS response regulation (recN, recQ, uvrB), suggesting stress-induced genetic adaptation. In vitro microevolutionary adaptation to potentiated sulphonamide exposure induced broad genetic changes in E. coli, potentially driving cross-resistance through co-selection. The MEGA-plate method proved to be a robust tool for tracking resistance development and dissecting complex resistance mechanisms.
Discussion: These findings underscore the need for cautious use of combination antimicrobials, as they may elicit pleiotropic resistance responses beyond their intended targets.

Graphical Abstract.
Introduction
The emergence of antibiotic-resistant bacterial pathogens has rapidly evolved into a global public health concern following the widespread use of antimicrobial therapy. In Gram-negative bacteria in particular, resistance is often attributed to the presence of specific genes associated with mobile genetic elements (1). The spread of multidrug-resistant (MDR) Gram-negative strains is now regarded as one of the most pressing global health threats (2).
Since their introduction in 1935, sulfonamides have been widely used for the treatment of infections caused by both Gram-positive and Gram-negative bacteria, as well as protozoa. Due to the rapid emergence of resistance, sulfonamides have typically been administered in combination with diaminopyrimidines — most commonly trimethoprim — since the 1970s (3, 4). In veterinary medicine, they are primarily used for the prevention and treatment of enteric diseases, either alone or in combination with other antimicrobials (5).
Sulfonamides, structural analogs of para-aminobenzoic acid (PABA), inhibit dihydropteroate synthase (DHPS), a key folate biosynthesis enzyme (6). In Escherichia coli, resistance to sulfonamides can arise through chromosomal mutations in the folP gene (7, 8), but more commonly results from the acquisition of alternative sul genes encoding variant DHPS enzymes with reduced affinity for sulfonamides (9, 10). In the second step of folate biosynthesis, dihydrofolate reductase (DHFR) catalyzes the reduction of dihydrofolate to tetrahydrofolate using NADPH, and it is at this enzymatic step that diaminopyrimidines exert their inhibitory action (11). DHFR is essential for the biosynthesis of thymidylate, purines, and several amino acids, including glycine, methionine, serine, and N-formyl-methionyl-tRNA (12, 13). Inhibition of folate-metabolizing enzymes disrupts thymidylate biosynthesis, impairs DNA replication, and ultimately leads to cell death (14).
Despite their continued importance in clinical settings — particularly for urinary tract infections — and in livestock production, widespread and persistent sulfonamide resistance has been documented globally in Gram-negative strains from both humans and animals (15–17). Pathogenic E. coli strains frequently exhibit resistance to sulfonamides (18). Three key genes underlie this resistance: sul1, sul2, and sul3 (5, 19). The sul1 gene is most commonly found on conjugative plasmids and class 1 integrons (17), while sul2 was initially identified on non-conjugative plasmids but is now increasingly detected on conjugative plasmids, often co-occurring with streptomycin resistance (16). The sul3 gene was first recorded in bacterial samples taken from pigs in Switzerland in 2003 and has since been identified in both human and animal isolates (20), and is frequently associated with non-classical integrons (5).
Resistance-conferring mutations are often associated with a so-called “fitness cost” syndrome. As many antibiotics target essential cellular functions, the acquisition of resistance may impair bacterial fitness by imposing additional energetic burdens or by interfering with core biological processes (21–23). However, resistance may persist even after the selective pressure has been removed, due to gene linkage, co-selection, or compensatory mutations (24).
E. coli was selected as the model organism for this study due to its well-characterized genetics, rapid growth, and wide use in laboratory evolution experiments. As one of the most prevalent Gram-negative facultative anaerobes, E. coli frequently serves as a host for the acquisition and dissemination of antimicrobial resistance (AMR) genes, including plasmid-borne elements. Its ability to readily accumulate and maintain diverse resistance determinants makes it a relevant and representative system for studying adaptive trajectories under antibiotic pressure (23, 25).
Mitigating resistance requires the implementation of preventive strategies, such as adherence to biosecurity protocols (26), and the stabilization of the host microbiome through the use of prebiotics and probiotics (27, 28). Furthermore, antimicrobial stewardship should be supported by the targeted application of antibiotics and, where feasible, by replacing them with alternative agents such as antimicrobial peptides, medium-chain fatty acids, plant extracts, or essential oils (29–35).
The aim of this study was to investigate the microevolutionary adaptation of E. coli to potentiated sulfonamides (trimethoprim-sulfamethoxazole) under in vitro conditions, using a controlled model system. Special attention was given to the identification of resistance-related genetic mutations and co-selection mechanisms that may contribute to the emergence of cross-resistance to other antimicrobials.
Materials and methods
Bacterial strain
Experiments used the E. coli reference strain ATCC 25922, a widely accepted quality control organism for antimicrobial susceptibility testing.
MEGA-plate preparation
To enable gradient-based selection, a custom MEGA-plate system was designed and constructed from polycarbonate sheets (dimensions: 60 × 30 cm; thickness: 5 mm) by Innoterm Ltd. (Budapest, Hungary). The design was based on the original evolutionary plate format (36), and refined using established protocols (37). For sterilization, a 7.5% hydrogen peroxide solution was prepared by diluting 30% stock hydrogen peroxide (VWR International Ltd., Hungary) with deionized water at a 1:3 ratio, as described in prior studies (38, 39). The plate chamber was placed under a laminar flow hood, filled with the disinfectant, and the inner surface of the lid was wiped with 0.9% sodium hypochlorite (NaOCl) (Merck KGaA, Darmstadt, Germany). After 15 min of exposure, the disinfectant was removed using vacuum suction, and the system was subjected to UV irradiation for 30 min to complete the decontamination process.
The MEGA-plate was assembled in three sequential layers. The growth medium was layered in three distinct phases. The base layer comprised nine separated zones containing incrementally increasing concentrations of the tested antimicrobial compound (0×, 1×, 10×, 100×, and 1,000×), arranged symmetrically from the edges toward the center. This was followed by a continuous solid agar layer that provided structural stability. The uppermost layer consisted of semi-solid agar to allow bacterial movement and gradual migration across the gradient. The antimicrobial concentration increments (0×, 1×, 10×, 100×, and 1,000×) were selected to model large-scale evolutionary transitions and to simulate strong, stepwise selection pressure across several orders of magnitude. This log-scale gradient approach is based on the original MEGA-plate concept introduced by Baym et al. (36), which demonstrated that logarithmic steps (e.g., 10-fold increases) allow observation of successive mutational sweeps and adaptation bottlenecks as bacterial populations advance through increasingly selective environments. Such a design allows the detection of both early and late resistance adaptations, including low-level tolerance mechanisms at 1 × to 10 × MIC, and high-level mutational resistance at extreme concentrations such as 1,000 ×.
Agar was prepared using 2% BD Bacto Agar (VWR International Ltd., Debrecen, Hungary) for all layers, except the top layer which contained 0.28% agar to allow for semi-solid consistency. Each liter of medium was supplemented with one LB-Lennox capsule (VWR International Ltd., Debrecen, Hungary), while the top layer received two capsules to optimize surface growth conditions. Cycloheximide (64 μg/mL final concentration; Merck KGaA, Darmstadt, Germany) was incorporated to inhibit fungal contamination. For enhanced visual contrast during imaging, black acrylic paint (Artmie, Budapest, Hungary) was added at 4 mL/L to the base and intermediate layers.
The E. coli inoculum was prepared by adjusting the culture to 0.5 McFarland turbidity (1.5 × 108 CFU/mL) using a nephelometer (Thermo Fisher Scientific, Waltham, MA, United States). The E. coli inoculum was applied to both lateral ends of the plate, by dispensing 100 μL of the standardized suspension evenly along the entire edge to form continuous inoculation lines. Inoculation was performed immediately after the top layer had solidified at room temperature. The plate was subsequently incubated at 37 °C.
Antibiotic susceptibility testing
Antimicrobial susceptibility was initially assessed by determining the MIC of each agent against the E. coli ATCC 25922 strain, following the Clinical and Laboratory Standards Institute (CLSI) guidelines (40). To facilitate the evolution of resistant clones, the 1 × baseline concentration of potentiated sulphonamide was defined as one-quarter of the MIC (2 μg/mL), with subsequent concentrations (10×, 100×, and 1,000×) scaled accordingly for MEGA-plate application.
After 10 days of incubation on the MEGA-plate — during which the bacterial population successfully expanded through all concentration gradients including 1,000 × — samples were taken from each antibiotic exposure zone. These were streaked onto ChromoBio Coliform agar (Biolab Zrt., Budapest, Hungary), a chromogenic selective medium, to confirm the absence of cross-contamination. Single colonies were subcultured onto tryptic soy agar and cryopreserved in Microbank vials at −80 °C for later analysis.
Three independent biological replicates were performed for each antibiotic concentration, using separate MEGA-plate assays. For each replicate, samples were collected from three parallel locations within the gradient region of interest. These parallel samples were not treated as technical replicates, as they originated from distinct spatial points and served to confirm intra-experimental consistency. All recovered isolates from a given MEGA-plate exhibited uniform susceptibility profiles. Frozen stocks were revived 24 h prior to testing by inoculating into 3 mL of cation-adjusted Mueller-Hinton broth (CAMHB; Biolab Zrt., Budapest, Hungary), followed by incubation at 37 °C for 18–24 h.
Broth microdilution assays were conducted in 96-well microtiter plates. Wells in columns 2 through 12 were filled with 90 μL of CAMHB (Step 1). Stock solutions of the test compounds — ceftriaxone, cefquinome, cefotaxime, ceftiofur, colistin, enrofloxacin, amoxicillin, neomycin, oxytetracycline, florfenicol, and potentiated sulfonamide (trimethoprim-sulfamethoxazole, 1:19) — were prepared at 1024 μg/mL according to CLSI standards, diluted 1:1 in CAMHB, and 180 μL of each solution was dispensed into column 1 (Step 2). Twofold serial dilutions were performed across the plate (Step 3). To standardize volume, pipette tips were discarded after column 10, ensuring a consistent 90 μL final volume in each well.
Bacterial suspensions were adjusted to a 0.5 McFarland standard using a nephelometer (ThermoFisher Scientific, Budapest, Hungary), and 10 μL of each suspension was added to the wells in reverse order from column 11 to column 2 (41). MIC endpoints were determined and analyzed using standardized methods.
Extended-spectrum beta-lactamase (ESBL) detection
Extended-spectrum beta-lactamase (ESBL) activity in E. coli isolates was assessed per CLSI protocols (40). MICs were determined for ceftazidime and cefotaxime, both alone and in combination with clavulanic acid, a β-lactamase inhibitor. A fixed clavulanic acid concentration of 4 μg/mL was maintained across all wells containing the antibiotic-inhibitor combinations.
To prepare the inhibitor solution, 10 mg of clavulanic acid was dissolved in 10 mL of distilled water and filtered through a sterile cellulose membrane. This solution was then mixed with 230 mL of sterile cation-adjusted Mueller-Hinton broth (CAMHB; Merck KGaA, Darmstadt, Germany), producing a final concentration equivalent to 10 mg per 240 mL.
For the working plates, wells designated for ESBL testing were pre-filled with 240 μL of this clavulanate-supplemented broth. A 10 μL aliquot of bacterial suspension was added to each well, followed by a 10 μL transfer into 90 μL of CAMHB in the test plates, resulting in a precise final clavulanic acid concentration of 4 μg/mL per well.
Plates were incubated at 37 °C for 18–24 h. Interpretation of results followed CLSI standards: an isolate was considered ESBL-positive if the MIC of the beta-lactam agent decreased by threefold or more in the presence of clavulanic acid. For example, a reduction in ceftazidime MIC from 8 μg/mL to 1 μg/mL in the clavulanic acid combination would indicate ESBL production.
Molecular genetic analysis
To assess the genomic effects of antibiotic exposure and screen for genes potentially involved in ESBL production, we performed comprehensive molecular genetic analyses. Genomic DNA was extracted using the Zymo Quick-DNA Fungal/Bacterial Miniprep Kit, per manufacturer’s instructions. Cell lysis and mechanical disruption were achieved by bead beating using a Qiagen TissueLyzer LT system (Qiagen GmbH, Hilden, Germany) at 50 Hz for 5 min. Extracted nucleic acids were stored at −20 °C until further use.
Sequencing was conducted on an Illumina NextSeq 500 platform, employing paired-end reads. Illumina’s sequencing-by-synthesis technology involves immobilization of DNA fragments via bridge amplification, removal of the reverse strand, and iterative nucleotide incorporation with fluorescent signal detection (42, 43).
For co-evolutionary assessments and comparative genomic analysis, whole-genome libraries were prepared using the Illumina® Nextera XT DNA Library Preparation Kit (Illumina, San Diego, USA). Fragment-specific dual-index barcoding was achieved using Nextera XT Index Kit v2 Set A. DNA input was standardized to 0.2 ng/μL in a final volume of 2.5 μL. This was mixed with 5 μL of Tagment DNA buffer and 2.5 μL of Amplicon Tagment Mix and incubated at 55 °C for 6 min using an Eppendorf Mastercycler nexus GX2 thermal cycler (Eppendorf SE, Hamburg, Germany). The reaction was cooled to 10 °C and neutralized with 2.5 μL of Neutralize Tagment buffer, followed by a 5-min room-temperature incubation.
For library amplification, 7.5 μL of Nextera PCR Master Mix was combined with 2.5 μL each of i5 and i7 index primers and added to the tagmented DNA. PCR conditions included an initial denaturation at 95 °C for 30 s, followed by 12 cycles (95 °C for 10 s, 55 °C for 30 s, 72 °C for 30 s), and a final extension at 72 °C for 5 min. Reactions were cooled to 10 °C. Libraries were purified using the Gel/PCR DNA Fragments Extraction Kit (Geneaid Biotech, Hsinchu, Taiwan) according to the column-based protocol, and DNA concentrations were quantified using the Qubit® dsDNA HS Assay Kit (Thermo Fisher Scientific, Waltham, MA, United States). Indexed libraries were pooled and normalized for sequencing.
To ensure high-confidence variant interpretation, all detected mutations were subjected to quality filtering prior to downstream analysis. Variants were classified as “confidently characterized” if they met the following criteria: (i) a minimum variant allele frequency (VAF) of ≥90%, (ii) a coverage depth of at least 30 × at the mutation site, (iii) consistent detection across at least two of the three biological replicates, and (iv) unambiguous functional annotation by both SnpEff and Prokka pipelines. Variants not fulfilling these thresholds were excluded from quantitative summary statistics and functional interpretation.
Results
Antimicrobial resistance gene set
Based on GenomeScope analyses used for general genome characterization, the k-mer frequency histograms consistently aligned with the expected size of E. coli genomes (Supplementary Figures 1–5). Furthermore, taxonomic classification performed using CheckM (v1.1.6) and Kraken (v1.1.1.1) confirmed a 100% match with E. coli across all samples.
Resistance gene annotation using the Comprehensive Antibiotic Resistance Database (CARD) identified 44 distinct genes with ≥90% sequence identity and coverage [additional data (potentiated sulphonamide)]. The identified ARG profile was identical across all samples and conferred resistance against 22 classes of antibiotics, disinfectants, and dyes; as in our previous similar studies (44, 45). Of particular note, the ampC and ampH genes — encoding β-lactamase enzymes — are critical for resistance to β-lactam antibiotics (e.g., penicillins and cephalosporins) via enzymatic inactivation (Table 1). The presence of acrA, acrB, and tolC genes indicates a functional multidrug efflux pump system, which is commonly found in E. coli. Similar efflux systems are formed by the combination of tolC with emrA and emrB, as well as mdtA, mdtB, and mdtC genes.

Table 1. The 44 antimicrobial resistance genes identified by next-generation sequencing, consistently detected across all samples.
Figure 1 presents the classification of the 44 identified ARGs according to drug class and resistance mechanism. Of these, 21 genes confer fluoroquinolone resistance, with 19 encoding efflux pumps. The second most prevalent group includes genes conferring resistance to penicillins (n = 17), of which 14 are efflux pump-related and two (ampC, ampH) are associated with enzymatic inactivation. Target modification resistance genes (n = 4) were exclusively associated with peptide (polymyxin) antibiotics.

Figure 1. Classification of the identified antimicrobial resistance genes by antimicrobial class and resistance mechanism.
The folP gene, associated with sulfonamide resistance, was detected but did not meet the CARD inclusion threshold. However, the identification of point mutations in folP suggests a potential key role in the observed resistance phenotype to potentiated sulfonamides (see Figure 2).
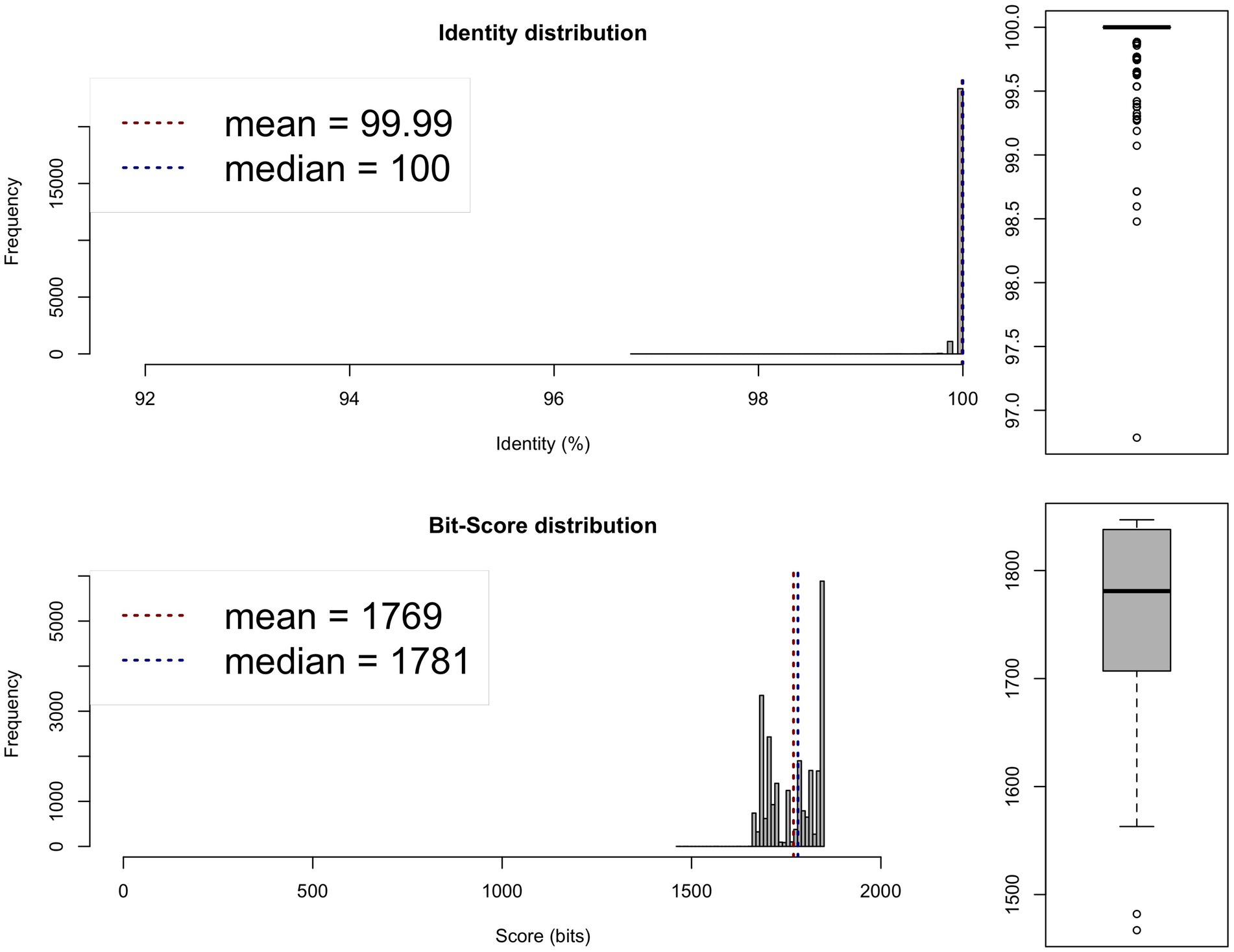
Figure 2. Average nucleotide identity (ANI) analysis between the genomes of the 0 × and 1,000 × potentiated sulphonamide-exposed samples, showing species-level similarity exceeding 95%. One-way ANI 1: 99.98% (SD: 0.44%), based on 24,651 fragments. One-way ANI 2: 99.99% (SD: 0.11%), based on 24,584 fragments. Two-way ANI: 99.99% (SD: 0.04%), based on 24,544 fragments.
Based on the sequencing data, serotyping of the studied strain was also possible, leading to the identification of O6 type-specific polysaccharide antigens (wzx, wzy) and H1 and H12 protein antigens (fliC).
Phenotypic results
When exposed to potentiated sulphonamide, bacteria grew across the highest antibiotic concentration within 7 days. Exposure to potentiated sulphonamide resulted in increased MIC values for all tested antibiotics at both 100 × and 1,000 × concentrations (Table 2).

Table 2. Effect of increasing potentiated sulphonamide concentrations on the minimum inhibitory concentration (MIC) values of the tested antimicrobial agents.
Across all three biological replicates, MIC values remained consistent, with no detectable variation between corresponding concentration endpoints. Intra-plate sampling from three parallel locations yielded identical susceptibility profiles, indicating high spatial uniformity. Given the full agreement among replicates, basic descriptive statistics (mean ± SD) are not presented for MIC changes, as standard deviation was effectively zero across all measurements. These results confirm the robustness and reproducibility of resistance acquisition patterns under sulphonamide exposure.
ESBL production assays yielded negative results (see Table 3).

Table 3. Results of the extended-spectrum β-lactamase (ESBL) test using ceftazidime (CTZ), cefotaxime (CTX), and their respective clavulanic acid (KLA) combinations, following the guidelines of the Clinical Laboratory Standards Institute (CLSI).
Genotypic results
All contigs passed quality control, ensuring consistent sequencing quality (see Table 4).
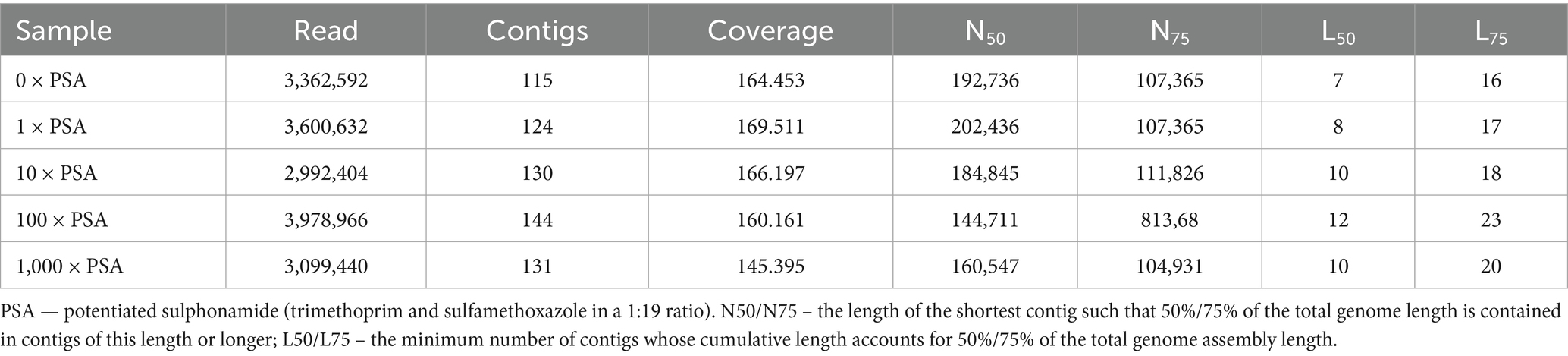
Table 4. Contig quality metrics based on QUAST software analysis for potentiated sulphonamide-exposed samples.
The bacA gene was detected on phage elements in all samples. The emrB gene was found on phage-associated sequences in both 10 and 1,000 potentiated sulphonamide-treated samples. Additionally, bacA was identified as a mobile genetic element (MGE) in all cases, while acrE and acrF were present as MGEs in the 1 × and 10 × samples.
The number of mutations detected per sample compared to the reference genome is summarized in Table 5. A total of 9,278 mutations were identified; 4,935 were classified with confidence (criteria detailed in Methods). The distribution of total mutations ranged from 1,752 to 1,958 per sample, while the number of identified mutations ranged from 923 to 1,074. Mutation overlap with the 0 baseline strain was: 1 sample, 101.6; 10 sample, 107.3; 100 sample, 109.4; and 1,000 sample, 116.4.

Table 5. Number of total and identified mutations per sample, classified by mutation type.
Table 6 summarizes the mutations that may have contributed to the phenotypic expression of antimicrobial resistance. An emrR gene deletion occurred only in the 1,000 concentration sample. This mutation may promote the expression of the emrAB-tolC multidrug efflux pump system, which is known to play a key role in resistance development (46). The mdtM gene encodes a multidrug resistance protein shown to interact with the acrAB-tolC efflux system. A deletion in mdtM was also identified in the 1,000 × treated sample. This gene encodes a transmembrane drug/H+ antiporter, which has been implicated in resistance to multiple antibiotics in E. coli (47). Additionally, yfiB, encoding the ompA outer membrane lipoprotein (48), also harbored a deletion in the 1,000 × sample. Mutations in yfiB have previously been associated with resistance phenotypes in E. coli (49, 50).

Table 6. Deletions, inversions, and single nucleotide polymorphisms (SNPs) detected in genes relevant to antimicrobial resistance.
An inversion in the acrF gene was observed in the 1,000 × sample. acrF encodes an inner membrane transporter functionally analogous to acrB, and is a component of the acrEF-tolC efflux pump complex, which plays a key role in multidrug efflux (51). The mdtF gene showed both a deletion and a single nucleotide polymorphism (SNP) in the 100 × sample. This gene encodes the inner membrane transporter of the mdtEF-tolC multidrug efflux system (52).
Among the identified SNPs, acrB mutations were detected at 1, 10, 100, and 1,000 concentrations; these merit particular attention. acrB is a key subunit of the acrAB-tolC efflux complex, functioning as a heterotrimeric drug-proton antiporter across the inner membrane (53–56). Notably, acrR—the acrAB-tolC repressor—mutated in several samples. These changes are known to result in high-level antibiotic resistance (57).
For potentiated sulphonamides, folA mutation was exclusive to the 1,000 sample, while folP mutations occurred at 10, 100, and 1,000. The folA encodes dihydrofolate reductase, a key enzyme in the synthesis of tetrahydrofolate, which is essential for the de novo synthesis of the DNA nucleotide thymidine. One known bacterial resistance mechanism is the overproduction of this enzyme, and the observed folA mutation may reflect such adaptation to trimethoprim exposure (58).
In the 100 sample, a marR mutation may have upregulated marA, activating acrAB-tolC efflux (59, 60).
An SNP in mdtB (mdtABC-tolC system) was found in the 100 sample. Loss-of-function mutations in mdtB can lead to compensatory expression of mdtC, preserving pump activity (61).
An additional SNP in mdtF was identified in the 1,000 × sample, potentially enhancing activity of the mdtEF-tolC system (52), which may explain the observed MIC increases for multiple antibiotics.
Potentiated sulphonamide exposure induced diverse mutations, especially in efflux regulation and folate metabolism genes. These genetic alterations likely underpin the observed increases in MIC not only to sulfamethoxazole and trimethoprim, but to other unrelated antibiotics as well. Furthermore, mutations were detected in key SOS-box regulatory genes—including recN, recQ, and uvrB—in the 100 × and 1,000 × samples, suggesting an induced stress response pathway.
Discussion
Bacterial resistance to sulfonamides usually involves gene mutation or gene substitution. In the mutation-based mechanism, alterations in the chromosomal folP gene, which encodes DHPS, confer resistance to sulfonamides (24). In the substitution-based pathway, sulfonamide resistance arises from the acquisition of alternative DHPS-encoding genes (sul1, sul2, sul3), which produce enzymes with lower affinity for sulfonamides. This latter mechanism is more prevalent in clinical and environmental isolates (62, 63).
Sulfonamides, competitive inhibitors of DHPS, block PABA conversion to dihydropteroate, disrupting folate biosynthesis. Folates function as essential cofactors in nucleotide and certain amino acid biosynthesis. Consequently, DHPS inhibition blocks bacterial proliferation, particularly in organisms unable to scavenge folates from their environment. However, sul genes encode DHPS variants that are not inhibited by sulfonamides, thereby conferring resistance (64–67).
Among the genetic changes induced by potentiated sulphonamide exposure, the most critical were mutations observed in the folP gene (in 10×, 100×, and 1,000 × samples) and in the folA gene (in the 1,000 × sample). The folA gene regulates the synthesis of DHFR, which converts dihydrofolate into tetrahydrofolate — a coenzyme essential for de novo thymidine nucleotide biosynthesis. One of the known bacterial resistance mechanisms involves overexpression of this enzyme, and the observed folA mutation may reflect an adaptation to reduce susceptibility to the diaminopyrimidine component of the drug combination (58).
Meanwhile, folP encodes DHPS itself, and point mutations in this gene can impair sulfonamide binding, thereby maintaining folate synthesis and promoting resistance (7). The combined mutations in folA and folP likely explain the elevated MIC values observed in response to potentiated sulphonamide exposure.
Multidrug efflux transporters are widespread and play a central role in antibiotic resistance mechanisms, as they enable bacteria to evade the action of many current therapeutic agents (68, 69). Among them, ATP-binding cassette (ABC) transporters are essential molecular transport systems that rely on ATP hydrolysis to provide the energy required for translocating various biomolecules across cellular membranes (69). Alterations in ABC transporter function are closely associated with the physiological state of the bacterial strain and environmental growth conditions (24).
Our results suggest that the increased MIC values observed for several antibiotics may be attributed to the activation of efflux pump systems. A deletion in the emrR gene, observed exclusively in the 1,000 × PSA-treated strain, could lead to upregulation of the emrAB-tolC multidrug efflux pump (46). The mdtM gene, which regulates the acrAB-tolC efflux system, also showed a deletion in the 1,000 × sample. This gene encodes a transmembrane drug/H+ antiporter involved in resistance against multiple agents (47). An inversion in the acrF gene, a homolog of acrB that contributes to the acrEF-tolC efflux complex, was also detected in the 1,000 × strain (51).
Additionally, a mutation in the acrR gene — encoding a repressor of the acrAB-tolC system—was associated with high-level resistance phenotypes (57). The marR gene, whose mutation was observed in the 100 × sample, triggers the expression of the marA activator protein, leading to enhanced acrAB-tolC pump activity (59, 60).
We also observed a deletion and SNP in the mdtF gene (100 × and 1,000 × samples), which encodes the inner membrane transporter of the mdtEF-tolC efflux system (52). Moreover, an SNP in mdtB, a component of the mdtABC-tolC pump, was found in the 100 × sample. If this mutation disrupts mdtB expression, it may indirectly activate mdtC expression, thus maintaining efflux pump functionality (61).
Several identified mutations occurred in well-characterized efflux regulators, including emrR, marR, and acrR. These genes encode local or global transcriptional repressors that modulate multidrug resistance efflux systems such as EmrAB, AcrAB-TolC, and others. Loss-of-function mutations in marR or acrR are known to derepress expression of their corresponding operons (marA and acrAB, respectively), leading to upregulation of the AcrAB-TolC efflux pump and increased tolerance to structurally unrelated antibiotics, including fluoroquinolones, chloramphenicol, and tetracyclines (60, 70). Similarly, inactivation of emrR results in overexpression of the EmrAB pump, which has been implicated in resistance to nalidixic acid and other hydrophobic agents (46). These regulatory disruptions likely contribute to the observed cross-resistance phenotypes in our evolved populations, highlighting the role of efflux derepression as a collateral pathway of adaptation under sulphonamide pressure.
These regulatory disruptions likely contribute to the observed cross-resistance phenotypes in our evolved populations, highlighting the role of efflux depression as a collateral pathway of adaptation under sulphonamide pressure. Comparable cross-resistance patterns have been reported in systems exposed to aminoglycosides and fluoroquinolones, where marR and acrR mutations similarly led to broad-spectrum efflux activation and multidrug tolerance (71, 72).
As for SOS-box related genes, point mutations were identified in recN, recQ, and uvrB in both 100 × and 1,000 × PSA-treated samples. These genes are involved in DNA damage response and may contribute to mutagenesis and adaptive stress responses under antimicrobial pressure.
Horizontal gene transfer (HGT) can occur under a variety of laboratory conditions and can be facilitated by physical means that enhance DNA exchange — such as immobilization on membranes or agar surfaces — and likely by various environmental factors that promote gene acquisition. In our study, E. coli strains exposed to increasing concentrations of PSA under controlled conditions accumulated point mutations, confirming the utility of the MEGA-plate system in capturing microevolutionary dynamics (36, 37).
Similar approaches have been successfully used to monitor evolutionary trajectories in response to diverse antimicrobials, including trimethoprim (36), and ciprofloxacin (73), consistently demonstrating that spatial drug gradients can unveil stepwise resistance acquisition and collateral adaptation patterns.
Compared to our previous investigations using β-lactams and florfenicol (44, 45), the present study demonstrates that sulphonamides induce a markedly different adaptive landscape. Unlike β-lactam exposure, which primarily selects resistance via target modification or efflux, sulphonamide exposure potentiates hypermutation. This is likely mediated through indirect interference with nucleotide synthesis, leading to replication stress and increased SOS response activation. This mechanistic divergence is critical, while β-lactams often select for resistance mutations within the target genes or membrane-associated functions, sulphonamides appear to globally elevate the mutation rate, possibly accelerating the emergence of de novo resistance mutations across unrelated loci. Such behavior highlights a previously underappreciated evolutionary pathway, in which sub-inhibitory sulphonamide concentrations function not only as selective agents, but also as mutagenic potentiators, thereby expanding the evolutionary trajectories available to bacterial populations under antimicrobial pressure.
Similar mutagenic effects have been documented for fluoroquinolones and β-lactams under subinhibitory conditions, where they activate the SOS response and drive error-prone polymerase expression (74, 75), albeit through distinct primary mechanisms. Our findings suggest that sulphonamides may exploit a comparable yet broader pathway via nucleotide pool disruption.
It is worth emphasizing that antibiotics — even at subinhibitory concentrations — can promote the emergence of resistance (76). They have been shown to enhance gene transfer and recombination events (77), partially through the activation of the bacterial SOS response (78, 79). These factors may significantly contribute to elevated gene exchange rates in environments such as farms, hospitals, and sewage systems, which offer ideal conditions for the acquisition and dissemination of resistance genes.
Conclusion
The results of this study confirm that potentiated sulfonamides—specifically the combination of sulfamethoxazole and trimethoprim—can induce substantial genetic adaptation in E. coli strains after short-term in vitro exposure. Mutations in folP and folA likely cause resistance to target compounds; efflux pump gene alterations contributed to broad-spectrum cross-resistance. Several of these genetic changes were associated with mobile genetic elements, indicating a potential for horizontal gene transfer and the wider dissemination of resistance traits. The MEGA-plate platform proved to be an effective experimental model for tracking co-evolutionary dynamics. Our findings underscore the need for caution in the clinical and veterinary use of antibiotic combinations and highlight the importance of further molecular investigations into the mechanisms underlying co-selection and multidrug resistance.
Data availability statement
The raw data supporting the conclusions of this article will be made available by the authors. The datasets analyzed for this study can be found in the National Library of Medicine: https://www.ncbi.nlm.nih.gov/bioproject/PRJNA1261795, accessed on 12 May 2025.
Author contributions
ÁK: Methodology, Conceptualization, Visualization, Supervision, Writing – review & editing, Project administration. BT: Resources, Writing – review & editing, Project administration. ÁS: Project administration, Writing – review & editing, Resources. LL: Formal analysis, Software, Validation, Writing – review & editing. GK: Resources, Writing – review & editing, Supervision. KBá: Writing – review & editing, Resources, Project administration, Supervision. EK: Formal analysis, Software, Validation, Writing – review & editing. KBa: Writing – original draft, Data curation, Investigation. ÁJ: Validation, Conceptualization, Funding acquisition, Writing – review & editing.
Funding
The author(s) declare that financial support was received for the research and/or publication of this article. The authors declare that financial support was received for the research, authorship, and/or publication of this article. This research was funded by the ÚNKP-20-3-I-ÁTE-1 and ÚNKP-21-2-I-ÁTE-3 New National Excellence Program of the Ministry for Innovation and Technology from the National Research, Development and Innovation Fund. Project no. RRF-2.3.1-21-2022-00001 was implemented with the support provided by the Recovery and Resilience Facility (RRF), financed under the National Recovery Fund budget estimate, RRF-2.3.1-21 funding scheme.
Acknowledgments
We would like to thank all the laboratory technical assistants, without whom this research would not have been possible.
Conflict of interest
The authors declare that the research was conducted in the absence of any commercial or financial relationships that could be construed as a potential conflict of interest.
Generative AI statement
The author(s) declare that no Gen AI was used in the creation of this manuscript.
Any alternative text (alt text) provided alongside figures in this article has been generated by Frontiers with the support of artificial intelligence and reasonable efforts have been made to ensure accuracy, including review by the authors wherever possible. If you identify any issues, please contact us.
Publisher’s note
All claims expressed in this article are solely those of the authors and do not necessarily represent those of their affiliated organizations, or those of the publisher, the editors and the reviewers. Any product that may be evaluated in this article, or claim that may be made by its manufacturer, is not guaranteed or endorsed by the publisher.
Supplementary material
The Supplementary material for this article can be found online at: https://www.frontiersin.org/articles/10.3389/fvets.2025.1697872/full#supplementary-material
References
1. Exner, M, Bhattacharya, S, Christiansen, B, Gebel, J, Goroncy-Bermes, P, Hartemann, P, et al. Antibiotic resistance: what is so special about multidrug-resistant gram-negative bacteria? GMS Hyg Infect Control. (2017) 12:Doc05. doi: 10.3205/dgkh000290
2. Shrivastava, SR, Shrivastava, PS, and Ramasamy, J. World health organization releases global priority list of antibiotic-resistant bacteria to guide research, discovery, and development of new antibiotics. J Med Soc. (2018) 32:76. doi: 10.4103/jms.jms_25_17
3. Huovinen, P, Sundström, L, Swedberg, G, and Sköld, O. Trimethoprim and sulfonamide resistance. Antimicrob Agents Chemother. (1995) 39:279–89. doi: 10.1128/AAC.39.2.279
4. Huovinen, P. Resistance to trimethoprim-sulfamethoxazole. Clin Infect Dis. (2001) 32:1608–14. doi: 10.1086/320532
5. Perreten, V, and Boerlin, P. A new sulfonamide resistance gene (sul3) in Escherichia coli is widespread in the pig population of Switzerland. Antimicrob Agents Chemother. (2003) 47:1169–72. doi: 10.1128/AAC.47.3.1169-1172.2003
6. Sköld, O. Sulfonamide resistance: mechanisms and trends. Drug Resist Updat. (2000) 3:155–60. doi: 10.1054/drup.2000.0146
7. Vedantam, G, Guay, GG, Austria, NE, Doktor, SZ, and Nichols, BP. Characterization of mutations contributing to sulfathiazole resistance in Escherichia coli. Antimicrob Agents Chemother. (1998) 42:88–93. doi: 10.1128/AAC.42.1.88
8. Swedberg, G, Fermér, C, and Sköld, O. Point mutations in the dihydropteroate synthase gene causing sulfonamide resistance. Adv Exp Med Biol. (1993) 338:555–8. doi: 10.1007/978-1-4615-2960-6_113
9. Rådström, P, and Swedberg, G. RSF1010 and a conjugative plasmid contain sulII, one of two known genes for plasmid-borne sulfonamide resistance dihydropteroate synthase. Antimicrob Agents Chemother. (1988) 32:1684–92. doi: 10.1128/AAC.32.11.1684
10. Swedberg, G, and Sköld, O. Characterization of different plasmid-borne dihydropteroate synthases mediating bacterial resistance to sulfonamides. J Bacteriol. (1980) 142:1–7. doi: 10.1128/jb.142.1.1-7.1980
11. Schweitzer, BI, Dicker, AP, and Bertino, JR. Dihydrofolate reductase as a therapeutic target. FASEB J. (1990) 4:2441–52. doi: 10.1096/fasebj.4.8.2185970
12. Blakley, RL. Eukaryotic dihydrofolate reductase. Adv Enzymol Relat Areas Mol Biol. (1995) 70:23–102. doi: 10.1002/9780470123164.ch2
13. Hong, W, Wang, Y, Chang, Z, Yang, Y, Pu, J, Sun, T, et al. The identification of novel Mycobacterium tuberculosis DHFR inhibitors and the investigation of their binding preferences by using molecular modelling. Sci Rep. (2015) 5:15328. doi: 10.1038/srep15328
14. Rashid, N, Thapliyal, C, and Chattopadhyay, P. Dihydrofolate reductase as a versatile drug target in healthcare. J Prot Prot (2016). Available online at: https://www.semanticscholar.org/paper/DIHYDROFOLATE-REDUCTASE-AS-A-VERSATILE-DRUG-TARGET-Rashid-Thapliyal/5ce97b91d207b38dd68a68fccb5dbd250a99909d [Accessed May 4, 2025]
15. Nicolle, LE. Urinary tract infection: traditional pharmacologic therapies. Dis Mon. (2003) 49:111–28. doi: 10.1067/mda.2003.11
16. Bean, DC, Livermore, DM, and Hall, LMC. Plasmids imparting sulfonamide resistance in Escherichia coli: implications for persistence. Antimicrob Agents Chemother. (2009) 53:1088–93. doi: 10.1128/AAC.00800-08
17. Hammerum, AM, Sandvang, D, Andersen, SR, Seyfarth, AM, Porsbo, LJ, Frimodt-Møller, N, et al. Detection of sul1, sul2 and sul3 in sulphonamide resistant Escherichia coli isolates obtained from healthy humans, pork and pigs in Denmark. Int J Food Microbiol. (2006) 106:235–7. doi: 10.1016/j.ijfoodmicro.2005.06.023
18. Lanz, R, Kuhnert, P, and Boerlin, P. Antimicrobial resistance and resistance gene determinants in clinical Escherichia coli from different animal species in Switzerland. Vet Microbiol. (2003) 91:73–84. doi: 10.1016/s0378-1135(02)00263-8
19. Sköld, O. Resistance to trimethoprim and sulfonamides. Vet Res. (2001) 32:261–73. doi: 10.1051/vetres:2001123
20. Antunes, P, Machado, J, and Peixe, L. Dissemination of sul3-containing elements linked to class 1 integrons with an unusual 3′ conserved sequence region among Salmonella isolates. Antimicrob Agents Chemother. (2007) 51:1545–8. doi: 10.1128/AAC.01275-06
21. Melnyk, AH, Wong, A, and Kassen, R. The fitness costs of antibiotic resistance mutations. Evol Appl. (2015) 8:273–83. doi: 10.1111/eva.12196
22. Stickland, HG, Davenport, PW, Lilley, KS, Griffin, JL, and Welch, M. Mutation of nfxB causes global changes in the physiology and metabolism of Pseudomonas aeruginosa. J Proteome Res. (2010) 9:2957–67. doi: 10.1021/pr9011415
23. Davies, J, and Davies, D. Origins and evolution of antibiotic resistance. Microbiol Mol Biol Rev. (2010) 74:417–33. doi: 10.1128/MMBR.00016-10
24. Zhou, Y, Fang, J, Davood, Z, Han, J, and Qu, D. Fitness cost and compensation mechanism of sulfonamide resistance genes (sul1, sul2, and sul3) in. Environ Microbiol. (2021) 23:7538–49. doi: 10.1111/1462-2920.15783
25. Lopatkin, AJ, Bening, SC, Manson, AL, Stokes, JM, Kohanski, MA, Badran, AH, et al. Clinically relevant mutations in core metabolic genes confer antibiotic resistance. Science. (2021) 371:eaba0862. doi: 10.1126/science.aba0862
26. Farkas, M, Könyves, L, Csorba, S, Farkas, Z, Józwiák, Á, Süth, M, et al. Biosecurity situation of large-scale poultry farms in Hungary according to the databases of National Food Chain Safety Office Centre for disease control and biosecurity audit system of poultry product Board of Hungary in the period of 2021–2022. Magy Allatorv Lapja. (2024) 146:723–42. doi: 10.56385/magyallorv.2024.12.723-742
27. Kovács, L, Hejel, P, Farkas, M, László, L, and Könyves, L. Study report on the effect of a litter treatment product containing bacillus licheniformis and zeolite in male fattening Turkey flock. Magy Allatorv Lapja. (2024) 146:291–305. doi: 10.56385/magyallorv.2024.05.291-305
28. Essősy, M, Fodor, I, Ihnáth, Z, Karancsi, Z, Kovács, D, Szalai, KV, et al. The possibilities of antibiotic-free broiler-hen fattening, with special reference to the use of pre- and probiotics. Magy Allatorv Lapja. (2020) 142:397–407.
29. Jerzsele, Á, Somogyi, Z, Szalai, M, and Kovács, D. Effects of fermented wheat germ extract on artificial Salmonella Typhimurium infection in broiler chickens. Magy Allatorv Lapja. (2020) 142:77–85.
30. Kerek, Á, Csanády, P, and Jerzsele, Á. Antibacterial efficiency of propolis – part 1. Magy Allatorv Lapja. (2022) 144:285–98.
31. Kerek, Á, Csanády, P, and Jerzsele, Á. Antiprotozoal and antifungal efficiency of propolis – part 2. Magy Allatorv Lapja. (2022) 144:691–704.
32. Kovács, L, Nagy, D, Könyves, L, Jerzsele, Á, and Kerek, Á. Antimicrobial properties of essential oils – animal health aspects. Magy Allatorv Lapja. (2023) 145:497–510. doi: 10.56385/magyallorv.2023.08.497-510
33. Olasz, Á, Jerzsele, Á, Balta, L, Dobra, PF, and Kerek, Á. In vivo efficacy of different extracts of propolis in broiler salmonellosis. Magy Allatorv Lapja. (2023) 145:461–75. doi: 10.56385/magyallorv.2023.08.461-475
34. Hetényi, N, Bersényi, A, and Hullár, I. Physiological effects of medium-chain fatty acids and triglycerides, and their potential use in poultry and swine nutrition: a literature review. Magy Allatorv Lapja. (2024) 146:651–9. doi: 10.56385/magyallorv.2024.11.651-659
35. Jócsák, G, Schilling-Tóth, B, Bartha, T, Tóth, I, Ondrašovičová, S, and Kiss, DS. Metal nanoparticles—immersion in the “tiny” world of medicine. Magy Allatorv Lapja. (2025) 147:115–27. doi: 10.56385/magyallorv.2025.02.115-127
36. Baym, M, Lieberman, TD, Kelsic, ED, Chait, R, Gross, R, Yelin, I, et al. Spatiotemporal microbial evolution on antibiotic landscapes. Science. (2016) 353:1147–51. doi: 10.1126/science.aag0822
37. Kerek, Á, Török, B, and Jerzsele, Á. MEGA-plate—new evolutionary and coselection microbiological method. Magy Allatorv Lapja. (2022) 144:429–39.
38. Sattar, SA, Taylor, YE, Paquette, M, and Rubino, J. In-hospital evaluation of 7.5% hydrogen peroxide as a disinfectant for flexible endoscopes. Can J Infect Control. (1996) 11:51–4.
39. Rutala, WA, and Weber, DJ. Disinfection of endoscopes: review of new chemical sterilants used for high-level disinfection. Infect Control Hosp Epidemiol. (1999) 20:69–76. doi: 10.1086/501544
40. Clinical and Laboratory Standards Institute (CLSI). Methods for dilution antimicrobial susceptibility tests for Bacteria that grow aerobically. 12. th ed. Wayne, PA: Clinical and Laboratory Standards Institute (2024).
41. Clinical and Laboratory Standards Institute. CLSI. Methods for dilution antimicrobial susceptibility tests for Bacteria that grow aerobically. 11. th ed. Wayne, PA: Clinical and Laboratory Standards Institute (2018).
42. Muzzey, D, Evans, EA, and Lieber, C. Understanding the basics of NGS: from mechanism to variant calling. Curr Genet Med Rep. (2015) 3:158–65. doi: 10.1007/s40142-015-0076-8
43. Mihály, Z, and Győrffy, B. Next generation sequencing technologies (NGST) development and applications. Orv Hetil. (2011) 152:55–62. doi: 10.1556/OH.2011.29007
44. Kerek, Á, Török, B, Laczkó, L, Kardos, G, Bányai, K, Somogyi, Z, et al. In vitro microevolution and co-selection assessment of florfenicol impact on Escherichia coli resistance development. Antibiotics. (2023) 12:1728. doi: 10.3390/antibiotics12121728
45. Kerek, Á, Török, B, Laczkó, L, Somogyi, Z, Kardos, G, Bányai, K, et al. In vitro microevolution and co-selection assessment of amoxicillin and cefotaxime impact on Escherichia coli resistance development. Antibiotics. (2024) 13:247. doi: 10.3390/antibiotics13030247
46. Lomovskaya, O, Lewis, K, and Matin, A. EmrR is a negative regulator of the Escherichia coli multidrug resistance pump EmrAB. J Bacteriol. (1995) 177:2328–34. doi: 10.1128/jb.177.9.2328-2334.1995
47. Holdsworth, SR, and Law, CJ. The major facilitator superfamily transporter MdtM contributes to the intrinsic resistance of Escherichia coli to quaternary ammonium compounds. J Antimicrob Chemother. (2013) 68:831–9. doi: 10.1093/jac/dks491
48. Malone, JG, Jaeger, T, Spangler, C, Ritz, D, Spang, A, Arrieumerlou, C, et al. YfiBNR mediates cyclic di-GMP dependent small colony variant formation and persistence in Pseudomonas aeruginosa. PLoS Pathog. (2010) 6:e1000804. doi: 10.1371/journal.ppat.1000804
49. Ruiz, J. Mechanisms of resistance to quinolones: target alterations, decreased accumulation and DNA gyrase protection. J Antimicrob Chemother. (2003) 51:1109–17. doi: 10.1093/jac/dkg222
50. Vila, J. Fluoroquinolone Resistance In: Frontiers in Antimicrobial Resistance. David G. White, Michael N. Alekshun, Patrick F. McDermott, és Stuart B. Levy editors. Washington, DC, USA John Wiley & Sons, Ltd (2005). 41–52.
51. Lau, SY, and Zgurskaya, HI. Cell division defects in Escherichia coli deficient in the multidrug efflux transporter AcrEF-TolC. J Bacteriol. (2005) 187:7815–25. doi: 10.1128/JB.187.22.7815-7825.2005
52. Nishino, K, and Yamaguchi, A. EvgA of the two-component signal transduction system modulates production of the yhiUV multidrug transporter in Escherichia coli. J Bacteriol. (2002) 184:2319–23. doi: 10.1128/JB.184.8.2319-2323.2002
53. Törnroth-Horsefield, S, Gourdon, P, Horsefield, R, Brive, L, Yamamoto, N, Mori, H, et al. Crystal structure of AcrB in complex with a single transmembrane subunit reveals another twist. Structure. (2007) 15:1663–73. doi: 10.1016/j.str.2007.09.023
54. Das, D, Xu, QS, Lee, JY, Ankoudinova, I, Huang, C, Lou, Y, et al. Crystal structure of the multidrug efflux transporter AcrB at 3.1A resolution reveals the N-terminal region with conserved amino acids. J Struct Biol. (2007) 158:494–502. doi: 10.1016/j.jsb.2006.12.004
55. Pos, KM. Drug transport mechanism of the AcrB efflux pump. Biochim Biophys Acta. (2009) 1794:782–93. doi: 10.1016/j.bbapap.2008.12.015
56. Eicher, T, Brandstätter, L, and Pos, KM. Structural and functional aspects of the multidrug efflux pump AcrB. Biol Chem. (2009) 390:693–9. doi: 10.1515/BC.2009.090
57. Pradel, E, and Pagès, J-M. The AcrAB-TolC efflux pump contributes to multidrug resistance in the nosocomial pathogen Enterobacter aerogenes. Antimicrob Agents Chemother. (2002) 46:2640–3. doi: 10.1128/AAC.46.8.2640-2643.2002
58. Myllykallio, H, Leduc, D, Filee, J, and Liebl, U. Life without dihydrofolate reductase FolA. Trends Microbiol. (2003) 11:220–3. doi: 10.1016/s0966-842x(03)00101-x
59. Delcour, AH. Outer membrane permeability and antibiotic resistance. Biochim Biophys Acta. (2009) 1794:808–16. doi: 10.1016/j.bbapap.2008.11.005
60. Alekshun, MN, and Levy, SB. Regulation of chromosomally mediated multiple antibiotic resistance: the mar regulon. Antimicrob Agents Chemother. (1997) 41:2067–75. doi: 10.1128/AAC.41.10.2067
61. Nagakubo, S, Nishino, K, Hirata, T, and Yamaguchi, A. The putative response regulator BaeR stimulates multidrug resistance of Escherichia coli via a novel multidrug exporter system, MdtABC. J Bacteriol. (2002) 184:4161–7. doi: 10.1128/JB.184.15.4161-4167.2002
62. Liang, P, Liu, S, Xu, F, Jiang, S, Yan, J, He, Q, et al. Corrigendum: powdery mildews are characterized by contracted carbohydrate metabolism and diverse effectors to adapt to obligate biotrophic lifestyle. Front Microbiol. (2019) 10:1. doi: 10.3389/fmicb.2019.00001
63. Suzuki, S, and Hoa, PTP. Distribution of quinolones, sulfonamides, tetracyclines in aquatic environment and antibiotic resistance in indochina. Front Microbiol. (2012) 3:67. doi: 10.3389/fmicb.2012.00067
64. Nunes, OC, Manaia, CM, Kolvenbach, BA, and Corvini, PF-X. Living with sulfonamides: a diverse range of mechanisms observed in bacteria. Appl Microbiol Biotechnol. (2020) 104:10389–408. doi: 10.1007/s00253-020-10982-5
65. Revuelta, JL, Serrano-Amatriain, C, Ledesma-Amaro, R, and Jiménez, A. Formation of folates by microorganisms: towards the biotechnological production of this vitamin. Appl Microbiol Biotechnol. (2018) 102:8613–20. doi: 10.1007/s00253-018-9266-0
66. Domínguez, M, Miranda, CD, Fuentes, O, de la Fuente, M, Godoy, FA, Bello-Toledo, H, et al. Occurrence of transferable Integrons and sul and dfr genes among Sulfonamide-and/or trimethoprim-resistant Bacteria isolated from Chilean salmonid farms. Front Microbiol. (2019) 10:748. doi: 10.3389/fmicb.2019.00748
67. Baran, W, Adamek, E, Ziemiańska, J, and Sobczak, A. Effects of the presence of sulfonamides in the environment and their influence on human health. J Hazard Mater. (2011) 196:1–15. doi: 10.1016/j.jhazmat.2011.08.082
68. Wasi, M, Khandelwal, NK, Moorhouse, AJ, Nair, R, Vishwakarma, P, Bravo Ruiz, G, et al. ABC transporter genes show upregulated expression in drug-resistant clinical isolates of Candida auris: a genome-wide characterization of ATP-binding cassette (ABC) transporter genes. Front Microbiol. (2019) 10:1445. doi: 10.3389/fmicb.2019.01445
69. Orelle, C, Mathieu, K, and Jault, J-M. Multidrug ABC transporters in bacteria. Res Microbiol. (2019) 170:381–91. doi: 10.1016/j.resmic.2019.06.001
70. Ma, D, Cook, DN, Alberti, M, Pon, NG, Nikaido, H, and Hearst, JE. Genes acrA and acrB encode a stress-induced efflux system of Escherichia coli. Mol Microbiol. (1995) 16:45–55. doi: 10.1111/j.1365-2958.1995.tb02390.x
71. Barbosa, C, Trebosc, V, Kemmer, C, Rosenstiel, P, Beardmore, R, Schulenburg, H, et al. Alternative evolutionary paths to bacterial antibiotic resistance cause distinct collateral effects. Mol Biol Evol. (2017) 34:2229–44. doi: 10.1093/molbev/msx158
72. Hernando-Amado, S, Blanco, P, Alcalde-Rico, M, Corona, F, Reales-Calderón, JA, Sánchez, MB, et al. Multidrug efflux pumps as main players in intrinsic and acquired resistance to antimicrobials. Drug Resist Updat. (2016) 28:13–27. doi: 10.1016/j.drup.2016.06.007
73. Sommer, LM, Johansen, HK, and Molin, S. Antibiotic resistance in Pseudomonas aeruginosa and adaptation to complex dynamic environments. Microb Genom. (2020) 6:e000370. doi: 10.1099/mgen.0.000370
74. Cirz, RT, Chin, JK, Andes, DR, de Crécy-Lagard, V, Craig, WA, and Romesberg, FE. Inhibition of mutation and combating the evolution of antibiotic resistance. PLoS Biol. (2005) 3:e176. doi: 10.1371/journal.pbio.0030176
75. Beaber, JW, Hochhut, B, and Waldor, MK. SOS response promotes horizontal dissemination of antibiotic resistance genes. Nature. (2004) 427:72–4. doi: 10.1038/nature02241
76. Davies, J, Spiegelman, GB, and Yim, G. The world of subinhibitory antibiotic concentrations. Curr Opin Microbiol. (2006) 9:445–53. doi: 10.1016/j.mib.2006.08.006
77. Couce, A, and Blázquez, J. Side effects of antibiotics on genetic variability. FEMS Microbiol Rev. (2009) 33:531–8. doi: 10.1111/j.1574-6976.2009.00165.x
78. Guerin, E, Cambray, G, Sanchez-Alberola, N, Campoy, S, Erill, I, Da Re, S, et al. The SOS response controls integron recombination. Science. (2009) 324:1034. doi: 10.1126/science.1172914
Keywords: microevolution, co-selection, MEGA-plate, Escherichia coli , potentiated sulphonamide, NGS, AMR, resistance development
Citation: Kerek Á, Török B, Szabó Á, Laczkó L, Kardos G, Bányai K, Kaszab E, Bali K and Jerzsele Á (2025) Resistance evolution under potentiated sulphonamide pressure in Escherichia coli. Front. Vet. Sci. 12:1697872. doi: 10.3389/fvets.2025.1697872
Edited by:
Kumar Siddharth Singh, Helmholtz Institute for One Health, GermanyReviewed by:
Gitartha Kaushik, Rangapara College, IndiaSantosh Anand, Birsa Agricultural University, India
Copyright © 2025 Kerek, Török, Szabó, Laczkó, Kardos, Bányai, Kaszab, Bali and Jerzsele. This is an open-access article distributed under the terms of the Creative Commons Attribution License (CC BY). The use, distribution or reproduction in other forums is permitted, provided the original author(s) and the copyright owner(s) are credited and that the original publication in this journal is cited, in accordance with accepted academic practice. No use, distribution or reproduction is permitted which does not comply with these terms.
*Correspondence: Ádám Kerek, a2VyZWsuYWRhbUB1bml2ZXQuaHU=